 Leonardo Cecotto1,2
Leonardo Cecotto1,2 Daphne A. C. Stapels2,3Kok P. M. van Kessel2Michiel Croes1Zeldali Lourens2H. Charles Vogely1
Daphne A. C. Stapels2,3Kok P. M. van Kessel2Michiel Croes1Zeldali Lourens2H. Charles Vogely1 Bart C. H. van der Wal1
Bart C. H. van der Wal1 Jos A. G. van Strijp2
Jos A. G. van Strijp2 Harrie Weinans1,4*
Harrie Weinans1,4* Saber Amin Yavari1,5
Saber Amin Yavari1,5- 1Department of Orthopedics, University Medical Center Utrecht, Utrecht, Netherlands
- 2Department of Medical Microbiology, University Medical Center Utrecht, Utrecht, Netherlands
- 3Infection Biology Group, Department of Biomolecular Health Sciences, Utrecht University, Utrecht, Netherlands
- 4Department of Biomechanical Engineering, Delft University of Technology, Delft, Netherlands
- 5Regenerative Medicine Centre Utrecht, Utrecht University, Utrecht, Netherlands
Background: Despite the extensive use of silver ions or nanoparticles in research related to preventing implant-associated infections (IAI), their use in clinical practice has been debated. This is because the strong antibacterial properties of silver are counterbalanced by adverse effects on host cells. One of the reasons for this may be the lack of comprehensive in vitro models that are capable of analyzing host-bacteria and host-host interactions.
Methods and results: In this study, we tested silver efficacy through multicellular in vitro models involving macrophages (immune system), mesenchymal stem cells (MSCs, bone cells), and S. aureus (pathogen). Our model showed to be capable of identifying each element of culture as well as tracking the intracellular survival of bacteria. Furthermore, the model enabled to find a therapeutic window for silver ions (AgNO3) and silver nanoparticles (AgNPs) where the viability of host cells was not compromised, and the antibacterial properties of silver were maintained. While AgNO3 between 0.00017 and 0.017 µg/mL retained antibacterial properties, host cell viability was not affected. The multicellular model, however, demonstrated that those concentrations had no effect on the survival of S. aureus, inside or outside host cells. Similarly, treatment with 20 nm AgNPs did not influence the phagocytic and killing capacity of macrophages or prevent S. aureus from invading MSCs. Moreover, exposure to 100 nm AgNPs elicited an inflammatory response by host cells as detected by the increased production of TNF-α and IL-6. This was visible only when macrophages and MSCs were cultured together.
Conclusions: Multicellular in vitro models such as the one used here that simulate complex in vivo scenarios can be used to screen other therapeutic compounds or antibacterial biomaterials without the need to use animals.
1 Introduction
Implant-related infections are one of the most frequent and severe complications associated with the use of biomaterials (Arciola et al., 2018). Despite best practice in medical and surgical management, infection occurs in approximately 5% of all operated orthopedic patients (Moriarty et al., 2016). The majority of orthopedic implant-associated infections (IAI) are caused by Staphylococci, in particular by Staphyloccocus aureus (Campoccia et al., 2006; Masters et al., 2022). This pathogen evolved multiple strategies to evade recognition and killing by our immune system and has developed resistance to commonly used antibiotics (Perret et al., 2012; Flannagan et al., 2015; de Jong et al., 2019; de Vor et al., 2020). Moreover, its ability to invade and survive within different host cells and tissues (Fraunholz and Sinha, 2012; Strobel et al., 2016; Horn et al., 2018) and infiltrate within the osteocyte canaliculi network (de Mesy Bentley et al., 2017) further complicates treatment. Altogether, these features make S. aureus one of the most challenging causes of IAI to treat with traditional antibacterial therapies.
Metallic silver (Ag) is a known, broad-spectrum antibacterial agent that was already used to treat infections before the introduction of antibiotics (Chopra, 2007). Over the years, numerous coating techniques have been developed to couple the antibacterial properties of silver to orthopedic implants (Amin Yavari et al., 2020; Djošić et al., 2021). Such Ag-coated implants have already been implemented in the clinic to decrease the incidence of infection after primary and revision surgeries, especially in oncologic patients who are susceptible to infections (Hardes et al., 2010; Wafa et al., 2015; Trovarelli et al., 2019; Diez-Escudero and Hailer, 2021; Fiore et al., 2021). However, the favorable antibacterial properties of silver are counterbalanced by adverse effects such as the skin blue coloring called argyria (Diez-Escudero and Hailer, 2021). Although most clinical studies do not report significant side effects, there is no consensus around the concentration at which silver may cause serious local or systemic damage (Eto et al., 2016; Diez-Escudero and Hailer, 2021; Fiore et al., 2021).
At a cellular level, many in vitro studies showed that silver exposure, both in free-ion (AgNO3) and nanoparticle (AgNP) form, correlates with DNA damage, increased production of inflammatory stimuli and reactive oxygen species, eventually leading to cell death (AshaRani et al., 2009; Park et al., 2010). However, whether AgNP effects are derived by the physical interaction with the nanoparticles, or the ions released from it is still under debate (Xiu et al., 2012; Durán et al., 2016; Liao et al., 2019). Nonetheless, nanoparticles might release lower concentrations of silver ions compared to AgNO3 (McQuillan et al., 2012; Ivask et al., 2014; Barros et al., 2019), therefore reducing silver toxicity. Moreover, as nanoparticles efficacy is shape-, size-, charge-, dose-, and time-dependent (Liao et al., 2019; Hamad et al., 2020), multiple studies tried to identify a therapeutic window where silver could retain its antibacterial activity while losing its toxicity to the host. Specifically, the optimal condition should not affect bone tissue and immune cells, which are key players involved in the post-operative (bone) healing process and control of infection around the implant.
Although the only place to start, simple in vitro studies are limited in the scope of their findings. For example, previous studies have shown that osteoblast viability, mesenchymal stem cell (MSC) proliferation and osteogenic differentiation were not negatively affected after prolonged incubation with silver at concentrations that still retain antibacterial properties (Qing et al., 2018; Rajendran et al., 2019; Souter et al., 2020). However, some signs of cell activation and toxicity were described at high silver concentrations (Hackenberg et al., 2011; Samberg et al., 2012; Pauksch et al., 2014), and other studies showed the negative impact of silver on osteoblast survival and MSCs osteogenic differentiation (Albers et al., 2013; Sengstock et al., 2014). In contrast to bone tissue, the viability of cells from the innate immune system is highly affected by silver. Micromolar concentrations of AgNO3 are already toxic to neutrophils (Croes et al., 2018), and macrophages could withstand exposure to silver for up to 24 h only at sub-antibacterial concentrations (Sarkar et al., 2015a). However, even such short exposure to silver negatively affected the metabolic activity of macrophages (Carrola et al., 2020) and their phagocytotic and bacterial killing properties did not improve (Sarkar et al., 2015b; Aurore et al., 2018). Although these studies provide useful insights into silver action against single types of host cells, they were incapable of verifying interactions between host cells and bacteria, and between host cell types that occur in the complex in vivo situation. Consequently, silver-coated implants with promising cytotoxicity and antibacterial properties in vitro generated contrasting results when tested in animal models (Chen et al., 2015; Croes et al., 2018).
Since the current in vitro models are not able to mimic a physiological environment, they consequently cannot predict in vivo behavior very well (Moriarty et al., 2014). Therefore, forced by regulatory authorities, medical device companies tend to undertake a vast amount of animal testing (Grainger et al., 2013). Not surprisingly, many of these animal trials are not conclusive due to variability between animals, highly scattered read-outs, and dead animals pre-/post-operation (Veening-Griffioen et al., 2019).
In order to better predict effectivity of antibacterial coating of implants and limit animal testing, we designed and built a multifaceted in vitro setup, primarily composed of a combination of host bone cells, bacteria, and immune cells. This model can precisely mimic the in vivo arena, test antibacterial properties and identify undesired foreign body responses to the developed implants. In our model, we chose to test the effectivity of two different forms of silver – AgNO3 or AgNP – due to their debatable effects on host-cell viability and bacterial survival. In our model, we co-cultured human bone marrow-derived MSCs, human monocyte-derived macrophages, and S. aureus in search of a therapeutic window for AgNO3 or AgNP that are used in implant coating, where neither the viability of bone nor immune cells was compromised. We here show that silver has its limitations as a therapeutic agent. This in vitro multicellular culture system represents a valid screening tool to predict the effects of various antibacterial compounds in an environment similar to the in vivo scenario without involving any animal testing.
2 Materials and methods
2.1 Human monocyte-derived macrophage culture
Blood from healthy human donors was supplied by the Dutch blood bank (Sanquin, Amsterdam, The Netherlands). Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats using Ficoll-Paque (Pharmacia, Uppsala, Sweden) density centrifugation. Monocytes were positively selected by magnetic-activated cell sorting (MACS) with anti-CD14 labelled microbeads (Miltenyi Biotec, 130050201) according to the manufacturer’s instructions.
Isolated monocytes were seeded in a 24-wells plate at a density of 3 x 105 cells/well, except when stated otherwise. When monocytes were seeded on top of 13-mm diameter titanium disks (Alfa Aesar, 10385-HP), these were placed in the 24-wells plate before seeding. Monocytes were differentiated into macrophages by culturing for 7 days at 37°C, 5% CO2 in α-Minimum Essential Medium (α-MEM, Gibco Paisley, 22561021) supplemented with 10% (v/v) hyFBSclone fetal bovine serum (hyFBS, Biowest, HYCLSV30160), 100 U/mL penicillin- streptomycin (1% p/s, Gibco, 15140122), and 40 ng/mL human recombinant M-CSF (Peprotech, 300-25). Culture media was refreshed after 3-4 days.
2.2 Human mesenchymal stem cell culture
Mesenchymal stem cells (MSCs) were isolated from human bone marrow aspirates upon informed consent. The aspiration procedure was approved by the local medical research ethics committee, University Medical Center Utrecht, under the protocols METC 08-001/K and METC 07-125/C.
Aspirates were diluted in PBS, filtered through a 100 µm cell strainer and the mononuclear cell layer was collected after Ficoll-Paque density centrifugation. Approximately 2.5 x 105 mononuclear cells were plated per cm² in MSC expansion medium consisting of α-MEM supplemented with 10% (v/v) heat-inactivated FBS (FBS, Biowest, S181H), 1% p/s, 0,2 mM L-ascorbic acid-2-phosphate (ASAP, Sigma-Aldrich, A8960) and incubated at 37°C, 5% CO2. Cells starting from passage 3 were used in the experimental setups.
2.3 Macrophage-MSC co-culture
To build the macrophage-MSC co-culture the following steps were adopted. Monocytes were seeded in 24-wells plates at a density of 3 x 105 cells/well and differentiated into macrophages by culturing for 7 days. Meanwhile, MSCs were cultured to reach ~70% confluency until monocytes fully differentiated into macrophages. Then, MSCs were fluorescently labelled with celltrace violet (Invitrogen, C34557) diluted in Hanks balanced salt solution (HBSS) according to the manufacturer’s instructions for labelling adherent cells. After staining, MSCs were detached with 0,25% trypsin/EDTA (Gibco, 25200056) and re-seeded together with macrophages at a density of 1 x 105 cells/well according to the experimental setup. Cells were allowed to adhere for about ~24 h before readouts started or bacteria were added, as explained below in the section “Multicellular infection model to study intracellular survival of bacteria”.
Cell seeding densities and culture plate format were adjusted according to the experimental setup. For instance, when measuring cytokine production, monocytes and MSCs were combined in a 96-well plate at a density of 1.5 and 0.5 x 105 cells/well, respectively. Moreover, when the identification of single cell types was not relevant for the experimental setup, fluorescence labelling was omitted.
2.4 Bacterial culture
All experiments used GFP-labelled Staphylococcus aureus (kind gift from Prof. Simon Foster) and Staphylococcus epidermidis (kind gift from Prof. Leo Koenderman) were transformed with a GFP-expressing plasmid pCM29 to constitutively express GFP, as previously described (Boero et al., 2021). Bacteria were grown overnight in Todd-Hewitt broth (THB) with 10 ng/mL chloramphenicol to reach stationary phase.
2.5 Silver ions and nanoparticles
A solution of silver ions (AgNO3) was prepared by dissolving silver nitrate (Sigma-Aldrich, S6506) in ultrapure water. Commercially available 20 nm and 100 nm silver nanoparticles (AgNP) (Alfa Aesar, J67067 and J67099), were used. Before each experiment, both sizes of nanoparticles were pelleted by centrifugation for 30 min at 4°C with 17000 x g (20 nm AgNP) or 300 x g (100 nm AgNP). A stock solution of 80 µg/mL was prepared for each type of nanoparticle in ultrapure water. AgNP and AgNO3 were diluted to various concentrations in α-MEM with 10% FBS or in THB to test their effects on host or bacterial cells.
2.6 Effect of silver on cell viability and cytokine production
Monocytes and MSCs were seeded in a 96-wells plate at a density of 1.5 and 0.5 x 105 cells/well, respectively. The same numbers of cells were combined in the co-culture, where macrophages were combined with MSCs as previously described.
Macrophages, MSCs, and the co-culture were incubated with fresh media containing 10 ng/mL LPS O111:B4 from Escherichia coli (Sigma-Aldrich), or various concentrations of AgNO3, 20 nm AgNP, and 100 nm AgNP. After 24 h stimulation, the culture medium was harvested to measure cytokine production and cells were processed for viability evaluation. When measuring cell viability, the culture medium was replaced by α-MEM with 10% FBS and 10% Alamar Blue solution prepared by dissolving Resazurin sodium salt (Sigma-Aldrich, R7017) in PBS. Then cells were incubated at 37°C for 2-3 h, in the dark. Next, the supernatant was transferred to a new plate and fluorescence was measured at 530-10/580-10 nm (Ex/Em) with a Clariostar plate reader (BMG labtech). Background fluorescence values were subtracted, and metabolic activity was normalized to the control sample. Production of cytokine TNF-α and IL-6 was measured in the collected supernatant by ELISA (Duoset, R&D Systems, DY210 and DY217B), according to the manufacturer’s instructions. ELISA values are expressed as fold-change over non-stimulated controls. Samples were analyzed in triplicates and experiments repeated three times with different monocytes and MSCs donors.
2.7 Direct antimicrobial properties of silver
The direct antimicrobial properties of silver were determined by measuring OD (600nm), combined with the broth microdilution method. Overnight bacterial culture was diluted in THB to reach a final inoculum of 5 x 105 colony-forming units per mL (CFU/mL). In a flat-bottom 96-well plate, the bacterial suspension was mixed with AgNO3, 20 nm AgNPs, or 100 nm AgNP in equal parts in triplicates, with a final volume of 200 μL. Bacterial growth was monitored at 37°C by measuring OD (600nm) every 5 min, for a total time of 12 h, on a Clariostar plate reader with gentle shaking before each measurement. Then, bacterial suspensions were serially diluted and plated on Todd-Hewitt agar (THA), and colonies counted after overnight incubation at 37°C. Samples were analyzed in triplicates.
2.8 Multicellular infection model to study intracellular survival of bacteria
An overnight bacterial culture was diluted in α-MEM to reach a final concentration of 1 x 107 CFU/mL. To mimic the in vivo situation, bacteria were incubated with 5% normal human serum (NHS) for 15 min at 37°C, which coats them with antibodies and complement (opsonization) to enable their uptake by immune cells. Serum was collected from blood obtained from healthy donors after informed consent, as previously described (Boero et al., 2021). Approval from the Medical Ethics Committee of the University Medical Center Utrecht was obtained (METC protocol 07-125/C approved on March 1, 2010).
Opsonized bacteria were added to the co-culture at various multiplicity of infection (MOI). To synchronize bacterial uptake, plates were centrifuged for 5 min with 110 x g at RT, and then incubated at 37°C, 5% CO2. To study intracellular bacterial survival, the cells were washed twice after 30 min of infection to remove free bacteria and cultured in media supplemented with 100 µg/mL gentamicin (Serva, 22185.02, to kill the bacteria) and 20 µg/mL lysostaphin (Bioconnect, MBS635842, to lyse the bacteria and lose the GFP signal) for 1 h. Afterwards, cells were washed twice and incubated in media with only 5 µg/mL gentamicin. All washing steps were performed with warm α-MEM. This treatment allows only intracellular bacteria to survive, as both gentamicin and lysostaphin are unable to penetrate mammalian cell membranes within short time periods (Hamza et al., 2013; Hamza and Li, 2014). To verify treatment efficacy in lysing bacteria, serum-opsonized S. aureus was incubated for 1 h at 37°C in α-MEM in presence of 100 µg/mL gentamicin and 20 µg/mL lysostaphin. Then, GFP expression was measured with a MACSquant VYB (Miltenyi Biotech) flow cytometer and data were analyzed with FlowJo (v.10.1., FlowJo LLC).
At the desired time points, co-culture samples were processed for flow cytometry analysis, microscopy observation, or quantification of intracellular bacteria via CFU count.
For flow cytometry, cells were detached by a combination of trypsin and eventually gentle scraping in 1 mM DPBS/EDTA if cells were still attached to the bottom of the culture plate. Cells were transferred to a 96-wells plate and stained with sytox orange dead cell stain for flow cytometry (Invitrogen, S34861) according to the manufacturer’s instructions. Samples were measured with a MACSquant VYB flow cytometer and data were analyzed with FlowJo. The gating strategy is summarized in Supplementary Figure 1. Briefly, a total of 10000 events were collected for each sample gated based on forward scatter (FSC) and side scatter (SSC) parameters. The two cell types were selected based on the signal of CellTrace violet. Non-infected samples from the sytox negative population were used to set GFP fluorescence baseline and define the proportion of infected, GFP-positive cells.
For confocal imaging, cells were collected as described for flow cytometry analysis and fixed in 1.5% paraformaldehyde. Cell membranes were stained with 3 µg/mL Alexa Fluor 647-conjugated Wheat Germ Agglutinin (WGA, Invitrogen, W32466) for 10 min at RT, on a shaking plate. Then, samples were transferred to CELLview slide (Greiner Bio-One, 543079) previously coated with poly-L-lysine (Sigma-Aldrich, P4707), and imaged on a Leica TCS SP5 microscope with a HCX PL AP CS 63x/1.40-0.60 OIL objective (Leica Microsystems). Images were adjusted for publication using Image J Fiji.
Finally, to quantify the number of intracellular bacteria, cells were lysed with 0.1% Triton X-100 and plated on THA plates in serial dilutions. Plates were incubated overnight at 37°C after which colonies were counted.
2.9 Multicellular infection model to study the effects of silver
A variation of this model was used to study silver effects. Briefly, cells were incubated for 24 h with different concentrations of AgNO3 and 20 nm AgNP before adding S. aureus at a MOI=10. Samples were analyzed by flow cytometry and CFU counting at 30 min and 4 h after infection. In this setup, when cells were incubated with silver, gentamicin and lysostaphin treatment was not employed for the 4 h time point. Samples were analyzed in triplicates and experiments repeated three times with different monocytes and MSCs donors.
2.10 Statistical analysis and graphics
GraphPad Prism 9 (version 9.3) was used to create the graphs and determine statistical significance via a two-way or one-way ANOVA, or t-test. p<0.05 was considered statistically significant. Illustrations were created with BioRender.com.
3 Results
3.1 Establishing a tunable, multicellular, in vitro model to study IAI
To build a comprehensive, multicellular in vitro model that mimics the main players in IAI, we co-cultured primary MSC and monocyte-derived human macrophages as summarized in Figure 1. First, monocytes were isolated from human blood and seeded on a surface to differentiate into macrophages (Figures 1A, B). Cells were seeded either on a cell culture plate or on any biomaterial mimicking an implant. When comparing plastic with titanium, the surface itself seemed to have minimal impact on the phagocytic capacity of the macrophages (Supplementary Figure 2A). After differentiation, macrophages were re-seeded together with MSCs, which can differentiate into osteoblasts, to create an IAI-like environment (Figure 1C). In order to distinguish each element in the co-culture, we labeled at least one cell type before mixing with the second one. The co-culture was viable for up to several days, enabling the verification of cytotoxicity, modulation of cell functions, and antibacterial effects over a relatively long-time span.
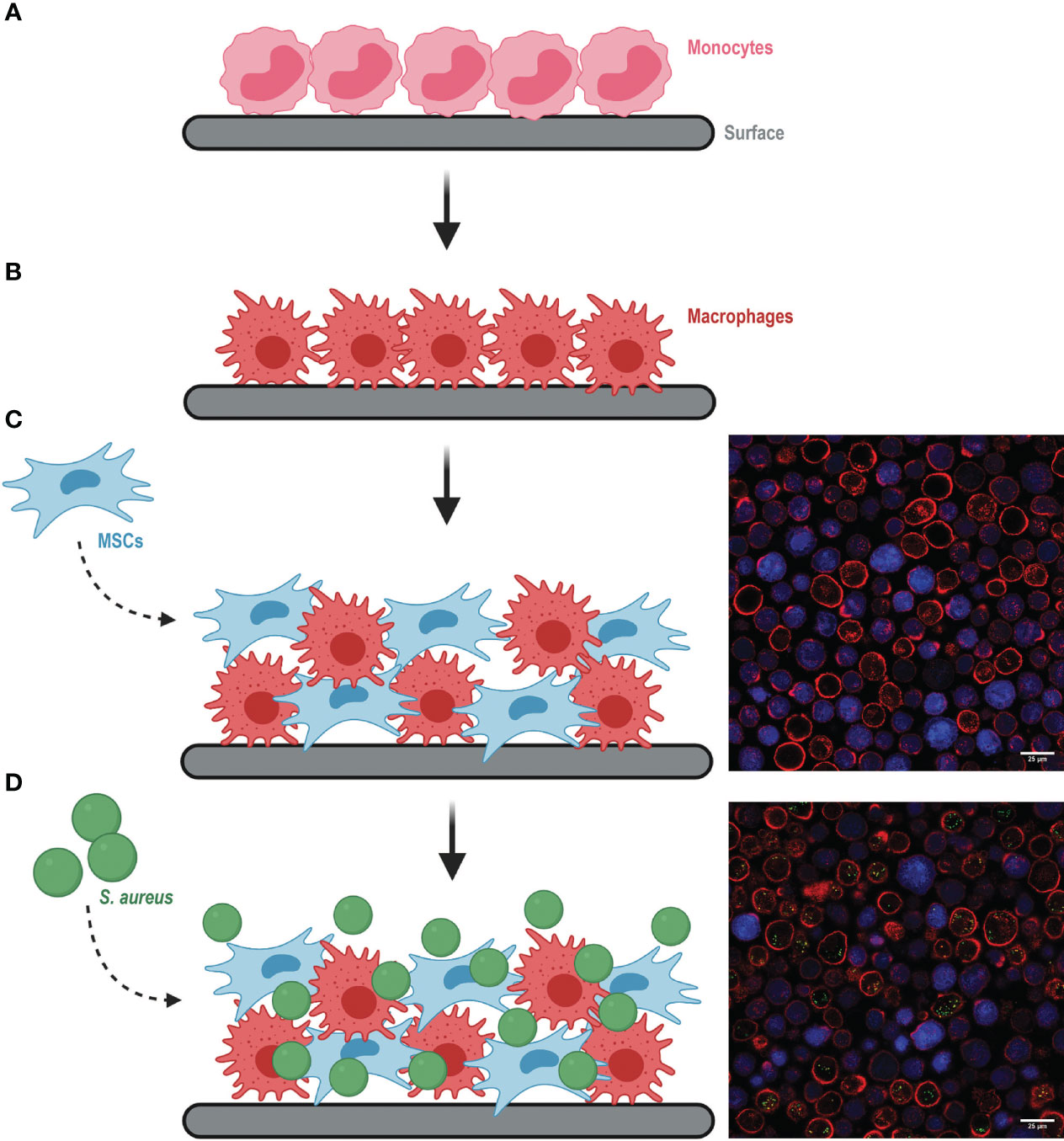
Figure 1 Schematic representation of building the multicellular in vitro model mimicking an implant-associated infection environment (images created with Biorender.com). (A) Monocytes were seeded on the selected surface and differentiated to macrophages (B) for 7 days. (C) Differentiated macrophages were re-seeded together with MSCs (left). Representative microscopy image of the co-culture (right), where MSCs were previously stained with CellTrace Violet (in blue) and all cell membranes were stained with Alexa Fluor 647-conjugated WGA (in red). (D) S. aureus was added to the co-culture (left). Representative image of the co-culture with intracellular S. aureus (green dots) (right).
The co-culture of macrophages and MSCs was then exposed to GFP-expressing bacteria (Figure 1D). We verified that this model could be used to study infections caused by different bacterial species relevant in the orthopedic field, such as S. aureus and S. epidermidis. Although macrophages were equally capable of associating with both bacterial species, we observed that MSCs were more susceptible to S. aureus than S. epidermidis infection (Supplementary Figure 2B). To assess the efficiency of antibacterial treatments in both macrophages and MSCs, all further experiments were conducted with S. aureus.
At the desired time points, the co-culture was processed for (confocal) microscopy (Figure 1), flow cytometry (Supplementary Figure 2), or bacterial enumeration via CFU plating. Flow cytometry offered the possibility to track intracellular bacterial survival per host-cell type over time by including gentamicin and lysostaphin treatment. These two compounds are not membrane-permeable and therefore selectively kill and lyse extracellular bacteria (Hamza et al., 2013; Hamza and Li, 2014). We confirmed by flow cytometry that the treatment efficiently lysed bacteria, as no GFP signal could be detected from S. aureus already after 1 h incubation with gentamicin and lysostaphin (Supplementary Figure 2C). Moreover, microscopy images confirmed that all GFP signal detected by the flow cytometer originated from intracellular bacteria and not from membrane-bound or extracellular S. aureus (Figure 1D). These results confirm the validity of the model we developed to study intracellular bacteria.
As expected, after 30 min of direct contact between bacteria and host cells, almost all macrophages had engulfed at least one S. aureus bacterium. In comparison, less than half of the MSCs population had engulfed bacteria. After 24 h, the proportion of infected macrophages was significantly reduced both when cultured alone, and in the co-culture. Interestingly, the proportion of infected MSCs was only reduced when cultured alone, but not in co-culture (Supplementary Figure 2D). This highlights distinct behaviors between cells cultured alone or together.
3.2 Finding a therapeutic window for silver treatment
We tested the validity of our co-culture model by using it to determine a possible therapeutic window for silver as an antibacterial treatment. For this purpose, silver must be at a concentration that is antibacterial, but not toxic to human cells. Therefore, we incubated S. aureus, macrophages, and MSCs with several concentrations of AgNO3, 20 nm AgNP, and 100 nm AgNP.
We observed that at the lowest concentration tested, AgNO3 reduced bacterial growth and significantly decreased the number of viable S. aureus,while complete inhibition of growth and killing was achieved after exposure to AgNO3 concentrations higher than 0.0017 µg/mL (Figures 2A, B). Although bacterial growth speed was affected by increasing the concentrations of 20 nm AgNP (Figure 2D), no bacteriostatic or bactericidal effects were observed after incubation with 20 nm AgNP and 100 nm AgNP (Figures 2D, E, G, H).
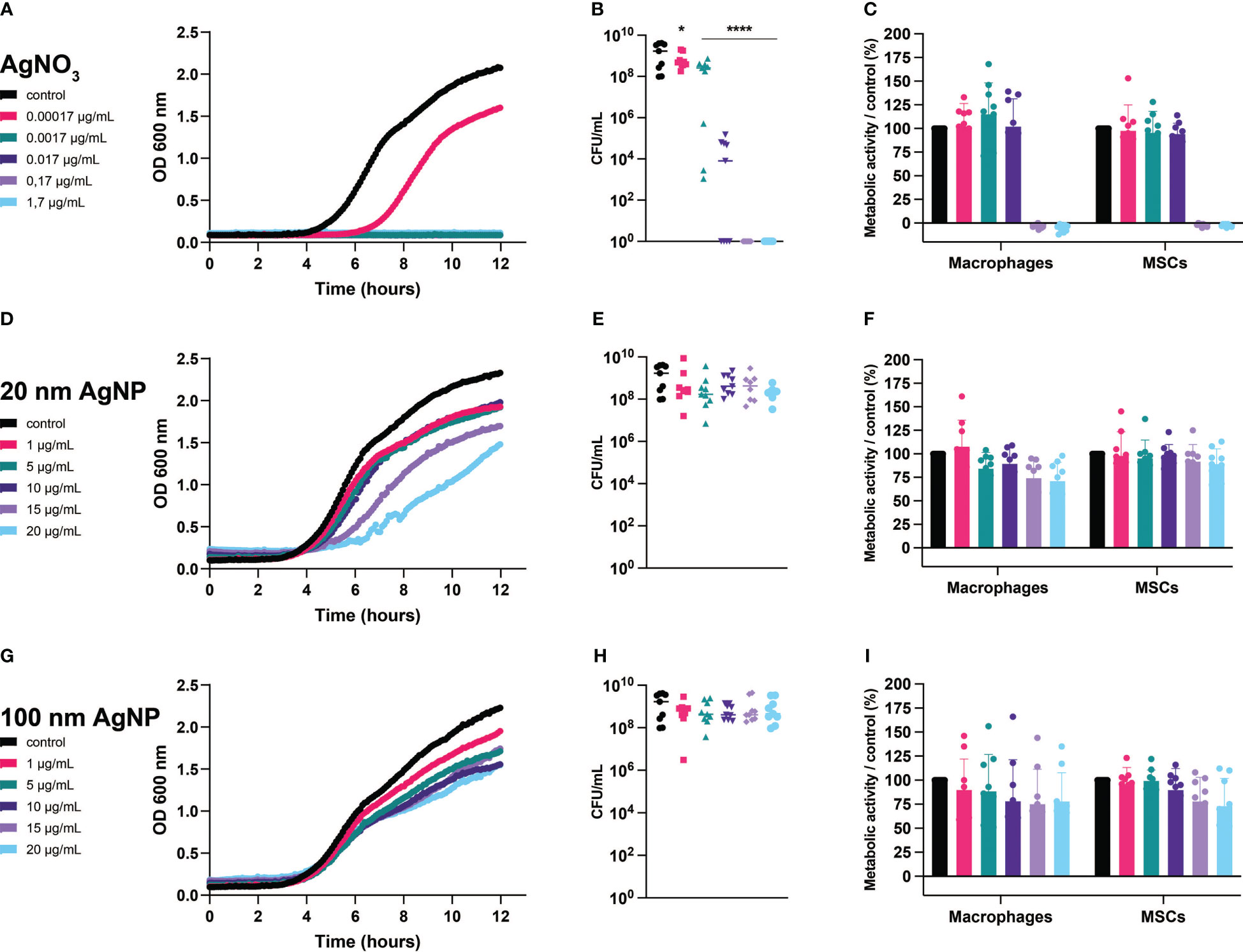
Figure 2 Evaluation of the effects of silver on S. aureus, macrophages, and MSCs. Bacteria and host cells were incubated with several concentrations of AgNO3 (A–C), 20 nm AgNP (D–F), and 100 nm AgNP (G–I). Bacterial growth (graphs on the left) was monitored by measuring OD (600 nm) continuously during 12 h and the number of surviving bacteria was quantified by CFU count (graphs on the center). (n=9, from 3 independent experiments). Data for bacterial growth were plotted with mean only, and data for CFU count were transformed into log10 and represented as mean +/- SD. Statistical significance compared to control was determined via one-way ANOVA. *p<0,05; ****p<0,0001 on the log-transformed data. The metabolic activity of macrophages and MSCs (graphs on the right) was determined by alamar blue assay after 24 h incubation with silver formulations. Assay values were normalized to untreated control cells. Assay values were normalized to untreated control cells. (n=9, from 3 independent experiments). Data were represented as mean +/- SD.
On the other hand, AgNO3 completely abolished the metabolic activity of host cells starting at 0.017 µg/mL (Figure 2C), while exposure to AgNP, regardless of size or concentration, partially affected the metabolism of both cell types. For instance, concentrations of 20 nm AgNP higher than 1 µg/mL reduced the total metabolic activity of the macrophage population by up to ~25% (Figure 2F). Incubation with any concentration of 100 nm AgNP reduced the viability of macrophages, while the metabolic activity of MSCs was affected only at concentrations higher than 10 µg/mL (Figure 2I). Nonetheless, we exclude a toxic effect derived from exposure to nanoparticles as more than 75% of the total metabolic activity remained for both the macrophages and MSCs populations.
In addition, we assessed whether silver triggered an inflammatory response by measuring the secretion of TNF-α and IL-6. Overall, non-toxic silver concentrations did not elicit any inflammatory response in monocultured cells compared to control samples (Figure 3). However, from this assay we could observe the added value of co-culturing different cell types. Addition of MSCs reduced LPS-mediated activation of macrophages, as observed by decreased production of TNF-α. In addition, including immune cells reduced IL-6 production by MSCs. Moreover, exposure to 100 nm AgNP at 20 µg/mL and at concentrations higher than 5 µg/mL significantly increased TNF-α (Figure 3C) and IL-6 (Figure 3F) levels respectively in the co-culture.
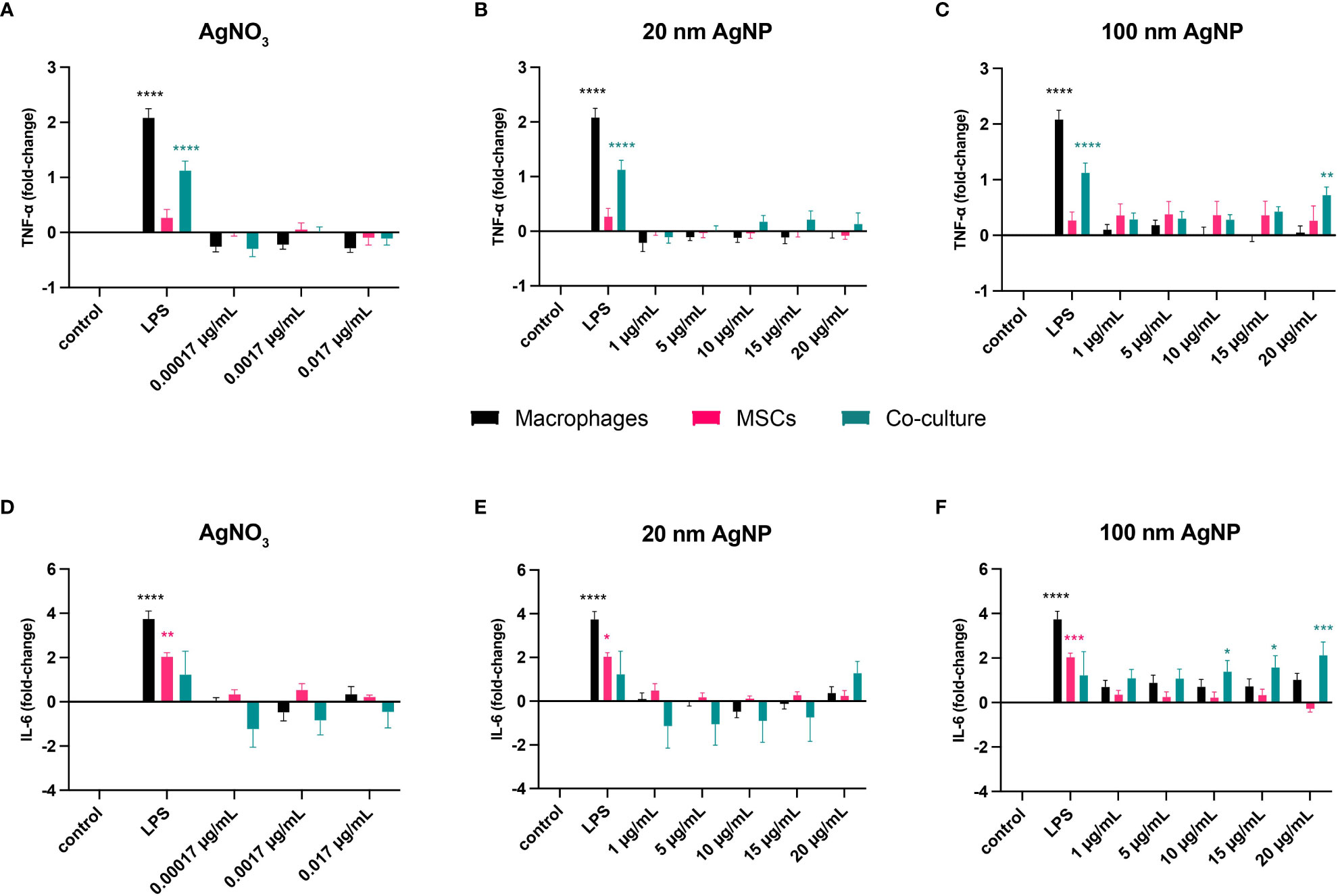
Figure 3 Evaluation of the inflammatory response caused by stimulation of host cells with non-toxic concentrations of silver. Macrophages, MSCs, and macrophages with MSCs (Co-culture) were incubated for 24 h with several concentrations of AgNO3, 20 nm AgNP, 100 nm AgNP, and 10 ng/mL LPS as positive control. Then, TNF-α (A–C) and IL-6 (D–F) levels in the supernatant were quantified by cytokine-specific ELISA and expressed as fold-change compared to control samples. (n=9, from 3 independent experiments). Data were transformed into 2-log and represented as mean +/- SEM. Statistical significance compared to control was determined via two-way ANOVA. ****p<0.0001; ***p<0.001; **p≤ 0.005; *p<0,05.
3.3 Evaluation of the effects of silver on the multicellular in vitro model
Once we established the effects of silver in simple in vitro models, we could further explore the consequences of its use in an IAI-like environment thanks to our multicellular model to study infection. Besides defining the antibacterial, non-toxic, and non-inflammatory concentrations of AgNO3 and AgNPs, we explored the impact of silver on the antibacterial functions of host cells (Figure 4A).
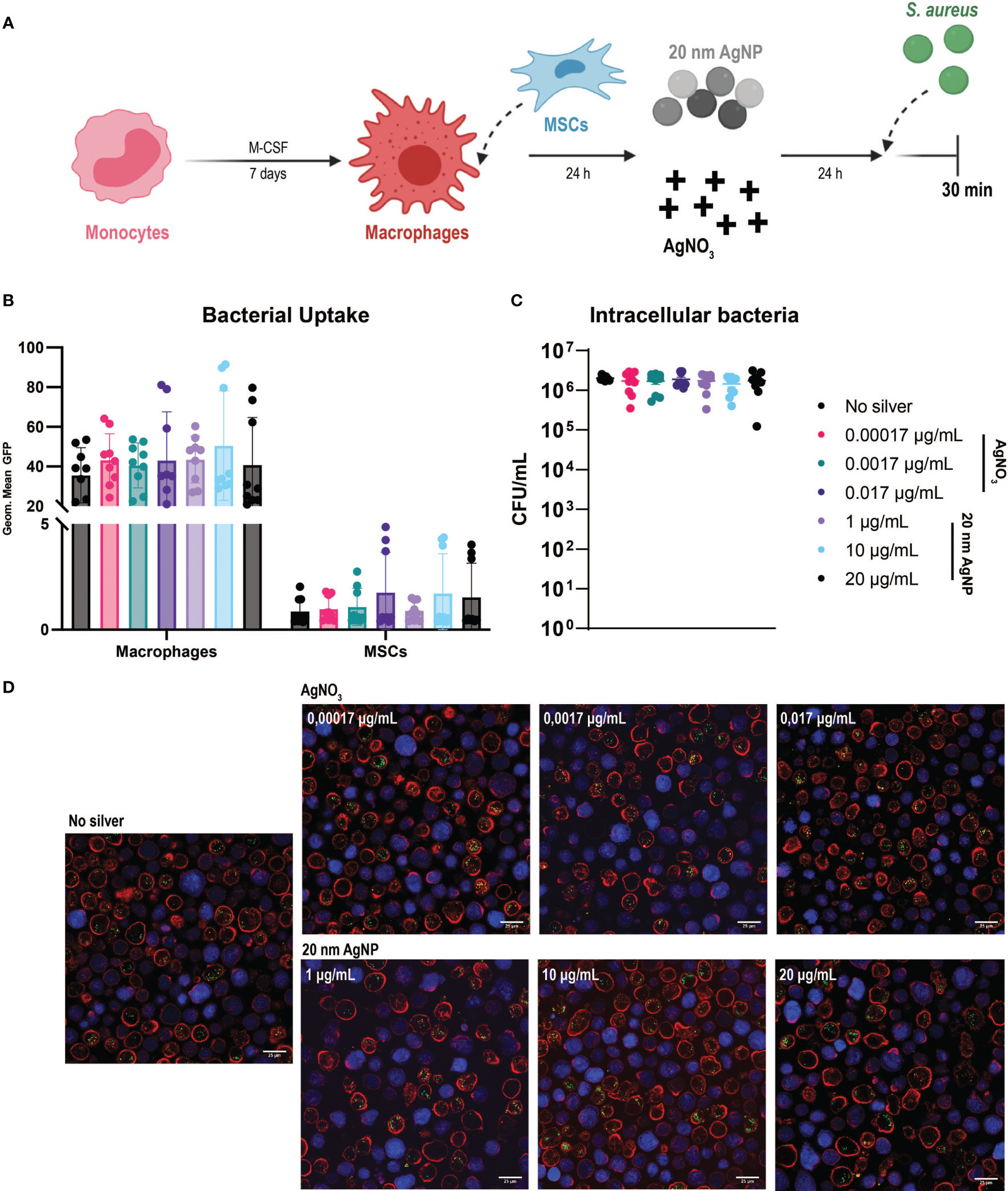
Figure 4 Evaluation of the effects of AgNO3 and 20 nm AgNP on the multicellular in vitro model (A) Schematic representation of the protocol used (images created with Biorender.com). Macrophages were combined with MSCs in culture and then incubated with AgNO3 and 20 nm AgNP for 24 h before infection with S. aureus for 30 min. (B) The amount of bacteria taken up by macrophages and MSCs is represented as the geometric mean of the GFP signal. (C) The number of intracellular bacteria was determined by counting CFU after cells lysis. (n = 9, from a total of 3 independent experiments). Data were represented as mean +/- SD. (D) Confocal imaging confirms flow cytometry observations. Macrophages (red), MSCs (blue), and S. aureus (green dots).
Due to their toxicity to host cells, we excluded AgNO3 at concentrations of 0.17 µg/mL and 1.7 µg/mL from the assays. Only three representative concentrations of 20 nm AgNP were selected, as similar cytotoxicity and antibacterial activity were observed. Analysis of samples by flow cytometry showed a remarkable shift in the side-scatter (SSC) values for macrophages in the co-culture when exposed to AgNP for 24 h (Supplementary Figure 3A). This increase in cellular granularity was concentration dependent and particularly evident in the presence of 100 nm AgNP. The effect was probably caused by the internalization of nanoparticles by macrophages, as shown by bright field microscopy (Supplementary Figure 3B). For this reason, 100 nm AgNPs were excluded from the analysis with the multicellular model.
Despite the previously observed antibacterial activity of AgNO3 (Figures 2A, B), this was not strong enough to counteract bacterial growth within the time frame tested in the multicellular model. In spite of the presence of AgNO3 or 20 nm AgNP, after 4 h incubation, the bacteria had already overtaken and killed most macrophages and MSCs in culture as observed by the increased amount of cells positive for sytox staining (Supplementary Figure 4).
Following S. aureus introduction into the co-culture, silver treatment did not influence bacterial uptake by host cells. We confirmed the presence of intracellular S. aureus in both cell types. As observed before, macrophages phagocytosed more bacteria than MSCs (Figure 4B). Depending on the donor, silver treatment had either no impact on phagocytosis or eventually led to an increased uptake of bacteria by both cell types. Moreover, exposure to silver did not reduce the amount of S. aureus able to survive intracellularly compared to the control samples (Figure 4C). Confocal microscopy imaging confirmed the flow cytometry results. Higher numbers of intracellular S. aureus were observed within macrophages than MSCs, while exposure to AgNO3 or AgNP did not influence bacterial uptake (Figure 4D).
4 Discussion
Despite the long-known antibacterial efficacy of silver, its clinical use has lagged. One cause might be ascribed to the lack of reliable in vitro models to test silver’s efficacy. Another limitation might be related to the need to identify a therapeutic window where pathogens are killed without affecting the viability of host cells. In this work, we developed a multicellular in vitro model that mimics an IAI-like scenario to investigate the efficacy of silver as a therapeutic treatment.
Traditionally, the effect of silver on bacteria or host cells was investigated separately, ignoring cell-cell interactions. This often rushed towards the identification of silver concentrations that were antibacterial and non-toxic for host cells (Priebe et al., 2017; Huang et al., 2022; Kuo et al., 2022; Selvamani et al., 2022) without addressing interactions between bacteria and host cells, and between host cell types. Although they did not address the antibacterial properties of silver, other research groups have shown that the effectiveness of novel treatments was different when assessed on cells cultured alone or in the presence of bacteria (Zaatreh et al., 2016; Luan et al., 2020). Similarly, encouraging results have been achieved after tests run on monocultures in vitro, but generated opposite outcomes when these treatments were tested in animal models (Chen et al., 2015; Tuchscherr et al., 2016). In a previous study, we showed that AgNP-coated surfaces completely killed S. aureus in vitro, while they did not exert any bactericidal effect in vivo. Moreover, the presence of coated implants worsened the healing process. Afterward, we learnt that these side effects derived from previously undetected Ag toxic levels against immune cells (Croes et al., 2018). Therefore, to improve the reliability of our in vitro tests, we designed a comprehensive multicellular model where cell-cell interactions can be investigated.
The interaction between MSCs and macrophages have already been showed to improve osteogenic differentiation of MSCs (Nicolaidou et al., 2012; Loi et al., 2016), resolution of infection (Krasnodembskaya et al., 2012; Johnson et al., 2017a; Chow et al., 2020) and inflammation (Lu et al., 2021). We also assessed the additive value of culturing macrophages and MSCs together. In fact, in the presence of LPS, cells in co-culture secreted a lower amount of cytokines compared to single cells (Figure 3). On the other hand, although immune cells captured most of the invading pathogens, a small fraction of MSCs was still infiltrated by S. aureus, via a process that could have been either active or passive (Fraunholz and Sinha, 2012; Josse et al., 2015). In co-culture, 24 h after infection, we observed a reduction of intracellular bacteria only within macrophages, with no changes in MSCs (Supplementary Figure 2D). While this could be explained in immune cells by the activation of their bactericidal activity (Pidwill et al., 2021), further studies are needed to understand the antibacterial mechanisms activated in MSCs when seeded alone but not in co-culture. Nonetheless, we have shown that our multicellular model allows us to identify each element involved in culture as well as track the intracellular survival of bacteria over time. As cells could also be seeded on different surfaces (Supplementary Figure 2A), this model can be extended to study the impact of new biomaterials and/or coatings on host cells and bacteria. Here, we used the model as a preliminary screening tool for finding the optimal silver concentrations to include in antibacterial coatings for orthopedic implants.
There are several reasons to favor silver formulations as nanoparticles rather than free ions. Use of higher-sized nanoparticles with a larger surface area is thought to reduce the amount of silver ions released, and therefore reduce AgNPs toxicity against host cells (McQuillan et al., 2012; Ivask et al., 2014; Barros et al., 2019). Accordingly, nanoparticles with larger sizes need to be used at higher concentrations to achieve similar efficacy to AgNO3 (Albers et al., 2013; Lu et al., 2013; Pazos-Ortiz et al., 2017; Qais et al., 2019). Besides a reduction in cytotoxicity, it should be considered that continuous uptake of non-degradable nanoparticles by host cells eventually leads to alterations in cell shape and morphology, as shown by our flow cytometry analysis (Supplementary Figure 3). Moreover, AshaRani et al., showed that nanoparticle uptake in vitro did not correlate with substantial cell death, even after incubation with higher concentrations of AgNP with a size distribution lower than 20 nm, but forced macrophages into a state of metabolic arrest (AshaRani et al., 2009). Instead, nanoparticle dimensions affected in vivo clearance and tissue accumulation, with the risk that particles larger than 40 nm reside indefinitely within the body (Gustafson et al., 2015). Furthermore, continuous exposure to nanoparticles might cause the onset of a local inflammatory response in the long term with detrimental consequences for implant-tissue integration (Zreiqat et al., 2003). According to our results, AgNPs were less toxic to host cells and bacteria than AgNO3 (Figure 2). Although we observed only a slight reduction in metabolically active macrophages and MSCs after exposure to AgNPs, previous studies suggest that use of higher concentrations might have induced cytotoxic effects on host cells as well (Arai et al., 2015; Sarkar et al., 2015a; Aurore et al., 2018; Carrola et al., 2020). For instance, the onset of harmful effects on host cells was evident from the inflammatory response caused by 100 nm AgNP (Figures 3C, F). Interestingly, this side effect was detected only with the use of the co-culture model rather than with single cell assays. Furthermore, treatment with neither 20 nm AgNP or AgNO3 reduced the number of intracellular bacteria found in macrophages and MSCs (Figure 4).
The multicellular model we used only allowed us to assess silver’s efficacy for a limited period of time due to the fact that bacteria grow faster than host cells. While AgNO3 displayed bacteriostatic and bactericidal effects after 12 h (Figures 2A, B), the same concentrations failed to inhibit S. aureus growth after 4 h in the multicellular model (Supplementary Figure 4). This limited our study to bacteria that survive intracellularly, while this multicellular model might be used to assess the efficacy of treatment on both extracellular and intracellular bacteria. The observed change in the efficacy of silver might be caused by the assay conditions in vitro. For instance, aerobic or anaerobic culture conditions (Yang et al., 2013), aggregation of nanoparticles (Brennan et al., 2015) or binding of silver ions to serum proteins (Gosheger et al., 2004; Shahabadi et al., 2012) could negatively impact the bio-functionality of silver. Our study might be affected by this, since different types of media were used to assess silver toxicity against S. aureus, macrophages, and MSCs. Moreover, silver exposure may even affect the response of macrophages to invading pathogens. For instance, Sarkar et al., showed that AgNP-treated macrophages had a reduced cytokine response and activation following Mycobacterium tuberculosis infection (Sarkar et al., 2015a). On the other hand, other research groups did not observe any positive impact on phagocytosis or oxidative burst in innate immune cells after treatment with silver (Liu et al., 2013; Haase et al., 2014; Huang et al., 2021).
Apart from testing the efficacy of silver as an antibacterial agent, this model could be used as an in vitro screening tool for several other therapeutic compounds (Cecotto et al., 2022) or antibacterial coatings. By developing culture models in which host-host and host-bacteria cells interact, we may gain greater insight into how implant surface features influence these cells, which has been investigated only on single elements so far (Leach and Whitehead, 2018; Liu and Segura, 2020; Zhu et al., 2021; Monteiro et al., 2022; Tan et al., 2022). In fact, this model could be adapted to study different scenarios of IAI. In accordance with the race for the surface theory (Gristina et al., 1989), bacterial colonization of the implant surface may prevent host cells adhesion and subsequently implant-tissue integration. Accordingly, Luan et al., showed that variations in gold nanoparticle-coatings were able to modulate macrophages functions and bone cells adhesion according to the presence or absence of bacteria on the coated surface (Luan et al., 2020). However, these and other studies, including ours, were performed under static conditions in vitro. By adding a flow system that simulates in vivo-like conditions such as shear stress and exchange of nutrients and molecules among cells in culture, outcomes can be closer to a real IAI scenario (Subbiahdoss et al., 2011; Zhao et al., 2014; da Silva Domingues et al., 2015). Finally, these multicellular models could include different cell types involved in IAI, such as osteocytes, osteoclasts, or cells from the bone marrow. Neutrophils and macrophages, for example, form the first line of defense against invading pathogens (Rosales, 2018; Rosowski, 2020; Pidwill et al., 2021). Despite previous reports about neutrophil interactions with single cell types and different biomaterials (Silva, 2010; Brandau et al., 2014; Jhunjhunwala, 2018; Abaricia et al., 2020; Wesdorp et al., 2023), little is known about their role in complex in vitro models that mimic the conditions of IAI.
5 Conclusion
Thanks to our multicellular model combining two host-cell types, bacteria, and an implant surface, we can predict the possible benefits and pitfalls derived from the use of silver as an antibacterial agent. Although our monoculture assays suggested the existence of a therapeutic window for the use of AgNO3 between 0.00017 and 0.017 µg/mL, our multicellular model revealed that neither the extracellular nor intracellular survival of S. aureus was affected. Moreover, we found that uptake of 20 nm AgNP had no impact on metabolic activity, phagocytic and killing capacities of macrophages, whereas increasing the size of the nanoparticles caused an inflammatory response that was only detectable when macrophages and MSCs were cultured together. By developing a model that accounts for the interactions among host cells and bacteria, in vitro screening tests can better simulate the complexity of in vivo models and better predict treatment outcomes.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
LC, DS, HW and SA contributed to the conception and design of the study. LC, DS, ZL performed the experiments. LC, DS, and SA wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
The collaboration project is cofounded by the PPP Allowance made available by Health~Holland, Top Sector Life Sciences & Health, to stimulate public-private partnerships. This research also received co-funding from the EU’s H2020 research and innovation program under Marie S. Curie cofund RESCUE grant agreement No 801540. This publication is also part of the project DARTBAC (with project number NWA.1292.19.354 of the research program NWA-ORC which is (partly) financed by the Dutch Research Council (NWO).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1186936/full#supplementary-material
References
Abaricia, J. O., Shah, A. H., Musselman, R. M., Olivares-Navarrete, R. (2020). Hydrophilic titanium surfaces reduce neutrophil inflammatory response and NETosis. Biomater Sci. 8, 2289–2299. doi: 10.1039/c9bm01474h
Albers, C. E., Hofstetter, W., Siebenrock, K. A., Landmann, R., Klenke, F. M. (2013). In vitro cytotoxicity of silver nanoparticles on osteoblasts and osteoclasts at antibacterial concentrations. Nanotoxicology 7, 30–36. doi: 10.3109/17435390.2011.626538
Amin Yavari, S., Castenmiller, S. M., van Strijp, J. A. G., Croes, M. (2020). Combating implant infections: shifting focus from bacteria to host. Advanced Materials 32, 1–25. doi: 10.1002/adma.202002962
Arai, Y., Miyayama, T., Hirano, S. (2015). Difference in the toxicity mechanism between ion and nanoparticle forms of silver in the mouse lung and in macrophages. Toxicology 328, 84–92. doi: 10.1016/J.TOX.2014.12.014
Arciola, C. R., Campoccia, D., Montanaro, L. (2018). Implant infections: adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 16, 397–409. doi: 10.1038/s41579-018-0019-y
AshaRani, P., Low Kah Mun, G., Hande, M., Valiyaveettil, S. (2009). Cytotoxicity and genotoxicity of silver nanomaterials. ACS Nano. 3 (2), 279–290. doi: 10.1021/nn800596w
Aurore, V., Caldana, F., Blanchard, M., Kharoubi Hess, S., Lannes, N., Mantel, P. Y., et al. (2018). Silver-nanoparticles increase bactericidal activity and radical oxygen responses against bacterial pathogens in human osteoclasts. Nanomedicine 14, 601–607. doi: 10.1016/j.nano.2017.11.006
Barros, D., Pradhan, A., Mendes, V. M., Manadas, B., Santos, P. M., Pascoal, C., et al. (2019). Proteomics and antioxidant enzymes reveal different mechanisms of toxicity induced by ionic and nanoparticulate silver in bacteria. Environ. Sci. Nano 6, 1207–1218. doi: 10.1039/c8en01067f
Boero, E., Brinkman, I., Juliet, T., van Yperen, E., van Strijp, J. A. G., Rooijakkers, S. H. M., et al. (2021). Use of flow cytometry to evaluate phagocytosis of staphylococcus aureus by human neutrophils. Front. Immunol. 12. doi: 10.3389/fimmu.2021.635825
Brandau, S., Jakob, M., Bruderek, K., Bootz, F., Giebel, B., Radtke, S., et al. (2014). Mesenchymal stem cells augment the anti-bacterial activity of neutrophil granulocytes. PloS One 9, e106903. doi: 10.1371/journal.pone.0106903
Brennan, S. A., Fhoghlú, C. N., DeVitt, B. M., O’Mahony, F. J., Brabazon, D., Walsh, A. (2015). Instructional review: general orthopaedics silver nanoparticles and their orthopaedic applications. Bone Joint J. 97-B, 582–589. doi: 10.1302/0301-620X.97B5.33336
Campoccia, D., Montanaro, L., Arciola, C. R. (2006). The significance of infection related to orthopedic devices and issues of antibiotic resistance. Biomaterials 27, 2331–2339. doi: 10.1016/j.biomaterials.2005.11.044
Carrola, J., Bastos, V., Daniel-da-Silva, A. L., Gil, A. M., Santos, C., Oliveira, H., et al. (2020). Macrophage metabolomics reveals differential metabolic responses to subtoxic levels of silver nanoparticles and ionic silver. Eur. J. Inorg Chem. 2020, 1867–1876. doi: 10.1002/ejic.202000095
Cecotto, L., van Kessel, K., Wolfert, M. A., Vogely, C., van der Wal, B., Weinans, H., et al. (2022). Antibacterial and anti-inflammatory properties of host defense peptides against staphylococcus aureus. iScience 25, 105211. doi: 10.1016/j.isci.2022.105211
Chen, Z., Yuen, J., Crawford, R., Chang, J., Wu, C., Xiao, Y. (2015). The effect of osteoimmunomodulation on the osteogenic effects of cobalt incorporated β-tricalcium phosphate. Biomaterials 61, 126–138. doi: 10.1016/j.biomaterials.2015.04.044
Chopra, I. (2007). The increasing use of silver-based products as antimicrobial agents: a useful development or a cause for concern? J. Antimicrobial Chemotherapy 59, 587–590. doi: 10.1093/jac/dkm006
Chow, L., Johnson, V., Impastato, R., Coy, J., Strumpf, A., Dow, S. (2020). Antibacterial activity of human mesenchymal stem cells mediated directly by constitutively secreted factors and indirectly by activation of innate immune effector cells. Stem Cells Transl. Med. 9, 235–249. doi: 10.1002/sctm.19-0092
Croes, M., Bakhshandeh, S., van Hengel, I. A. J., Lietaert, K., van Kessel, K. P. M., Pouran, B., et al. (2018). Antibacterial and immunogenic behavior of silver coatings on additively manufactured porous titanium. Acta Biomater 81, 315–327. doi: 10.1016/j.actbio.2018.09.051
da Silva Domingues, J. F., Roest, S., Wang, Y., van der Mei, H. C., Libera, M., van Kooten, T. G., et al. (2015). Macrophage phagocytic activity toward adhering staphylococci on cationic and patterned hydrogel coatings versus common biomaterials. Acta Biomater 18, 1–8. doi: 10.1016/j.actbio.2015.02.028
de Jong, N. W. M., van Kessel, K. P. M., van Strijp, J. A. G. (2019). Immune evasion by staphylococcus aureus. Gram-Positive Pathog. 618–639. doi: 10.1128/9781683670131.ch39
de Mesy Bentley, K. L., Trombetta, R., Nishitani, K., Bello-Irizarry, S. N., Ninomiya, M., Zhang, L., et al. (2017). Evidence of staphylococcus aureus deformation, proliferation, and migration in canaliculi of live cortical bone in murine models of osteomyelitis. J. Bone Mineral Res. 32, 985–990. doi: 10.1002/jbmr.3055
de Vor, L., Rooijakkers, S. H. M., van Strijp, J. A. G. (2020). Staphylococci evade the innate immune response by disarming neutrophils and forming biofilms. FEBS Lett. 594 (16), 2556–2569. doi: 10.1002/1873-3468.13767
Diez-Escudero, A., Hailer, N. P. (2021). The role of silver coating for arthroplasty components. Bone Joint J. 103, 423–429. doi: 10.1302/0301-620X.103B3.BJJ-2020-1370.R1
Djošić, M., Janković, A., Mišković-Stanković, V. (2021). Electrophoretic deposition of biocompatible and bioactive hydroxyapatite-based coatings on titanium. Materials 14 (18), 5391. doi: 10.3390/ma14185391
Durán, N., Durán, M., de Jesus, M. B., Seabra, A. B., Fávaro, W. J., Nakazato, G. (2016). Silver nanoparticles: a new view on mechanistic aspects on antimicrobial activity. Nanomedicine 12, 789–799. doi: 10.1016/j.nano.2015.11.016
Eto, S., Kawano, S., Someya, S., Miyamoto, H., Sonohata, M., Mawatari, M. (2016). First clinical experience with thermal-sprayed silver oxide-containing hydroxyapatite coating implant. J. Arthroplasty 31, 1498–1503. doi: 10.1016/j.arth.2015.12.034
Fiore, M., Sambri, A., Zucchini, R., Giannini, C., Donati, D. M., de Paolis, M. (2021). Silver-coated megaprosthesis in prevention and treatment of peri-prosthetic infections: a systematic review and meta-analysis about efficacy and toxicity in primary and revision surgery. Eur. J. Orthopaedic Surg. Traumatology 31, 201–220. doi: 10.1007/s00590-020-02779-z
Flannagan, R. S., Heit, B., Heinrichs, D. E. (2015). Antimicrobial mechanisms of macrophages and the immune evasion strategies of staphylococcus aureus. Pathogens 4 (4), 826–868. doi: 10.3390/pathogens4040826
Fraunholz, M., Sinha, B. (2012). Intracellular staphylococcus aureus: live-in and let die. Front. Cell Infect. Microbiol. 2. doi: 10.3389/fcimb.2012.00043
Gosheger, G., Hardes, J., Ahrens, H., Streitburger, A., Buerger, H., Erren, M., et al. (2004). Silver-coated megaendoprostheses in a rabbit model - an analysis of the infection rate and toxicological side effects. Biomaterials 25, 5547–5556. doi: 10.1016/j.biomaterials.2004.01.008
Grainger, D. W., van der Mei, H. C., Jutte, P. C., van den Dungen, J. J. A. M., Schultz, M. J., van der Laan, B. F. A. M., et al. (2013). Critical factors in the translation of improved antimicrobial strategies for medical implants and devices. Biomaterials 34, 9237–9243. doi: 10.1016/J.BIOMATERIALS.2013.08.043
Gristina, A. G., Naylor, P., Myrvik, Q. (1989). Infections from biomaterials and implants: a race for the surface. Med. Prog. Technol. 14, 205–224.
Gustafson, H. H., Holt-Casper, D., Grainger, D. W., Ghandehari, H. (2015). Nanoparticle uptake: the phagocyte problem. Nano Today 10, 487–510. doi: 10.1016/j.nantod.2015.06.006
Haase, H., Fahmi, A., Mahltig, B. (2014). Impact of silver nanoparticles and silver ions on innate immune cells. J. BioMed. Nanotechnol 10, 1146–1156. doi: 10.1166/jbn.2014.1784
Hackenberg, S., Scherzed, A., Kessler, M., Hummel, S., Technau, A., Froelich, K., et al. (2011). Silver nanoparticles: evaluation of DNA damage, toxicity and functional impairment in human mesenchymal stem cells. Toxicol. Lett. 201, 27–33. doi: 10.1016/j.toxlet.2010.12.001
Hamad, A., Khashan, K. S., Hadi, A. (2020). Silver nanoparticles and silver ions as potential antibacterial agents. J. Inorg Organomet Polym Mater 30, 4811–4828. doi: 10.1007/s10904-020-01744-x
Hamza, T., Dietz, M., Pham, D., Clovis, N., Danley, S., Li, B. (2013). Intra-cellular staphylococcus aureus alone causes infection in vivo. Eur. Cell Mater 25, 341–350. doi: 10.22203/ecm.v025a24
Hamza, T., Li, B. (2014). Differential responses of osteoblasts and macrophages upon staphylococcus aureus infection. BMC Microbiol. 14, 1–11. doi: 10.1186/s12866-014-0207-5
Hardes, J., von Eiff, C., Streitbuerger, A., Balke, M., Budny, T., Henrichs, M. P., et al. (2010). Reduction of periprosthetic infection with silver-coated megaprostheses in patients with bone sarcoma. J. Surg. Oncol. 101, 389–395. doi: 10.1002/jso.21498
Horn, J., Stelzner, K., Rudel, T., Fraunholz, M. (2018). Inside job: staphylococcus aureus host-pathogen interactions. Int. J. Med. Microbiol. 308, 607–624. doi: 10.1016/j.ijmm.2017.11.009
Huang, M., Ye, K., Hu, T., Liu, K., You, M., Wang, L., et al. (2021). Silver nanoparticles attenuate the antimicrobial activity of the innate immune system by inhibiting neutrophil-mediated phagocytosis and reactive oxygen species production. Int. J. Nanomedicine 16, 1345–1360. doi: 10.2147/IJN.S292482
Huang, Y., Zhang, Y., Li, M., Yang, H., Liang, J., Chen, Y., et al. (2022). Physicochemical, osteogenic and antimicrobial properties of graphene oxide reinforced silver/strontium-doped hydroxyapatite on titanium for potential orthopedic applications. Surf Coat Technol. 446, 128788. doi: 10.1016/J.SURFCOAT.2022.128788
Ivask, A., Elbadawy, A., Kaweeteerawat, C., Boren, D., Fischer, H., Ji, Z., et al. (2014). Toxicity mechanisms in escherichia coli vary for silver nanoparticles and differ from ionic silver. ACS Nano 8, 374–386. doi: 10.1021/nn4044047
Jhunjhunwala, S. (2018). Neutrophils at the biological-material interface. ACS Biomater Sci. Eng. 4, 1128–1136. doi: 10.1021/acsbiomaterials.6b00743
Johnson, V., Webb, T., Norman, A., Coy, J., Kurihara, J., Regan, D., et al. (2017). Activated mesenchymal stem cells interact with antibiotics and host innate immune responses to control chronic bacterial infections. Sci. Rep. 7, 1–18. doi: 10.1038/s41598-017-08311-4
Josse, J., Velard, F., Gangloff, S. C. (2015). Staphylococcus aureus vs. osteoblast: relationship and consequences in osteomyelitis. Front. Cell Infect. Microbiol. 5. doi: 10.3389/fcimb.2015.00085
Krasnodembskaya, A., Samarani, G., Song, Y., Zhuo, H., Su, X., Lee, J. W., et al. (2012). Human mesenchymal stem cells reduce mortality and bacteremia in gram-negative sepsis in mice in part by enhancing the phagocytic activity of blood monocytes. Am. J. Physiol. Lung Cell Mol. Physiol. 302, 1003–1013. doi: 10.1152/ajplung.00180.2011
Kuo, Y. J., Chen, C. H., Dash, P., Lin, Y. C., Hsu, C. W., Shih, S. J., et al. (2022). Angiogenesis, osseointegration, and antibacterial applications of polyelectrolyte multilayer coatings incorporated with Silver/Strontium containing mesoporous bioactive glass on 316L stainless steel. Front. Bioeng Biotechnol. 10. doi: 10.3389/fbioe.2022.818137
Leach, J. K., Whitehead, J. (2018). Materials-directed differentiation of mesenchymal stem cells for tissue engineering and regeneration. ACS Biomater Sci. Eng. 4, 1115–1127. doi: 10.1021/acsbiomaterials.6b00741
Liao, C., Li, Y., Tjong, S. C. (2019). Bactericidal and cytotoxic properties of silver nanoparticles. Int. J. Mol. Sci. 20 (2), 449. doi: 10.3390/ijms20020449
Liu, Y., Segura, T. (2020). Biomaterials-mediated regulation of macrophage cell fate. Front. Bioeng Biotechnol. 8. doi: 10.3389/fbioe.2020.609297
Liu, H., Yang, D., Yang, H., Zhang, H., Zhang, W., Fang, Y. J., et al. (2013). Comparative study of respiratory tract immune toxicity induced by three sterilisation nanoparticles: silver, zinc oxide and titanium dioxide. J. Hazard Mater 248–249, 478–486. doi: 10.1016/j.jhazmat.2013.01.046
Loi, F., Córdova, L. A., Ruth, Z., Jukka, P., Lin, T. H., Goodman, S. B., et al. (2016). The effects of immunomodulation by macrophage subsets on osteogenesis in vitro. Stem Cell Res. Ther. 7, 1–11. doi: 10.1186/s13287-016-0276-5
Lu, Z., Rong, K., Li, J., Yang, H., Chen, R. (2013). Size-dependent antibacterial activities of silver nanoparticles against oral anaerobic pathogenic bacteria. J. Mater Sci. Mater Med. 24, 1465–1471. doi: 10.1007/s10856-013-4894-5
Lu, D., Xu, Y., Liu, Q., Zhang, Q. (2021). Mesenchymal stem cell-macrophage crosstalk and maintenance of inflammatory microenvironment homeostasis. Front. Cell Dev. Biol. 9. doi: 10.3389/fcell.2021.681171
Luan, Y., van der Mei, H. C., Dijk, M., Geertsema-Doornbusch, G. I., Atema-Smit, J., Ren, Y., et al. (2020). Polarization of macrophages, cellular adhesion, and spreading on bacterially contaminated gold nanoparticle-coatings in vitro. ACS Biomater Sci. Eng. 6, 933–945. doi: 10.1021/acsbiomaterials.9b01518
Masters, E. A., Ricciardi, B. F., Bentley, K. L., de, M., Moriarty, T. F., Schwarz, E. M., et al. (2022). Skeletal infections: microbial pathogenesis, immunity and clinical management. Nat. Rev. Microbiol. 20, 385–400. doi: 10.1038/s41579-022-00686-0
McQuillan, J. S., Groenaga Infante, H., Stokes, E., Shaw, A. M. (2012). Silver nanoparticle enhanced silver ion stress response in escherichia coli K12. Nanotoxicology 6, 857–866. doi: 10.3109/17435390.2011.626532
Monteiro, N. O., Casanova, M. R., Quinteira, R., Fangueiro, J. F., Reis, R. L., Neves, N. M. (2022). Biomimetic surface topography as a potential modulator of macrophages inflammatory response to biomaterials. Biomaterials Adv. 141, 213128. doi: 10.1016/J.BIOADV.2022.213128
Moriarty, T. F., Grainger, D. W., Richards, R. G. (2014). Challenges in linking preclinical anti-microbial research strategies with clinical outcomes for device-associated infections. Eur. Cell Mater 28, 112–128. doi: 10.22203/eCM.v028a09
Moriarty, T. F., Kuehl, R., Coenye, T., Metsemakers, W. J., Morgenstern, M., Schwarz, E. M., et al. (2016). Orthopaedic device-related infection: current and future interventions for improved prevention and treatment. EFORT Open Rev. 1, 89–99. doi: 10.1302/2058-5241.1.000037
Nicolaidou, V., Wong, M. M., Redpath, A. N., Ersek, A., Baban, D. F., Williams, L. M., et al. (2012). Monocytes induce STAT3 activation in human mesenchymal stem cells to promote osteoblast formation. PloS One 7, e39871. doi: 10.1371/journal.pone.0039871
Park, E. J., Yi, J., Kim, Y., Choi, K., Park, K. (2010). Silver nanoparticles induce cytotoxicity by a Trojan-horse type mechanism. Toxicol. Vitro 24, 872–878. doi: 10.1016/j.tiv.2009.12.001
Pauksch, L., Hartmann, S., Rohnke, M., Szalay, G., Alt, V., Schnettler, R., et al. (2014). Biocompatibility of silver nanoparticles and silver ions in primary human mesenchymal stem cells and osteoblasts. Acta Biomater 10, 439–449. doi: 10.1016/j.actbio.2013.09.037
Pazos-Ortiz, E., Roque-Ruiz, J. H., Hinojos-Márquez, E. A., López-Esparza, J., Donohué-Cornejo, A., Cuevas-González, J. C., et al. (2017). Dose-dependent antimicrobial activity of silver nanoparticles on polycaprolactone fibers against gram-positive and gram-negative bacteria. J. Nanomater 2017. doi: 10.1155/2017/4752314
Perret, M., Badiou, C., Lina, G., Burbaud, S., Benito, Y., Bes, M., et al. (2012). Cross-talk between staphylococcus aureus leukocidins-intoxicated macrophages and lung epithelial cells triggers chemokine secretion in an inflammasome-dependent manner. Cell Microbiol. 14, 1019–1036. doi: 10.1111/j.1462-5822.2012.01772.x
Pidwill, G. R., Gibson, J. F., Cole, J., Renshaw, S. A., Foster, S. J. (2021). The role of macrophages in staphylococcus aureus infection. Front. Immunol. 11. doi: 10.3389/fimmu.2020.620339
Priebe, M., Widmer, J., Suhartha Löwa, N., Abram, S. L., Mottas, I., Woischnig, A. K., et al. (2017). Antimicrobial silver-filled silica nanorattles with low immunotoxicity in dendritic cells. Nanomedicine 13, 11–22. doi: 10.1016/J.NANO.2016.08.002
Qais, F. A., Shafiq, A., Khan, H. M., Husain, F. M., Khan, R. A., Alenazi, B., et al. (2019). Antibacterial effect of silver nanoparticles synthesized using murraya koenigii (L.) against multidrug-resistant pathogens. Bioinorg Chem. Appl. 2019. doi: 10.1155/2019/4649506
Qing, Y., Cheng, L., Li, R., Liu, G., Zhang, Y., Tang, X., et al. (2018). Potential antibacterial mechanism of silver nanoparticles and the optimization of orthopedic implants by advanced modification technologies. Int. J. Nanomedicine 13, 3311–3327. doi: 10.2147/IJN.S165125
Rajendran, A., Kapoor, U., Jothinarayanan, N., Lenka, N., Pattanayak, D. K. (2019). Effect of silver-containing Titania layers for bioactivity, antibacterial activity, and osteogenic differentiation of human mesenchymal stem cells on Ti metal. ACS Appl. Bio Mater 2, 3808–3819. doi: 10.1021/acsabm.9b00420
Rosales, C. (2018). Neutrophil: a cell with many roles in inflammation or several cell types? Front. Physiol. 9. doi: 10.3389/fphys.2018.00113
Rosowski, E. E. (2020). Determining macrophage versus neutrophil contributions to innate immunity using larval zebrafish. DMM Dis. Models Mech. 13, dmm041889. doi: 10.1242/dmm.041889
Samberg, M. E., Loboa, E. G., Oldenburg, S. J., Monteiro-Riviere, N. A. (2012). Silver nanoparticles do not influence stem cell differentiation but cause minimal toxicity. Nanomedicine 7, 1197–1209. doi: 10.2217/nnm.12.18
Sarkar, S., Leo, B. F., Carranza, C., Chen, S., Rivas-Santiago, C., Porter, A. E., et al. (2015). Modulation of human macrophage responses to mycobacterium tuberculosis by silver nanoparticles of different size and surface modification. PloS One 10, 1–28. doi: 10.1371/journal.pone.0143077
Selvamani, V., Kadian, S., Detwiler, D. A., Zareei, A., Woodhouse, I., Qi, Z., et al. (2022). Laser-assisted nanotexturing and silver immobilization on titanium implant surfaces to enhance bone cell mineralization and antimicrobial properties. Langmuir 38, 4014–4027. doi: 10.1021/acs.langmuir.2c00008
Sengstock, C., Diendorf, J., Epple, M., Schildhauer, T. A., Köller, M. (2014). Effect of silver nanoparticles on human mesenchymal stem cell differentiation. Beilstein J. Nanotechnology 5, 2058–2069. doi: 10.3762/bjnano.5.214
Shahabadi, N., Maghsudi, M., Ahmadipour, Z. (2012). Study on the interaction of silver(I) complex with bovine serum albumin by spectroscopic techniques. Spectrochim Acta A Mol. Biomol Spectrosc 92, 184–188. doi: 10.1016/j.saa.2012.02.071
Silva, M. T. (2010). When two is better than one: macrophages and neutrophils work in concert in innate immunity as complementary and cooperative partners of a myeloid phagocyte system. J. Leukoc. Biol. 87, 93–106. doi: 10.1189/jlb.0809549
Souter, P., Vaughan, J., Butcher, K., Dowle, A., Cunningham, J., Dodd, J., et al. (2020). Identification of mesenchymal stromal cell survival responses to antimicrobial silver ion concentrations released from orthopaedic implants. Sci. Rep. 10, 1–14. doi: 10.1038/s41598-020-76087-1
Strobel, M., Pförtner, H., Tuchscherr, L., Völker, U., Schmidt, F., Kramko, N., et al. (2016). Post-invasion events after infection with staphylococcus aureus are strongly dependent on both the host cell type and the infecting s. aureus strain. Clin. Microbiol. Infection 22, 799–809. doi: 10.1016/j.cmi.2016.06.020
Subbiahdoss, G., Fernández, I. C. S., da Silva Domingues, J. F., Kuijer, R., van der Mei, H. C., Busscher, H. J. (2011). In vitro interactions between bacteria, osteoblast-like cells and macrophages in the pathogenesis of biomaterial-associated infections. PloS One 6, e24827. doi: 10.1371/journal.pone.0024827
Tan, L., Liu, X., Dou, H., Hou, Y. (2022). Characteristics and regulation of mesenchymal stem cell plasticity by the microenvironment [[/amp]]mdash; specific factors involved in the regulation of MSC plasticity. Genes Dis. 9, 296–309. doi: 10.1016/J.GENDIS.2020.10.006
Trovarelli, G., Angelini, A., Pala, E., Cappellari, A., Breda, A., Ruggieri, P. (2019). Infection in orthopaedic oncology: crucial problem in modern reconstructive techniques. Eur. Rev. Med. Pharmacol. Sci. 23, 271–278. doi: 10.26355/EURREV_201904_17501
Tuchscherr, L., Kreis, C. A., Hoerr, V., Flint, L., Hachmeister, M., Geraci, J., et al. (2016). Staphylococcus aureus develops increased resistance to antibiotics by forming dynamic small colony variants during chronic osteomyelitis. J. Antimicrobial Chemotherapy 71, 438–448. doi: 10.1093/jac/dkv371
Veening-Griffioen, D. H., Ferreira, G. S., van Meer, P. J. K., Boon, W. P. C., Gispen-de Wied, C. C., Moors, E. H. M., et al. (2019). Are some animal models more equal than others? a case study on the translational value of animal models of efficacy for alzheimer’s disease. Eur. J. Pharmacol. 859, 172524. doi: 10.1016/J.EJPHAR.2019.172524
Wafa, H., Grimer, R. J., Reddy, K., Jeys, L., Abudu, A., Carter, S. R., et al. (2015). Retrospective evaluation of the incidence of early periprosthetic infection with silver-treated endoprostheses in high-risk patients: case-control study. Bone Joint J. 97-B, 252–257. doi: 10.1302/0301-620X.97B2.34554
Wesdorp, M. A., Schwab, A., Bektas, E. I., Narcisi, R., Eglin, D., Stoddart, M. J., et al. (2023). A culture model to analyze the acute biomaterial-dependent reaction of human primary neutrophils in vitro. Bioact Mater 20, 627–637. doi: 10.1016/J.BIOACTMAT.2022.05.036
Xiu, Z. M., Zhang, Q. B., Puppala, H. L., Colvin, V. L., Alvarez, P. J. J. (2012). Negligible particle-specific antibacterial activity of silver nanoparticles. Nano Lett. 12, 4271–4275. doi: 10.1021/nl301934w
Yang, Y., Zhang, C., Hu, Z. (2013). Impact of metallic and metal oxide nanoparticles on wastewater treatment and anaerobic digestion. Environ. Sciences: Processes Impacts 15, 39–48. doi: 10.1039/c2em30655g
Zaatreh, S., Wegner, K., Strauß, M., Pasold, J., Mittelmeier, W., Podbielski, A., et al. (2016). Co-Culture of s. epidermidis and human osteoblasts on implant surfaces: an advanced in vitro model for implant-associated infections. PloS One 11, 1–16. doi: 10.1371/journal.pone.0151534
Zhao, B., van der Mei, H. C., Subbiahdoss, G., de Vries, J., Rustema-Abbing, M., Kuijer, R., et al. (2014). Soft tissue integration versus early biofilm formation on different dental implant materials. Dental Materials 30, 716–727. doi: 10.1016/J.DENTAL.2014.04.001
Zhu, Y., Liang, H., Liu, X., Wu, J., Yang, C., Man Wong, T., et al. (2021) Regulation of macrophage polarization through surface topography design to facilitate implant-to-bone osteointegration. Available at: https://www.science.org.
Keywords: biomaterial-related infection, antimicrobial, co-culture, cytotoxicity, immune response
Citation: Cecotto L, Stapels DAC, van Kessel KPM, Croes M, Lourens Z, Vogely HC, van der Wal BCH, van Strijp JAG, Weinans H and Amin Yavari S (2023) Evaluation of silver bio-functionality in a multicellular in vitro model: towards reduced animal usage in implant-associated infection research. Front. Cell. Infect. Microbiol. 13:1186936. doi: 10.3389/fcimb.2023.1186936
Received: 11 April 2023; Accepted: 16 May 2023;
Published: 05 June 2023.
Edited by:
Yugal Kishore Mohanta, University of Science and Technology, Meghalaya, IndiaReviewed by:
Debasis Nayak, North Orissa University, IndiaArne Kienzle, Brigham and Women’s Hospital and Harvard Medical School, United States
Copyright © 2023 Cecotto, Stapels, van Kessel, Croes, Lourens, Vogely, van der Wal, van Strijp, Weinans and Amin Yavari. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Harrie Weinans, aC5oLndlaW5hbnNAdW1jdXRyZWNodC5ubA==