 Albert S. Lee
Albert S. Lee Olivia K. Lamanna2†
Olivia K. Lamanna2† Kenji Ishida
Kenji Ishida Andrew Nguyen
Andrew Nguyen Michael H. Hsieh
Michael H. Hsieh
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol., 01 February 2022
Sec. Microbiome in Health and Disease
Volume 12 - 2022 | https://doi.org/10.3389/fcimb.2022.794323
This article is part of the Research TopicThe Urogenital Microbiota in Urinary Tract DiseasesView all 10 articles
Background: Polymerase chain reaction (PCR) is an important means by which to study the urine microbiome and is emerging as possible alternative to urine cultures to identify pathogens that cause urinary tract infection (UTI). However, PCR is limited by its inability to differentiate DNA originating from viable, metabolically active versus non-viable, inactive bacteria. This drawback has led to concerns that urobiome studies and PCR-based diagnosis of UTI are confounded by the presence of relic DNA from non-viable bacteria in urine. Propidium monoazide (PMA) dye can penetrate cells with compromised cell membranes and covalently bind to DNA, rendering it inaccessible to amplification by PCR. Although PMA has been shown to differentiate between non-viable and viable bacteria in various settings, its effectiveness in urine has not been previously studied. We sought to investigate the ability of PMA to differentiate between viable and non-viable bacteria in urine.
Methods: Varying amounts of viable or non-viable uropathogenic E. coli (UTI89) or buffer control were titrated with mouse urine. The samples were centrifuged to collect urine sediment or not centrifuged. Urine samples were incubated with PMA and DNA cross-linked using blue LED light. DNA was isolated and uidA gene-specific PCR was performed. For in vivo studies, mice were inoculated with UTI89, followed by ciprofloxacin treatment or no treatment. After the completion of ciprofloxacin treatment, an aliquot of urine was plated on non-selective LB agar and another aliquot was treated with PMA and subjected to uidA-specific PCR.
Results: PMA’s efficiency in excluding DNA signal from non-viable bacteria was significantly higher in bacterial samples in phosphate-buffered saline (PBS, dCT=13.69) versus bacterial samples in unspun urine (dCT=1.58). This discrepancy was diminished by spinning down urine-based bacterial samples to collect sediment and resuspending it in PBS prior to PMA treatment. In 3 of 5 replicate groups of UTI89-infected mice, no bacteria grew in culture; however, there was PCR amplification of E. coli after PMA treatment in 2 of those 3 groups.
Conclusion: We have successfully developed PMA-based PCR methods for amplifying DNA from live bacteria in urine. Our results suggest that non-PMA bound DNA from live bacteria can be present in urine, even after antibiotic treatment. This indicates that viable but non-culturable E. coli can be present following treatment of UTI, and may explain why some patients have persistent symptoms but negative urine cultures following UTI treatment.
The existence of the urinary microbiome, the presence of bacterial communities within the urinary tract, is challenging the paradigm that this organ system is normally sterile (Siddiqui et al., 2012; Wolfe et al., 2012; Hilt et al., 2014; Brubaker and Wolfe, 2015; Thomas-White et al., 2016). Furthermore, several studies have shown an association between the urine microbiome and numerous urological diseases (Fouts et al., 2012; Siddiqui et al., 2012; Whiteside et al., 2015; Bajic et al., 2018; Bučević Popović et al., 2018; Magistro and Stief, 2019; Neugent et al., 2020). Therefore, it is imperative to accurately characterize the urinary microbiome as it may inform overall urinary tract health and aid in the diagnosis of urinary conditions, i.e., urinary tract infection (UTI) (Perez-Carrasco et al., 2021).
Numerous clinical studies of patients with UTI feature assessments of both microbiologic and clinical cure, which are based on negative urine cultures and resolution/improvement of symptoms, respectively (Raz et al., 2002; Wunderink et al., 2018; Miller et al., 2019). Some patients in these studies have featured discordance between microbiologic and clinical cure (Hilt et al., 2014; Price et al., 2018; Swamy et al., 2019). One possible interpretation of this discordance is that conventional urine cultures may be missing residual bacteria causing persistent symptoms following antibiotic therapy. An alternative to urine cultures for detection of urinary microorganisms is polymerase chain reaction (PCR). PCR identifies organisms through the amplification of DNA material present in urine and many studies on the urinary microbiome rely on this molecular approach (Lewis et al., 2013; Brubaker and Wolfe, 2015; Ackerman et al., 2019). These methods do not discriminate between relic DNA (DNA from non-viable bacteria) versus DNA from viable bacteria. This is an important limitation of conventional PCR because the confounding effects of relic DNA have been reported in various microbiologic settings (Carini et al., 2016; Nagler et al., 2021; Ren et al., 2021). Viable, metabolically active bacteria presumably exert much more influence over the clinical course of UTI than dead bacteria. Thus, the amplification of total DNA without selection for DNA from viable bacteria may bias conventional PCR-derived results (Carini et al., 2016; Lennon et al., 2018). Given that relic DNA influences conventional molecular measurements of microbial abundance and diversity, we posit that a method to detect viable, metabolically active bacteria is needed for more accurate urobiome studies (Lennon et al., 2018).
A method of identifying metabolically active bacteria via PCR has been recently developed. Propidium monoazide (PMA) dye penetrates cells with compromised cell membranes (non-viable cells) and covalently binds to DNA, rendering it unable to be amplified by PCR (Deshmukh et al., 2020). PMA-based PCR has previously been shown to differentiate between non-viable and viable bacteria in many settings (Fittipaldi et al., 2010; Cattani et al., 2016; Gobert et al., 2018; Brauge et al., 2019; Lu et al., 2019). However, the efficiency of PMA in urine has not been previously investigated. Here we investigated the ability of PMA dye in urine to detect DNA derived from viable bacteria.
All bacteria work was performed under sterile conditions in a BSL-2 biosafety cabinet. Bacteria were prepared by previously reported methods (Hung, 2009). Briefly, glycerol stock containing the uropathogenic Escherichia coli strain UTI89 was used to inoculate a Miller Luria Broth (LB) agar plate (Sigma-Aldrich, St. Louis, MO). The plate was incubated for 24 hours at 37°C. A single colony was picked and transferred to 10 mL LB broth. The culture was incubated overnight at 37°C in a stationary flask. Twenty-five microliters of the 10 mL culture were transferred to 25mL of LB broth. The culture was incubated again overnight at 37°C in a stationary flask. The culture was centrifuged at 5000 x g for 5 minutes at 4°C. The supernatant was decanted and the bacteria pellet was resuspended in 10 mL of sterile phosphate buffered saline (PBS) (Thermofisher Scientific, Waltham, MA). This suspension was diluted tenfold in sterile PBS. The optical density (OD) 600nm value was analyzed using the NanoDrop-1000 (Thermofisher Scientific, Waltham, MA) and the suspension diluted until the OD was 0.50, corresponding to 1-2x107 colony-forming units (CFU) per 50 μL. Experiments involving Klebsiella pneumoniae (ATCC reference strain 13883) followed the same procedure described above for UTI89.
Mice were scruffed and held with their pelvises above sterile parafilm (Sigma-Aldrich, St. Louis, MO) until they voided. Urine was aspirated from the parafilm. New parafilm was used for each mouse. Urine was then placed on ice and immediately processed.
Patient urine was collected according to George Washington University IRB protocol # NCR213442
Five hundred μL of E. coli with OD value 0.5 was mixed with isopropanol (Sigma-Aldrich, St. Louis, MO) to achieve a final concentration of 70% v/v. After 10 minutes, the mixture was centrifuged at 8000 x g for 10 min. The supernatant was removed and the pellet was resuspended in 100 μL of PBS. The suspensions of non-viable bacteria were plated on LB with agar plates and incubated at 37°C overnight to confirm successful killing.
Mouse urine was serially diluted to a ratio of 1:2, 1:4, 1:8, and 1:12 with PBS. Subsequently, 50 μL of all viable or all non-viable bacteria was added to 50 μL of the various titrations of urine, or undiluted urine, with resultant ratio of 1:2, 1:4, 1: 8, 1:16, and 1:24. The samples were then either treated with PMA or left untreated.
Under minimal light, PMAxx Dye (hereafter referred to as PMA) (Biotium, Fremont, CA) with a concentration of 20 mM was diluted with nuclease-free water (Sigma-Aldrich, St. Louis, MO) to a final concentration of 10 mM. It was then added to the bacterial mixture in a 1:100 ratio. Next, samples were incubated for 15 minutes in the dark with gentle agitation. The samples were placed in an LED lightbox with LED output wavelength of 465-475 nm (Biotium, Fremont, CA) for 20 minutes to induce PMA crosslinking of DNA. The supernatant was removed and the pellet was reconstituted with PBS to its original volume of 100 μL.
DNA was isolated using the DNeasy PowerSoil Pro Kit (Qiagen, Germantown, MD) according to kit instructions, except that DNA was eluted from the column with 25 μL of nuclease-free water. PCR was performed targeting the E. coli uidA gene with TaqMan polymerase (Invitrogen, Waltham, MA) according to previously described methods (Taskin et al., 2011). Delta CT (dCT) values were calculated to quantify the differences between CT values of PMA-treated and untreated samples. For the DNA extraction in the comparison between non-DNA-extracted samples and DNA-extracted samples, 50 µL of cells (0.5-1x106 cells) were subjected to DNA extraction, ending in an elution of 50 µL of C6 elution buffer. The PCR quantification was performed in a 25 µL volume in technical triplicates using 2.5 µL of cells (2.5-5x105 cells) or 2.5 µL of extracted DNA (which represents the DNA material from 2.5-5x105 cells) for each reaction.
Urea (Sigma-Aldrich, St. Louis, MO) was diluted with PBS in to two concentrations; 285 mM, corresponding to the urine urea level in humans and 1800 mM, corresponding to the urine urea level in mice (Yang and Bankir, 2005). Fifty μL of 100% viable or 100% non-viable bacteria, corresponding to 1-2x107 colony-forming units (CFU) of E. Coli was added to 50 μL of the urea solutions. The DNA of the PMA treated and untreated samples was extracted and amplified as described above.
Defined quantities of viable bacteria, isopropanol-killed non-viable bacteria, or PBS were mixed to a total volume of 100 μL. With a fixed amount of viable bacteria (1-2x107 CFU), non-viable bacteria were added to achieve 1:10 (1x 106 non-viable), 1:100 (1x 105 non-viable) and 1:1000 (1x 104 non-viable) non-viable to viable bacterial dilutions. In a similar manner, various amounts of viable bacteria (1x 106,1 x 105, 1x 104) were added to a fixed amount of non-viable bacteria (1-2x107 CFU) to generate a standard curve. Undiluted viable and non-viable cultures were also used. Fifty μL of these bacterial solutions were added to urine. The mixture was then centrifuged at 5000xg, resuspended with 100 μL of sterile PBS, treated with PMA, and the DNA was extracted as outlined above.
All animal work was approved by The Institutional Animal Care and Use Committee of Children’s National Hospital under Animal Use Protocol #00030764. Procedures were performed in an ethical fashion. Prior to use, all animals were acclimated for 7 days after arrival to the animal facility. 24-week-old female C3H/HeOuJ mice (stock no: 000635, The Jackson Laboratory, Bar Harbor, ME) were used in this study.
Mice were anesthetized using 2% isoflurane. Any urine in the bladder was expressed by gently pressing on the lower abdomen. A 24g x ¾ inch angiocatheter (Clint Pharmaceuticals, Old Hickory, TN) was attached to a prepared 1 ml syringe containing the inoculant. The angiocath was lubricated (DynaLub Sterile Lubricating Jelly, Amazon, Seattle, WA) and transurethrally inserted into the bladder. 100 μL of the inoculant was instilled slowly into the bladder and the angiocatheter kept inserted for 30 seconds to prevent leakage of the inoculant.
Five days after inoculation, mice were intraperitoneally injected with 10 mg/kg ciprofloxacin twice a day. This regimen was selected as it recapitulates the human plasma peak levels achieved with the commonly used 500 mg oral dose, and has been shown previously to adequately treat UTI in mice (Guillard et al., 2013).
One day after completion of ciprofloxacin treatment, mouse urine was collected on ice. Individual urine samples in the same treatment groups (3-4 mice/group) were pooled. The urine was either serially diluted and plated in triplicate on LB agar or prepared for PMA treatment. Fifty μL of PBS was added to the urine and the solution was centrifuged and treated with PMA as outlined above. DNA was extracted and the E. coli uidA gene was amplified as described above (Taskin et al., 2011).
Initial PMA-based PCR experiments using mouse urine spiked with 1-2x107 CFU of isopropanol killed UPEC UTI89 yielded little differences in amplification of PMA-treated vs. untreated DNA samples (Figure 1A). This led us to consider the possibility that urine was exerting a matrix effect which interferes with downstream molecular processes such as PMA crosslinking (Taylor et al., 2012). The dCT value of PMA-treated vs. untreated samples that contained 100% non-viable bacteria resuspended in mouse urine was 1.58, which was about one tenth of the dCT of the same sample resuspended in PBS (13.69). When urine was diluted with PBS, the dCTs of PMA-treated vs. untreated samples increased, indicating improved PMA efficiency (Figure 1B). However, the increase in dCT plateaued at a dilution of 1:8. These findings indicate that urine inhibits PMA activity.
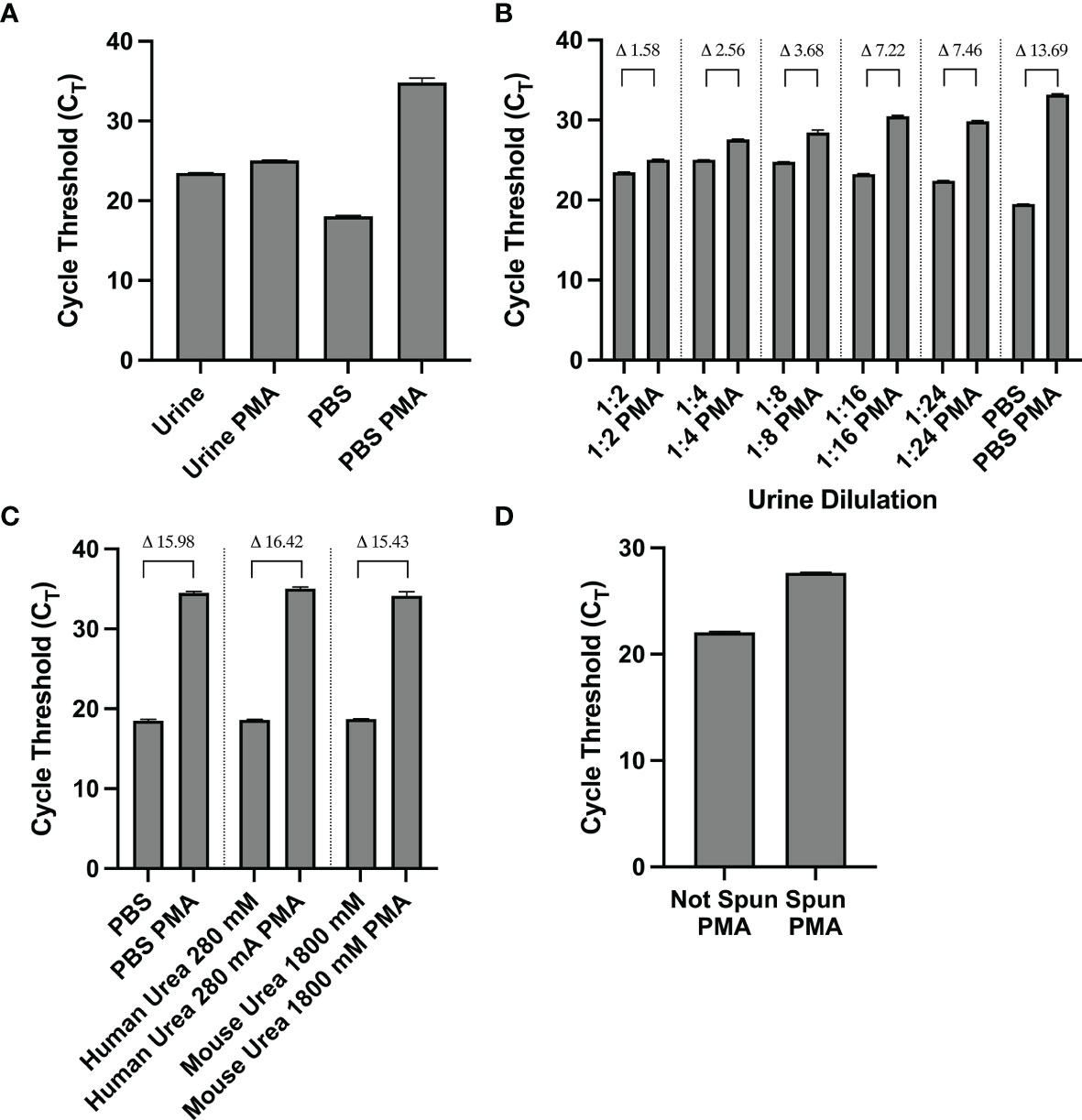
Figure 1 (A–D) The matrix effect of urine on PMA crosslinking of DNA is not urea-based and is eliminated by centrifugation and resuspension in PBS. (A) CT values of PMA-treated and untreated samples of non-viable bacteria resuspended in mouse urine or PBS. (B) CT (and indicated dCT) values of nonviable bacteria resuspended in various dilutions of urine or PBS. (C) CT (and indicated dCT) values of non-viable bacteria treated with PMA in PBS or urea concentrations corresponding to human and mouse urine. (D) CT values of non-viable bacteria in urine with or without resuspending the contents in PBS before PMA treatment. Data shown is representative of three replicate experiments.
Urea is the most abundant solute present in urine and is known to influence molecular structure and function (Yang and Bankir, 2005; Wei et al., 2010). Thus, we sought to investigate urea’s potential effect on PMA’s function in crosslinking DNA and subsequently inhibiting its amplification. We analyzed PMA’s efficiency at two different urea concentrations: 280 mM and 1800 mM, the approximate concentration of urea in human and mouse urine, respectively. The dCTs of PMA-treated vs. untreated samples with 100% nonviable bacteria (1-2x107 CFU of E Coli) suspended in either urea concentration was similar to that of the PBS control. Namely, the dCT of the PBS, human urea concentration, and mouse urea concentration samples were 15.98, 16.42, and 15.43, respectively (Figure 1C). This suggests that urea does not cause urine’s matrix effect on PMA efficiency.
We observed that for a solution with 100% nonviable bacteria, the CT value increases when the urine supernatant was removed and the pellet is resuspended in PBS prior to PMA crosslinking compared to when PMA crosslinking is performed in unspun urine (Figure 1D). Furthermore, upon removal of the urine supernatant and subsequent pellet resuspension in PBS, PMA treatment and downstream PCR was most efficient in differentiating viability when there was a greater proportion of nonviable cells in the solution. Across titrations of viable and non-viable bacteria where viable bacteria make up the majority of the solution, CT values did not significantly differ, with all values close to 16 (Figure 2A). This suggests the amount of non-viable bacteria does not influence the detection of a fixed amount of viable bacteria. Conversely, when the majority of the cells are non-viable, the CT values decrease as the amount of viable bacteria in the solution increases. For instance, the CT is approximately 18 with 100% viable cells (1-2x107 CFU of E Coli) in solution and increases to ~ 27 when viable cells make up only 0.1% of the solution (Figure 2B).
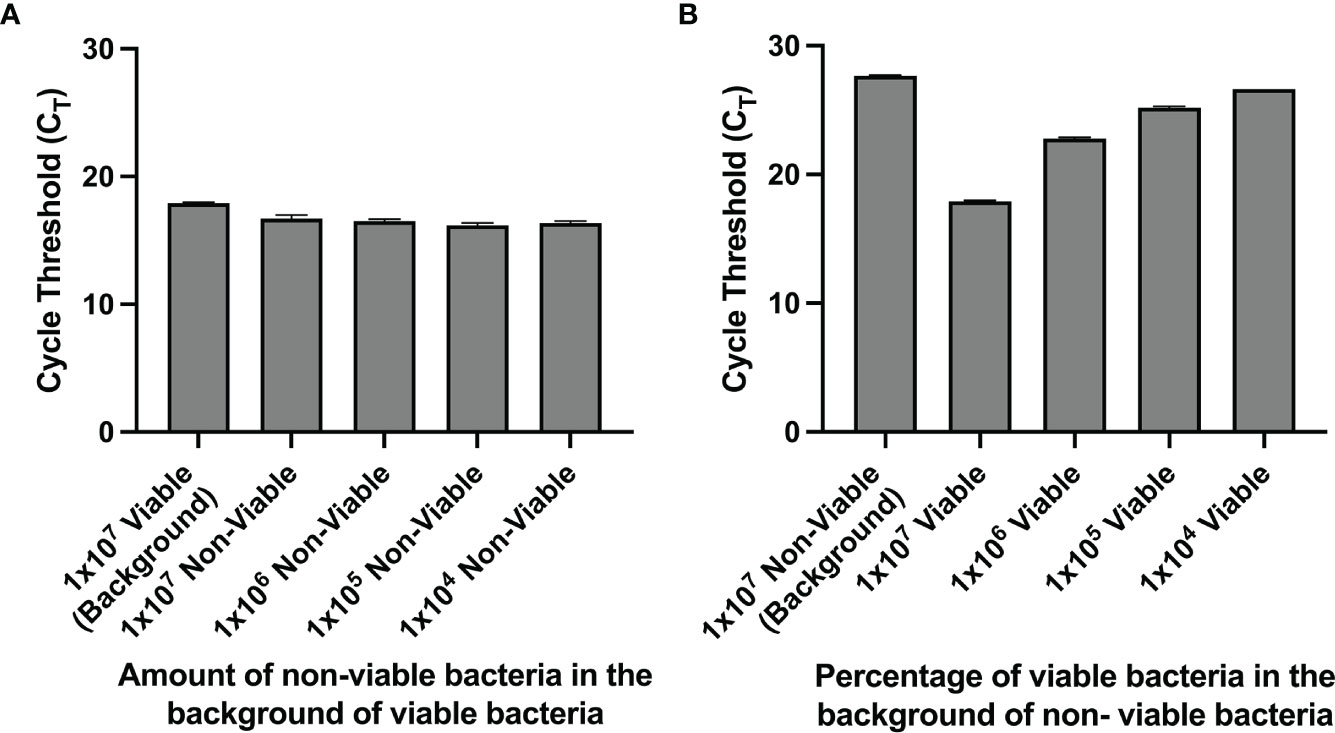
Figure 2 (A, B) PMA crosslinking of DNA is most efficient when most of the bacteria is non-viable. Bacterial samples were centrifuged and resuspended in PBS before PMA treatment. (A) CT values of samples with a fixed amount of viable bacteria and titrated amounts of nonviable bacteria. (B) CT values of samples with a fixed amount of non-viable bacteria and titrated amounts of viable bacteria.
Given that the number of colony forming units present in urine cultures remains the mainstay of clinical diagnosis of UTI, we sought to determine whether urine cultures with various titrations of viable cells and a fixed amount of nonviable cells yielded cfu and CT values that correlated with each other. Indeed, the correlation between cfu and CT values was strongly negative with an r2 of 0.955 (Figure 3).
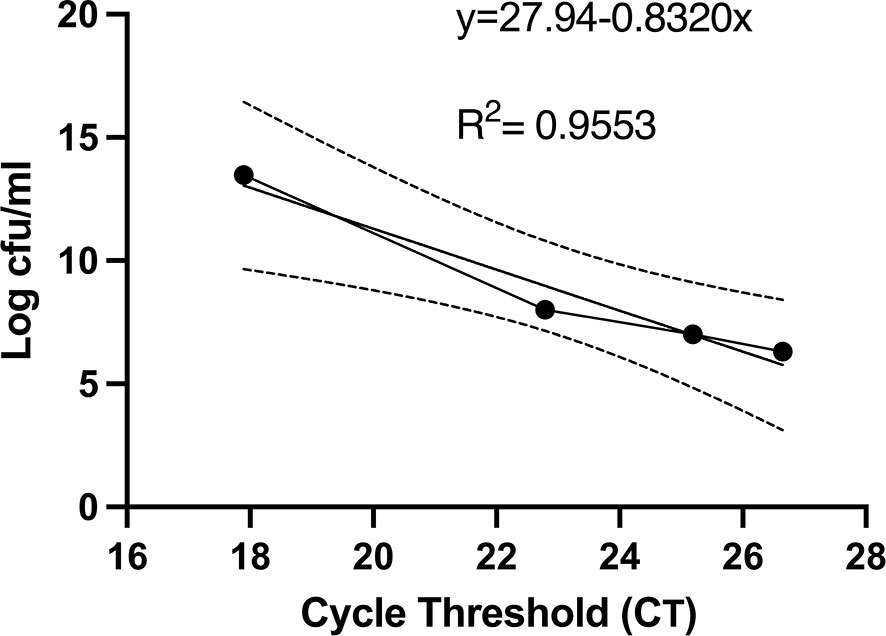
Figure 3 Colony forming units and PMA-based viability PCR cycle threshold number has a strong negative correlation. Before PMA treatment, an aliquot of bacterial samples was serial diluted, plated on LB agar plates, and cfu were counted after 24 hours. cfu per ml were plotted against the respective CT values. The linear regression and 95% confidence interval band is shown. All experiments were repeated 3 times.
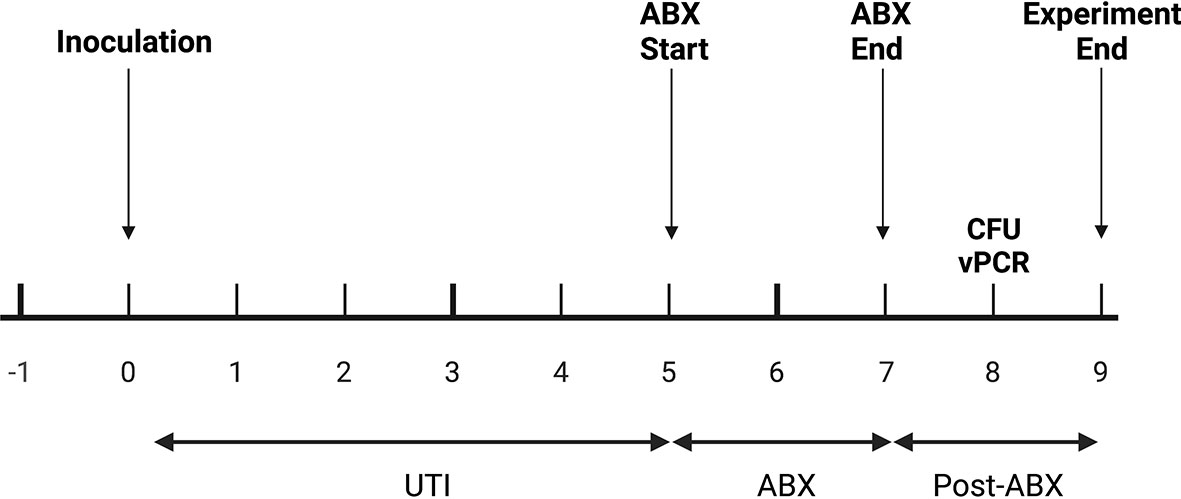
Figure 4 In vivo experimental plan. Mice were inoculated with UTI89 via transurethral catheterization on day 0 and given ciprofloxacin (“ABX”) twice daily on days 5, 6, and 7 post-infection. On day 8, one day after the completion of antibiotics, urine was collected to measure cfu/mL and to perform PMA-based viability PCR (“vPCR”).
PCR-based detection of bacterial DNA in urine from patients with persistent UTI symptoms and negative cultures following antibiotic therapy has been criticized as being confounded by the presence of relic DNA (Lehmann et al., 2011). To investigate whether non-culturable, viable bacteria can still be present in urine after antibiotic treatment of UTI, we administered uropathogenic E. coli (UTI89) to mice and treated them with ciprofloxacin according to established protocols (Hung, 2009). One day after the completion of antibiotic treatment, 3 out of the 5 replicate groups had no bacterial growth on non-selective LB agar (Figure 5). However, after PMA treatment of these samples, PCR successfully amplified the E. coli uidA gene in 2 out of the 3 culture-negative groups, indicating the presence of viable, nonculturable bacteria. Based on our standard curve (Figure 3), these 2 groups contained 1 and 6×105 cfu/ml E. coli.
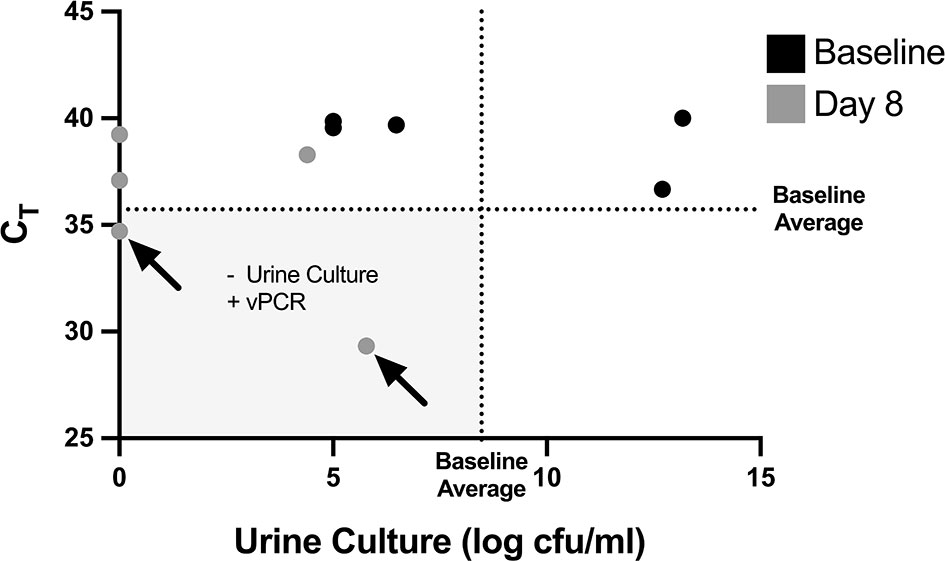
Figure 5 Non-culturable, live bacteria detected in mouse urine by PMA-based PCR after antibiotic treatment. Mice were given UTIs on day 0 and were administered ciprofloxacin starting on day 5 and ending after day 7 (see Figure 4). Graph depicts CT values and log transformed urine cfu/mL values before bacterial inoculation, labeled as baseline (black dots), and at day 8 after antibiotic treatment (grey dots). Each dot represents a pooled cohort of 2-3 mice. The average CT and log cfu/ml of the baseline samples are indicated by the dotted lines. The greyed quadrant (Q3) represents values that are considered as having a negative urine culture and positive PMA-based viability PCR (“vPCR”). Data shown is pooled from two set of experiments.
To further investigate the relevance and feasibility of PMA-based urine PCR for human urine, with considerations for polymicrobial infections and the possibility to decrease the required time and resources for the assay, we spiked culture-negative human urine with different combinations of viable and non-viable E. coli and viable Klebsiella pneumoniae, followed by PMA treatment and PCR, with or without DNA extraction. We found that, as we observed with mouse urine, in the presence of non-soluble components of human urine collected with the bacterial pellet during centrifugation, PMA treatment could exclude signal from non-viable E. coli with dCT values of at least 8.66 (Figure 6; samples 1-2, 6-7, 10-11). Similarly, we observed that the presence of K. pneumoniae, another commonly found uropathogen, did not strongly affect the signal from E. coli, for both PMA-treated and non-PMA-treated samples (Figure 6; samples 6-9). Interestingly, DNA-extracted samples showed higher CT values compared to their respective non-extracted samples, indicating loss of some DNA material during the DNA extraction process; however, this loss of material did not strongly affect PMA function (Figure 6; samples 1-4, 6-13).
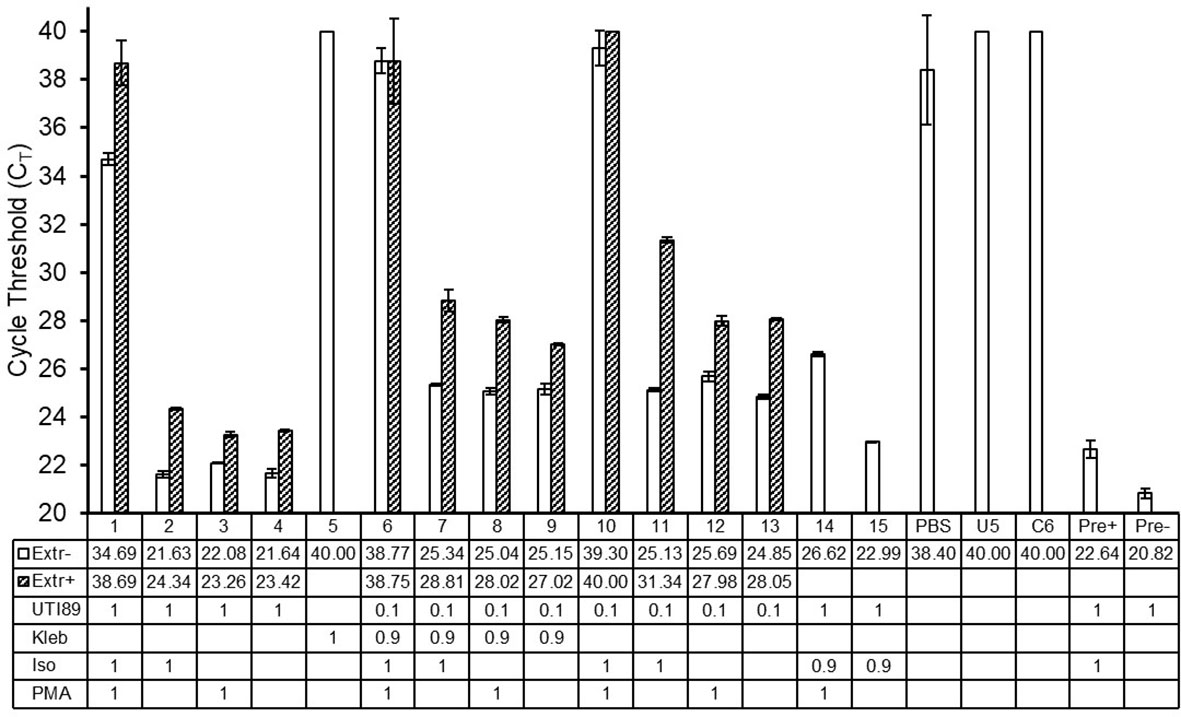
Figure 6 PMA-PCR can distinguish viable E. coli from non-viable E. coli in the presence of non-soluble components of human urine, presence of Klebsiella pneumoniae, and with or without DNA extraction. Bars represent the arithmetic average of CT values +/- standard deviation of technical triplicate reactions. Open bars represent samples that did not undergo DNA extraction, while hatched bars represent samples that did undergo DNA extraction. Sample numbers/names and CT values for non-DNA-extracted and DNA-extracted samples are shown in the table below the graph. PCR samples that did not reach the signal threshold before 40 cycles of PCR, i.e., samples whose CT could not be determined, were assigned a CT value of 40. The numbers in the rows labeled “UTI89” and “Kleb” indicate the respective number of bacteria represented in the PCR of each sample, with a value of 1 equating to 2.5-5x105 cells. The numbers in the rows labeled “Iso” and “PMA” indicate whether a given sample received the respective treatment, where a value of 1 represents treatment; the value of 0.9 in the “Iso” treatment for samples 14 and 15 indicates that this proportion of cells were treated. In samples 6 and 7, only the UTI89 cells were treated with isopropanol. UTI89, UTI89 strain of E. coli; Kleb, Klebsiella pneumoniae; Iso, isopropanol; PMA, propidium monoazide; PBS, phosphate buffered saline; U5, culture-negative human urine; C6, DNA extraction elution buffer; Pre+ and Pre-, pre-PMA samples.
Our findings indicate that a PMA-based urine PCR is an appropriate method to distinguish viable and non-viable E. coli in the urine for both in vitro and in vivo applications. We were able to eliminate the signal from soluble relic DNA and DNA from nonviable cells. By resuspending urine contents in PBS before PMA treatment we established an easy and reproducible method to eliminate soluble relic DNA while preserving E. coli cells. This approach yielded dCT values similar to that of non-urine exposed E. coli resuspended in PBS and those reported in the literature (Taskin et al., 2011). Thus, we demonstrate a novel method to utilize PMA in urine that will allow for PCR-based studies to selectively identify viable bacteria in urine.
Our preliminary experiments pointed to a matrix effect of murine urine that inhibited PMA crosslinking of DNA (Chamberlain et al., 2019). We initially focused on urea as a potential cause of this effect. Urea is a by-product of amino acid metabolism and one of the most abundant urine solutes. Mouse urine has a higher urea concentration compared to human urine, which led us to speculate that urea could be influencing PMA crosslinking function in mouse urine (Yang and Bankir, 2005). The similar dCT of E. coli in urea vs. PBS suggested urea is not the substance in urine that inhibits PMA’s crosslinking efficiency. Thus, the inhibitory effect of urine is likely due to a non-urea-related matrix effect. The simple steps of centrifuging bacteria-containing samples and resuspending them in buffer may eliminate any matrix effect of other biofluids and environmental samples of interest, enabling PMA-based PCR amplification of DNA from viable bacteria in other settings. While potentially a small number of bacteria can be lost in the supernatant, the standardized centrifuging force and pellet resuspension process used across the various samples should yield similar number of bacteria lost and thus still allow us to perform comparative studies across the samples.
Identifying viable, but potentially unculturable bacteria may improve understanding of bacterial biology in patients with recurrent UTI. We identified non-culturable but viable UTI E. coli in the urine of infected mice given ciprofloxacin. Non-culturable but viable bacteria in settings other than the urinary tract are a recognized phenomenon (Coutard et al., 2005; Oliver, 2005). However, the presence of such bacteria in urine is poorly characterized. It may be that these bacteria represent intracellular UTI E. coli which have formed intracellular communities and become quiescent reservoirs of infection (Mulvey et al., 2001; Rosen et al., 2007). Our findings may explain why, despite patients having undergone susceptibility-guided antibiotic treatment and a subsequent negative test-of-cure by urine culture, some of these patients experience recurrent UTI.
Compared to urine culture, PMA-based urine PCR has the clinical advantages of a more rapid time to UTI diagnosis and broader organism detection. While enhanced quantitative urine culture has demonstrated greater sensitivity for uropathogen detection than conventional culture (Price et al., 2016), it is still time-intensive. In contrast, a uropathogen-specific PCR platform based on PMA could detect multiple viable organisms quickly.
This is a preliminary study using a new molecular method for identification of bacteria in urine. A potential limitation to our study is the use of PMA dye prior to PCR. Specifically, studies have shown that PMA results can be skewed by specific primers. However, the primers used in this study have been shown to be effective in multiple studies for E. coli without loss of viability data (Taskin et al., 2011; van Frankenhuyzen et al., 2013).
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by Children’s National and the Children’s National Research Institute- Institutional Animal Care and Use Committee. The human urine study protocol was approved by George Washington University Institutional Review Board.
AL, OL, and MH contributed to conception and design of the study. AL and OL organized the database. OL performed the statistical analysis. AL and OL wrote the first draft of the manuscript. KI, EH, AN, and MH wrote sections of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
This work was supported by NIH-R01DK113504 (MH) and George Washington University Cancer Biology Training Program NIH-T32CA247756 (KI).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
CFU, Colony forming units; CT, Threshold Cycle; dCT, Delta Threshold Cycle; E. Coli, Escherichia coli; LB, Luria-Bertani; OD, Optical Density; PBS, Phosphate-buffered saline; PMA, Propidium Monoazide; RT, PCR- Real-time polymerase chain reaction; UTI, Urinary Tract Infection.
Ackerman, A. L., Anger, J. T., Khalique, M. U., Ackerman, J. E., Tang, J., Kim, J., et al. (2019). Optimization of DNA Extraction From Human Urinary Samples for Mycobiome Community Profiling. PLoS One 14 (4), e0210306. doi: 10.1371/journal.pone.0210306
Bajic, P., Van Kuiken, M. E., Burge, B. K., Kirshenbaum, E. J., Joyce, C. J., Wolfe, A. J., et al. (2018). Male Bladder Microbiome Relates to Lower Urinary Tract Symptoms. Eur. Urol. Focus. 6 (2), 376–382. doi: 10.1016/j.euf.2018.08.001
Brauge, T., Midelet-Bourdin, G., Soumet, C. (2019). Viability Detection of Foodborne Bacterial Pathogens in Food Environment by PMA-qPCR and by Microscopic Observation. Methods Mol. Biol. 1918, 117–128. doi: 10.1007/978-1-4939-9000-9_9
Brubaker, L., Wolfe, A. J. (2015). The New World of the Urinary Microbiota in Women. Am. J. Obstet. Gynecol. 213, 644–649. doi: 10.1016/j.ajog.2015.05.032
Bučević Popović, V., Šitum, M., Chow, C.-E. T., Chan, L. S., Roje, B., Terzić, J. (2018). The Urinary Microbiome Associated With Bladder Cancer. Sci. Rep. 8, 12157. doi: 10.1038/s41598-018-29054-w
Carini, P., Marsden, P. J., Leff, J. W., Morgan, E. E., Strickland, M. S., Fierer, N. (2016). Relic DNA Is Abundant in Soil and Obscures Estimates of Soil Microbial Diversity. Nat. Microbiol. 2, 16242. doi: 10.1038/nmicrobiol.2016.242
Cattani, F., Barth, V. C. J., Nasário, J. S. R., Ferreira, C. A. S., Oliveira, S. D. (2016). Detection and Quantification of Viable Bacillus Cereus Group Species in Milk by Propidium Monoazide Quantitative Real-Time PCR. J. Dairy Sci. 99, 2617–2624. doi: 10.3168/jds.2015-10019
Chamberlain, C. A., Rubio, V. Y., Garrett, T. J. (2019). Impact of Matrix Effects and Ionization Efficiency in Non-Quantitative Untargeted Metabolomics. Metabolomics 15, 135. doi: 10.1007/s11306-019-1597-z
Coutard, F., Pommepuy, M., Loaec, S., Hervio-Heath, D. (2005). mRNA Detection by Reverse Transcription-PCR for Monitoring Viability and Potential Virulence in a Pathogenic Strain of Vibrio Parahaemolyticus in Viable But Nonculturable State. J. Appl. Microbiol. 98, 951–961. doi: 10.1111/j.1365-2672.2005.02534.x
Deshmukh, R., Bhand, S., Roy, U. (2020). A Novel Method for Rapid and Sensitive Detection of Viable Escherichia Coli Cells Using UV-Induced PMA-Coupled Quantitative PCR. Braz. J. Microbiol. 51, 773–778. doi: 10.1007/s42770-019-00161-8
Fittipaldi, M., Rodriguez, N. J. P., Codony, F., Adrados, B., Peñuela, G. A., Morató, J. (2010). Discrimination of Infectious Bacteriophage T4 Virus by Propidium Monoazide Real-Time PCR. J. Virol. Methods 168, 228–232. doi: 10.1016/j.jviromet.2010.06.011
Fouts, D. E., Pieper, R., Szpakowski, S., Pohl, H., Knoblach, S., Suh, M., et al. (2012). Integrated Next-Generation Sequencing of 16S rDNA and Metaproteomics Differentiate the Healthy Urine Microbiome From Asymptomatic Bacteriuria in Neuropathic Bladder Associated With Spinal Cord Injury. J. Transl. Med. 10, 1. doi: 10.1186/1479-5876-10-174
Gobert, G., Cotillard, A., Fourmestraux, C., Pruvost, L., Miguet, J., Boyer, M. (2018). Droplet Digital PCR Improves Absolute Quantification of Viable Lactic Acid Bacteria in Faecal Samples. J. Microbiol. Methods 148, 64–73. doi: 10.1016/j.mimet.2018.03.004
Guillard, T., Cambau, E., Chau, F., Massias, L., de Champs, C., Fantin, B. (2013). Ciprofloxacin Treatment Failure in a Murine Model of Pyelonephritis Due to an AAC(6’)-Ib-Cr-Producing Escherichia Coli Strain Susceptible to Ciprofloxacin In Vitro. Antimicrob. Agents Chemother. 57, 5830–5835. doi: 10.1128/AAC.01489-13
Hilt, E. E., McKinley, K., Pearce, M. M., Rosenfeld, A. B., Zilliox, M. J., Mueller, E. R., et al. (2014). Urine Is Not Sterile: Use of Enhanced Urine Culture Techniques to Detect Resident Bacterial Flora in the Adult Female Bladder. J. Clin. Microbiol. 52, 871–876. doi: 10.1128/JCM.02876-13
Hung, C. (2009). A Murine Model of Urinary Tract Infection. Nat. Protoc. 4, 1230–1243. doi: 10.1038/nprot.2009.116.A
Lehmann, L. E., Hauser, S., Malinka, T., Klaschik, S., Weber, S. U., Schewe, J.-C., et al. (2011). Rapid Qualitative Urinary Tract Infection Pathogen Identification by SeptiFast Real-Time PCR. PLoS One 6, e17146. doi: 10.1371/journal.pone.0017146
Lennon, J. T., Muscarella, M. E., Placella, S. A., Lehmkuhl, B. K. (2018). How, When, and Where Relic DNA Affects Microbial Diversity. MBio 9 (3), e00637-18. doi: 10.1128/mBio.00637-18
Lewis, D. A., Brown, R., Williams, J., White, P., Jacobson, S. K., Marchesi, J. R., et al. (2013). The Human Urinary Microbiome; Bacterial DNA in Voided Urine of Asymptomatic Adults. Front. Cell. Infect. Microbiol. 3, 41. doi: 10.3389/fcimb.2013.00041
Lu, J., Zheng, H., Chu, P., Han, S., Yang, H., Wang, Z., et al. (2019). Direct Detection From Clinical Sputum Samples to Differentiate Live and Dead Mycobacterium Tuberculosis. J. Clin. Lab. Anal. 33, e22716. doi: 10.1002/jcla.22716
Magistro, G., Stief, C. G. (2019). The Urinary Tract Microbiome: The Answer to All Our Open Questions? Eur. Urol. Focus 5, 36–38. doi: 10.1016/j.euf.2018.06.011
Miller, L. G., Friedland, I., Dwyer, J. P. (2019). Once-Daily Plazomicin for Complicated Urinary Tract Infections. N. Engl. J. Med. 380 (8), 729–740. doi: 10.1056/NEJMoa1801467
Mulvey, M. A., Schilling, J. D., Hultgren, S. J. (2001). Establishment of a Persistent Escherichia Coli Reservoir During the Acute Phase of a Bladder Infection. Infect. Immun. 69, 4572–4579. doi: 10.1128/IAI.69.7.4572-4579.2001
Nagler, M., Podmirseg, S. M., Mayr, M., Ascher-Jenull, J., Insam, H. (2021). The Masking Effect of Extracellular DNA and Robustness of Intracellular DNA in Anaerobic Digester NGS Studies: A Discriminatory Study of the Total DNA Pool. Mol. Ecol. 30, 438–450. doi: 10.1111/mec.15740
Neugent, M. L., Hulyalkar, N. V., Nguyen, V. H., Zimmern, P. E., De Nisco, N. J. (2020). Advances in Understanding the Human Urinary Microbiome and Its Potential Role in Urinary Tract Infection. MBio 11, e00218–e00220. doi: 10.1128/mBio.00218-20
Oliver, J. D. (2005). The Viable But Nonculturable State in Bacteria. J. Microbiol. 43 Spec No, 93–100.
Perez-Carrasco, V., Soriano-Lerma, A., Soriano, M., Gutiérrez-Fernández, J., Garcia-Salcedo, J. A. (2021). Urinary Microbiome: Yin and Yang of the Urinary Tract. Front. Cell. Infect. Microbiol. 11, 617002. doi: 10.3389/fcimb.2021.617002
Price, T. K., Dune, T., Hilt, E. E., Thomas-White, K. J., Kliethermes, S., Brincat, C., et al. (2016). The Clinical Urine Culture: Enhanced Techniques Improve Detection of Clinically Relevant Microorganisms. J. Clin. Microbiol. 54, 1216–1222. doi: 10.1128/JCM.00044-16
Price, T. K., Hilt, E. E., Dune, T. J., Mueller, E. R., Wolfe, A. J., Brubaker, L. (2018). Urine Trouble: Should We Think Differently About UTI? Int. Urogynecol. J. 29, 205–210. doi: 10.1007/s00192-017-3528-8
Raz, R., Chazan, B., Kennes, Y., Colodner, R., Rottensterich, E., Dan, M., et al. (2002). Empiric Use of Trimethoprim-Sulfamethoxazole (TMP-SMX) in the Treatment of Women With Uncomplicated Urinary Tract Infections, in a Geographical Area With a High Prevalence of TMP-SMX-Resistant Uropathogens. Clin. Infect. Dis. 34, 1165–1169. doi: 10.1086/339812
Ren, Q., Wei, F., Yuan, C., Zhu, C., Zhang, Q., Quan, J., et al. (2021). The Effects of Removing Dead Bacteria by Propidium Monoazide on the Profile of Salivary Microbiome. BMC Oral. Health 21, 460. doi: 10.1186/s12903-021-01832-5
Rosen, D. A., Hooton, T. M., Stamm, W. E., Humphrey, P. A., Hultgren, S. J. (2007). Detection of Intracellular Bacterial Communities in Human Urinary Tract Infection. PLoS Med. 4, e329. doi: 10.1371/journal.pmed.0040329
Siddiqui, H., Lagesen, K., Nederbragt, A. J., Jeansson, S. L., Jakobsen, K. S. (2012). Alterations of Microbiota in Urine From Women With Interstitial Cystitis. BMC Microbiol. 12, 205. doi: 10.1186/1471-2180-12-205
Swamy, S., Kupelian, A. S., Khasriya, R., Dharmasena, D., Toteva, H., Dehpour, T., et al. (2019). Cross-Over Data Supporting Long-Term Antibiotic Treatment in Patients With Painful Lower Urinary Tract Symptoms, Pyuria and Negative Urinalysis. Int. Urogynecol. J. 30, 409–414. doi: 10.1007/s00192-018-3846-5
Taskin, B., Gozen, A. G., Duran, M. (2011). Selective Quantification of Viable Escherichia Coli Bacteria in Biosolids by Quantitative PCR With Propidium Monoazide Modification. Appl. Environ. Microbiol. 77, 4329–4335. doi: 10.1128/AEM.02895-10
Taylor, T. P., Janech, M. G., Slate, E. H., Lewis, E. C., Arthur, J. M., Oates, J. C. (2012). Overcoming the Effects of Matrix Interference in the Measurement of Urine Protein Analytes. Biomark. Insights 7, 1–8. doi: 10.4137/BMI.S8703
Thomas-White, K., Brady, M., Wolfe, A. J., Mueller, E. R. (2016). The Bladder Is Not Sterile: History and Current Discoveries on the Urinary Microbiome. Curr. Bladder. Dysfunct. Rep. 11, 18–24. doi: 10.1007/s11884-016-0345-8
van Frankenhuyzen, J. K., Trevors, J. T., Flemming, C. A., Lee, H., Habash, M. B. (2013). Optimization, Validation, and Application of a Real-Time PCR Protocol for Quantification of Viable Bacterial Cells in Municipal Sewage Sludge and Biosolids Using Reporter Genes and Escherichia Coli. J. Ind. Microbiol. Biotechnol. 40, 1251–1261. doi: 10.1007/s10295-013-1319-x
Wei, H., Fan, Y., Gao, Y. Q. (2010). Effects of Urea, Tetramethyl Urea, and Trimethylamine N-Oxide on Aqueous Solution Structure and Solvation of Protein Backbones: A Molecular Dynamics Simulation Study. J. Phys. Chem. B. 114, 557–568. doi: 10.1021/jp9084926
Whiteside, S. A., Razvi, H., Dave, S., Reid, G., Burton, J. P. (2015). The Microbiome of the Urinary Tract–A Role Beyond Infection. Nat. Rev. Urol. 12, 81–90. doi: 10.1038/nrurol.2014.361
Wolfe, A. J., Toh, E., Shibata, N., Rong, R., Kenton, K., Fitzgerald, M., et al. (2012). Evidence of Uncultivated Bacteria in the Adult Female Bladder. J. Clin. Microbiol. 50, 1376–1383. doi: 10.1128/JCM.05852-11
Wunderink, R. G., Giamarellos-Bourboulis, E. J., Rahav, G., Mathers, A. J., Bassetti, M., Vazquez, J., et al. (2018). Effect and Safety of Meropenem-Vaborbactam Versus Best-Available Therapy in Patients With Carbapenem-Resistant Enterobacteriaceae Infections: The TANGO II Randomized Clinical Trial. Infect. Dis. Ther. 7, 439–455. doi: 10.1007/s40121-018-0214-1
Keywords: propidium monoazide, viability, urine, microbiome, non-culturable bacteria, urobiome, urinary tract infection, relic DNA
Citation: Lee AS, Lamanna OK, Ishida K, Hill E, Nguyen A and Hsieh MH (2022) A Novel Propidium Monoazide-Based PCR Assay Can Measure Viable Uropathogenic E. coli In Vitro and In Vivo. Front. Cell. Infect. Microbiol. 12:794323. doi: 10.3389/fcimb.2022.794323
Received: 13 October 2021; Accepted: 11 January 2022;
Published: 01 February 2022.
Edited by:
Amanda L. Lewis, University of California, San Diego, United StatesReviewed by:
Gianni Prosseda, Sapienza University of Rome, ItalyCopyright © 2022 Lee, Lamanna, Ishida, Hill, Nguyen and Hsieh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael H. Hsieh, bWhzaWVoQGNoaWxkcmVuc25hdGlvbmFsLm9yZw==
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.