
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Cell. Infect. Microbiol., 05 December 2023
Sec. Microbes and Innate Immunity
Volume 13 - 2023 | https://doi.org/10.3389/fcimb.2023.1321315
This article is part of the Research TopicMicrobiota-Immune System Interplay: Insights into Inflammatory and Autoimmune DiseasesView all 6 articles
 Teresa Quintero-Puerta1
Teresa Quintero-Puerta1 Juan Alberto Lira-Lucio1
Juan Alberto Lira-Lucio1 Ramcés Falfán-Valencia1
Ramcés Falfán-Valencia1 Ángel E. Vega-Sánchez2
Ángel E. Vega-Sánchez2 Eduardo Márquez-García3
Eduardo Márquez-García3 Mayra Mejía2
Mayra Mejía2 Brandon Bautista-Becerril1
Brandon Bautista-Becerril1 Jorge Rojas-Serrano2
Jorge Rojas-Serrano2 Espiridión Ramos-Martínez4
Espiridión Ramos-Martínez4 Ivette Buendía-Roldán5
Ivette Buendía-Roldán5 Gloria Pérez-Rubio1*
Gloria Pérez-Rubio1*Aim: To characterize the lung microbiome in the bronchoalveolar lavage fluid (BALF) of patients with Antisynthetase Syndrome (ASSD) according to anti-Jo1 autoantibody positivity and evaluate the correlation with differential cell count and other bacterial genera in BALF.
Methods: We sequenced the 16S ribosomal RNA gene in the BALF of anti-Jo1-positive (JoP, n=6) and non-Jo1-positive (NJo, n=17) patients, and the differential cell count in BALF was evaluated. The Spearman’s correlation was calculated for the quantitative variables and abundance of bacterial species.
Results: The Veillonella genus showed a significant decrease (p<0.01) in JoP (2.2%) in comparison to NJo (4.1%) patients. The correlation analysis showed several high (rho ≥ ± 0.7) and significant (p < 0.05) correlations. We analyzed the results obtained for the Veillonella genera and other study variables. The JoP group showed that the abundance of Veillonella had a high negative correlation with macrophages (rho = - 0.77) and a positive correlation with eosinophils (rho = 0.77), lymphocytes (rho = 0.77), and Prevotella (rho = 1).
Conclusions: The lung microbiome in ASSD patients differs and may affect cell composition, contributing to lung damage mechanisms. The presence of anti-Jo1 autoantibodies showed a low abundance of Veillonella. This genus had a strong and positive correlation with Prevotella abundance and levels of eosinophils and lymphocytes, and it showed a strong negative correlation with the percentage of macrophages.
Antisynthetase syndrome (ASSD) is a rare autoimmune disease (1.5 cases per 100, 000) that commonly exists with extra-muscular manifestations, including fever, Raynaud’s syndrome, arthritis, mechanic’s hands, and interstitial lung disease (ILD) (Hamaguchi et al., 2013; Marie et al., 2013b; Rojas-Serrano et al., 2015; Pinal-Fernandez et al., 2017). This pathology is characterized by the presence of autoantibodies against various aminoacyl-transfer RNA (tRNA) synthetases (Witt et al., 2016; Pinal-Fernandez et al., 2017), including anti-Jo1 (most common), anti-PL7, anti-PL12, anti-OJ, anti-EJ, anti-KS, anti-Zo, anti-SC, anti-JS, and anti-YRS (Lega et al., 2010; Aggarwal et al., 2014; Rojas-Serrano et al., 2015; Pinal-Fernandez et al., 2017; Ponce-Gallegos et al., 2020). Independent of the autoantibody subtype, the leading predictive factors for a progressive pulmonary disease are a low diffusing capacity of the lungs for carbon monoxide (DLCO) at diagnosis, a low forced vital capacity (FVC) at baseline evaluation (Rojas-Serrano et al., 2015), and muscle weakness (Kamiya et al., 2018; Gasparotto et al., 2019; Liu et al., 2020). The abnormalities most often observed using high-resolution computed tomography (HRCT) are organized pneumonia (OP), followed by nonspecific interstitial pneumonia (NSIP) (González-Pérez et al., 2020).
Differences exist between anti-Jo1-positive (JoP) and non-Jo1-positive (NJo) patients, with the latter often demonstrating ILD in the absence of muscle involvement and having worse survival (Aggarwal et al., 2014); however, JoP patients present higher Goh inflammation score (Lega et al., 2010; Aggarwal et al., 2014; Rojas-Serrano et al., 2015; Pinal-Fernandez et al., 2017; Ponce-Gallegos et al., 2020). The Goh score is a semi-quantitative CT (SQCT) assessment that can estimate inflammation associated with ILD (Rojas-Serrano et al., 2015). Lung inflammation is a mechanism that has a crucial role in tissue repair, but if unregulated, it can be an underlying cause of chronic lung diseases (Huffnagle et al., 2017). Interactions between the host and lung microbiome, primarily at the mucosal surfaces, are fundamental to developing and regulating immunity mechanisms (Neish, 2014).
The lungs are constantly exposed to diverse communities of microbes from the oropharynx and other sources. The most abundant phyla of healthy lungs are Bacteroidetes and Firmicutes. At the genera level, Prevotella, Veillonella, and Streptococcus are predominant (Dickson et al., 2016). The disruption of the lung microbiome (dysbiosis) contributes to acute inflammation, progression, or exacerbations in diseases like asthma, chronic obstructive pulmonary disease (COPD), and interstitial lung disease (ILD) (Ren et al., 2018; Paudel et al., 2020). The most frequent ILD is idiopathic pulmonary fibrosis (IPF), where there is clear evidence of the microbiome’s contribution to collagen deposition in the lungs (O’Dwyer et al., 2019). In the microbiome of patients with stable IPF predominates Streptococcus, Prevotella, Veillonella, Haemophilus, and Pseudomonas. In contrast, IPF patients with acute exacerbation demonstrate increased Campylobacter sp. and Stenotrophomonas sp., associated with a significant decrease in Veillonella sp (Molyneaux et al., 2017). Disease progression has been associated with a high abundance of Streptococcus and Staphylococcus and a loss of diversity in the lungs (Han et al., 2014).
The dysbiosis of the lung microbiome is a major factor contributing to developing and progressing ILD associated with dermatomyositis, IPF, and rheumatoid arthritis. However, in ASSD, the lung microbiome has not been explored. This study aimed to characterize the lung microbiome in the bronchoalveolar lavage fluid (BALF) of patients with ASSD according to the presence or absence of anti-Jo1 autoantibodies and to evaluate correlation with differential cell count in BALF.
We included 23 patients with a confirmed diagnosis of ASSD who were recruited from the Interstitial Lung Disease and Rheumatology Unit of Instituto Nacional de Enfermedades Respiratorias Ismael Cosío Villegas (INER) in Mexico. A multidisciplinary group (pulmonologists, rheumatologists, and radiologists) performed the diagnosis. All patients were newly diagnosed. We registered the patients’ demographic variables, tobacco consumption, results of pulmonary function tests (including spirometry and diffusion capacity of the lungs for carbon monoxide, DLCO), levels of creatine-phosphokinase (CPK), differential cell count in BALF, clinical manifestations, and autoantibody patterns, which include anti-Mi-2α, anti-Mi-2β, anti-TIF1γ, anti-MDA5, anti-NXP2, anti-SAE1, anti-Ku, anti-PM-Scl100, anti-PM-Scl75, anti-Jo1, anti-SRP, anti-PL7, anti-PL12, anti-EJ, anti-OJ, and anti-Ro-52 (EUROIMMUN. Lübeck, Germany), as well as specific HRCT (Rojas-Serrano et al., 2015; González-Pérez et al., 2020). All participants were invited to donate their remnant BALF sample, signed an informed consent letter, and were provided with a personal data protection document. The exclusion criteria were the presence of an infectious disease, the use of antibiotics or immunosuppressive medications in the past three months, and evidence of acute upper respiratory symptoms in the previous four weeks. The patients were divided into anti-Jo1-positive (JoP) and non-Jo1-positive (NJo) groups. The Institutional Committees for Research, Biosecurity, and Ethics in Research of the INER approved this study (approval number B28-18).
The BALF samples were centrifuged at 3,000 rpm at 20˚C for 10 minutes, and the supernatant was stored at -30°C. The cell pellet was employed for bacterial DNA extraction and purified using the ZymoBIOMICS™ DNA Miniprep Kit (Zymo Research Corp. CA, EU), following the manufacturer’s instructions for liquid samples (200 µL). DNA was quantitated using the Qubit dsDNA High Sensitivity (HS) Assay Kit (Invitrogen) and visually assessed for integrity via electrophoresis on a 1% agarose gel.
We prepared the libraries for targeted amplicon sequencing following the “16S Metagenomic Sequencing Library Preparation” guide (Part# 15044223 Rev. B, Illumina). We used the primer pair sequence (forward primer = 5’ TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG; reverse primer = 5’ GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC) for the V3 and V4 regions of the 16S ribosomal RNA gene (16S rRNA) and Nextera XT indices. The sequencing was run on the MiSeq platform using paired 300 bp reads and MiSeq v3 reagents. We employed MiSeq Control Software v 3.1.1.13 (Illumina) to generate the FASTQ file. We included two control saline samples to identify sources of bacterial contamination; these controls were treated under the same conditions as the BALF.
We analyzed the FASTQ file using RStudio (R Studio, 2019), with the packages DADA2 (Callahan et al., 2016a), Phyloseq (McMurdie and Holmes, 2013), DESeq2 (Love et al., 2014), ggplot 2 (Lin Pedersen, 2019), and microbiome (Lahti et al., 2017), and followed the workflow of Bioconductor (Callahan et al., 2016b) and CastroLab (Castro Nallar, 2020). To assign taxonomy, we used the database of silva nr v138 and 97% similarity level (Quast et al., 2013). Alpha diversity was evaluated using the Chao1, Shannon, and Simpson indexes; beta diversity was evaluated using Bray–Curtis dissimilarity and principal coordinate analysis (PCoA). The readings were normalized to obtain the amplicon sequence variants (ASVs).
According to the results obtained using 16S metagenomics, we selected three bacterial species belonging to Veillonella with clinical relevance. The specificity of each primer set and probe was evaluated using the basic local alignment search tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) ([[NoAuthor]]) (Supplementary Table 1). A real-time PCR assay was performed using the StepOnePlus system (Applied Biosystems™). We used TaqMan custom assay to evaluate V. parvula and V. dispar. The reactions were performed using a total volume of 10 µl that contained 5 µl of Maxima Probe/ROX qPCR Master Mix (2X) (Thermo Scientific, Vilnius, Lithuania), 0.5 µl each of the forward and reverse primers (final concentration, 300 µM), 0.03 µl of TaqMan custom probe (final concentration 70 µM), 2 µl of the template DNA solution (20 ng/µl), and 2 µl of nuclease-free water. The thermal cycling conditions for all real-time PCR assays were as follows: 1 cycle at 50°C for 10 minutes, 1 cycle at 95°C for 10 minutes, followed by 45 cycles at 95°C for 15 seconds and at 60°C for 1 minute. For V. atypica, we used SYBR green qPCR in a total volume of 25 μL that contained 4 μl of the template DNA solution (20 ng/μL), 3.5 μL of nuclease-free water, 12.5 mL of Maxima SYBR Green/ROX qPCR Master Mix (2X) (Thermo Scientific, Vilnius, Lithuania), and 3 μL of the forward and 3 μL of reverse primers (final concentration 375 μM). The thermal cycling conditions for all real-time PCR assays were 1 cycle at 95°C for 10 minutes, 40 at 95°C for 30 seconds, and 1 at 56°C for 1 minute.
We employed the Shapiro–Wilk test to assess the normality of demographic characteristics, smoking status, pulmonary function test results, cells present in BALF, clinical manifestations, autoantibody profile, and HRCT pattern. For quantitative variables, we presented the means and standard deviations or medians and interquartile ranges (IQR); according to the distribution of the variables, we used parametric or non-parametric tests. The categorical data were analyzed using a contingency table and Fisher’s exact test. The identified ASVs were compared in both study groups; we employed a negative binomial generalized linear model (GLM) and obtained maximum likelihood estimates for an ASV log-fold change between two study groups; the p-value was adjusted based on the false discovery rate (FDR) measured using the Benjamin–Hochberg method. Alpha diversity was assessed using the Mann–Whitney U test. Beta diversity was assessed using the Wilcoxon signed-rank test. A p-value < 0.05 or an FDR < 0.05 was considered statistically significant. Spearman’s correlation was determined for the quantitative variables and abundance of bacteria. The results were presented using the ggplot2 package (Wickham, 2016) and corrplot (Wei and Simko, 2021). The dataset supporting the results of this article has been deposited in the NCBI Short Read Archive database under BioProject accession code PRJNA715451.
We included 23 ASSD patients; six were JoP and 17 NJo (Table 1). Age and sex did not differ significantly between the two groups; however, JoP patients were older in comparison to the NJo group (62 vs. 55 years), and the female sex was predominant in both groups (83.3% vs. 58.3%) compared to the male sex (16.7% and 41.7%). The proportion of smokers was similar (33.3% vs. 35.3%), and the tobacco index was higher in JoP patients compared to NJo patients (7.1 pack-years vs. 4.5 pack-years), although this difference was not statistically significant. DLCO was lower in JoP patients compared to NJo patients, but there was no statistical difference (42% vs. 49%). Both groups had moderate DLCO reduction (Ponce et al., 2022). Patients with JoP status had a smaller value in the 6MWT than NJo patients (120 vs. 405, respectively, p=0.01). Patients JoP showed high levels of CPK compared to the NJo group (242.5 vs. 67.5 U/L, respectively, p=0.001). The tomography patterns were inflammatory in both groups (JoP = 66.7% and NJo = 70.6%). The inflammatory component was defined based on ground-glass opacities, while the fibrotic component was defined based on reticular opacities (Rojas-Serrano et al., 2015). In the BALF samples, macrophages and lymphocytes were the predominant cells.
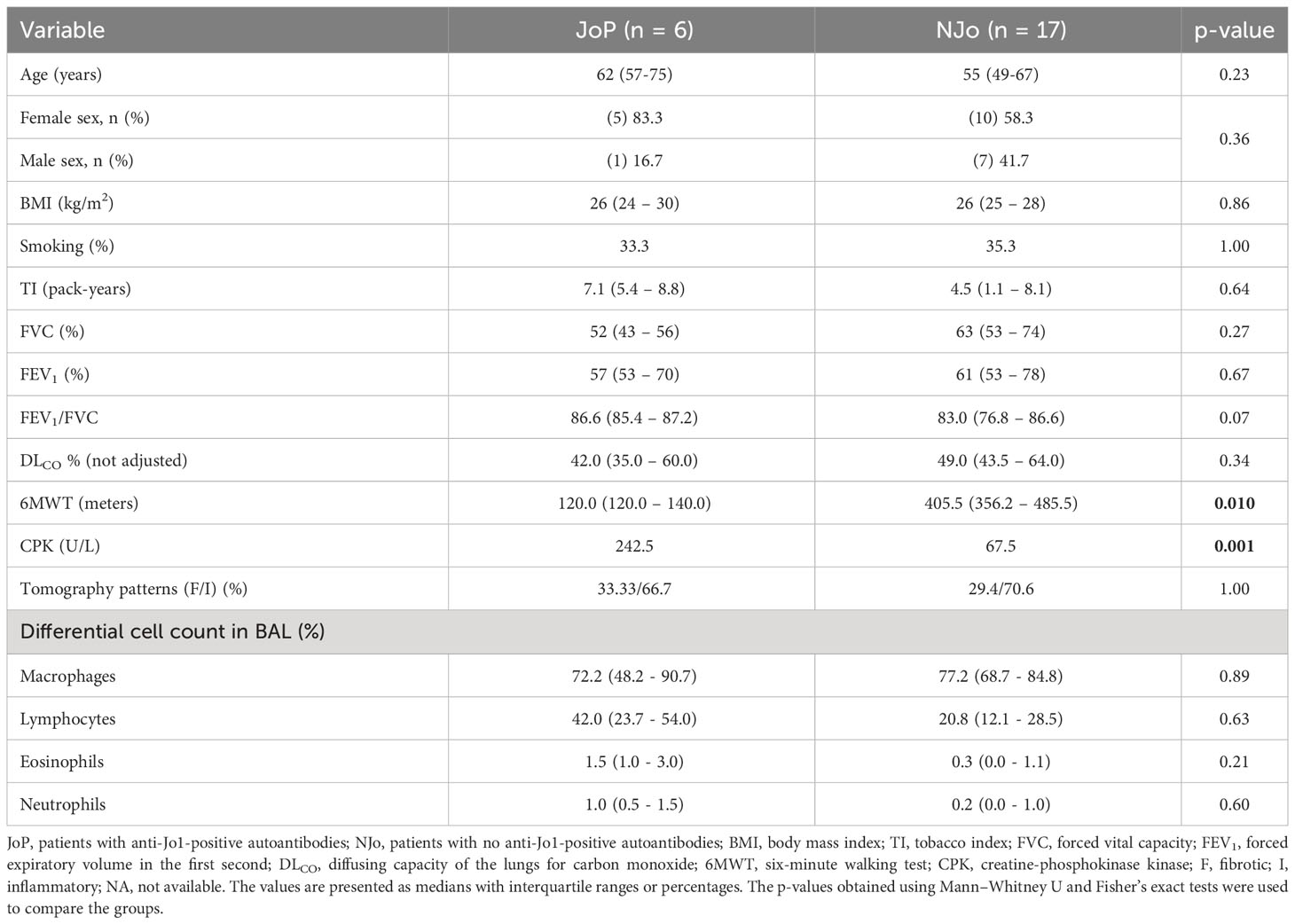
Table 1 Demographic and clinical characteristics of the patients in this study.
The most frequent clinical manifestations in all patients were mechanic’s hands (43.5%), dyspnea (43.5%), and cough (43.5%). In addition, only the NJo group presented fever (35.3%) and weight loss (23.5%) (Supplementary Table 2). The patients in this study showed an overlap of antibodies typical of myositis and sclerosis. JoP patients showed a predominance of anti-OJ (33.3%), anti-MI-2β (33.3%), and anti- Th/To (33.3%), whereas NJo patients showed a predominance of anti-Ro52, anti-PL7, anti-PL12 (35.3% in each autoantibody), and Th/To (23.5%) (Supplementary Table 3).
We obtained 115 ASVs, and after performing quality control, we identified 10 ASVs with an abundance of >1,000 reads present in ≥ 10% of the samples analyzed. The datasets analyzed in this study (FASTQ file) can be found in online repositories (Sequence Read Archive, SRA; submission number: SUB9326500). The analysis of alpha diversity showed that there were no significant differences in the Shannon (p = 0.92), Simpson (p = 0.43), and Chao1 (p = 1) indexes (Supplementary Figure 1) between JoP and NJo groups. Beta diversity showed a statistically significant difference in divergence (p = 0.016), and using PCoA, we observed that overlap existed between the JoP and NJo groups (Supplementary Figures 2A, B). Both groups of patients predominantly showed phyla Firmicutes (71.32%), Bacteroidota (17.11%), and Proteobacteria (11.29%) (Figure 1A). The top five genera in all patients were Streptococcus (57.8%), Sediminibacterium (11.34%), Veillonella (8.92%), Sphingomonas (6.02%), and Prevotella (5.77%) (Figure 1B). A comparison of relative abundance in the two groups identified a statistically significant difference in the relative abundance of Veillonella (p < 0.01); this genus showed a low abundance in JoP patients (2.2%) in comparison to NJo (4.1%) patients (Figure 1C).
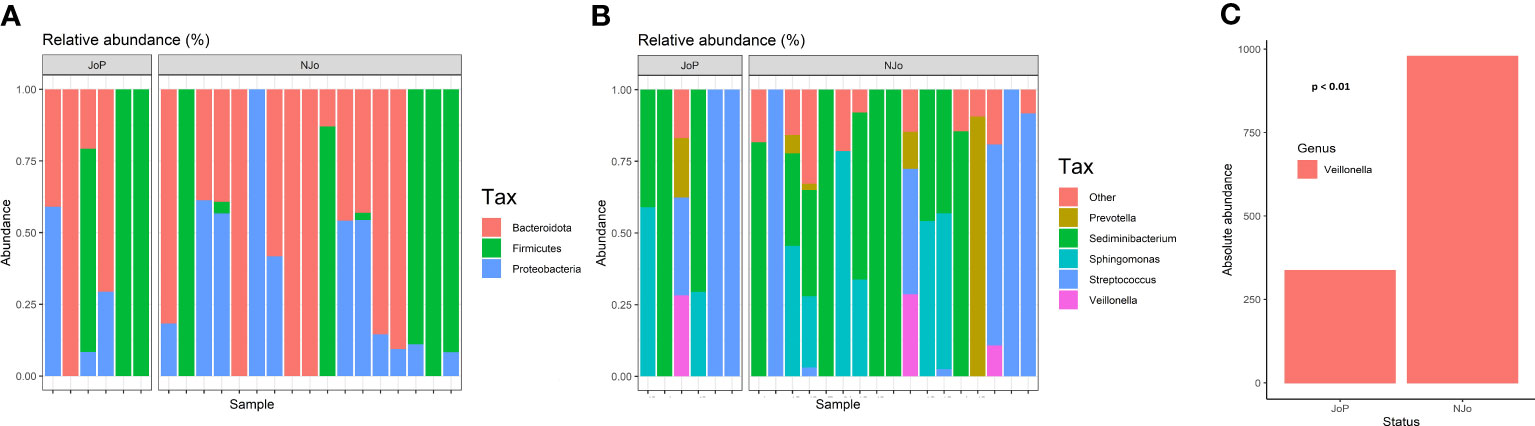
Figure 1 Composition of the lung microbiome in antisynthetase syndrome (ASSD) patients: (A) relative abundance of different phyla in both study groups; (B) top five genera in ASSD patients (according to relative abundance); and (C) Absolute abundance of Veillonella in JoP and NJo patients. This genus is lower in JoP patients (2.2%) than in NJo patients (4.1%).
The correlation analysis showed several high (rho ≥ 0.7) and significant (p < 0.05) correlations. We analyzed the results obtained for the Veillonella genus and other study variables. In the JoP group, Veillonella had a high negative correlation with macrophages (rho = - 0.77) and a positive correlation with eosinophils (rho = 0.77), lymphocytes (rho = 0.77) and Prevotella (rho = 1). In contrast, NJo patients did not show high correlations (Figures 2A, B).
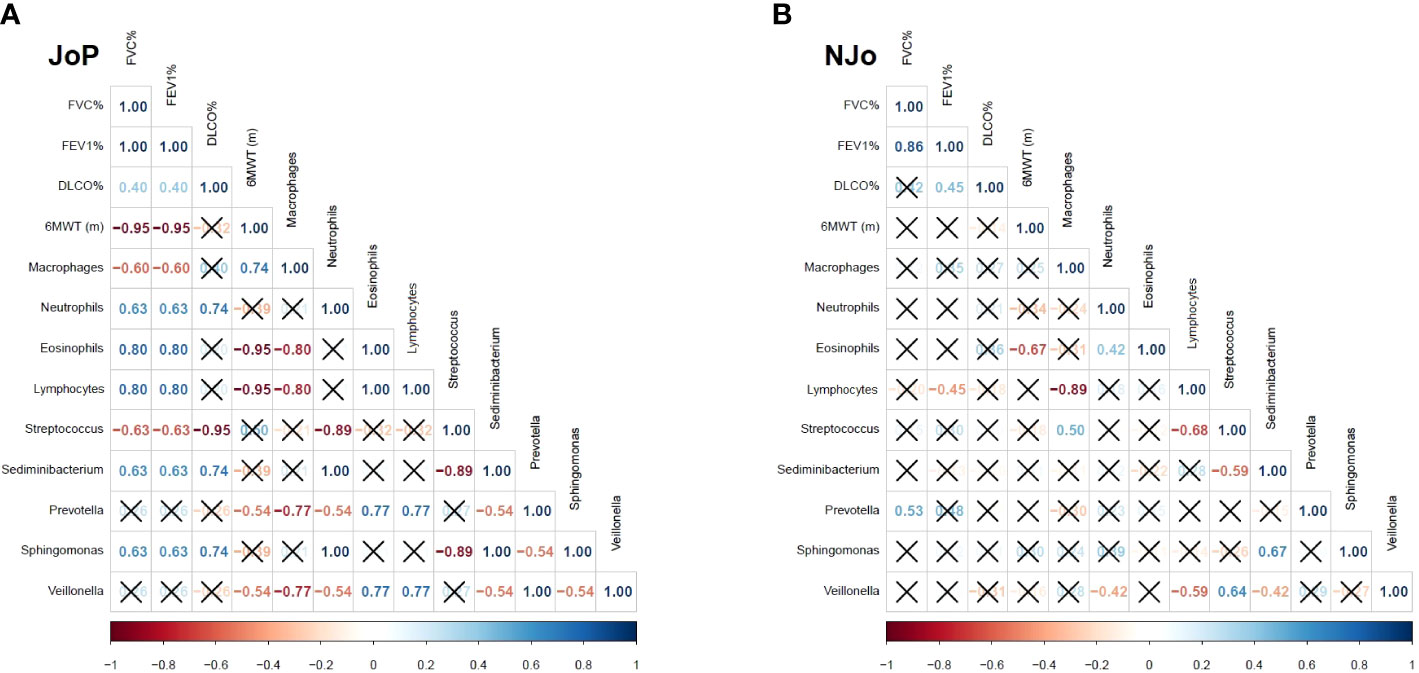
Figure 2 Spearman’s correlation in antisynthetase syndrome (ASSD) patients: (A) correlation analysis in the JoP group and (B) correlation analysis in the NJo group. Only the rho coefficients of significative correlations are shown.
Identifying specific species of Veillonella showed that V. parvula, V. dispar, and V. atypica had no significant difference between the study groups. We observed more patients who were positive for V. dispar in the NJo group (40.7%) compared to the JoP group (25.0%) (Table 2).
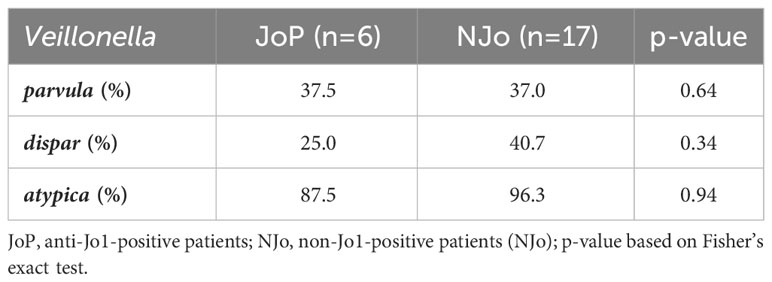
Table 2 Species identification in the study sample.
Previous reports have evaluated the lung microbiome and its influences on the course of various diseases, like asthma, cystic fibrosis, and chronic obstructive pulmonary disease; however, this is the first study to characterize the bacterial communities in patients with ASSD and ILD, which is the most frequent and severe extra-muscular manifestation (Gasparotto et al., 2019). The present study used the presence or absence of anti-aminoacyl-tRNA synthetase (anti-Jo1) autoantibodies with the strongest association with the development and severity of ILD in ASSD (Cavagna et al., 2019) to stratify the study patients.
We included a population of patients with ASSD. There were no significant differences in age, sex, BMI, TI, or pulmonary function test (FVC, FEV1, FEV1/FVC, and DLCO). Patients in the anti-Jo1-positive group walked significantly shorter distances during the 6MWT compared to the anti-Jo1-negative patients (p=0.01, 120 m vs. 405.5 m, respectively). The anti-Jo1-positive group had a tendency to show decreased lung function compared to the anti-Jo1-negative group; this finding was similar to that reported in a Mayo Clinic cohort study, where 53% of its patients with anti-Jo1-positive ASSD had decreased lung function, regardless of treatment (Zamora et al., 2016). In Caucasian patients, the functional outcomes of the anti-Jo1 patients with ASSD were poor, as shown by the marked reduction of activities evaluated according to the disability scale of the Health Assessment Questionnaire (HAQ > 0.75) (Marie et al., 2013a). In our study group, ASSD patients positive for anti-Jo1 antibody present more important muscle involvement, represented by higher levels of CPK (Ponce-Gallegos et al., 2020).
The predominant tomographic pattern was inflammatory, regardless of autoantibodies classification (66.7% in anti-Jo1-positive and 70.6% in anti-Jo1-negative patients). Cells showing a higher abundance in BAL were macrophages and lymphocytes. These findings correspond to the findings previously reported in ASSD (Lang et al., 2021) and other interstitial diseases, such as rheumatoid arthritis associated with ILD (Lee et al., 2005).
Interestingly, no significant difference was found in alpha diversity (Chao1, Shannon, and Simpson indices); we assume this is because both groups were patients diagnosed with ASSD. The loss of bacterial diversity has been linked to pulmonary diseases with increased profibrotic and proinflammatory cytokines (O’Dwyer et al., 2019), contributing to the severe outcomes of diseases such as COPD, asthma, cystic fibrosis, and idiopathic pulmonary fibrosis (Tunney et al., 2013; Yagi et al., 2021). The alpha diversity of the lung microbiome did not show differences between dermatomyositis and rheumatoid arthritis associated with ILD (Lou et al., 2022) or compared with patients with rheumatoid arthritis and sarcoidosis (Scher et al., 2016).
In the analysis of beta diversity, we found a divergence between the two study groups (p=0.016); this is probably due to the heterogeneity in the clinical manifestations of ASSD, as well as the overlap in autoantibodies that exists in the participants, regardless of the presence or absence of anti-Jo1 positivity.
At the phylum level, the lung microbiome of patients with ASSD comprises Firmicutes, Bacteroidota, and Proteobacteria. In healthy individuals, previous reports indicate a predominance of the genera Prevotella, Streptococcus, Veillonella, Neisseria, Haemophilus, and Fusobacterium (Segal et al., 2016; Yu et al., 2016). In our study, patients with ASSD showed a predominance of Streptococcus, Sediminibacterium, Veillonella, Sphingomonas, and Prevotella; the genus Veillonella was found in lower abundance in the JoP group compared to the NJo group (p<0.01). This bacterial genus is anaerobic and can tolerate environments with higher oxidative stress because all species of Veillonella have the rbr gene that codes for ruberythrins; these enzymes have an antioxidant mechanism that gives them the ability to catalyze hydrogen peroxide and, consequently, reactive oxygen species in the microenvironment decrease (Zhou et al., 2017).
Patients in the JoP group showed a very strong positive correlation between the abundance levels of Veillonella and Prevotella, which are anaerobic bacteria. In cystic fibrosis, it has been reported that Prevotella can produce short-chain fatty acids (acetic, i-butyric, 2-methyl butyric, and i-valeric) that activate epithelial cells in the airways to produce cytokines (Mirković et al., 2015), act as chemotactic agents for neutrophils (Vinolo et al., 2011), and affect phagocytosis (Vinolo et al., 2009). There is also a strong positive correlation between the levels of Sediminibacterium and Sphingomonas. Previous in vitro assays have shown that Sphingomonas can induce apoptosis in human lung cells via the activation of caspases 3 and 9 (Asghar et al., 2021). Sediminibacterium is abundant in the blood of patients with type 2 diabetes (Qiu et al., 2019); however, it has not been reported in lung diseases. In addition, a strong negative correlation was found between the levels of Streptococcus and neutrophils; previous research with mouse models has shown that the M1 protein of Streptococcus forms complexes with fibrinogen that binds to β2 integrin on the surface of neutrophils, which causes degranulation, increased vascular permeability, lung damage, and remodeling (Soehnlein et al., 2008).
As for ASSD patients who were anti-Jo1 negative, those with better parameters in terms of respiratory function and significantly greater 6MWT presented moderate positive correlations between the relative abundance levels of Veillonella and Streptococcus. It should be noted that certain species of Streptococcus produce hydrogen peroxide, which can be used by Veillonella, thus generating a reducing microenvironment that improves the conditions for the establishment of other bacteria that are unable to tolerate an environment with reactive oxygen species (Zhou et al., 2017). Furthermore, a less inflammatory microenvironment could be generated due to the abundance of Veillonella in anti-Jo1-negative patients. This could explain the finding of the strong correlation between the abundance of Streptococcus and DLCO (p<0.001, rho=-0.95). We did not find a strong correlation and statistically significant differences between FVC%, FEV1%, DLCO%, or 6MWT and Veillonella, Sphingomonas, Prevotella, and Sediminebacterium.
When we evaluated Veillonella parvula, atypica, and dispar species, there were no statistically significant differences between the study groups; however, V. dispar was less abundant in the JoP patients group than in the NJo group. Veillonella has been associated with different diseases, such as asthma (Espuela-Ortiz et al., 2019), COPD, and IPF exacerbations (Molyneaux et al., 2017).
Several reports conclude that the presence of Veillonella is good for the health of humans. In patients with non-small cell lung cancer, V. dispar is predominated in groups with high levels of programmed cell death-ligand 1 (PD-L1), which are responders to immunotherapy, proving that the composition of the lung microbiome may influence disease development as well as treatment efficacy (Jang et al., 2021).
In evaluating the gut microbiome of elite athletes, the presence of V. atypica improves runtime via its metabolic conversion of exercise-induced lactate into propionate, thereby identifying a natural enzymatic process that enhances athletic performance (Scheiman et al., 2019). In diseases related to chronic hypoxemia, there is a high metabolism of lactate (Iscra et al., 2002), which Veillonella sp. could metabolize and shows a similar effect as that at the intestinal level; these findings could explain why patients included in this study who had a higher abundance of Veillonella exhibited more time walking in the 6MWT.
The knowledge of the bacterial communities in patients with ASSD, as well as the interaction mechanisms related to the immune response and remodeling of the pulmonary epithelium, will allow the identification of possible bacterial genera as biomarkers of ASSD, with the purpose of a better the understanding, management, and treatment of ASSD-ILD. In patients with ASSD, greater caspase-1, and higher LDH activity were observed in BAL, suggesting cell death due to pyroptosis and activation of the inflammasome pathway (Ramos-Martinez et al., 2022). Serum levels of 18 cytokines from baseline and after six months of treatment were quantified in patients with anti-tRNA associated ILD (anti-tRNA-ILD) and estimated the association between these and ILD improvement and progression; patients were classified as with or without ILD progression at six months. Only three patients had ILD progression (progressors patients, PP) and showed statistically higher levels in IL-4, IL-10, IL-17A, IL-22, GM-CSF, IL-1β, IL-6, IL-12, IL-18, and TNF-α, compared to patients without disease progression (no progressors patients, NPP). IL-17A, IL-1β, and IL-6 (T-helper-lymphocyte (Th)17 inflammatory cytokine profile) were elevated and had a high discriminatory capacity in distinguishing ILD PP of those NPP at follow-up (Ramos-Martinez et al., 2020).
Our findings show that the composition of the lung microbiome is different according to the profile of autoantibodies present in patients with ASSD-ILD, and in turn, there are correlations between bacterial genera and cells in the BAL that depend on anti-Jo1 autoantibodies. The methodology used to evaluate the lung microbiome only allows the exact identification up to bacterial genus. Our results provide evidence that patients with ASSD and positive anti-Jo1 autoantibodies have decreased Veillonella abundance, correlated with an increase in the count of lymphocytes, eosinophils, and other bacteria relevant to lung diseases.
The main limitation of this study was the lack of a control group. In the literature, there are reports about lung microbiome composition in healthy subjects. Firmicutes and Bacteroidota are the predominant phyla in healthy lung microbiota, with Prevotella, Veillonella, and Streptococcus being the most common genera (Mathieu et al., 2018; Yi et al., 2022). Pathological conditions lead to a loss of that diversity, with increased concentrations of some bacterial genera (Costa et al., 2018). We found a low abundance of Veillonella in patients with ASDD and anti-Jo1 autoantibodies; however, because of the heterogeneity of the lung microbiome between individuals, the characterization of the lung microbiome should be verified in a larger study.
The lung microbiome in ASSD patients with anti-Jo1 autoantibodies showed a low abundance of Veillonella compared to non-Jo1-positive ASSD patients. This genus had a strong and positive correlation with Prevotella abundance and levels of eosinophils and lymphocytes, and it showed a strong negative correlation with the percentage of macrophages. These correlations were not observed in non-Jo1-positive patients.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, BioProject accession code PRJNA715451.
The Institutional Committee on Ethics and Research reviewed and approved the study (ethical protocol number B28-18). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
TQ-P: Formal analysis, Resources, Writing – original draft. JL-L: Methodology, Software, Writing – original draft. RF-V: Funding acquisition, Supervision, Validation, Writing – review & editing. ÁV-S: Data curation, Validation, Writing – review & editing. EM-G: Resources, Validation, Writing – review & editing. MM: Supervision, Visualization, Writing – review & editing. BB-B: Formal analysis, Investigation, Writing – review & editing. JR-S: Validation, Visualization, Writing – review & editing. ER-M: Data curation, Writing – review & editing. IB-R: Data curation, Visualization, Writing – review & editing. GP-R: Formal analysis, Investigation, Project administration, Writing – original draft.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work is supported by the allocated budget to research (HLA Laboratory) from the Instituto Nacional de Enfermedades Respiratorias Ismael Cosío Villegas (INER).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1321315/full#supplementary-material
Aggarwal, R., Cassidy, E., Fertig, N., Koontz, D. C., Lucas, M., Ascherman, D. P., et al. (2014). Patients with non-Jo-1 anti-tRNA-synthetase autoantibodies have worse survival than Jo-1 positive patients. Ann. Rheum Dis. 73 (1), 227–232. doi: 10.1136/annrheumdis-2012-201800
Asghar, M. T., Khurshid, M., Nazir, J., Shakoori, A. R. (2021). Induction of apoptosis in human lung epithelial cell by sphingomonas sp. Shah, a recently identified cell culture contaminant. Crit. Rev. Eukaryot Gene Expr. 31 (2), 55–62. doi: 10.1615/CritRevEukaryotGeneExpr.2021037677
(2021) BLAST: Basic Local Alignment Search Tool. Available at: https://blast.ncbi.nlm.nih.gov/Blast.cgi.
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J., Holmes, S. P. (2016a). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13 (7), 581–583. doi: 10.1038/nmeth.3869
Callahan, B. J., Sankaran, K., Fukuyama, J. A., McMurdie, P. J., Holmes, S. P. (2016b). Bioconductor Workflow for Microbiome Data Analysis: from raw reads to community analyses. F1000Res 5, 1492. doi: 10.12688/f1000research.8986.2
Castro Nallar, E. (2020). Introducción al análisis de diversidad | Diseño experimental y análisis de datos (Santiago, Chile: Castro Lab). Available at: http://www.castrolab.org/teaching/data_analysis/intro-analisis-diversidad.html.
Cavagna, L., Trallero-Araguás, E., Meloni, F., Cavazzana, I., Rojas-Serrano, J., Feist, E., et al. (2019). Influence of antisynthetase antibodies specificities on antisynthetase syndrome clinical spectrum time course. J. Clin. Med. 8 (11), 2013. doi: 10.3390/jcm8112013
Costa, A. N., Costa, F. M. D., Campos, S. V., Salles, R. K., Athanazio, R. A. (2018). The pulmonary microbiome: challenges of a new paradigm. J. Bras. Pneumol. 44 (5), 424–432. doi: 10.1590/s1806-37562017000000209
Dickson, R. P., Erb-Downward, J. R., Martinez, F. J., Huffnagle, G. B. (2016). The microbiome and the respiratory tract. Annu. Rev. Physiol. 78, 481–504. doi: 10.1146/annurev-physiol-021115-105238
Espuela-Ortiz, A., Lorenzo-Diaz, F., Baez-Ortega, A., Eng, C., Hernandez-Pacheco, N., Oh, S. S., et al. (2019). Bacterial salivary microbiome associates with asthma among african american children and young adults. Pediatr. Pulmonol. 54 (12), 1948–1956. doi: 10.1002/ppul.24504
Gasparotto, M., Gatto, M., Saccon, F., Ghirardello, A., Iaccarino, L., Doria, A. (2019). Pulmonary involvement in antisynthetase syndrome. Curr. Opin. Rheumatol. 31 (6), 603–610. doi: 10.1097/BOR.0000000000000663
González-Pérez, M. I., Mejía-Hurtado, J. G., Pérez-Román, D. I., Buendía-Roldán, I., Mejía, M., Falfán-Valencia, R., et al. (2020). Evolution of pulmonary function in a cohort of patients with interstitial lung disease and positive for antisynthetase antibodies. J. Rheumatol. 47 (3), 415–423. doi: 10.3899/jrheum.181141
Hamaguchi, Y., Fujimoto, M., Matsushita, T., Kaji, K., Komura, K., Hasegawa, M., et al. (2013). Common and distinct clinical features in adult patients with anti-aminoacyl-tRNA synthetase antibodies: heterogeneity within the syndrome. PloS One 8 (4), e60442. doi: 10.1371/journal.pone.0060442
Han, M. K., Zhou, Y., Murray, S., Tayob, N., Noth, I., Lama, V. N., et al. (2014). Lung microbiome and disease progression in idiopathic pulmonary fibrosis: an analysis of the COMET study. Lancet Respir. Med. 2 (7), 548–556. doi: 10.1016/S2213-2600(14)70069-4
Huffnagle, G. B., Dickson, R. P., Lukacs, N. W. (2017). The respiratory tract microbiome and lung inflammation: a two-way street. Mucosal Immunol. 10 (2), 299–306. doi: 10.1038/mi.2016.108
Iscra, F., Gullo, A., Biolo, G. (2002). Bench-to-bedside review: lactate and the lung. Crit. Care 6 (4), 327–329. doi: 10.1186/cc1519
Jang, H. J., Choi, J. Y., Kim, K., Yong, S. H., Kim, Y. W., Kim, S. Y., et al. (2021). Relationship of the lung microbiome with PD-L1 expression and immunotherapy response in lung cancer. Respir. Res. 22 (1), 322. doi: 10.1186/s12931-021-01919-1
Kamiya, H., Panlaqui, O. M., Izumi, S., Sozu, T. (2018). Systematic review and meta-analysis of prognostic factors for idiopathic inflammatory myopathy-associated interstitial lung disease. BMJ Open 8 (12), e023998. doi: 10.1136/bmjopen-2018-023998
Lahti, L., Sudarshan, S., et al. (2017) Introduction to the microbiome R package [Internet]. Bioconductor. Available at: https://microbiome.github.io/tutorials/.
Lang, D., Akbari, K., Horner, A., Hepp, M., Kaiser, B., Pieringer, H., et al. (2021). Computed tomography findings as determinants of local and systemic inflammation biomarkers in interstitial lung diseases: A retrospective registry-based descriptive study. Lung 199 (2), 155–164. doi: 10.1007/s00408-021-00434-w
Lee, H. K., Kim, D. S., Yoo, B., Seo, J. B., Rho, J. Y., Colby, T. V., et al. (2005). Histopathologic pattern and clinical features of rheumatoid arthritis-associated interstitial lung disease. Chest 127 (6), 2019–2027. doi: 10.1378/chest.127.6.2019
Lega, J. C., Cottin, V., Fabien, N., Thivolet-Béjui, F., Cordier, J. F. (2010). Interstitial lung disease associated with anti-PM/Scl or anti-aminoacyl-tRNA synthetase autoantibodies: a similar condition? J. Rheumatol 37 (5), 1000–1009. doi: 10.3899/jrheum.090652
Lin Pedersen, T. (2019) Package “ggplot2” Title Create Elegant Data Visualisations Using the Grammar of Graphics. Available at: https://cran.r-project.org/web/packages/ggplot2/ggplot2.pdf.
Liu, Y., Liu, X., Xie, M., Chen, Z., He, J., Wang, Z., et al. (2020). Clinical characteristics of patients with anti-EJ antisynthetase syndrome associated interstitial lung disease and literature review. Respir. Med. 165, 105920. doi: 10.1016/j.rmed.2020.105920
Lou, Y., Wei, Q., Fan, B., Zhang, L., Wang, X., Chen, Z., et al. (2022). The composition of the lung microbiome differs between patients with dermatomyositis and rheumatoid arthritis associated with interstitial lung disease. FEBS Open Bio 12 (1), 258–269. doi: 10.1002/2211-5463.13334
Love, M. I., Huber, W., Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15 (12), 550. doi: 10.1186/s13059-014-0550-8
Marie, I., Hatron, P. Y., Cherin, P., Hachulla, E., Diot, E., Vittecoq, O., et al. (2013a). Functional outcome and prognostic factors in anti-Jo1 patients with antisynthetase syndrome. Arthritis Res. Ther. 15 (5), R149. doi: 10.1186/ar4332
Marie, I., Josse, S., Hatron, P. Y., Dominique, S., Hachulla, E., Janvresse, A., et al. (2013b). Interstitial lung disease in anti-Jo-1 patients with antisynthetase syndrome. Arthritis Care Res. (Hoboken). 65 (5), 800–808. doi: 10.1002/acr.21895
Mathieu, E., Escribano-Vazquez, U., Descamps, D., Cherbuy, C., Langella, P., Riffault, S., et al. (2018). Paradigms of lung microbiota functions in health and disease, particularly, in asthma. Front. Physiol. 9, 1168. doi: 10.3389/fphys.2018.01168
McMurdie, P. J., Holmes, S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS One 8 (4), e61217. doi: 10.1371/journal.pone.0061217
Mirković, B., Murray, M. A., Lavelle, G. M., Molloy, K., Azim, A. A., Gunaratnam, C., et al. (2015). The role of short-chain fatty acids, produced by anaerobic bacteria, in the cystic fibrosis airway. Am. J. Respir. Crit. Care Med. 192 (11), 1314–1324. doi: 10.1164/rccm.201505-0943OC
Molyneaux, P. L., Cox, M. J., Wells, A. U., Kim, H. C., Ji, W., Cookson, W. O., et al. (2017). Changes in the respiratory microbiome during acute exacerbations of idiopathic pulmonary fibrosis. Respir. Res. 18 (1), 29. doi: 10.1186/s12931-017-0511-3
Neish, A. S. (2014). Mucosal immunity and the microbiome. Ann. Am. Thorac. Soc 11 Suppl 1 (Suppl 1), S28–S32. doi: 10.1513/AnnalsATS.201306-161MG
O’Dwyer, D. N., Ashley, S. L., Gurczynski, S. J., Xia, M., Wilke, C., Falkowski, N. R., et al. (2019). Lung microbiota contribute to pulmonary inflammation and disease progression in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 199 (9), 1127–1138. doi: 10.1164/rccm.201809-1650OC
Paudel, K. R., Dharwal, V., Patel, V. K., Galvao, I., Wadhwa, R., Malyla, V., et al. (2020). Role of lung microbiome in innate immune response associated with chronic lung diseases. Front. Med. (Lausanne). 7, 554. doi: 10.3389/fmed.2020.00554
Pinal-Fernandez, I., Casal-Dominguez, M., Huapaya, J. A., Albayda, J., Paik, J. J., Johnson, C., et al. (2017). A longitudinal cohort study of the anti-synthetase syndrome: increased severity of interstitial lung disease in black patients and patients with anti-PL7 and anti-PL12 autoantibodies. Rheumatol. (Oxford). 56 (6), 999–1007. doi: 10.1093/rheumatology/kex021
Ponce, M. C., Sankari, A., Sharma, S. (2022). “Pulmonary Function Tests,” in StatPearls (Treasure Island (FL: StatPearls Publishing).
Ponce-Gallegos, M. A., Ramos-Martínez, E., García-Carmona, A., Mejía, M., Nava-Quiroz, K. J., Pérez-Rubio, G., et al. (2020). Genetic susceptibility to antisynthetase syndrome associated with single-nucleotide variants in the IL1B gene that lead variation in IL-1β Serum levels. Front. Med. (Lausanne). 7, 547186. doi: 10.3389/fmed.2020.547186
Qiu, J., Zhou, H., Jing, Y., Dong, C. (2019). Association between blood microbiome and type 2 diabetes mellitus: A nested case-control study. J. Clin. Lab. Anal. 33 (4), e22842. doi: 10.1002/jcla.22842
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41 (Database issue), D590–D596. doi: 10.1093/nar/gks1219
Ramos-Martinez, E., Falfán-Valencia, R., Pérez-Rubio, G., Mejia, M., Buendía-Roldán, I., González-Pérez, M. I., et al. (2020). Anti-aminoacyl transfer-RNA-synthetases (Anti-tRNA) autoantibodies associated with interstitial lung disease: pulmonary disease progression has a persistent elevation of the th17 cytokine profile. J. Clin. Med. 9 (5), 1356. doi: 10.3390/jcm9051356
Ramos-Martinez, E., Vega-Sánchez, A. E., Pérez-Rubio, G., Mejia, M., Buendía-Roldán, I., González-Pérez, M. I., et al. (2022). Enhanced activity of NLRP3 inflammasome in the lung of patients with anti-synthetase syndrome. Cells 12 (1), 60. doi: 10.3390/cells12010060
Ren, L., Zhang, R., Rao, J., Xiao, Y., Zhang, Z., Yang, B., et al. (2018). Transcriptionally active lung microbiome and its association with bacterial biomass and host inflammatory status. mSystems 3 (5), e00199–e00118. doi: 10.1128/mSystems.00199-18
Rojas-Serrano, J., Herrera-Bringas, D., Mejía, M., Rivero, H., Mateos-Toledo, H., Figueroa, J. E. (2015). Prognostic factors in a cohort of antisynthetase syndrome (ASS): serologic profile is associated with mortality in patients with interstitial lung disease (ILD). Clin. Rheumatol. 34 (9), 1563–1569. doi: 10.1007/s10067-015-3023-x
R Studio (2019) RStudio | Open source & professional software for data science teams - RStudio [Internet]. Available at: https://www.rstudio.com/.
Scheiman, J., Luber, J. M., Chavkin, T. A., MacDonald, T., Tung, A., Pham, L. D., et al. (2019). Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 25 (7), 1104–1109. doi: 10.1038/s41591-019-0485-4
Scher, J. U., Joshua, V., Artacho, A., Abdollahi-Roodsaz, S., Öckinger, J., Kullberg, S., et al. (2016). The lung microbiota in early rheumatoid arthritis and autoimmunity. Microbiome 4 (1), 60. doi: 10.1186/s40168-016-0206-x
Segal, L. N., Clemente, J. C., Tsay, J. C., Koralov, S. B., Keller, B. C., Wu, B. G., et al. (2016). Enrichment of the lung microbiome with oral taxa is associated with lung inflammation of a Th17 phenotype. Nat. Microbiol. 1, 16031. doi: 10.1038/nmicrobiol.2016.31
Soehnlein, O., Oehmcke, S., Ma, X., Rothfuchs, A. G., Frithiof, R., van Rooijen, N., et al. (2008). Neutrophil degranulation mediates severe lung damage triggered by streptococcal M1 protein. Eur. Respir. J. 32 (2), 405–412. doi: 10.1183/09031936.00173207
Tunney, M. M., Einarsson, G. G., Wei, L., Drain, M., Klem, E. R., Cardwell, C., et al. (2013). Lung microbiota and bacterial abundance in patients with bronchiectasis when clinically stable and during exacerbation. Am. J. Respir. Crit. Care Med. 187 (10), 1118–1126. doi: 10.1164/rccm.201210-1937OC
Vinolo, M. A., Ferguson, G. J., Kulkarni, S., Damoulakis, G., Anderson, K., Bohlooly-Y, M., et al. (2011). SCFAs induce mouse neutrophil chemotaxis through the GPR43 receptor. PloS One 6 (6), e21205. doi: 10.1371/journal.pone.0021205
Vinolo, M. A., Hatanaka, E., Lambertucci, R. H., Newsholme, P., Curi, R. (2009). Effects of short chain fatty acids on effector mechanisms of neutrophils. Cell Biochem. Funct. 27 (1), 48–55. doi: 10.1002/cbf.1533
Wei, T., Simko, V. (2021) R package “corrplot”: Visualization of a Correlation Matrix. Available at: https://github.com/taiyun/corrplot.
Wickham, H. (2016). ggplot2: Elegant Graphics for Data Analysis (New York: Springer-Verlag). Available at: https://ggplot2.tidyverse.org.
Witt, L. J., Curran, J. J., Strek, M. E. (2016). The diagnosis and treatment of antisynthetase syndrome. Clin. Pulm Med. 23 (5), 218–226. doi: 10.1097/CPM.0000000000000171
Yagi, K., Huffnagle, G. B., Lukacs, N. W., Asai, N. (2021). The lung microbiome during health and disease. Int. J. Mol. Sci. 22 (19), 10872. doi: 10.3390/ijms221910872
Yi, X., Gao, J., Wang, Z. (2022). The human lung microbiome—A hidden link between microbes and human health and diseases. iMeta 1, e33. doi: 10.1002/imt2.33
Yu, G., Gail, M. H., Consonni, D., Carugno, M., Humphrys, M., Pesatori, A. C., et al. (2016). Characterizing human lung tissue microbiota and its relationship to epidemiological and clinical features. Genome Biol. 17 (1), 163. doi: 10.1186/s13059-016-1021-1
Zamora, A. C., Hoskote, S. S., Abascal-Bolado, B., White, D., Cox, C. W., Ryu, J. H., et al. (2016). Clinical features and outcomes of interstitial lung disease in anti-Jo-1 positive antisynthetase syndrome. Respir. Med. 118, 39–45. doi: 10.1016/j.rmed.2016.07.009
Keywords: lung microbiome, 16S ribosomal subunit, antisynthetase syndrome, interstitial lung disease, Veillonella
Citation: Quintero-Puerta T, Lira-Lucio JA, Falfán-Valencia R, Vega-Sánchez ÁE, Márquez-García E, Mejía M, Bautista-Becerril B, Rojas-Serrano J, Ramos-Martínez E, Buendía-Roldán I and Pérez-Rubio G (2023) Lung microbiome alterations in patients with anti-Jo1 antisynthetase syndrome and interstitial lung disease. Front. Cell. Infect. Microbiol. 13:1321315. doi: 10.3389/fcimb.2023.1321315
Received: 13 October 2023; Accepted: 21 November 2023;
Published: 05 December 2023.
Edited by:
Dong Tang, Yangzhou University, ChinaCopyright © 2023 Quintero-Puerta, Lira-Lucio, Falfán-Valencia, Vega-Sánchez, Márquez-García, Mejía, Bautista-Becerril, Rojas-Serrano, Ramos-Martínez, Buendía-Roldán and Pérez-Rubio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gloria Pérez-Rubio, Z3BlcmV6cnViaW9AaW5lci5nb2IubXg=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.