
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol., 17 April 2023
Sec. Intestinal Microbiome
Volume 13 - 2023 | https://doi.org/10.3389/fcimb.2023.1167116
This article is part of the Research TopicInsights of Probiotics and Fermented Diet on Gut MicrobiomeView all 4 articles
 Jie Gao1†Longyou Zhao2†Yiwen Cheng1,3†Wenhui Lei3,4Yu Wang1
Jie Gao1†Longyou Zhao2†Yiwen Cheng1,3†Wenhui Lei3,4Yu Wang1 Xia Liu5Nengneng Zheng6
Xia Liu5Nengneng Zheng6 Li Shao7Xulei Chen8Yilai Sun8
Li Shao7Xulei Chen8Yilai Sun8 Zongxin Ling1,3*‡Weijie Xu8*
Zongxin Ling1,3*‡Weijie Xu8*Depression is one of the most common psychiatric conditions, characterized by significant and persistent depressed mood and diminished interest, and often coexists with various comorbidities. The underlying mechanism of depression remain elusive, evidenced by the lack of an appreciate therapy. Recent abundant clinical trials and animal studies support the new notion that the gut microbiota has emerged as a novel actor in the pathophysiology of depression, which partakes in bidirectional communication between the gut and the brain through the neuroendocrine, nervous, and immune signaling pathways, collectively known as the microbiota-gut-brain (MGB) axis. Alterations in the gut microbiota can trigger the changes in neurotransmitters, neuroinflammation, and behaviors. With the transition of human microbiome research from studying associations to investigating mechanistic causality, the MGB axis has emerged as a novel therapeutic target in depression and its comorbidities. These novel insights have fueled idea that targeting on the gut microbiota may open new windows for efficient treatment of depression and its comorbidities. Probiotics, live beneficial microorganisms, can be used to modulate gut dysbiosis into a new eubiosis and modify the occurrence and development of depression and its comorbidities. In present review, we summarize recent findings regarding the MGB axis in depression and discuss the potential therapeutic effects of probiotics on depression and its comorbidities.
Depression is a psychiatric syndrome that characterized by slowed thinking, depressed mood, and reduced volitional activity as its main symptoms, and is often accompanied by suicidal tendencies and somatization symptoms, which pose a great threat to human health. Depression causes a huge economic burden on both families and society. Mounting evidence has shown a predominant increasing trend in the prevalence of depression in the general population (Weinberger et al., 2018), ranking 3rd leading cause of global disability in healthy life years (Friedrich, 2017). Currently, depression become the 4th most common illness in the world, with a prevalence rate of nearly 4.4% in both developed and developing countries (World Health Organization, 2017). According to the data from WHO, it is estimated that depression influences nearly 350 million people worldwide (Ledford, 2014), resulting in more than 800,000 suicide deaths annually. By 2030, depression is projected to rank as the first disease burden worldwide. Recently, the current COVID-19 pandemic has led to a great increase in depression, with an increase of nearly 53 million cases globally, 27.6% above the pre-pandemic levels (Salari et al., 2020). In China, depression is a major public health issue which has been reported to be the second leading cause of disability-adjusted life year (DALY) (Lu et al., 2021). According to the Blue Book on Depression in China (2022), the China Mental Health Survey reported that more than 95 million people suffering from depressive disorders account for 6.8% of the total population in China (Kessler and Bromet, 2013; Lu et al., 2021). Depression results in annual medical and social costs of up to 49.4 billion yuan, which has gradually become a major public health concern. Depression not only increases emotional suffering in patients but is also associated with an elevated prevalence of substantial present and future complications, such as irritable bowel syndrome (IBS), inflammatory bowel disease (IBD), heart disease, high cholesterol, obesity, diabetes mellitus (DM), and Alzheimer’s disease (AD), which will affect the quality of life (QoL) of these patients (Gold et al., 2020). Thus, there is an urgent need for scientists worldwide to address depression and its complications.
However, the pathogenesis of depression, which is important for its prevention and treatment, has not yet been clarified. Depression is caused by numerous environmental, genetic, and psychological factors. Currently, the major hypotheses for the development of depression include the monoamine reduction hypothesis, overactivation of the hypothalamus-pituitary-adrenal (HPA) axis, and the decrease in brain-derived neurotrophic factor (BDNF) levels (Sonali et al., 2022). All these hypotheses are closely related to the interaction of the gut-brain axis with the gut microbiota, named the microbiota-gut-brain (MGB) axis. In recent years, accumulating evidence has shown a close link between the gut and the brain, and that the gut microbiota can be involved in regulating brain development, anxiety, depression, cognitive function, and other central nervous system (CNS) activities (Adak and Khan, 2019; Ling et al., 2020; Simpson et al., 2021; Cheng Y. et al., 2022; Khan et al., 2022; Li and Chen, 2022; Li et al., 2022; Ling et al., 2022a; Ling et al., 2022b; Ling et al., 2022c; Ling ZX. et al., 2022). In patients with depression, neuronal apoptosis occurs in the frontal cortex, hippocampus, and amygdala, while the abnormalities of the metabolism, secretion, inter-synaptic transmission, and reuptake of monoamine transmitters such as norepinephrine, dopamine, and 5-hydroxytryptamine (5-HT) take place in the synaptic gap (Farooq et al., 2022; Suda and Matsuda, 2022; Zhu et al., 2022). Structural imaging techniques have revealed that the dorsolateral prefrontal cortex, superior parietal lobule hippocampus, and other parts of the hippocampus are reduced in volume (Doney et al., 2022). These responses are achieved through the immune, neuroendocrine, and vagal pathways of the MGB axis, which can be affected by the gut microbiota. In the immune pathway, the metabolites of the gut microbiota, such as short-chain fatty acids (SCFAs) and indole, can stimulate epithelial enterochromaffin cells (ECCs) to produce glucagon-like peptide-1 (GLP-1), which plays a vital role in reducing neuroinflammation (Cheng et al., 2019; Peirce and Alviña, 2019; Farooq et al., 2022; Sonali et al., 2022). In addition, gut dysbiosis-induced immune activation increases pro-inflammatory cytokines and reactive oxygen/nitrogen species levels, which leads to oxidative stress and causes hyperactivation of the HPA axis (Bravo et al., 2011; Simpson et al., 2021; Sonali et al., 2022). In the neuroendocrine pathway, various products of the gut microbiota can influence brain function, including neurotransmitters such as γ-aminobutyric acid (GABA), dopamine, serotonin, SCFAs, and tryptophan metabolites (Alli et al., 2022; Sonali et al., 2022). In the vagal pathway, the gut microbiota can modulate brain function through the vagus nerve. Vagal sensory neurons form various mechanosensory and chemosensory endings along the gastrointestinal tract that receive enterocephalic signals (Liu et al., 2021a; Décarie-Spain et al., 2023). In addition, ECCs can form synapses with adjacent nerves to assist the vagus nerve in receiving intestinal signals (Bravo et al., 2011; Sun et al., 2019; Snigdha et al., 2022). It is clear from the above that the gut microbiota and its metabolites regulate brain function through multiple pathways, which provides us with the novel idea of treating CNS diseases by modulating the gut microbiota.
Gut microbiota is a reservoir of trillions of bacteria, archaea, viruses, parasites, and fungi that live in the gut, which has been considered a forgotten organ of the human body. The gut microbiota plays a crucial role in a wide array of host processes, such as growth, development, physiology, immune regulation (Wastyk et al., 2021), intestinal mucosal barrier (Paone and Cani, 2020), nutrition (Valdes et al., 2018), colonization resistance (Ducarmon et al., 2019), and alterations in the gut microbiota are related to the development of various intestinal and extraintestinal diseases. During the past decades, scientists have made great efforts to explore how large-scale disruptions and dynamic shifts in the gut microbiota can drive phenotypic changes and disease states. With the strong evidence displayed by multi-directional evidence, recent findings confirm that the gut microbiota composition can affect brain development and behavior (Loughman et al., 2020; Carlson et al., 2021; Kelsey et al., 2021). Currently, the gut microbiota is no longer forgotten and has been called the “second brain” of the body (Ridaura and Belkaid, 2015). It is now well accepted that the gut microbiota is important for various brain processes such as neurogenesis, myelination, and microglial activation, which can also effectively regulate behavior and affect psychological processes such as mood and cognition (Dinan and Cryan, 2016). Gut dysbiosis can disturb the bidirectional crosstalk between the gut microbiota and the CNS, leading to numerous neurological conditions, including chronic pain, stress, anxiety, depression, autism spectrum disorder (ASD), AD, and Parkinson’s disease.
Recently, increasing evidence has shown a link between gut microbiota and depression via the MGB axis. In people with depression, alterations in gut microbiota patterns are evident, suggesting that the gut microbiota plays important roles in the pathogenesis and onset of depression (Jiang et al., 2015; Zheng et al., 2016; Ling et al., 2022a). The use of antibiotics can lead to gut dysbiosis, disrupt intestinal homeostasis, and increase the risk of depression (Hao et al., 2020). Changes in bacterial diversity and richness in patients with depression have been observed in several previous studies, although there was no consistent directional alteration in microbial diversity. Our previous study found that α-diversity increased significantly in adult patients with depression, whereas bacterial β-diversity did not change significantly (Jiang et al., 2015). However, the opposite patterns were found in childhood depression (Ling et al., 2022a). Previous studies have reported that several key functional bacteria at different taxonomic levels are associated with depression. Our previous study found that the decreased butyrate-producing bacteria (e.g., Faecalibacterium) were associated with the enrichment of intestinal pathogenic bacteria or opportunistic pathogens such as Enterobacteriaceae (Jiang et al., 2015). Interestingly, the key functional genus, Faecalibacterium, was correlated with the severity of depression symptoms and the difficulty in abstract thinking negatively (Valles-Colomer et al., 2019). Barandouzi et al. observed that depressed patients had a lower abundance of Bacillus spp., Proteus spp., E. faecalis, and Sartorius spp. and a higher abundance of Actinomyces and Eggerthella spp (Barandouzi et al., 2020).. These key functional bacteria were found to be correlated with the development of depression significantly, but their causal effects on depression remain unclear. Recently, animal models have emerged as valuable experimental tools for host-microbiome interaction research. Zheng et al. demonstrated that transferring the gut microbiota from depressed humans to germ-free (GF) mice could increase depression-like behavior in the recipient mice, supporting a causal relationship between gut microbiota and depression (Zheng et al., 2016). After successful antidepressant treatment, the depression-associated key functional differential bacteria decreased significantly compared to healthy controls, indicating that the depressed gut microbiota tended to reconstitute (Li N. et al., 2019; Farooq et al., 2022). Based on these clinical and preclinical findings, we tentatively inferred that maintaining or restoring the normal condition of the gut microbiota is associated with regression depression. These interactions are often shown to be produced by the immune, neuroendocrine, and vagal pathways with the gut microbiota and its metabolites via the MGB axis. Bacterial groups associated with gastrointestinal inflammation (e.g., Enterobacteriaceae, Eggerthella, and Desulfovibrio) were relatively abundant in patients with depression, and these patients had fewer anti-inflammatory SCFA-producing bacterial species, including butyrate-producing E. faecalis and Clostridium XIVa (Simpson et al., 2021). With the advent of multi-omics techniques, such as metagenomics, metabolomics, proteomics, and culturomics, depression-associated key functional bacteria can be identified into species or strain levels, which allows us to clarify the specific functions of the depressed bacteria and screen these species as biomarkers for the intervention and treatment of depression and related complications.
Traditionally, depression treatments target the brain with psychotherapy and/or different drugs, such as 5-HT reuptake inhibitors (SSRI), 5-HT, and norepinephrine reuptake inhibitors (SNRI). However, the prevalence and burden of depression remained unchanged (Malhi and Mann, 2018). Recent advances have observed that these conventional treatments not only regulate the brain directly (Cipriani et al., 2018), but also affect the gut microbiota (Davey et al., 2013). Correcting abnormal gut microbiota could alleviate depression, suggesting that targeting on the gut microbiota could be considered as a promising and tractable therapy for depression. The modulation of the gut microbiota has been highlighted in the treatment of mental disorders, including depression and its comorbidities. To explore more potential possibilities for the treatment of depression, this review explored the use of probiotics in depression and for diseases in which depression is a co-morbidity. Here, we describe the pathogenesis associated with the MGB axis in depression and discuss the potential therapeutic effects of probiotics on depression and its comorbidities, with the expectation that probiotics will become a new and effective treatment for depression and its comorbidities in the future.
There is growing evidence to support the role of the gut microbiota in regulating host behavior and brain function. Although the exact mechanisms by which the gut microbiota causes or alters depression are not fully understood, the vast evidence from previous clinical and preclinical studies supports the hypothesis that the gut microbiota can affect the development of depression, mainly through the HPA axis, inflammation, and modify the abundance of BDNF. Clarifying the underlying mechanisms between gut microbiota and depression can contribute to the effective prevention and treatment of depression (Figure 1).
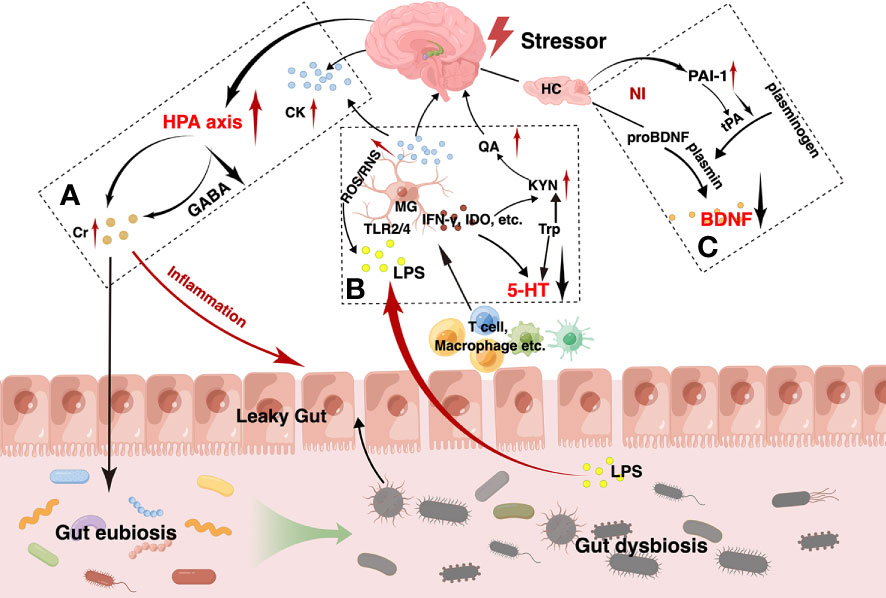
Figure 1 The main pathogenesis hypotheses for depression via the gut-brain axis. As shown, the hypothalamus-pituitary-adrenal (HPA) axis of the brain is activated by various stressors such as psychological stress and the increased pro-inflammatory cytokines (A). Due to the disruption of the g-aminobutyric acid (GABA) negative feedback in depression, however, the HPA axis abnormally activated, leading to persistent elevation of cortisol. High level of cortisol contributes to peripheral inflammation and can disrupt the normal gut microbiota. Gut dysbiosis, inflammation and other factors in combination lead to impaired intestinal barrier function, called leaky gut, which manifests as increased intestinal permeability, decreased intestinal villi length and colonic crypts depth. Intestinal immune cells such as T cells, macrophages, and so on, and gram-negative bacteria-produced lipopolysaccharide (LPS) in the gut can translocate from gut into brain, which can act on the microglia and lead to neuroinflammation. In turn, the over-activated microglia can release amount of indoleamine 2,3-dioxygenase (IDO), interferon γ (IFN-γ) and so on, resulting in a decrease in tryptophan metabolism to 5-hydroxytryptamine (5-HT) and an increase in the neurotoxic kynurenine, quinolinic acid (B). Meanwhile, neuroinflammation can reduce the level of brain-derived neurotrophic factor (BDNF) that is produced in the atrophied hippocampus and prefrontal cortex. Abnormally reduced plasminogen activator inhibitor-1 (PAI-1) is also considered as a possible causative agent (C). All these factors interact with each other and contribute to the onset and worsening of depression.
The HPA axis is a feedback pathway consisting of the hypothalamus, pituitary, and adrenal glands and is involved in the control of the stress response. The HPA axis is an important component of the MGB interaction (Cryan and O'mahony, 2011; Frankiensztajn et al., 2020). Under normal conditions, environmental, emotional, and physiological stressors increase the systemic levels of pro-inflammatory cytokines, which in turn secrete corticotropin-releasing hormone (CRH) by triggering secretion from the paraventricular nucleus of the hypothalamus. Increased CRH levels activate adrenocorticotropic hormone (ACTH) secretion from the anterior pituitary. ACTH subsequently results in the release of glucocorticoids from the adrenal cortex. When glucocorticoid levels reach their peak, negative feedback regulation is triggered via the GABA pathway, resulting in a decrease in cortisol secretion. This constitutes a complete HPA axis pathway, which ensures the stability of cortisol levels in vivo (Suda and Matsuda, 2022). However, this mechanism is disrupted in patients with depression. As we know before, the HPA axis is negatively regulated through the GABA pathway, which proceeds by increasing glutamate reuptake by presynaptic neurons when cortisol levels peak, and within the neuron, glutamate is converted to GABA as a precursor, followed by GABA exiting the neuron into the synaptic vesicles, crossing the synaptic gap and binding to GABA receptors on the postsynaptic membrane, thus establishing an aminobutyric acidergic synapse (Herman et al., 2004; Cullinan et al., 2008; Sarkar et al., 2011; Colmers and Bains, 2018; Duman et al., 2019). GABA acts as an inhibitory neurotransmitter that inhibits CRF neurons, ultimately reducing the secretion of cortisol (Giordano et al., 2006). That is, the GABA content is elevated during normal action. However, the dexamethasone suppression tests observed lesser levels of GABA levels in depressed patients which are unable to regulate excessive cortisol levels (Carroll et al., 1981), while excessive cortisol levels can lead to gut dysbiosis eventually (Łaniewski and Herbst-Kralovetz, 2022; Couch et al., 2023). Gut dysbiosis promotes the growth of gram-negative bacteria, releases increased immunogenic lipopolysaccharide (LPS), disrupts intestinal permeability, leads to leaky gut and endotoxemia, and causes translocation of bacterial components in the gut lumen due to the inflammatory response. These inflammatory mediators in turn stimulate the HPA axis, exacerbating its hyperactivation and promoting neuroinflammation, leading to depressive behavior (Zheng et al., 2016; Liu et al., 2021b; Sonali et al., 2022).
As mentioned above, high cortisol levels lead to a compromised intestinal mucosal barrier, which allows T cells, giant cells, and antigens to flow into the brain, activate microglia, and trigger inflammation (Doney et al., 2022). It can also lead to inflammation through the binding of effluent LPS to toll-like receptor (TLR) on microglia (Qin et al., 2007). Indeed, excessive microglial activation is the cornerstone of neuroinflammation. Multiple factors can lead to its overactivation to produce neuroinflammation and eventually lead to depression.
Inflammation is a key causative factor in the development of depression. However, the exact mechanisms underlying inflammation-related depression are still elusive. Typically, over-activation of the HPA axis by stress can cause abnormally high cortisol levels and disruption of the intestinal barrier, allowing the outflow of gut microbiota-derived endotoxins into circulation. This causes increased pro-inflammatory cytokines and decreased anti-inflammatory cytokines, leading to peripheral inflammation, which is associated with depressive symptoms. Some studies have reported increased interleukin-1β (IL-1β) and interleukin-6 (IL-6) and decreased IL-4 and IL-10 in patients with depression (Berk et al., 2013; Wong et al., 2016; Ling et al., 2022a). Then, pro-inflammatory factors cross the blood-brain barrier (BBB) into the brain through blood circulation, activating microglia, releasing reactive nitrogen and oxygen, and damaging brain epithelial cells leading to neuroinflammation, which can lead to mental illness (Peirce and Alviña, 2019; Doney et al., 2022). At the same time, indoleamine 2,3-dioxygenase (IDO) is a key enzyme in inflammation-induced depression. The pro-inflammatory factors also activate IDO enzyme activity, which causes tryptophan to be broken down into kynurenine rather than 5-HT, reducing 5-HT concentrations (Peirce and Alviña, 2019; Walker et al., 2019). Furthermore, pro-inflammatory factors disrupt tetrahydrobiopterin, an essential factor for monoamine synthesis, leading to impaired synthesis of 5-HT, dopamine, and other neurotransmitters (Dantzer et al., 2008; Miller and Raison, 2016).
As mentioned earlier, fecal microbiota transplantation (FMT) from depressed patients to GF mice causes morbidity in the mice. Compared with normal mice, GF mice are immunocompromised and lack regulatory T cells (Tregs) (Zheng et al., 2016). Thus, these findings suggest a mediating role of the gut microbiota in immune response and depression. Mason et al. found that anti-inflammatory bacterial groups such as Bacteroides and the Clostridium leptum subgroup are reduced in those diagnosed with depression (Mason et al., 2020), suggesting that gut dysbiosis, especially the decrease of these anti-inflammatory regulatory bacteria, plays an important role in the pathophysiology of depression. Given the key role of inflammation in depression, strong evidence has supported the novel concept of the gut microbiota-inflammation-brain axis, in which the gut microbiota can alter brain function through inflammatory signaling pathways and affect depression-like behavior. Thus, targeting the gut microbiota to modulate the inflammatory response in individuals with depression may represent a useful therapeutic approach for depression.
BDNF is a neurotrophin that can regulate the growth and plasticity of neurons and synapses (Bercik et al., 2011a). It is distributed in the hippocampus widely and has been considered as a key transducer of antidepressant effects. Generally, BDNF is first synthesized a precursor protein, pro-brain-derived neurotrophic factor (pro-BDNF), which is further processed into the mature form by fibrinolytic enzymes. BDNF can increase synaptic plasticity, promote neurogenesis, especially in the hippocampus, and maintain and promote the developmental differentiation and regeneration of various neurons, especially pentraxin and dopaminergic neurons, whereas pro-BDNF induces neuronal death and synaptic pruning (Bai et al., 2016). Fibrinolytic enzymes are converted from fibrinogen in the presence of tissue plasminogen activator, which can be inhibited by plasminogen activator inhibitor-1 (PAI-1), leading to the accumulation of pro-BDNF. Previous studies found that PAI-1 increases in the prefrontal cortex and hippocampus of chronically stressed rats (Party et al., 2019; Zhang W. et al., 2022). The decreased BDNF can be found in patients with depression (Jiang et al., 2015; Youssef et al., 2018), whereas antidepressant treatment can increase the levels of BDNF (Martinotti et al., 2016). Kuhlmann et al. observed that the levels of BDNF were negatively correlated with the severity of depression (Kuhlmann et al., 2017). Serum BDNF levels correlate with hippocampal volume, and insufficient BDNF levels can impair neurogenesis and lead to the onset of depression (Erickson et al., 2012; Von Bohlen Und Halbach and Von Bohlen Und Halbach, 2018). These findings suggest the possibility of using serum BDNF level as an indicator of disease activity and treatment response. Evidence from animal studies using GF, antibiotic-treated, depression models, and FMT mice has also demonstrated lower levels of BDNF in the hippocampus and cortex than in healthy controls (Clarke et al., 2013; Jang et al., 2021; Suda and Matsuda, 2022). Gut eubiosis can increase the activation of cAMP response element binding (CREB) in the hippocampus, prefrontal cortex, and amygdala, leading to an increase in BDNF production (Gerhard et al., 2016; Methiwala et al., 2021). In addition, the gut microbiota can modulate the conversion of pro-BDNF into BDNF. Administration of microbiota products to mice increases BDNF levels in the hippocampus (Carabotti et al., 2015; Maqsood and Stone, 2016). Previous interventional studies have revealed that probiotics can restore the levels of pro-BDNF and BDNF in various brain regions which are related to the development of depression-like behavioral phenotypes (Sun et al., 2018; Mohammed et al., 2020). These findings suggest that re-establishment of the gut microbiota may contribute to the increased levels of brain BDNF and modulate host behavior.
The aforementioned mechanisms provide an introduction to the pathogenesis of depression briefly. In fact, the causes of depression are inconclusive and there are numerous hypotheses about its pathogenesis, of which the above-mentioned are few and incomplete. The premise we are dealing with is related to the gut microbiota and can be treated with probiotics and other related mechanisms. Based on the above mechanisms, we will explore the existing targets that can act through probiotic therapy and explore the possible applications of probiotics in the future, which often act through the MGB axis interaction, reflecting the great role and potential of the gut microbiota in regulating neurological diseases.
Various antidepressant agents are available to treat depression, including monoamine oxidase inhibitors, tricyclic antidepressants, selective SSRI, nonselective SNRI, selective norepinephrine reuptake inhibitors, and other miscellaneous agents such as mirtazapine (Keller et al., 2002; Cryan and Dinan, 2015). However, nearly 30% of patients with depression are resistant to any treatment. Thus, novel antidepressant agents and strategies are required (Miyanishi and Nitta, 2021). Recent microbiota studies have demonstrated a strong link between depression and the gut microbiota. Miyaoka et al. has observed that the combination of antidepressants and probiotics is more effective to treat drug-resistant depression (Miyaoka et al., 2018). Preclinical studies and clinical trials suggest that modifying the composition of the gut microbiota via probiotic supplementation have been proven to be beneficial in treating or preventing human diseases (Zhao et al., 2018; Sanders et al., 2019; Edwards et al., 2020; Sun et al., 2021), which may be a viable adjuvant treatment option for patients with depression.
Probiotics are live microorganisms that, when administered in adequate amounts, confer a health benefit to the host by changing the composition of the host’s microbiota in a certain area (Hill et al., 2014). They play a vital role in maintaining a healthy gut by regulating the host mucosa and systemic immune function or by regulating the balance of microbiota in the gut (Sánchez et al., 2017; Clemente et al., 2018). Probiotics can tolerate stomach acid and bile salts, adhere to host intestinal epithelial cells, and remove or reduce the adhesion of pathogenic bacteria (Selle and Klaenhammer, 2013; Ducarmon et al., 2019; Mazloom et al., 2019). Owing to their non-toxic side effects and high stability, probiotics are increasingly used in the prevention and treatment of intestinal disorders such as IBS, IBD, antibiotic-associated diarrhea, and other disorders (Park et al., 2018; Glassner et al., 2020). Gut microbiota modulation with probiotics has become a hot topic in the treatment of mental disorders, including depression, although it is still in its infancy stage. Probiotics acting via the MGB axis can influence brain development, function, and behavior (Desbonnet et al., 2014; Buffington et al., 2016). This has prompted growing interest in the possibility of targeting the gut microbiota to beneficially impact depression. The concept of “psychobiotics”, proposed by Dinan et al., emphasizes the potential of probiotics in mental disorders treatment (Dinan et al., 2013). Psychobiotics can convey benefits to the host’s mental health via dynamic MGB crosstalk. An emerging body of evidence suggests possible antidepressant effects resulting from probiotic supplementation, which can normalize depression-associated physiological outputs, such as corticosterone, noradrenaline, BDNF, and immune function. The promising role of probiotics in depression via in vivo and in vitro studies have laid a strong foundation for clinical application. Several recent randomized controlled trials (RCTs) have demonstrated that probiotics can alleviate depressive symptoms in participants both with and without a clinical diagnosis of depression effectively (Goh et al., 2019; Amirani et al., 2020; Chao et al., 2020; Dehghani et al., 2022). Probiotics exhibit antidepressant properties in the absence of other therapeutic options (Nikolova et al., 2021). Thus, microbiota-based interventions with probiotics may possess greater therapeutic potential for depression treatment, which can be used as an adjunct to current approaches (Table 1). Common probiotic strains, such as Lactobacillus spp., Bifidobacterium spp., Akkermansia spp., Clostridium spp., and Enterococcus spp., have been used to treat depression in clinical and animal studies, either as a single agent or in combination with other potential psychobiotics (Figure 2). However, it is important to note that these benefits are strain-specific. We selected some strains that have already played an effective role in the treatment of depression to illustrate the specific mechanism of its action, clarify its dosage, periodicity, and other key information in the current treatment regimen, and pave the way for the further role of probiotics, which are expected to become new options for the treatment of depression (Table 2).
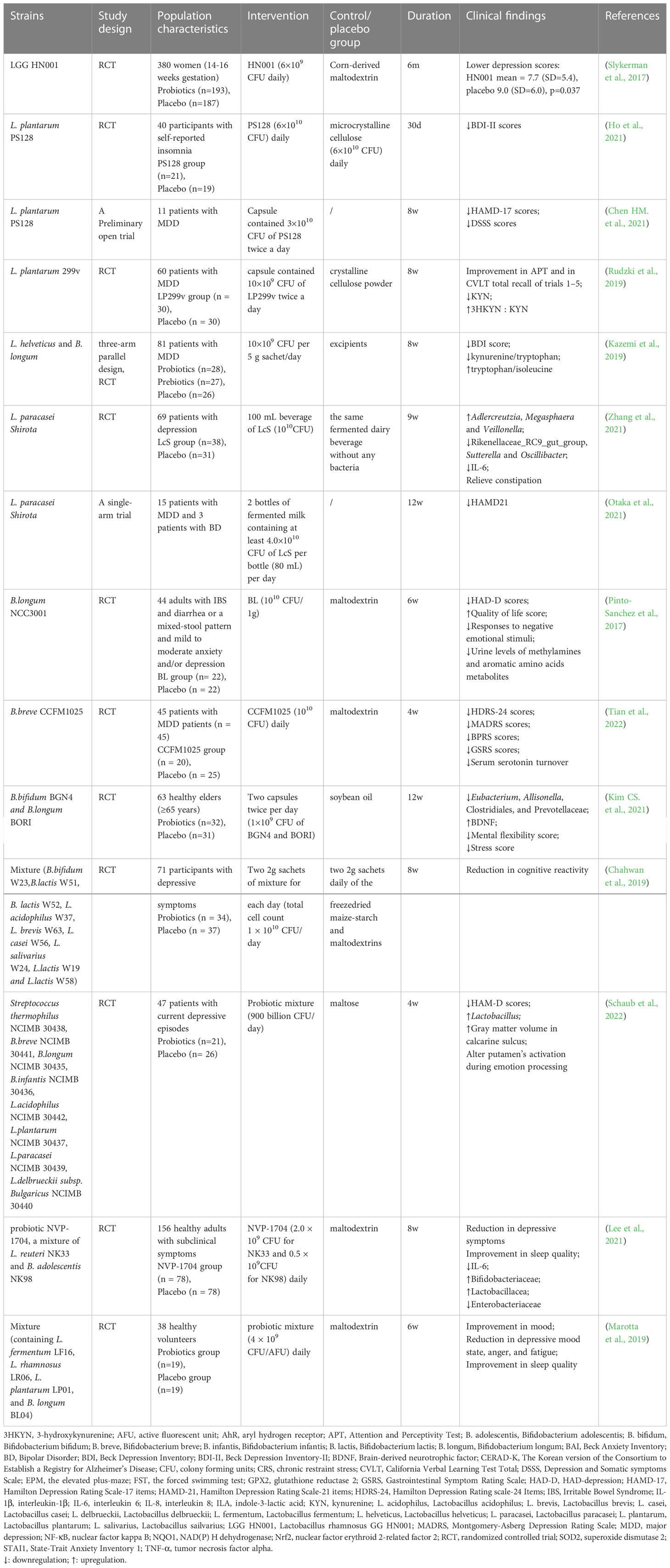
Table 1 Clinical evaluation of probiotics on depression and its comorbidities.
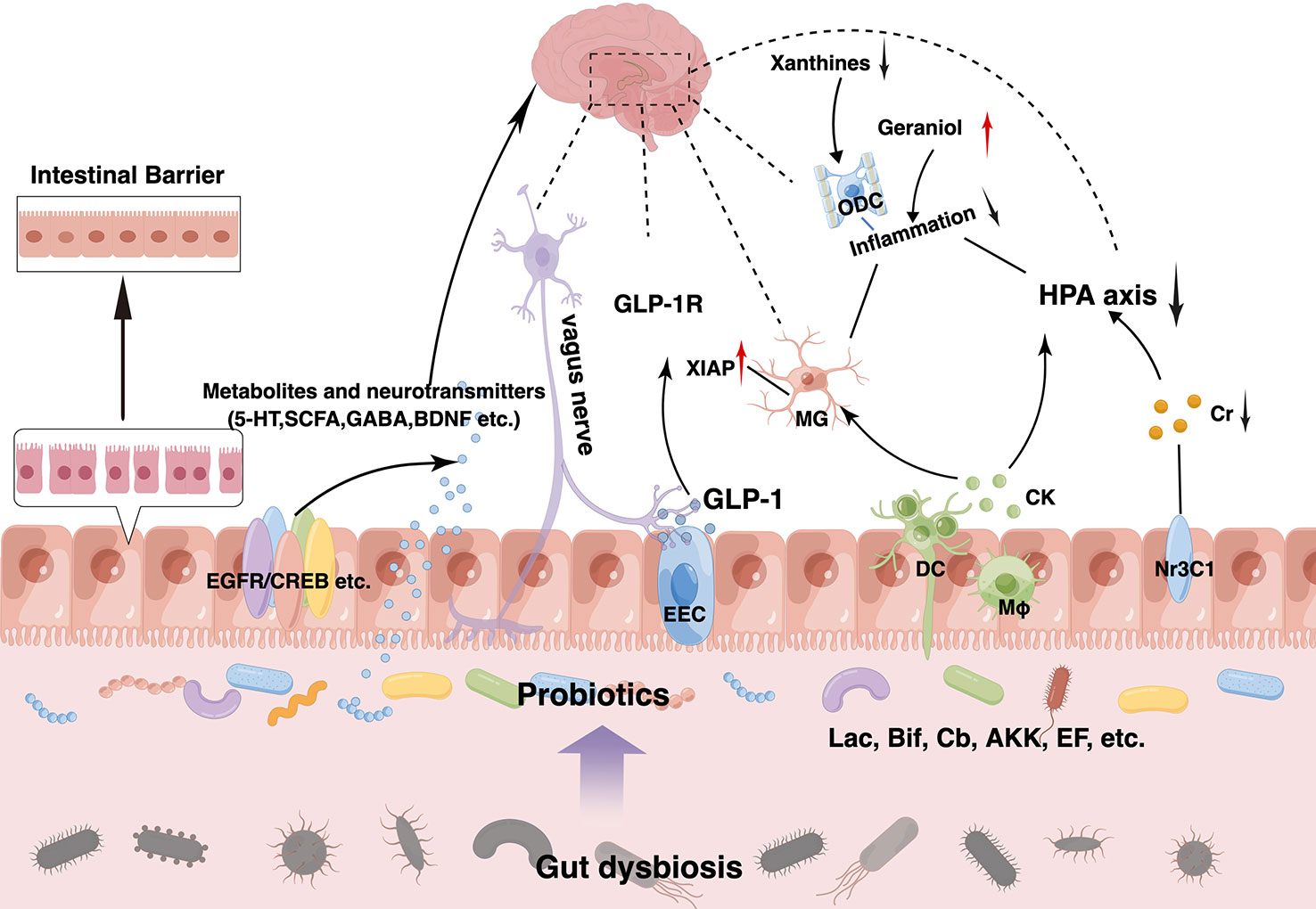
Figure 2 Therapeutic effects of probiotics on depression and its comorbidities. Probiotics exert antidepressant effects at multiple targets. Supplementation of probiotics can increase beneficial microorganisms and reduce harmful ones to achieve new gut eubiosis. Meanwhile, they can produce beneficial substances such as 5-hydroxytryptamine (5-HT), short-chain fatty acids (SCFAs) and brain-derived neurotrophic factor (BDNF) and so on by themselves or indirectly through upregulation of beneficial microbiota, which can act on various receptors on the intestinal epithelium such as EGFR, CREB, Nr3C1, etc. Moreover, probiotics can enhance intestinal barrier function in a variety of ways, such as upregulating mucus production by goblet cells, enhancing zonula occludens-1 (ZO-1), occludin, and claudin-1 expression, and reducing inflammation. Probiotics can also regulate brain function and improve neurological function through the vagus nerve, glucagon-like peptide-1 (GLP-1) pathway, etc. And it can activate the immune system to produce anti-inflammatory factors, alleviating circulatory inflammation and neuroinflammation while downregulating the hyperactive microglia and hypothalamus-pituitary-adrenal (HPA) axis. Similarly, decreased xanthines levels and elevated geraniol levels can also reduce inflammation.
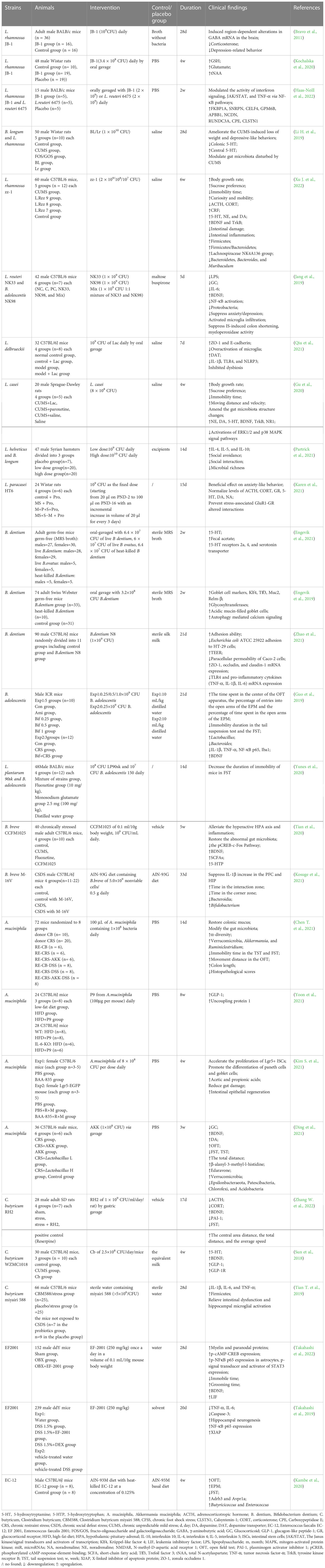
Table 2 Animal studies of probiotics on depression and its comorbidities.
Lactobacillus is one of the most widely used and intensively studied probiotic bacteria in gut microbiota. Lactobacillus spp. are anaerobic, gram-positive, peroxidase-negative, non-spore-forming rods that grow better under the microaerobic condition. As one of the inhabitants of the healthy microbiota in the human gut, vagina, and oral cavity, Lactobacilli have been considered safe microorganisms for the host health, with low pathogenic potential, and lack the ability to transmit antibiotic resistance factors to pathogens (Saarela et al., 2000). Thus, Lactobacilli strains isolated from natural products have been proposed as promising probiotic candidates. Several Lactobacilli strains have been used as probiotics, including L. plantarum, L. fermentum, L. rhamnosus and L. casei, which are isolated from the gut and exert various benefits to the host, including attenuation of anxiety and cognitive improvement (Goldstein et al., 2015). Many previous studies have demonstrated the beneficial effects of these Lactobacilli strains on mood, anxiety, and cognition, which can be considered as potential psychobiotics.
Among these Lactobacilli strains, L. rhamnosus must be mentioned, which is acid- and bile-stable and has a strong affinity for human intestinal mucosal cells. Many animal studies have found that oral administration with one L. rhamnosus strain, L. rhamnosus JB-1 (JB-1), demonstrates psychoactive and neuroactive properties. JB-1 can change the levels of neurotransmitters in the brains of mice, which in turn reduces stress-induced anxiety- and depression-related behaviors (Bravo et al., 2011; Janik et al., 2016). JB-1 consistently regulates the expression of GABAA and GABAB receptors in a region-dependent manner in mice, restoring metabolites such as GABA and glutamate to normal levels and reducing corticosterone levels (Tette et al., 2022). Janik et al. reported that 25% of central GABA levels could be elevated by four weeks of treatment with the JB-1 strain in BALB/c mice (Janik et al., 2016). Interestingly, the antidepressant effects of JB-1 depend on the intact vagus nerve connection between the gut and brain (Bravo et al., 2011). Subphrenic vagotomy prevented this effect, suggesting that the modulatory effect of JB-1 on GABA proceeds through the vagus nerve. JB-1 can directly stimulate vagal afferent neurons in the gut, with signals uploaded to the solitary bundle nucleus, followed by projections to the paraventricular nucleus, ultimately activating the GABAergic system, creating negative feedback, and lowering cortisol levels (Bravo et al., 2011). In addition, JB-1 also results in modulating the immune system and induces regulatory T cells, which have been found to be both necessary and sufficient to mediate the behavioral effects of bacteria (Liu et al., 2020). However, JB-1 can attenuate stress-induced behavioral deficits successfully but fails to re-establish the diversity and richness of the gut microbiota or correct the relative abundances of specific bacteria that altered by stress. This suggests that the neuroactive properties of beneficial microbes may not be mediated by gut microbiota restoration, but be determined by their functional activity (bacterial metabolites) and direct modulation of host signaling pathways (Bharwani et al., 2017).
Another L. rhamnosus strain, L. rhamnosus GG (LGG), which originates indigenously in the human gut, became available for use as a probiotic in Finland in 1990. LGG colonization early in life increases tight junction protein expression and immunoglobulin A production, upregulates host immune responses, increases intestinal villus length and colonic crypt depth, and enriches beneficial bacteria such as Bifidobacterium and Akkermansia (Zhou et al., 2022). With its effects on the increase in SCFAs-producing bacteria such as Bifidobacterium, mice had significantly higher levels of acetate, which helped alleviate anxiety (Strati et al., 2016). LGG has been found to increase GABA concentrations in fermented adzuki bean milk under optimized culture conditions (Song and Yu, 2018). In addition, LGG implantation can also activate epithelial growth factor receptor expression, enhance serotonin transporter protein expression, modulate the serotonergic system in the gut, and increase the levels of BDNF and GABA receptors in the amygdala and hippocampus (Cui et al., 2014; Johnson and Foster, 2018), which can alleviate anxiety and depression symptoms (Xu J. et al., 2022). Neufeld et al. observed that dietary supplementation with the probiotic LGG alone or in combination with the prebiotics polydextrose and galactooligosaccharide can ameliorate stress-induced increases in anxiety-like behavior (Mcvey Neufeld et al., 2019). However, the anti-depressive benefits are dependent on live LGG, while treatment with the heat-inactivated form of LGG had no effect.
Two other strains of Lactobacillus, L. rhamnosus CCFM1228 and L. paracasei CCFM1229, can alleviate anxiety- and depression-related behaviors in animal models, which may be achieved by regulating the activity of xanthine oxidase (XO) in brain (Xu M. et al., 2022). In fact, several anxiety- and depression-related indicators such as immobility time in the forced swimming test (FST), serum corticosterone level, and hippocampal BDNF concentration were significantly associated with XO activity in the cerebral cortex. XO activity is significantly increased in patients with depression, and xanthine and XO produce superoxide anions and free radicals that generate oxidative stress, leading to cellular damage and death. Xanthines synthesized via the pentose phosphate pathway can cross the BBB easily to the amygdala and act on oligodendrocytes via purine receptors on the cell surface, causing abnormal activation and proliferation of oligodendrocytes, leading to local neuronal hyperactivation in the fear center. L. paracasei CCFM1229 significantly upregulated the expression of Grin1, Grin2a, and Grin2b, and enhanced synaptic plasticity in depressed mice (Xu M. et al., 2022). It could also maintain the structural and functional stability of myelin by upregulating Mbp mRNA expression. Myelin loss and oligodendrocyte dysfunction may be involved in depression pathogenesis. In contrast to LGG, both live and heat-killed L. paracasei PS23 can reverse chronic corticosterone-induced anxiety- and depression-like behaviors (Wei et al., 2019). L. rhamnosus CCFM1228 can enhance astrocyte function in depressed mice by upregulating Gfap mRNA expression significantly. L. rhamnosus CCFM1228 significantly downregulates CD36 mRNA expression, which is upregulated in depressed mice, and CD36 deficiency may influence depression-like behavior by altering the gut microbiota and inflammatory pathways (Xu M. et al., 2022). In addition, taurine deficiency may lead to oxidative stress as well as reduced total N-acetylaspartate levels in neurodegenerative diseases, which can be restored by supplementation with L. rhamnosus JB-1 (Devkota et al., 2012; Kochalska et al., 2020). Of course, other Lactobacillus strains, alone or in combination with other microorganisms, have also been investigated for their roles and possible mechanisms in anti-depressive behavior.
Like Lactobacillus, Bifidobacterium is a commonly used probiotic bacterium. Bifidobacterium spp. is a genus of gram-positive, non-motile, rod-shaped, sometimes bifurcated at one end, strictly anaerobic bacteria that are widely found in the human and animal digestive tract. Most probiotic Bifidobacterium strains have shown positive effects on human health. Bifidobacterium inhibits the proliferation of harmful bacteria, ameliorates the function of the gastrointestinal mucosal barrier, and protects against pathogens. With the growing recognition of the existence of the MGB axis, recent studies have identified that Bifidobacterium can affect the functioning of the brain and CNS, leading to alterations in behavior, and cognitive abilities of humans and animals. Aizawa et al. found that lower fecal counts of Bifidobacterium in depressed patients than that in healthy controls, suggesting that Bifidobacterium plays an important role in the pathogenesis of depression (Aizawa et al., 2016). In GF rats, Bifidobacterium can successfully inhibit elevated HPA axis and depression-like behaviors (Messaoudi et al., 2011). Kazem et al. demonstrated that a significant improvement in depression and well-being status was obtained after the administration of probiotic Bifidobacterium spp. for 8 weeks (Kazem et al., 2021).
Several strains of Bifidobacterium such as B. adolescentis, B. dentium, and B. infantis exert beneficial effects in reducing anxiety- and depression-like behaviors, which are related with the production of GABA. These GABA producers can bioconvert monosodium glutamate to GABA (Barrett et al., 2012). Bifidobacterium can biosynthesize GABA from glutamate by the action of glutamate decarboxylase, and then transport it extracellularly by the action of the glutamate GABA antiporter. One efficient GABA producer, B. adolescentis 150, can attenuate depression-like behavior during the FST conducted on BALB/c mice (Yunes et al., 2020). Guo et al. also found that B. adolescentis exhibits antidepressant and anxiolytic effects, which are associated with a reduction in inflammatory cytokines and re-establishment the gut microbiota (Guo et al., 2019). Another Bifidobacterium strain, B. breve CCFM1025, exerts an antidepressant-like effect by reshaping the gut microbiota, increasing the production of beneficial metabolites, attenuating the HPA axis and inflammation, upregulating BDNF expression, and downregulating c-Fos expression in the brain (Tian et al., 2020). In one RCT, Tian et al. observed that B. breve CCFM1025 can attenuate depression and associated gastrointestinal disorders by altering the gut microbiota and gut tryptophan metabolism (Tian et al., 2022), suggesting that B. breve CCFM1025 is a promising psychobiotic candidate. Interestingly, B. breve CCFM1025 can normalize the abundance of SCFA-producing species such as Heterobacterium spp., Clostridium faecium, and Clostridium tumefaciens in patients with depression, whereas B. longum subsp. infantis E41 significantly reduced the elevated abundance of Veillonellaceae and Desulfovibrio (Tian et al., 2022). 5-hydroxytryptophan (5-HTP), a precursor substance of 5-HT and a key neurotransmitter that can cross the BBB, links bidirectional gut-brain communication, making it possible for the brain and gut to maintain a host’s health jointly. Intestinal and serum 5-HTP were positively correlated with brain 5-HT levels. Tian also found that B. breve M2CF22M7 and B. longum subsp. infantis E41 have antidepressant effects in mice, partly by improving the secretion of 5-HTP, reversing the deficits in hippocampal 5-HT, and increasing BDNF levels in the prefrontal cortex (Tian P. et al., 2019). A previous study also found that chronic stress impaired the negative feedback of corticosterone in the HPA axis by down-regulating the glucocorticoid receptors (Nr3c1), leading to glucocorticoid resistance, which is also coincident with a high level of inflammation. B. breve CCFM1025 normalizes the expression of Nr3c1 to reduce cortisol levels (Tian et al., 2020). In addition, the presence of glutamate decarboxylase genes in Bifidobacteria may mediate GABA production, which can complement GABA levels and enhance the negative feedback regulatory mechanism of cortisol, as exemplified by B. adolescentum (Duranti et al., 2020). Higher levels of pro-inflammatory cytokines lead to the onset and progression of depression-related behaviors, whereas Bifidobacteria are known for their ability to counter inflammation (Dyakov et al., 2020). An increase in pro-inflammatory cytokines, namely IL-6, IL-1β, tumor necrosis factor α, and C-reactive protein (CRP), participate actively in the development of depression. B. bifidum TMC3115 suppresses the stress-induced increase in IL-6 levels and reduces inflammation, which can alleviate stress-induced inflammatory responses (Yoda et al., 2022). In addition, Bifidobacterium can promote the production of Foxp3+ Tregs mediated by SCFAs to limit the inflammatory response in peripheral tissues (Arpaia et al., 2013; Smith et al., 2013). In contrast to the findings in LGG, Kosuge et al. observed that heat-sterilized B. breve M-16V can prevent depression-like behavior and IL-1β expression induced by chronic social defeat stress through modulation of gut microbiota composition in mice (Kosuge et al., 2021). These findings suggest that Bifidobacterium could be used as a potential psychobiotics for the prevention and treatment of depression.
A. muciniphila is a gram-negative anaerobic bacterium, representative of the phylum Verrucomicrobia, a symbiotic bacterium widely distributed in the mucus layer of the human intestine, which lives by breaking down mucin (Derrien et al., 2008). Due to its beneficial effects in many diseases, it has attracted much attention and research from the academic community and has become a new generation of probiotics. Several studies have shown that A. muciniphila can influence glucose and lipid metabolism, and intestinal immunity, while certain food ingredients, like polyphenols, can increase the abundance of A. muciniphila in the gut. The abundance of A. muciniphila is significantly reduced under certain medical conditions. A. muciniphila is negatively associated with DM and obesity. Recent studies indicate that several neurological disorders, including amyotrophic lateral sclerosis, depression, AD, and ASD, disrupt the abundance of A. muciniphila (Wang et al., 2011; Blacher et al., 2019; Li B. et al., 2019; Mcgaughey et al., 2019). A. muciniphila has emerged as the “sentinel of the gut”, which can promote gut barrier integrity, enrich butyrate-producing bacteria modulate immune responses, and inhibit inflammation (Ouyang et al., 2020). Given its higher abundance in healthy mucosa, Akkermansia has been suggested as biomarker of healthy intestines (Mcgaughey et al., 2019).
A. muciniphila feeds on mucin in the mucus layer of the intestine, thereby settling in the intestine and protecting it from pathogens via competitive rejection (Remely et al., 2015; Kim et al., 2022). Colonic mucus is heavily dependent on the release of MUC2, a hydrated glycosylated protein produced by cupped cells that adheres to the colonic surface to prevent invasion by luminal microbes and pathogens (Mcgaughey et al., 2019). A. muciniphila can improve MUC2 expression and increase the number of cupped and MUC2 cells to strengthen the colonic mucosal barrier in recipient mice (Chen T. et al., 2021). At the same time, A. muciniphila breaks down mucin to produce SCFAs, such as acetate and propionate, to play a regulatory role. SCFAs can alleviate oxidative stress and inflammatory responses through the GLP-1 pathway, as well as act in intestinal Treg homeostasis or directly through the BBB to increase the levels of neurotransmitters such as glutamine, glutamate, and GABA in the hypothalamus of mice, exerting an antidepressant effect (Zhai et al., 2019). In socially defeated animals, the decrease of Akkermansia spp. was correlated with the behavioral metrics of both anxiety and depression positively. McGaughey et al. found that reduction of fecal Akkermansia spp. in mice exhibited decreased center time during the open field test (OFT), indicating increased anxiety-like behavior, as well as decreased sucrose preference, suggesting increased anhedonia (Mcgaughey et al., 2019). Generally, supplementation with A. muciniphila may improve gut dysbiosis due to depression. Ding et al. found that A. muciniphila reduces depression-like behaviors induced by chronic stress by regulating gut dysbiosis and metabolic disorders related to the gut microbiota (Ding et al., 2021). A. muciniphila increases the level of Akkermansia and decreases the relative abundance of Helicobacter, Lachnoclostridium, Candidatus_Saccharimonas, and Eubacterium_brachy, promoting the re-establishment of gut microbiota (Ding et al., 2021). Increased Clostridium tumefaciens abundance after A. muciniphila treatment is negatively correlated with the number of microglia, which may exert an anti-neuroinflammatory effect. In addition to gut microbiota modulation, the antidepressant effect of A. muciniphila positively correlates with an increase in metabolites, such as edaravone and β-alanyl-3-methyl-l-histidine (Ding et al., 2021). Edaravone increased serotonin concentrations significantly, whereas β-alanyl-3-methyl-l-histidine increased the levels of dopamine (Ding et al., 2021). A. muciniphila administration may also reduce the degradation of geraniol, which has neuroprotective and anti-inflammatory activities and may alleviate depression (Deng et al., 2015; Ding et al., 2021). In addition, Amuc_1100, the outer membrane protein of A. muciniphila, also plays an important and direct role in the crosstalk between A. muciniphila and its host. Cheng et al. found that Amuc_1100 can improve chronic unpredictable mild stress (CUMS)-induced depression-like behavior and CUMS-induced downregulation of serotonin in the serum and colon of mice, restore gut dysbiosis, upregulate BDNF, and inhibit inflammation in the hippocampus (Cheng et al., 2021). Recently, they also found that the Amuc_1100Δ80, a truncated protein with 80 amino acids truncated at the N-terminus of Amuc_1100, shows a better antidepressant effect on modifying CUMS-induced depression-like behavior in mice than Amuc_1100 does (Cheng R. et al., 2022). Taken together, both human and animal studies have consistently reported that increasing A. muciniphila abundance can be a potential method for treating depression- and anxiety-like behaviors. Direct and compelling evidence from future comprehensive pre-clinical analyses and well-designed clinical studies is necessary to explore the therapeutic potential of A. muciniphila in depression.
Clostridium butyricum is a gram-positive anaerobic bacterium that can generate SCFAs by consuming undigested dietary fiber, mainly butyrate and acetate. It has a strong ability to survive independently of stomach and bile acids. C. butyricum plays an important role in regulating the gut microbiota, which has been safely used as a probiotic for decades. It is increasingly utilized in the treatment of various human diseases, including intestinal injury, gut-acquired infection, IBS, IBD, neurodegenerative diseases, metabolic diseases, and colorectal cancer (Lynch and Pedersen, 2016). Generally, C. butyricum promotes the proliferation of beneficial Bifidobacteria, Lactobacilli, and anthropoid bacteria, reducing pathogens and providing benefits to the intestinal microbial ecosystem (Hagihara et al., 2018; Miao et al., 2018).
Previous studies have found that exposure to chronic or acute psychosocial stress decreases the level of Clostridium spp. in the cecum (Bailey et al., 2011), while mental stress increases the levels of peripheral IL-6 and chemokine CCL2, and the expression of TLR3 and TLR4 in the prefrontal cortex of suicidal patients with depression, leading to hyperactivation of microglia (García Bueno et al., 2016; Park et al., 2018). Recently, many preclinical studies have shown that Clostridium spp. alone or in combination with other antidepressants can be used to treat depression (Sun et al., 2018; Tian T. et al., 2019). Liu et al. found that C. butyricum can be considered as a safe and economical therapeutic option to treat mental disorders, which can influence the gut microbiota-butyrate-brain axis in mice (Liu et al., 2015). A specific phenotype of C. butyricum, C. butyricum MIYAIRI 588 (CBM588), was isolated from the feces of a Japanese person firstly. CBM588 has been used as a probiotic in humans and domestic animals, exerting a variety of beneficial health effects. Hagihara et al. first reported that the administration of CBM588 improved the ecosystem of the gastrointestinal tract in mice significantly, modulating the gut microbiota composition, increasing the numbers of Bifidobacterium, Coprococcus, and Bacteroides, enhancing butyrate production, and reducing epithelial damage (Hagihara et al., 2018). They also found that CBM588 treatment caused a functional shift of the gut microbiota toward increased carbohydrate metabolism (Hagihara et al., 2019). Tian et al. found that 28-day preventive treatment with CBM588 improved depression-like behaviors in mice with chronic social defeat stress (Tian T. et al., 2019). CBM588 can alter the composition of the gut microbiota, induce a higher abundance of C. perfringens, and produce more butyrate, exerting a regulatory effect of SCFAs on the MGB axis and reducing depression-like behavior. Simultaneously, CBM588 prevents stress-induced activation of hippocampal inflammatory microglia by reducing cytokines, including IL-1β, IL-6, and TNF-α (Tian T. et al., 2019). In a model of CUMS-induced depression, Sun et al. observed that the administration of C. butyricum CGMCC 9830 reversed depression-like behavior, increased hippocampal BDNF and 5-HT levels, and improved intestinal GLP-1 levels (Sun et al., 2018). GLP-1 is secreted by intestinal L cells distributed in the ileum and colon, and changes in the gut microbiota can affect GLP-1 levels, which are closely related to CNS function (Reimann et al., 2008). By producing metabolic butyrate, Clostridium spp. has been implicated in depression through HPA-axis perturbation and damage to intestinal permeability by combining G-protein receptor 43 (GPR43) and GPR41 and regulating NF-κB and PPARγ signaling (Lührs et al., 2002; Van De Wouw et al., 2018). In a recent prospective open-label trial, Miyaoka et al. found that subjects with treatment-resistant major depressive disorder (MDD) receiving CBM588 (60 mg/d) in combination with antidepressants (flvoxamine, paroxetine, escitalopram, duroxetine, and sertraline) reported significantly lower median scores across several indices (BDI, HAMD-17, and BAI scores) than those in the control group (Miyaoka et al., 2018). Although these preclinical and clinical studies have shown the efficacy and safety of C. butyricum in depression, future larger-scale RCT on depression should be conducted to provide clearer recommendations for C. butyricum application and evaluate its possible mechanisms.
Enterococcus faecalis is a gram-positive, facultative anaerobic, lactic acid bacterium belonging to the phylum Firmicutes. E. faecalis is a normal resident of the gut in many hosts (García-Solache and Rice, 2019) and is generally believed to be harmless. Some strains of E. faecalis with beneficial effects are used as probiotics and starter cultures in the dairy industry, whereas other strains of E. faecalis contribute to the development of nosocomial infections and cause bacteremia, endocarditis, or urinary tract infections. Strain-specific differences in probiotic, pathogenic, and commensal E. faecalis may depend on their interaction with the host. Thus, E. faecalis has received substantial attention owing to its ‘dualistic’ behavior toward human health.
E. faecalis 2001 (EF-2001), one biogenic lactic acid bacterium, has been used as a probiotic to improve immunity and exert antitumor activity in mice (Choi et al., 2016; Gu et al., 2017). Takahashi et al. demonstrated that 20-day administration of EF-2001 can prevent colitis-induced depression-like behavior via the MGB axis in mice, which can reduce rectal and hippocampal inflammatory cytokines such as TNF-α and IL-6 and facilitate the NF-κB p65/XIAP pathway in the hippocampus (Takahashi et al., 2019). Olfactory bulbectomized (OBX) mice is a valuable experimental animal model for MDD, which expresses various depression-like behaviors such as anhedonia, memory impairment, and reduction in sexual contact (Takahashi et al., 2018). Recently, EF-2001 was shown to prevent OBX-induced depression-like behaviors through the regulation of prefrontal cortical myelination via the enhancement of CREB/BDNF and NF-κB p65/LIF/STAT3 pathways (Takahashi et al., 2022). In addition, this group also found that the anti-dementia effects of EF-2001 in OBX mice are associated with the enhancement of hippocampal neurogenesis via the ERK-CREB-BDNF pathway (Takahashi et al., 2020). Another Japanese group observed that male mice fed a diet supplemented with heat-killed E. faecalis strain EC-12 showed decreased anxiety-like behavior in OFT and elevated plus-maze test, which can increase the expression of neurotransmitter receptor genes such as Adrb3, Avpr1a, and Drd5, and improve the gut dysbiosis (Kambe et al., 2020). These findings suggest that some strains of E. faecalis have the potential to alleviate depressive symptoms. Future studies are required to explore the antidepressant effects and the exact mechanism of action of E. faecalis in the human brain.
A large body of evidence has demonstrated that depression is not only more common with other psychiatric disorders; for example, up to 90% of patients with anxiety disorders present with co-morbid depression (Tiller, 2013), but it is also highly comorbid and occurs together with many physical diseases, such as IBS, IBD, and metabolic syndromes such as DM and obesity. A common pathological mechanism of co-morbidity is a prerequisite for the administration of the same drug to achieve common remission. To understand if probiotics have this potential, we have addressed recent advances in the application of probiotics in depression-related disorders, which will provide novel therapeutic options for these co-morbid disorders associated with depression.
IBS is a chronic dysfunction of the gastrointestinal system characterized by altered bowel habits and abdominal pain in the absence of biochemical or structural abnormalities, primarily manifesting as diarrhea, constipation, or both. IBS affects approximately 7% to 21% of the global population (Chey et al., 2015). Based on the Rome IV criteria, which was updated in June 2016, the gold standard for the diagnosis of IBS is the exclusion of other diseases (Mearin et al., 2016). Patients with IBS are categorized into four subtypes based on their predominant stool habits: IBS-C (constipation, 20-30%), IBS-D (diarrhea, 38-50%), IBS-M (mixed type, 6-16%), and IBS-U (unclassified, 24-60%). The pathophysiology of IBS is not fully understood, but evidence suggested that abnormalities in the composition or metabolic activity of the gut microbiota are associated with its progression (Han et al., 2022). Increasing studies have suggested that the MGB axis plays a role in IBS.
IBS is frequently associated with psychiatric comorbidities such as depression and gut-brain crosstalk is thought to contribute to its development (Ray, 2017). A significant association between IBS and MDD has been reported previously, and most patients with IBS identify stress and anxiety as symptom aggravators (Lacy et al., 2007). Lee et al. reported significantly higher levels of depression and anxiety in patients with IBS than in healthy controls (Lee et al., 2017). Midenfjord et al. also found that patients with IBS who suffered from psychological distress reported more severe gastrointestinal symptoms (Midenfjord et al., 2019). Recent evidence suggests that gut dysbiosis can be considered one of the fundamental theories that can explain both physical and mental symptoms in patients with IBS. Alterations in the gut microbiota, MGB axis, and neuro-immune system may be the cornerstone of the association between IBS and depression (Mudyanadzo et al., 2018). A systematic review identified that patients with comorbid IBS and anxiety/depression had lower α-diversity, higher levels of Proteobacteria, Prevotella/Prevotellaceae, and Bacteroides, and lower abundance of Lachnospiraceae than the controls (Simpson et al., 2020). This suggests that microbiota modulation with specific probiotics or other microecological regulators may be a useful therapeutic approach for depression-related disorders, such as IBS.
In fact, recent findings suggest that probiotics may improve host health in patients with IBS both physically and mentally (Le Morvan De Sequeira et al., 2021; Zhang T. et al., 2022). The safety and efficacy of probiotics in the treatment of IBS are supported by numerous clinical studies. A systematic review and network meta-analysis of 43 RCTs involving 5,531 IBS patients provided data regarding the best probiotic species used in the treatment of IBS (Zhang T. et al., 2022). Several probiotic strains, such as B. bifidum, B. infantis, L. casei, L. acidophilus, L. plantarum, Bacillus coagulans, C. butyricum, and Saccharomyces boulardii alone or in combination, have been used to treat depression in patients with IBS. Among these probiotic strains, B. coagulans exhibited the highest probability of being the optimal probiotic species for improving IBS symptom relief rate, as well as global symptoms, bloating, abdominal pain, and straining scores. L. plantarum ranked first in improving the QoL of patients with IBS and had the lowest incidence of adverse events. B. coagulans can promote intestinal digestion, maintain host microbiota homeostasis, and regulate the host immune system, and has been studied in the treatment of several human diseases. Recently, a double-blind RCT reported that B. coagulans MTCC 5856 as a single probiotic agent at a dose of 2 × 109 spores (CFU) per day showed robust efficacy in the treatment of patients with IBS symptoms and MDD (Majeed et al., 2018). B. coagulans MTCC 5856 could improve the Montgomery-Asberg Depression Rating Scale (MADRS), and Hamilton Rating Scale for Depression (HAM-D) scores, indicating that it may be a new optional approach for the management of depression in patients with IBS. B. coagulans MTCC 5856 can produce SCFAs (such as acetic, propionic, and butyric acid), neurotransmitters, and antimicrobial and anti-inflammatory substances, which could be the possible mechanism of action in alleviating depression symptoms. In addition, this probiotic significantly reduced the level of myeloperoxidase (an inflammatory biomarker), which is responsible for the production of free radicals. Another placebo-controlled trial observed that the probiotic B. longum NCC3001 reduced depression, but not anxiety scores, and increased the QoL in patients with IBS (Pinto-Sanchez et al., 2017). These improvements were associated with changes in the brain activation patterns, indicating that this probiotic reduces limbic reactivity. Despite these promising findings, there is still limited evidence for the efficacy of probiotic intervention in patients with IBS and depression, as the benefit of probiotics tends to be symptom- and strain-specific. Further prospective, larger-scale trials with extended follow-up durations, as well as a detailed assessment of the therapeutic effects of specific probiotic supplementation, are critical prior to managing depression in patients with IBS with probiotics in clinical practice.
IBD, including Crohn’s disease (CD) and ulcerative colitis (UC), is an idiopathic, lifelong, and destructive chronic inflammatory condition of the gastrointestinal tract that affects tens of millions of people worldwide (Mulder et al., 2014). The pathogenesis of IBD is incompletely understood, although the major factors influencing IBD may include genetic, environmental, and microbial determinants (Jakubczyk et al., 2020). Recently, gut microbiota has been increasingly recognized as a critical and central factor in IBD. There is a growing consensus that inappropriate activation of the immune system by commensal bacteria underlies IBD.
The disease burden of IBD includes not only the physiological manifestations of the disease but also psychological and social burdens. Although the link between IBD and psychological disorders remains unclear, patients with IBD have a high prevalence of depression and anxiety. Psychological symptoms appear to be more prevalent in active disease states, with no difference in prevalence between CD and UC. The comorbid prevalence of depressive symptoms was 25.2%, and 38.9% of patients with active IBD had depression (Barberio et al., 2021). Compared to those who do not show psychiatric symptoms, patients with IBD suffering from depression have a decreased remission, and patients demonstrating depression show a more severe illness over a longer period of time (Stasi and Orlandelli, 2008). Chen et al. have summarized that depression in IBD may arise through an “IBD-inflammation-kynurenine pathway-depression” association (Chen LM. et al., 2021). Thus, encouraging screening and treatment of these comorbid psychiatric disorders may improve the prognosis of patients with IBD.
Gut dysbiosis is commonly observed in IBD patients with depression, generally with increased Firmicutes and reduced Proteobacteria (Chen DL. et al., 2021; Yuan et al., 2021). Weis et al. also observed that disorganized gut microbiota and disturbed metabolism were found in patients with active UC accompanied by depression and anxiety, which could increase systematic inflammation (Weis et al., 2019). The altered gut microbiota in patients with IBD can disturb bidirectional communication in the gut-brain axis, which might be associated with potential consequences for the CNS. The gut dysbiosis in IBD patients with depression represents a potential therapeutic target. The modulation of the gut microbiota using probiotics can alter the behavioral response in IBD, which has been increasingly studied in mouse models. Mice subjected to dextran sodium sulfate (DSS)-induced acute IBD-like colitis demonstrate behavioral changes, including anxiety-like behaviors and cognitive deficits. In DSS-induced animal models, probiotics, such as B. longum NCC3001, can reduce anxiety-like behaviors induced in rats in response to DSS-induced colitis (Bercik et al., 2011b). The probiotic B. longum NCC3001 normalizes behavior by decreasing the excitability of enteric neurons, but does not affect MPO activity, histological scores, or BDNF levels. In another previously mentioned behavioral study, Emge et al. found that recognition memory deficits and anxiety-like behavior during acute inflammation in murine IBD were improved by the administration of a probiotic mixture containing L. rhamnosus R0011 and L. helveticus R0052 (Emge et al., 2016). The improvements in behavior after probiotic administration were broadly correlated with the restoration of the microbiota and modulated hippocampal c-Fos expression. Inflammation may serve as a common trigger for the altered cognitive function observed in these models. Yoo et al. found that oral administration of a probiotic mixture containing L. plantarum NK151, B. longum NK173, and B. bifidum NK175 could also alleviate stress-induced fatigue, depression, and IBD by modulating inflammatory cytokines and gut microbiota byproducts such as LPS (Yoo et al., 2022). A previous study found that E. coli K1 significantly caused psychiatric disorders, such as depression and memory impairment, and IBD-like colitis in SPF and GF mice (Jang et al., 2018). Yun et al. observed that oral gavage of L. gasseri NK109 significantly alleviated E. coli K1-induced depression-like behaviors in GF and SPF mice by regulating the immune response through NF-κB-mediated BDNF expression, IL-1β expression, and vagus nerve-mediated gut-brain signaling (Yun et al., 2020). In addition, this group demonstrated that L. reuteri NK33 and B. adolescentis NK98 synergistically improved the occurrence and development of anxiety/depression and colitis by regulating gut immune responses and microbiota composition (Jang et al., 2019). However, E. faecium and Pediococcus acidilactici, known as probiotic strains, deteriorated Enterobacteriaceae-induced depression and colitis in mice (Jang et al., 2022). Another probiotic strain, Weissella paramesenteroides WpK4, can reduce anxiety-like and depression-like behaviors in murine models of ulcerative colitis by regulating the MGB axis and reducing gut permeability (Sandes et al., 2020). One Chinese group also observed that L. plantarum DMDL 9010 intake could reduce colitis and depression-like behavior in mice with DSS-induced colitis by upregulating the levels of neurotransmitters (especially 5-HT, DA, NE, and 5-HIAA) and SCFAs (such as butyric acid and propionic acid) (Huang et al., 2022). Considering these encouraging data from preclinical trials, microbiota modulation with probiotics might offer a novel therapeutic approach to combat behavioral comorbidities, such as depression, in patients suffering from IBD. However, the benefits of these treatments are limited because of the scarcity of interventional studies. Further well-designed clinical trials should be conducted to confirm the benefits of these probiotics in patients with IBD and comorbid depression.
Metabolic syndrome is defined as a cluster of obesity, hypertension, dyslipidemia, and dysglycemia. Over the past few decades, the prevalence of metabolic syndrome has increased markedly worldwide, which may be explained by urbanization, aging, lifestyle changes, and nutritional transition. Metabolic syndrome has become a serious public health problem, highlighting the urgent need to tackle metabolic syndrome in China and other populations (Li et al., 2018). Obesity is defined as abnormal or excessive fat accumulation that presents a risk to health and is a well-known cause of cardiovascular disease burden and premature death. DM is a chronic disease caused by an inherited or acquired lack of insulin produced by the pancreas or the inability of the body to fully utilize the insulin produced. A growing body of evidence suggests that co-morbid metabolic syndrome and depression are common and are often considered “metabolic depression” (Demakakos et al., 2010; Forsythe et al., 2010). Major depression and the exacerbation of depression symptoms have been reported in 11% and 31% of patients with DM, respectively (Semenkovich et al., 2015). The lifetime prevalence of major depression is approximately 28.5%. It has been reported that the prevalence of depression in patients with diabetes is higher than in the normal population (Anderson et al., 2001). In a meta-analysis of 17 community-based studies with 204,507 participants, there was a significant association between depression and obesity (De Wit et al., 2010). There is evidence of a dose-response effect of obesity severity on the odds of depression (Onyike et al., 2003). Obesity increases the incidence of depression as a function of metabolic dysfunction (Opel et al., 2015; Delgado et al., 2018). Depression is linked to a higher rate of complications in metabolic syndrome, to more disability, and to loss of years of life. Depression worsens glycemic control and causes greater severity of DM complications, resulting in poorer adherence to DM self-care (adherence to diet, checking blood sugar level), increased risk of retinopathy and macrovascular complications, decreased QoL, and increased disability burden (De Groot et al., 2016). Obesity leads to poorer performance in diverse cognitive tasks, and these deficits are exacerbated in instances of comorbid depressive disorder (Restivo et al., 2017). Thus, a better understanding of the link between comorbid depression and metabolic syndrome is critical to inform appropriate preventive and intervention strategies. Simultaneous therapeutic option for depression and metabolic syndrome is merited to enhance treatment outcomes in both conditions.
The exact mechanisms underlying the association between depression and metabolic syndrome are poorly understood, and possible pathophysiological overlap has been proposed. However, whether there is a causal relationship between both diseases and the nature of that causal relationship is still unclear. Recent advances have reported that both depression and these metabolic diseases are related to chronic low-grade inflammation and gut dysbiosis (Chan et al., 2019). The gut microbiota plays a vital role in regulating both the metabolic and brain functions of the host by reducing inflammatory activation and affecting the regulation of energy balance and release of neurotransmitters, suggesting that the gut microbiota can be considered a promising target to treat metabolic depression. To date, there have been no specific pharmacotherapies for metabolic depression. Current therapies for both depression and metabolic syndrome remain suboptimal for many patients, thus making improvements and advances in intervention options in high demand. Increasing evidence indicates that probiotics play a promising role in the management of these comorbidities. Probiotics appear to be effective in reducing depressive symptoms and can improve some of the clinical components of metabolic syndrome, making it a potential new therapeutic option or patient-specific strategy to treat both metabolic and depressive disorders.
Numerous animal and clinical studies have provided evidence for the treatment of metabolic depression with different probiotics. Patterson et al. found that daily administration of GABA-producing L. brevis DSM32386 and L. brevis DPC6108 ameliorated both metabolic abnormalities and depression-like behavior associated with metabolic syndrome in mice (Patterson et al., 2019). Specifically, L. brevis attenuated various abnormalities associated with metabolic dysfunction, causing a reduction in the accumulation of mesenteric adipose tissue, increased insulin secretion following glucose challenge, improved plasma cholesterol clearance, and reduced despair-like behavior and basal corticosterone production during FST. This exploratory study suggested that increased microbial GABA production could affect both host metabolism and behavior. Another study found that A. muciniphila subtype improves olanzapine-induced glucose homeostasis in mice by downregulating G6Pase and phosphoenolpyruvate carboxykinase overexpression, attenuating insulin resistance, and reducing systemic inflammation by restoring intestinal barrier function (Corb Aron et al., 2021). In addition, one recent ongoing RCT conducted by Gawlik-Kotelnicka et al. found that probiotic administration (including L. casei, L. acidophilus, and B. bifidum) in patients with depression for eight weeks had beneficial effects on the Beck Depression Inventory score, insulin, hs-CRP, and glutathione concentrations (Gawlik-Kotelnicka et al., 2021). They also evaluated the influence of supplementation with a probiotic mixture including B. longum Rosell®-175 and L. helveticus Rosell®-52 on depressive symptoms, QoL, inflammation, oxidative stress indices, and fecal microbiota in patients with depression, depending on the metabolic syndrome comorbidity (Gawlik-Kotelnicka et al., 2021). If successful, the trial will establish an easy-to-use and safe treatment option (probiotic supplement) as an adjunctive therapy in patients who are only partially responsive to pharmacological treatment. In a 2-month clinical trial, treatment with a synbiotic formula of L. acidophilus PBS066, L. plantarum PBS067, and L. reuteri PBS072 with active prebiotics could decrease the prevalence of metabolic syndrome and several cardiovascular risk factors and markers of insulin resistance in older patients, which improved their QoL (Cicero et al., 2021). The above-mentioned evidence suggests that some specific probiotics could be used as adjunctive treatment options to treat the comorbidity of depression and metabolic syndrome. Thus, future probiotic intervention studies in large cohorts of patients with comorbid depression and metabolic syndrome and more rigorous RCTs are needed to determine whether probiotics can provide benefits for comorbidity treatment in clinical practice.
Accumulating evidence has identified that the gut microbiota actively participates in bidirectional gut-brain communication, which is considered the “second brain” of the human body. The gut microbiota has significant impact on the immune system, brain development, and behavior, and its alterations lead to the onset and progression of several neuropsychiatric disorders, including depression. The exact mechanisms by which the gut microbiota causes or alters depression are not fully understood, although current evidence demonstrates that the gut microbiota can affect the development of depression, mainly through the HPA axis, and inflammation, and modify the level of BDNF. Based on the present scientific discoveries, the gut microbiota may be a novel therapeutic target for the prevention and treatment of depression, with pre-clinical and clinical studies suggesting that several strains of probiotics can provide critical benefits for preventing and treating depression. Due to the lack of direct clinical evidence, it still cannot be recommended that probiotics can replace antidepressant medications as the primary treatment for patients with depression. In addition, depression often coexists with IBS, IBD, and metabolic syndromes, further increasing the risk of mortality. The pathophysiological overlap between depression and its comorbidity makes it feasible to treat these diseases with specific probiotics, which can improve both depressive symptoms and comorbid abnormalities. However, it is noteworthy that these benefits are strain-specific, while other influencing factors, including the type of intervention (add-on versus standalone), intervention content (strain combinations and dosing), delivery modes (tablets, capsules, powders, and freeze-dried formulations), patient population, and disease severity threshold for inclusion should also be considered. There is an urgent need to identify safe and effective novel probiotic strains to prevent and treat depression and its comorbidities. In the future, more rigorous RCTs in larger samples of patients diagnosed with depression with/without comorbidity should explore the optimal probiotic supplement content and dosage, long-term safety, and efficacy, along with an appropriate follow-up to assess relapse rates. Probiotic supplementation may serve as a simple and effective dietary intervention to promote mental well-being among patients with depression and depressed comorbidities.
ZL, and WX conceived and designed the review. JG, LZ, YC, ZL, and WX analyzed the data. JG, LZ, YC, WL, YW, XL, NZ, LS, XC, YS, ZL, and WX discussed the contents, wrote, reviewed, and edited the manuscript. All authors contributed to the article and approved the submitted version.
This present work was funded by the grants of Key R&D Program of Zhejiang (2022C03060), Zhejiang Basic Public Welfare Research Project (LGF19H010001), Lishui Basic Public Welfare Research Project (2023GYX03), the Research Project of Jinan Microecological Biomedicine Shandong Laboratory (JNL-2022033C), the Taishan Scholar Foundation of Shandong Province (tsqn202103119), the Fundamental Research Funds for the Central Universities (2022ZFJH003) and the Foundation of China’s State Key Laboratory for Diagnosis and Treatment of Infectious Diseases.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adak, A., Khan, M. R. (2019). An insight into gut microbiota and its functionalities. Cell Mol. Life Sci. 76 (3), 473–493. doi: 10.1007/s00018-018-2943-4
Aizawa, E., Tsuji, H., Asahara, T., Takahashi, T., Teraishi, T., Yoshida, S., et al. (2016). Possible association of bifidobacterium and lactobacillus in the gut microbiota of patients with major depressive disorder. J. Affect. Disord. 202, 254–257. doi: 10.1016/j.jad.2016.05.038
Alli, S. R., Gorbovskaya, I., Liu, J. C. W., Kolla, N. J., Brown, L., Müller, D. J. (2022). The gut microbiome in depression and potential benefit of prebiotics, probiotics and synbiotics: A systematic review of clinical trials and observational studies. Int. J. Mol. Sci. 23 (9), 4494. doi: 10.3390/ijms23094494
Amirani, E., Milajerdi, A., Mirzaei, H., Jamilian, H., Mansournia, M. A., Hallajzadeh, J., et al. (2020). The effects of probiotic supplementation on mental health, biomarkers of inflammation and oxidative stress in patients with psychiatric disorders: A systematic review and meta-analysis of randomized controlled trials. Complement Ther. Med. 49, 102361. doi: 10.1016/j.ctim.2020.102361
Anderson, R. J., Freedland, K. E., Clouse, R. E., Lustman, P. J. (2001). The prevalence of comorbid depression in adults with diabetes: a meta-analysis. Diabetes Care 24 (6), 1069–1078. doi: 10.2337/diacare.24.6.1069
Arpaia, N., Campbell, C., Fan, X., Dikiy, S., van der Veeken, J., Deroos, P., et al. (2013). Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 504 (7480), 451–455. doi: 10.1038/nature12726
Bai, Y. Y., Ruan, C. S., Yang, C. R., Li, J. Y., Kang, Z. L., Zhou, L., et al. (2016). ProBDNF signaling regulates depression-like behaviors in rodents under chronic stress. Neuropsychopharmacology 41 (12), 2882–2892. doi: 10.1038/npp.2016.100
Bailey, M. T., Dowd, S. E., Galley, J. D., Hufnagle, A. R., Allen, R. G., Lyte, M. (2011). Exposure to a social stressor alters the structure of the intestinal microbiota: implications for stressor-induced immunomodulation. Brain Behav. Immun. 25 (3), 397–407. doi: 10.1016/j.bbi.2010.10.023
Barandouzi, Z. A., Starkweather, A. R., Henderson, W. A., Gyamfi, A., Cong, X. S. (2020). Altered composition of gut microbiota in depression: A systematic review. Front. Psychiatry 11. doi: 10.3389/fpsyt.2020.00541
Barberio, B., Zamani, M., Black, C. J., Savarino, E. V., Ford, A. C. (2021). Prevalence of symptoms of anxiety and depression in patients with inflammatory bowel disease: a systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 6 (5), 359–370. doi: 10.1016/s2468-1253(21)00014-5
Barrett, E., Ross, R. P., O'toole, P. W., Fitzgerald, G. F., Stanton, C. (2012). γ-aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 113 (2), 411–417. doi: 10.1111/j.1365-2672.2012.05344.x
Bercik, P., Denou, E., Collins, J., Jackson, W., Lu, J., Jury, J., et al. (2011a). The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 141 (2), 599–609. doi: 10.1053/j.gastro.2011.04.052
Bercik, P., Park, A. J., Sinclair, D., Khoshdel, A., Lu, J., Huang, X., et al. (2011b). The anxiolytic effect of bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol. Motil. 23 (12), 1132–1139. doi: 10.1111/j.1365-2982.2011.01796.x
Berk, M., Williams, L. J., Jacka, F. N., O'neil, A., Pasco, J. A., Moylan, S., et al. (2013). So depression is an inflammatory disease, but where does the inflammation come from? BMC Med. 11, 200. doi: 10.1186/1741-7015-11-200
Bharwani, A., Mian, M. F., Surette, M. G., Bienenstock, J., Forsythe, P. (2017). Oral treatment with lactobacillus rhamnosus attenuates behavioural deficits and immune changes in chronic social stress. BMC Med. 15 (1), 7. doi: 10.1186/s12916-016-0771-7
Blacher, E., Bashiardes, S., Shapiro, H., Rothschild, D., Mor, U., Dori-Bachash, M., et al. (2019). Potential roles of gut microbiome and metabolites in modulating ALS in mice. Nature 572 (7770), 474–480. doi: 10.1038/s41586-019-1443-5
Bravo, J. A., Forsythe, P., Chew, M. V., Escaravage, E., Savignac, H. M., Dinan, T. G., et al. (2011). Ingestion of lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. U.S.A. 108 (38), 16050–16055. doi: 10.1073/pnas.1102999108
Buffington, S. A., Di Prisco, G. V., Auchtung, T. A., Ajami, N. J., Petrosino, J. F., Costa-Mattioli, M. (2016). Microbial reconstitution reverses maternal diet-induced social and synaptic deficits in offspring. Cell 165 (7), 1762–1775. doi: 10.1016/j.cell.2016.06.001
Carabotti, M., Scirocco, A., Maselli, M. A., Severi, C. (2015). The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 28 (2), 203–209.
Carlson, A. L., Xia, K., Azcarate-Peril, M. A., Rosin, S. P., Fine, J. P., Mu, W., et al. (2021). Infant gut microbiome composition is associated with non-social fear behavior in a pilot study. Nat. Commun. 12 (1), 3294. doi: 10.1038/s41467-021-23281-y
Carroll, B. J., Feinberg, M., Greden, J. F., Tarika, J., Albala, A. A., Haskett, R. F., et al. (1981). A specific laboratory test for the diagnosis of melancholia. standardization, validation, and clinical utility. Arch. Gen. Psychiatry 38 (1), 15–22. doi: 10.1001/archpsyc.1981.01780260017001
Chahwan, B., Kwan, S., Isik, A., Van Hemert, S., Burke, C., Roberts, L. (2019). Gut feelings: A randomised, triple-blind, placebo-controlled trial of probiotics for depressive symptoms. J. Affect. Disord. 253, 317–326. doi: 10.1016/j.jad.2019.04.097
Chan, K. L., Cathomas, F., Russo, S. J. (2019). Central and peripheral inflammation link metabolic syndrome and major depressive disorder. Physiol. (Bethesda) 34 (2), 123–133. doi: 10.1152/physiol.00047.2018
Chao, L., Liu, C., Sutthawongwadee, S., Li, Y., Lv, W., Chen, W., et al. (2020). Effects of probiotics on depressive or anxiety variables in healthy participants under stress conditions or with a depressive or anxiety diagnosis: A meta-analysis of randomized controlled trials. Front. Neurol. 11. doi: 10.3389/fneur.2020.00421
Chen, L. M., Bao, C. H., Wu, Y., Liang, S. H., Wang, D., Wu, L. Y., et al. (2021). Tryptophan-kynurenine metabolism: a link between the gut and brain for depression in inflammatory bowel disease. J. Neuroinflamm. 18 (1), 135. doi: 10.1186/s12974-021-02175-2
Chen, D. L., Dai, Y. C., Zheng, L., Chen, Y. L., Zhang, Y. L., Tang, Z. P. (2021). Features of the gut microbiota in ulcerative colitis patients with depression: A pilot study. Med. (Baltimore) 100 (7), e24845. doi: 10.1097/md.0000000000024845
Chen, H. M., Kuo, P. H., Hsu, C. Y., Chiu, Y. H., Liu, Y. W., Lu, M. L., et al. (2021). Psychophysiological effects of lactobacillus plantarum PS128 in patients with major depressive disorder: A preliminary 8-week open trial. Nutrients 13 (11), 3731. doi: 10.3390/nu13113731
Chen, T., Wang, R., Duan, Z., Yuan, X., Ding, Y., Feng, Z., et al. (2021). Akkermansia muciniphila protects against psychological disorder-induced gut microbiota-mediated colonic mucosal barrier damage and aggravation of colitis. Front. Cell Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.723856
Cheng, Y., Liu, J., Ling, Z. (2022). Short-chain fatty acids-producing probiotics: A novel source of psychobiotics. Crit. Rev. Food Sci. Nutr. 62 (28), 7929–7959. doi: 10.1080/10408398.2021.1920884
Cheng, L. H., Liu, Y. W., Wu, C. C., Wang, S., Tsai, Y. C. (2019). Psychobiotics in mental health, neurodegenerative and neurodevelopmental disorders. J. Food Drug Anal. 27 (3), 632–648. doi: 10.1016/j.jfda.2019.01.002
Cheng, R., Xu, W., Wang, J., Tang, Z., Zhang, M. (2021). The outer membrane protein Amuc_1100 of akkermansia muciniphila alleviates the depression-like behavior of depressed mice induced by chronic stress. Biochem. Biophys. Res. Commun. 566, 170–176. doi: 10.1016/j.bbrc.2021.06.018
Cheng, R., Zhu, H., Sun, Y., Hang, T., Zhang, M. (2022). The modified outer membrane protein Amuc_1100 of akkermansia muciniphila improves chronic stress-induced anxiety and depression-like behavior in mice. Food Funct. 13 (20), 10748–10758. doi: 10.1039/d2fo01198k
Chey, W. D., Kurlander, J., Eswaran, S. (2015). Irritable bowel syndrome: a clinical review. JAMA 313 (9), 949–958. doi: 10.1001/jama.2015.0954
Choi, E. J., Iwasa, M., Han, K. I., Kim, W. J., Tang, Y., Han, W. C., et al. (2016). Effect of enterococcus faecalis EF-2001 on experimentally induced atopic eczema in mice. Food Sci. Biotechnol. 25 (4), 1087–1093. doi: 10.1007/s10068-016-0175-7
Cicero, A. F. G., Fogacci, F., Bove, M., Giovannini, M., Borghi, C. (2021). Impact of a short-term synbiotic supplementation on metabolic syndrome and systemic inflammation in elderly patients: a randomized placebo-controlled clinical trial. Eur. J. Nutr. 60 (2), 655–663. doi: 10.1007/s00394-020-02271-8
Cipriani, A., Furukawa, T. A., Salanti, G., Chaimani, A., Atkinson, L. Z., Ogawa, Y., et al. (2018). Comparative efficacy and acceptability of 21 antidepressant drugs for the acute treatment of adults with major depressive disorder: A systematic review and network meta-analysis. Lancet 391 (10128), 1357–1366. doi: 10.1016/s0140-6736(17)32802-7
Clarke, G., Grenham, S., Scully, P., Fitzgerald, P., Moloney, R. D., Shanahan, F., et al. (2013). The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 18 (6), 666–673. doi: 10.1038/mp.2012.77
Clemente, J. C., Manasson, J., Scher, J. U. (2018). The role of the gut microbiome in systemic inflammatory disease. Bmj 360, j5145. doi: 10.1136/bmj.j5145
Colmers, P. L. W., Bains, J. S. (2018). Balancing tonic and phasic inhibition in hypothalamic corticotropin-releasing hormone neurons. J. Physiol. 596 (10), 1919–1929. doi: 10.1113/jp275588
Corb Aron, R. A., Abid, A., Vesa, C. M., Nechifor, A. C., Behl, T., Ghitea, T. C., et al. (2021). Recognizing the benefits of pre-/Probiotics in metabolic syndrome and type 2 diabetes mellitus considering the influence of akkermansia muciniphila as a key gut bacterium. Microorganisms 9 (3), 618. doi: 10.3390/microorganisms9030618
Couch, C. E., Neal, W. T., Herron, C. L., Kent, M. L., Schreck, C. B., Peterson, J. T. (2023). Gut microbiome composition associates with corticosteroid treatment, morbidity, and senescence in Chinook salmon (Oncorhynchus tshawytscha). Sci. Rep. 13 (1), 2567. doi: 10.1038/s41598-023-29663-0
Cryan, J. F., Dinan, T. G. (2015). More than a gut feeling: the microbiota regulates neurodevelopment and behavior. Neuropsychopharmacology 40 (1), 241–242. doi: 10.1038/npp.2014.224
Cryan, J. F., O'mahony, S. M. (2011). The microbiome-gut-brain axis: from bowel to behavior. Neurogastroenterol Motil. 23 (3), 187–192. doi: 10.1111/j.1365-2982.2010.01664.x
Cui, X. F., Zhou, W. M., Yang, Y., Zhou, J., Li, X. L., Lin, L., et al. (2014). Epidermal growth factor upregulates serotonin transporter and its association with visceral hypersensitivity in irritable bowel syndrome. World J. Gastroenterol. 20 (37), 13521–13529. doi: 10.3748/wjg.v20.i37.13521
Cullinan, W. E., Ziegler, D. R., Herman, J. P. (2008). Functional role of local GABAergic influences on the HPA axis. Brain Struct. Funct. 213 (1-2), 63–72. doi: 10.1007/s00429-008-0192-2
Dantzer, R., O'connor, J. C., Freund, G. G., Johnson, R. W., Kelley, K. W. (2008). From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 9 (1), 46–56. doi: 10.1038/nrn2297
Davey, K. J., Cotter, P. D., O'sullivan, O., Crispie, F., Dinan, T. G., Cryan, J. F., et al. (2013). Antipsychotics and the gut microbiome: olanzapine-induced metabolic dysfunction is attenuated by antibiotic administration in the rat. Transl. Psychiatry 3 (10), e309. doi: 10.1038/tp.2013.83
Décarie-Spain, L., Hayes, A. M. R., Lauer, L. T., Kanoski, S. E. (2023). The gut-brain axis and cognitive control: A role for the vagus nerve. Semin. Cell Dev. Biol. doi: 10.1016/j.semcdb.2023.02.004
De Groot, M., Crick, K. A., Long, M., Saha, C., Shubrook, J. H. (2016). Lifetime duration of depressive disorders in patients with type 2 diabetes. Diabetes Care 39 (12), 2174–2181. doi: 10.2337/dc16-1145
Dehghani, F., Abdollahi, S., Shidfar, F., Clark, C. C. T., Soltani, S. (2022). Probiotics supplementation and brain-derived neurotrophic factor (BDNF): A systematic review and meta-analysis of randomized controlled trials. Nutr. Neurosci. doi: 10.1080/1028415x.2022.2110664
Delgado, I., Huet, L., Dexpert, S., Beau, C., Forestier, D., Ledaguenel, P., et al. (2018). Depressive symptoms in obesity: Relative contribution of low-grade inflammation and metabolic health. Psychoneuroendocrinology 91, 55–61. doi: 10.1016/j.psyneuen.2018.02.030
Demakakos, P., Pierce, M. B., Hardy, R. (2010). Depressive symptoms and risk of type 2 diabetes in a national sample of middle-aged and older adults: The English longitudinal study of aging. Diabetes Care 33 (4), 792–797. doi: 10.2337/dc09-1663
Deng, X. Y., Xue, J. S., Li, H. Y., Ma, Z. Q., Fu, Q., Qu, R., et al. (2015). Geraniol produces antidepressant-like effects in a chronic unpredictable mild stress mice model. Physiol. Behav. 152 (Pt A), 264–271. doi: 10.1016/j.physbeh.2015.10.008
Derrien, M., Collado, M. C., Ben-Amor, K., Salminen, S., De Vos, W. M. (2008). The mucin degrader akkermansia muciniphila is an abundant resident of the human intestinal tract. Appl. Environ. Microbiol. 74 (5), 1646–1648. doi: 10.1128/aem.01226-07
Desbonnet, L., Clarke, G., Shanahan, F., Dinan, T. G., Cryan, J. F. (2014). Microbiota is essential for social development in the mouse. Mol. Psychiatry 19 (2), 146–148. doi: 10.1038/mp.2013.65
Devkota, S., Wang, Y., Musch, M. W., Leone, V., Fehlner-Peach, H., Nadimpalli, A., et al. (2012). Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 487 (7405), 104–108. doi: 10.1038/nature11225
De Wit, L., Luppino, F., Van Straten, A., Penninx, B., Zitman, F., Cuijpers, P. (2010). Depression and obesity: A meta-analysis of community-based studies. Psychiatry Res. 178 (2), 230–235. doi: 10.1016/j.psychres.2009.04.015
Dinan, T. G., Cryan, J. F. (2016). Mood by microbe: towards clinical translation. Genome Med. 8 (1), 36. doi: 10.1186/s13073-016-0292-1
Dinan, T. G., Stanton, C., Cryan, J. F. (2013). Psychobiotics: a novel class of psychotropic. Biol. Psychiatry 74 (10), 720–726. doi: 10.1016/j.biopsych.2013.05.001
Ding, Y., Bu, F., Chen, T., Shi, G., Yuan, X., Feng, Z., et al. (2021). A next-generation probiotic: Akkermansia muciniphila ameliorates chronic stress-induced depressive-like behavior in mice by regulating gut microbiota and metabolites. Appl. Microbiol. Biotechnol. 105 (21-22), 8411–8426. doi: 10.1007/s00253-021-11622-2
Doney, E., Cadoret, A., Dion-Albert, L., Lebel, M., Menard, C. (2022). Inflammation-driven brain and gut barrier dysfunction in stress and mood disorders. Eur. J. Neurosci. 55 (9-10), 2851–2894. doi: 10.1111/ejn.15239
Ducarmon, Q. R., Zwittink, R. D., Hornung, B. V. H., Van Schaik, W., Young, V. B., Kuijper, E. J. (2019). Gut microbiota and colonization resistance against bacterial enteric infection. Microbiol. Mol. Biol. Rev. 83 (3), e00007–e00019. doi: 10.1128/mmbr.00007-19
Duman, R. S., Sanacora, G., Krystal, J. H. (2019). Altered connectivity in depression: GABA and glutamate neurotransmitter deficits and reversal by novel treatments. Neuron 102 (1), 75–90. doi: 10.1016/j.neuron.2019.03.013
Duranti, S., Ruiz, L., Lugli, G. A., Tames, H., Milani, C., Mancabelli, L., et al. (2020). Bifidobacterium adolescentis as a key member of the human gut microbiota in the production of GABA. Sci. Rep. 10 (1), 14112. doi: 10.1038/s41598-020-70986-z
Dyakov, I. N., Mavletova, D. A., Chernyshova, I. N., Snegireva, N. A., Gavrilova, M. V., Bushkova, K. K., et al. (2020). FN3 protein fragment containing two type III fibronectin domains from b. longum GT15 binds to human tumor necrosis factor alpha in vitro. Anaerobe 65, 102247. doi: 10.1016/j.anaerobe.2020.102247
Edwards, P. T., Kashyap, P. C., Preidis, G. A. (2020). Microbiota on biotics: probiotics, prebiotics, and synbiotics to optimize growth and metabolism. Am. J. Physiol. Gastrointest Liver Physiol. 319 (3), G382–G390. doi: 10.1152/ajpgi.00028.2020
Emge, J. R., Huynh, K., Miller, E. N., Kaur, M., Reardon, C., Barrett, K. E., et al. (2016). Modulation of the microbiota-gut-brain axis by probiotics in a murine model of inflammatory bowel disease. Am. J. Physiol. Gastrointest Liver Physiol. 310 (11), G989–G998. doi: 10.1152/ajpgi.00086.2016
Engevik, M. A., Luck, B., Visuthranukul, C., Ihekweazu, F. D., Engevik, A. C., Shi, Z., et al. (2021). Human-derived bifidobacterium dentium modulates the mammalian serotonergic system and gut-brain axis. Cell Mol. Gastroenterol. Hepatol. 11 (1), 221–248. doi: 10.1016/j.jcmgh.2020.08.002
Engevik, M. A., Luk, B., Chang-Graham, A. L., Hall, A., Herrmann, B., Ruan, W., et al. (2019). Bifidobacterium dentium fortifies the intestinal mucus layer via autophagy and calcium signaling pathways. mBio 10 (3), e01087–e01019. doi: 10.1128/mBio.01087-19
Erickson, K. I., Miller, D. L., Roecklein, K. A. (2012). The aging hippocampus: interactions between exercise, depression, and BDNF. Neuroscientist 18 (1), 82–97. doi: 10.1177/1073858410397054
Farooq, R. K., Alamoudi, W., Alhibshi, A., Rehman, S., Sharma, A. R., Abdulla, F. A. (2022). Varied composition and underlying mechanisms of gut microbiome in neuroinflammation. Microorganisms 10 (4), 705. doi: 10.3390/microorganisms10040705
Forsythe, P., Sudo, N., Dinan, T., Taylor, V. H., Bienenstock, J. (2010). Mood and gut feelings. Brain Behav. Immun. 24 (1), 9–16. doi: 10.1016/j.bbi.2009.05.058
Frankiensztajn, L. M., Elliott, E., Koren, O. (2020). The microbiota and the hypothalamus-pituitary-adrenocortical (HPA) axis, implications for anxiety and stress disorders. Curr. Opin. Neurobiol. 62, 76–82. doi: 10.1016/j.conb.2019.12.003
Friedrich, M. J. (2017). Depression is the leading cause of disability around the world. Jama 317 (15), 1517. doi: 10.1001/jama.2017.3826
García Bueno, B., Caso, J. R., Madrigal, J. L., Leza, J. C. (2016). Innate immune receptor toll-like receptor 4 signalling in neuropsychiatric diseases. Neurosci. Biobehav. Rev. 64, 134–147. doi: 10.1016/j.neubiorev.2016.02.013
García-Solache, M., Rice, L. B. (2019). The enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 32 (2), e00058–e00018. doi: 10.1128/cmr.00058-18
Gawlik-Kotelnicka, O., Skowrońska, A., Margulska, A., Czarnecka-Chrebelska, K. H., Łoniewski, I., Skonieczna-Żydecka, K., et al. (2021). The influence of probiotic supplementation on depressive symptoms, inflammation, and oxidative stress parameters and fecal microbiota in patients with depression depending on metabolic syndrome comorbidity-PRO-DEMET randomized study protocol. J. Clin. Med. 10 (7), 1342. doi: 10.3390/jcm10071342
Gerhard, D. M., Wohleb, E. S., Duman, R. S. (2016). Emerging treatment mechanisms for depression: focus on glutamate and synaptic plasticity. Drug Discov. Today 21 (3), 454–464. doi: 10.1016/j.drudis.2016.01.016
Giordano, R., Pellegrino, M., Picu, A., Bonelli, L., Balbo, M., Berardelli, R., et al. (2006). Neuroregulation of the hypothalamus-pituitary-adrenal (HPA) axis in humans: Effects of GABA-, mineralocorticoid-, and GH-secretagogue-receptor modulation. Sci. World J. 6, 1–11. doi: 10.1100/tsw.2006.09
Glassner, K. L., Abraham, B. P., Quigley, E. M. M. (2020). The microbiome and inflammatory bowel disease. J. Allergy Clin. Immunol. 145 (1), 16–27. doi: 10.1016/j.jaci.2019.11.003
Goh, K. K., Liu, Y. W., Kuo, P. H., Chung, Y. E., Lu, M. L., Chen, C. H. (2019). Effect of probiotics on depressive symptoms: A meta-analysis of human studies. Psychiatry Res. 282, 112568. doi: 10.1016/j.psychres.2019.112568
Gold, S. M., Köhler-Forsberg, O., Moss-Morris, R., Mehnert, A., Miranda, J. J., Bullinger, M., et al. (2020). Comorbid depression in medical diseases. Nat. Rev. Dis. Primers 6 (1), 69. doi: 10.1038/s41572-020-0200-2
Goldstein, E. J., Tyrrell, K. L., Citron, D. M. (2015). Lactobacillus species: taxonomic complexity and controversial susceptibilities. Clin. Infect. Dis. 60 Suppl 2, S98–107. doi: 10.1093/cid/civ072
Gu, Y. H., Choi, H., Yamashita, T., Kang, K. M., Iwasa, M., Lee, M. J., et al. (2017). Pharmaceutical production of anti-tumor and immune-potentiating enterococcus faecalis-2001 β-glucans: Enhanced activity of macrophage and lymphocytes in tumor-implanted mice. Curr. Pharm. Biotechnol. 18 (8), 653–661. doi: 10.2174/1389201018666171002130428
Gu, F., Wu, Y., Liu, Y., Dou, M., Jiang, Y., Liang, H. (2020). Lactobacillus casei improves depression-like behavior in chronic unpredictable mild stress-induced rats by the BDNF-TrkB signal pathway and the intestinal microbiota. Food Funct. 11 (7), 6148–6157. doi: 10.1039/d0fo00373e
Guo, Y., Xie, J. P., Deng, K., Li, X., Yuan, Y., Xuan, Q., et al. (2019). Prophylactic effects of bifidobacterium adolescentis on anxiety and depression-like phenotypes after chronic stress: A role of the gut microbiota-inflammation axis. Front. Behav. Neurosci. 13. doi: 10.3389/fnbeh.2019.00126
Haas-Neill, S., Iwashita, E., Dvorkin-Gheva, A., Forsythe, P. (2022). Effects of two distinct psychoactive microbes, lacticaseibacillus rhamnosus JB-1 and limosilactobacillus reuteri 6475, on circulating and hippocampal mRNA in Male mice. Int. J. Mol. Sci. 23 (17), 9653. doi: 10.3390/ijms23179653
Hagihara, M., Yamashita, R., Matsumoto, A., Mori, T., Inagaki, T., Nonogaki, T., et al. (2019). The impact of probiotic clostridium butyricum MIYAIRI 588 on murine gut metabolic alterations. J. Infect. Chemother. 25 (8), 571–577. doi: 10.1016/j.jiac.2019.02.008
Hagihara, M., Yamashita, R., Matsumoto, A., Mori, T., Kuroki, Y., Kudo, H., et al. (2018). The impact of clostridium butyricum MIYAIRI 588 on the murine gut microbiome and colonic tissue. Anaerobe 54, 8–18. doi: 10.1016/j.anaerobe.2018.07.012
Han, L., Zhao, L., Zhou, Y., Yang, C., Xiong, T., Lu, L., et al. (2022). Altered metabolome and microbiome features provide clues in understanding irritable bowel syndrome and depression comorbidity. ISME J. 16 (4), 983–996. doi: 10.1038/s41396-021-01123-5
Hao, W. Z., Li, X. J., Zhang, P. W., Chen, J. X. (2020). A review of antibiotics, depression, and the gut microbiome. Psychiatry Res. 284, 112691. doi: 10.1016/j.psychres.2019.112691
Herman, J. P., Mueller, N. K., Figueiredo, H. (2004). Role of GABA and glutamate circuitry in hypothalamo-pituitary-adrenocortical stress integration. Ann. N. Y. Acad. Sci. 1018, 35–45. doi: 10.1196/annals.1296.004
Hill, C., Guarner, F., Reid, G., Gibson, G. R., Merenstein, D. J., Pot, B., et al. (2014). Expert consensus document. the international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11 (8), 506–514. doi: 10.1038/nrgastro.2014.66
Ho, Y. T., Tsai, Y. C., Kuo, T. B. J., Yang, C. C. H. (2021). Effects of lactobacillus plantarum PS128 on depressive symptoms and sleep quality in self-reported insomniacs: A randomized, double-blind, placebo-controlled pilot trial. Nutrients 13 (8), 2820. doi: 10.3390/nu13082820
Huang, Y. Y., Wu, Y. P., Jia, X. Z., Lin, J., Xiao, L. F., Liu, D. M., et al. (2022). Lactiplantibacillus plantarum DMDL 9010 alleviates dextran sodium sulfate (DSS)-induced colitis and behavioral disorders by facilitating microbiota-gut-brain axis balance. Food Funct. 13 (1), 411–424. doi: 10.1039/d1fo02938j
Jakubczyk, D., Leszczyńska, K., Górska, S. (2020). The effectiveness of probiotics in the treatment of inflammatory bowel disease (IBD)-a critical review. Nutrients 12 (7), 1973. doi: 10.3390/nu12071973
Jang, H. M., Kim, J. K., Joo, M. K., Shin, Y. J., Lee, C. K., Kim, H. J., et al. (2021). Transplantation of fecal microbiota from patients with inflammatory bowel disease and depression alters immune response and behavior in recipient mice. Sci. Rep. 11 (1), 20406. doi: 10.1038/s41598-021-00088-x
Jang, H. M., Kim, J. K., Joo, M. K., Shin, Y. J., Lee, K. E., Lee, C. K., et al. (2022). Enterococcus faecium and pediococcus acidilactici deteriorate enterobacteriaceae-induced depression and colitis in mice. Sci. Rep. 12 (1), 9389. doi: 10.1038/s41598-022-13629-9
Jang, H. M., Lee, K. E., Kim, D. H. (2019). The preventive and curative effects of lactobacillus reuteri NK33 and bifidobacterium adolescentis NK98 on immobilization stress-induced Anxiety/Depression and colitis in mice. Nutrients 11 (4), 819. doi: 10.3390/nu11040819
Jang, H. M., Lee, K. E., Lee, H. J., Kim, D. H. (2018). Immobilization stress-induced escherichia coli causes anxiety by inducing NF-κB activation through gut microbiota disturbance. Sci. Rep. 8 (1), 13897. doi: 10.1038/s41598-018-31764-0
Janik, R., Thomason, L., Stanisz, A. M., Forsythe, P., Bienenstock, J., Stanisz, G. J. (2016). Magnetic resonance spectroscopy reveals oral lactobacillus promotion of increases in brain GABA, n-acetyl aspartate and glutamate. Neuroimage 125, 988–995. doi: 10.1016/j.neuroimage.2015.11.018
Jiang, H., Ling, Z., Zhang, Y., Mao, H., Ma, Z., Yin, Y., et al. (2015). Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 48, 186–194. doi: 10.1016/j.bbi.2015.03.016
Johnson, K. V., Foster, K. R. (2018). Why does the microbiome affect behaviour? Nat. Rev. Microbiol. 16 (10), 647–655. doi: 10.1038/s41579-018-0014-3
Kambe, J., Watcharin, S., Makioka-Itaya, Y., Inoue, R., Watanabe, G., Yamaguchi, H., et al. (2020). Heat-killed enterococcus fecalis (EC-12) supplement alters the expression of neurotransmitter receptor genes in the prefrontal cortex and alleviates anxiety-like behavior in mice. Neurosci. Lett. 720, 134753. doi: 10.1016/j.neulet.2020.134753
Karen, C., Shyu, D. J. H., Rajan, K. E. (2021). Lactobacillus paracasei supplementation prevents early life stress-induced anxiety and depressive-like behavior in maternal separation model-possible involvement of microbiota-Gut-Brain axis in differential regulation of MicroRNA124a/132 and glutamate receptors. Front. Neurosci. 15. doi: 10.3389/fnins.2021.719933
Kazem, Y. I., Mahmoud, M. H., Essa, H. A., Azmy, O., Kandeel, W. A., Al-Moghazy, M., et al. (2021). Role of bifidobacterium spp. intake in improving depressive mood and well-being and its link to kynurenine blood level: an interventional study. J. Complement Integr. Med. 20 (1), 223–232. doi: 10.1515/jcim-2021-0351
Kazemi, A., Noorbala, A. A., Azam, K., Eskandari, M. H., Djafarian, K. (2019). Effect of probiotic and prebiotic vs placebo on psychological outcomes in patients with major depressive disorder: A randomized clinical trial. Clin. Nutr. 38 (2), 522–528. doi: 10.1016/j.clnu.2018.04.010
Keller, M. B., Hirschfeld, R. M., Demyttenaere, K., Baldwin, D. S. (2002). Optimizing outcomes in depression: focus on antidepressant compliance. Int. Clin. Psychopharmacol. 17 (6), 265–271. doi: 10.1097/00004850-200211000-00001
Kelsey, C. M., Prescott, S., Mcculloch, J. A., Trinchieri, G., Valladares, T. L., Dreisbach, C., et al. (2021). Gut microbiota composition is associated with newborn functional brain connectivity and behavioral temperament. Brain Behav. Immun. 91, 472–486. doi: 10.1016/j.bbi.2020.11.003
Kessler, R. C., Bromet, E. J. (2013). The epidemiology of depression across cultures. Annu. Rev. Public Health 34, 119–138. doi: 10.1146/annurev-publhealth-031912-114409
Khan, I., Bai, Y. R., Ullah, N., Liu, G. L., Rajoka, M. S. R., Zhang, C. J. (2022). Differential susceptibility of the gut microbiota to DSS treatment interferes in the conserved microbiome association in mouse models of colitis and is related to the initial gut microbiota difference. Adv. Gut Microbiome Res. 2022, 7813278. doi: 10.1155/2022/7813278
Kim, C. S., Cha, L., Sim, M., Jung, S., Chun, W. Y., Baik, H. W., et al. (2021). Probiotic supplementation improves cognitive function and mood with changes in gut microbiota in community-dwelling older adults: A randomized, double-blind, placebo-controlled, multicenter trial. J. Gerontol A Biol. Sci. Med. Sci. 76 (1), 32–40. doi: 10.1093/gerona/glaa090
Kim, J. S., Kang, S. W., Lee, J. H., Park, S. H., Lee, J. S. (2022). The evolution and competitive strategies of akkermansia muciniphila in gut. Gut Microbes 14 (1), 2025017. doi: 10.1080/19490976.2021.2025017
Kim, S., Shin, Y. C., Kim, T. Y., Kim, Y., Lee, Y. S., Lee, S. H., et al. (2021). Mucin degrader akkermansia muciniphila accelerates intestinal stem cell-mediated epithelial development. Gut Microbes 13 (1), 1–20. doi: 10.1080/19490976.2021.1892441
Kochalska, K., Oakden, W., Słowik, T., Chudzik, A., Pankowska, A., Łazorczyk, A., et al. (2020). Dietary supplementation with lactobacillus rhamnosus JB-1 restores brain neurochemical balance and mitigates the progression of mood disorder in a rat model of chronic unpredictable mild stress. Nutr. Res. 82, 44–57. doi: 10.1016/j.nutres.2020.06.019
Kosuge, A., Kunisawa, K., Arai, S., Sugawara, Y., Shinohara, K., Iida, T., et al. (2021). Heat-sterilized bifidobacterium breve prevents depression-like behavior and interleukin-1β expression in mice exposed to chronic social defeat stress. Brain Behav. Immun. 96, 200–211. doi: 10.1016/j.bbi.2021.05.028
Kuhlmann, S. L., Tschorn, M., Arolt, V., Beer, K., Brandt, J., Grosse, L., et al. (2017). Serum brain-derived neurotrophic factor and stability of depressive symptoms in coronary heart disease patients: A prospective study. Psychoneuroendocrinology 77, 196–202. doi: 10.1016/j.psyneuen.2016.12.015
Lacy, B. E., Weiser, K., Noddin, L., Robertson, D. J., Crowell, M. D., Parratt-Engstrom, C., et al. (2007). Irritable bowel syndrome: patients' attitudes, concerns and level of knowledge. Aliment Pharmacol. Ther. 25 (11), 1329–1341. doi: 10.1111/j.1365-2036.2007.03328.x
Łaniewski, P., Herbst-Kralovetz, M. M. (2022). Connecting microbiome and menopause for healthy ageing. Nat. Microbiol. 7 (3), 354–358. doi: 10.1038/s41564-022-01071-6
Ledford, H. (2014). Medical research: if depression were cancer. Nature 515 (7526), 182–184. doi: 10.1038/515182a
Lee, C., Doo, E., Choi, J. M., Jang, S. H., Ryu, H. S., Lee, J. Y., et al. (2017). The increased level of depression and anxiety in irritable bowel syndrome patients compared with healthy controls: Systematic review and meta-analysis. J. Neurogastroenterol Motil. 23 (3), 349–362. doi: 10.5056/jnm16220
Lee, H. J., Hong, J. K., Kim, J. K., Kim, D. H., Jang, S. W., Han, S. W., et al. (2021). Effects of probiotic NVP-1704 on mental health and sleep in healthy adults: An 8-week randomized, double-blind, placebo-controlled trial. Nutrients 13 (8), 2660. doi: 10.3390/nu13082660
Le Morvan De Sequeira, C., Kaeber, M., Cekin, S. E., Enck, P., Mack, I. (2021). The effect of probiotics on quality of life, depression and anxiety in patients with irritable bowel syndrome: A systematic review and meta-analysis. J. Clin. Med. 10 (16), 3497. doi: 10.3390/jcm10163497
Li, W., Chen, T. T. (2022). An insight into the clinical application of gut microbiota during anticancer therapy. Adv. Gut Microbiome Res. 2022, 8183993. doi: 10.1155/2022/8183993
Li, B., He, Y., Ma, J., Huang, P., Du, J., Cao, L., et al. (2019). Mild cognitive impairment has similar alterations as alzheimer's disease in gut microbiota. Alzheimers Dement 15 (10), 1357–1366. doi: 10.1016/j.jalz.2019.07.002
Li, H., Wang, P., Huang, L., Li, P., Zhang, D. (2019). Effects of regulating gut microbiota on the serotonin metabolism in the chronic unpredictable mild stress rat model. Neurogastroenterol Motil. 31 (10), e13677. doi: 10.1111/nmo.13677
Li, N., Wang, Q., Wang, Y., Sun, A., Lin, Y., Jin, Y., et al. (2019). Fecal microbiota transplantation from chronic unpredictable mild stress mice donors affects anxiety-like and depression-like behavior in recipient mice via the gut microbiota-inflammation-brain axis. Stress 22 (5), 592–602. doi: 10.1080/10253890.2019.1617267
Li, P., Zhang, Y. X., Xu, Y. P., Cao, H. C., Li, L. J. (2022). Characteristics of CD8+ and CD4+ tissue-resident memory lymphocytes in the gastrointestinal tract. Adv. Gut Microbiome Res. 2022, 9157455. doi: 10.1155/2022/9157455
Li, Y., Zhao, L., Yu, D., Wang, Z., Ding, G. (2018). Metabolic syndrome prevalence and its risk factors among adults in China: A nationally representative cross-sectional study. PloS One 13 (6), e0199293. doi: 10.1371/journal.pone.0199293
Ling, Z., Cheng, Y., Chen, F., Yan, X., Liu, X., Shao, L., et al. (2022a). Changes in fecal microbiota composition and the cytokine expression profile in school-aged children with depression: A case-control study. Front. Immunol. 13. doi: 10.3389/fimmu.2022.964910
Ling, Z., Cheng, Y., Yan, X., Shao, L., Liu, X., Zhou, D., et al. (2020). Alterations of the fecal microbiota in Chinese patients with multiple sclerosis. Front. Immunol. 11. doi: 10.3389/fimmu.2020.590783
Ling, Z., Jin, G., Yan, X., Cheng, Y., Shao, L., Song, Q., et al. (2022b). Fecal dysbiosis and immune dysfunction in Chinese elderly patients with schizophrenia: An observational study. Front. Cell Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.886872
Ling, Z., Liu, X., Cheng, Y., Yan, X., Wu, S. (2022c). Gut microbiota and aging. Crit. Rev. Food Sci. Nutr. 62 (13), 3509–3534. doi: 10.1080/10408398.2020.1867054
Ling, Z. X., Xiao, H., Chen, W. (2022). Gut microbiome: The cornerstone of life and health. Adv. Gut Microbiome Res. 2022, 9894812. doi: 10.1155/2022/9894812
Liu, Y., Mian, M. F., Mcvey Neufeld, K. A., Forsythe, P. (2020). CD4(+)CD25(+) T cells are essential for behavioral effects of lactobacillus rhamnosus JB-1 in Male BALB/c mice. Brain Behav. Immun. 88, 451–460. doi: 10.1016/j.bbi.2020.04.014
Liu, Y., Sanderson, D., Mian, M. F., Mcvey Neufeld, K. A., Forsythe, P. (2021a). Loss of vagal integrity disrupts immune components of the microbiota-gut-brain axis and inhibits the effect of lactobacillus rhamnosus on behavior and the corticosterone stress response. Neuropharmacology 195, 108682. doi: 10.1016/j.neuropharm.2021.108682
Liu, J., Sun, J., Wang, F., Yu, X., Ling, Z., Li, H., et al. (2015). Neuroprotective effects of clostridium butyricum against vascular dementia in mice via metabolic butyrate. BioMed. Res. Int. 2015, 412946. doi: 10.1155/2015/412946
Liu, Y., Wang, H., Gui, S., Zeng, B., Pu, J., Zheng, P., et al. (2021b). Proteomics analysis of the gut-brain axis in a gut microbiota-dysbiosis model of depression. Transl. Psychiatry 11 (1), 568. doi: 10.1038/s41398-021-01689-w
Loughman, A., Ponsonby, A. L., O'hely, M., Symeonides, C., Collier, F., Tang, M. L. K., et al. (2020). Gut microbiota composition during infancy and subsequent behavioural outcomes. EBioMedicine 52, 102640. doi: 10.1016/j.ebiom.2020.102640
Lu, J., Xu, X., Huang, Y., Li, T., Ma, C., Xu, G., et al. (2021). Prevalence of depressive disorders and treatment in China: A cross-sectional epidemiological study. Lancet Psychiatry 8 (11), 981–990. doi: 10.1016/s2215-0366(21)00251-0
Lührs, H., Gerke, T., Müller, J. G., Melcher, R., Schauber, J., Boxberge, F., et al. (2002). Butyrate inhibits NF-kappaB activation in lamina propria macrophages of patients with ulcerative colitis. Scand. J. Gastroenterol. 37 (4), 458–466. doi: 10.1080/003655202317316105
Lynch, S. V., Pedersen, O. (2016). The human intestinal microbiome in health and disease. N Engl. J. Med. 375 (24), 2369–2379. doi: 10.1056/NEJMra1600266
Majeed, M., Nagabhushanam, K., Arumugam, S., Majeed, S., Ali, F. (2018). Bacillus coagulans MTCC 5856 for the management of major depression with irritable bowel syndrome: A randomised, double-blind, placebo controlled, multi-centre, pilot clinical study. Food Nutr. Res. 62, 1218. doi: 10.29219/fnr.v62.1218
Malhi, G. S., Mann, J. J. (2018). Depression. Lancet 392 (10161), 2299–2312. doi: 10.1016/s0140-6736(18)31948-2
Maqsood, R., Stone, T. W. (2016). The gut-brain axis, BDNF, NMDA and CNS disorders. Neurochem. Res. 41 (11), 2819–2835. doi: 10.1007/s11064-016-2039-1
Marotta, A., Sarno, E., Del Casale, A., Pane, M., Mogna, L., Amoruso, A., et al. (2019). Effects of probiotics on cognitive reactivity, mood, and sleep quality. Front. Psychiatry 10. doi: 10.3389/fpsyt.2019.00164
Martinotti, G., Pettorruso, M., De Berardis, D., Varasano, P. A., Lucidi Pressanti, G., De Remigis, V., et al. (2016). Agomelatine increases BDNF serum levels in depressed patients in correlation with the improvement of depressive symptoms. Int. J. Neuropsychopharmacol. 19 (5), pyw003. doi: 10.1093/ijnp/pyw003
Mason, B. L., Li, Q., Minhajuddin, A., Czysz, A. H., Coughlin, L. A., Hussain, S. K., et al. (2020). Reduced anti-inflammatory gut microbiota are associated with depression and anhedonia. J. Affect. Disord. 266, 394–401. doi: 10.1016/j.jad.2020.01.137
Mazloom, K., Siddiqi, I., Covasa, M. (2019). Probiotics: How effective are they in the fight against obesity? Nutrients 11 (2), 258. doi: 10.3390/nu11020258
Mcgaughey, K. D., Yilmaz-Swenson, T., Elsayed, N. M., Cruz, D. A., Rodriguiz, R. M., Kritzer, M. D., et al. (2019). Relative abundance of akkermansia spp. and other bacterial phylotypes correlates with anxiety- and depressive-like behavior following social defeat in mice. Sci. Rep. 9 (1), 3281. doi: 10.1038/s41598-019-40140-5
Mcvey Neufeld, K. A., O'mahony, S. M., Hoban, A. E., Waworuntu, R. V., Berg, B. M., Dinan, T. G., et al. (2019). Neurobehavioural effects of lactobacillus rhamnosus GG alone and in combination with prebiotics polydextrose and galactooligosaccharide in male rats exposed to early-life stress. Nutr. Neurosci. 22 (6), 425–434. doi: 10.1080/1028415x.2017.1397875
Mearin, F., Lacy, B. E., Chang, L., Chey, W. D., Lembo, A. J., Simren, M., et al. (2016). Bowel disorders. Gastroenterology. doi: 10.1053/j.gastro.2016.02.031
Messaoudi, M., Violle, N., Bisson, J. F., Desor, D., Javelot, H., Rougeot, C. (2011). Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes 2 (4), 256–261. doi: 10.4161/gmic.2.4.16108
Methiwala, H. N., Vaidya, B., Addanki, V. K., Bishnoi, M., Sharma, S. S., Kondepudi, K. K. (2021). Gut microbiota in mental health and depression: Role of pre/pro/synbiotics in their modulation. Food Funct. 12 (10), 4284–4314. doi: 10.1039/d0fo02855j
Miao, R. X., Zhu, X. X., Wan, C. M., Wang, Z. L., Wen, Y., Li, Y. Y. (2018). Effect of clostridium butyricum supplementation on the development of intestinal flora and the immune system of neonatal mice. Exp. Ther. Med. 15 (1), 1081–1086. doi: 10.3892/etm.2017.5461
Midenfjord, I., Polster, A., Sjövall, H., Törnblom, H., Simrén, M. (2019). Anxiety and depression in irritable bowel syndrome: Exploring the interaction with other symptoms and pathophysiology using multivariate analyses. Neurogastroenterol. Motil. 31 (8), e13619. doi: 10.1111/nmo.13619
Miller, A. H., Raison, C. L. (2016). The role of inflammation in depression: From evolutionary imperative to modern treatment target. Nat. Rev. Immunol. 16 (1), 22–34. doi: 10.1038/nri.2015.5
Miyanishi, H., Nitta, A. (2021). A role of BDNF in the depression pathogenesis and a potential target as antidepressant: The modulator of stress sensitivity "Shati/Nat8l-BDNF system" in the dorsal striatum. Pharm. (Basel) 14 (9), 889. doi: 10.3390/ph14090889
Miyaoka, T., Kanayama, M., Wake, R., Hashioka, S., Hayashida, M., Nagahama, M., et al. (2018). Clostridium butyricum MIYAIRI 588 as adjunctive therapy for treatment-resistant major depressive disorder: A prospective open-label trial. Clin. Neuropharmacol 41 (5), 151–155. doi: 10.1097/wnf.0000000000000299
Mohammed, S. K., Magdy, Y. M., El-Waseef, D. A., Nabih, E. S., Hamouda, M. A., El-Kharashi, O. A. (2020). Modulation of hippocampal TLR4/BDNF signal pathway using probiotics is a step closer towards treating cognitive impairment in NASH model. Physiol. Behav. 214, 112762. doi: 10.1016/j.physbeh.2019.112762
Mudyanadzo, T. A., Hauzaree, C., Yerokhina, O., Architha, N. N., Ashqar, H. M. (2018). Irritable bowel syndrome and depression: A shared pathogenesis. Cureus 10 (8), e3178. doi: 10.7759/cureus.3178
Mulder, D. J., Noble, A. J., Justinich, C. J., Duffin, J. M. (2014). A tale of two diseases: The history of inflammatory bowel disease. J. Crohns Colitis 8 (5), 341–348. doi: 10.1016/j.crohns.2013.09.009
Nikolova, V. L., Cleare, A. J., Young, A. H., Stone, J. M. (2021). Updated review and meta-analysis of probiotics for the treatment of clinical depression: Adjunctive vs. stand-alone treatment. J. Clin. Med. 10 (4), 647. doi: 10.3390/jcm10040647
Onyike, C. U., Crum, R. M., Lee, H. B., Lyketsos, C. G., Eaton, W. W. (2003). Is obesity associated with major depression? results from the third national health and nutrition examination survey. Am. J. Epidemiol. 158 (12), 1139–1147. doi: 10.1093/aje/kwg275
Opel, N., Redlich, R., Grotegerd, D., Dohm, K., Heindel, W., Kugel, H., et al. (2015). Obesity and major depression: Body-mass index (BMI) is associated with a severe course of disease and specific neurostructural alterations. Psychoneuroendocrinology 51, 219–226. doi: 10.1016/j.psyneuen.2014.10.001
Otaka, M., Kikuchi-Hayakawa, H., Ogura, J., Ishikawa, H., Yomogida, Y., Ota, M., et al. (2021). Effect of lacticaseibacillus paracasei strain shirota on improvement in depressive symptoms, and its association with abundance of actinobacteria in gut microbiota. Microorganisms 9 (5), 1026. doi: 10.3390/microorganisms9051026
Ouyang, J., Lin, J., Isnard, S., Fombuena, B., Peng, X., Marette, A., et al. (2020). The bacterium akkermansia muciniphila: A sentinel for gut permeability and its relevance to HIV-related inflammation. Front. Immunol. 11. doi: 10.3389/fimmu.2020.00645
Paone, P., Cani, P. D. (2020). Mucus barrier, mucins and gut microbiota: the expected slimy partners? Gut 69 (12), 2232–2243. doi: 10.1136/gutjnl-2020-322260
Park, C., Brietzke, E., Rosenblat, J. D., Musial, N., Zuckerman, H., Ragguett, R. M., et al. (2018). Probiotics for the treatment of depressive symptoms: An anti-inflammatory mechanism? Brain Behav. Immun. 73, 115–124. doi: 10.1016/j.bbi.2018.07.006
Partrick, K. A., Rosenhauer, A. M., Auger, J., Arnold, A. R., Ronczkowski, N. M., Jackson, L. M., et al. (2021). Ingestion of probiotic (Lactobacillus helveticus and bifidobacterium longum) alters intestinal microbial structure and behavioral expression following social defeat stress. Sci. Rep. 11 (1), 3763. doi: 10.1038/s41598-021-83284-z
Party, H., Dujarrier, C., Hébert, M., Lenoir, S., Martinez De Lizarrondo, S., Delépée, R., et al. (2019). Plasminogen activator inhibitor-1 (PAI-1) deficiency predisposes to depression and resistance to treatments. Acta Neuropathol. Commun. 7 (1), 153. doi: 10.1186/s40478-019-0807-2
Patterson, E., Ryan, P. M., Wiley, N., Carafa, I., Sherwin, E., Moloney, G., et al. (2019). Gamma-aminobutyric acid-producing lactobacilli positively affect metabolism and depressive-like behaviour in a mouse model of metabolic syndrome. Sci. Rep. 9 (1), 16323. doi: 10.1038/s41598-019-51781-x
Peirce, J. M., Alviña, K. (2019). The role of inflammation and the gut microbiome in depression and anxiety. J. Neurosci. Res. 97 (10), 1223–1241. doi: 10.1002/jnr.24476
Pinto-Sanchez, M. I., Hall, G. B., Ghajar, K., Nardelli, A., Bolino, C., Lau, J. T., et al. (2017). Probiotic bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: A pilot study in patients with irritable bowel syndrome. Gastroenterology 153 (2), 448–459.e448. doi: 10.1053/j.gastro.2017.05.003
Qin, L., Wu, X., Block, M. L., Liu, Y., Breese, G. R., Hong, J. S., et al. (2007). Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 55 (5), 453–462. doi: 10.1002/glia.20467
Qiu, X., Wu, G., Wang, L., Tan, Y., Song, Z. (2021). Lactobacillus delbrueckii alleviates depression-like behavior through inhibiting toll-like receptor 4 (TLR4) signaling in mice. Ann. Transl. Med. 9 (5), 366. doi: 10.21037/atm-20-4411
Ray, K. (2017). IBS: Mindful of probiotics for psychiatric comorbidities in IBS. Nat. Rev. Gastroenterol. Hepatol. 14 (7), 386–387. doi: 10.1038/nrgastro.2017.70
Reimann, F., Habib, A. M., Tolhurst, G., Parker, H. E., Rogers, G. J., Gribble, F. M. (2008). Glucose sensing in l cells: a primary cell study. Cell Metab. 8 (6), 532–539. doi: 10.1016/j.cmet.2008.11.002
Remely, M., Hippe, B., Geretschlaeger, I., Stegmayer, S., Hoefinger, I., Haslberger, A. (2015). Increased gut microbiota diversity and abundance of faecalibacterium prausnitzii and akkermansia after fasting: A pilot study. Wien Klin Wochenschr 127 (9-10), 394–398. doi: 10.1007/s00508-015-0755-1
Restivo, M. R., Mckinnon, M. C., Frey, B. N., Hall, G. B., Syed, W., Taylor, V. H. (2017). The impact of obesity on neuropsychological functioning in adults with and without major depressive disorder. PloS One 12 (5), e0176898. doi: 10.1371/journal.pone.0176898
Ridaura, V., Belkaid, Y. (2015). Gut microbiota: The link to your second brain. Cell 161 (2), 193–194. doi: 10.1016/j.cell.2015.03.033
Rudzki, L., Ostrowska, L., Pawlak, D., Małus, A., Pawlak, K., Waszkiewicz, N., et al. (2019). Probiotic lactobacillus plantarum 299v decreases kynurenine concentration and improves cognitive functions in patients with major depression: A double-blind, randomized, placebo controlled study. Psychoneuroendocrinology 100, 213–222. doi: 10.1016/j.psyneuen.2018.10.010
Saarela, M., Mogensen, G., Fondén, R., Mättö, J., Mattila-Sandholm, T. (2000). Probiotic bacteria: Safety, functional and technological properties. J. Biotechnol. 84 (3), 197–215. doi: 10.1016/s0168-1656(00)00375-8
Salari, N., Hosseinian-Far, A., Jalali, R., Vaisi-Raygani, A., Rasoulpoor, S., Mohammadi, M., et al. (2020). Prevalence of stress, anxiety, depression among the general population during the COVID-19 pandemic: A systematic review and meta-analysis. Global Health 16 (1), 57. doi: 10.1186/s12992-020-00589-w
Sánchez, B., Delgado, S., Blanco-Míguez, A., Lourenço, A., Gueimonde, M., Margolles, A. (2017). Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 61 (1), 1600240. doi: 10.1002/mnfr.201600240
Sanders, M. E., Merenstein, D. J., Reid, G., Gibson, G. R., Rastall, R. A. (2019). Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 16 (10), 605–616. doi: 10.1038/s41575-019-0173-3
Sandes, S., Figueiredo, N., Pedroso, S., Sant'anna, F., Acurcio, L., Abatemarco Junior, M., et al. (2020). Weissella paramesenteroides WpK4 plays an immunobiotic role in gut-brain axis, reducing gut permeability, anxiety-like and depressive-like behaviors in murine models of colitis and chronic stress. Food Res. Int. 137, 109741. doi: 10.1016/j.foodres.2020.109741
Sarkar, J., Wakefield, S., Mackenzie, G., Moss, S. J., Maguire, J. (2011). Neurosteroidogenesis is required for the physiological response to stress: Role of neurosteroid-sensitive GABAA receptors. J. Neurosci. 31 (50), 18198–18210. doi: 10.1523/jneurosci.2560-11.2011
Schaub, A. C., Schneider, E., Vazquez-Castellanos, J. F., Schweinfurth, N., Kettelhack, C., Doll, J. P. K., et al. (2022). Clinical, gut microbial and neural effects of a probiotic add-on therapy in depressed patients: A randomized controlled trial. Transl. Psychiatry 12 (1), 227. doi: 10.1038/s41398-022-01977-z
Selle, K., Klaenhammer, T. R. (2013). Genomic and phenotypic evidence for probiotic influences of lactobacillus gasseri on human health. FEMS Microbiol. Rev. 37 (6), 915–935. doi: 10.1111/1574-6976.12021
Semenkovich, K., Brown, M. E., Svrakic, D. M., Lustman, P. J. (2015). Depression in type 2 diabetes mellitus: prevalence, impact, and treatment. Drugs 75 (6), 577–587. doi: 10.1007/s40265-015-0347-4
Simpson, C. A., Diaz-Arteche, C., Eliby, D., Schwartz, O. S., Simmons, J. G., Cowan, C. S. M. (2021). The gut microbiota in anxiety and depression - a systematic review. Clin. Psychol. Rev. 83, 101943. doi: 10.1016/j.cpr.2020.101943
Simpson, C. A., Mu, A., Haslam, N., Schwartz, O. S., Simmons, J. G. (2020). Feeling down? a systematic review of the gut microbiota in anxiety/depression and irritable bowel syndrome. J. Affect. Disord. 266, 429–446. doi: 10.1016/j.jad.2020.01.124
Slykerman, R. F., Hood, F., Wickens, K., Thompson, J. M. D., Barthow, C., Murphy, R., et al. (2017). Effect of lactobacillus rhamnosus HN001 in pregnancy on postpartum symptoms of depression and anxiety: A randomised double-blind placebo-controlled trial. EBioMedicine 24, 159–165. doi: 10.1016/j.ebiom.2017.09.013
Smith, P. M., Howitt, M. R., Panikov, N., Michaud, M., Gallini, C. A., Bohlooly, Y. M., et al. (2013). The microbial metabolites, short-chain fatty acids, regulate colonic treg cell homeostasis. Science 341 (6145), 569–573. doi: 10.1126/science.1241165
Snigdha, S., Ha, K., Tsai, P., Dinan, T. G., Bartos, J. D., Shahid, M. (2022). Probiotics: Potential novel therapeutics for microbiota-gut-brain axis dysfunction across gender and lifespan. Pharmacol. Ther. 231, 107978. doi: 10.1016/j.pharmthera.2021.107978
Sonali, S., Ray, B., Ahmed Tousif, H., Rathipriya, A. G., Sunanda, T., Mahalakshmi, A. M., et al. (2022). Mechanistic insights into the link between gut dysbiosis and major depression: An extensive review. Cells 11 (8), 1362. doi: 10.3390/cells11081362
Song, H. Y., Yu, R. C. (2018). Optimization of culture conditions for gamma-aminobutyric acid production in fermented adzuki bean milk. J. Food Drug Anal. 26 (1), 74–81. doi: 10.1016/j.jfda.2016.11.024
Stasi, C., Orlandelli, E. (2008). Role of the brain-gut axis in the pathophysiology of crohn's disease. Dig Dis. 26 (2), 156–166. doi: 10.1159/000116774
Strati, F., Cavalieri, D., Albanese, D., De Felice, C., Donati, C., Hayek, J., et al. (2016). Altered gut microbiota in rett syndrome. Microbiome 4 (1), 41. doi: 10.1186/s40168-016-0185-y
Suda, K., Matsuda, K. (2022). How microbes affect depression: Underlying mechanisms via the gut-brain axis and the modulating role of probiotics. Int. J. Mol. Sci. 23 (3), 1172. doi: 10.3390/ijms23031172
Sun, J., Li, H., Jin, Y., Yu, J., Mao, S., Su, K. P., et al. (2021). Probiotic clostridium butyricum ameliorated motor deficits in a mouse model of parkinson's disease via gut microbiota-GLP-1 pathway. Brain Behav. Immun. 91, 703–715. doi: 10.1016/j.bbi.2020.10.014
Sun, J., Wang, F., Hu, X., Yang, C., Xu, H., Yao, Y., et al. (2018). Clostridium butyricum attenuates chronic unpredictable mild stress-induced depressive-like behavior in mice via the gut-brain axis. J. Agric. Food Chem. 66 (31), 8415–8421. doi: 10.1021/acs.jafc.8b02462
Sun, J., Xu, J., Ling, Y., Wang, F., Gong, T., Yang, C., et al. (2019). Fecal microbiota transplantation alleviated alzheimer's disease-like pathogenesis in APP/PS1 transgenic mice. Transl. Psychiatry 9 (1), 189. doi: 10.1038/s41398-019-0525-3
Takahashi, K., Kurokawa, K., Hong, L., Miyagawa, K., Mochida-Saito, A., Iwasa, M., et al. (2022). Antidepressant effects of enterococcus faecalis 2001 through the regulation of prefrontal cortical myelination via the enhancement of CREB/BDNF and NF-κB p65/LIF/STAT3 pathways in olfactory bulbectomized mice. J. Psychiatr. Res. 148, 137–148. doi: 10.1016/j.jpsychires.2022.01.047
Takahashi, K., Kurokawa, K., Miyagawa, K., Mochida-Saito, A., Nemoto, Y., Iwasa, H., et al. (2020). Antidementia effects of enterococcus faecalis 2001 are associated with enhancement of hippocampal neurogenesis via the ERK-CREB-BDNF pathway in olfactory bulbectomized mice. Physiol. Behav. 223, 112997. doi: 10.1016/j.physbeh.2020.112997
Takahashi, K., Nakagawasai, O., Nemoto, W., Kadota, S., Isono, J., Odaira, T., et al. (2018). Memantine ameliorates depressive-like behaviors by regulating hippocampal cell proliferation and neuroprotection in olfactory bulbectomized mice. Neuropharmacology 137, 141–155. doi: 10.1016/j.neuropharm.2018.04.013
Takahashi, K., Nakagawasai, O., Nemoto, W., Odaira, T., Sakuma, W., Onogi, H., et al. (2019). Effect of enterococcus faecalis 2001 on colitis and depressive-like behavior in dextran sulfate sodium-treated mice: involvement of the brain-gut axis. J. Neuroinflamm. 16 (1), 201. doi: 10.1186/s12974-019-1580-7
Tette, F. M., Kwofie, S. K., Wilson, M. D. (2022). Therapeutic anti-depressant potential of microbial GABA produced by lactobacillus rhamnosus strains for GABAergic signaling restoration and inhibition of addiction-induced HPA axis hyperactivity. Curr. Issues Mol. Biol. 44 (4), 1434–1451. doi: 10.3390/cimb44040096
Tian, P., Chen, Y., Zhu, H., Wang, L., Qian, X., Zou, R., et al. (2022). Bifidobacterium breve CCFM1025 attenuates major depression disorder via regulating gut microbiome and tryptophan metabolism: A randomized clinical trial. Brain Behav. Immun. 100, 233–241. doi: 10.1016/j.bbi.2021.11.023
Tian, P., O'riordan, K. J., Lee, Y. K., Wang, G., Zhao, J., Zhang, H., et al. (2020). Towards a psychobiotic therapy for depression: Bifidobacterium breve CCFM1025 reverses chronic stress-induced depressive symptoms and gut microbial abnormalities in mice. Neurobiol. Stress 12, 100216. doi: 10.1016/j.ynstr.2020.100216
Tian, P., Wang, G., Zhao, J., Zhang, H., Chen, W. (2019). Bifidobacterium with the role of 5-hydroxytryptophan synthesis regulation alleviates the symptom of depression and related microbiota dysbiosis. J. Nutr. Biochem. 66, 43–51. doi: 10.1016/j.jnutbio.2019.01.007
Tian, T., Xu, B., Qin, Y., Fan, L., Chen, J., Zheng, P., et al. (2019). Clostridium butyricum miyairi 588 has preventive effects on chronic social defeat stress-induced depressive-like behaviour and modulates microglial activation in mice. Biochem. Biophys. Res. Commun. 516 (2), 430–436. doi: 10.1016/j.bbrc.2019.06.053
Tiller, J. W. (2013). Depression and anxiety. Med. J. Aust. 199 (S6), S28–S31. doi: 10.5694/mja12.10628
Valdes, A. M., Walter, J., Segal, E., Spector, T. D. (2018). Role of the gut microbiota in nutrition and health. Bmj 361, k2179. doi: 10.1136/bmj.k2179
Valles-Colomer, M., Falony, G., Darzi, Y., Tigchelaar, E. F., Wang, J., Tito, R. Y., et al. (2019). The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 4 (4), 623–632. doi: 10.1038/s41564-018-0337-x
Van De Wouw, M., Boehme, M., Lyte, J. M., Wiley, N., Strain, C., O'sullivan, O., et al. (2018). Short-chain fatty acids: microbial metabolites that alleviate stress-induced brain-gut axis alterations. J. Physiol. 596 (20), 4923–4944. doi: 10.1113/jp276431
Von Bohlen Und Halbach, O., Von Bohlen Und Halbach, V. (2018). BDNF effects on dendritic spine morphology and hippocampal function. Cell Tissue Res. 373 (3), 729–741. doi: 10.1007/s00441-017-2782-x
Walker, A. K., Wing, E. E., Banks, W. A., Dantzer, R. (2019). Leucine competes with kynurenine for blood-to-brain transport and prevents lipopolysaccharide-induced depression-like behavior in mice. Mol. Psychiatry 24 (10), 1523–1532. doi: 10.1038/s41380-018-0076-7
Wang, L., Christophersen, C. T., Sorich, M. J., Gerber, J. P., Angley, M. T., Conlon, M. A. (2011). Low relative abundances of the mucolytic bacterium akkermansia muciniphila and bifidobacterium spp. in feces of children with autism. Appl. Environ. Microbiol. 77 (18), 6718–6721. doi: 10.1128/aem.05212-11
Wastyk, H. C., Fragiadakis, G. K., Perelman, D., Dahan, D., Merrill, B. D., Yu, F. B., et al. (2021). Gut-microbiota-targeted diets modulate human immune status. Cell 184 (16), 4137–4153.e4114. doi: 10.1016/j.cell.2021.06.019
Wei, C. L., Wang, S., Yen, J. T., Cheng, Y. F., Liao, C. L., Hsu, C. C., et al. (2019). Antidepressant-like activities of live and heat-killed lactobacillus paracasei PS23 in chronic corticosterone-treated mice and possible mechanisms. Brain Res. 1711, 202–213. doi: 10.1016/j.brainres.2019.01.025
Weinberger, A. H., Gbedemah, M., Martinez, A. M., Nash, D., Galea, S., Goodwin, R. D. (2018). Trends in depression prevalence in the USA from 2005 to 2015: widening disparities in vulnerable groups. Psychol. Med. 48 (8), 1308–1315. doi: 10.1017/s0033291717002781
Weis, S., Schwiertz, A., Unger, M. M., Becker, A., Faßbender, K., Ratering, S., et al. (2019). Effect of parkinson's disease and related medications on the composition of the fecal bacterial microbiota. NPJ Parkinsons Dis. 5, 28. doi: 10.1038/s41531-019-0100-x
Wong, M. L., Inserra, A., Lewis, M. D., Mastronardi, C. A., Leong, L., Choo, J., et al. (2016). Inflammasome signaling affects anxiety- and depressive-like behavior and gut microbiome composition. Mol. Psychiatry 21 (6), 797–805. doi: 10.1038/mp.2016.46
World Health Organization. (2017). Depression and other common mental disorders: Global health estimates. Available at: http://apps.who.int/iris/bitstream/10665/254610/1/WHO-MSD-MER-2017.2-eng.pdf?ua=1.
Xu, J., Tang, M., Wu, X., Kong, X., Liu, Y., Xu, X. (2022). Lactobacillus rhamnosus zz-1 exerts preventive effects on chronic unpredictable mild stress-induced depression in mice via regulating the intestinal microenvironment. Food Funct. 13 (8), 4331–4343. doi: 10.1039/d1fo03804d
Xu, M., Tian, P., Zhu, H., Zou, R., Zhao, J., Zhang, H., et al. (2022). Lactobacillus paracasei CCFM1229 and lactobacillus rhamnosus CCFM1228 alleviated depression- and anxiety-related symptoms of chronic stress-induced depression in mice by regulating xanthine oxidase activity in the brain. Nutrients 14 (6), 1294. doi: 10.3390/nu14061294
Yoda, K., Harata, G., Sato, M., Miyazawa, K., Ohsawa, N., He, F., et al. (2022). Effect of probiotic bifidobacterium bifidum TMC3115 supplementation on psychosocial stress using a Sub-chronic and mild social defeat stress in mice. Nutrients 14 (5), 970. doi: 10.3390/nu14050970
Yoo, J. W., Shin, Y. J., Ma, X., Son, Y. H., Jang, H. M., Lee, C. K., et al. (2022). The alleviation of gut microbiota-induced depression and colitis in mice by anti-inflammatory probiotics NK151, NK173, and NK175. Nutrients 14 (10), 2080. doi: 10.3390/nu14102080
Yoon, H. S., Cho, C. H., Yun, M. S., Jang, S. J., You, H. J., Kim, J. H., et al. (2021). Akkermansia muciniphila secretes a glucagon-like peptide-1-inducing protein that improves glucose homeostasis and ameliorates metabolic disease in mice. Nat. Microbiol. 6 (5), 563–573. doi: 10.1038/s41564-021-00880-5
Youssef, M. M., Underwood, M. D., Huang, Y. Y., Hsiung, S. C., Liu, Y., Simpson, N. R., et al. (2018). Association of BDNF Val66Met polymorphism and brain BDNF levels with major depression and suicide. Int. J. Neuropsychopharmacol. 21 (6), 528–538. doi: 10.1093/ijnp/pyy008
Yuan, X., Chen, B., Duan, Z., Xia, Z., Ding, Y., Chen, T., et al. (2021). Depression and anxiety in patients with active ulcerative colitis: Crosstalk of gut microbiota, metabolomics and proteomics. Gut Microbes 13 (1), 1987779. doi: 10.1080/19490976.2021.1987779
Yun, S. W., Kim, J. K., Lee, K. E., Oh, Y. J., Choi, H. J., Han, M. J., et al. (2020). A probiotic lactobacillus gasseri alleviates escherichia coli-induced cognitive impairment and depression in mice by regulating IL-1β expression and gut microbiota. Nutrients 12 (11), 3441. doi: 10.3390/nu12113441
Yunes, R. A., Poluektova, E. U., Vasileva, E. V., Odorskaya, M. V., Marsova, M. V., Kovalev, G. I., et al. (2020). A multi-strain potential probiotic formulation of GABA-producing lactobacillus plantarum 90sk and bifidobacterium adolescentis 150 with antidepressant effects. Probiotics Antimicrob. Proteins 12 (3), 973–979. doi: 10.1007/s12602-019-09601-1
Zhai, Q., Feng, S., Arjan, N., Chen, W. (2019). A next generation probiotic, akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 59 (19), 3227–3236. doi: 10.1080/10408398.2018.1517725
Zhang, X., Chen, S., Zhang, M., Ren, F., Ren, Y., Li, Y., et al. (2021). Effects of fermented milk containing lacticaseibacillus paracasei strain shirota on constipation in patients with depression: A randomized, double-blind, placebo-controlled trial. Nutrients 13 (7), 2238. doi: 10.3390/nu13072238
Zhang, W., Ding, T., Zhang, H., Chen, Y., Liu, L., Jiang, J., et al. (2022). Clostridium butyricum RH2 alleviates chronic foot shock stress-induced behavioral deficits in rats via PAI-1. Front. Pharmacol. 13. doi: 10.3389/fphar.2022.845221
Zhang, T., Zhang, C., Zhang, J., Sun, F., Duan, L. (2022). Efficacy of probiotics for irritable bowel syndrome: A systematic review and network meta-analysis. Front. Cell Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.859967
Zhao, L., Xie, Q., Etareri Evivie, S., Liu, D., Dong, J., Ping, L., et al. (2021). Bifidobacterium dentium N8 with potential probiotic characteristics prevents LPS-induced intestinal barrier injury by alleviating the inflammatory response and regulating the tight junction in caco-2 cell monolayers. Food Funct. 12 (16), 7171–7184. doi: 10.1039/d1fo01164b
Zhao, L., Zhang, F., Ding, X., Wu, G., Lam, Y. Y., Wang, X., et al. (2018). Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 359 (6380), 1151–1156. doi: 10.1126/science.aao5774
Zheng, P., Zeng, B., Zhou, C., Liu, M., Fang, Z., Xu, X., et al. (2016). Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host's metabolism. Mol. Psychiatry 21 (6), 786–796. doi: 10.1038/mp.2016.44
Zhou, B., Jin, G., Pang, X., Mo, Q., Bao, J., Liu, T., et al. (2022). Lactobacillus rhamnosus GG colonization in early life regulates gut-brain axis and relieves anxiety-like behavior in adulthood. Pharmacol. Res. 177, 106090. doi: 10.1016/j.phrs.2022.106090
Keywords: comorbidity, depression, dysbiosis, microbiota-gut-brain axis, probiotics
Citation: Gao J, Zhao L, Cheng Y, Lei W, Wang Y, Liu X, Zheng N, Shao L, Chen X, Sun Y, Ling Z and Xu W (2023) Probiotics for the treatment of depression and its comorbidities: A systemic review. Front. Cell. Infect. Microbiol. 13:1167116. doi: 10.3389/fcimb.2023.1167116
Received: 16 February 2023; Accepted: 15 March 2023;
Published: 17 April 2023.
Edited by:
Deguang Song, Yale University, United StatesReviewed by:
Xin Chen, Stanford University, United StatesCopyright © 2023 Gao, Zhao, Cheng, Lei, Wang, Liu, Zheng, Shao, Chen, Sun, Ling and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zongxin Ling, bGluZ3pvbmd4aW5Aemp1LmVkdS5jbg==; Weijie Xu, d2Vpal94dUAxMjYuY29t
†These authors have contributed equally to this work
‡ORCID: Zongxin Ling, orcid.org/0000-0001-9662-099X
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.