
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol., 17 August 2022
Sec. Microbes and Innate Immunity
Volume 12 - 2022 | https://doi.org/10.3389/fcimb.2022.834895
This article is part of the Research TopicThe Role of Autophagy in Infectious DiseasesView all 15 articles
 Amanda Demeter1,2,3
Amanda Demeter1,2,3 Anne-Claire Jacomin4†
Anne-Claire Jacomin4† Lejla Gul1
Lejla Gul1 Ashleigh Lister1
Ashleigh Lister1 James Lipscombe1
James Lipscombe1 Rachele Invernizzi2,5†
Rachele Invernizzi2,5† Priscilla Branchu2†Iain Macaulay1
Priscilla Branchu2†Iain Macaulay1 Ioannis P. Nezis4
Ioannis P. Nezis4 Robert A. Kingsley2,5
Robert A. Kingsley2,5 Tamas Korcsmaros1,2,6*‡
Tamas Korcsmaros1,2,6*‡ Isabelle Hautefort1
Isabelle Hautefort1Macroautophagy is a ubiquitous homeostasis and health-promoting recycling process of eukaryotic cells, targeting misfolded proteins, damaged organelles and intracellular infectious agents. Some intracellular pathogens such as Salmonella enterica serovar Typhimurium hijack this process during pathogenesis. Here we investigate potential protein-protein interactions between host transcription factors and secreted effector proteins of Salmonella and their effect on host gene transcription. A systems-level analysis identified Salmonella effector proteins that had the potential to affect core autophagy gene regulation. The effect of a SPI-1 effector protein, SopE, that was predicted to interact with regulatory proteins of the autophagy process, was investigated to validate our approach. We then confirmed experimentally that SopE can directly bind to SP1, a host transcription factor, which modulates the expression of the autophagy gene MAP1LC3B. We also revealed that SopE might have a double role in the modulation of autophagy: Following initial increase of MAP1LC3B transcription triggered by Salmonella infection, subsequent decrease in MAP1LC3B transcription at 6h post-infection was SopE-dependent. SopE also played a role in modulation of the autophagy flux machinery, in particular MAP1LC3B and p62 autophagy proteins, depending on the level of autophagy already taking place. Upon typical infection of epithelial cells, the autophagic flux is increased. However, when autophagy was chemically induced prior to infection, SopE dampened the autophagic flux. The same was also observed when most of the intracellular Salmonella cells were not associated with the SCV (strain lacking sifA) regardless of the autophagy induction status before infection. We demonstrated how regulatory network analysis can be used to better characterise the impact of pathogenic effector proteins, in this case, Salmonella. This study complements previous work in which we had demonstrated that specific pathogen effectors can affect the autophagy process through direct interaction with autophagy proteins. Here we show that effector proteins can also influence the upstream regulation of the process. Such interdisciplinary studies can increase our understanding of the infection process and point out targets important in intestinal epithelial cell defense.
Invasion and survival of intracellular bacterial pathogens within mammalian cells results from the timely expression of an arsenal of virulence factors, often horizontally acquired (Marcus et al., 2000; Hansen-Wester and Hensel, 2001; Nieto et al., 2016). The zoonotic Salmonella enterica subsp. enterica serovar Typhimurium pathogen (S. Typhimurium for short) is no exception to that rule (Srikumar et al., 2015; Colgan et al., 2016; Powers et al., 2021). Like many other enteric pathogens, S. Typhimurium hijacks the host cell machinery to enter the host cells, to hide from the innate immune system and ultimately to survive and spread to the next host (Alpuche Aranda et al., 1992; Lucas and Lee, 2000; Kröger et al., 2013). For that, Salmonella expresses and coordinates the secretion of its effectors directly into the host cell cytosol through type three secretion systems (T3SS), encoded on two Salmonella Pathogenicity Islands (SPIs) (Marcus et al., 2000; Schlumberger and Hardt, 2005; Lou et al., 2019).
Upon entry, S. Typhimurium will reside in a membrane-bound vacuole called the Salmonella-containing vacuole (SCV), where it may replicate. Salmonella entry is mediated by molecular mimics of host proteins (such as Guanine Exchange Factors (GEFs)) through effectors, including SopE, SopE2 and SopB, that activate host Rho-GTPases, RAC1, CDC42 (Schlumberger and Hardt, 2005), and the GTPase activating protein (GAP) SptP that subsequently deactivates Rho-GTPases, resolving host cell apical changes (Srikanth et al., 2011). In 20% of the cases entry into non-phagocytic cells is followed by an escape of Salmonella from the SCV (Brumell et al., 2002; Beuzón et al., 2002; Castanheira and García-Del Portillo, 2017). The resulting cytosolic Salmonella population must then adapt to the cytosol environment by mechanisms that are not fully elucidated, although SPI-1 effectors, including SopB, SptP, SipA, SopA, SopB, SopD and SopE may play additional roles impacting cytosolic Salmonella (Kubori and Galán, 2003; Drecktrah et al., 2005; Giacomodonato et al., 2007). Consistent with the idea that SopE could play a critical role in the adaptation of the pathogen to the host cell cytosol, SopE and SopE2 remain detectable on the SCV membrane up to 6 hours post-infection (Vonaesch et al., 2014).
Autophagy is a ubiquitous process crucial for cell homeostasis and stress survival of eukaryotic cells. Double membrane structures called autophagosomes are generated inside the cells and engulf superfluous organelles and proteins as well as invading pathogens. Autophagosomes then fuse with lysosomes, leading to degradation of the content (Feng et al., 2014; Cicchini et al., 2015; Sorbara and Girardin, 2015). By eliminating intracellular pathogens, such as bacteria and viruses (Sorbara and Girardin, 2015), autophagy assists the immune system in fighting infectious agents.
Over 38 proteins are involved in the autophagy process, each being temporally regulated throughout the different stages of the process: initiation, cargo recognition by the ATG ubiquitination system, membrane nucleation permitting the double membrane autophagosome formation, maturation of the compartment and fusion with the lysosome (Quan and Lee, 2013; Cicchini et al., 2015; Türei et al., 2015). We have previously described a computational pipeline allowing identification of interactions between secreted effector proteins of bacterial pathogens and autophagy core proteins (Sudhakar et al., 2019). In another previous study, we had generated and manually curated an autophagy protein interaction network in which we grouped the core autophagy proteins based on the following phases of autophagy: induction; cargo recognition and packaging; Atg protein cycling; vesicle nucleation; vesicle expansion and completion; transport of autophagosome; fusion with the lysosome (Kubisch et al., 2013).
Among those proteins, the MAP1LC3B (LC3-II) receptor, a ubiquitin-like protein, plays a critical role in the capture of the cargo into the autophagosome. When the autophagy process is activated, MAP1LC3B gets lipidated and associated with the double membrane of the autophagosome. In concert with autophagy adaptors (e.g. p62, NDP52, OPTN), MAP1LC3B binds to the targeted protein, ensuring its capture for lysosomal degradation (Shaid et al., 2013; Khaminets et al., 2016).
Although autophagy is a robust clearing process against intracellular pathogens, some infectious agents, including S. Typhimurium have developed ways to escape or hijack autophagy for their own benefit. S. Typhimurium can subvert host autophagy at several stages of this process (Baxt et al., 2013; Baxt and Xavier, 2015; Sorbara et al., 2018). Salmonella secretes several effectors that have been proposed to interact with the ubiquitin pathway, such as the E3 ligases SopA, SspH1, SspH2 and SlrP, and the deubiquitinases SseL and AvrA (Baxt et al., 2013; Herhaus and Dikic, 2018). SopB can prevent fusion of the autophagosome with the lysosome (Weigele and Alto, 2010). Furthermore, cytosolic S. Typhimurium interacts with autophagy proteins, particularly MAP1LC3B and p62 proteins (Yu et al., 2014). Autophagy can also promote bacterial growth by sealing damaged SCVs maintaining a suitable environment for replication (Kreibich et al., 2015). It is therefore apparent that Salmonella-mediated modulation of autophagy is a more complicated and dynamic process that remains to be fully elucidated.
In this study, we present a network approach and describe its application to predict interactions of Salmonella effectors with host transcription factors, potentially resulting in changes in expression of key autophagy genes. We experimentally validate this prediction for the GEF-mimicking effector SopE and show that it can also modulate the flux of autophagy later on during infection.
Protein-protein interactions (PPIs) were inferred by domain-domain interaction (DDI) prediction using the MicrobioLink pipeline (Andrighetti et al., 2020). Briefly, DDIs with high confidence values (interactions predicted by at least two different in silico methods or using multiple sources) were collected from the DOMINE resource (Yellaboina et al., 2011). We assumed that Salmonella and human proteins are connected if the interacting domains were represented in the database. The interaction prediction was merged with already existing predictions (Krishnadev and Srinivasan, 2011; Kshirsagar et al., 2012) and experimentally validated transcription factor-gene interactions related to autophagy.
Scripts for processing the interaction predictions, databases and other tables were written in R (See Table S1 for a list of host transcription factors and bacterial effectors found to interact). For the network analysis the Cytoscape network visualization program was used (Shannon et al., 2003). Autophagy core genes and their direct transcriptional interactions were downloaded from the following databases: Autophagy Regulatory Network (ARN, http://autophagyregulation.org/, (Türei et al., 2015), HTRIdb (http://www.lbbc.ibb.unesp.br/htri, (Bovolenta et al., 2012) and TRRUST (https://www.grnpedia.org/trrust/, (Han et al., 2015). All scripts are available at https://github.com/korcsmarosgroup/HMIpipeline.
HEK293 cells grown to 50% confluence were transfected with plasmids for overexpression of BirA-Myc-SP1 (gift from Markku Varjosalo; Addgene plasmid #167726) and GFP or GFP-SopE (Yuki et al., 2019) using GeneJuice transfection reagent according to the manufacturer’s recommendations. Protein extracts were recovered 24-hour post-transfection in lysis buffer (20mM Tris ph7.5, 0.5% Triton-X100, 150mM NaCl, 2mM EDTA) supplemented with EDTA-free proteases inhibitors cocktail (04693132001, Roche) and benzonase (1.03773.1010, Millipore). Co-immunoprecipitations were performed on cleared lysates with GFP-Trap Magnetic beads (gtd, Chromotek) overnight at 4°C. Four consecutive washes with the lysis buffer were performed before the bead suspension was added in sample loading buffer (Sigma) and incubated for 5-10 min at 95°C. Inputs and IP samples were loaded onto 4-20% polyacrylamide gels and were transferred onto PVDF membranes (cold wet transfer in 10% ethanol for 1h at 100V). Membranes were blocked in 5% non-fat milk in TBST (0.1% Tween-20 in TBS) for 1 h. Primary antibodies anti-Myc (Cell Signaling #2276) and anti-GFP (sc-9996, Santa Cruz Biotechnology) diluted in TBST were incubated overnight at 4°C with gentle agitation. HRP-coupled secondary antibodies binding was done at room temperature (RT) for 45 min in 1% non-fat milk dissolved in TBST. All washes of the membranes were performed for 10 min in TBST at RT. Probed membranes were developed using a Chemidoc imaging system (BioRad) and signal intensity quantification was performed by measuring the densitometry of appropriate bands on not overexposed membranes using FiJi/ImageJ.
All bacterial strains are listed below in Table 1. Gene deletion mutant strains were developed from the JH3009 (here referred to as wild type, wt) strain. The sopE gene was replaced in S. Typhimurium with the aphII gene conferring resistance to kanamycin, using a method LambdaRed recombination method (Datsenko and Wanner, 2000), with the exception that the red recombinase was supplied on the pSIM18 plasmid (Chan et al., 2007). The aphII gene from pKD4 was amplified using oligonucleotide primers 5’- CCTGCTATCTATATATAAATGAATTATGTACATATAAAAGGATCATTACCgtgtaggctggagctgcttcg-3’ and 5’- GGTTCATATTAATCAGGAAGAGGCTCCGCATATTTTTTGGTTTTTCAGTGTcatatgaatatcctccttagt-3’ and introduced into strain SL1344 containing the pSIM18 plasmid. Recombinant transformants were selected on LB agar plates containing 50 µg ml-1 kanamycin. The sopE gene deletion was reintroduced into SL1344 by P22 transduction to minimise off-target mutations, as described previously (Kingsley et al., 1999). The genotype was confirmed by PCR amplification across the sopE locus using oligonucleotides 5’-CAGATGGACATAGCATTTGC-3’ and 5’-ATGACGGTTTAGCTCCGGAG-3’.

Table 1 Salmonella enterica Typhimurium strains, generated and used in this study.
Combination of the ɸ(ssaG’-gfp+) intracellular reporter fusion with both the sifA and sopE gene deletions was carried out as follows in the SL1344 genetic background. The Kanamycin resistance cassette replacing the sifA gene in TK0021 was excised from the chromosome using the yeast Flp recombinase expressed from the thermosensitive replicon pCP20. The pCP20 replicon was subsequently removed from TK0021 after culture at 40°C in non-selective medium. The sopE, KmR deletion and the ɸ(ssaG’-gfp+), CmR transcriptional fusion were transduced into the sifA, KmS new strain by P22 phage transduction, generating the strain TK0026 used in this study (See Table 1) (Gemski and Stocker, 1967). The generated the strain TK0026, lacking both SifA and SopE and carrying a GFP reporter system to indicate intracellular Salmonella location in infected epithelial cells.
Bacterial strains were grown in 5 ml of LB broth overnight (Sambrook and Russell, 2001) at 37°C at 250 rpm. For invasion assays, a 1:100 dilution of the overnight bacterial culture was grown in 25 ml of LBS (LB containing a total of 0.3 M NaCl) in 250 ml conical flasks until an optical density of 1.2 was obtained at 600 nm (A600). Antibiotics were added as required at the following final concentrations (kanamycin, 50 mg ml-1; chloramphenicol, 10 mg ml-1).
HT-29 human colon cancer epithelial line (HTB-38, ATCC) was cultured in DMEM supplemented with 10% heat-inactivated Fetal Bovine Serum (FBS) and 2 mM L-Glutamine at 37°C, 5% CO2. HT-29 epithelial cells were seeded into 6- and 24-well cell culture plates at a density of 3.2x106 and 2x105 cells/well, respectively, to obtain 80% confluency by the day of the invasion assays (48h later). Six well-plates were used to generate enough infected epithelial cells for RNA extraction and qPCR autophagy gene expression analysis. Twenty four-well plates were used for immunofluorescence microscopy monitoring of autophagy flux upon infection. In those plates, HT-29 cells were seeded on 13mm diameter glass round coverslips. Three biological replicates were used for each condition and time point. Where necessary, autophagy was induced by treating the cells with 30 µg ml-1 rapamycin or DMSO only prior to and during the 6h long infection (17h total). On the day of the invasion assay, cells were washed twice in non-supplemented DMEM followed by the infection. Bacterial suspensions were prepared in DMEM from the LBS sub-culture of the Salmonella strains (see Bacterial strains and culture conditions) at a multiplicity of infection (MOI) of 10 bacterial cells per mammalian cell. Infected cells were incubated for 30 minutes at 37°C, 5% CO2. The infection medium was then replaced with a complete medium containing 30 µg ml-1 gentamicin for 30 minutes to kill the remaining extracellular Salmonella cells. For the rest of the experiment, the gentamicin concentration was then reduced to and maintained at 5 µg ml-1.
For the assay allocated to autophagy flux bioimaging, the medium was removed at 6h post infection (p.i.). Cells were washed twice in Dubelcco’s phosphate-buffered saline (DPBS; D8537, Sigma Aldrich), fixed in 4% paraformaldehyde at room temperature for 20 min, and washed twice for 5 min at room temperature in DPBS prior to immunofluorescence labelling.
At 2h and 6h post-infection, the medium was removed and cells were washed twice in DPBS. Each sample well was trypsinised in 50% Trypsin-Versene (EDTA), 49.6% DPBS, 2mM EDTA and 5 µg ml-1 gentamicin for 5 minutes at 37°C. The trypsin reaction was stopped in 89.6% DPBS, 10% FBS, 2mM EDTA and 5 µg ml-1 gentamicin (i.e. FACS buffer). Single-cell suspensions were obtained by pipetting several times. Cells were washed and resuspended in FACS buffer. Infected cells and bystanders were separated by Fluorescence-Activated Cell Sorting on a BD FACSMelody machine (BD Biosciences). Sorting gates were set based on negative cells (from an uninfected well) and positive control cells (from an infected well) based on the level of GFP present in each cell. Eight pools of 50 epithelial cells were sorted from each condition and time point (4 x GFP- as non-infected cells or “bystanders’’ and 4 x GFP+ as “Salmonella-containing epithelial cells”) into 96-well plates containing 2 μl of lysis buffer (0.2% Triton X-100 and 2 U µl-1 RNase inhibitor). Samples were then processed as detailed in the RNA samples extraction and processing section.
The low input RNA extraction and reverse transcription were carried out following the SmartSeq2 protocol previously described (Picelli et al., 2013). Reverse transcription was performed as in the following steps. First, Oligo-dT30VN primer was added to the sample lysates at 2.5 µM, 2.5 nM each dNTP final, and priming reactions were incubated at 72°C for 3 mins. First strand synthesis was subsequently initiated by addition of 1.03 µM custom template switching oligo, 6.18 µM MgCl2, 1.03 M Betaine, 5.12 mM DTT, 1.03x Superscript First Strand Buffer, 0.52 U µl-1 RNase Inhibitor, 10.3 U µl-1 SuperScript II reverse transcriptase in nuclease-free water. The reverse transcription reactions followed the successive steps: 42°C 90 mins, 10×(50°C 2 mins, 42°C 2 mins), and 70°C 15 mins. Finally, 15 ul PCR mastermix (1.6x KAPA HiFi Hotstart Readymix, KAPABIOSYSTEMS, 0.16 µM IS primers, nuclease-free water) was added to each sample. PCR cycles were as follows, 98°C 3 mins, 21×(98°C 20 secs, 67°C 15 secs, 72°C 6 mins), 72°C 5 mins). PCR products were cleaned up with 0.8x volumes of Ampure XP and 80% ethanol. Samples were then eluted in 20 µl 10 mM Tris-HCl.
Taqman gene expression analysis was conducted on the SmartSeq2 amplified cDNA. In brief, TaqMan™ Fast Advanced Master Mix (4444557, ThermoFisher Scientific) was used for all qPCR reactions, assay probes and samples were dispensed into 384-well Roche-style qPCR plates (4titude, 4ti-1381) using a Mosquito HV automated liquid handling instrument (SPT Labtech), to a final reaction volume of 1.6µl (80nl 20× TaqMan® Gene Expression Assay, 800nl 2× TaqMan® Gene Expression Master Mix, 720nl normalised cDNA). Samples were then analysed on a Roche Lightcycler 480. The Taqman gene expression assay probes were used for the MAP1LC3B test gene and the beta-2-microglobulin B2M reference gene (Hs00797944_s1 4453320 and Hs00984230_m1 4453320, respectively, ThermoFisher Scientific).
Gene expression was normalised to the B2M internal reference gene and plotted as log2^-deltaCT (Rao et al., 2013). First, technical replicates with the smallest and largest CT values (raw CT in case of potential reference gene and normalised CT values in case of genes of interest) were excluded for each gene within each condition.
MAP1LC3 and p62 were labelled as previously done (Sudhakar et al., 2019). In brief, for MAP1LC3B/LC3B immunostaining, cells were quenched at room temperature in 50 mM NH4Cl in DPBS for 10 min, then permeabilized in methanol for 5 min and washed in DPBS 3 times for 5 min. This was followed by blocking the samples at room temperature in 1% bovine serum albumin (BSA) Fraction V (05479, Sigma-Aldrich) in DPBS for 30 min. The rabbit anti-MAP1LC3B/LC3B (ab48394, Abcam) antibody was applied overnight at 4°C. The antibody labelling solution was diluted at a 1:2000 dilution in DPBS containing 1% BSA fraction V (05479, Sigma Aldrich). SQSTM1/p62 immunolabeling was performed as follows. Fixed cells were permeabilized and blocked in a solution containing 1% BSA and 0.1% saponin (84-510, Fluka), in DPBS for 30 min at room temperature. The rabbit anti-SQSTM1/p62 antibody (ab91526, Abcam) was applied overnight at 4°C at a f 1:6000 dilution in DPBS containing 1% BSA and 0.1% Saponin.
The FITC-conjugated anti-GFP antibody (ab6662, Abcam) was applied overnight at 4°C in all samples at a 1:200 dilution in all primary solutions.
All primary antibodies were washed 3 times in either 1% BSA in DPBS (MAP1LC3B/LC3B) or 1% BSA and 0.1% saponin in DPBS (SQSTM1/p62). Alexa Fluor 594-conjugated anti-rabbit secondary IgG (ab91526, Abcam) was diluted 1 in 1000 and applied to all samples at room temperature for 1 h. All samples were counterstained with DAPI at a dilution of 1:2000 in the buffer respective to each primary antibody. Samples were then washed 3 times in their respective buffers, once in water and finally mounted on microscopy glass slides. Coverslips were mounted in Aqua-poly/mount anti-fading compound (18606, Polysciences Inc.). Coverslips were left to set, sealed using nail varnish and stored at -20°C until observation. Slides were imaged on a Zeiss LSM710 microscope, using a 100x Apochromat (100x/1.4 Oil DIC plan Apo) oil immersion objective. Focal plan and laser power/gain was kept constant throughout the acquisition process. Over 500 epithelial cells were imaged per condition tested. The acquisition was semi-blind with conditions revealed post-analysis.
The analysis of SQSTM1/p62 and LC3 staining was done using semi-automated macros within FiJi (Image J2) software. To avoid unconscious-bias, imaged areas were chosen randomly based solely on the DAPI staining. DAPI staining was used for the identification of nuclei and individual cells were identified by extension of the nuclei mask. Segmented cells touching the edge of the images or artefacts (small objects) were eliminated. Individual regions of interest (ROIs) were saved for each image and subsequently used for quantification of SQSTM1/p62 and LC3 staining. Intensity and puncta information for individual cells in each fluorescent channel was exported into Excel spreadsheet and used for statistical analysis.
Distribution normality of data points was determined by the Shapiro-Wilk test and the equality of variance was determined by the Levene’s test. To compare 3 or more groups, one-way ANOVA (for normal distribution) or Kruskal-Wallis (for non-normal distribution) tests were performed using R. On statistically different samples the appropriate post hoc test was applied: Tukey following ANOVA and kruskalmc following Kruskal-Wallis. T-tests and kruskalmc tests were performed to compare LC3 and p62 protein levels in wild type Salmonella-infected cells and cells infected by the ΔsopE deletion strain derivative.
The potential effect of S. Typhimurium effector proteins on autophagy gene expression was analysed using a network of predicted and experimentally validated interactions. We predicted pathogen-host interactions based on known domain-domain interactions and supplemented this with two predictions previously described (Krishnadev and Srinivasan, 2011; Kshirsagar et al., 2012) making up our merged list of interactions (See Table S1). The source proteins were filtered for secreted S. Typhimurium effectors (Miao and Miller, 2000; Ehrbar and Hardt, 2005; Haraga et al., 2008; Figueira et al., 2013), and the target proteins were filtered for those that had previously been validated experimentally as transcription factors of autophagy-core genes (Bovolenta et al., 2012; Han et al., 2015; Türei et al., 2015). The core autophagy network was extended with an additional gene, ATG4D and its connection to GABARAPL1 (Betin and Lane, 2009). As further filtering steps, transcription factors were kept if they were predicted to connect to only one or few Salmonella effectors (eliminating some potential false positives) while at the same time regulating more than one of the core autophagy genes.
The first layer of the constructed network was centred around the interactions between S. Typhimurium virulence proteins (excluding SlrP) and human autophagy core-regulating transcription factors. Regarding these interactions there was no overlap between the three sources, and only three interactions of SlrP with CTCF, YY1 and SP1 were overlapping between the prediction of Kshirsagar et al. and our prediction. This layer of the final network contains 71 connections from Kshirsagar, 0 connections from Krishnadev and 4 connections from our prediction (Krishnadev and Srinivasan, 2011; Kshirsagar et al., 2012). On the human side there were 154 transcription factor-autophagy core interactions from ARN, 105 from HTRI and TRRUST database, with altogether 33 transcription factors, 35 core autophagy genes and 159 regulatory interactions between them.
Our network analysis highlights that several Salmonella effectors can impact several transcription factors. For example, effectors such as SseI, SseL, SlrP and SspH2 target 9, 10, 16 and 33 transcription factors, respectively (Figure 1A). This reflects the multiple routes Salmonella has evolved to subvert host intracellular clearance mechanisms, here by affecting all stages of the autophagy process (from induction, through to the autophagosome formation to the fusion with lysosomes; Figure 1). We selected six S. Typhimurium proteins that were not predicted to connect to too many of the human transcription factors (except SspH2) to simplify our further analysis. One of these selected effectors is SlrP. SlrP, that can be secreted through the SPI-1 and -2 apparatus as well, and is an E3 ubiquitin ligase, hence potentially able to interact with ubiquitin-mediated autophagy. Moreover, it has overlapping predicted interactions in two of the PPI predictions, highlighting the power of our computational approach. Two additional effectors, SopE and SopE2, are SPI-1 effectors and mimic host GEFs. They are particularly important in the membrane ruffling associated with Salmonella entry into the non-phagocytic epithelial cells. Yet, they are also expressed in later stages of the invasion and present in our network possible interactions with two autophagy transcription factors. The remaining of the selected proteins, SseI, SseL and SspH2 are SPI-2 virulence effectors, which means that they are probably secreted in the later stages of invasion and SseL and SspH2 are also among the proteins that can alter ubiquitination.
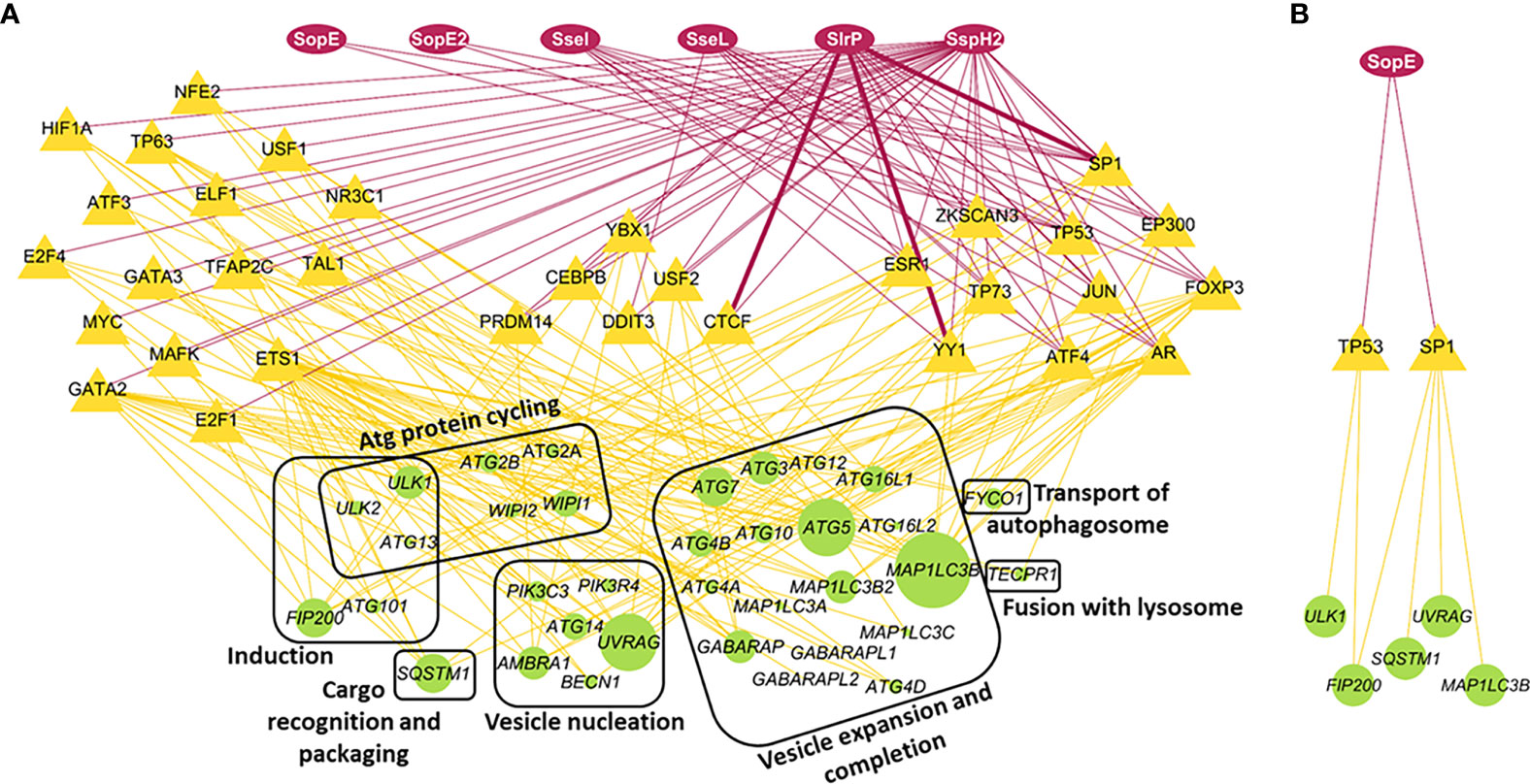
Figure 1 Interaction network between selected Salmonella Typhimurium effectors, host cell transcription factors and autophagy core genes. (A) Network analysis of potential interactions between Salmonella and host autophagy. Red ovoid nodes are the selected Salmonella effector proteins. Red edges are PPI predictions. Thin edges were predicted by one of the three methods. Thick edges were predicted by two of the three methods. Yellow triangular nodes are host transcription factors that Salmonella effectors can influence. These are clustered according to the number of Salmonella effectors they are targeted by. Yellow edges reflect transcriptional regulation of core autophagy genes (round green nodes) by the transcription factors. The size of the green nodes is proportional to the number of transcription factors they are connected to. (B) Subnetwork illustrating the potential interaction of the Salmonella effector SopE with transcription factors affecting specific autophagy core genes. The same layout was used here as for the large network.
Considering SopE’s role in altering RhoGTPases downstream effect and persisting long enough in the host cells to be exposed to the host cell machinery, including the autophagy process (Schlumberger and Hardt, 2005; Vonaesch et al., 2014), it was subsequently selected for experimental validation of our predictions. When focusing on the SopE potential interactions with autophagy regulators, we predicted that SopE can interact with only two key transcription factors, TP53 regulating genes in the autophagy induction phase, and SP1, controlling the formation and expansion of the autophagosome compartment (Figure 1B). These two steps of autophagy are likely to be influenced differently depending on the location of the intracellular Salmonella cells, either within a damaged SCV or cytosolic. We investigated further if the GEF-mimicking SopE influenced the autophagy process at different levels, focusing only on the SopE and SP1 potential interaction.
Our network analysis suggests that one way Salmonella can modulate autophagy flux is through altering the regulation of autophagy-related gene expression. For example, we predicted that SopE can interact with SP1 and TP53 (Figure 1B). Our study focuses later on the modulation of the autophagy process through MAP1LC3B, so we centred our experimental validation on the potential SopE-SP1 interaction. To validate this hypothesis, we performed a co-immunoprecipitation assay between GFP-SopE and BirA-Myc-SP1 recombinant proteins, ectopically expressed in HEK293 cells (Figure 2A). We observed that SP1 is enriched over sixfold with GFP-SopE compared to GFP alone (Figure 2B), confirming the direct interaction of the Salmonella SopE protein with the SP1 autophagy regulator. This suggests that through interaction with SP1, SopE can alter the expression of SP1-target genes, such as MAP1LC3B.
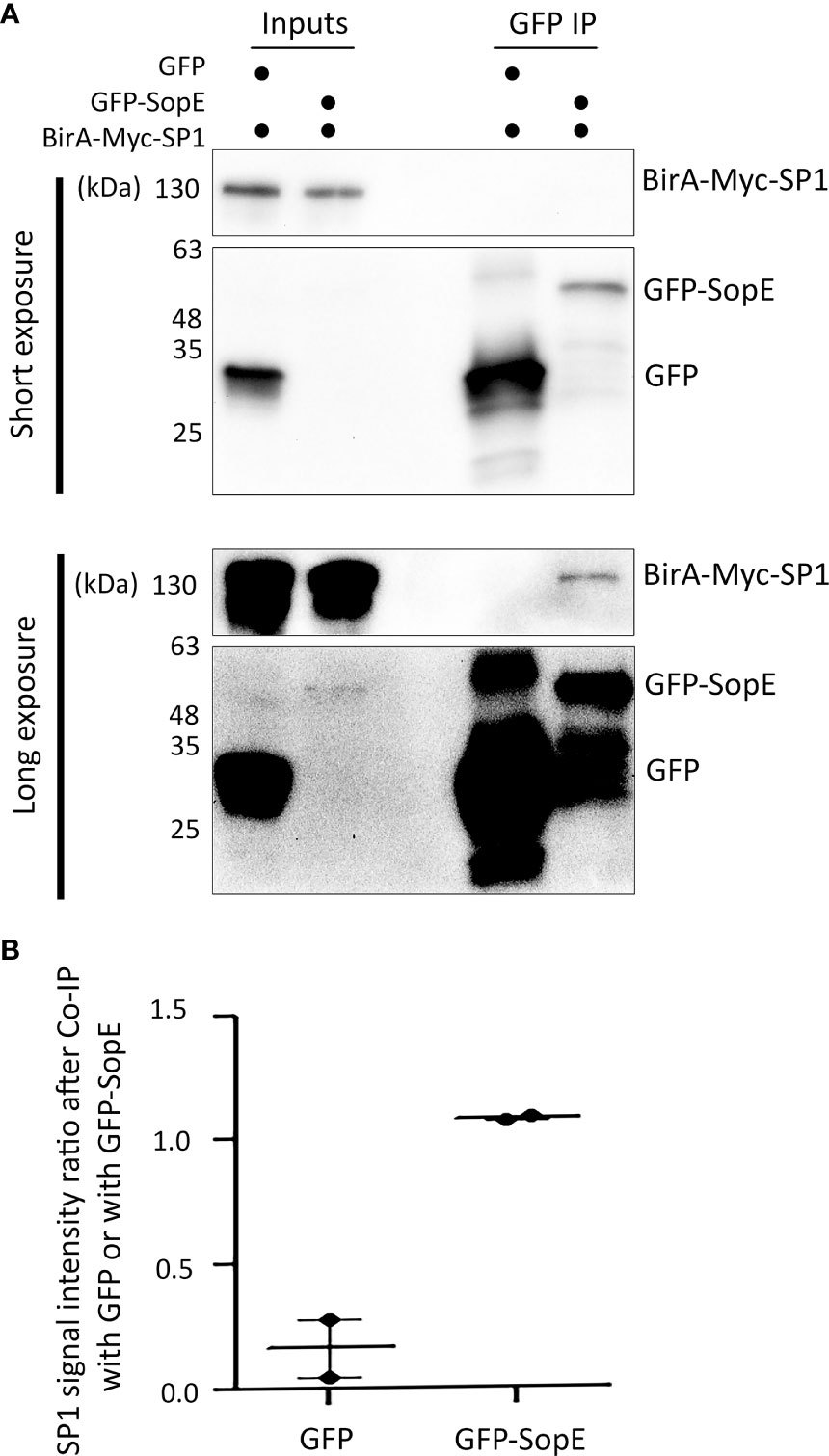
Figure 2 The Salmonella effector SopE can bind to the human transcription factor SP1, as predicted. (A) Co-immunoprecipitation of GFP-SopE and BirA-Myc-SP1 ectopically expressed in HEK293 cells. (B) SP1 signal intensity ratio after Co-IP with GFP or with GFP-SopE showing enrichment of SP1 when GFP-SopE is used compared with GFP alone, based on two independent measurements.
SopE was shown to be essential in the SCV formation during invasion and it was also shown that some of the intracellular SopE protein remains associated with the SCV membrane at later time point during infection at a time when autophagy process is induced (Schlumberger and Hardt, 2005; Vonaesch et al., 2014). We first tested the functional importance of the direct interaction of SopE with SP1 by monitoring whether SopE influences the expression of the key MAP1LC3B gene, directly downstream of the SP1 transcription factor. For that, we infected HT-29 epithelial cell monolayers with either a sopE+ strain of S. Typhimurium (=wt), or its ΔsopE deletion strain derivative. Both strains contain a ɸ(ssaG’-gfp+) transcriptional fusion, allowing intracellular GFP+ Salmonella containing HT-29 cells identifiable in a pool of infected and non-infected cells. At 2 and 6h p.i., MAP1LC3B RNA levels were quantified in Salmonella-containing cells or in epithelial cells that did not contain Salmonella despite being part of the infected cells (from then on called “bystanders”). MAP1LC3B RNA levels revealed that SopE impacts on the transcriptional levels of MAP1LC3B compared with non-infected cells (Figure 3), consistent with this pathogen-derived protein interacting with SP1 transcription factor (Figure 2).
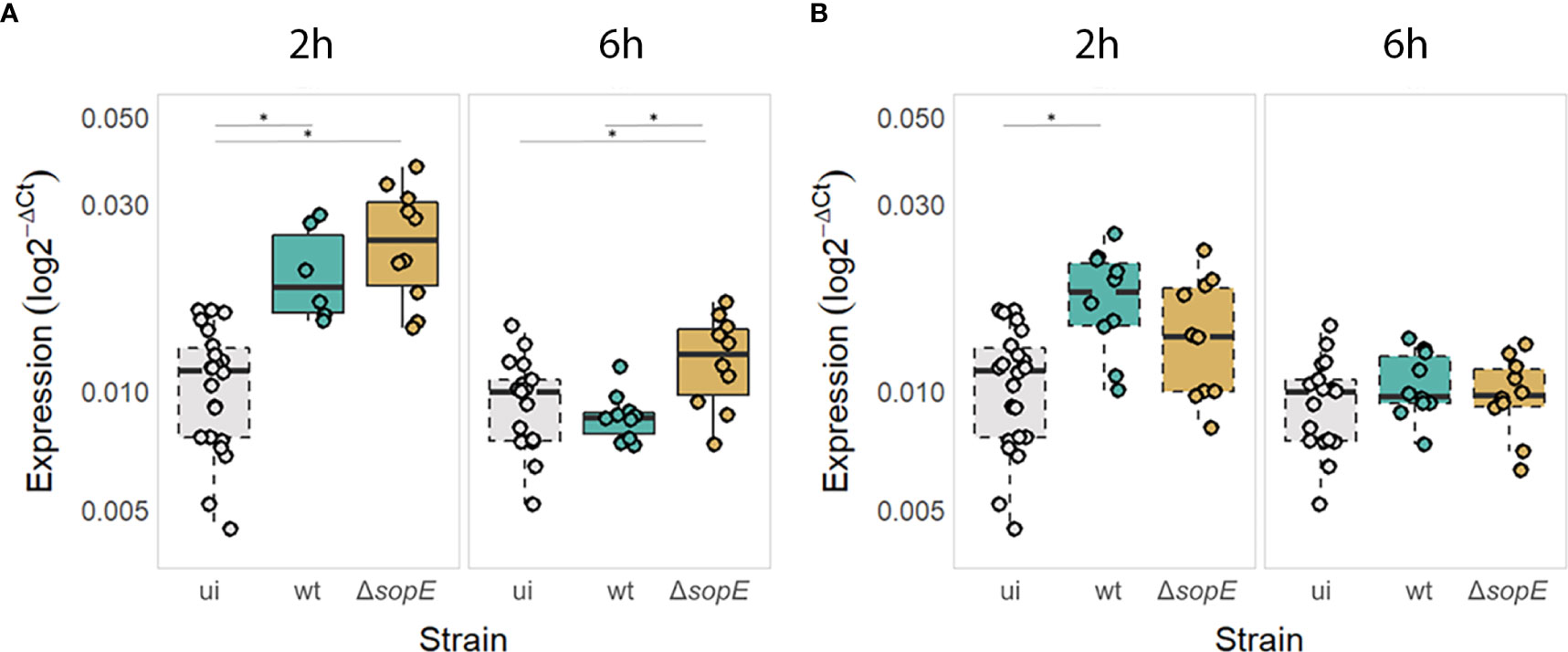
Figure 3 MAP1LC3B gene expression is increased at the early stages of Salmonella infection but decreased at later stages in a SopE-dependent manner in Salmonella-containing epithelial cells (A) but not in bystander cells (B). MAP1LC3B expression levels expressed as log2-ΔCT so that uninfected cells (ui) are also displayed. Continuous borders = infected cells containing Salmonella (wt or mutant) and dashed borders = bystander epithelial cells not containing Salmonella or uninfected cells. * p<0.05.
Surprisingly, although SopE is essential for the early internalisation of Salmonella inside epithelial cells, it is not required for upregulating autophagy early during infection, e.g. 2h p.i. (Figure 3A). Epithelial cells upregulated MAP1LC3B expression irrespective of the presence or absence of SopE. Interestingly, MAP1LC3B gene expression was upregulated significantly even in bystander epithelial cells, suggesting that either the bystander cells respond to Salmonella-derived compounds that are sensed when the pathogen is not internalised or that they respond to mediators produced by the Salmonella-containing epithelial cells directly (Figure 3B).
However, at a later time point, when Salmonella started proliferating intracellularly (6h p.i.), we observed that the level of MAP1LC3B expression returned to that of non-infected cells in a SopE-dependent manner. This was particularly the case in HT-29 cells that contained Salmonella and not so for bystander epithelial cells that did not contain Salmonella cells (Figure 3B).
As well as modulating the regulation of core autophagy gene expression through specific transcription factors, we hypothesise that SopE could also influence the autophagy flux as it is retained at the SCV membrane site several hours after Salmonella internalisation. MAP1LC3, once lipidated and recruited to the membrane nucleation site, connects the cargo to the vesicle membrane through the SQSTM1/p62 adaptor protein. We chose to follow MAP1LC3 and SQSTM1/p62 associated with the autophagosome as these two proteins are good indicators of the autophagy flux taking place in a mammalian cell (Xu et al., 2018). To address this, LC3 number of puncta and p62 intensity were assessed by immunofluorescence confocal bioimaging 6h p.i., at a similar time when SopE is still retained at the SCV membrane (See Materials & Methods section).
When autophagy is not pre-induced before infection by Salmonella, no significant difference in the number of LC3 puncta is observed at 6h p.i. (Figure 4A). However, the p62 dots intensity is significantly greater in cells infected with Salmonella that lack sopE (Figure 4B). This suggests that SopE, although dampening the expression of autophagy core genes at 6h p.i., simultaneously modulates the autophagy flux locally, at intracellular autophagy sites.
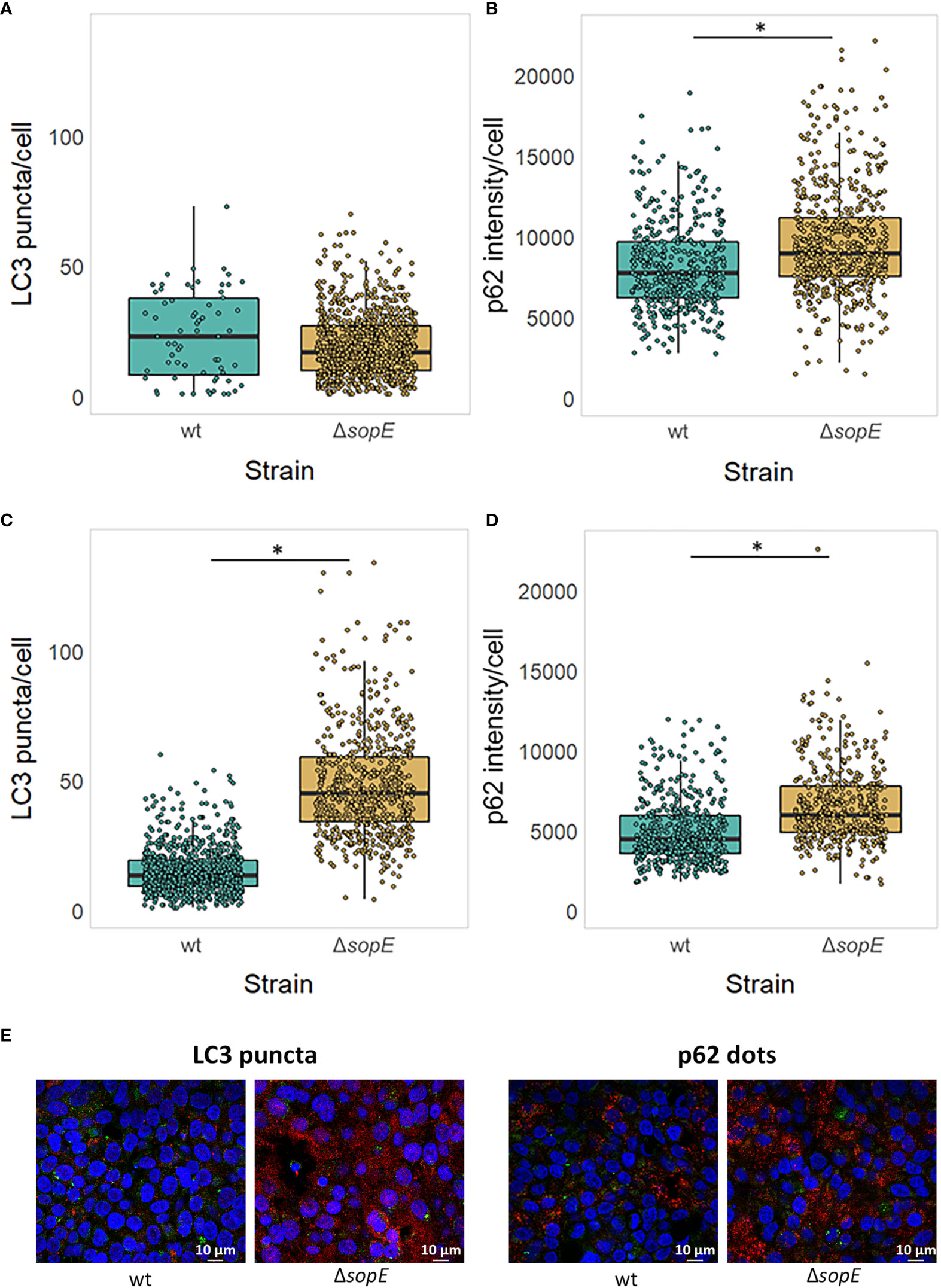
Figure 4 SopE dampens the autophagic flux by 6h post infection. LC3 puncta number and p62 dot intensity in HT-29 epithelial cells infected for 6h with either wt Salmonella strain or its ΔsopE gene deletion derivative. (A, B) HT-29 epithelial cells were infected with wt Salmonella and its ΔsopE gene deletion derivative strain as indicated before. (C, D) HT-29 cells were pre-treated with Rapamycin for 11h prior to and during the 6h-long infection (Maximum 17h). LC3 puncta (A, C) and p62 dot intensity (B, D) was quantified from HT-29 cells containing Salmonella. *p=0.05. (E) Micrographs of HT-29 cells pre-treated with Rapamycin showing LC3 puncta (red, left) and p62 dots (red, right), intracellular Salmonella cells (green) and nuclei (blue) illustrating panels (C, D).
Whether SopE acts on the autophagy flux, before or after that process is induced, is not yet clear. LC3 puncta number and p62 dots intensity were therefore monitored also after autophagy was experimentally triggered. Figures 4C, D clearly show that both LC3 puncta (C) and p62 dot intensity (D) increase significantly when SopE is absent, confirming that the dampening effect SopE normally has on autophagy by the time Salmonella starts to replicate intracellularly. SopE’s effect is even amplified when autophagy is already activated.
Finally, we questioned whether the fraction of SopE retained at the SCV membrane implies that Salmonella benefits from the SopE-dampening autophagy only when residing within an SCV.
To address the impact of Salmonella intracellular localisation on the role of SopE as modulator of the autophagy flux, we constructed a strain lacking SopE that would essentially be located in the epithelial cell cytosol. SifA effector protein of Salmonella is well known for being instrumental to the evolution and maintenance of the SCV (Beuzón et al., 2002), recruiting vacuolar ATPase to the SCV, permitting the SCV luminal environment to acidify (Beuzón et al., 1999; Martin-Orozco et al., 2006), and the formation and extension in certain host cells to Salmonella-induced Tubules (SITs) from the endosomal system for the intracellular survival of Salmonella in the host (Liss and Hensel, 2015). It was shown in diverse studies that Salmonella strains lacking SifA escape the SCV and start proliferating anarchically in the cytosol of non-phagocytic cells such as epithelial cells. We therefore constructed the strain TK0026 carrying the double ΔsifA and ΔsopE genes deletion (See Materials & Methods). Similarly done to the experiment shown on Figure 4, HT-29 epithelial cell monolayers were infected this time with either the strain lacking SifA only or the strain lacking both SifA and SopE. Figure 5 shows a mild, yet significantly different increase in the number of LC3 puncta in cells infected with Salmonella lacking SifA and SopE compared with the cells infected with Salmonella lacking SifA only, suggesting SopE could mildly influence MAP1LC3B lipidation and recruitment to the autophagosome membrane. However, the autophagy flux does not seem to be differing whether SopE is present or not once Salmonella is cytosolic, emphasising the specificity of SopE’s modulatory effect.
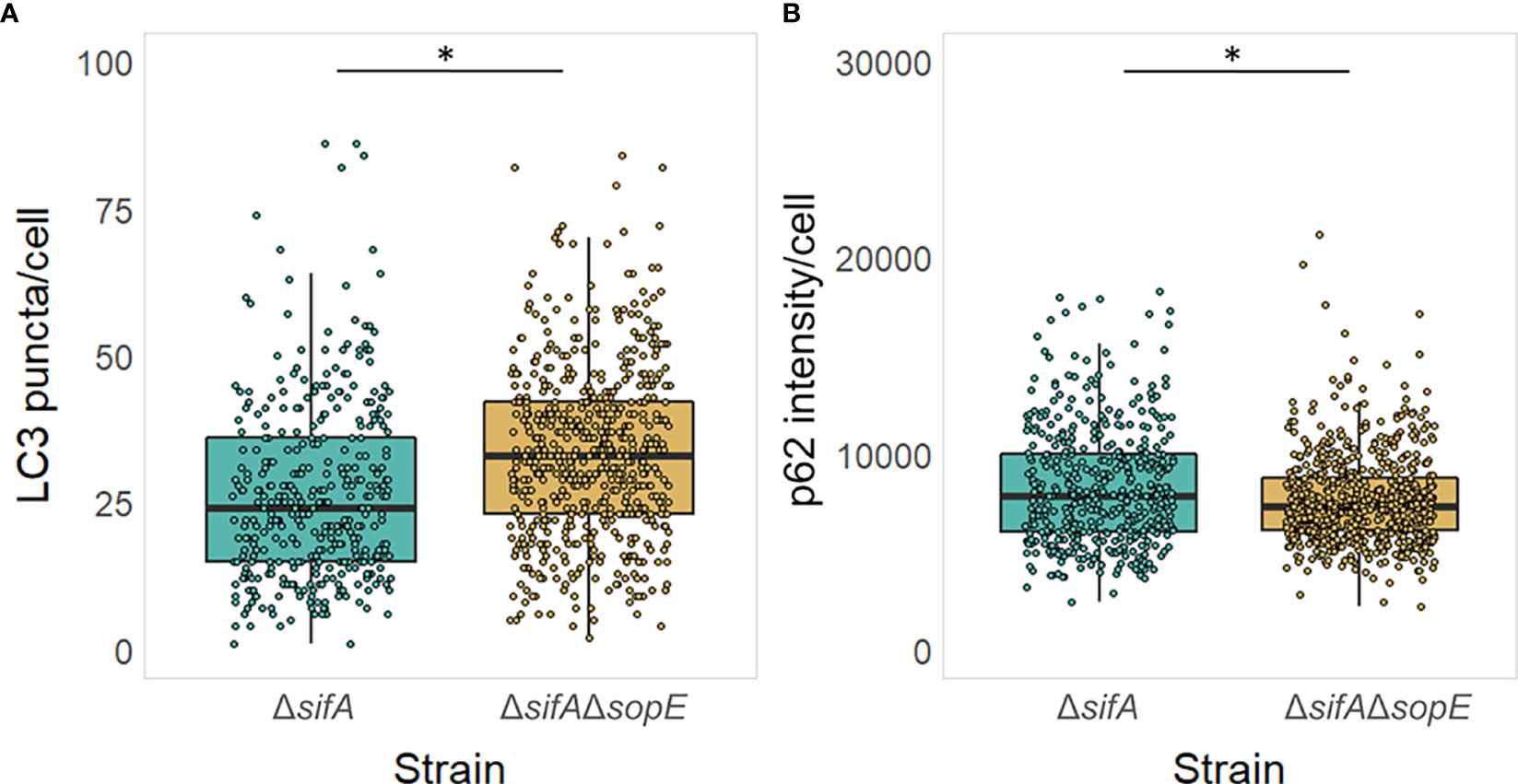
Figure 5 Cytosolic Salmonella no longer can dampen the autophagy flux in a SopE-dependent manner. (A, B) Autophagy was pre-induced with Rapamycin in HT-29 epithelial cell monolayers and maintained during the 6h infection with Salmonella, as it exacerbates the impact of SopE on modulating autophagy, making it easier to visualise. LC3 puncta (A) and p62 intensity (B) was quantified from HT-29 cells containing Salmonella. *p=0.05.
Our study predicts that several Salmonella virulence effectors proteins interact directly with some transcription factors involved in regulating autophagy, impacting the subsequent expression of core autophagy genes, such as MAP1LC3B. We also observed that Salmonella influences the autophagy flux at certain stages of infection; it does so in epithelial cells where Salmonella is still associated with the SCV. We had previously shown through protein-protein interaction network analysis that autophagy preferentially targets virulence effectors secreted by intracellular pathogens, and that pathogens have evolved mechanisms that conversely compromise the autophagy process. Autophagy-associated proteins are directly targeted by pathogens at different stages of the process (Sudhakar et al., 2019). Here we asked whether S. Typhimurium virulence effector proteins can influence this clearing process by acting on the regulatory level above the core autophagy genes, i.e. on the transcription factors that normally regulate them. Using protein-protein interaction predictions and transcription factor-gene interaction databases we built a network that links several S. Typhimurium virulence effectors to key transcription factors and predicted that S. Typhimurium can modulate the regulation of autophagy core gene expression. We showed that six SPI-1 effectors could interact with overall 33 transcription factors that normally regulate the expression of core autophagy genes (Figure 1A). As expected, SPI-2 SseL and SspH2 effectors showed a greater number of interactions with transcription factors, with 10 and all 33 regulatory targets, respectively. This reflects the adaptation of Salmonella to the host cell intracellular environment where SPI-2 secreted effectors are the main contributors to Salmonella survival and virulence (Haraga et al., 2008; Srikanth et al., 2011). Yet our study also highlighted new autophagy interactors among the Salmonella effector arsenal; for example, we showed that the Salmonella SseI [also called SrfH; (Thornbrough and Worley, 2012)] cysteine hydrolase can interact with 9 transcription factors modulating autophagy gene expression in epithelial cells. This effector might play an important role later in the infection, affecting the migration of macrophages and DCs (Jennings et al., 2017) although a single nucleotide polymorphism in the sseI gene that occurs naturally in some Salmonella strains prevents SseI from stimulating monocyte migration (Thornbrough and Worley, 2012). Our study suggests that SseI essentially impacts autophagy through its interaction with SP1 and TP53 transcription factors. We can envisage that, in addition to its regulatory role, it could also interfere with proteolytic processes taking place within epithelial cells through its cysteine hydrolase activity; initiation, execution, or inhibition of the autophagy process being highly dependent on proteases activity (Kaminskyy and Zhivotovsky, 2012).
Fewer SPI-1 effectors seem to interact with autophagy regulators (Figure 1A). This is in agreement with SPI-1 effectors being mostly involved in the entry of the pathogen into non-phagocytic mammalian cells (Lou et al., 2019). However, certain SPI-1 effectors have been shown to persist within the host cells, even past the point when Salmonella starts replicating. For instance, the SPI1 SlrP E3 ligase can also bind to a large number of transcription factors (Figure 1A) highlighting the importance of interfering with the ubiquitin pathway of the host, and possibly assisting the interaction of E3 ligases normally secreted through SPI-2 T3SS (e.g. SspH2; (Herhaus and Dikic, 2018).
The two Salmonella SPI-1 GEFs-mimicking effectors SopE and SopE2 seem to have a more specific impact on autophagy gene expression as they can interact with much fewer transcription factors (Figures 1A, B). SopE and SopE2 effectors can both target two transcription factors, TP53 and SP1, affecting autophagy induction through ULK1 and FIP200, respectively, and the autophagosome formation through SQSTM1/p62, UVRAG and MAP1LC3B, rather than the late lysosome-mediated clearance of the cargo. This restricted number of transcription factors potentially interacting with SopE and SopE2, as shown here for SP1 and SopE, suggests a very specific role of these effectors in the hijacking of autophagy by Salmonella. TP53 was also predicted to be a binding target of SopE and, although the role of this interaction was not studied here, its role in Salmonella-mediated autophagy gene expression regulation should be further explored. Indeed, when the transcription factor TP53 is inhibited or absent, increased autophagosome formation and overall autophagy flux is observed, suggesting that cytosolic TP53 reduces autophagy initiation. TP53 transcription factor acts upon autophagy modulation even when Salmonella remains cytosolic (Tasdemir et al., 2008; Kroemer et al., 2015). We suggest that SopE inhibits autophagy induction through its binding to TP53, maintaining this transcription factor in the cytosol and exacerbating its autophagy dampening effect. sopE2 is present in all pathogenic strains of Salmonella, while sopE is assumed to have appeared later in the Salmonella evolution by duplication of sopE2 and is present only in a subset of strains such as SL1344 used in this study (Table 1) (Bakshi et al., 2000; Mirold et al., 2001), suggesting a gained function in these strains. Indeed, SopE was also obtained from the strain responsible for a major epidemic in the 1970s and 1980s (Mirold et al., 1999). We focussed particularly on SopE to study its added impact on core autophagy gene expression and autophagy flux modulation by these Salmonella strains.
We first validated experimentally that SopE can bind to the SP1 transcription factor regulating MAP1LC3B expression (Figure 2), validating our prediction (Figure 1B). We then showed that, following the early increase in autophagy associated with the infection by Salmonella (2h p.i.), SopE contributes to the dampening of MAP1LC3B gene expression (Figure 3), and of the autophagy flux (Figure 4) in the HT-29 cell line. Indeed, at 6h p.i., this effect observed with the wildtype strain was no longer observed when SopE was missing. Acting on key transcription factors directly is likely to be an evolutionary selected mechanism for Salmonella to control host clearance function. We propose that SopE induces autophagosome formation through SP1. Our regulatory network analysis showed that SP1 transcription factor regulates several core autophagy genes, including MAP1LC3B and p62/SQSTM1 (Figures 1A, B). SP1 has also been shown to repress the autophagy process in malignant epithelial cells by dampening p62 expression (Xu et al., 2018). Here, we show that SopE dampens the autophagy flux. Indeed, accumulation of p62 protein is a commonly used indication for autophagy impairment or decrease. Here, we observed a decrease in p62/SQSTM1 protein associated with autophagosomes that depended on SopE being present (Figures 4C, D, E). We suggest that SopE, through its interaction with the SP1 transcription factor, negatively modulates the autophagy flux, protecting vacuolar Salmonella from lysosomal degradation.
We observed however that the effect of SopE in dampening autophagy only happens when Salmonella is associated with the SCV. Conversely, when Salmonella was mostly cytosolic due to the deletion of sifA, SopE no longer seemed to have the striking effect on autophagy dampening we saw when Salmonella remained mostly associated with the SCV compartment (Figures 4 vs 5). A previous study showed that infection-induced autophagy also contributes to the healing of damaged SCV by SPI1 proteins secreted through theT3SS, helping the endosomal/SCV maturation (Kreibich et al., 2015). This benefits consequently intravacuolar Salmonella cells, as these can proceed with SPI-2 effector-mediated intracellular survival and proliferation. In our study, the SopE protein no longer dampens autophagy flux in host cells where Salmonella has escaped the vacuolar compartment or resides within damaged SCV that will require part of the autophagy flux to repair the endosomal/SCV membrane. This process could take place in parallel to epithelial cell compartment size regulation which determines whether the pathogens will escape the vacuole or trigger enlargement of the vacuole by fusion with infection-associated macropinosomes as previously described (Stévenin et al., 2019).
The prediction that both SopE and SopE2 effectors can similarly affect autophagy highlights even more the importance of these molecular mimics across a broad spectrum of S. Typhimurium strains. In strains expressing both sopE and sopE2 genes, modulation of autophagy induction and autophagosome formation might play a key role in strongly hijacking host functions that respond to autophagy modulation, such as antimicrobial production, possibly to help maintaining intestinal Salmonella populations and ensure spreading of the pathogen. However, as harbouring the evolutionarily newer sopE suggests a gained function in these strains compared to having sopE2, it is crucial to do follow up experiments confirming or contradicting the predictions.
Our experimental strategy to physically separate bystanders from epithelial cells that contain Salmonella revealed clear differences between these two categories of host cells as well as between bystanders and uninfected cells. Indeed, bystanders seem to participate in the host response to Salmonella infection as already shown for other pathogens such as Shigella flexneri, where bystanders were responding to infection in an effector-independent manner (Lippmann et al., 2015). Cross-talk between infected and uninfected neighbouring cells has previously been described for many bacterial or viral pathogens, for example through cytokines signalling (Milivojevic et al., 2017; Bost et al., 2020), or directly through uptake of pathogen effector proteins from infected to uninfected cells (Guidi et al., 2013). S. Typhimurium was also shown to cause a reprogramming of microRNAs in infected cells, affecting also bystander cells (Aguilar et al., 2021). Endoplasmic reticulum stress response is activated in bystander cells, affecting many genes’ expression; in particular downregulating key transcription factors, such as E2F1. E2F1 normally activates autophagy and was shown to work in synergy with the SP1 transcription factor (Lin et al., 1996) propose that SP1 is one target of the Salmonella-derived SopE GEF-mimicking protein, affecting autophagy regulation, possibly in combination with E2F1 down-regulation, although to a lesser degree in bystanders cells. This is in agreement with SopE acting locally within infected cells without affecting the surrounding cells, possibly as a hiding mechanism from host innate defense. It is also possible that bystanders would follow the same pattern at a later time point if there is a delay in their response to what is happening in Salmonella-containing cells.
The HT-29 cell line we used is a cancer-derived cell line, and like many lines it carries several mutations that affect the normal functioning of the cells. However, along with other cell lines used to study Salmonella-host cell interactions, HT-29 cells carry no mutation in core-autophagy genes (unpublished data). Cancer cell lines present the advantage to study the pathogen’s behaviour in a very homogenous host cell population, yet they only partially mirror what is happening in native tissue. With the growing evidence of how useful intestinal organoid-based models are to study host-microbe interactions, including Salmonella-host interactions, using them for validation of network-based predictions of how pathogens’ effectors interfere with host cellular pathways is the obvious next step to pursue.
Overall, our study emphasises the power of network analysis approach in identifying potential interactions between pathogen effector proteins and host cellular machinery regulation. We had previously demonstrated that protein-protein interactions between secreted pathogen effectors and core autophagy proteins is a conserved strategy for many intracellular pathogens (including Salmonella) to modulate the host autophagic clearance mechanisms (Sudhakar et al., 2019). Here, using a similar approach, we showed that Salmonella targets also the regulation of those core autophagy genes. Our experimental validation emphasises the possible role of SopE effector protein as such a local regulator of autophagy flux. Future complementary work investigating the mechanism behind SopE/SP1 or SopE/TP53 interactions will add to our understanding of the complexity and fine tuning of how host-pathogens crosstalk has developed.
The list of Salmonella effectors, relevant host transcription factors and their first neighbours used in the study have is available as supplementary data. The scripts used in the study are available at: https://github.com/korcsmarosgroup/HMIpipeline.
AD has conducted the majority of the computational and experimental work. She contributed to the data analysis as well as the writing and editing of the manuscript. A-CJ has performed all microscopy work, image analysis for this study and the writing of the relevant methods section. LG has contributed to the computational work of this study. IM, JL and AL have contributed to the Flow cytometry work, RNA extraction from low input samples and the writing of the corresponding method section. PB, RI and RK have generated the initial Salmonella sopE gene deletion strain used in this study, and to the writing of the manuscript. IN has contributed to the writing and editing of the manuscript. TK contributed to the conception of the study and the writing and editing of this manuscript. Finally, IH contributed to the conception of the study, to some experimental work, to the data analysis. She led the writing and editing of the manuscript. All authors contributed to the article and approved the submitted version.
We are truly grateful to Jost Enninga (Department of Cell Biology & Infection, Pasteur Institute, Paris, France) for his instrumental advice and discussion. The work of AD, RI, TK and IH were supported by the UKRI BBSRC Gut Microbes and Health Institute Strategic Programme BB/R012490/1 and its constituent projects BBS/E/F/000PR10353 and BBS/E/F/000PR10355 as well as a BBSRC Core Strategic Programme Grant for Genomes to Food Security (BB/CSP1720/1) and its constituent work packages, BBS/E/T/000PR9819 and BBS/E/T/000PR9817. LG was supported by a BBSRC - Norwich Research Park Biosciences Doctoral Training Partnership grant (BB/M011216/1). RK was supported by research grants BB/N007964/1 and BB/M025489/1, and by the BBSRC Institute Strategic Programme Microbes in the Food Chain BB/R012504/1 and its constituent projects BBS/E/F/000PR10348 and BBS/E/F/000PR10349. ACJ and IPN were supported by BBSRC grants BB/L006324/1 and BB/P007856/1.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2022.834895/full#supplementary-material
Aguilar, C., Costa, S., Maudet, C., Vivek-Ananth, R. P., Zaldívar-López, S., Garrido, J. J., et al. (2021). Reprogramming of microRNA expression via E2F1 downregulation promotes salmonella infection both in infected and bystander cells. Nat. Commun. 12, 3392. doi: 10.1038/s41467-021-23593-z
Alpuche Aranda, C. M., Swanson, J. A., Loomis, W. P., Miller, S. I. (1992). Salmonella typhimurium activates virulence gene transcription within acidified macrophage phagosomes. Proc. Natl. Acad. Sci. U.S.A. 89, 10079–10083. doi: 10.1073/pnas.89.21.10079
Andrighetti, T., Bohar, B., Lemke, N., Sudhakar, P., Korcsmaros, T. (2020). MicrobioLink: An Integrated Computational Pipeline to Infer Functional Effects of Microbiome-Host Interactions. Cells. 9, 1278–93. doi: 10.3390/cells9051278
Bakshi, C. S., Singh, V. P., Wood, M. W., Jones, P. W., Wallis, T. S., Galyov, E. E. (2000). Identification of SopE2, a salmonella secreted protein which is highly homologous to SopE and involved in bacterial invasion of epithelial cells. J. Bacteriol. 182, 2341–2344. doi: 10.1128/JB.182.8.2341-2344.2000
Baxt, L. A., Garza-Mayers, A. C., Goldberg, M. B. (2013). Bacterial subversion of host innate immune pathways. Science 340, 697–701. doi: 10.1126/science.1235771
Baxt, L. A., Xavier, R. J. (2015). Role of autophagy in the maintenance of intestinal homeostasis. Gastroenterology 149, 553–562. doi: 10.1053/j.gastro.2015.06.046
Betin, V. M. S., Lane, J. D. (2009). Caspase cleavage of Atg4D stimulates GABARAP-L1 processing and triggers mitochondrial targeting and apoptosis. J. Cell Sci. 122, 2554–2566. doi: 10.1242/jcs.046250
Beuzón, C. R., Banks, G., Deiwick, J., Hensel, M., Holden, D. W. (1999). pH-dependent secretion of SseB, a product of the SPI-2 type III secretion system of salmonella typhimurium. Mol. Microbiol. 33, 806–816. doi: 10.1046/j.1365-2958.1999.01527.x
Beuzón, C. R., Salcedo, S. P., Holden, D. W. (2002). Growth and killing of a salmonella enterica serovar typhimurium sifA mutant strain in the cytosol of different host cell lines. Microbiol. (Reading Engl) 148, 2705–2715. doi: 10.1099/00221287-148-9-2705
Bost, P., Giladi, A., Liu, Y., Bendjelal, Y., Xu, G., David, E., et al. (2020). Host-viral infection maps reveal signatures of severe COVID-19 patients. Cell 181, 1475–1488.e12. doi: 10.1016/j.cell.2020.05.006
Bovolenta, L. A., Acencio, M. L., Lemke, N. (2012). HTRIdb: an open-access database for experimentally verified human transcriptional regulation interactions. BMC Genomics 13, 405. doi: 10.1186/1471-2164-13-405
Brumell, J. H., Tang, P., Zaharik, M. L., Finlay, B. B. (2002). Disruption of the salmonella-containing vacuole leads to increased replication of salmonella enterica serovar typhimurium in the cytosol of epithelial cells. Infect. Immun. 70, 3264–3270. doi: 10.1128/IAI.70.6.3264-3270.2002
Castanheira, S., García-Del Portillo, F. (2017). Salmonella populations inside host cells. Front. Cell. Infect. Microbiol. 7. doi: 10.3389/fcimb.2017.00432
Chan, W., Costantino, N., Li, R., Lee, S. C., Su, Q., Melvin, D., et al. (2007). A recombineering based approach for high-throughput conditional knockout targeting vector construction. Nucleic Acids Res. 35, e64. doi: 10.1093/nar/gkm163
Cicchini, M., Karantza, V., Xia, B. (2015). Molecular pathways: autophagy in cancer–a matter of timing and context. Clin. Cancer Res. 21, 498–504. doi: 10.1158/1078-0432.CCR-13-2438
Colgan, A. M., Kröger, C., Diard, M., Hardt, W.-D., Puente, J. L., Sivasankaran, S. K., et al. (2016). The impact of 18 ancestral and horizontally-acquired regulatory proteins upon the transcriptome and sRNA landscape of salmonella enterica serovar typhimurium. PloS Genet. 12, e1006258. doi: 10.1371/journal.pgen.1006258
Datsenko, K. A., Wanner, B. L. (2000). One-step inactivation of chromosomal genes in escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A. 97, 6640–6645. doi: 10.1073/pnas.120163297
Drecktrah, D., Knodler, L. A., Galbraith, K., Steele-Mortimer, O. (2005). The salmonella SPI1 effector SopB stimulates nitric oxide production long after invasion. Cell. Microbiol. 7, 105–113. doi: 10.1111/j.1462-5822.2004.00436.x
Ehrbar, K., Hardt, W.-D. (2005). Bacteriophage-encoded type III effectors in salmonella enterica subspecies 1 serovar typhimurium. Infect. Genet. Evol. 5, 1–9. doi: 10.1016/j.meegid.2004.07.004
Feng, Y., He, D., Yao, Z., Klionsky, D. J. (2014). The machinery of macroautophagy. Cell Res. 24, 24–41. doi: 10.1038/cr.2013.168
Figueira, R., Watson, K. G., Holden, D. W., Helaine, S. (2013). Identification of salmonella pathogenicity island-2 type III secretion system effectors involved in intramacrophage replication of s. enterica serovar typhimurium: implications for rational vaccine design. MBio 4, e00065. doi: 10.1128/mBio.00065-13
Gemski, P., Stocker, B. A. (1967). Transduction by bacteriophage P22 in nonsmooth mutants of salmonella typhimurium. J. Bacteriol. 93, 1588–1597. doi: 10.1128/jb.93.5.1588-1597.1967
Giacomodonato, M. N., Uzzau, S., Bacciu, D., Caccuri, R., Sarnacki, S. H., Rubino, S., et al. (2007). SipA, SopA, SopB, SopD and SopE2 effector proteins of salmonella enterica serovar typhimurium are synthesized at late stages of infection in mice. Microbiol. (Reading Engl) 153, 1221–1228. doi: 10.1099/mic.0.2006/002758-0
Guidi, R., Levi, L., Rouf, S. F., Puiac, S., Rhen, M., Frisan, T. (2013). Salmonella enterica delivers its genotoxin through outer membrane vesicles secreted from infected cells. Cell. Microbiol. 15, 2034–2050. doi: 10.1111/cmi.12172
Hansen-Wester, I., Hensel, M. (2001). Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 3, 549–559. doi: 10.1016/s1286-4579(01)01411-3
Han, H., Shim, H., Shin, D., Shim, J. E., Ko, Y., Shin, J., et al. (2015). TRRUST: a reference database of human transcriptional regulatory interactions. Sci. Rep. 5, 11432. doi: 10.1038/srep11432
Haraga, A., Ohlson, M. B., Miller, S. I. (2008). Salmonellae interplay with host cells. Nat. Rev. Microbiol. 6, 53–66. doi: 10.1038/nrmicro1788
Hautefort, I., Proença, M. J., Hinton, J. C. D. (2003). Single-copy green fluorescent protein gene fusions allow accurate measurement of salmonella gene expression in vitro and during infection of mammalian cells. Appl. Environ. Microbiol. 69, 7480–7491. doi: 10.1128/aem.69.12.7480-7491.2003
Herhaus, L., Dikic, I. (2018). Regulation of salmonella-host cell interactions via the ubiquitin system. Int. J. Med. Microbiol. 308, 176–184. doi: 10.1016/j.ijmm.2017.11.003
Hoiseth, S. K., Stocker, B. A. (1981). Aromatic-dependent salmonella typhimurium are non-virulent and effective as live vaccines. Nature 291, 238–239. doi: 10.1038/291238a0
Jennings, E., Thurston, T. L. M., Holden, D. W. (2017). Salmonella SPI-2 type III secretion system effectors: Molecular mechanisms and physiological consequences. Cell Host Microbe 22, 217–231. doi: 10.1016/j.chom.2017.07.009
Kaminskyy, V., Zhivotovsky, B. (2012). Proteases in autophagy. Biochim. Biophys. Acta 1824, 44–50. doi: 10.1016/j.bbapap.2011.05.013
Khaminets, A., Behl, C., Dikic, I. (2016). Ubiquitin-dependent and independent signals in selective autophagy. Trends Cell Biol. 26, 6–16. doi: 10.1016/j.tcb.2015.08.010
Kingsley, R. A., Reissbrodt, R., Rabsch, W., Ketley, J. M., Tsolis, R. M., Everest, P., et al. (1999). Ferrioxamine-mediated Iron(III) utilization by salmonella enterica. appl. environ. Microbiol. 65, 1610–1618. doi: 10.1128/AEM.65.4.1610-1618.1999
Kreibich, S., Emmenlauer, M., Fredlund, J., Rämö, P., Münz, C., Dehio, C., et al. (2015). Autophagy proteins promote repair of endosomal membranes damaged by the salmonella type three secretion system 1. Cell Host Microbe 18, 527–537. doi: 10.1016/j.chom.2015.10.015
Krishnadev, O., Srinivasan, N. (2011). Prediction of protein-protein interactions between human host and a pathogen and its application to three pathogenic bacteria. Int. J. Biol. Macromol 48, 613–619. doi: 10.1016/j.ijbiomac.2011.01.030
Kroemer, G., Bravo-San Pedro, J. M., Galluzzi, L. (2015). Novel function of cytoplasmic p53 at the interface between mitochondria and the endoplasmic reticulum. Cell Death Dis. 6, e1698. doi: 10.1038/cddis.2015.70
Kröger, C., Colgan, A., Srikumar, S., Händler, K., Sivasankaran, S. K., Hammarlöf, D. L., et al. (2013). An infection-relevant transcriptomic compendium for salmonella enterica serovar typhimurium. Cell Host Microbe 14, 683–695. doi: 10.1016/j.chom.2013.11.010
Kshirsagar, M., Carbonell, J., Klein-Seetharaman, J. (2012). Techniques to cope with missing data in host-pathogen protein interaction prediction. Bioinformatics 28, i466–i472. doi: 10.1093/bioinformatics/bts375
Kubisch, J., Türei, D., Földvári-Nagy, L., Dunai, Z. A., Zsákai, L., Varga, M., et al. (2013). ). complex regulation of autophagy in cancer - integrated approaches to discover the networks that hold a double-edged sword. Semin. Cancer Biol. 23, 252–261. doi: 10.1016/j.semcancer.2013.06.009
Kubori, T., Galán, J. E. (2003). Temporal regulation of salmonella virulence effector function by proteasome-dependent protein degradation. Cell 115, 333–342. doi: 10.1016/s0092-8674(03)00849-3
Lin, S. Y., Black, A. R., Kostic, D., Pajovic, S., Hoover, C. N., Azizkhan, J. C. (1996). Cell cycle-regulated association of E2F1 and SP1 is related to their functional interaction. Mol. Cell. Biol. 16, 1668–1675. doi: 10.1128/MCB.16.4.1668
Lippmann, J., Gwinner, F., Rey, C., Tamir, U., Law, H. K. W., Schwikowski, B., et al. (2015). Bacterial internalization, localization, and effectors shape the epithelial immune response during shigella flexneri infection. infect. Immun. 83, 3624–3637. doi: 10.1128/IAI.00574-15
Liss, V., Hensel, M. (2015). Take the tube: remodelling of the endosomal system by intracellular salmonella enterica. Cell. Microbiol. 17, 639–647. doi: 10.1111/cmi.12441
Lou, L., Zhang, P., Piao, R., Wang, Y. (2019). Salmonella pathogenicity island 1 (SPI-1) and its complex regulatory network. Front. Cell. Infect. Microbiol. 9. doi: 10.3389/fcimb.2019.00270
Lucas, R. L., Lee, C. A. (2000). Unravelling the mysteries of virulence gene regulation in salmonella typhimurium. Mol. Microbiol. 36, 1024–1033. doi: 10.1046/j.1365-2958.2000.01961.x
Marcus, S. L., Brumell, J. H., Pfeifer, C. G., Finlay, B. B. (2000). Salmonella pathogenicity islands: big virulence in small packages. Microbes Infect. 2, 145–156. doi: 10.1016/s1286-4579(00)00273-2
Martin-Orozco, N., Touret, N., Zaharik, M. L., Park, E., Kopelman, R., Miller, S., et al. (2006). Visualization of vacuolar acidification-induced transcription of genes of pathogens inside macrophages. Mol. Biol. Cell 17, 498–510. doi: 10.1091/mbc.e04-12-1096
Miao, E. A., Miller, S. I. (2000). A conserved amino acid sequence directing intracellular type III secretion by salmonella typhimurium. Proc. Natl. Acad. Sci. U.S.A. 97, 7539–7544. doi: 10.1073/pnas.97.13.7539
Milivojevic, M., Dangeard, A.-S., Kasper, C. A., Tschon, T., Emmenlauer, M., Pique, C., et al. (2017). ALPK1 controls TIFA/TRAF6-dependent innate immunity against heptose-1,7-bisphosphate of gram-negative bacteria. PloS Pathog. 13, e1006224. doi: 10.1371/journal.ppat.1006224
Mirold, S., Ehrbar, K., Weissmüller, A., Prager, R., Tschäpe, H., Rüssmann, H., et al. (2001). Salmonella host cell invasion emerged by acquisition of a mosaic of separate genetic elements, including salmonella pathogenicity island 1 (SPI1), SPI5, and sopE2. J. Bacteriol. 183, 2348–2358. doi: 10.1128/JB.183.7.2348-2358.2001
Mirold, S., Rabsch, W., Rohde, M., Stender, S., Tschäpe, H., Rüssmann, H., et al. (1999). Isolation of a temperate bacteriophage encoding the type III effector protein SopE from an epidemic salmonella typhimurium strain. Proc. Natl. Acad. Sci. U.S.A. 96, 9845–9850. doi: 10.1073/pnas.96.17.9845
Nieto, P. A., Pardo-Roa, C., Salazar-Echegarai, F. J., Tobar, H. E., Coronado-Arrázola, I., Riedel, C. A., et al. (2016). New insights about excisable pathogenicity islands in salmonella and their contribution to virulence. Microbes Infect. 18, 302–309. doi: 10.1016/j.micinf.2016.02.001
Picelli, S., Björklund, Å.K., Faridani, O. R., Sagasser, S., Winberg, G., Sandberg, R. (2013). Smart-seq2 for sensitive full-length transcriptome profiling in single cells. Nat. Methods 10, 1096–1098. doi: 10.1038/nmeth.2639
Powers, T. R., Haeberle, A. L., Predeus, A. V., Hammarlöf, D. L., Cundiff, J. A., Saldaña-Ahuactzi, Z., et al. (2021). Intracellular niche-specific profiling reveals transcriptional adaptations required for the cytosolic lifestyle of salmonella enterica. PloS Pathog. 17, e1009280. doi: 10.1371/journal.ppat.1009280
Quan, W., Lee, M.-S. (2013). Role of autophagy in the control of body metabolism. Endocrinol. Metab. (Seoul) 28, 6–11. doi: 10.3803/EnM.2013.28.1.6
Rao, X., Huang, X., Zhou, Z., Lin, X. (2013). An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat Bioinforma Biomath 3, 71–85.
Sambrook, J., Russell, D.W. (2001). Molecular Cloning: A Laboratory Manual. 3rd Edition, Vol. 1 (New York: Cold Spring Harbor Laboratory Press).
Schlumberger, M. C., Hardt, W. D. (2005). Triggered phagocytosis by salmonella: bacterial molecular mimicry of RhoGTPase activation/deactivation. Curr. Top. Microbiol. Immunol. 291, 29–42. doi: 10.1007/3-540-27511-8_3
Shaid, S., Brandts, C. H., Serve, H., Dikic, I. (2013). Ubiquitination and selective autophagy. Cell Death Differ. 20, 21–30. doi: 10.1038/cdd.2012.72
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Sorbara, M. T., Foerster, E. G., Tsalikis, J., Abdel-Nour, M., Mangiapane, J., Sirluck-Schroeder, I., et al. (2018). Complement C3 drives autophagy-dependent restriction of cyto-invasive bacteria. Cell Host Microbe 23, 644–652.e5. doi: 10.1016/j.chom.2018.04.008
Sorbara, M. T., Girardin, S. E. (2015). Emerging themes in bacterial autophagy. Curr. Opin. Microbiol. 23, 163–170. doi: 10.1016/j.mib.2014.11.020
Srikanth, C. V., Mercado-Lubo, R., Hallstrom, K., McCormick, B. A. (2011). Salmonella effector proteins and host-cell responses. Cell. Mol. Life Sci. 68, 3687–3697. doi: 10.1007/s00018-011-0841-0
Srikumar, S., Kröger, C., Hébrard, M., Colgan, A., Owen, S. V., Sivasankaran, S. K., et al. (2015). RNA-Seq brings new insights to the intra-macrophage transcriptome of salmonella typhimurium. PloS Pathog. 11, e1005262. doi: 10.1371/journal.ppat.1005262
Stévenin, V., Chang, Y.-Y., Le Toquin, Y., Duchateau, M., Gianetto, Q. G., Luk, C. H., et al. (2019). Dynamic growth and shrinkage of the salmonella-containing vacuole determines the intracellular pathogen niche. Cell Rep. 29, 3958–3973.e7. doi: 10.1016/j.celrep.2019.11.049
Sudhakar, P., Jacomin, A.-C., Hautefort, I., Samavedam, S., Fatemian, K., Ari, E., et al. (2019). Targeted interplay between bacterial pathogens and host autophagy. Autophagy 15, 1620–1633. doi: 10.1080/15548627.2019.1590519
Tasdemir, E., Maiuri, M. C., Galluzzi, L., Vitale, I., Djavaheri-Mergny, M., D’Amelio, M., et al. (2008). Regulation of autophagy by cytoplasmic p53. Nat. Cell Biol. 10, 676–687. doi: 10.1038/ncb1730
Thornbrough, J. M., Worley, M. J. (2012). A naturally occurring single nucleotide polymorphism in the salmonella SPI-2 type III effector srfH/sseI controls early extraintestinal dissemination. PloS One 7, e45245. doi: 10.1371/journal.pone.0045245
Türei, D., Földvári-Nagy, L., Fazekas, D., Módos, D., Kubisch, J., Kadlecsik, T., et al. (2015). Autophagy regulatory network - a systems-level bioinformatics resource for studying the mechanism and regulation of autophagy. Autophagy 11, 155–165. doi: 10.4161/15548627.2014.994346
Vonaesch, P., Sellin, M. E., Cardini, S., Singh, V., Barthel, M., Hardt, W.-D. (2014). The salmonella typhimurium effector protein SopE transiently localizes to the early SCV and contributes to intracellular replication. Cell. Microbiol. 16, 1723–1735. doi: 10.1111/cmi.12333
Weigele, B. A., Alto, N. M. (2010). Salmonella taking charge. Cell Host Microbe 7, 421–422. doi: 10.1016/j.chom.2010.06.001
Xu, X.-W., Pan, C.-W., Yang, X.-M., Zhou, L., Zheng, Z.-Q., Li, D.-C. (2018). SP1 reduces autophagic flux through activating p62 in gastric cancer cells. Mol. Med. Rep. 17, 4633–4638. doi: 10.3892/mmr.2018.8400
Yellaboina, S., Tasneem, A., Zaykin, D. V., Raghavachari, B., Jothi, R. (2011). DOMINE: a comprehensive collection of known and predicted domain-domain interactions. Nucleic Acids Res. 39, D730–D735. doi: 10.1093/nar/gkq1229
Yu, H. B., Croxen, M. A., Marchiando, A. M., Ferreira, R. B. R., Cadwell, K., Foster, L. J., et al. (2014). Autophagy facilitates salmonella replication in HeLa cells. MBio 5, e00865–e00814. doi: 10.1128/mBio.00865-14
Keywords: Salmonella Typhimurium, autophagy, SopE, network biology, MAP1LC3B, Host-microbe interactions
Citation: Demeter A, Jacomin A-C, Gul L, Lister A, Lipscombe J, Invernizzi R, Branchu P, Macaulay I, Nezis IP, Kingsley RA, Korcsmaros T and Hautefort I (2022) Computational prediction and experimental validation of Salmonella Typhimurium SopE-mediated fine-tuning of autophagy in intestinal epithelial cells. Front. Cell. Infect. Microbiol. 12:834895. doi: 10.3389/fcimb.2022.834895
Received: 13 December 2021; Accepted: 28 July 2022;
Published: 17 August 2022.
Edited by:
Hua Niu, Affiliated Hospital of Guilin Medical University, ChinaReviewed by:
Mindy Engevik, Medical University of South Carolina, United StatesCopyright © 2022 Demeter, Jacomin, Gul, Lister, Lipscombe, Invernizzi, Branchu, Macaulay, Nezis, Kingsley, Korcsmaros and Hautefort. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tamas Korcsmaros, VGFtYXMuS29yY3NtYXJvc0BlYXJsaGFtLmFjLnVr
†Present address: Anne-Claire Jacomin, Institute of Biochemistry II, Faculty of Medicine, Goethe University, Frankfurt am Main, Germany
Rachele Invernizzi,Broad Institute of MIT & Harvard, Merkin Building, Cambridge, MA, United States, Institut de Recherche en Santé Digestive (IRSD) INSERM U1220, INRA UMR1416, ENVT, UT3 Microbiology and the Food Chain Division (MICA) Bat B, CHU Purpan, Toulouse, France
Priscilla Branchu, French National Institute for Agricultural and Environmental Research, France
‡ORCID: Tamas Korcsmaros, orcid.org/0000-0003-1717-996X
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.