 Jiajia Yang1†
Jiajia Yang1† Ying Wang
Ying Wang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol. , 17 May 2023
Sec. Clinical Microbiology
Volume 13 - 2023 | https://doi.org/10.3389/fcimb.2023.1189417
This article is part of the Research Topic Pathogen-Host Interaction in the Development of Viral Hepatitis View all 9 articles
Viral hepatitis is a major worldwide public health issue, affecting hundreds of millions of people and causing substantial morbidity and mortality. The majority of the worldwide burden of viral hepatitis is caused by five biologically unrelated hepatotropic viruses: hepatitis A virus (HAV), hepatitis B virus (HBV), hepatitis C virus (HCV), hepatitis D virus (HDV), and hepatitis E virus (HEV). Metabolomics is an emerging technology that uses qualitative and quantitative analysis of easily accessible samples to provide information of the metabolic levels of biological systems and changes in metabolic and related regulatory pathways. Alterations in glucose, lipid, and amino acid levels are involved in glycolysis, the tricarboxylic acid cycle, the pentose phosphate pathway, and amino acid metabolism. These changes in metabolites and metabolic pathways are associated with the pathogenesis and medication mechanism of viral hepatitis and related diseases. Additionally, differential metabolites can be utilized as biomarkers for diagnosis, prognosis, and therapeutic responses. In this review, we present a thorough overview of developments in metabolomics for viral hepatitis.
Viral hepatitis is a significant global public health issue with hundreds of millions of victims and significant morbidity and mortality rates. Hepatitis A virus (HAV), hepatitis B virus (HBV), hepatitis C virus (HCV), hepatitis D virus (HDV), and hepatitis E virus (HEV) are the five biologically unrelated hepatotropic viruses that cause the majority of viral hepatitis worldwide (Xiang et al., 2022). Although HAV does not progress into a chronic infection, HBV, HCV, HDV, and HEV occasionally do. HBV and HCV in particular show a strong correlation with the occurrence of chronic infections. HBV and HCV infections cause the majority of viral hepatitis-related deaths. The statistics indicate that 296 million individuals were estimated to have hepatitis B, 58 million to have hepatitis C, and 1.1 million died as a result of viral hepatitis infections in 2019. As the body’s primary metabolic organ, the liver will inevitably change the linked metabolic network once sickness arises, which will have an impact on the levels of many endogenous small molecule metabolites in the body. The detection of these endogenous small molecules in tissues or fluids can be used as sensitive indicators of liver injury and can aid in the analysis and understanding of the etiology and pathogenesis of liver diseases (Xiang et al., 2022; Wolfe et al., 2023).
Metabolomics is a new systems biology field that uses methods from analytical chemistry to characterize the endogenous small-molecule metabolites that are found in tissues and biofluids. A variety of biological activities, such as mitochondrial activity, lipid biosynthesis and metabolism, glucose and glutamine metabolism, and nucleotide biosynthesis, can all be simultaneously analyzed using metabolomics. As metabolites are the downstream end products of gene and protein expression, they most closely correspond to phenotype (Manchester and Anand, 2017). In recent years, through the use of metabolomics research, the pathogenic mechanisms of infectious disorders, such as coronavirus disease 2019 (COVID-19) (Hasan et al., 2021), human immunodeficiency virus (HIV) disease (Olund Villumsen et al., 2021), and herpesviruses (HPV) infection (Abudula et al., 2020) have been successfully understood. Many researchers have suggested etiology and potential biomarkers for viral hepatitis and related diseases utilizing metabolomics resources, demonstrating the promise of the metabolomics method in increasingly complicated disorders (Cui et al., 2021). Therefore, this review covers the applications of metabolomics in various forms of viral hepatitis and associated diseases to shed light on the progression of these disorders.
In 1971, Pauling et al. performed the first large-scale quantitative detection of hundreds of substances in various biological fluids to reflect the functional state of biological systems, ushering in a new era of metabolomics research (Cui et al., 2018). The concept of metabolomics was first proposed by Nicholson et al. in 1999 (Nicholson et al., 1999). It is a science that analyzes the changes of small molecule metabolites (relative molecular mass <1 000) in biological tissues. Metabolomics research can be divided into non-targeted and targeted analysis methods, which differ in the quantitative level of research targets, sample preparation, accuracy and precision of obtained results, and the number of identified metabolites (Dunn et al., 2011). Untargeted metabolomics enables the investigation of complex interactions between a variety of metabolites from various pathways before the formulation of hypotheses, resulting in the discovery of new metabolites but also producing large data sets and experimental challenges in overcoming the identification of previously unidentified metabolites. Targeted metabolomics involves measuring or quantifying metabolites whose pertinent chemical structure and biological activity are known. Targeted metabolomics can be used to more accurately and selectively explain how some chemicals are associated with particular disease conditions (Roberts et al., 2012).
The classic metabolomics process includes the collection of samples (plasma, serum, urine, saliva, cell and tissue extracts and feces, etc.) and sample pretreatment, metabolite measurement, data processing, and statistical analysis, in order to obtain small molecules of biological significance that are closely related to metabolic characteristics (Roca et al., 2021). The commonly used measurement methods of metabolomics include nuclear magnetic resonance (NMR), liquid chromatography-mass spectrometry (LC-MS), and gas chromatography-mass spectrometry (GC-MS) (Carbone et al., 2021; Edison, 2021). NMR is simple to pretreat samples, does not damage the sample structure, requires a small number of samples, has a short measurement time, has high repeatability, can perform non-selective detection of specific metabolites, and can also classify the phenotype, but the sensitivity is not as good as MS, for some compounds with low content may not be detected. The volatility and thermal stability of the measured components are not required by LC-MS technology, and the preliminary processing is simple, allowing for rapid and efficient detection. However, there is a lack of a reference standard spectrum library, and identifying metabolites requires the use of standard substances, which is relatively difficult. GC-MS technology has a high resolution and detection sensitivity, as well as a standard spectrum library for reference, which can be used for qualitative metabolite characterization. However, derivation processing is required for samples with low volatility, and the early processing is complicated and may result in the loss of signals of some substances. As there is no single technology that can simultaneously measure and identify all metabolites in samples due to the diversity and complexity of metabolites, it is required to combine the data from different analytic platforms to gain more thorough metabolite information (Pomyen et al., 2020; Spina and Saliba, 2021). Bioanalytical procedures are used to get the raw data, which is then analyzed utilizing several stoichiometric approaches (Ghosh et al., 2020). The principal component analysis (PCA), hierarchical cluster analysis (HCA), and other unsupervised pattern recognition techniques, as well as supervised pattern recognition techniques like orthogonal partial least squares (OPLS), partial least squares-discriminant analysis (PIS-DA), and random forest analysis (RFA), are currently the most widely used pattern recognition techniques. The most often utilized of these are PCA, PLS-DA, and OPLS-DA (Trainor et al., 2017). The general workflow of metabolomics analysis was shown in Figure 1.
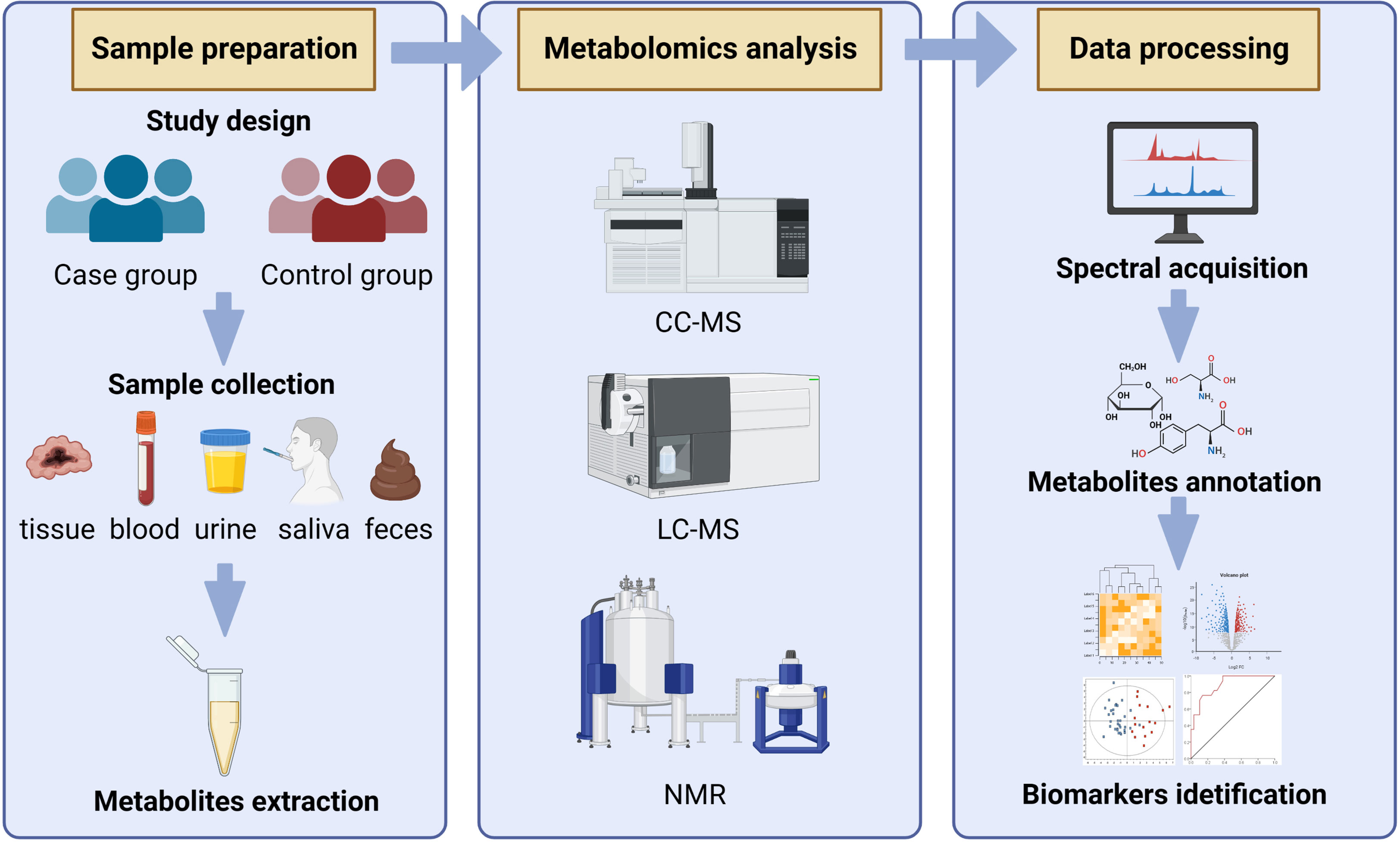
Figure 1 The general workflow of metabolomics analysis.
The key metabolic pathways and associated enzymes in the occurrence and development of diseases can be screened through the systematic characterization of metabolic profiles in the biological samples of subjects, and from there, the regulatory data of upstream related proteins and genes can be gathered (Schmidt et al., 2021). A succession of biological events in the entire biomolecular network during the diseased process can thus be understood by individual metabolomics analysis (Alarcon-Barrera et al., 2022). In terms of disease diagnosis, prognosis, drug effects, and knowledge of disease pathogenesis and progression, metabolomics has special advantages in the field of medicine.
HAV is a positive-strand RNA virus that spreads by the fecal-oral route. HAV epidemics are frequently linked to bad hygiene, crowded living conditions, or tainted food and water (Abutaleb and Kottilil, 2020). Children with HAV infections typically show no symptoms, whereas adults with HAV infections may exhibit signs such as hyperbilirubinemia, stomach discomfort, and jaundice. Hepatitis A is a self-limiting disease that primarily causes acute hepatitis and has a favorable prognosis (Xiang et al., 2022). Yamane et al. (1994) conducted phosphorus-31-31 NMR analysis on 26 patients with acute hepatitis A to assess the changes in phospholipid metabolism in hepatocytes during acute hepatitis A virus infection. Six signaling components corresponding to phosphomonoester (PME), inorganic phosphate (Pi), phosphodiester (PDE), and adenosine triphosphate isomers (α-, β-, and γ-ATP) were detected in the liver using spectroscopy in all patients and healthy subjects. Their results showed that the PME-PDE ratios in the patients significantly increased compared to the ratios in the controls during the early stages of the disease. The PME/PDE ratios reverted to controls within six weeks of the disease’s onset. According to the time course study, the PME/PDE ratio and the length of time after onset are inversely correlated.
In addition, Yamane et al. (1994) discovered that the spectral alterations shown in acute viral hepatitis A-infected human livers were comparable to those seen in rat livers that had undergone regeneration. Their findings demonstrated that metabolic profiling could provide non-invasive real-time information on human liver metabolism and could be a key method for monitoring liver regeneration (Yamane et al., 1994). At present, there are few studies on the metabolomics of hepatitis A, but this study provides a certain perspective and foundation for future research.
Globally, the hepatitis B virus (HBV) is the most frequent cause of both acute and chronic liver disorders, infecting around 4 million individuals annually, primarily in Asia and Africa (Khan et al., 2021). In about 10% of HBV-infected people, persistent infections like liver fibrosis and cirrhosis will appear (Lu et al., 2018). A million people per year pass away from chronic liver diseases due to hepatitis B. Most chronic hepatitis B (CHB) patients don’t have any noticeable symptoms at first, but as the condition worsens, they eventually get cirrhosis of the liver and hepatocellular carcinoma (HCC) (Su et al., 2022). The development of metabolomics investigations has sped up our understanding of the pathogenesis, early diagnosis and treatment of HBV infection.
As of right now, there are no drugs or forms of treatment that will completely eradicate HBV, therefore treatment must last a lifetime (McNaughton et al., 2021; Lian et al., 2022). HBV reinfection, particularly HBV infection in extrahepatic organs, has grown to be a significant risk factor for prognosis (Yue et al., 2002). The prospective therapeutics will become more clear if the mechanisms of HBV infection-associated liver diseases (HBV-CLD) is understood.
The microbiota can produce metabolites and active ingredients that participate in the regulation of host metabolism and immunity and is closely related to the pathological processes of HBV-CLD. Infection with HBV has been linked to intestinal microbial dysregulation. By combining investigations of the fecal microbiome and metabolome, a recent study (Shen et al., 2023) revealed a striking shift in the gut microbiota and metabolites in HBV-CLD patients. The progression of the disease and antiviral therapy were identified as the two main contributors to the shift. Through fecal metabolomics technology, Li et al (Li, 2021). also found that intestinal amino acid and glucose metabolism become aberrant when the HBV infection progresses.
Additionally, oral microbiota and metabolites may be involved in the development of HBV-CLD. Zhao et al. (2018) reported that the relative compositions of the tongue coating microbiotas and metabolites in the CHB patients were significantly different from those of the healthy controls, and the number of amino acid metabolites was greater in the CHB yellow tongue coating patients than in the white tongue coating patients. The researchers inferred that the metagenomic pathways that were enriched in the CHB yellow tongue coating patients were mainly those involved in amino acid metabolism. Another study by Zhao et al. (2013) showed significant differences in 17 urinary metabolites among CHB patients with different tongue coatings, and these metabolites may be the material basis of the two tongue coatings differentiation. Compared with the CHB white tongue coating patients, the CHB yellow tongue coating patients had higher HBV-DNA titers that is considered to be the most direct etiological basis of hepatitis B virus replication and infection (Gu et al., 2009).
Serum and urine are the simplest body fluid samples to collect. The detection of metabolite levels in serum and urine can provide new insights into disease pathogenesis and progression. Yang et al. (2016) used GC-MS metabolic profiles in serum and urine samples from CHB patients to conduct a covariation analysis to investigate metabolic anomalies in CHB. According to their findings, CHB patients may have altered pathways for a) glycine metabolism and b) the bridge that joins fatty acids with the tricarboxylic acid (TCA) cycle. Simillion et al. (2017) used GC-MS to analyze the plasma and urine of groups of HBV patients and healthy controls. They discovered 10 metabolite correlations for the HBV plasma and urine, but none for the controls. Their findings showed that increased glucose uptake, glycolysis, and pentose phosphate pathway metabolism were all consistent with HBV hepatitis; the latter uses xylitol and generates threonic acid, which may also be imported by glucose transporters. Xie et al (Xie, 2016). used serum metabolomics technology to screen out the metabolites isocitrate and isocitrate and lysophosphatidyl ethanolamine with significant differences in their expression in HBV DNA positive and negative groups, suggesting that the trilboic acid cycle pathway and the phospholipid metabolism pathway may be related to the pathogenesis of hepatitis B. Based on the serum metabolic profile, the metabolic processes identified by Du et al. (2011) that are related to the occurrence and development of HBV-related cirrhosis include amino acid metabolism, phospholipid metabolism, bile acid metabolism, tribolic acid cycle, and nicoculiamine metabolism. The biological processes involved include energy metabolism, protein synthesis and degradation, cell necrosis, inflammatory response, neurotransmitter synthesis, and degradation, etc. It can be seen that the metabolic abnormalities of liver cirrhosis are involved in all the main biochemical processes, among which amino acid metabolism is the most obvious. This may be because amino acids are widely involved in proteins, energy, and other processes in the body, most of which take place in the liver.
Gao et al (Gao, 2016). performed a paired comparative analysis of the metabolome of liver cancer tissue, paracancer tissue, and distant paracancer tissue from 39 patients with liver cancer, revealing that in order to balance the energy demands of rapid growth and proliferation of liver cancer cells as well as the material demands for biomacromolecule synthesis, on the one hand, glucose intake was increased and the glycolysis pathway was activated, while the tridellic acid pathway was deactivated. On the other hand, it significantly activates the pentose phosphate pathway (PPP), which, in addition to providing energy, primarily provides a variety of raw materials for anabolic metabolism and also produces a large amount of NADPH as a reducing power for biosynthesis and antioxidant stress damage. Furthermore, it increases glutamiculamine metabolism. Glutamiculamine is another non-mitochondrial ATP energy source for liver cancer cells, and it can provide energy and biomacromolecular synthesis raw materials for tumor cell growth and proliferation, as well as help maintain tumor cell microenvironment homeostasis. Furthermore, it improved the oxidative utilization of fatty acids and amino acid metabolism, providing enough energy for the growth and proliferation of liver cancer cells. By using untargeted NMR spectroscopy, Lan et al (2022). found that HBV-transgenic mice had significantly different metabolites from their wild-type littermates in the heart, liver, spleen, lung, kidney, pancreas, and intestine. They also discovered several metabolites, including branched-chain amino acids (BCAAs), a general potential biomarker in HBV-transgenic mice, which showed an increasing trend in most tissues, and choline, which may be a liver-specific biomarker for HBV detection. In HBV transgenic mice, the elevated choline concentrations may be associated with biofilm synthesis and lipid metabolism, providing lecithin for HBV replication. In addition, in response to oxidative stress caused by HBV replication, in the absence of glutathione, the concentration of BCAAs is increased to produce more alternative antioxidants.
HepG2.2.15 cells have a similar phenotype to HBV infected liver cells and have become a common model for the study of HBV infection (Wang et al., 2009). Through NMR spectroscopy, Li et al. (2015) conducted NMR metabolomics to detect and analyze metabolome differences between HepG2 and HepG2.2.15 cells, and found that HBV infection upregates hexoamine and phosphatidylcholine synthesis in host cells, generates oxidative stress and promotes central carbon metabolism and nucleoside synthesis. Using NMR metabolomics spectroscopy, Wan et al. (2017) discovered that HBV infection enhanced glycolysis, promoted cellular uptake of glucose, and improved the pentose phosphate and hexosamine pathways for the production of RNA, DNA, and nucleotide sugars to promote HBV replication. Furthermore, Li et al. (2017) confirmed through GC-MS analysis that HBV infection can alter the metabolism of amino acids and lipids in host cells, which is connected to numerous metabolic pathways including glycolysis, fatty acid metabolism, and amino acid metabolism. In addition, the information of metabolites obtained by GC-MS analysis is a good complement to that obtained by NMR. Recently, Hu et al. (2021) performed a metabolomics experiment via LC-MS, which demonstrated that host antiviral reaction to HBV is positively regulated by the hexosamine biosynthesis pathway by enhancing O-GlcNAc transferase-mediated protein O-GlcNAcylation in vitro and in vivo. Hepatitis B virus X protein (HBx) plays an important role in HBV-related HCC. To thoroughly research how HBx affects cell metabolism, Yue et al (Dan et al., 2016). used an NMR-based metabolomics approach. The findings suggested that HBx causes DNA damage at first, affects nucleic acid metabolism later, and then prevents DNA repair, leading to the development of HCC. Taking into account of the above evidence, metabolomics can be applied to explore the pathogenesis and progression of hepatitis B and related diseases.
Prolonged HBV infection may lead to chronic hepatitis, liver fibrosis, cirrhosis, and hepatocyte (HCC), and some patients may progress to liver failure, which seriously endangers the health and quality of life of patients. Therefore, early diagnosis and grading evaluation of hepatitis B and related diseases are of great significance (Table 1).
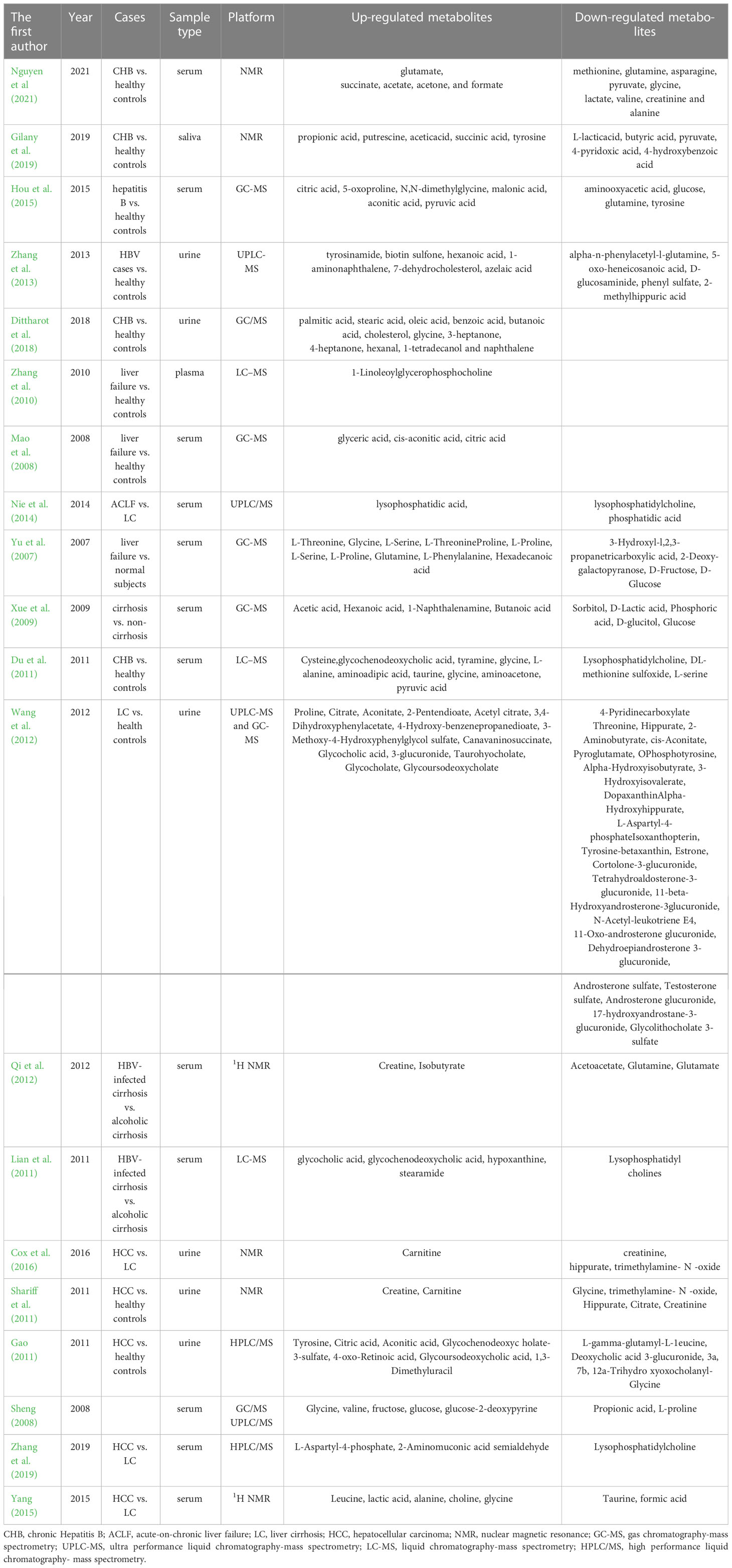
Table 1 Metabolites as the diagnostic biomarkers of hepatitis B and related diseases.
Metabolomics can be used to distinguish CHB patients from healthy controls and to differentiate between the various phases of CHB. Nguyen et al (2021) analyzed and compared the serum metabolites of patients at different stages of CHB and comparing them to a healthy individual, and found that perturbations in ammonia detoxification, glutamine and glutamate metabolism, methionine metabolism, dysregulation of branched-chain amino acids, and the TCA cycle are the main factors involved in the progression of the disease, and fluctuations increasing in aspartate, glutamate, glutamine, and methionine are fingerprints of progression. Their findings revealed the effectiveness of the metabolomics approach in detecting CHB transitioning from the immune tolerance to the immune clearance phase and can provide a more detailed decision basis for starting medical treatment. Hou et al. (2015) reported significant differences in serum metabolites between HBV patients and healthy controls. The area under the curve (AUC) of the total metabolite variables consisting of citric acid, aconitonic acid, glutamine, N,N- dimethyl glycine, malonic acid in the diagnosis of HBV infections was 0.975, which is of high diagnostic value. Gao et al. (2015) found that serum metabolites alanine, malic acid, and 5-methoxytryptamine could accurately distinguish the CHB group from the healthy control group. Zhang et al (Zhang et al., 2013). conducted metabolomics analysis of urine samples from HBV-infected patients and healthy controls, and observed that biotin sulfone, 5-oxo-heneicosanoic acid, d-Glucosaminide, and 2-methylhippuric acid were significantly different in the HBV patients and controls. Additionally, the ROC curve study of these four marker metabolites produced an AUC of 0.807, which was higher than that of ALT (0.619), AST (0.624), and LB (0.628), respectively. The unsupervised grouping further demonstrated that we can distinguish between controls and HBV cases by using the metabolites as biomarkers. Dittharot et al. (2018) also reported that GC/MS-based urine metabolomics in combination with multivariate statistical analysis distinguished CHB patients from healthy controls with excellent sensitivity (95%) and specificity (85%). Using NMR technology, Gilany et al. (2019) confirmed that salivary metabolites including propionic acid, putrescine, acetic acid, succini cacid, tyrosine, lactic acid, butyric acid, pyruvic acid, 4-pyridoxic acid, and 4-hydroxybenzoic acid were different between CHB patients and healthy subjects, which can be used as markers for the diagnosis of CHB.
Hepatitis B cirrhosis is a serious and irreversible liver disease. It is very important to accurately judge the severity and development trend of the disease and carry out targeted treatment to effectively control the development of the disease. In recent years, many scholars have used metabolomics to differentiate, grade, and forecast HBV-associated cirrhosis. Seven oxylipins derived from omega-6 were discovered to be altered in patients with HBV-related liver diseases, according to the research by Lu et al (Lu et al., 2018). These seven different oxylipins, when combined with AFP, age, and sex, greatly increased the prediction of HBV-related cirrhosis. The research by Xue et al. (2009) demonstrated a distinction in serum metabolites between HBV-infected non-cirrhosis and cirrhosis patients and verified the viability of GC-MS technology for the selection of biomarkers for cirrhosis after hepatitis B virus infection.
At present, the Child-Pugh grading system is commonly used to indicate the status of liver reserve function to judge the prognosis of chronic liver disease. However, there are still some limitations in the practical application, such as the subjective interference of score calculation is strong, and the response is not sensitive enough. Du et al. (2011) found that clinical stages of hepatitis B cirrhosis can be identified using serum metabolic profiles with high accuracy. Another study (Wang et al., 2012) revealed six urinary metabolites (α-hydroxy hippurate, tyrosine-betaxanthin, 3-hydroxy isovalerate, canavaninosuccinate, estrone, and glycoursodeoxycholate) were significantly altered among cirrhotic patients with CP A, B, and C, and demonstrated the diagnostic and prognostic potential for the development of cirrhosis.
The etiology of cirrhosis is various, and some scholars distinguished the etiology by metabolomics. Qi et al. (2012) detected serum metabolites of 21 patients with hepatitis B cirrhosis, 20 patients with alcoholic cirrhosis, and 20 healthy controls by the NMR method. The results showed that creatine, acetoacetic acid, isobutyrate, glutamine, and glutamate were significantly different between patients with hepatitis B cirrhosis and patients with alcoholic cirrhosis, and could be used as specific markers for the diagnosis of the two diseases. In the study of Lian et al. (2011), oleamide and myristamide were elevated in the serum of patients with alcoholic cirrhosis but reduced in those with HBV-induced cirrhosis, suggesting that they may be useful biomarkers for differentiating between these two types of cirrhosis.
HCC usually develops gradually, with an insidious start, late onset of clinical symptoms, and a high mortality rate. Early detection, diagnosis, and therapy are therefore essential for HCC patients. HCC is clinically identified through tissue-based histopathological results, distinctive radiologic characteristics, or blood-based assays like the AFP test. It would seem that the sensitivity and specificity of such clinically applied methodologies may not be sufficient in detecting early-stage HCC, greatly decreasing the reliability. Metabolomics has recently grown in prominence in the field of oncology due to the possibility of detecting homeostasis perturbations associated with the emergence of cancer (Lee et al., 2020). Through urinary nuclear magnetic resonance spectroscopy, Cox et al. (2016) studied patients with hepatitis B hepatocellular carcinoma in Bangladesh, and the results showed that carnitine was significantly increased in HCC, and creatinine, hippurate, and trimethylamine-N-oxide were significantly reduced in HCC compared to CHB, HBV-related cirrhosis, and healthy controls. The results may help in the creation of a low-cost HCC urinary dipstick screening test. Shariff et al. (2011) explored the urinary metabolic biomarkers of HBV-related HCC in an Egyptian Population. The discriminatory metabolites they found included glycine, trimethylamine-N-oxide, hippurate, citrate, creatinine, creatine, and carnitine. The sensitivity and specificity of the technique for distinguishing patients with HCC from healthy controls and patients with cirrhosis were 100%/94% and 81%/71%, respectively. In the Chinese population, Gao et al (Gao, 2011). also showed the usefulness of urine metabolomics techniques in the differential diagnosis of HBV-related HCC from cirrhosis and normal subjects. Moreover, there are numerous studies (Sheng, 2008; Zhang et al., 2013; Huang, 2014; Gao et al., 2015; Yang, 2015; Zhang et al., 2019) demonstrated that serum metabolites can distinguish HBV-related HCC from normal subjects. Additionally, a recent study (Lee et al., 2020) on the comprehensive metabolomic profile revealed that HCC with normal alpha-fetoprotein had serum levels of O-acetylcarnitine that were significantly higher than those with higher levels of AFP, and HCC with microscopic vascular invasion (VI) had preoperative serum levels of formate that were significantly higher than those with HCC without microscopic VI. Therefore, serum metabolites could be used to help with the early diagnosis of HCC patients who tested negative for AFP and the identification of microvascular invasion to aid in preoperative surgical planning and postoperative follow-up.
As HBV-related liver failure is a serious condition with a high mortality rate, early detection, precise diagnosis, and prompt treatment are crucial. Hyperbilirubinemia and a prolonged clotting time are the main criteria used in clinical practice to diagnose liver failure. The aforementioned two indices, however, do not accurately reflect liver function, which has limitations for the early detection of the liver failure. The early diagnosis of liver failure can significantly benefit from metabolomics. By using the PCA method to analyze the raw data (roughly 1000 compounds) from the plasma of patients with HBV-related liver failure and healthy controls, Zhang et al. (2010) found that a pattern recognition profile that distinguished significantly between patients with liver failure and healthy controls could be created. The ability of this model to predict liver failure has a 94.3 percent specificity and a 100 percent sensitivity, respectively. Mao et al. (2008) used GC/MS technology to perform serum metabolomics studies on 24 patients with HBV-related liver failure and 23 healthy controls, and found that glycolic acid, cisolmonic acid, and citric acid could be used as markers for the diagnosis of liver failure. Nie et al. (2014) discovered that the HBV-related acute-on-chronic failure (ACLF) group had substantially lower levels of lysophosphatidic acid and lecithin compared to the CHB group, the hepatitis B cirrhosis group, and the normal control group. The levels of lysophosphatidyllecithin, lysophosphatidic acid, and phosphatidic acid were consistent with the severity of liver disease and could be used as potential markers to forecast the outcome of acute-on-chronic liver failure. Yu et al. (2007) observed that the amino acid and glucose chromatograms in the HBV-induced liver failure group were significantly different from those in healthy people and that the identification of amino acid and glucose chromatograms could be used to distinguish early liver failure patients. Sheng et al (Sheng, 2008). also showed that serum metabolites can distinguish hepatitis B-associated liver failure from normal subjects. Li et al (Li, 2014). found that serum metabolite profiles of hepatitis B-related ACLF and chronic liver failure (CLF) were significantly different, and linocarnitine could be used as a specific biomarker for the diagnosis of ACLF and CLF. Another study showed that when compared to the group with a poor prognosis, the group with an improved prognosis had substantially higher levels of arginine and short- and long-chain coolyl carnitine, which were potential indicators of liver failure prognosis (Gao, 2019). The artificial liver support system (ALSS) is essential for the treatment of ACLF. Nowadays, several studies have shown that serum and urine metabolomics approaches could examine the dynamic change process of biomarkers before and after ALSS treatment, and some metabolites can be utilized as indicators to evaluate the prognosis of liver failure (Guo, 2011; Hao, 2011; Huang et al., 2021). These findings indicate that the metabolites in serum, urine and saliva can be used as markers for the diagnosis, development and treatment of hepatitis B and related diseases.
Some Chinese medicines have been used to treat hepatitis B, but the mechanism of action is unclear. Metabolomics is helpful to elucidate the efficacy and mechanism of TCM. Using the methods of epigenetics and metabolomics, Zhang et al. (2022) showed that the action mechanism of Chrysanthemi indici C (CIC) against HBV may be the synergistic action of multiple pathways and pathways multiple targets, including related inflammatory pathways, immune pathways and lipid metabolism, through regulating epigenetic expression balance and restoring the balance of cell microenvironment. Chai et al. (2018) confirmed that Tonkinensis may mainly act on 16 target proteins to exert anti-HBV effects, and its mechanism may be related to the regulation of retinol metabolism, peroxisome proliferator activate-receptors (PPAR) signaling pathway, transcription disorders in cancer and other processes, so as to control HBV replication and regulate immune and metabolic disorders in the body. Zheng et al. (2022) found that Kushen Wumei Decoction can resist HBV. Metabolomics and network pharmacology found that the mechanism of its action of anti-HBV may be related to the regulation of sphingolipid metabolism pathway, the biosynthesis of phenylalanine, tyrosine and tryptophan, and phenylalanine metabolism pathway, and PI3K-Akt signaling pathway. Recently, the results of the LC-MS/MS analysis revealed that C. nutans significantly upregulated the HBV mouse markers hippuric acid, L-histidine, trehalose, D-threitol, and stachyose while significantly downregulating uridine 5’-diphosphate, cholic acid, trimethylamine N-oxide CDP-ethanolamine, and phosphorylcholine. The correlation analysis showed that C nutans modulates essential bacteria (Alistipes), exhibiting specific anti-inflammatory effects on the levels of associated metabolites hippuric acid and cholic acid. The therapeutic potential for anti-HBV infection was demonstrated by these findings, which revealed that C. nutans exhibits protective effects in HBV model mice (Zhang et al.,2023). Hence, metabolomics plays an important role in exploring the mechanism of drug therapy for hepatitis B and related diseases.
HCV virions are spherical and are single-stranded positive-stranded RNA viruses. More than 71 million people globally are affected by HCV. HCV infection is an significant contributing factor to end-disease. Chronic HCV (CHC) infection is related to advanced liver disease and can cause hepatocellular carcinoma, which generates a variety of extrahepatic symptoms.
The majority of people with HCV infection (around 85%) acquire chronic infections. As with other types of chronic liver diseases, HCV infections are accompanied by liver fibrosis. If treatment is not timely, it can even develop into HCC, which seriously endangers the life and health of patients. A MS-based lipidomic investigation of 30 patients with chronic HCV infection and 30 healthy blood donor controls was undertaken, and identified 34 downregulated metabolites and 21 upregulated metabolites. Anandamide and eight fatty acid amides were elevated, which probably stimulating the cannabinoid receptor GPR55, a crucial host component for HCV replication. In addition, several lysophosphatidylinositols, which are necessary for the formation of phosphatidylinositol 4-phosphate pools required for HCV replication and can also activate the GPR55 receptor, were markedly downregulated (Beyoğlu et al., 2023). Ma et al. (2018) demonstrated that the changes in core fucosylation are protein and site specific during the progression of fibrotic liver disease and independent of the changes in the quantity of N-glycoproteins. Simillion et al. (2017) used GC-MS to examine the plasma and urine of groups of HCV cases. For the HCV plasma and urine, they found 18 metabolite associations. Their research demonstrated that HCV hepatitis was consistent with impaired glucose uptake, glycolysis, and pentose phosphate pathway metabolism, with the TCA pathway fueled by branched-chain amino acids feeding gluconeogenesis and the loss of glucose from hepatocytes, which most likely contributed to hyperglycemia.
Roe et al. (2011) investigated how HCV affects the hepatocyte metabolome. They discovered that during early HCV infection, several metabolites implicated in nucleotide synthesis and RNA replication significantly increased. Along with several amino acids, NAD levels were also noticeably raised. The effects of HCV infection included altered phospholipid metabolism, possible disruptions in mitochondrial fatty acid transport, a rise in cholesterol and sphingolipid levels, and disruptions in a number of lipid metabolic pathways. Changes in the glutathione synthesis route and fluctuations in 5’-methylthioadenosine levels were also observed. Aldose reductase activity caused by AKR1B10 is elevated in hepatitis C virus infection, according to research by Semmo et al. (2015) using metabolomics. Elevated glucose, mannose, and oleamide were detected in the plasma metabolomic phenotype of HCV-positive individuals, along with decreased plasma lactate. Reduced urine excretion of fructose and galactose was present in HCV-positive individuals, while urinary excretion of 6-deoxygalactose (fructose) and the polyols sorbitol, galactitol, and xylitol was raised. These findings suggested increased aldose reductase activity, and real-time quantitative polymerase chain reaction results with increased AKR1B10 gene expression in the liver supported this conclusion. It’s interesting to note that those who had previously been infected with HCV maintained their HCV infection’s metabolomic phenotype rather than reverting to an HCV-negative phenotype, suggesting that the effects of HCV on hepatic metabolism may be long lived. A comprehensive transcriptome, proteome, and metabolome analysis of liver tissues from HCV-infected mice and hepatocyte-like cells revealed that the STAT3 signaling pathway and glucose metabolism are both increased by HCV infection, which affects speroxisome activity. Alterations in peroxisome gene expression were also connected to the outcomes of patients with liver disorders (Lupberger et al., 2019). Therefore, these metabolomics studies contribute to our understanding of the pathogenesis and development of hepatitis C and related diseases.
The aminotransferase to platelet ratio index (APRI) and four-factor fibrosis index (FIB-4) are the most commonly used non-invasive fibrosis serologic biomarkers, both of which have high accuracy in ruling out significant fibrosis and confirming cirrhosis (Wai et al., 2003; Sterling et al., 2006). However, they were unable to identify intermediate fibrosis (Archer et al., 2022). Recently, there has been an increase in interest in metabolomics to find new biomarkers of hepatitis C, HCV-related cirrhosis, hepatic fibrosis, and HCC that are more accessible, specific, sensitive, and reproducible in a clinical context (Table 2).
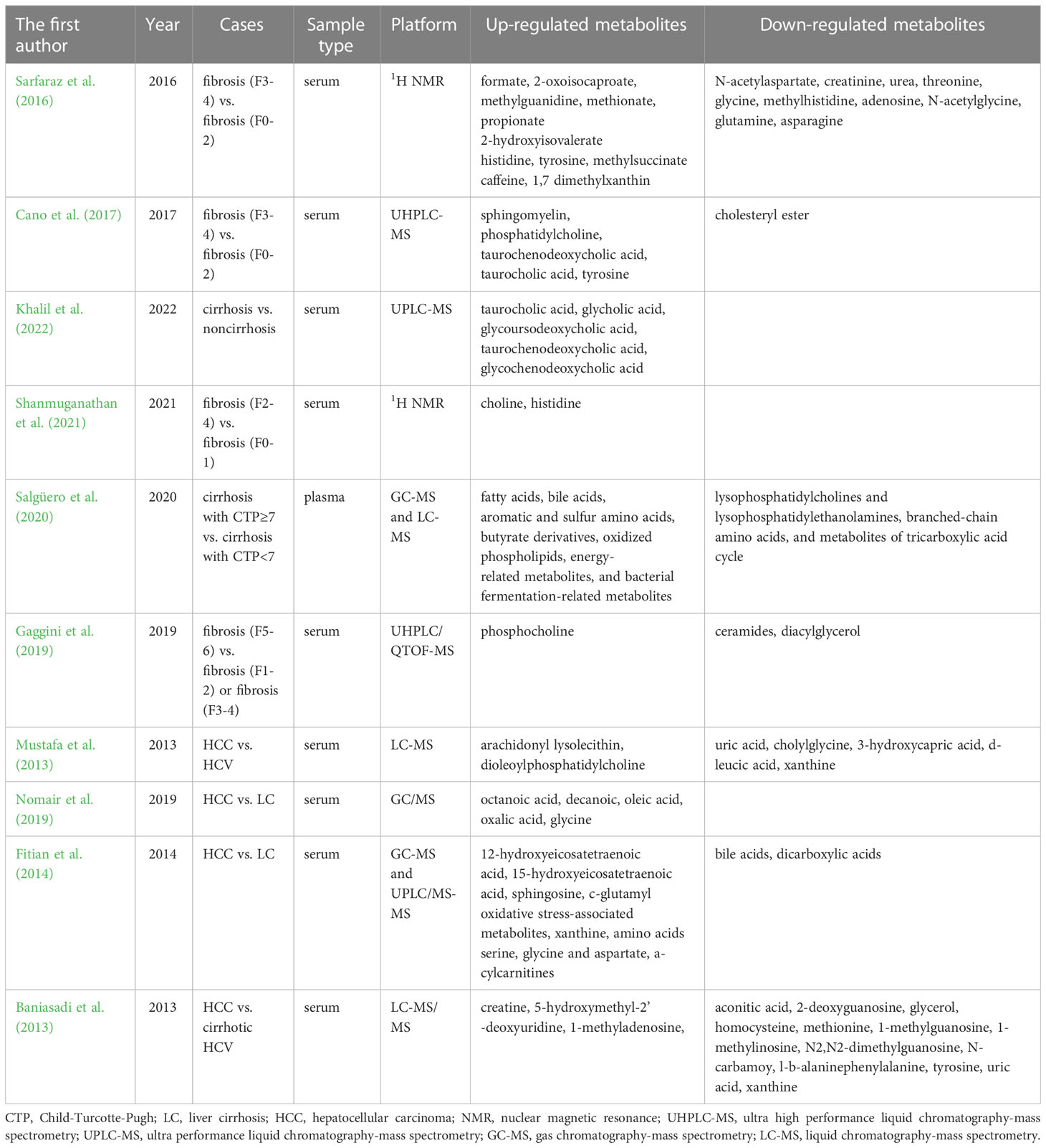
Table 2 Metabolites as the diagnostic biomarkers of hepatitis C and related diseases.
Godoy et al. (2010) reported that the metabolomics model based on NMR spectra of urine samples discriminated patients with HCV infection with high sensitivity and specificity. Additionally, the virology response of hepatitis C patients to treatment can be anticipated based on the metabolomics findings. Serum tryptophan levels in hepatitis C patients prior to treatment are considerably higher than those of patients who do not respond to treatment. These patients have a sustained virological response to the combination of PEG-IFN and RBV treatment (Saito et al., 2013).
Using NMR spectra of serum, Batista et al. (2018) developed metabolomic models to predict significant liver fibrosis, and cirrhosis in CHC in patients, which had excellent sensitivities and specificities. In addition, the models accurately classified the patients who had intermediate APRI and FIB-4 values, which might help avoid the need for these patients to have a liver biopsy. According to Salgüero et al. (2020), more advanced cirrhosis stages were characterized by an increase in plasma fatty acids, bile acids, aromatic and sulfur amino acids, butyrate derivatives, oxidized phospholipids, energy-related metabolites, and metabolites related to bacterial fermentation, while a decrease in branched-chain amino acids, TCA cycle metabolites, among other things, was observed. Lysophosphatidylcholine, taurocholic acid, and glycolic acid showed high accuracy in distinguishing patients with decompensated cirrhosis. Additionally, lower levels of sera ceramides, diacylglycerol, and phosphocholine were reported to be linked to greater levels of hepatic fibrosis and may be used as biomarkers of hepatic fibrosis (Gaggini et al., 2019). Shanmuganathan et al. (2021) observed that when compared to HCV patients with early-stage (F0-F1) liver fibrosis, serum levels of choline and histidine were reliably higher in late-stage (F2-F4) liver fibrosis patients. Using a receiver operating characteristic curve, the ratio of serum choline to uric acid allowed for the best differentiation of the severity of liver disease, and it was favorably correlated with ultrasound imaging measurements of liver stiffness. Sarfaraz et al. (2016) also demonstrated the utility of a metabolomics profiling approach to non-invasively identify biomarkers of liver fibrosis, steatosis and inflammation in patients with chronic HCV. Cnao et al (Cano et al., 2017). examined serum metabolomics in transplanted Hepatitis C patients and discovered that a model consisting of two sphingomyelins and two phosphatidylcholines accurately classifies rapid and slow fibrosers after transplantation, with an AUROC of 0.92, sensitivity of 71%, specificity of 85%, and accuracy of 84%. Fitian et al. (2014) observed the highly correlation of elevations in bile acids and dicarboxylic acids with HCV-associated cirrhosis, and these serum markers were highly sensitive and specific for cirrhosis.
Mustafa et al. (2013) conducted a study in which serum global metabolite profiles from patients with HCC and HCV were obtained using high performance liquid chromatography-mass spectrometry (HPLC-MS) techniques, making it simple to distinguish between patients with HCC and HCV and those with HCV alone. In addition, metabolomics could be applied in distinguishing HCV-related HCC from liver cirrhosis. In the study of Nomair et al. (2019), total of 34 known plasma metabolites were detected, and five of them—octanoic acid (caprylic acid), decanoic acid (capric acid), oleic acid, oxalic acid, and glycine—were found to have the strongest discriminatory potential for separating the HCC and cirrhosis groups, and ROC curve analysis revealed that oleic acid, octanoic acid and glycine had higher positive predictive value than AFP. An integrated metabolomic profiling analysis (Fitian et al., 2014) through GC/MS and UPLC/MS-MS revealed that the presence of HCV-related HCC was highly correlated with elevated levels of 12-hydroxyeicosatetraenoic acid (12-HETE), 15-HETE, sphingosine, c-glutamyl oxidative stress-associated metabolites, xanthine, amino acids serine, glycine, and aspartate, and a-cylcarnitines, and these serum markers were highly sensitive and specific for HCC. Baniasadi et al. (2013) noticed that 16 serum metabolites were substantially different between HCC and cirrhotic HCV patients. A categorization model was created and internally validated using PLS-DA analysis according to the four of the metabolites (methionine, 5-hydroxymethyl-2’-deoxyuridine, N2,N2-dimethylguanosine, and uric acid) that had the lowest p values. With sensitivity, specificity, and AUC of 97%, 95%, and 0.98, respectively, the model demonstrated excellent classification accuracy for differentiating the two groups. Furthermore, through GC-MS -based metabolomic analyses, Liu et al. (2018) suggested that the combination of glutamate and aspartate and the combination of glycerol and proline may be potential biomarkers for the prediction of HCC recurrence before and after radiofrequency ablation (RFA) treatment, respectively. According to a recent study, the fold changes in many serum bile acid concentrations showed a linear trend with hepatocellular carcinoma > cirrhosis > noncirrhosis > healthy controls, and ROC curve analysis revealed five conjugated acids, TCA, GCA, GUDCA, TCDCA, and GCDCA, that distinguished HCC from noncirrhotic liver patients (Khalil et al., 2022). Overall, metabolites in blood and urine can be used as markers to evaluate the occurrence, progression, and therapeutic efficacy of hepatitis C and related diseases.
At present, antiviral therapy is the main treatment for hepatitis C. The effectiveness of pharmacological therapy and potential biochemical mechanisms can be better understood through the use of metabolite analysis.
Direct-acting antivirals (DAAs) and ribavirin (RBV), which lead to a shorter medication period and a sustained virologic response up to 98%, are the most widely utilized antiviral treatment against the hepatitis C virus. However, a number of RBV adverse effects that are dose-related may restrict its uses (Tsubota et al., 2003). In a recent study (Giampaoliet al., 2022), RBV and its metabolites were measured in the urine of HCV patients receiving DAAs+RBV therapy in order to assess the ability of HCV patients to metabolize drugs, as well as the adverse effects such as anemia in relation to RBV metabolite levels. Some HCV patients were discovered to maintain significant levels of RBV during both the TW4 and EOT stages, while another several patients were discovered to maintain high levels of RBV proactive metabolites, most likely because of nucleosidase activity. Hepatic fibrosis and metabolic changes are caused by oxidative stress and a protracted inflammatory response brought on by HCV infection in the liver microenvironment. Liver damage is only partially repaired even though DAAs cause HCV-clearance. According to the study of Biliotti et al. (2021),a catabolic intermediate of nicotinamide-adenine-dinucleotide, 1-methylnicotinamide, was considerably elevated in HCV patients and restored following HCV clearance, most likely as a result of the diminished hepatic inflammation. An enhancement in skeletal muscle protein synthesis was suggested by 3-hydroxy-3-methylbutyrate, a leucine-catabolism intermediary that was completely restored following HCV elimination. Glycine and choline momentarily rose throughout therapy, as did 3-hydroxyisobutyrate and 2, 3-dihydroxy-2-methylbutyrate, intermediates of valine catabolism. Using LC-MS/MS and lipoprotein electrophoresis assays, Chang et al. (2018) demonstrated that at 24 weeks post-anti-HCV-therapy, accelerated cholesterol biosynthesis, hepatic lipid export, ω-oxidation and decreased systemic inflammation were noted in CHC patients with sustained virological response (SVR); while HCV-associated lipid metabolic alterations required >24 weeks for restoration or were incompletely reversible after SVR. Jun et al (Wang et al., 2013). showed the effectiveness of metabolomics in capturing and elucidating metabolic characteristics of HCV and the therapeutic benefits of the bear bile powder (PBBP). Keogh et al. (2017) conducted a GCMS-based metabolomic investigation to clarify the mechanisms underlying the efficacy of a novel anti-Hepatitis C virus and anti proliferative agent in altering metabolic networks in hepG2 and hep3B Cells, which showed that metabolomics can provide mechanistic insights into the efficacy of novel drug candidates prior to the appearance of their pharmacological effect. Therefore, metabolomics is an effective method to explore the mechanism of drug treatment of hepatitis C and related diseases.
HEV is a significant zoonotic virus that can affect a variety of hosts. It is mainly transmitted through the fecal-oral route and has obvious seasonality. It has eight major genotypes. Most HEV-infected individuals have no symptoms, but some may have jaundice and other acute hepatitis symptoms (Wu et al., 2019; Wu and Huang, 2020). HEV infection can also lead to a variety of extrahepatic symptoms (Wu et al., 2021). Although HEV infection typically results in acute and self-limiting diseases, acute liver failure (ALF) brought on by HEV infection has increased in prevalence over the past few years, with serious implications for patient outcomes (Wu et al., 2020).
Using ultra-performance liquid chromatography-mass spectroscopy (UPLC-MS), a recent study showed that there were significant differences in the serum metabolite components between AHE patients and healthy controls, or between HEV-ALF and acute hepatitis E (AHE) patients. Their research indicated that dynamic changes in serum metabolites were related to the occurrence and severity of AHE, and can be used as diagnostic and prognostic markers for HEV-ALF (Wu et al., 2022).
Munshi et al. (2011) discovered variations in the concentrations of several metabolites in the plasma and urine of hepatitis E patients. According to pathway analysis methods, glycolysis, urea cycle, TCA cycle, and amino acid metabolism may be involved in acute hepatitis E patients. These discoveries could contribute to a better understanding of the pathophysiologic mechanisms at play in this disease as well as its clinical and biochemical manifestations. Currently, researches on metabolomics in hepatitis E and related diseases are still scarce, and the in-depth mechanism discussion and markers studies may be the hot topics in the future.
Metabolomics is still a developing field, and as such, it still has several limitations, like the variety of detection samples. Emphasis must be placed on the value of reliable metabolite measurement and standardization of biological materials. The type and quantity of metabolites can also be affected by a variety of variables, including sampling time, collection method, treatment, storage stability, extraction, sample dilution, measurement methods, and statistical analysis. In addition, outside variables like body mass index, diet, physical activity, and treatment may also have an impact on the progression of the disease and consequently metabolite levels.
The use of metabolomics offers a fresh approach and viewpoint for understanding the mechanisms of occurrence and development, diagnosis, prognosis, and therapy of viral hepatitis and related diseases. The metabolomics of HBV, HCV, and associated disorders have been extensively studied. Further associations have emerged and several biomarkers have been discovered. The efficacy of these screening indicators in patient diagnosis and prognosis, however, is still up for debate. Furthermore, the metabolomics of other hepatitis is insufficient and needs further investigation.
All the authors contributed substantially to this review. YW: conceptualization and design. JY, DW and YL: manuscript drafting. HW and QH: revising, checking, and approving of the final version of the manuscript.
We sincerely appreciate the relevant published researches for their support, meanwhile, thank the reviewers and editors for their valuable time and helpful suggestions. The Figure 1 was created with biorender.com.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abudula, A., Rouzi, N., Xu, L., Yang, Y., Hasimu, A. (2020). Tissue-based metabolomics reveals potential biomarkers for cervical carcinoma and hpv infection. Bosnian J. basic Med. Sci. 20 (1), 78–87. doi: 10.17305/bjbms.2019.4359
Abutaleb, A., Kottilil, S. (2020). Hepatitis a: epidemiology, natural history, unusual clinical manifestations, and prevention. Gastroenterol. Clinics North America 49 (2), 191–199. doi: 10.1016/j.gtc.2020.01.002
Alarcon-Barrera, J. C., Kostidis, S., Ondo-Mendez, A., Giera, M. (2022). Recent advances in metabolomics analysis for early drug development. Drug Discovery Today 27 (6), 1763–1773. doi: 10.1016/j.drudis.2022.02.018
Archer, A. J., Belfield, K. J., Orr, J. G., Gordon, F. H., Abeysekera, K. W. (2022). Easl clinical practice guidelines: non-invasive liver tests for evaluation of liver disease severity and prognosis. Frontline Gastroenterol. 13 (5), 436–439. doi: 10.1136/flgastro-2021-102064
Baniasadi, H., Gowda, G. A., Gu, H., Zeng, A., Zhuang, S., Skill, N., et al. (2013). Targeted metabolic profiling of hepatocellular carcinoma and hepatitis c using lc-Ms/Ms. Electrophoresis 34 (19), 2910–2917. doi: 10.1002/elps.201300029
Batista, A. D., Barros, C. J. P., Costa, T., de Godoy, M. M. G., Silva, R. D., Santos, J. C., et al. (2018). Proton nuclear magnetic resonance-based metabonomic models for non-invasive diagnosis of liver fibrosis in chronic hepatitis c: optimizing the classification of intermediate fibrosis. World J. Hepatol. 10 (1), 105–115. doi: 10.4254/wjh.v10.i1.105
Beyolu, D., Schwalm, S., Semmo, N., Huwiler, A., Idle, J. R. (2023). Hepatitis C Virus Infection Upregulates Plasma Phosphosphingolipids and Endocannabinoids and Downregulates Lysophosphoinositols. International journal of molecular sciences 24(2), 1407.
Biliotti, E., Giampaoli, O., Sciubba, F., Marini, F., Tomassini, A., Palazzo, D., et al. (2021). Urinary metabolomics of hcv patients with severe liver fibrosis before and during the sustained virologic response achieved by direct acting antiviral treatment. Biomed pharmacother = Biomed pharmacother 143, 112217. doi: 10.1016/j.biopha.2021.112217
Cano, A., Mariño, Z., Millet, O., Martínez-Arranz, I., Navasa, M., Falcón-Pérez, J. M., et al. (2017). A metabolomics signature linked to liver fibrosis in the serum of transplanted hepatitis c patients. Sci. Rep. 7 (1), 10497. doi: 10.1038/s41598-017-10807-y
Carbone, F., Bruzzaniti, S., Fusco, C., Colamatteo, A., Micillo, T., De Candia, P., et al. (2021). Metabolomics, lipidomics, and immunometabolism. Methods Mol. Biol. (Clifton NJ) 2285, 319–328. doi: 10.1007/978-1-0716-1311-5_24
Chai, X., Meng, Y., Bo, Z., Zhang, Y., Xiao, X., Niu, M. (2018). A preliminary study on the mechanism of action against hepatitis b virus by rhizoma vulgaris based on biological target network analysis. Acta Pharm. Sin. 53 (3), 396–402. doi: 10.16438/j.0513-4870.2017-1124
Chang, S. W., Cheng, M. L., Shiao, M. S., Yeh, C. T., Wang, C. H., Fan, C. M., et al. (2018). Recovery of lipid metabolic alterations in hepatitis c patients after viral clearance: incomplete restoration with accelerated Ω-oxidation. J. Clin. lipidol 12 (3), 756–766. doi: 10.1016/j.jacl.2018.02.011
Cox, I. J., Aliev, A. E., Crossey, M. M., Dawood, M., Al-Mahtab, M., Akbar, S. M., et al. (2016). Urinary nuclear magnetic resonance spectroscopy of a Bangladeshi cohort with hepatitis-b hepatocellular carcinoma: a biomarker corroboration study. World J. Gastroenterol. 22 (16), 4191–4200. doi: 10.3748/wjg.v22.i16.4191
Cui, D., Li, W., Jiang, D., Wu, J., Xie, J., Wu, Y. (2021). Advances in multi-omics applications in hbv-associated hepatocellular carcinoma. Front. Med. 8. doi: 10.3389/fmed.2021.754709
Cui, L., Lu, H., Lee, Y. H. (2018). Challenges and emergent solutions for lc-Ms/Ms based untargeted metabolomics in diseases. Mass spectrom Rev. 37 (6), 772–792. doi: 10.1002/mas.21562
Dan, Y., Zhang, Y., Cheng, L., Ma, J., Xi, Y., Yang, L., et al. (2016). Hepatitis b virus X protein (Hbx)-induced abnormalities of nucleic acid metabolism revealed by (1)H-Nmr-Based metabonomics. Sci. Rep. 6, 24430. doi: 10.1038/srep24430
Dittharot, K., Jittorntam, P., Wilairat, P., Sobhonslidsuk, A. (2018). Urinary metabolomic profiling in chronic hepatitis b viral infection using gas Chromatography/Mass spectrometry. Asian Pac J. Cancer prevent: APJCP 19 (3), 741–748. doi: 10.22034/apjcp.2018.19.3.741
Du, Z., Zhang, L., Liu, S. (2011). Application of liquid chromatography-mass spectrometry in the study of metabolic profile at different stages of liver cirrhosis. Chin. J. Chromatogr. 29 (4), 314–319. doi: 10.3724/sp.j.1123.2011.00314
Dunn, W. B., Broadhurst, D. I., Atherton, H. J., Goodacre, R., Griffin, J. L. (2011). Systems level studies of mammalian metabolomes: the roles of mass spectrometry and nuclear magnetic resonance spectroscopy. Chem. Soc. Rev. 40 (1), 387–426. doi: 10.1039/b906712b
Edison, A. S., Colonna, M., Gouveia, G. J., Holderman, N. R., Judge, M. T., Shen, X., et al (2021). NMR: Unique strengths that enhance modern metabolomics research. Analytical Chemistry 93 (1), 478–499. doi: 10.1021/acs.analchem.0c04414
Fitian, A. I., Nelson, D. R., Liu, C., Xu, Y., Ararat, M., Cabrera, R. (2014). Integrated metabolomic profiling of hepatocellular carcinoma in hepatitis c cirrhosis through Gc/Ms and Uplc/Ms-Ms. Liver Int. 34 (9), 1428–1444. doi: 10.1111/liv.12541
Gaggini, M., Carli, F., Rosso, C., Younes, R., D'Aurizio, R., Bugianesi, E., et al (2019). Altered metabolic profile and adipocyte insulin resistance mark severe liver fibrosis in patients with chronic liver disease. International Journal of Molecular Sciences 20 (24), 6333. doi: 10.3390/ijms20246333
Gao, C. (2011). Metabolomics study of urine in patients with hepatocellular carcinoma (Tianjin: Tianjin Medical University).
Gao, R. (2016). GC-MS based metabolomics study and biomarkers screening of hepatocellular carcinoma (Beijing:Academy of Military Medical Sciences).
Gao, P. (2019). Metabolomic analysis of hepatitis b virus-associated chronic hepatitis, cirrhosis, liver cancer and liver failure (Shandong:Shandong University).
Gao, R., Cheng, J., Fan, C., Shi, X., Cao, Y., Sun, B., et al. (2015). Serum metabolomics to identify the liver disease-specific biomarkers for the progression of hepatitis to hepatocellular carcinoma. Sci. Rep. 5, 18175. doi: 10.1038/srep18175
Ghosh, T., Zhang, W., Ghosh, D., Kechris, K. (2020). Predictive modeling for metabolomics data. Methods Mol. Biol. (Clifton NJ) 2104, 313–336. doi: 10.1007/978-1-0716-0239-3_16
Giampaoli, O., Sciubba, F. (2022). Precision medicine: determination of ribavirin urinary metabolites in relation to drug adverse effects in hcv patients. International Journal of Molecular Sciences 23 (17), 10043. doi: 10.3390/ijms231710043
Gilany, K., Mohamadkhani, A., Chashmniam, S., Shahnazari, P., Amini, M., Arjmand, B., et al. (2019). Metabolomics analysis of the saliva in patients with chronic hepatitis b using nuclear magnetic resonance: a pilot study. Iranian J. basic Med. Sci. 22 (9), 1044–1049. doi: 10.22038/ijbms.2019.36669.8733
Godoy, M. M., Lopes, E. P., Silva, R. O., Hallwass, F., Koury, L. C., Moura, I. M., et al. (2010). Hepatitis c virus infection diagnosis using metabonomics. J. Viral hepa 17 (12), 854–858. doi: 10.1111/j.1365-2893.2009.01252.x
Gu, X. B., Yang, X. J., Wang, D., Hua, Z., Xu, Y. Q., Lu, Z. H. (2009). Relationship between serum hbv DNA level and hbv-specific, nonspecific cytotoxic T lymphocytes and natural killer cells in patients with chronic hepatitis b. Chin. Med. J. 122 (18), 2129–2132. doi: 10.3760/cma,j.issn0366-6999.2009.18.010
Guo, Y. (2011). Urine metabolomics in patients with hepatitis b liver failure treated with alss (Zhejiang: Zhejiang University).
Hao, S. (2011). Metabonomics of artificial liver in the treatment of chronic acute (Subacute) hepatitis b liver failure (Zhejiang: Zhejiang University).
Hasan, M. R., Suleiman, M., Pérez-López, A. (2021). Metabolomics in the diagnosis and prognosis of covid-19. Front. Genet. 12. doi: 10.3389/fgene.2021.721556
Hou, Y., Zhu, W., Chen, C., Wang, Y., Duan, Z., Yan, C. (2015). Exploration of hepatitis b serum metabolic markers based on gas chromatography-mass spectrometry technology. Chin. J. Chromatogr. 4), 383–388. doi: 10.3724/sp.j.1123.2014.12007
Hu, J., Gao, Q., Yang, Y., Xia, J., Zhang, W., Chen, Y., et al. (2021). Hexosamine biosynthetic pathway promotes the antiviral activity of Samhd1 by enhancing O-glcnac transferase-mediated protein O-glcnacylation. Theranostics 11 (2), 805–823. doi: 10.7150/thno.50230
Huang, Y. (2014). Analysis of serum metabolic profile in patients with hepatocellular carcinoma of different degrees of differentiation (Tianjin: Tianjin Medical University).
Huang, S., Hu, D., Yuan, S., He, Y., Li, C., Zhu, Y., et al. (2021). The serum metabolomics study of liver failure and artificial liver therapy intervention. Med. Sci. monitor 27, e930638. doi: 10.12659/msm.930638
Keogh, A., Şenkardeş, S., Idle, J. R., Küçükgüzel, ŞG., Beyoğlu, D. (2017). A novel anti-hepatitis c virus and antiproliferative agent alters metabolic networks in Hepg2 and Hep3b cells. Metabolites 7 (2), 23. doi: 10.3390/metabo7020023
Khalil, A., ElSheashaey, A., Abdelsameea, E., Obada, M., Bayomy, F. F. M., El-Said, H. (2022). Value of bile acids in diagnosing hepatitis c virus-induced liver cirrhosis and hepatocellular carcinoma. Br. J. Biomed. Sci. 79. doi: 10.3389/bjbs.2021.10191
Khan, A., Ahsan, O., Wei, D. Q. (2021). Computational evaluation of abrogation of hbx-Bcl-Xl complex with high-affinity carbon nanotubes (Fullerene) to halt the hepatitis B virus replication Molecules (Basel, Switzerland) 26 (21), 6433. doi: 10.3390/molecules26216433
Lan, W., Wang, Y. (2022). Metabolic regulation of hepatitis b virus infection in hbv-transgenic mice. Metabolites 12 (4), 287. doi: 10.3390/metabo12040287
Lee, C. W., Yu, M. C., Lin, G., Chiu, J. C., Chiang, M. H., Sung, C. M., et al. (2020). Serum metabolites may be useful markers to assess vascular invasion and identify normal alpha-fetoprotein in hepatocellular carcinoma undergoing liver resection: a pilot study. World J. Surg. Oncol. 18 (1), 121. doi: 10.1186/s12957-020-01885-w
Li, X. (2014). Preliminary study on serum metabolomics of chronic hepatitis b plus acute liver failure, chronic liver failure and pre-liver failure (Zhejiang: Zhejiang University).
Li, Y. (2021). Metagenomics and metabonomics-based study on the association between specific gut microbiota and the dysfunction of NK cells in the immune-tolerant phase of chronic hepatitis B virus infection (Fujian medical university), Fujian.
Li, J., Wan, Q., Gan, M., Tang, H. (2017). Effects of hepatitis b virus on metabolism of Hepg2 cells by gas chromatography-mass spectrometry. J. Shanghai Jiao Tong Univ. 37 (10), 1322–1326. doi: 10.3969/j.issn.1674-8115.2017.10.002
Li, H., Zhu, W., Zhang, L., Lei, H., Wu, X., Guo, L., et al. (2015). The metabolic responses to hepatitis b virus infection shed new light on pathogenesis and targets for treatment. Sci. Rep. 5, 8421. doi: 10.1038/srep08421
Lian, J., Lin, F., Lu, Y., Hao, S., Cai, H., Zhang, X., et al. (2022). Alterations in cirrna expression profile during the development of acute-on-Chronic liver failure induced by hepatitis b. Adv Gut Microbiom Res. 1179, 71–107. doi: 10.1007/978-981-13-9151-4_4
Lian, J. S., Liu, W., Hao, S. R., Guo, Y. Z., Huang, H. J., Chen, D. Y., et al. (2011). A serum metabonomic study on the difference between alcohol- and hbv-induced liver cirrhosis by ultraperformance liquid chromatography coupled to mass spectrometry plus quadrupole time-of-Flight mass spectrometry. Chin. Med. J. 124 (9), 1367–1373. doi: 10.3760/cma.jissn.0366-6999.2011.09.018
Liu, Z., Nahon, P., Li, Z., Yin, P., Li, Y., Amathieu, R., et al. (2018). Determination of candidate metabolite biomarkers associated with recurrence of hcv-related hepatocellular carcinoma. Oncotarget 9 (5), 6245–6258. doi: 10.18632/oncotarget.23500
Lu, Y., Fang, J., Zou, L., Cui, L., Liang, X., Lim, S. G., et al. (2018). Omega-6-Derived oxylipin changes in serum of patients with hepatitis b virus-related liver diseases. Metabolomics 14 (3), 26. doi: 10.1007/s11306-018-1326-z
Lu, D., Zhuo, J., Yang, M., Wang, C., Pan, L., Xie, H., et al. (2018). The association between donor genetic variations in one-carbon metabolism pathway genes and hepatitis b recurrence after liver transplantation. Gene 663, 121–125. doi: 10.1016/j.gene.2018.03.071
Lupberger, J., Croonenborghs, T., Roca Suarez, A. A., Van Renne, N., Jühling, F., Oudot, M. A., et al. (2019). Combined analysis of metabolomes, proteomes, and transcriptomes of hepatitis c virus-infected cells and liver to identify pathways associated with disease development. Gastroenterology 157 (2), 537–51.e9. doi: 10.1053/j.gastro.2019.04.003
Ma, J., Sanda, M., Wei, R., Zhang, L., Goldman, R. (2018). Quantitative analysis of core fucosylation of serum proteins in liver diseases by lc-Ms-Mrm. J. Proteomics 189, 67–74. doi: 10.1016/j.jprot.2018.02.003
Manchester, M., Anand, A. (2017). Metabolomics: strategies to define the role of metabolism in virus infection and pathogenesis. Adv. Virus Res. 98, 57–81. doi: 10.1016/bs.aivir.2017.02.001
Mao, Y., Huang, X., Yu, K., Qu, H. B., Liu, C. X., Cheng, Y. Y. (2008). Metabonomic analysis of hepatitis b virus-induced liver failure: identification of potential diagnostic biomarkers by fuzzy support vector machine. J. Zhejiang Univ. Sci. B 9 (6), 474–481. doi: 10.1631/jzus.B0820044
McNaughton, A. L., Lemoine, M., van Rensburg, C., Matthews, P. C. (2021). Extending treatment eligibility for chronic hepatitis b virus infection. Gastroenterology & Hepatology 18 (3), 146–147. doi: 10.1038/s41575-020-00398-x
Munshi, S. U., Taneja, S., Bhavesh, N. S., Shastri, J., Aggarwal, R., Jameel, S. (2011). Metabonomic analysis of hepatitis e patients shows deregulated metabolic cycles and abnormalities in amino acid metabolism. J. Viral hepa 18 (10), e591–e602. doi: 10.1111/j.1365-2893.2011.01488.x
Mustafa, M. G., Petersen, J. R., Ju, H., Cicalese, L., Snyder, N., Haidacher, S. J., et al. (2013). Biomarker discovery for early detection of hepatocellular carcinoma in hepatitis c-infected patients. Mol. Cell. proteomics: MCP 12 (12), 3640–3652. doi: 10.1074/mcp.M113.031252
Nguyen, H. T. T., Wimmer, R., Le, V. Q., Krarup, H. B. (2021). Metabolic fingerprint of progression of chronic hepatitis b: changes in the metabolome and novel diagnostic possibilities. Metabolomics : Official journal of the Metabolomic Society 17 (2), 16. doi: 10.1007/s11306-020-01767-y
Nicholson, J. K., Lindon, J. C., Holmes, E. (1999). ‘Metabonomics’: understanding the metabolic responses of living systems to pathophysiological stimuli Via multivariate statistical analysis of biological nmr spectroscopic data. Xenobiotica; fate foreign compounds Biol. Syst. 29 (11), 1181–1189. doi: 10.1080/004982599238047
Nie, C. Y., Han, T., Zhang, L., Li, Y., Liu, H., Xiao, S. X., et al. (2014). Cross-sectional and dynamic change of serum metabolite profiling for hepatitis b-related acute-on-Chronic liver failure by Uplc/Ms. J. Viral hepa 21 (1), 53–63. doi: 10.1111/jvh.12122
Nomair, A. M., Madkour, M. A., Shamseya, M. M., Elsheredy, H. G., Shokr, A. (2019). Profiling of plasma metabolomics in patients with hepatitis c-related liver cirrhosis and hepatocellular carcinoma. Clin. Exp. Hepatol. 5 (4), 317–326. doi: 10.5114/ceh.2019.89478
Olund Villumsen, S., Benfeitas, R., Knudsen, A. D., Gelpi, M., Høgh, J., Thomsen, M. T., et al. (2021). Integrative lipidomics and metabolomics for system-level understanding of the metabolic syndrome in long-term treated hiv-infected individuals. Front. Immunol. 12. doi: 10.3389/fimmu.2021.742736
Pomyen, Y., Wanichthanarak, K., Poungsombat, P., Fahrmann, J., Grapov, D., Khoomrung, S. (2020). Deep metabolome: applications of deep learning in metabolomics. Comput. Struct. Biotechnol. J. 18, 2818–2825. doi: 10.1016/j.csbj.2020.09.033
Qi, S., Tu, Z., Ouyang, X., Wang, L., Peng, W., Cai, A., et al. (2012). Comparison of the metabolic profiling of hepatitis b virus-infected cirrhosis and alcoholic cirrhosis patients by using (1) h nmr-based metabonomics. Hepatol. Res. 42 (7), 677–685. doi: 10.1111/j.1872-034X.2011.00964.x
Roberts, L. D., Souza, A. L., Gerszten, R. E., Clish, C. B. (2012). Targeted metabolomics. Curr. Protoc. Mol. Biol. Chapter 30, Unit30.2–30.2.24. doi: 10.1002/0471142727.mb3002s98
Roca, M., Alcoriza, M. I., Garcia-Cañaveras, J. C., Lahoz, A. (2021). Reviewing the metabolome coverage provided by lc-Ms: focus on sample preparation and chromatography-a tutorial. Anal chimica Acta 1147, 38–55. doi: 10.1016/j.aca.2020.12.025
Roe, B., Kensicki, E., Mohney, R., Hall, W. W. (2011). Metabolomic profile of hepatitis c virus-infected hepatocytes. PloS One 6 (8), e23641. doi: 10.1371/journal.pone.0023641
Saito, T., Sugimoto, M., Igarashi, K., Saito, K., Shao, L., Katsumi, T., et al. (2013). Dynamics of serum metabolites in patients with chronic hepatitis c receiving pegylated interferon plus ribavirin: a metabolomics analysis. Metab: Clin. Exp. 62 (11), 1577–1586. doi: 10.1016/j.metabol.2013.07.002
Salgüero, S., Rojo, D., Berenguer, J., González-García, J., Fernández-Rodríguez, A., Brochado-Kith, O., et al (2020). Plasma metabolomic fingerprint of advanced cirrhosis stages among Hiv/Hcv-coinfected and hcv-monoinfected patients. Liver International : Official Journal of the International Association for the Study of the Liver 40 (9), 2215–2227. doi: 10.1111/liv.14580
Sarfaraz, M. O., Myers, R. P., Coffin, C. S., Gao, Z. H., Shaheen, A. A., Crotty, P. M., et al. (2016). A quantitative metabolomics profiling approach for the noninvasive assessment of liver histology in patients with chronic hepatitis c. Clin. Trans. Med. 5 (1), 33. doi: 10.1186/s40169-016-0109-2
Schmidt, D. R., Patel, R., Kirsch, D. G., Lewis, C. A., Vander Heiden, M. G., Locasale, J. W. (2021). Metabolomics in cancer research and emerging applications in clinical oncology. CA: Cancer J. Clin. 71 (4), 333–358. doi: 10.3322/caac.21670
Semmo, N., Weber, T., Idle, J. R., Beyoğlu, D. (2015). Metabolomics reveals that aldose reductase activity due to Akr1b10 is upregulated in hepatitis c virus infection. J. Viral hepa 22 (7), 617–624. doi: 10.1111/jvh.12376
Shanmuganathan, M., Sarfaraz, M. O., Kroezen, Z., Philbrick, H., Poon, R., Don-Wauchope, A., et al. (2021). A cross-platform metabolomics comparison identifies serum metabolite signatures of liver fibrosis progression in chronic hepatitis c patients. Front. Mol. Biosci. 8. doi: 10.3389/fmolb.2021.676349
Shariff, M. I., Gomaa, A. I., Cox, I. J., Patel, M., Williams, H. R., Crossey, M. M., et al. (2011). Urinary metabolic biomarkers of hepatocellular carcinoma in an Egyptian population: a validation study. J. Proteome Res. 10 (4), 1828–1836. doi: 10.1021/pr101096f
Shen, Y., Wu, S. D., Chen, Y., Li, X. Y., Zhu, Q., Nakayama, K., et al. (2023). Alterations in gut microbiome and metabolomics in chronic hepatitis B infection-associated liver disease and their impact on peripheral immune response. Gut Microbes 15 (1), 2155018. doi: 10.1080/19490976.2022.2155018
Sheng, G. (2008). Metabolomics studies of liver diseases based on Gc/Ms and Lc/Ms techniques (Zhejiang: Zhejiang University Health Science Center, Zhejiang University, School of Medicine, Zhejiang University).
Simillion, C., Semmo, N., Idle, J. R., Beyoğlu, D. (2017). Robust regression analysis of gcms data reveals differential rewiring of metabolic networks in Hepatitis B and C patients. Metabolites 7 (4), 51. doi: 10.3390/metabo7040051
Spina, R., Saliba, S., Dupire, F., Ptak, A., Hehn, A., Piutti, S., et al. (2021). Molecular identification of endophytic bacteria in leucojum aestivum in vitro culture, nmr-based metabolomics study and lc-Ms analysis leading to potential amaryllidaceae alkaloid production. International Journal of Molecular Sciences 22 (4), 1773. doi: 10.3390/ijms22041773
Sterling, R. K., Lissen, E., Clumeck, N., Sola, R., Correa, M. C., Montaner, J., et al. (2006). Development of a simple noninvasive index to predict significant fibrosis in patients with Hiv/Hcv coinfection. Hepatol. (Baltimore Md) 43 (6), 1317–1325. doi: 10.1002/hep.21178
Su, R. Y., Ling, S. B., Shan, Q. N., Wei, X. Y., Wang, R., Jia, C. K., et al. (2022). Efficacy and safety of sirolimus early conversion protocol in liver transplant patients with hepatocellular carcinoma: a single-arm, multicenter, prospective study. Hepatobil pancreatic Dis. international: HBPD Int. 21 (2), 106–112. doi: 10.1016/j.hbpd.2021.09.001
Trainor, P. J., DeFilippis, A. P., Rai, S. N. (2017). Evaluation of classifier performance for multiclass phenotype discrimination in untargeted metabolomics. Metabolites 7 (2), 30. doi: 10.3390/metabo7020030
Tsubota, A., Hirose, Y., Izumi, N., Kumada, H. (2003). Pharmacokinetics of ribavirin in combined interferon-alpha 2b and ribavirin therapy for chronic hepatitis c virus infection. Br. J. Clin. Pharmacol. 55 (4), 360–367. doi: 10.1046/j.1365-2125.2003.01780.x
Wai, C. T., Greenson, J. K., Fontana, R. J., Kalbfleisch, J. D., Marrero, J. A., Conjeevaram, H. S., et al. (2003). A simple noninvasive index can predict both significant fibrosis and cirrhosis in patients with chronic hepatitis c. Hepatol. (Baltimore Md) 38 (2), 518–526. doi: 10.1053/jhep.2003.50346
Wan, Q., Wang, Y., Tang, H. (2017). Quantitative (13)C traces of glucose fate in hepatitis b virus-infected hepatocytes. Anal Chem. 89 (6), 3293–3299. doi: 10.1021/acs.analchem.6b03200
Wang, J., Jiang, D., Zhang, H., Lv, S., Rao, H., Fei, R., et al. (2009). Proteome responses to stable hepatitis b virus transfection and following interferon alpha treatment in human liver cell line Hepg2. Proteomics 9 (6), 1672–1682. doi: 10.1002/pmic.200800621
Wang, X., Wang, X., Xie, G., Zhou, M., Yu, H., Lin, Y., et al. (2012). Urinary metabolite variation is associated with pathological progression of the post-hepatitis b cirrhosis patients. J. Proteome Res. 11 (7), 3838–3847. doi: 10.1021/pr300337s
Wang, X. J., Yan, G. L., Zhang, A. H., Sun, H., Piao, C. Y., Li, W. Y., et al. (2013). Metabolomics and proteomics approaches to characterize and assess proteins of bear bile powder for hepatitis c virus. Chin. J. Natural Medicines 11 (6), 653–665. doi: 10.1016/s1875-5364(13)60076-x
Wolfe, W., Xiang, Z., Yu, X., Li, P., Chen, H., Yao, M., et al. (2023). The challenge of applications of probiotics in gastrointestinal diseases. Adv Gut Microbiom Res. 6 (1), 94. doi: 10.1038/s41392-020-00443-w
Wu, J., Guo, Y., Lu, X., Huang, F., Lv, F., Wei, D., et al. (2020). Th1/Th2 cells and associated cytokines in acute hepatitis e and related acute liver failure. Journal of Immunology Research 2020, 6027361. doi: 10.1155/2020/6027361
Wu, J., Guo, N., Zhang, X., Xiong, C., Liu, J., Xu, Y., et al. (2019). Hev-Lf(S): a novel scoring model for patients with hepatitis e virus-related liver failure 26 (11), 1334–1343. doi: 10.1111/jvh.13174
Wu, J., Huang, F., Ling, Z., Liu, S., Liu, J., Fan, J. (2020). Altered faecal microbiota on the expression of Th cells responses in the exacerbation of patients with hepatitis e infection. Journal of Viral Hepatitis 27 (11), 1243–1252. doi: 10.1111/jvh.13344
Wu, J., Xiang, Z., Zhu, C., Yao, Y., Bortolanza, M., Cao, H., et al. (2021). Extrahepatic manifestations related to hepatitis e virus infection and their triggering mechanisms. J. infect 83 (3), 298–305. doi: 10.1016/j.jinf.2021.07.021
Wu, J., Xu, Y., Cui, Y., Bortolanza, M., Wang, M., Jiang, B., et al. (2022). Dynamic changes of serum metabolites associated with infection and severity of patients with acute hepatitis e infection. Journal of Medical Virology 94 (6), 2714–2726. doi: 10.1002/jmv.27669
Xiang, Z., Li, J., Lu, D., Wei, X., Xu, X. (2022). Advances in multi-omics research on viral hepatitis. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.987324
Xie, Q. (2016). Metabolic Profiling Analysis of Different HBV DNA levels inCHB patients Based on UPLC-MS (Dalian Medical University), Dalian.
Xue, R., Dong, L., Wu, H., Liu, T., Wang, J., Shen, X. (2009). Gas Chromatography/Mass spectrometry screening of serum metabolomic biomarkers in hepatitis b virus infected cirrhosis patients. Clin. Chem. Lab. Med. 47 (3), 305–310. doi: 10.1515/cclm.2009.083
Yamane, Y., Umeda, M., O’Uchi, T., Mitsushima, T., Nakata, K., Nagataki, S. (1994). Phosphorus-31 nuclear magnetic resonance in vivo spectroscopy of human liver during hepatitis a virus infection. Digest Dis. Sci. 39 (1), 33–38. doi: 10.1007/bf02090057
Yang, W. (2015). Serum metabolomics of hbv-related liver cirrhosis and hepatocellular carcinoma (Nanchang: Nanchang University).
Yang, L., Yang, X., Kong, X., Cao, Z., Zhang, Y., Hu, Y., et al. (2016). Covariation analysis of serumal and urinary metabolites suggests aberrant glycine and fatty acid metabolism in chronic hepatitis b. PloS One 11 (5), e0156166. doi: 10.1371/journal.pone.0156166
Yu, K., Sheng, G., Sheng, J., Chen, Y., Xu, W., Liu, X., et al. (2007). A metabonomic investigation on the biochemical perturbation in liver failure patients caused by hepatitis b virus. J. Proteome Res. 6 (7), 2413–2419. doi: 10.1021/pr060591d
Yue, Y., Meng, J., Zhang, S. (2002). Mechanism of peripheral blood mononuclear cell invasion by hbv on artificial immunization in newborns. Chin. Med. J. 115 (9), 1380–1382.
Zhang, L., Fan, Z., Kang, H., Wang, Y., Liu, S., Shan, Z. (2019). Analysis of serum metabolic profile of hepatitis b-related hepatocellular carcinoma by Hplc/Ms. J. South. Med. Univ. 39 (1), 49–56. doi: 10.12122/j.issn.1673-4254.2019.01.08
Zhang, L., Jia, X., Peng, X., Ou, Q., Zhang, Z., Qiu, C., et al. (2010). Development and validation of a liquid chromatography-mass spectrometry metabonomic platform in human plasma of liver failure caused by hepatitis b virus. Acta Biochim. Biophys. Sin. 42 (10), 688–698. doi: 10.1093/abbs/gmq078
Zhang, R., Lu, Z. (2023). Protective effects of clinacanthus nutans (Burm.F.) lindau aqueous extract on hbv mouse model by modulating gut microbiota and liver metabolomics. Evidence-based complementary and alternative medicine : eCAM 2023, 5625222. doi: 10.1155/2023/5625222
Zhang, L., Ma, Y., Zhang, l., Wang, Y., Wu, J., Huang, Y., et al. (2013). Analysis of serum metabolic profile and screening of characteristic metabolites in hbv-related hepatocellular carcinoma before and after surgery. Chin. J. Hepatobil Surg. 19 (2), 81–87. doi: 10.3760/cma.j.issn.1007-8118.2013.02.001
Zhang, A., Sun, H., Han, Y., Yan, G., Wang, X. (2013). Urinary metabolic biomarker and pathway study of hepatitis b virus infected patients based on uplc-Ms system. PloS One 8 (5), e64381. doi: 10.1371/journal.pone.0064381
Zhang, F., Wang, Y., Cheng, X., Wang, D., Li, Y., Liu, T., et al. (2022). Molecular mechanism of global action of active parts of chrysanthemum chrysanthemums against hepatitis b virus based on epigenetics and metabolomics. Acta Pharm. Sin. 57 (8), 2352–2363. doi: 10.16438/j.0513-4870.2022-0179
Zhao, Y., Gou, X. J., Dai, J. Y., Peng, J. H., Feng, Q., Sun, S. J., et al. (2013). Differences in metabolites of different tongue coatings in patients with chronic hepatitis b. Evidence-Based complement Altern. med: eCAM 2013, 204908. doi: 10.1155/2013/204908
Zhao, Y., Mao, Y. F., Tang, Y. S., Ni, M. Z., Liu, Q. H., Wang, Y., et al. (2018). Altered oral microbiota in chronic hepatitis b patients with different tongue coatings. World J. Gastroenterol. 24 (30), 3448–3461. doi: 10.3748/wjg.v24.i30.3448
Keywords: viral hepatitis, metabolomics, pathogenesis, biomarkers, treatment
Citation: Yang J, Wang D, Li Y, Wang H, Hu Q and Wang Y (2023) Metabolomics in viral hepatitis: advances and review. Front. Cell. Infect. Microbiol. 13:1189417. doi: 10.3389/fcimb.2023.1189417
Received: 19 March 2023; Accepted: 28 April 2023;
Published: 17 May 2023.
Edited by:
Min Yao, Nantong University, ChinaCopyright © 2023 Yang, Wang, Li, Wang, Hu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ying Wang, d2FuZ3k4MzAyMjRAc2luYS5jb20=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.