 Qian Chen
Qian Chen Lu Liu
Lu Liu Jing Si
Jing Si Josef Vlasák
Josef Vlasák- 1College of Architecture and Urban Planning, Chongqing Jiaotong University, Chongqing, China
- 2College of Architecture and Urban Planning, Tongji University, Shanghai, China
- 3Institute of Microbiology, School of Ecology and Nature Conservation, Beijing Forestry University, Beijing, China
- 4Biology Centre, Institute of Plant Molecular Biology, Czech Academy of Sciences, Branišovská, České Budějovice, Czechia
Fuscoporia is a cosmopolitan, poroid, wood-decaying genus, belonging to the Hymenochaetales. During a study of wood-inhabiting fungi in the USA, four unknown specimens were collected from Hawaii. Both morphological criteria and molecular genetic analyses based on the ITS+nLSU+EF1-α datasets and the nLSU dataset confirmed that these four specimens represent two new species of Fuscoporia, and they are described as F. hawaiiana and F. minutissima. Fuscoporia hawaiiana is characterized by pileate basidiocarps, the absence of cystidioles, hooked hymenial setae, broadly ellipsoid to subglobose basidiospores measuring 4−6 × 3.5−4.5 μm. Fuscoporia minutissima is distinguished by small pores (10−13 per mm) and basidiospores (3.4−4 × 2.4−3 μm). The taxonomic status of the two new species is briefly discussed. A key to the North American species of Fuscoporia is provided.
1 Introduction
Most wood-rotting fungi, belonging to Basidiomycetes, can use different types of wood as their nutritional source and are indispensable participants and important biological regeneration resources in natural ecosystems. The genus Fuscoporia Murrill (Hymenochaetales, Basidiomycota), erected by Murrill (1907) with F. ferruginosa (Schrad.) as generic type, is a cosmopolitan fungal group that encompasses also important species with medicinal effects, such as F. gilva (Schwein.) T. Wagner & M. Fisch. and F. torulosa (Pers.) T. Wagner & M. Fisch. (Wu et al., 2019). Fuscoporia is widely distributed in Asia, Europe, Oceania, and America. The genus has been considered a synonym of Poria Adans. or Phellinus Quél. for a long time (Overholts, 1953; Lowe, 1966; Ryvarden and Johansen, 1980; Gilbertson and Ryvarden, 1987; Larsen and Cobb-Poulle, 1990). However, Fiasson and Niemela (1984) recognized the genus as monophyletic morphologically, characterized by annual to perennial and resupinate to pileate basidiomata, a dimitic hyphal system with crystal encrustations on generative hyphae, the presence of hymenial setae, and hyaline, thin-walled and smooth basidiospores. Later, Wagner and Fischer, 2001; Wagner and Fischer, 2002) by means of nLSU sequence data and morphological studies of European Hymenochaetales reconfirmed the taxonomic status of the genus of Fuscoporia, with six species of Phellinus being classified into it.
Large number of Fuscoporia new species and new combinations have been discovered in recent years, and at present, more than 90 species are accepted in the genus (Chen et al., 2020; Tchoumi et al., 2020; Vlasák et al, 2020; Yuan et al., 2020; Dai et al., 2021; Chen et al., 2022; Hussain et al., 2022; Wu et al., 2022a, b). Fuscoporia is considered to be a complex genus with still unsettled taxonomy.
Striking diversity in the Hawaiian Islands with multiple co-occurring species leads to many species unreported in the literature. During a study of wood-inhabiting fungi, four unknown specimens were collected from Hawaii, with the features of Fuscoporia. To explore their relationships, phylogenetic analyses based on the ITS+nLSU+EF1-α datasets and the nLSU dataset were carried out. Both morphological criteria and molecular genetic analyses confirmed that these four specimens represent two new species of Fuscoporia. So, we describe them as F. hawaiiana and F. minutissima in the present paper. A key to the North American species of Fuscoporia is also provided.
2 Materials and methods
2.1 Morphological studies
Some studied specimen materials are deposited in the fungoria of the Institute of Microbiology, Beijing Forestry University (BJFC). The other ones are deposited in private fungoria of Josef Vlasak (JV) and then will be transferred into Prague Museum Herbarium (PRM). Morphological descriptions are based on field notes and herbarium specimens. The macroscopic color terms cited from Anonymous (1969) and Petersen (1996). Sections were studied at a magnification up to × 1,000 using a Nikon Eclipse 80i microscope with phase contrast illumination (Nikon, Tokyo, Japan). The microscopic analyses were performed accroding to Liu et al. (2022) and Si et al. (2023). Microscopic features, measurements, and drawings were prepared from slides stained with Cotton Blue. The following abbreviations are used: KOH = 5% potassium hydroxide, CB− = acyanophilous in Cotton Blue, IKI− = neither amyloid nor dextrinoid in Melzer’s reagent, L = arithmetic average of all spore length, W = arithmetic average of all spore width, Q = variation in the L/W ratios, and (n = x/y) = number of measured spores (x) measured from a given number of specimens (y).
2.2 DNA extraction, PCR, and sequencing
A CTAB rapid plant genome extraction kit (Aidlab Biotechnologies Co., Ltd., Beijing, China) was used to extract total genomic DNA from dried specimens following the manufacturer’s instructions with some modifications (Chen et al., 2020; Zhang et al., 2023; Zhou et al., 2023). For generation of PCR amplicons, the following primer pairs were used: ITS5 (GGA AGT AAA AGT CGT AAC AAG G) and ITS4 (TCC TCC GCT TAT TGATAT GC) for internal transcribed spacer (ITS, White et al., 1990); LR0R (ACC CGC TGA ACT TAA GC) and LR7 (TAC TAC CAC CAA GAT CT) for nuclear large subunit rDNA (nLSU, Vilgalys and Hester, 1990); EF1-983F (GCY CCY GGH CAY CGT GAY TTY AT) and EF1-1567R (ACH GTR CCR ATA CCA CCR ATC TT) for translation elongation factor-1 alpha (EF1-α, Rehner and Buckley, 2005). The PCR procedures for ITS and EF1-α were as follows: initial denaturation at 95°C for 3 min, followed by 35 cycles of denaturation at 94°C for 40 s, annealing at 54°C for 45 s and extension at 72°C for 1 min, and a final extension at 72°C for 10 min. The PCR procedure for nLSU was as follows: initial denaturation at 94°C for 1 min, followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 50°C for 1 min and extension at 72°C for 1.5 min, and a final extension at 72°C for 10 min. The PCR products were purified and sequenced at the Beijing Genomics Institute with the same primers and the sequences are deposited in GenBank. All newly generated sequences were deposited in GenBank (http://www.ncbi.nlm.nih.gov ) and are listed in Table 1.
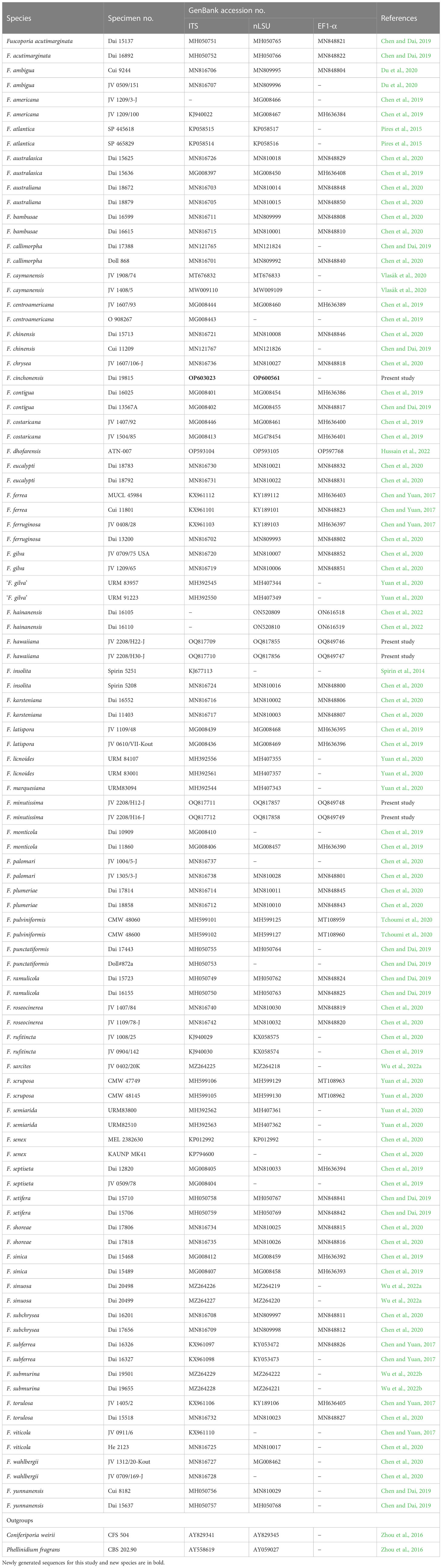
Table 1 Species, specimens, and GenBank accession numbers of sequences used in ITS+nLSU+EF1-α phylogenetic analyses.
2.3 Phylogenetic analysis
The following softwares were used for data processing and phylogenetic analyses: Mesquite, MAFFT 7.110, BioEdit 7.0.1 (Hall, 1999), ClustalX 2.0P10 (Thompson et al., 1997), MrModeltest 2.3 (Posada and Crandall, 1998; Nylander, 2004), PAUP* 4.0b10 (MP, Swofford, 2002), raxmlGUI 1.2 (ML, Stamatakis, 2006; Silvestro and Michalak, 2012), TreeView 1.5.0, and PowerPoint. Maximum parsimony (MP) and maximum likelihood (ML) methods were adopted to perform phylogenetic analyses of the two aligned datasets. The two phylogenetic methods resulted in similar topology for each data set. Thus, only the topology of the MP analysis appears along and branches that received bootstrap supports greater than or equal to 75% (MP and ML) were considered as significantly supported at the nodes.
To explore the phylogenetic position of Fuscoporia in Hymenochaetaceae, representatives of 28 genera of Hymenochaetaceae were included in nLSU dataset (not shown in Table 1 except for Fuscoporia; shown in Figure 1). Oxyporus populinus (Schumach.) Donk and Hyphodontia pallidula (Bres.) J. Erikss. were used as outgroups (Larsson et al., 2006; Zmitrovich and Malysheva, 2014; Zhou et al., 2016; Chen et al., 2019).
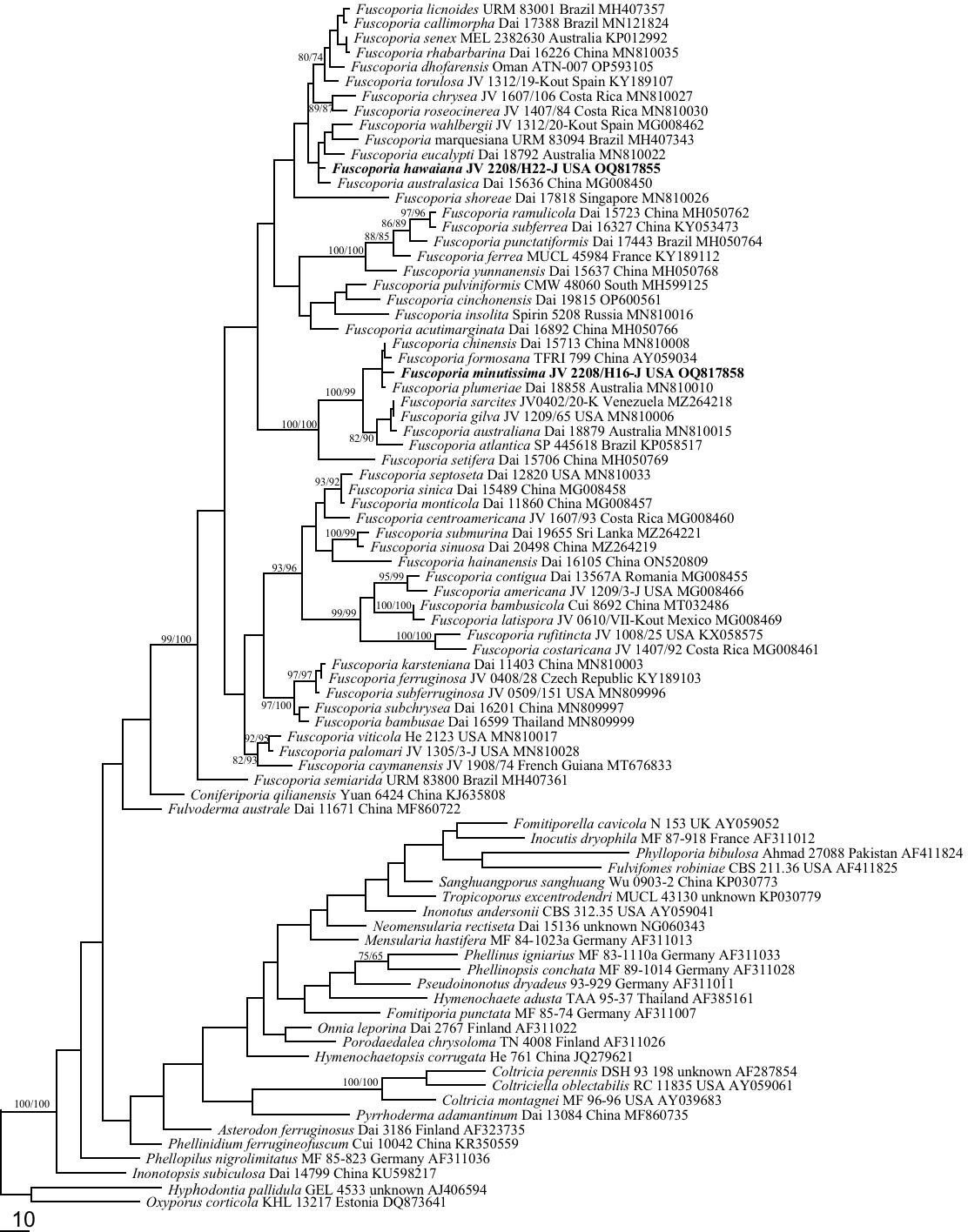
Figure 1 Phylogeny of Fuscoporia and related species generated inferred from nLSU sequences. Statistical values (MP/ML) are indicated for each node. Names of new species are in bold.
In addition to the newly generated sequences, additional ITS, nLSU, and EF1-α sequences of Fuscoporia based on formerly studies (Chen et al., 2019; Chen et al., 2020; Du et al., 2020; Wu et al., 2022a) were obtained from GenBank (Table 1) to explore the phylogenetic position of our specimens in Fuscoporia. A total of 251 sequences, 100 samples of Fuscoporia were analyzed in the ITS+nLSU+EF1-α dataset (Figure 2). Coniferiporia weirii (Murrill) L.W. Zhou et al (2016) and Phellinidium fragrans (M.J. Larsen & Lombard) Nuss (1986) were selected as outgroups (Zhou et al., 2016; Chen and Yuan, 2017).
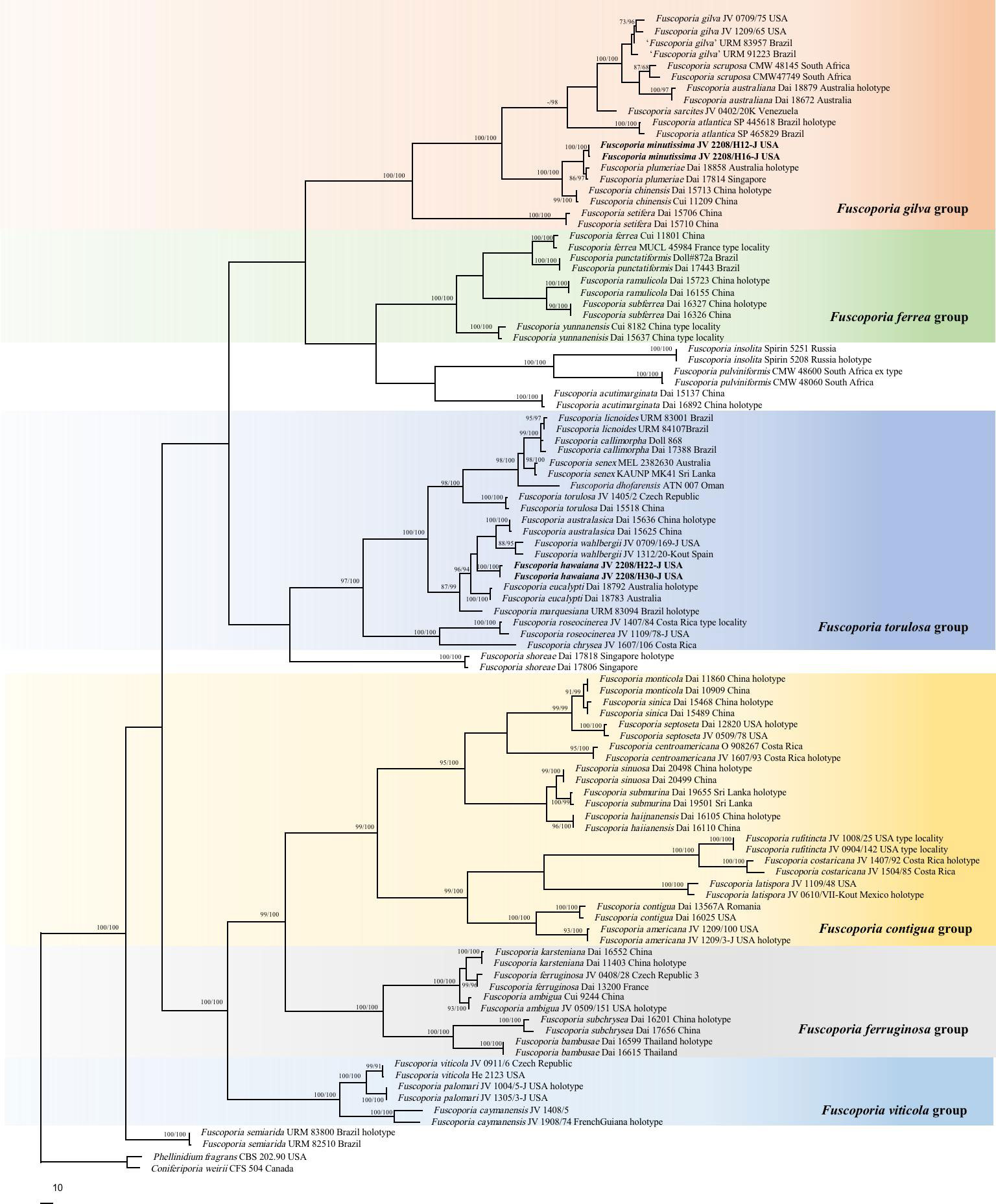
Figure 2 Phylogeny of Fuscoporia and related species generated inferred from ITS+nLSU+EF1-α dataset. Statistical values (MP/ML) are indicated for each node. Names of new species are in bold.
3 Results
3.1 Phylogeny
To explore the phylogenetic position of Fuscoporia in Hymenochaetaceae, 81 taxa of the family Hymenochaetaceae were included in nLSU-alone dataset (Figure 1). The dataset had an aligned length of 1436 characters, of which 1019 were constant, 98 variable but parsimony-uninformative, and 291 parsimony-informative. Maximum parsimony analysis yielded 140 equally topologies (TL = 1723, CI = 0.327, RI = 0.672, RC = 0.220, HI = 0.673). Fuscoporia is a powerfully supported lineage (100/100) within the Hymenochaetaceae family based on phylogenetic tree inferred from the nLSU dataset (Figure 1).
The ITS+nLSU+EF1-α dataset (Figure 2) included 98 ITS, 92 nLSU, and 62 EF1-α sequences from 100 fungal specimens representing 52 pecies of Fuscoporia. The dataset had an aligned length of 2705 characters, of which 1721 were constant, 99 variable but parsimony-uninformative, and 885 parsimony-informative. Maximum parsimony analysis yielded six equally topologies (TL = 4489, CI = 0.389, RI = 0.807, RC = 0.314, HI = 0.611). Two well-supported lineages (100/100) clustered within Fuscoporia and distincted from other species in phylogeny (Figure 2). Taking morphological characters into consideration, these two new lineages represent two new species, F. hawaiiana and F. minutissima, which are described below.
3.2 Taxonomy
Fuscoporia hawaiiana Q. Chen, Jing Si & Vlasák, sp. nov., Figures 3, 4

Figure 3 Basidiocarps of Fuscoporia hawaiiana (holotype, JV 2208/H22-J). Bars: (A) = 1 cm. (B) = 1 mm.
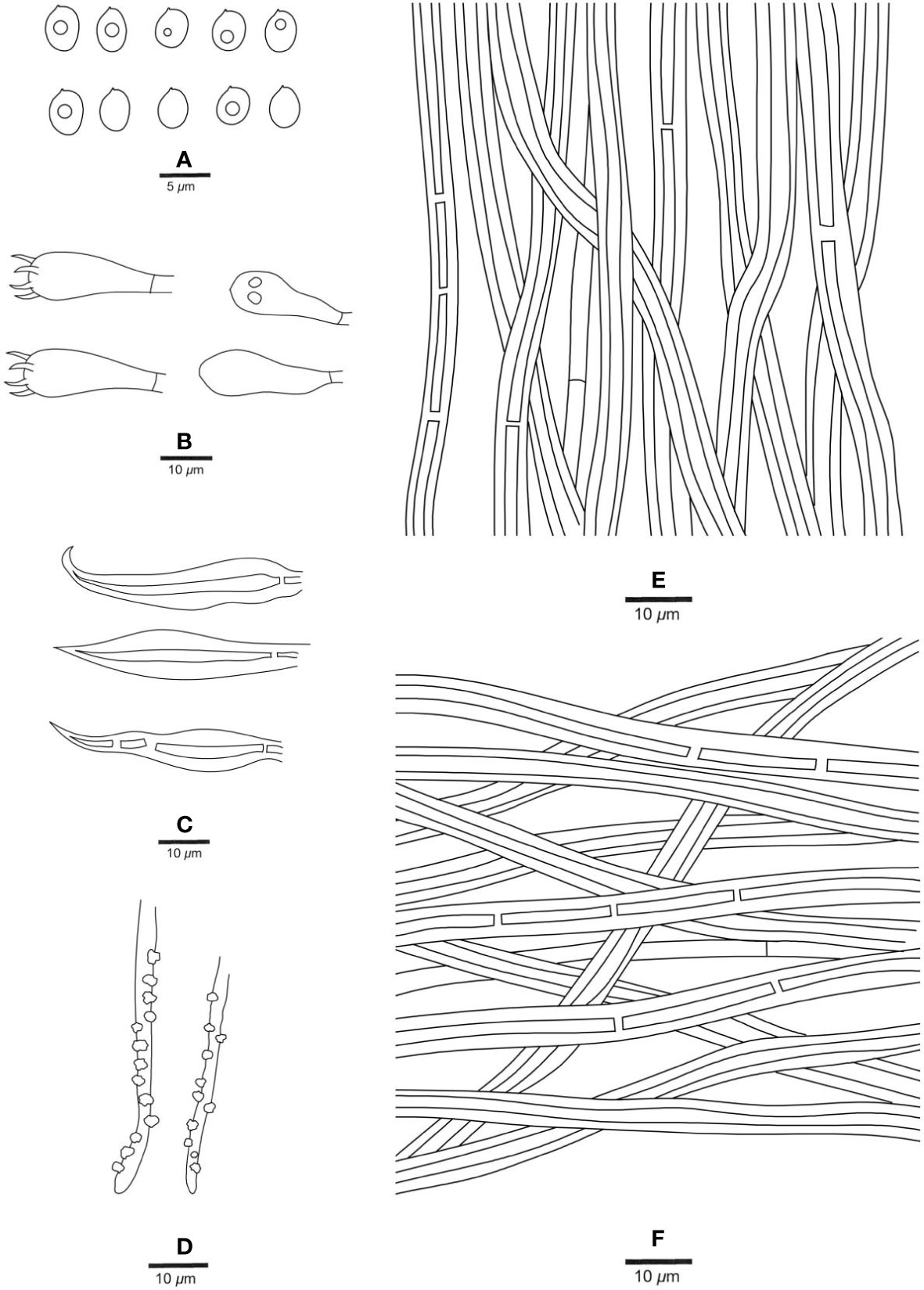
Figure 4 Microscopic structures of Fuscoporia hawaiiana (holotype, JV 2208/H22-J). (A) Basidiospores. (B) Basidia and basidioles. (C) Hymenial setae. (D) Generative hyphae at dissepiment edge. (E) Hyphae from tube trama. (F) Hyphae from subiculum.
MycoBank no. — MB 847960
Holotype. — USA. Hawaii, Big Island, Kalopa State Park, on living tree of Eucalyptus, August 2022, JV 2208/H22-J (PRM, isotype JV, BJFC 039915).
Etymology. — Hawaiiana (Lat.): refers to the place (Hawaii) where the species was collected.
Fruiting body. — Basidiocarps perennial, pileate, laterally fused to imbricate, without odor or taste when fresh, hard corky when dry. Pilei mostly semicircular, projecting up to 11 cm long, 5 cm wide, and 5 cm thick at the base, more or less convex towards margin. Pileal surface reddish brown, concentrically sulcate with zones, glabrous, sometimes covered with mosses; margin obtuse, yellowish brown, up to 5 mm wide. Pore surface honey-yellow to deep olive, slight glancing; margin narrow, olivaceous buff, up to 1 mm wide; pores circular, 5−7 per mm; dissepiments thin to fairly thick, entire, abundant hymenial setae in tube cavities (under anatomical lens). Context clay-buff, hard corky, about 8 mm thick. Tubes olivaceous buff, hard corky, up to 2 cm long.
Hyphal structure. — Hyphal system dimitic; generative hyphae simple septate; tissue darkening but otherwise unchanged in KOH.
Context. — Generative hyphae rare, hyaline, thin- to slightly thick-walled, branched, frequently simple septate, 2−2.5 μm in diam; skeletal hyphae dominant, rust-brown, thick-walled with a medium to wide lumen, unbranched, occasionally septate, straight, more or less straight and regularly arranged, 2.5−3.5 μm in diam.
Tubes. — Generative hyphae rare, mostly present at dissepiment edges and subhymenium, hyaline, thin-walled, frequently branched and simple septate, 1.5−2.5 μm in diam, some of them encrusted at dissepiment edges and in the hymenium; skeletal hyphae dominant, yellowish brown, thick-walled with a medium to wide lumen, frequently septate, more or less straight, subparallel along the tubes, 2.5−3.5 μm in diam. Hymenial setae subulate, occasionally hooked, mostly originating from tramal hyphae, dark brown, thick-walled, 30−45 × 4−7 μm; basidia broadly clavate, with four sterigmata and a simple septum at the base, 12−18 × 5−7 μm; basidioles dominating the hymenium, in shape similar to basidia, but slightly smaller.
Spores. — Basidiospores broadly ellipsoid, hyaline, thin-walled, smooth, IKI−, CB−, some of them bearing a guttule, 4−6 × (3.4−)3.5−4.5(−4.7) μm, L = 4.77 μm, W = 3.90 μm, Q = 1.14−1.31 (n = 60/2).
Other material examined (paratype). — USA. Hawaii, Big Island, Volcano, on dead tree of Metrosideros polymorpha, August 2022, JV 2208/H30-J (JV, BJFC 039918).
Fuscoporia minutissima Q. Chen, Jing Si & Vlasák, sp. nov., Figures 5, 6 MycoBank no. — MB 847964

Figure 5 Basidiocarps of Fuscoporia minutissima. (A–C)JV 2208/H16-J (holotype). (D) JV 2208/H12-J. Bars: (A, B, D) = 1 cm. (C) = 1 mm.
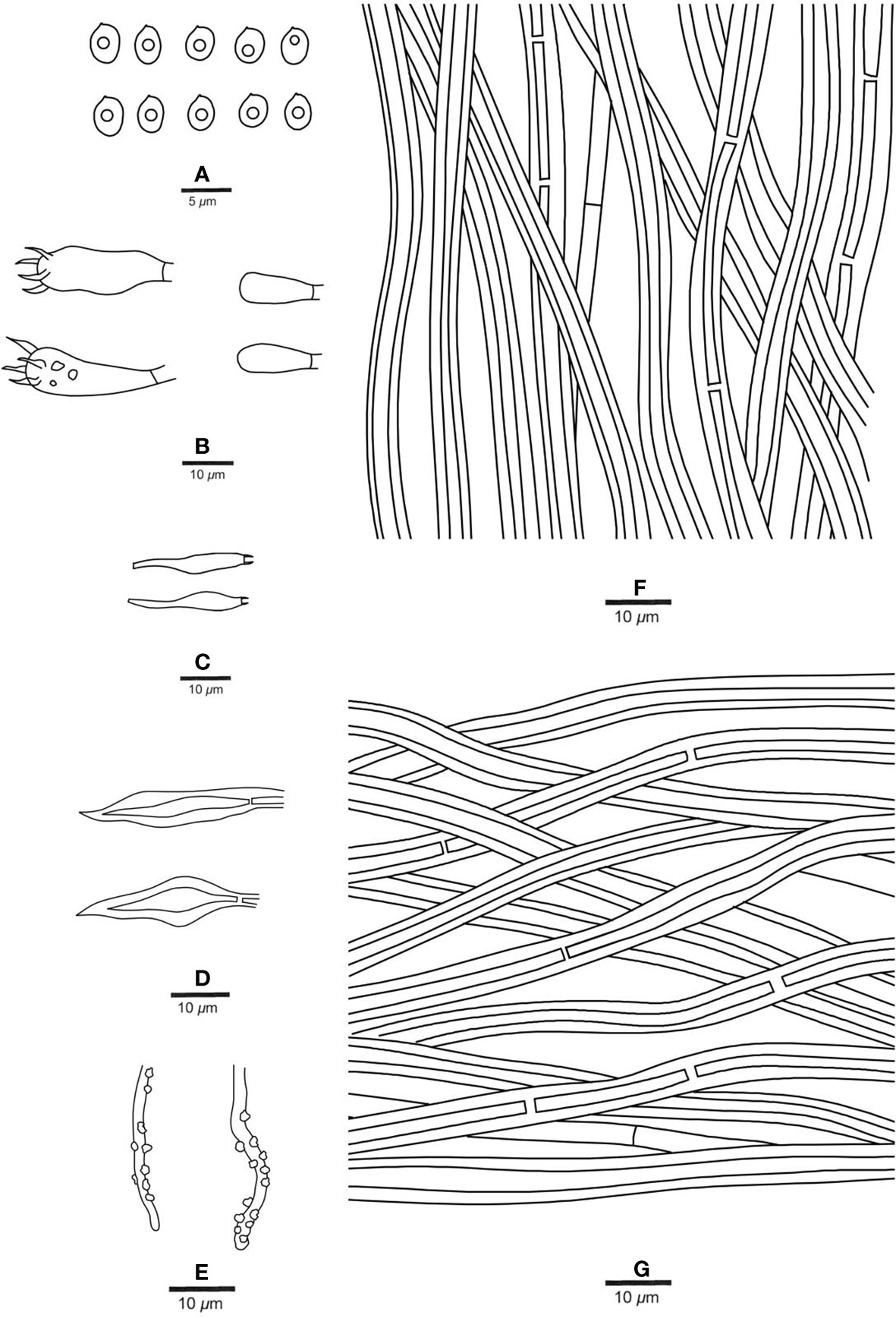
Figure 6 Microscopic structures of Fuscoporia minutissima (holotype, JV 2208/H16-J). (A) Basidiospores. (B) Basidia and basidioles. (C) Cystidioles. (D) Hymenial setae. (E) Generative hyphae at dissepiment edge. (F) Hyphae from tube trama. (G) Hyphae from subiculum.
Holotype. — USA. Hawaii, Big Island, Makuala O’oma Trail, August 2022, JV 2208/H16-J (PRM, isotype JV, BJFC 039911).
Etymology. — Minutissima (Lat.): refers to the very small size of the pores exhibiting by these species.
Fruiting body. — Basidiocarps perennial, pileate, imbricate, without odor or taste when fresh, hard corky when dry. Pilei mostly imbricate, projecting up to 5 cm long, 3 cm wide, and 1 cm thick at the base. Pileal surface reddish brown, concentrically sulcate with zones, nodulose; margin obtuse to slightly acute, honey-yellow, up to 1 mm wide. Pore surface greyish brown to deep olive, glancing; margin narrow, honey-yellow, up to 1 mm wide; pores circular, 10−13 per mm; dissepiments fairly thick, entire and matted, abundant hymenial setae in tube cavities (under anatomical lens). Context honey-yellow, hard corky, up to 5 mm thick. Tubes olivaceous buff, paler than pores, hard corky, up to 4 mm long.
Hyphal structure. — Hyphal system dimitic; generative hyphae simple septate; tissue darkening but otherwise unchanged in KOH.
Context. — Generative hyphae rare, hyaline, thin- to slightly thick-walled, unbranched, frequently simple septate, 2−3 μm in diam; skeletal hyphae dominant, rust-brown, thick-walled with a medium to narrow lumen, unbranched, occasionally septate, straight, regularly arranged, 3−4 μm in diam.
Tubes. — Generative hyphae rare, mostly present at dissepiment edges and subhymenium, hyaline, thin-walled, frequently branched and simple septate, 2−3 μm in diam, some of them encrusted at dissepiment edges and in the hymenium; skeletal hyphae dominant, yellowish brown, thick-walled with a medium to narrow lumen, frequently septate, more or less straight, subparallel along the tubes, 3−4 μm in diam. Hymenial setae subulate, mostly originating from tramal hyphae, dark brown, thick-walled, 18−40 × 6−9 μm; fusoid cystidioles hyaline and thin-walled, 9.5−12 × 4−5.5 μm; basidia short clavate to barrel-shaped, with four sterigmata and a simple septum at the base, 10−15 × 4−6.5 μm; basidioles dominating the hymenium, in shape similar to basidia, but slightly smaller.
Spores. — Basidiospores broadly ellipsoid to subglobose, hyaline, thin-walled, smooth, IKI−, CB−, bearing a guttule, (3−)3.4−4 × (2.2−)2.4−3(−3.8) μm, L = 3.60 μm, W = 2.79 μm, Q = 1.24−1.30 (n = 52/2).
Other material examined (paratype). — USA. Hawaii, Kauai Island, Koke’e State Park, on dead tree of Acacia koa, August 2022, JV 2208/H12-J (JV, BJFC 039910).
4 Discussion
The islands far from the mainland have attracted scientific researchers for a long time, due to the close combination of ecological environment and evolution process here that deepen our understanding of the formation process of biological diversity (Cotoras et al., 2018). Hawaii is a biological hotspot with a variety of climates and habitats. 160 genera and 400 species of wood-rotting basidiomycetes are reported from 110 native and exotic substrate species on the Hawaiian Islands (Gilbertson et al., 2002; Ashiglar et al., 2015). There are few natural forests on the Hawaii islands (Chambers et al., 2007). Most forests are filled with introduced trees, and our new species may be introduced as well. The two new wood-rotting fungal species Fuscoporia hawaiiana and F. minutissima were collected from the northernmost and oldest Kauai Island, and the largest island Big Island in the Hawaiian Islands, both with a tropical sea climate.
Fuscoporia hawaiiana is characterized by perennial and pileate basidiocarps, circular and medium pores (5−7 per mm), the absence of cystidioles, hooked hymenial setae, broadly ellipsoid to subglobose basidiospores measuring 4−6 × 3.5−4.5 μm. Fuscoporia hawaiiana may also be distributed in Yunnan Province, China, considered to be F. torulosa derived from CLZhao 10146 (OM959398) on GenBank. The ITS of CLZhao 10146 shows that only 2 base differences from our new samples, but unfortunately, it lacks nLSU or EF1-α sequence. Furthermore, F. torulosa can be easily distinguished from F. hawaiiana by its subungulate basidiocarps and straight hymenial setae. Morphologically, seven species, F. semihispida (Ryvarden) Y.C. Dai & F. Wu, F. australasica Q. Chen, F. Wu & Y.C. Dai, F. marquesiana Gibertoni & C.R.S. de Lira, F. atlantica Motato-Vásq., R.M. Pires & Gugliotta, F. wahlbergii (Fr.) T. Wagner & M. Fisch., F. eucalypti Q. Chen, F. Wu & Y.C. Dai, and F. rufa (Bres.) Y.C. Dai & F. Wu., are similar to F. hawaiiana by sharing similar pileate basidiocarps and hooked hymenial setae. However, F. hawaiiana is distant from F. atlantica, F. marquesiana, F. wahlbergii, F. eucalypti, and F. australasica both in the phylogenetic analyses (Figures 1, 2) and morphology. F. atlantica differs from F. hawaiiana by its annual basidiocarps (Pires et al., 2015); F. australasica is described from Southern China and Viet Nam and differs from F. hawaiiana by its glabrous basidiocarps and the presence of cystidioles (Chen et al., 2020); F. wahlbergii is described from Europe, East-African, Australia, and USA, and distinguishes from F. hawaiiana by its smaller pores (7−9 per mm) and the presence of cystidioles (Chen et al., 2020); F. eucalypti is distributed Australia, and differs from F. hawaiiana by its spores effused-reflexed to pileate and wider (4.3−5.5 × 4−4.5 μm) (Chen et al., 2020); F. marquesiana is a South American species, and can be easily distinguished from F. hawaiiana by its smaller pores (8−9 per mm) (Wu et al., 2022b). Even without molecular data, F. semihispida and F. rufa can be easily distinguished from F. hawaiiana by narrower or wider spores (2−2.5 μm in F. semihispida; 4−4.5 μm in F. rufa) (Wu et al., 2022a).
Fuscoporia minutissima stands out in the genus by its perennial and pileate basidiocarps, and very small pores (10−13 per mm) and spores (3.4−4 × 2.4−3 μm). Fuscoporia minutissima is usually sterile after collecting, and such specimens are very similar to F. gilva by pileate basidiocarps, glabrous to rugose sometimes with nodulose pileal surface and broadly ellipsoid to subglobose spores, but the later differs by having bigger pores (6−8 per mm). Fuscoporia plumeriae Q. Chen, F. Wu & Y.C. Dai and F. chinensis Q. Chen, F. Wu & Y.C. Dai clustered together with F. minutissima into a group with a strong support (100/100, Figures 1, 2). Morphologically, these species share the effused-reflexed to pileate basidiocarps with lacerate dissepiments and ellipsoid basidiospores; but F. plumeriae and F. chinensis have annual basidiocarps and bigger pores (10−13 per mm in F. minutissima; 8−10 per mm in F. plumeriae; 7−8 per mm in F. chinensis; Chen et al., 2020). Fuscoporia rhabarbarina (Berk.) Groposo, Log.-Leite & Góes-Neto is a common fungus on hardwoods along streams in South America and subtropical and tropical Asia. But the type locality of this species remains unknown, probably in Brazil (Wu et al., 2022b). Fuscoporia minutissima may be confused with F. rhabarbarina in field, but the pileal surface of F. rhabarbarina is glabrous and dark brown crust at the base. Moreover, the two species are not closely related in the phylogenetic tree (Figure 1).
Fuscoporia hawaiiana and F. minutissima are described in this study. Traditionally, they were most probably treated as F. wahlbergii and F. gilva, respectively. The phylogenetic analyses provide molecular evidence to support these two new species derive from the F. wahlbergii complex and the F. gilva complex (Figures 1, 2). The similar results are demonstrated in many polypores complex, for instances, Megasporoporia setulosa (Henn.) Rajchenb. (Wang et al., 2021; Wang et al., 2022), Heterobasidion annosum (Fr.) Bref. (Yuan et al., 2021), Porogramme epimiltina (Berk. & Broome) Y.C. Dai (Mao et al., 2023), Sidera vulgaris (Fr.) Miettinen (Liu et al., 2021), and Phaeolus schweinitzii (Fr.) Pat. (Yuan et al., 2022). It seems that the diversity of wood-inhabiting fungi is extremely rich in tropics, some traditional definition on tropical species should be re-evaluated, and the concepts of taxa in tropics should be modified after molecular phylogeny.
A key to North American species of Fuscoporia.
1. Basidiocarps completely resupinate...........................................1
- Basidiocarps pileate, effused-reflexed or substipitate................7
2. Mycelial setae absent .........................F. ferrea (Pers.) G. Cunn.
- Mycelial setae present.....................................................................3
3. Pores 5−9 per mm..........................................................................4
- Pores 2−4 per mm...........................................................................5
4. Pores 7−9 per mm, skeletal hyphae septate ................ ............................................................................................................ ......F. rufitincta (Berk. & M.A. Curtis ex A.L. Sm.) Murrill
- Pores 5−6 per mm, skeletal hyphae aseptate F. ambigua P. Du, Q. Chen & J. Vlasák
5. Basidiocarps perennial ..........................F. contigua (Pers.) G. Cunn.
- Basidiocarps annual.................................................................................6
6. Spores broadly ellipsoid, 4.8−6.0 × 3.2−4.2 μm ....................................F. americana Y.C. Dai, Q. Chen & J. Vlasák
- Spores cylindric, 6.0−7.0 × 2.0−3.0 μm ............................F. septiseta Y.C. Dai, Q. Chen & J. Vlasák
7. Hymenial setae hooked ........................................................................8
- Hymenial setae straight...........................................................................9
8. Pores 7−9 per mm, cystidioles present..........................F. wahlbergii
- Pores 5−7 per mm, cystidioles absent.............................F. hawaiiana
9. Basidiocarps annual.........................F. palomari Vlasák & Ryvarden
- Basidiocarps perennial..........................................................................10
10. Spores cylindric, 7−9 × 1.5−2 μm, Q > 2...........................F. viticola (Schwein.) Murrill
- Spores broadly ellipsoid to ellipsoid, Q < 2.......................................11
11. Pileal surface velutinate to glabrous.......................F. rhabarbarina
- Pileal surface tomentose, hispid, rugose............................................12
12. Pores 10−13 per mm...................................................F. minutissima
- Pores 6−9 per mm..................................................................................13
13. Pileal surface not sulcate...........................................................F. gilva
- Pileal surface concentrically sulcate....................................................14
14. Basidiocarps subungulate, pileal surface grayish brown.....................................................................................F. torulosa
- Basidiocarps usually applanate, pileal surface fuscous to black........................................F. senex (Nees & Mont.) Ghob.-Nejh
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: F. hawaiiana JV 2208/H22-J: OQ817709; OQ817855; OQ849746. F. hawaiiana JV 2208/H30-J: OQ817710; OQ817856; OQ849747. F. minutissima JV 2208/H12-J: OQ817711; OQ817857; OQ849748. F. minutissima JV 2208/H16-J: OQ817712; OQ817858; OQ849749. F. cinchonensis OP603023; OP600561.
Author contributions
QC, LL, JS, and JV designed the research and contributed to data analysis and interpretation. JV prepared the samples. QC and LL conducted the molecular experiments and analyzed the data. QC prepared the samples and drafted the manuscript. JV and JS discussed the results and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was financed by the National Natural Science Foundation of China (Nos. 32100014 and 32070016), the Science and Technology Research Program of Chongqing Municipal Education Commission (No. KJQN202100737), and the Academy of Sciences of the Czech Republic RVO (No. 60077344).
Acknowledgments
The authors would like to express their deep appreciations to Prof. Yu-Cheng Dai (Beijing Forestry University, China) for forwarding specimens on loan and Dr. Josef Vlasák Jr. for collecting fresh specimens on Hawaii islands.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Anonymous (1969). Flora of British fungi. colour identification chart (Edinburgh, UK: Her Majesty’s Stationery Office).
Ashiglar, S. M., Brooks, F., Cannon, P. G., Klopfenstein, N. B. (2015). “Preliminary survey of wood-associated fungi in southeast o’ahu of HawaiI using DNA-based identification,” in Proceedings of the 62nd annual Western international forest disease work conference. Eds. Murray, M., Palacios, P. (Cedar City, Utah, US), 67–69.
Chambers, J. Q., Asner, G. P., Morton, D. C., Anderson, L. O., Saatchi, S. S., Espírito-Santo, F. D. B., et al. (2007). Regional ecosystem structure and function: ecological insights from remote sensing of tropical forests. Trends Ecol. Evol. 22, 414–423. doi: 10.1016/j.tree.2007.05.001
Chen, Q., Dai, Y. C. (2019). Two new species of Fuscoporia (Hymenochaetales, basidiomycota) from southern China based on morphological characters and molecular evidence. MycoKeys 61, 75–89. doi: 10.3897/mycokeys.61.46799
Chen, Q., Du, P., Vlasak, J., Wu, F., Dai, Y. C. (2020). Global diversity and phylogeny of Fuscoporia (Hymenochaetales, basidiomycota). Mycosphere 11, 1477–1513. doi: 10.5943/mycosphere/11/1/10
Chen, Q., Liu, L., Zhang, D. S., Dong, L. L. (2022). Fuscoporia hainanensis sp. nov. (Hymenochaetales, basidiomycota), a new member of the F. contigua group. Phytotaxa 558, 251–262. doi: 10.11646/phytotaxa.558.3.1
Chen, Q., Wu, F., Ji, X. H., Si, J., Zhou, L. W., Tian, X. M., et al. (2019). Phylogeny of the genus Fuscoporia and taxonomic assessment of the F. contigua group. Mycologia 111, 423–444. doi: 10.1080/00275514.2019.1570749
Chen, Q., Yuan, Y. (2017). A new species of Fuscoporia (Hymenochaetales, basidiomycota) from southern China. Mycosphere 8, 1238–1245. doi: 10.5943/mycosphere/8/6/9
Cotoras, D. D., Bi, K., Brewer, M. S., Lindberg, D. R., Prost, S., Gillespie, R. G. (2018). Co-Occurrence of ecologically similar species of Hawaiian spiders reveals critical early phase of adaptive radiation. BMC Evol. Biol. 18, 100. doi: 10.1186/s12862-018-1209-y
Dai, Y. C., Yang, Z. L., Cui, B. K., Wu, G., Yuan, H. S., Zhou, L. W., et al. (2021). Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 40, 770–805. doi: 10.13346/j.mycosystema.210036
Du, P., Chen, Q., Vlasák, J. (2020). Fuscoporia ambigua sp. nov., a new species from America and China. Phytotaxa 456, 175–185. doi: 10.11646/phytotaxa.456.2.5
Fiasson, J. L., Niemela, T. (1984). The hymenochaetales: a revision of the European poroid taxa. Karstenia 24, 14–28. doi: 10.29203/ka.1984.224
Gilbertson, R. L., Bigelow, D. M., Hemmes, D. E., Desjardin, D. E. (2002). Annotated check list of wood-rotting basidiomycetes of Hawaii. Mycotaxon 82, 215–239.
Hall, T. A. (1999). Bioedit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 41, 95–98. doi: 10.1021/bk-1999-0734.ch008
Hussain, S., Al-Kharousi, M., Al-Muharabi, M. A., Al-Maqbali, D., Al-Shabibi, Z., Al-Balushi, A. H., et al. (2022). Phylogeny, distribution and time divergence of Fuscoporia (Hymenochaetaceae, basidiomycota) with the description of a new species from dhofar region, southern part of Oman. Phytotaxa 570, 150–164. doi: 10.11646/phytotaxa.570.2.3
Larsen, M. J., Cobb-Poulle, L. A. (1990). Phellinus (Hymenochaetaceae). a survey of the world taxa. Synop. Fungorum. 3, 1–206. doi: 10.2307/3760175
Larsson, K. H., Parmasto, E., Fischer, M., Langer, E., Nakasone, K. K., Redhead, S. A. (2006). Hymenochaetales: a molecular phylogeny for the hymenochaetoid clade. Mycologia 98, 926–936. doi: 10.1080/15572536.2006.11832622
Liu, Z. B., Wu, Y. D., Zhao, H., Lian, Y. P., Wang, Y. R., Wang, C. G., et al. (2022). Outline, divergence times, and phylogenetic analyses of Trechisporales (Agaricomycetes, basidiomycota). Front. Microbiol. 13. doi: 10.3389/fmicb.2022.818358
Liu, Z. B., Zhou, M., Yuan, Y., Dai, Y. C. (2021). Global diversity and taxonomy of Sidera (Hymenochaetales, basidiomycota): four new species and keys to species of the genus. J. Fungi 7, 251. doi: 10.3390/jof7040251
Lowe, J. L. (1966). Polyporaceae of north america. the genus poria Vol. 90 (New York, US: Technical Publication of the State University College of Forestry at Syracuse University), 1–183.
Mao, W. L., Wu, Y. D., Liu, H. G., Yuan, Y., Dai, Y. C. (2023). A contribution to Porogramme (Polyporaceae, agaricomycetes) and related genera. IMA Fungus 14, 5. doi: 10.1186/s43008-023-00110-z
Nylander, J. A. A. (2004). MrModeltest v2. uppsala: program distributed by the author (Sweden: Evolutionary Biology Centre, Uppsala University).
Overholts, L. D. (1953). The polyporaceae of the united states, Alaska and Canada (Ann Arbor: University of Michigan Press). doi: 10.2307/2481836
Petersen, J. H. (1996). The Danish mycological society’s colour-chart (Greve: Foreningen til Svampekundskabens Fremme), 1–6.
Pires, R. M., Motato-Vásquez, V., de Gugliotta, A. M. (2015). Fuscoporia atlantica sp. nov., a new polypore from the Brazilian Atlantic rainforest. Mycotaxon 130, 843–855. doi: 10.5248/130.843
Posada, D., Crandall, K. A. (1998). Modeltest: testing the model of DNA substitution. Bioinformatics 14, 817–818. doi: 10.1093/bioinformatics/14.9.817
Rehner, S. A., Buckley, E. (2005). A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 97, 84–98. doi: 10.1080/15572536.2006.11832842
Ryvarden, L., Johansen, I. (1980). A preliminary polypore flora of East Africa (Oslo: Fungiflora). doi: 10.2307/3759822
Si, J., Zhang, Y. Z., Liang, J. Q., Li, H. J. (2023). Morphology and phylogeny identify two new species and one new subspecies of Podoscypha from yunnan province, southwest China. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1151365
Silvestro, D., Michalak, I. (2012). raxmlGUI: a graphical front-end for rAxML. Org. Divers. Evol. 12, 335–337. doi: 10.1007/s13127-011-0056-0
Spirin, V., Vlasák, J., Niemelä, T. (2014). Fuscoporia insolita (Hymenochaetales, basidiomycota), a new species from Russian far East. Ann. Bot. Fenn. 51, 403–406. doi: 10.5735/085.051.0607
Stamatakis, A. (2006). rAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690. doi: 10.1093/bioinformatics/btl446
Swofford, D. L. (2002). PAUP*: phylogenetic analysis using parsimony (*and other methods). version 4.0b10 (Sunderland, MA: Sinauer Associates). doi: 10.1002/0471650129.dob0522
Tchoumi, J. M. T., Coetzee, M. P. A., Rajchenberg, M., Roux, J. (2020). Poroid hymenochaetaceae associated with trees showing wood-rot symptoms in the garden route national park of south Africa. Mycologia 112, 722–741. doi: 10.1080/00275514.2020.1753160
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., Higgins, D. G. (1997). The Clustal_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25, 4876–4882. doi: 10.1093/nar/25.24.4876
Vilgalys, R., Hester, M. (1990). Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 172, 4238–4246. doi: 10.1128/jb.172.8.4238-4246.1990
Vlasák, J., Kout, J., Chen, Q., Dai, Y. C. (2020). Fuscoporia caymanensis sp. nov. (Basidiomycota, hymenochaetaceae), a new species from tropical America. Phytotaxa 472, 135–146. doi: 10.11646/phytotaxa.472.2.4
Wagner, T., Fischer, M. (2001). Natural groups and a revised system for the European poroid hymenochaetales (Basidiomycota) supported by nLSU rDNA sequence data. Mycol. Res. 105, 773–782. doi: 10.1017/S0953756201004257
Wagner, T., Fischer, M. (2002). Proceedings towards a natural classification of the worldwide taxa Phellinus s.l. and Inonotus s.l., and phylogenetic relationships of allied genera. Mycologia 94, 998–1016. doi: 10.1080/15572536.2003.11833156
Wang, Y. R., Dai, Y. C., Liu, H. G., Vlasák, J., Buchanan, P., Yuan, Y., et al. (2022). A new contribution to Megasporoporia sensu lato: six new species and three new combinations. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.1046777
Wang, Y. R., Wu, Y. D., Vlasak, J., Yuan, Y., Dai, Y. C. (2021). Phylogenetic analysis demonstrating four new species in Megasporoporia sensu lato (Polyporales, basidiomycota). Mycosphere 12, 1012–1037. doi: 10.5943/mycosphere/12/1/11
White, T. J., Bruns, T. D., Lee, S., Taylor, J. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR protocols: a guide to methods and applications. Eds. Innis, M. A., Gelfand, D. H., Sninsky, J. J., White, T. J. (US: New York Academic Press), 315–322. doi: 10.1016/B978-0-12-372180-8.50042-1
Wu, F., Man, X. W., Tohtirjap, A., Dai, Y. C. (2022a). A comparison of polypore funga and species composition in forest ecosystems of China, north America, and Europe. For. Ecosyst. 9, 100051. doi: 10.1016/j.fecs.2022.100051
Wu, F., Zhou, L. W., Vlasák, J., Dai, Y. C. (2022b). Global diversity and systematics of hymenochaetaceae with poroid hymenophore. Fungal Divers. 113, 1–192. doi: 10.1007/s13225-021-00496-4
Wu, F., Zhou, L. W., Yang, Z. L., Bau, T., Li, T. H., Dai, Y. C. (2019). Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species. Fungal Divers. 98, 1–76. doi: 10.1007/s13225-019-00432-7
Yuan, Y., Chen, J. J., Korhonen, K., Martin, F., Dai, Y. C. (2021). An updated global species diversity and phylogeny in the forest pathogenic genus Heterobasidion (Basidiomycota, russulales). Front. Microbiol. 11. doi: 10.3389/fmicb.2020.596393
Yuan, H. S., Lu, X., Dai, Y. C., Hyde, K. D., Kan, Y. H., Kušan, I., et al. (2020). Fungal diversity notes 1277–1386: taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 104, 1–266. doi: 10.1007/s13225-020-00461-7
Yuan, Y., Wu, Y. D., Wang, Y. R., Zhou, M., Qiu, J. Z., Li, D. W., et al. (2022). Two new forest pathogens in Phaeolus (Polyporales, basidiomycota) on Chinese coniferous trees were confirmed by molecular phylogeny. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.942603
Zhang, Q. Y., Liu, Z. B., Liu, H. G., Si, J. (2023). Two new corticioid species of phanerochaetaceae (Polyporales, basidiomycota) from southwest China. Front. Cell. Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1105918
Zhou, H. M., Bau, T., Si, J. (2023). Morphological and phylogenetic evidence reveal three new Pseudohydnum (Auriculariales, basidiomycota) species from north China. Front. Cell. Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1139449
Zhou, L. W., Vlasák, J., Dai, Y. C. (2016). Taxonomy and phylogeny of Phellinidium (Hymenochaetales, basidiomycota): a redefinition and the segregation of Coniferiporia gen. nov. for forest pathogens. Fungal Biol. 120, 988–1001. doi: 10.1016/j.funbio.2016.04.008
Keywords: Hymenochaetaceae, phylogeny, polypore, taxonomy, new taxa
Citation: Chen Q, Liu L, Si J and Vlasák J (2023) Taxonomic and phylogenetic contributions to Fuscoporia (Hymenochaetales, Basidiomycota): two new species from Hawaii with a key to North American species. Front. Cell. Infect. Microbiol. 13:1205669. doi: 10.3389/fcimb.2023.1205669
Received: 14 April 2023; Accepted: 23 May 2023;
Published: 21 June 2023.
Edited by:
Tingting Dai, Nanjing Forestry University, ChinaReviewed by:
Sandrine Giraud, Université d’Angers, FranceHaonan Wang, Zhejiang Agriculture and Forestry University, China
Copyright © 2023 Chen, Liu, Si and Vlasák. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jing Si, amluZ3NpMTc4OEAxMjYuY29t; Josef Vlasák, dmxhc2FrQHVtYnIuY2FzLmN6