 Peng Qiu
Peng Qiu Takatsugu Ishimoto
Takatsugu Ishimoto Lingfeng Fu2,3
Lingfeng Fu2,3 Jun Zhang
Jun Zhang Yang Liu
Yang Liu- 1Department of Anesthesiology, Shengjing Hospital of China Medical University, Shenyang, China
- 2Department of Gastroenterological Surgery, Graduate School of Medical Sciences, Kumamoto University, Kumamoto, Japan
- 3Gastrointestinal Cancer Biology, International Research Center for Medical Sciences, Kumamoto University, Kumamoto, Japan
- 4Department of Oncology, Shengjing Hospital of China Medical University, Shenyang, China
Epidemiological surveys indicate that the incidence of inflammatory bowel disease (IBD) is increasing rapidly with the continuous growth of the economy. A large number of studies have investigated the relationship between the genetic factors related to the susceptibility to IBD and the gut microbiota of patients by using high-throughput sequencing. IBD is considered the outcome of the interaction between host and microorganisms, including intestinal microbial factors, abnormal immune response, and a damaged intestinal mucosal barrier. The imbalance of microbial homeostasis leads to the colonization and invasion of opportunistic pathogens in the gut, which increases the risk of the host immune response and promotes the development of IBD. It is critical to identify the specific pathogens related to the pathogenesis of IBD. An in-depth understanding of various pathogenic factors is of great significance for the early detection of IBD. This review highlights the role of gut microbiota in the pathogenesis of IBD and provides a theoretical basis for the personalized approaches that modulate the gut microbiota to treat IBD.
Introduction
Inflammatory bowel disease (IBD) consists of 2 subtypes: Crohn’s disease (CD) and ulcerative colitis (UC), which affects 0.3%–0.5% of the global population (Ng et al., 2017). The occurrence and development of IBD, which is a type of idiopathic inflammatory gastrointestinal disease, are influenced by multiple etiologies, including genetic susceptibility, immune factors, and the gut microbiota (Manichanh et al., 2012; Schaubeck et al., 2016). Several studies have confirmed that the composition of gut microbiota in IBD patients is significantly different from that of healthy individuals (Mosca et al., 2016; Oligschlaeger et al., 2019).
The human microbiota comprises 10–100 trillion microorganisms (Mukhopadhya et al., 2015; Lopetuso et al., 2016; Marchesi et al., 2016), including bacteria, viruses, protozoa, and fungi, among which bacteria are the most abundant, with a density of 1011–1012 cells/ml. More than 99% of the bacteria belong to the phyla Firmicutes, Bacteroides, Proteus, and Actinomycetes, whereas Firmicutes and Bacteroides are dominant in the gut flora of the healthy host (Kostic et al., 2014; Lloyd-Price et al., 2019). Over 1,000 species of bacteria in the gastrointestinal tract play a fundamental role in several aspects of host homeostasis: nutrition, immune, metabolism, and defense against pathogens (Wilson and Nicholson, 2017). The gut microbiota can decompose carbohydrates and indigestible oligosaccharides in food, synthesize short-chain fatty acids (SCFAs), such as butyric acid, propionic acid, and acetate, and provide abundant energy for the intestinal epithelium (Ramakrishna, 2013). Beneficial bacteria in the gut microbiota can play an immunosuppressive role by regulating host immune cells (Allen-Vercoe and Coburn, 2020). Some harmful bacteria can also induce inflammatory cytokines by immune cell interactions or their metabolites to promote the intestinal damage (Stappenbeck and Virgin, 2016). Herein, we review the pathogenic interaction of the microbial communities with the intestinal epithelial barrier, metabolome, and immune system of patients with IBD, and discuss the practical strategies used by microbiota-based therapies to treat IBD patients.
Microbiota Composition and IBD
The composition and diversity of the gut microbiota are key factors leading to the development of IBD (Lavelle et al., 2019; Schirmer et al., 2019). The composition of the gut microbiota can change in the early stages of IBD. The fluctuation of gut microbiota composition in IBD patients is greater than that in healthy individuals (Halfvarson et al., 2017). Some studies have found that the degree of dysbiosis in CD patients is greater than that in individuals with UC (Cekin, 2017; Kriss et al., 2018; Yilmaz et al., 2018). Compared with healthy controls, the levels of Bifidobacterium longum in UC, Eubacterium rectale, Faecalibacterium prausnitzii, Roseburia intestinalis, and other beneficial bacteria in CD and UC were significantly reduced, while the relative abundance and growth rate of harmful bacteria such as Bacteroides fragilis are increased (Vich Vila et al., 2018). Ruminococcus torques and Ruminococcus are also enriched in CD and UC at the onset of the disease. The transcriptional activity of a small number of strains increases as well, as shown by the significant differences in the abundance of Clostridium hathewayi, Clostridium bolteae, and Ruminococcus gnavus (Lloyd-Price et al., 2019). The families of Christensenellaceae, Coriobacteriaceae, and, in particular, Clostridium leptum decrease, while Actinomyces spp., Veillonella spp., and also Escherichia coli increase in patients with CD. For patients with UC, there is an enrichment of Eubacterium rectum and Akkermansia muciniphila decreases, while levels of E. coli increase (Pittayanon et al., 2020). A comparative study showed that the abundance of Intestinibacter spp. increases in both CD and UC, while the abundance of Coprococcus spp. significantly decreases in CD (Forbes et al., 2018). Hall et al. found that R. gnavus is significantly more abundant in patients with IBD.
A total of 199 IBD-specific genes have been identified that are involved in adhesion, oxidative stress responses, and utilization of the mucus, which favor the colonization of R. gnavus in IBD (Hall et al., 2017). A. muciniphila was demonstrated to be a pathobiont that promotes the development of IBD and NOD-like receptor 6 (NLRP6) and was identified as a key regulator of the abundance of A. muciniphila (Seregin et al., 2017). IBD-related genes Caspase recruitment domain family member 9 (CARD9), Nucleotide binding oligomerization domain containing 2 (NOD2), Autophagy related 16 like 1(ATG16L), Immunity related GTPase M (IRGM), and Fucosyltransferase 2 (FUT2) are significantly associated with the low abundance in the genus Roseburia (Imhann et al., 2018). The prevalence of Blastocystis spp. was reduced in patients with active IBD compared with healthy individuals (Tito et al., 2019). Weersma et al. used 12 exome-wide microbial quantitative trait loci (mbQTL) analyses, identified variants in several genes, including Myelin gene regulatory factor (MYRF), SEC16 homolog A (SEC16A), Interleukin 17 receptor E-like (IL17REL), and WD repeat domain 78 (WDR78), which were related with IBD. The genetic variants affecting the immune system play a vital role in shaping the microbiota in the etiology of IBD (Hu et al., 2021). There is increasing evidence that the influence of genetic susceptibility and the environment on gut microbiota is also related to the pathogenesis of IBD (Figure 1 and Table 1).
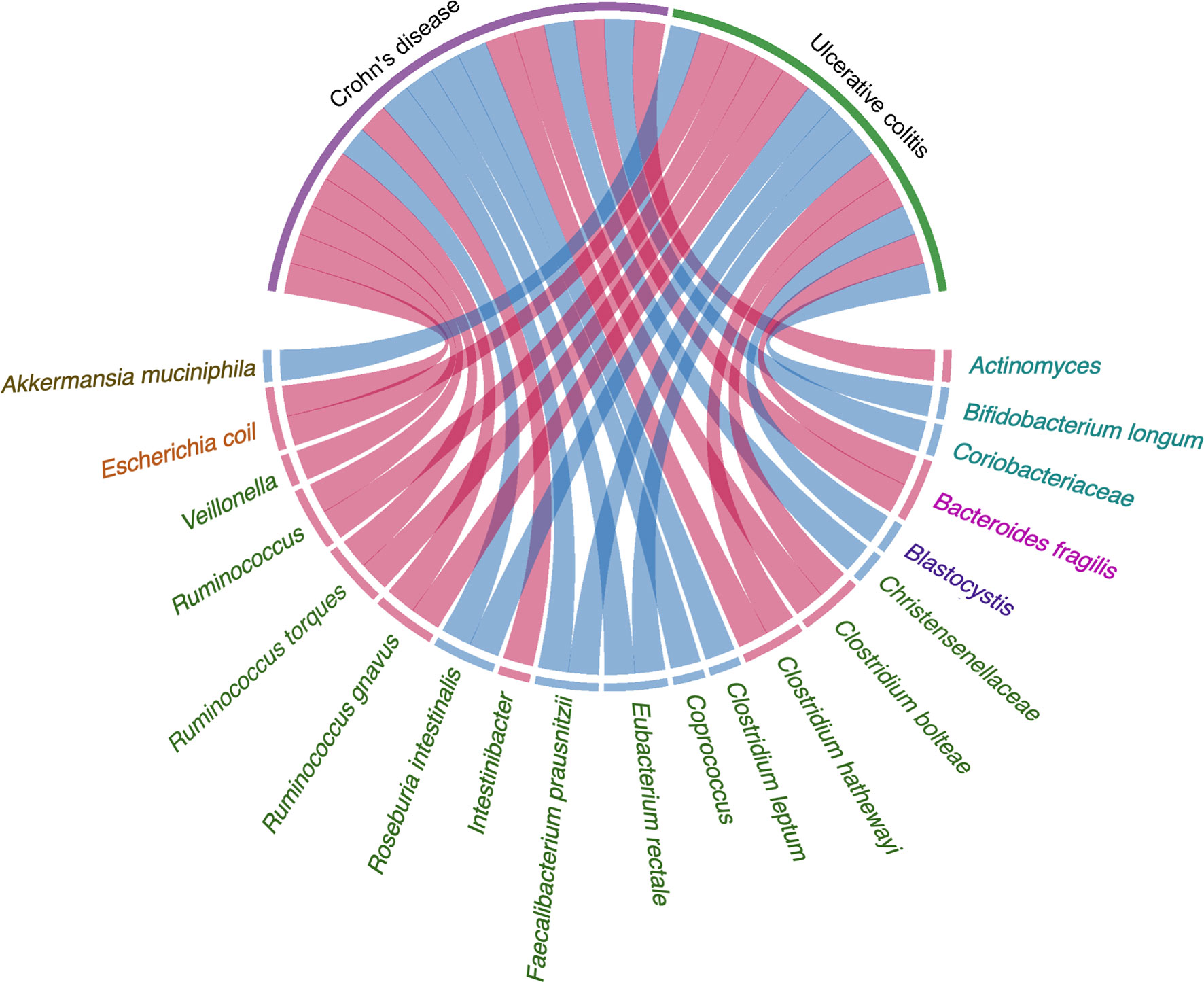
Figure 1 Evidence of the gut microbiota enriched in UC and CD playing a vital role in pathogenesis. Circos plots showing the correlation of bacteria with pathogenesis in IBD. The red ribbons represent the higher production of bacteria enriched in IBD development. The blue ribbons represent the lower production of bacteria enriched in IBD. The causality of the microbiota in IBD has not yet been fully elucidated. Different taxa are divided into six groups and colored by the phylum.
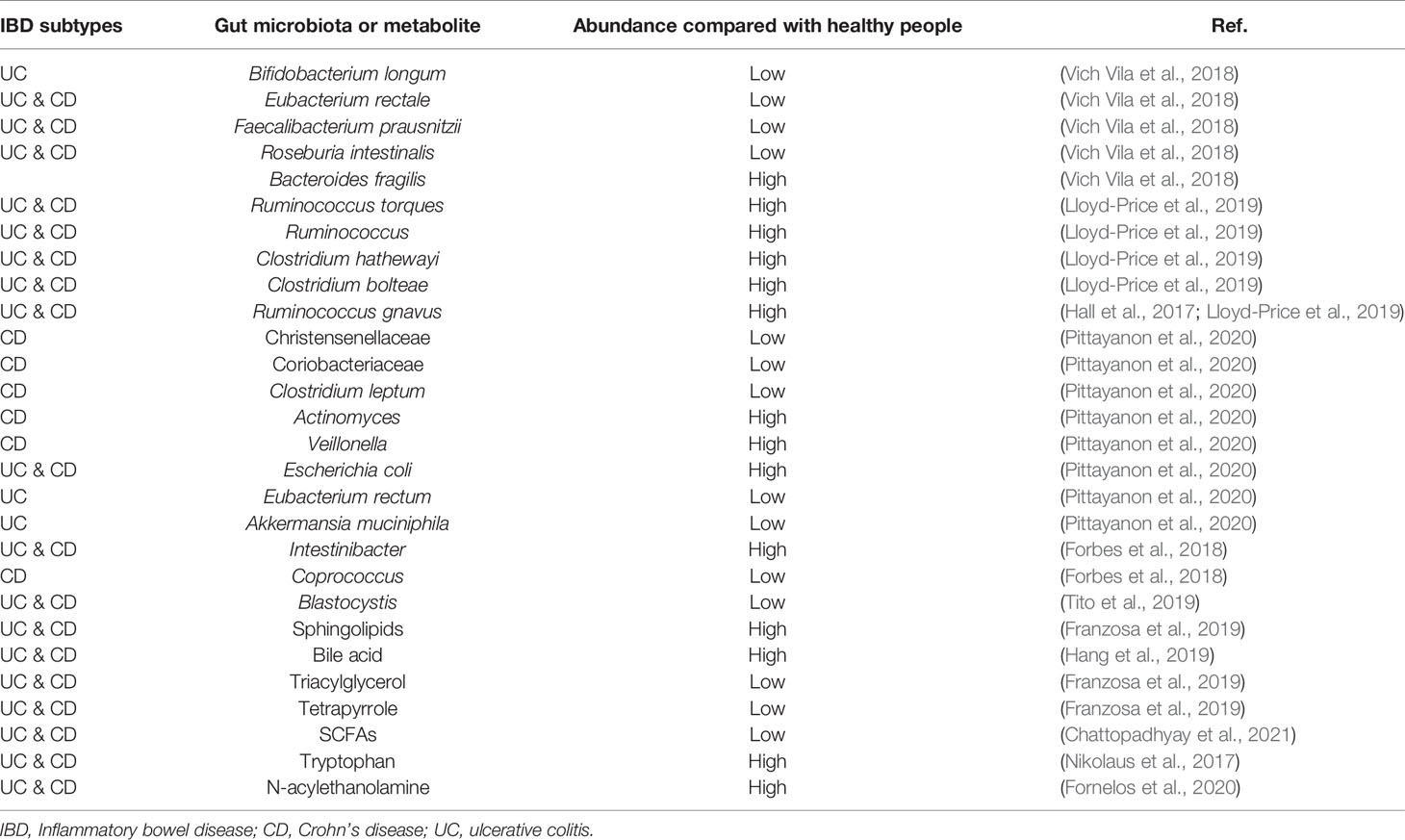
Table 1 The classification and abundance of gut microbiota and microbiota-derived metabolites in IBD.
Microbiota Interact With the Intestinal Epithelial Barrier in IBD
The intestinal barrier includes mechanical, chemical, immune, and microbial barriers. Roy et al. demonstrated that a distinct intestinal microbial community associated with the development of IBD is likely through its associated damage to the intestinal barrier via immune cells (Roy et al., 2017). Increasing evidence demonstrates that glycosylation of intestinal epithelial cells leads to increased expression of truncated O-glycans with altered expression of terminal glycan structures. Variability in glycan composition can disrupt the mucosal layer and immunity and ultimately contribute to the onset of IBD (Kudelka et al., 2020). Increased intestinal permeability has been proven to play a role in the pathogenesis of IBD (Llewellyn et al., 2018). The destruction of tight junction proteins leads to the damage of the intestinal mechanical barriers (Coyne and Comstock, 2019). A typical feature of IBD is dysbiosis of the gut microbiota, which results in the imbalance between beneficial and harmful bacteria taxa, and results in damage to the intestinal microbial barrier (Chelakkot et al., 2018). Some studies have linked the gut microbiota to metabolic defects of intestinal epithelial cells, especially through the Nod-like receptor (NLR) family. NLRX1 (nucleotide-binding oligomeric domain, X1 rich leucine-rich repeat) is a mitochondria-related NLR, which has a potential anti-inflammatory effect on colitis. NLRX1 is essential to maintain a balanced glutamine metabolism and barrier function in intestinal epithelial cells. The abundance of colitis-associated pathogens such as Veillonella spp. and Clostridium spp. in NLRX1 knockout mice increased. Supplementary feeding of glutamine can alleviate inflammation and induce changes in the gastrointestinal flora in NLRX1 knockout mice, while NLRX1 deletion affects the SIRT1 signaling pathway (Leber et al., 2018). Paneth cells are activated in SIRT1 knockout mice and promote nuclear factor kappa-B (NF-κB) pathway activation and ileum inflammation. The fecal microbiota is altered in SIRT1 knockout mice due to changes in bile acid metabolism. Moreover, SIRT1 knockout mice with gut microbiota dysbiosis have developed more serious colitis than control mice (Wellman et al., 2017). Previous studies have also shown a link between mitochondrial dysfunction and IBD (Rath et al., 2018; Mancini et al., 2021). In addition, mitochondrial dysfunction in intestinal epithelial cells and Paneth cells can induce ileal wall inflammation in mice. Mitochondrial respiratory dysfunction forces intestinal epithelial cells to acquire the abnormal phenotype of Paneth cells, resulting in metabolic imbalance and inflammation (Khaloian et al., 2020). Mitochondrial damage in patients with CD is also related to the decrease of H2S detoxification and the increase in the relative abundance of H2S-producing bacteria. The abundance of Atopobium parvulum, a type of H2S-producing bacteria, is related to the severity of CD patients. In CD, over 50% of patients present adherent-invasive E. coli (AIEC) colonization in the intestinal mucosa (Shawki and McCole, 2017). AIEC penetrate the mucus layer and adhere to intestinal epithelial cells through FimH and cell adhesion molecule 6 (Ceacam 6), and then colonize in the intestinal mucosa (Palmela et al., 2018). Enterobacteriaceae can use soluble factors released by apoptotic intestinal epithelial cells to promote the growth and colonization by driving the pyruvate formate-lyase-encoding pflB gene to induce the deterioration of IBD (Anderson et al., 2021). Klebsiella pneumoniae invades intestinal epithelial cells and interacts with macrophages to drive the release of interleukin (IL)-1β and tumor necrosis factor (TNF) (Read et al., 2021). Fusobacterium nucleatum can upregulate caspase recruitment domain 3 (CARD3) by Nucleotide-binding oligomerization domains 2 (NOD2) in colonic epithelial cells and thereby activates IL-17F/NF-κB signaling pathway and promotes the occurrence of intestinal inflammation (Chen et al., 2020).
The gut microbiota can produce various metabolites to prevent the invasion of pathogenic bacteria and promote intestinal homeostasis (Deleu et al., 2021). The gut microbiota and mucus secreted by Paneth cells play a vital role in the chemical barrier of the intestinal tract. SCFAs are an important metabolite produced by intestinal flora during dietary fiber fermentation (Louis et al., 2014; Lee and Chang, 2021). In addition, gut microbiota will compete with the pathogenic bacteria for nutrition, and the normal flora will play an antagonistic role against the pathogenic bacteria. F. prausnitzii can produce butyrate, which plays an anti-inflammatory role by inhibiting the IL-6/signal transducer and activator of transcription 3 (STAT3)/IL-17 pathway and promoting forkhead box protein P3 (Foxp3) (Zhou et al., 2018). Bacteriocin is an important antimicrobial agent. The bacteriocin-producing bacteria inhibits or competes with bacteria of the same species or related species. For example, Lactobacillus in humans produce lactobacillin and inhibit the infection of Listeria monocytogenes (Rolhion et al., 2019). Enterotoxin secreted by increased Enterotoxigenic E. coli (ETEC) increased the permeability of intestinal epithelium, which inhibits the uptake of ascorbic acid via the NF-κB pathway (Subramenium et al., 2019).
Immunoglobulin A (IgA) is the most common antibody subtype in the intestine. It is transported by polyclonal immunoglobulin receptor (pIgR) on host epithelial cells and is then released into the intestinal cavity in the form of secretory IgA (sIgA). In a stable environment, sIgA is important for the dynamic balance within the gut microbiota. When the pathogen secretes immunosuppressive protein, it can lead to the destruction of the intestinal immune barrier. IgA-coated bacteria from IBD patients are shown to invade into the mucus layer and exacerbate dextran sulfate sodium (DSS)-induced colitis in mouse models including Prevotellaceae, Helicobacter, and Segmented filamentous bacteria (SFB) (Kayama et al., 2020). The IgA coating patterns of gut microbiota in the IBD cohort were analyzed by using IgA-SEQ profiling, which found that the IgA coating levels of 43 bacterial taxa in IBD patients were higher than those in healthy individuals. The IgA coating levels of Vibrio can predict disease progression. Shapiro et al. showed that the analysis of the IgA response to microbiota could be used as a biomarker in the treatment of IBD (Shapiro et al., 2021). As IgA can bind to intestinal bacteria, breastfeeding can reduce the incidence rate of newborn enterocolitis since the specific component IgA in breast milk may mediate this protective effect (Gopalakrishna et al., 2019). Early intervention with FMT could stimulate sIgA secretion and modulate the gut microbiota composition (Cheng et al., 2019).
In addition to the excessive inflammatory response, there are other factors, such as flora imbalance and intestinal barrier disorder, involved in IBD development. Treatment aimed at ameliorating the intestinal barrier and gut microbiota may represent an important objective for the treatment of IBD in the future (Figure 2).
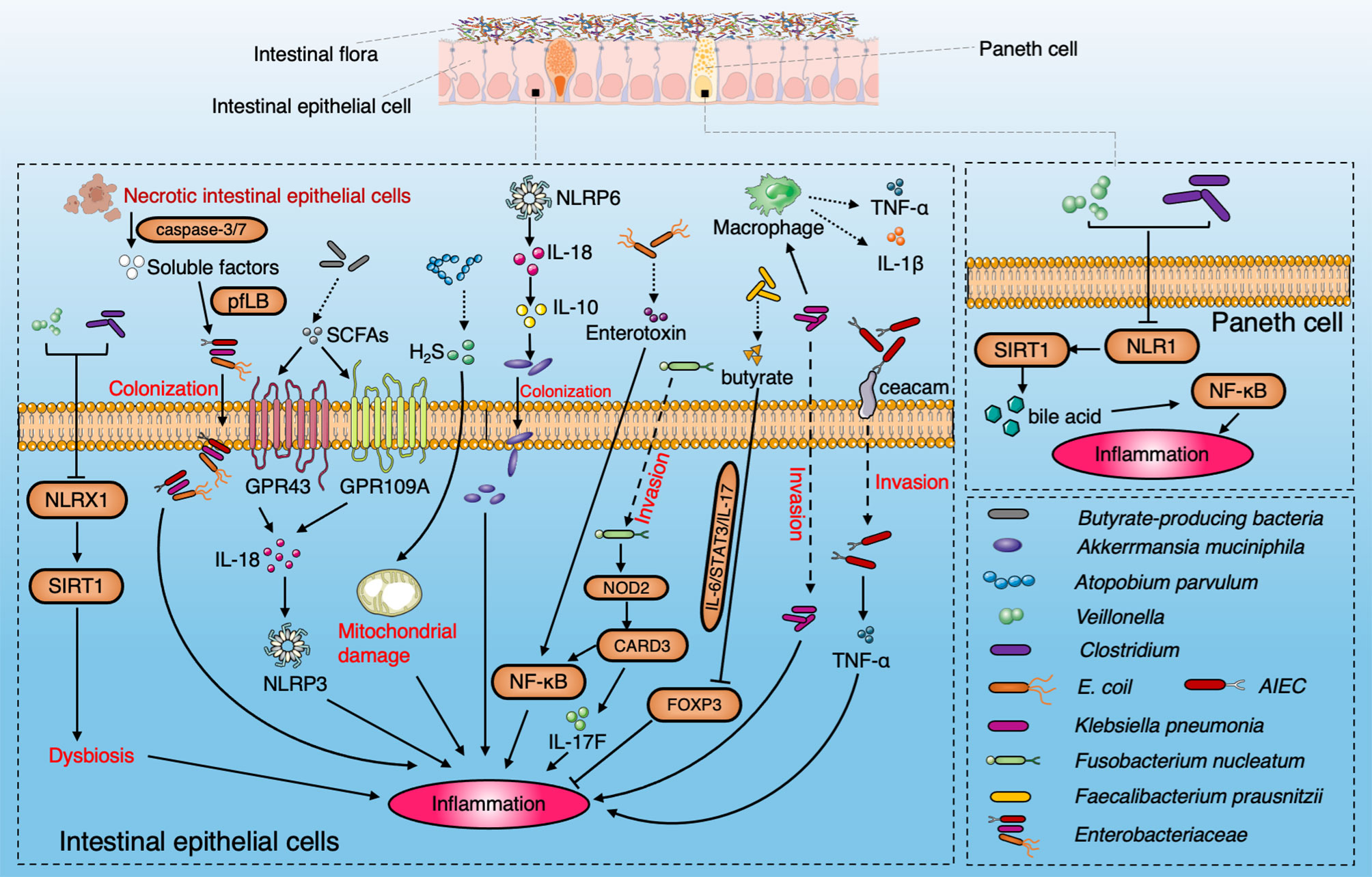
Figure 2 Diagram summarizing the pathogenic interaction between the gut microbiome and intestinal epithelial barrier in IBD. NLRX1 boosts the dysbiosis to induce the ileac inflammation via SIRT1 signaling pathway with enrichment of Veillonella and Clostridium. SCFAs produced by microbiota induce the activation of NLRP3 through GPR43 and GPR109A and protect colitis through IL-18. A. parvulum produce H2S to induce ileac inflammation by mitochondrial damage. NLRP6 is a key regulator to facilitate the colonization of A. muciniphila via IL-18 and IL-10 to promote IBD onset. AIEC penetrate the mucus layer and adhere to intestinal epithelial cells through FimH and ceacam6. Enterotoxin secreted by E. coli induce IBD by the NF-κB pathway. SIRT1 participates in the inflammation by stimulating Paneth cell to reflect the bile acids metabolism via the NF-κB signaling pathway. Klebsiella pneumoniae invades intestinal epithelial cells and promotes the secretion of IL-1β and TNF by interacting with macrophages. Fusobacterium nucleatum upregulates CARD3 via NOD2 in colonic epithelial cells to activate the IL-17F/NF-κB signaling pathway. Faecalibacterium prausnitzii produce butyrate to inhibit the IL-6/STAT3/IL-17 pathway to activate Foxp3.The soluble factors released by apoptotic intestinal epithelial cells through caspase3/7 facilitate the colonization of Enterobacteriaceae by driving the pyruvate formate-lyase-encoding pflB gene.
Microbiota-Derived Metabolites in IBD
The intestinal metabolome of IBD patients is disordered and is characterized by an imbalance of SCFA, bile acids, and tryptophan. Xavier et al. identified more than 2,700 bacteria metabolites with altered abundance in IBD. In addition, the production of sphingolipids and bile acid were upregulated in IBD patients, while triacylglycerol and tetrapyrrole were reduced. Overall, more than 50% of previously uncharacterized metabolites were detected, some of which might derive directly from the gut flora (Franzosa et al., 2019). SCFAs regulate mucosal immunity by promoting the development of B cells and the differentiation and expansion of regulatory T cells (Treg), and may activate the production of inflammatory cytokines. Butyric acid can act on immune cells in the intestinal mucosa and may increase the amount and activity of Treg, and inhibit the activity of neutrophils, macrophages, and dendritic cells. The dysbacteriosis and the increase of intestinal inflammatory cells in IBD patients are related to the decrease in SCFAs (Goncalves et al., 2018). The presence of butyrate-producing bacteria, such as E. coli, is consistent with the decrease in fecal SCFA levels (Chattopadhyay et al., 2021). SCFAs induce the activation of the NOD-like receptor family, containing three pyridine domains (NLRP3) through its G-protein-coupled receptor 43 (GPR43) and G protein-coupled receptor 109 (GPR109A), which induce ion efflux (K+ and Ca2+) and promote epithelial repair in colitis by regulating interleukin (IL)-18 (Macia et al., 2015). The effects of SCFA on macrophage polarization is associated with IBD. For example, antibiotic-induced SCFA depletion facilitates the transition to the M1 hyper-reactive phenotype, leading to the production of pro-inflammatory cytokines and promoting intestinal inflammation (Michaudel and Sokol, 2020).
Intestinal bacteria modulate bile acid levels to regulate host immunity (Campbell et al., 2020). Modified bile acids then activate two types of immune cells: Tregs and effector helper T cells, especially Th17, which regulate the immune response by inhibiting or promoting inflammation (Hang et al., 2019). Kasper et al. showed that the gut microbiota could modify bile acids and influence the levels of Treg cells in the mice colon (Song et al., 2020). These researchers speculated that bile acids played an immunomodulatory role by activating Farnesoid X receptor (FXR) and other receptors and decreased the activity of bile salt hydrolase in IBD patients, resulting in an imbalance between primary and secondary bile acids (Gadaleta et al., 2011). Heineken et al. analyzed 693 human gut microbiota genomes and revealed that each bacterium could produce 6 of 13 types of secondary bile acids, while the pairing of two microorganisms could produce 12 kinds of secondary bile acids, suggesting that the biotransformation of bile acids was achieved through interaction with bacteria. The bile acid production of the gut microbiota was significantly reduced in IBD children (Heinken et al., 2019).
Tryptophan is an essential aromatic amino acid. Dietary tryptophan is metabolized by host pathways (casein and serotonin pathway) and microbial pathways (indole pathway). Tryptophan can be metabolized to biologically activate indole by bacteria, which activates aryl hydrocarbon receptors and inhibits the production of inflammatory cytokines (Alexeev et al., 2018; Langan et al., 2021). In the cohort of 535 patients with IBD, the tryptophan metabolism level was associated with the severity of the disease (Nikolaus et al., 2017). In addition, CARD9 promotes the recovery of colitis by activating the IL-22 pathway. The microbiota cannot metabolize tryptophan as a ligand of the aromatic hydrocarbon receptor in the CARD9 knockout mouse model (Lamas et al., 2018). The lack of tryptophan in the diet is associated with the deterioration of colitis in a mouse model (Meisel et al., 2017).
N-acylethanolamine, an endogenous signal lipid, is a metabolite found in IBD, which is associated with Proteus enrichment and a reduction in Bacteroides levels. Targeting N-acylethanolamine may help to ameliorate IBD-related gut microbiota disorders (Fornelos et al., 2020).
The gut microbiota and its metabolites can regulate the innate and adaptive immune responses. The intervention of microbial metabolites may represent a potential approach for IBD treatment (Table 1).
Interaction Between Immune Cells and the Microbiota in IBD
Stronger antibody and T-cell responses to microbial antigens are common in patients with IBD (Moayyedi et al., 2015; Castellanos et al., 2018; Britton et al., 2020). A variety of immune cells and inflammatory factors participate in the initiation of IBD. The cooperation between immune response mediated by T-cell differentiation subsets and the gut microbiota may affect the occurrence of IBD (Inoue et al., 2005). The inflammatory response driven by Th cells protects the host from harmful pathogens, but the over-activation of Th cells is related to the onset and development of intestinal inflammation. It is generally believed that the occurrence of CD is mainly related to Th1 cells and Th17 cell activation, while the occurrence of UC was the result of the interaction between Th1 and Th2 cells (Chao et al., 2014; Lopetuso et al., 2018). Increasing studies have shown that the incidence of IBD is more correlated with Th17 cell activity. The cytokines secreted by Th17 cells are mainly IL-17 and IL-22, which play a vital role in mediating immune damage and autoimmune diseases. IL-22 is considered to play a protective role when acute colitis occurs; however, it is also found that IL-22 cooperates with IL17A to mediate pathogenicity in chronic colitis (Powell et al., 2020). Necrotic intestinal mucosal cells activate macrophages to produce IL-6 and Transforming growth factor-β (TGF-β) through Signal transducer and activator of transcription 3 (STAT3) and Retinoid related orphan receptors γ (RORγt), which induce the differentiation of Th17 cells (Owaga et al., 2015). Treg cells are mainly used to maintain the balance between T cells. In line with human data, the occurrence of IBD is related to the decrease or abnormal function of Treg cells. Due to the lack of immunosuppressive regulation of Treg cells, effector T cells can trigger an exaggerated immune response in the intestine that will eventually lead to intestinal mucosal injury (Sun et al., 2017). Th17 and Tregs are in equilibrium under normal conditions. The excessive increase of Th17 and the decrease of Tregs cells lead to a disorder in the Th17/Tregs balance. The destruction of this balance can lead to intestinal mucosal damage. IL-6 and low levels of TGF-β can stimulate T cells to differentiate into Th17. High levels of TGF-β can inhibit the production of Th17 cells and promote the production of Treg cells. Th17 cells can also inhibit the proliferation of Treg cells. Th17 cells increase and Tregs decrease in the peripheral blood of IBD patients, indicating that the imbalance of Th17/Tregs plays an important role in the development and maintenance of IBD. The gut microbiota can induce T cells to differentiate into Th17, Treg, and other phenotypes by shaping the intestinal microenvironment. SFB colonized in mouse small intestine can induce Th17 cells in the intestinal lamina propria to secrete IL-17 and IL-22 and promote intestinal inflammation (Lin and Zhang, 2017). Conversely, intestinal bacteria can also promote an increase in Treg cells with anti-inflammatory activity. Honda et al. found that the consumption of Clostridium spp. resulted in a significant reduction in intestinal Tregs in germ-free mice, and Clostridium spp. colonization promoted the aggregation of RORγ T+FOXP3T+pTreg cells, which, conversely, resulted in inhibiting the response of colonic Th2 and Th17 cells (Ohnmacht et al., 2015). B. fragilis, which is symbiotic with human beings, transmits immune regulatory molecules to immune cells through the secretion of outer membrane vesicles (OMVs). This mechanism of OMVs is closely associated with the expression of the IBD-related genes Atg16L1 and NOD2. OMVs play a protective role in IBD by activating the non-classical autophagy pathway. Atg16L1 deficient dendritic cells are unable to induce Tregs to inhibit mucosal inflammation. Human immune cells with Atg16L1 mutation present deficiencies in the Tregs response to OMV (Chu et al., 2016). B. thetaiotaomicron recapitulate the effects of gut microbiota and induces Tregs to influence the immune system in IBD (Hoffmann et al., 2016). F. prausnitzii-specific circulating CD4 and CD8a (DP8α) T cells have been identified in human colonic mucosa and serum, which exhibit similar features of Treg cells (Godefroy et al., 2018). The ectopic colonization of K. pneumoniae in the intestine did not improve the production of anti-inflammatory and regulatory T cells, but preferentially promoted the induction of Th1, showing its unique mechanism of inducing colitis (Atarashi et al., 2017).
Collectively, the dysbiosis of the gut microbiota, the imbalance of cytokines and the destruction of the mucosal barrier, contribute to induce mucosal inflammation and IBD development (Figure 3). Various susceptible genes and environmental factors can interfere with gut microbiota and the host immune system (Zhang et al., 2017).
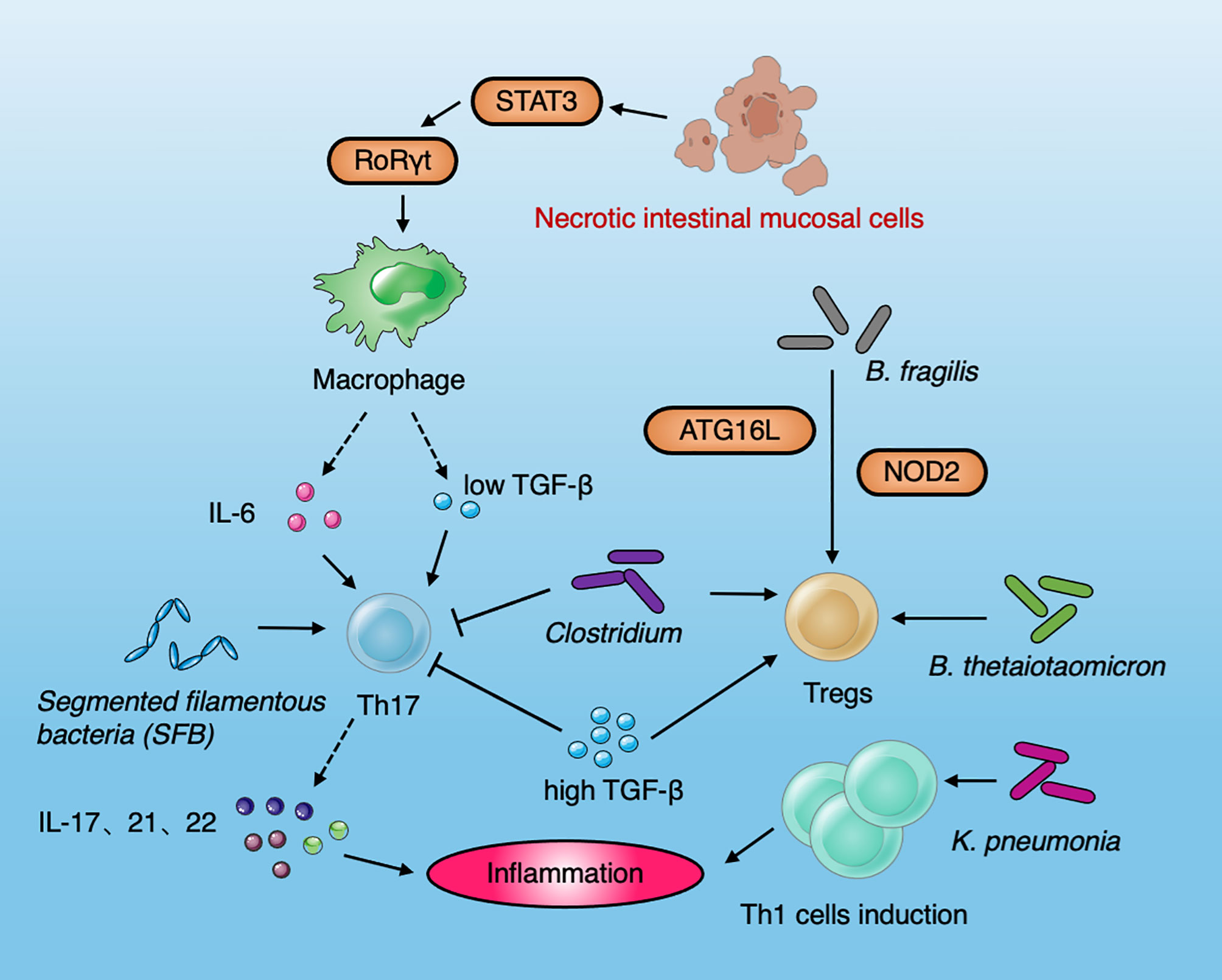
Figure 3 Diagram summarizing the interaction between immune cells and the microbiota in IBD. Necrotic intestinal mucosal cells activate macrophages to produce IL-6 and TGF-β through STAT3 and RORγt, which induce the differentiation of Th17 cells. IL-6 and low levels of TGF-β can stimulate T cells to differentiate into Th 17. High-levels of TGF-β can inhibit the production of Th 17 cells and promote the production of Treg cells. SFB can induce Th17 cells to secrete IL-17 and IL-22 and promote intestinal inflammation. The Clostridium spp. resulted in the production of Tregs. B. fragilis induced Tregs by the IBD-related genes Atg16L1 and NOD2. B. thetaiotaomicron recapitulate the effects of gut microbiota and induce Tregs to influence the immune system in IBD. Klebsiella pneumoniae improve the induction of Th1 cell to induce the occurrence of inflammation.
Microbiota-Targeted Treatment in IBD
There is a complex dynamic linkage between dysbiosis and the development of IBD, rather than a simple causal relationship (Sartor and Wu, 2017). Probiotics supplementation (especially butyrate-producing bacteria) and fecal microbiota transplantation (FMT) may be used to treat IBD. E. coli Nissle 1917 is a probiotic, which can inhibit the growth of Salmonella and other pathogenic bacteria. The H1 flagella of E. coli Nissle 1917 forms a tight network structure of the intestinal epithelium through the interaction between individual bacteria and anchoring recognition, which can inhibit the adhesion and invasion of pathogenic bacteria to intestinal epithelial cells. E. coli Nissle 1917 gene encodes three kinds of fimbriae, F1A, F1C, and curly fimbriae, which are powerful boosters for their continuous colonization and adhesion to intestinal epithelium. In addition, it secretes bacteriocin by competing with pathogens for nutrients in the intestine and antagonize the adhesion of pathogens to intestinal epithelium (Scaldaferri et al., 2016). A large clinical trial studied the efficacy of E. coli Nissle 1917 in maintaining the remission after clinical treatment in patients with UC (n = 120) compared with mesalazine treatment (1,500 mg/day). It was shown that E. coli Nissle 1917 and mesalazine were equivalent in maintaining the remission of UC (Nishida et al., 2021). IBD patients generally present symptoms of iron deficiency, and the iron-deficient intestinal microenvironment is more conducive to the intestinal colonization and curative effect of E. coli Nissle 1917. An iron-rich diet may weaken the efficacy of E. coli Nissle 1917, and the combination of treatment and diet will achieve twofold results with half the effort (Sassone-Corsi et al., 2016). The genetically engineered E. coli Nissle strain was used as a local probiotic to protect mice from chemically induced colitis and promote the mucosal healing (Praveschotinunt et al., 2019). A retrospective analysis of 200 patients with IBD found that probiotics reduced the incidence of side effects such as steroid medication, hospitalization, and surgery (Dore et al., 2020).
Based on the key role of the gut microbiota in the pathogenesis of IBD, FMT can restore the intestinal mucosal immune homeostasis for patients with IBD, which is also a current research hotspot. FMT can be applied for moderate to severe IBD complicated with recurrent or refractory Clostridium difficile infection (Colman and Rubin, 2014; Nishida et al., 2017; Goyal et al., 2018; Saha et al., 2021). Physiological and genetic factors related to donors or recipients will affect the curative effect. In addition, the use of specific flora and the method of administration should be reasonably designed and controlled. The factors to be considered in the selection of the flora include the abundance, classification, and composition of the bacterial community and whether to carry out antibiotic pretreatment. At the same time, drug formulation and administration are long-standing problems of FMT (Ng et al., 2020). It was found that triple antibiotics could cooperate with FMT to alleviate the intestinal ecological imbalance caused by the loss of Bacteroidetes species diversity in UC patients (Ishikawa et al., 2018). To date, several clinical trials have shown that intensive dose and multi-donor FMT could induce clinical and endoscopic remission (Moayyedi et al., 2015). However, one study found that the transferred microbes in the recipient who underwent FMT 10 weeks later were far from the donor bacteria, which provide a critical issue that the colonization dynamics of gut microbiota should be tracked and manipulated in the recipient of FMT (Chu et al., 2021). In addition, FMT is currently being investigated in registration and ethical clinical research, and it has not been approved for clinical application. Further research is needed to better understand the effective mechanisms underlying the effects of FMT, and to develop more effective regimens to treat IBD (Costea et al., 2017; Allegretti et al., 2019; Caruso et al., 2020).
Anti-tumor necrosis factor (anti-TNF) is a commonly used biological agent, which can prevent the development of inflammation by inhibiting the activity of TNF. At present, there are five kinds of anti-TNF agents: infliximab, adalimumab, etanercept, certolizumab, and golimumab. A German cohort study showed that anti-TNF treatment rendered fecal flora diversity similar to that of the control group. Fecal metabonomics analysis confirmed that the predicted levels of butyrate were significantly correlated with clinical remission after anti-TNF treatment (Aden et al., 2019). Zhuang et al. showed that the diversity and richness of the fecal microbiota in CD patients increased significantly after infliximab treatment, the number of bacteria-producing SCFAs increased, and the number of pathogenic bacteria decreased. Although anti-TNF therapy has changed the drug regimen for treatment of IBD, up to 30% of patients achieved no clinical benefit after the induction period, and up to 50% of patients have to withdraw from treatment (Zhuang et al., 2020).
The diet plays a crucial role in shaping the composition of the microbiota and can be regulated to control IBD symptoms. The effects of the diet on IBD occurs mainly through three mechanisms. First, some diets can change the composition of the intestinal microecology and its metabolic function, thus indirectly affecting intestinal immune function. Second, some dietary components can directly affect the intestinal mucosal barrier and induce a disorder of intestinal mucosal innate immunity indirectly. Finally, some components of the diet can directly participate in the intestinal immune response (Lewis and Abreu, 2017). Exclusive enteral nutrition (EEN) is a basic and polymerized diet without solid food, which is a dietary intervention widely studied in IBD (Levine et al., 2018). EEN has similar effects as corticosteroids in inducing remission in pediatric patients with CD without the known side effects of corticosteroid therapy (Heerasing et al., 2017; Pigneur et al., 2019). EEN is independent of other environmental factors and can rapidly change the composition of the microbiota and effectively reduce the intestinal inflammation of children with CD (Jongsma et al., 2020). The mechanisms by which EEN induces the remission of CD is unclear, but it may promote the growth of beneficial bacteria or the consumption of pathogens. Importantly, host susceptibility to IBD is highly dependent on the combination of specific fiber or protein components that constitute the microbiota, suggesting the importance of dietary composition in IBD.
Vedolizumab is a humanized monoclonal antibody, which can specifically bind to lymphocyte integrin α4β7, and thus inhibits the migration of lymphocytes from vascular endothelium to intestinal tissue (Fleisher et al., 2018). Xavier et al. found that 85 IBD patients receiving vedolizumab responded with high levels of Roseburia inulinivorans and Burkholderiales as the therapeutic effect. The authors identified 13 metabolic pathways that were more active in the intestinal bacteria of CD patients after remission (Stevens et al., 2017).
Gut-103 and Gut-108 are two living biotherapy products, which are mainly used to supplement the function of missing or representative deficiency in the dysbiosis of the gut microbiota of IBD patients. Gut-103, composed of 17 bacterial strains, cooperatively provides protection and continuous supplementation to the IBD inflammatory environment to prevent and treat chronic immune-mediated colitis. Gut-108 is an optimized version of Gut-103, which utilizes 11 human bacteria associated with 17 strains, which allow bacteria to remain in the colon longer than other probiotics that normally survive in the gut (van der Lelie et al., 2021).
HABN, a hyaluronic acid-coated bilirubin nanodrug, can accumulate in the inflamed colonic epithelium of mice with acute colitis after oral administration. The strong antioxidant effect of bilirubin allows HABN to protect the epithelial cells of the colon against apoptosis and promotes epithelial barrier recovery. HABN has been shown to regulate the gut microbiota, improve the richness and diversity of the flora, and increase A. muciniphila and Clostridium XIVα, which were beneficial to intestinal homeostasis. HABN interacts with pro-inflammatory macrophages through hyaluronic acid-CD44 to regulate the innate immune response and to reduce the release of inflammatory cytokines (Lee et al., 2020).
Recently, new drugs for the treatment of IBD have been developed, including agents targeting SMAD7 (Mongersen), IL-12 and IL-23 (ustekinumab), and Janus kinases (tofacitinib, filgotinib, and upadacitinib) (Monteleone et al., 2015; Feagan et al., 2016; Bradley, 2017; Hanauer et al., 2020; Feagan et al., 2021; Parigi et al., 2021). Since Enterobacteriaceae are implicated in a unique metabolic pathway whereby its overgrowth interferes with levels of beneficial bacteria, Winter et al. used the heavy metal tungsten to inhibit the proliferation of Enterobacteriaceae during IBD inflammation development (Zhu et al., 2018).
Although some regimens have been shown to have the potential or promising effect on IBD (Table 2). It is urgent to develop personalized treatment strategies and to identify which treatment regimens would be most beneficial to IBD patients. Using a combination of in vivo real-time molecular endoscopy technology, tissue transcriptome analysis, genetic research, gut microbiota analysis, and immune response analysis, we can comprehensively predict the response of patients to specific drugs.

Table 2 The impact mechanism of gut microbiota-targeted treatment in IBD.
Future Direction
Metabolites derived from the gut microbiota, especially SCFAs, are the key molecules of communication between the gut microbiota and host (Nikolaus et al., 2017; Osaka et al., 2017; Lavelle and Sokol, 2020). It is challenging to decipher all the effects of microbiota on immune metabolism. Part of this complexity is due to the final effects of microbial products, which may vary depending on the environment or the cell type. Functional metagenomics (e.g., expression of a metagenomic DNA library in bacteria such as E. coli), synthetic biology and bioinformatics, and the combination of these technologies in a strategic way have proved to be a very powerful approach, emphasizing the widespread presence of biologically active molecules or gene clusters in the population. These methods, using data and technology from different sources to mine the microbial community for new targeting compounds, provide a promising new direction for identifying bioactive molecules related to IBD. It is worth noting that family members of IBD patients can present changes in the metabolic spectrum similar to IBD characteristics, which may help to identify high-risk patients before clinical symptoms occur. In addition, high-resolution time mapping of the microbiome and metabolite markers may provide new biomarkers for predicting progression in diagnosis and treatment. By detecting 16S rRNA and other practical methods, we can target the gut microbiota and carry out the individualized treatment. Multiomics, such as genomics, transcriptomics, proteomics, immunomics, and microbiomics, can accurately analyze the onset of diseases (Lavelle and Sokol, 2018). New therapeutic drugs are constantly being developed, traditional therapeutic drugs are being optimized, and AI algorithms will be applied for the treatment of IBD (Rodrigues et al., 2018). International multi-center collaborative research and large-scale cohort studies will be implemented. Ultimately, an integrated personalized diseases profile (iPOP) will be developed by analyzing the specific disease context of patients at the molecular, cellular, tissue, organ, and body levels, and integrating all these data will help establish an “individual disease” model of each patient. Therefore, we will be able to develop precise therapeutic strategies for patients having similar individual disease spectrums to treat IBD.
Conclusion
IBD treatment is a long-term and complex process due to the complicated pathogenesis of IBD. Currently, a comprehensive evaluation system involving clinical, biochemical, endoscopic, and other indicators can direct optimal individualized treatment with the aim of predicting drug response. In the future, the combination of intestinal microbiology, gastroenterology epidemiology with rapid analysis of gut microbiota, metabolites, molecular signaling, and genetic engineering is expected to be the new therapeutic direction for IBD.
Author Contributions
Conception and design: TI, LF, JZ, and YL. Drafting of the manuscript: PQ and YL. Drawing of figures: YL and PQ. Conceiving and critical revision of the manuscript for important intellectual content: TI, ZZ, and YL. All authors contributed to the article and approved the submitted version.
Funding
This work is supported by the National Natural Science Foundation of China (Grant No. 82103468).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Charlesworth Author Service (https://www.cwauthors.com.cn/) for editing this manuscript.
Glossary

References
Aden, K., Rehman, A., Waschina, S., Pan, W. H., Walker, A., Lucio, M., et al. (2019). Metabolic Functions of Gut Microbes Associate With Efficacy of Tumor Necrosis Factor Antagonists in Patients With Inflammatory Bowel Diseases. Gastroenterology 157 (5), 1279–1292.e1211. doi: 10.1053/j.gastro.2019.07.025
Alexeev, E. E., Lanis, J. M., Kao, D. J., Campbell, E. L., Kelly, C. J., Battista, K. D., et al. (2018). Microbiota-Derived Indole Metabolites Promote Human and Murine Intestinal Homeostasis Through Regulation of Interleukin-10 Receptor. Am. J. Pathol. 188 (5), 1183–1194. doi: 10.1016/j.ajpath.2018.01.011
Allegretti, J. R., Mullish, B. H., Kelly, C., Fischer, M. (2019). The Evolution of the Use of Faecal Microbiota Transplantation and Emerging Therapeutic Indications. Lancet 394 (10196), 420–431. doi: 10.1016/S0140-6736(19)31266-8
Allen-Vercoe, E., Coburn, B. (2020). A Microbiota-Derived Metabolite Augments Cancer Immunotherapy Responses in Mice. Cancer Cell 38 (4), 452–453. doi: 10.1016/j.ccell.2020.09.005
Anderson, C. J., Medina, C. B., Barron, B. J., Karvelyte, L., Aaes, T. L., Lambertz, I., et al. (2021). Microbes Exploit Death-Induced Nutrient Release by Gut Epithelial Cells. Nature 596 (7871), 262–267. doi: 10.1038/s41586-021-03785-9
Atarashi, K., Suda, W., Luo, C., Kawaguchi, T., Motoo, I., Narushima, S., et al. (2017). Ectopic Colonization of Oral Bacteria in the Intestine Drives TH1 Cell Induction and Inflammation. Science 358 (6361), 359–365. doi: 10.1126/science.aan4526
Bradley, C. A. (2017). IBD: Tofacitinib Effective in Ulcerative Colitis. Nat. Rev. Gastroenterol. Hepatol. 14 (7), 388. doi: 10.1038/nrgastro.2017.66
Britton, G. J., Contijoch, E. J., Spindler, M. P., Aggarwala, V., Dogan, B., Bongers, G., et al. (2020). Defined Microbiota Transplant Restores Th17/RORgammat(+) Regulatory T Cell Balance in Mice Colonized With Inflammatory Bowel Disease Microbiotas. Proc. Natl. Acad. Sci. U. S. A. 117 (35), 21536–21545. doi: 10.1073/pnas.1922189117
Campbell, C., McKenney, P. T., Konstantinovsky, D., Isaeva, O. I., Schizas, M., Verter, J., et al. (2020). Bacterial Metabolism of Bile Acids Promotes Generation of Peripheral Regulatory T Cells. Nature 581 (7809), 475–479. doi: 10.1038/s41586-020-2193-0
Caruso, R., Lo, B. C., Nunez, G. (2020). Host-Microbiota Interactions in Inflammatory Bowel Disease. Nat. Rev. Immunol. 20 (7), 411–426. doi: 10.1038/s41577-019-0268-7
Castellanos, J. G., Woo, V., Viladomiu, M., Putzel, G., Lima, S., Diehl, G. E., et al. (2018). Microbiota-Induced TNF-Like Ligand 1A Drives Group 3 Innate Lymphoid Cell-Mediated Barrier Protection and Intestinal T Cell Activation During Colitis. Immunity 49 (6), 1077–1089.e1075. doi: 10.1016/j.immuni.2018.10.014
Cekin, A. H. (2017). A Microbial Signature for Crohn’s Disease. Turk. J. Gastroenterol. 28 (3), 237–238. doi: 10.5152/tjg.2017.24031
Chao, K., Zhang, S., Yao, J., He, Y., Chen, B., Zeng, Z., et al. (2014). Imbalances of CD4(+) T-Cell Subgroups in Crohn’s Disease and Their Relationship With Disease Activity and Prognosis. J. Gastroenterol. Hepatol. 29 (10), 1808–1814. doi: 10.1111/jgh.12592
Chattopadhyay, I., Dhar, R., Pethusamy, K., Seethy, A., Srivastava, T., Sah, R., et al. (2021). Exploring the Role of Gut Microbiome in Colon Cancer. Appl. Biochem. Biotechnol. 193 (6), 1780–1799. doi: 10.1007/s12010-021-03498-9
Chelakkot, C., Ghim, J., Ryu, S. H. (2018). Mechanisms Regulating Intestinal Barrier Integrity and its Pathological Implications. Exp. Mol. Med. 50 (8), 1–9. doi: 10.1038/s12276-018-0126-x
Chen, Y., Chen, Y., Cao, P., Su, W., Zhan, N., Dong, W. (2020). Fusobacterium nucleatum Facilitates Ulcerative Colitis Through Activating IL-17F Signaling to NF-kappaB via the Upregulation of CARD3 Expression. J. Pathol. 250 (2), 170–182. doi: 10.1002/path.5358
Cheng, C. S., Wei, H. K., Wang, P., Yu, H. C., Zhang, X. M., Jiang, S. W., et al. (2019). Early Intervention With Faecal Microbiota Transplantation: An Effective Means to Improve Growth Performance and the Intestinal Development of Suckling Piglets. Animal 13 (3), 533–541. doi: 10.1017/S1751731118001611
Chu, N. D., Crothers, J. W., Nguyen, L. T. T., Kearney, S. M., Smith, M. B., Kassam, Z., et al. (2021). Dynamic Colonization of Microbes and Their Functions After Fecal Microbiota Transplantation for Inflammatory Bowel Disease. mBio 12 (4), e0097521. doi: 10.1128/mBio.00975-21
Chu, H., Khosravi, A., Kusumawardhani, I. P., Kwon, A. H., Vasconcelos, A. C., Cunha, L. D., et al. (2016). Gene-Microbiota Interactions Contribute to the Pathogenesis of Inflammatory Bowel Disease. Science 352 (6289), 1116–1120. doi: 10.1126/science.aad9948
Colman, R. J., Rubin, D. T. (2014). Fecal Microbiota Transplantation as Therapy for Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis. J. Crohns Colitis 8 (12), 1569–1581. doi: 10.1016/j.crohns.2014.08.006
Costea, P. I., Zeller, G., Sunagawa, S., Pelletier, E., Alberti, A., Levenez, F., et al. (2017). Towards Standards for Human Fecal Sample Processing in Metagenomic Studies. Nat. Biotechnol. 35 (11), 1069–1076. doi: 10.1038/nbt.3960
Coyne, M. J., Comstock, L. E. (2019). Type VI Secretion Systems and the Gut Microbiota. Microbiol. Spectr. 7 (2). doi: 10.1128/microbiolspec.PSIB-0009-2018
Deleu, S., Machiels, K., Raes, J., Verbeke, K., Vermeire, S. (2021). Short Chain Fatty Acids and its Producing Organisms: An Overlooked Therapy for IBD? EBioMedicine 66, 103293. doi: 10.1016/j.ebiom.2021.103293
Dore, M. P., Rocchi, C., Longo, N. P., Scanu, A. M., Vidili, G., Padedda, F., et al. (2020). Effect of Probiotic Use on Adverse Events in Adult Patients With Inflammatory Bowel Disease: A Retrospective Cohort Study. Probiotics Antimicrob. Proteins 12 (1), 152–159. doi: 10.1007/s12602-019-9517-0
Feagan, B. G., Danese, S., Loftus, E. V., Jr., Vermeire, S., Schreiber, S., Ritter, T., et al. (2021). Filgotinib as Induction and Maintenance Therapy for Ulcerative Colitis (SELECTION): A Phase 2b/3 Double-Blind, Randomised, Placebo-Controlled Trial. Lancet 397 (10292), 2372–2384. doi: 10.1016/S0140-6736(21)00666-8
Feagan, B. G., Sandborn, W. J., Gasink, C., Jacobstein, D., Lang, Y., Friedman, J. R., et al. (2016). Ustekinumab as Induction and Maintenance Therapy for Crohn’s Disease. N. Engl. J. Med. 375 (20), 1946–1960. doi: 10.1056/NEJMoa1602773
Fleisher, M., Marsal, J., Lee, S. D., Frado, L. E., Parian, A., Korelitz, B. I., et al. (2018). Effects of Vedolizumab Therapy on Extraintestinal Manifestations in Inflammatory Bowel Disease. Dig. Dis. Sci. 63 (4), 825–833. doi: 10.1007/s10620-018-4971-1
Forbes, J. D., Chen, C. Y., Knox, N. C., Marrie, R. A., El-Gabalawy, H., de Kievit, T., et al. (2018). A Comparative Study of the Gut Microbiota in Immune-Mediated Inflammatory Diseases-Does a Common Dysbiosis Exist? Microbiome 6 (1), 221. doi: 10.1186/s40168-018-0603-4
Fornelos, N., Franzosa, E. A., Bishai, J., Annand, J. W., Oka, A., Lloyd-Price, J., et al. (2020). Growth Effects of N-Acylethanolamines on Gut Bacteria Reflect Altered Bacterial Abundances in Inflammatory Bowel Disease. Nat. Microbiol. 5 (3), 486–497. doi: 10.1038/s41564-019-0655-7
Franzosa, E. A., Sirota-Madi, A., Avila-Pacheco, J., Fornelos, N., Haiser, H. J., Reinker, S., et al. (2019). Gut Microbiome Structure and Metabolic Activity in Inflammatory Bowel Disease. Nat. Microbiol. 4 (2), 293–305. doi: 10.1038/s41564-018-0306-4
Gadaleta, R. M., van Erpecum, K. J., Oldenburg, B., Willemsen, E. C., Renooij, W., Murzilli, S., et al. (2011). Farnesoid X Receptor Activation Inhibits Inflammation and Preserves the Intestinal Barrier in Inflammatory Bowel Disease. Gut 60 (4), 463–472. doi: 10.1136/gut.2010.212159
Godefroy, E., Alameddine, J., Montassier, E., Mathe, J., Desfrancois-Noel, J., Marec, N., et al. (2018). Expression of CCR6 and CXCR6 by Gut-Derived CD4(+)/CD8alpha(+) T-Regulatory Cells, Which are Decreased in Blood Samples From Patients With Inflammatory Bowel Diseases. Gastroenterology 155 (4), 1205–1217. doi: 10.1053/j.gastro.2018.06.078
Goncalves, P., Araujo, J. R., Di Santo, J. P. (2018). A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflammation Bowel Dis. 24 (3), 558–572. doi: 10.1093/ibd/izx029
Gopalakrishna, K. P., Macadangdang, B. R., Rogers, M. B., Tometich, J. T., Firek, B. A., Baker, R., et al. (2019). Maternal IgA Protects Against the Development of Necrotizing Enterocolitis in Preterm Infants. Nat. Med. 25 (7), 1110–1115. doi: 10.1038/s41591-019-0480-9
Goyal, A., Yeh, A., Bush, B. R., Firek, B. A., Siebold, L. M., Rogers, M. B., et al. (2018). Safety, Clinical Response, and Microbiome Findings Following Fecal Microbiota Transplant in Children With Inflammatory Bowel Disease. Inflammation Bowel Dis. 24 (2), 410–421. doi: 10.1093/ibd/izx035
Halfvarson, J., Brislawn, C. J., Lamendella, R., Vazquez-Baeza, Y., Walters, W. A., Bramer, L. M., et al. (2017). Dynamics of the Human Gut Microbiome in Inflammatory Bowel Disease. Nat. Microbiol. 2, 17004. doi: 10.1038/nmicrobiol.2017.4
Hall, A. B., Yassour, M., Sauk, J., Garner, A., Jiang, X., Arthur, T., et al. (2017). A Novel Ruminococcus gnavus Clade Enriched in Inflammatory Bowel Disease Patients. Genome Med. 9 (1), 103. doi: 10.1186/s13073-017-0490-5
Hanauer, S. B., Sandborn, W. J., Feagan, B. G., Gasink, C., Jacobstein, D., Zou, B., et al. (2020). IM-UNITI: Three-Year Efficacy, Safety, and Immunogenicity of Ustekinumab Treatment of Crohn’s Disease. J. Crohns Colitis 14 (1), 23–32. doi: 10.1093/ecco-jcc/jjz110
Hang, S., Paik, D., Yao, L., Kim, E., Trinath, J., Lu, J., et al. (2019). Bile Acid Metabolites Control TH17 and Treg Cell Differentiation. Nature 576 (7785), 143–148. doi: 10.1038/s41586-019-1785-z
Heerasing, N., Thompson, B., Hendy, P., Heap, G. A., Walker, G., Bethune, R., et al. (2017). Exclusive Enteral Nutrition Provides an Effective Bridge to Safer Interval Elective Surgery for Adults With Crohn’s Disease. Aliment. Pharmacol. Ther. 45 (5), 660–669. doi: 10.1111/apt.13934
Heinken, A., Ravcheev, D. A., Baldini, F., Heirendt, L., Fleming, R. M. T., Thiele, I. (2019). Systematic Assessment of Secondary Bile Acid Metabolism in Gut Microbes Reveals Distinct Metabolic Capabilities in Inflammatory Bowel Disease. Microbiome 7 (1), 75. doi: 10.1186/s40168-019-0689-3
Hoffmann, T. W., Pham, H. P., Bridonneau, C., Aubry, C., Lamas, B., Martin-Gallausiaux, C., et al. (2016). Microorganisms Linked to Inflammatory Bowel Disease-Associated Dysbiosis Differentially Impact Host Physiology in Gnotobiotic Mice. ISME J. 10 (2), 460–477. doi: 10.1038/ismej.2015.127
Hu, S., Vich Vila, A., Gacesa, R., Collij, V., Stevens, C., Fu, J. M., et al. (2021). Whole Exome Sequencing Analyses Reveal Gene-Microbiota Interactions in the Context of IBD. Gut 70 (2), 285–296. doi: 10.1136/gutjnl-2019-319706
Imhann, F., Vich Vila, A., Bonder, M. J., Fu, J., Gevers, D., Visschedijk, M. C., et al. (2018). Interplay of Host Genetics and Gut Microbiota Underlying the Onset and Clinical Presentation of Inflammatory Bowel Disease. Gut 67 (1), 108–119. doi: 10.1136/gutjnl-2016-312135
Inoue, S., Nakase, H., Chiba, T. (2005). Etiopathogenesis and Aggravating Factors in Ulcerative Colitis. Nihon Rinsho 63 (5), 757–762.
Ishikawa, D., Sasaki, T., Takahashi, M., Kuwahara-Arai, K., Haga, K., Ito, S., et al. (2018). The Microbial Composition of Bacteroidetes Species in Ulcerative Colitis is Effectively Improved by Combination Therapy With Fecal Microbiota Transplantation and Antibiotics. Inflammation Bowel Dis. 24 (12), 2590–2598. doi: 10.1093/ibd/izy266
Jongsma, M. M. E., Aardoom, M. A., Cozijnsen, M. A., van Pieterson, M., de Meij, T., Groeneweg, M., et al. (2020). First-Line Treatment With Infliximab Versus Conventional Treatment in Children With Newly Diagnosed Moderate-to-Severe Crohn’s Disease: An Open-Label Multicentre Randomised Controlled Trial. Gut 71 (1), 34–42. doi: 10.1136/gutjnl-2020-322339
Kayama, H., Okumura, R., Takeda, K. (2020). Interaction Between the Microbiota, Epithelia, and Immune Cells in the Intestine. Annu. Rev. Immunol. 38, 23–48. doi: 10.1146/annurev-immunol-070119-115104
Khaloian, S., Rath, E., Hammoudi, N., Gleisinger, E., Blutke, A., Giesbertz, P., et al. (2020). Mitochondrial Impairment Drives Intestinal Stem Cell Transition Into Dysfunctional Paneth Cells Predicting Crohn’s Disease Recurrence. Gut 69 (11), 1939–1951. doi: 10.1136/gutjnl-2019-319514
Kostic, A. D., Xavier, R. J., Gevers, D. (2014). The Microbiome in Inflammatory Bowel Disease: Current Status and the Future Ahead. Gastroenterology 146 (6), 1489–1499. doi: 10.1053/j.gastro.2014.02.009
Kriss, M., Hazleton, K. Z., Nusbacher, N. M., Martin, C. G., Lozupone, C. A. (2018). Low Diversity Gut Microbiota Dysbiosis: Drivers, Functional Implications and Recovery. Curr. Opin. Microbiol. 44, 34–40. doi: 10.1016/j.mib.2018.07.003
Kudelka, M. R., Stowell, S. R., Cummings, R. D., Neish, A. S. (2020). Intestinal Epithelial Glycosylation in Homeostasis and Gut Microbiota Interactions in IBD. Nat. Rev. Gastroenterol. Hepatol. 17 (10), 597–617. doi: 10.1038/s41575-020-0331-7
Lamas, B., Michel, M. L., Waldschmitt, N., Pham, H. P., Zacharioudaki, V., Dupraz, L., et al. (2018). Card9 Mediates Susceptibility to Intestinal Pathogens Through Microbiota Modulation and Control of Bacterial Virulence. Gut 67 (10), 1836–1844. doi: 10.1136/gutjnl-2017-314195
Langan, D., Perkins, D. J., Vogel, S. N., Moudgil, K. D. (2021). Microbiota-Derived Metabolites, Indole-3-Aldehyde and Indole-3-Acetic Acid, Differentially Modulate Innate Cytokines and Stromal Remodeling Processes Associated With Autoimmune Arthritis. Int. J. Mol. Sci. 22 (4). doi: 10.3390/ijms22042017
Lavelle, A., Hoffmann, T. W., Pham, H. P., Langella, P., Guedon, E., Sokol, H. (2019). Baseline Microbiota Composition Modulates Antibiotic-Mediated Effects on the Gut Microbiota and Host. Microbiome 7 (1), 111. doi: 10.1186/s40168-019-0725-3
Lavelle, A., Sokol, H. (2018). Gut Microbiota: Beyond Metagenomics, Metatranscriptomics Illuminates Microbiome Functionality in IBD. Nat. Rev. Gastroenterol. Hepatol. 15 (4), 193–194. doi: 10.1038/nrgastro.2018.15
Lavelle, A., Sokol, H. (2020). Gut Microbiota-Derived Metabolites as Key Actors in Inflammatory Bowel Disease. Nat. Rev. Gastroenterol. Hepatol. 17 (4), 223–237. doi: 10.1038/s41575-019-0258-z
Leber, A., Hontecillas, R., Tubau-Juni, N., Zoccoli-Rodriguez, V., Abedi, V., Bassaganya-Riera, J. (2018). NLRX1 Modulates Immunometabolic Mechanisms Controlling the Host-Gut Microbiota Interactions During Inflammatory Bowel Disease. Front. Immunol. 9. doi: 10.3389/fimmu.2018.00363
Lee, M., Chang, E. B. (2021). Inflammatory Bowel Diseases (IBD) and the Microbiome-Searching the Crime Scene for Clues. Gastroenterology 160 (2), 524–537. doi: 10.1053/j.gastro.2020.09.056
Lee, Y., Sugihara, K., Gillilland, M. G., 3rd, Jon, S., Kamada, N., Moon, J. J. (2020). Hyaluronic Acid-Bilirubin Nanomedicine for Targeted Modulation of Dysregulated Intestinal Barrier, Microbiome and Immune Responses in Colitis. Nat. Mater. 19 (1), 118–126. doi: 10.1038/s41563-019-0462-9
Levine, A., Sigall Boneh, R., Wine, E. (2018). Evolving Role of Diet in the Pathogenesis and Treatment of Inflammatory Bowel Diseases. Gut 67 (9), 1726–1738. doi: 10.1136/gutjnl-2017-315866
Lewis, J. D., Abreu, M. T. (2017). Diet as a Trigger or Therapy for Inflammatory Bowel Diseases. Gastroenterology 152 (2), 398–414, e396. doi: 10.1053/j.gastro.2016.10.019
Lin, L., Zhang, J. (2017). Role of Intestinal Microbiota and Metabolites on Gut Homeostasis and Human Diseases. BMC Immunol. 18 (1), 2. doi: 10.1186/s12865-016-0187-3
Llewellyn, S. R., Britton, G. J., Contijoch, E. J., Vennaro, O. H., Mortha, A., Colombel, J. F., et al. (2018). Interactions Between Diet and the Intestinal Microbiota Alter Intestinal Permeability and Colitis Severity in Mice. Gastroenterology 1541037-1046 (4), e1032. doi: 10.1053/j.gastro.2017.11.030
Lloyd-Price, J., Arze, C., Ananthakrishnan, A. N., Schirmer, M., Avila-Pacheco, J., Poon, T. W., et al. (2019). Multi-Omics of the Gut Microbial Ecosystem in Inflammatory Bowel Diseases. Nature 569 (7758), 655–662. doi: 10.1038/s41586-019-1237-9
Lopetuso, L. R., Ianiro, G., Scaldaferri, F., Cammarota, G., Gasbarrini, A. (2016). Gut Virome and Inflammatory Bowel Disease. Inflammation Bowel Dis. 22 (7), 1708–1712. doi: 10.1097/MIB.0000000000000807
Lopetuso, L. R., Napoli, M., Rizzatti, G., Gasbarrini, A. (2018). The Intriguing Role of Rifaximin in Gut Barrier Chronic Inflammation and in the Treatment of Crohn’s Disease. Expert Opin. Investig. Drugs 27 (6), 543–551. doi: 10.1080/13543784.2018.1483333
Louis, P., Hold, G. L., Flint, H. J. (2014). The Gut Microbiota, Bacterial Metabolites and Colorectal Cancer. Nat. Rev. Microbiol. 12 (10), 661–672. doi: 10.1038/nrmicro3344
Macia, L., Tan, J., Vieira, A. T., Leach, K., Stanley, D., Luong, S., et al. (2015). Metabolite-Sensing Receptors GPR43 and GPR109A Facilitate Dietary Fibre-Induced Gut Homeostasis Through Regulation of the Inflammasome. Nat. Commun. 6, 6734. doi: 10.1038/ncomms7734
Mancini, N. L., Rajeev, S., Jayme, T. S., Wang, A., Keita, A. V., Workentine, M. L., et al. (2021). Crohn’s Disease Pathobiont Adherent-Invasive E. coli Disrupts Epithelial Mitochondrial Networks With Implications for Gut Permeability. Cell Mol. Gastroenterol. Hepatol. 11 (2), 551–571. doi: 10.1016/j.jcmgh.2020.09.013
Manichanh, C., Borruel, N., Casellas, F., Guarner, F. (2012). The Gut Microbiota in IBD. Nat. Rev. Gastroenterol. Hepatol. 9 (10), 599–608. doi: 10.1038/nrgastro.2012.152
Marchesi, J. R., Adams, D. H., Fava, F., Hermes, G. D., Hirschfield, G. M., Hold, G., et al. (2016). The Gut Microbiota and Host Health: A New Clinical Frontier. Gut 65 (2), 330–339. doi: 10.1136/gutjnl-2015-309990
Meisel, M., Mayassi, T., Fehlner-Peach, H., Koval, J. C., O’Brien, S. L., Hinterleitner, R., et al. (2017). Interleukin-15 Promotes Intestinal Dysbiosis With Butyrate Deficiency Associated With Increased Susceptibility to Colitis. ISME J. 11 (1), 15–30. doi: 10.1038/ismej.2016.114
Michaudel, C., Sokol, H. (2020). The Gut Microbiota at the Service of Immunometabolism. Cell Metab. 32 (4), 514–523. doi: 10.1016/j.cmet.2020.09.004
Moayyedi, P., Surette, M. G., Kim, P. T., Libertucci, J., Wolfe, M., Onischi, C., et al. (2015). Fecal Microbiota Transplantation Induces Remission in Patients With Active Ulcerative Colitis in a Randomized Controlled Trial. Gastroenterology 149 (1), 102–109.e106. doi: 10.1053/j.gastro.2015.04.001
Monteleone, G., Neurath, M. F., Ardizzone, S., Di Sabatino, A., Fantini, M. C., Castiglione, F., et al. (2015). Mongersen, an Oral SMAD7 Antisense Oligonucleotide, and Crohn’s Disease. N. Engl. J. Med. 372 (12), 1104–1113. doi: 10.1056/NEJMoa1407250
Mosca, A., Leclerc, M., Hugot, J. P. (2016). Gut Microbiota Diversity and Human Diseases: Should We Reintroduce Key Predators in Our Ecosystem? Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00455
Mukhopadhya, I., Hansen, R., Meharg, C., Thomson, J. M., Russell, R. K., Berry, S. H., et al. (2015). The Fungal Microbiota of De-Novo Paediatric Inflammatory Bowel Disease. Microbes Infect. 17 (4), 304–310. doi: 10.1016/j.micinf.2014.12.001
Ng, S. C., Kamm, M. A., Yeoh, Y. K., Chan, P. K. S., Zuo, T., Tang, W., et al. (2020). Scientific Frontiers in Faecal Microbiota Transplantation: Joint Document of Asia-Pacific Association of Gastroenterology (APAGE) and Asia-Pacific Society for Digestive Endoscopy (APSDE). Gut 69 (1), 83–91. doi: 10.1136/gutjnl-2019-319407
Ng, S. C., Shi, H. Y., Hamidi, N., Underwood, F. E., Tang, W., Benchimol, E. I., et al. (2017). Worldwide Incidence and Prevalence of Inflammatory Bowel Disease in the 21st Century: A Systematic Review of Population-Based Studies. Lancet 390 (10114), 2769–2778. doi: 10.1016/S0140-6736(17)32448-0
Nikolaus, S., Schulte, B., Al-Massad, N., Thieme, F., Schulte, D. M., Bethge, J., et al. (2017). Increased Tryptophan Metabolism is Associated With Activity of Inflammatory Bowel Diseases. Gastroenterology 153 (6), 1504–1516.e1502. doi: 10.1053/j.gastro.2017.08.028
Nishida, A., Imaeda, H., Ohno, M., Inatomi, O., Bamba, S., Sugimoto, M., et al. (2017). Efficacy and Safety of Single Fecal Microbiota Transplantation for Japanese Patients With Mild to Moderately Active Ulcerative Colitis. J. Gastroenterol. 52 (4), 476–482. doi: 10.1007/s00535-016-1271-4
Nishida, A., Nishino, K., Sakai, K., Owaki, Y., Noda, Y., Imaeda, H. (2021). Can Control of Gut Microbiota be a Future Therapeutic Option for Inflammatory Bowel Disease? World J. Gastroenterol. 27 (23), 3317–3326. doi: 10.3748/wjg.v27.i23.3317
Ohnmacht, C., Park, J. H., Cording, S., Wing, J. B., Atarashi, K., Obata, Y., et al. (2015). The Microbiota Regulates Type 2 Immunity Through RORgammat(+) T Cells. Science 349 (6251), 989–993. doi: 10.1126/science.aac4263
Oligschlaeger, Y., Yadati, T., Houben, T., Condello Olivan, C. M., Shiri-Sverdlov, R. (2019). Inflammatory Bowel Disease: A Stressed “Gut/Feeling”. Cells 8 (7). doi: 10.3390/cells8070659
Osaka, T., Moriyama, E., Arai, S., Date, Y., Yagi, J., Kikuchi, J., et al. (2017). Meta-Analysis of Fecal Microbiota and Metabolites in Experimental Colitic Mice During the Inflammatory and Healing Phases. Nutrients 9 (12). doi: 10.3390/nu9121329
Owaga, E., Hsieh, R. H., Mugendi, B., Masuku, S., Shih, C. K., Chang, J. S. (2015). Th17 Cells as Potential Probiotic Therapeutic Targets in Inflammatory Bowel Diseases. Int. J. Mol. Sci. 16 (9), 20841–20858. doi: 10.3390/ijms160920841
Palmela, C., Chevarin, C., Xu, Z., Torres, J., Sevrin, G., Hirten, R., et al. (2018). Adherent-Invasive Escherichia coli in Inflammatory Bowel Disease. Gut 67 (3), 574–587. doi: 10.1136/gutjnl-2017-314903
Parigi, T. L., D’Amico, F., Danese, S. (2021). Upadacitinib for Crohn’s Disease and Ulcerative Colitis Treatment: Hitting the Selective JAKpot. Gastroenterology 160 (5), 1472–1474. doi: 10.1053/j.gastro.2020.04.034
Pigneur, B., Lepage, P., Mondot, S., Schmitz, J., Goulet, O., Dore, J., et al. (2019). Mucosal Healing and Bacterial Composition in Response to Enteral Nutrition Vs Steroid-Based Induction Therapy-A Randomised Prospective Clinical Trial in Children With Crohn’s Disease. J. Crohns Colitis 13 (7), 846–855. doi: 10.1093/ecco-jcc/jjy207
Pittayanon, R., Lau, J. T., Leontiadis, G. I., Tse, F., Yuan, Y., Surette, M., et al. (2020). Differences in Gut Microbiota in Patients With vs Without Inflammatory Bowel Diseases: A Systematic Review. Gastroenterology 158 (4), 930–946.e931. doi: 10.1053/j.gastro.2019.11.294
Powell, N., Pantazi, E., Pavlidis, P., Tsakmaki, A., Li, K., Yang, F., et al. (2020). Interleukin-22 Orchestrates a Pathological Endoplasmic Reticulum Stress Response Transcriptional Programme in Colonic Epithelial Cells. Gut 69 (3), 578–590. doi: 10.1136/gutjnl-2019-318483
Praveschotinunt, P., Duraj-Thatte, A. M., Gelfat, I., Bahl, F., Chou, D. B., Joshi, N. S. (2019). Engineered E. coli Nissle 1917 for the Delivery of Matrix-Tethered Therapeutic Domains to the Gut. Nat. Commun. 10 (1), 5580. doi: 10.1038/s41467-019-13336-6
Ramakrishna, B. S. (2013). Role of the Gut Microbiota in Human Nutrition and Metabolism. J. Gastroenterol. Hepatol. 28 Suppl 4, 9–17. doi: 10.1111/jgh.12294
Rath, E., Moschetta, A., Haller, D. (2018). Mitochondrial Function - Gatekeeper of Intestinal Epithelial Cell Homeostasis. Nat. Rev. Gastroenterol. Hepatol. 15 (8), 497–516. doi: 10.1038/s41575-018-0021-x
Read, E., Curtis, M. A., Neves, J. F. (2021). The Role of Oral Bacteria in Inflammatory Bowel Disease. Nat. Rev. Gastroenterol. Hepatol. 18 (10), 731–742. doi: 10.1038/s41575-021-00488-4
Rodrigues, R. R., Shulzhenko, N., Morgun, A. (2018). Transkingdom Networks: A Systems Biology Approach to Identify Causal Members of Host-Microbiota Interactions. Methods Mol. Biol. 1849, 227–242. doi: 10.1007/978-1-4939-8728-3_15
Rolhion, N., Chassaing, B., Nahori, M. A., de Bodt, J., Moura, A., Lecuit, M., et al. (2019). A Listeria monocytogenes Bacteriocin can Target the Commensal Prevotella copri and Modulate Intestinal Infection. Cell Host Microbe 26 (5), 691–701.e695. doi: 10.1016/j.chom.2019.10.016
Roy, U., Galvez, E. J. C., Iljazovic, A., Lesker, T. R., Blazejewski, A. J., Pils, M. C., et al. (2017). Distinct Microbial Communities Trigger Colitis Development Upon Intestinal Barrier Damage via Innate or Adaptive Immune Cells. Cell Rep. 21 (4), 994–1008. doi: 10.1016/j.celrep.2017.09.097
Saha, S., Mara, K., Pardi, D. S., Khanna, S. (2021). Long-Term Safety of Fecal Microbiota Transplantation for Recurrent Clostridioides difficile Infection. Gastroenterology 160 (6), 1961–1969.e1963. doi: 10.1053/j.gastro.2021.01.010
Sartor, R. B., Wu, G. D. (2017). Roles for Intestinal Bacteria, Viruses, and Fungi in Pathogenesis of Inflammatory Bowel Diseases and Therapeutic Approaches. Gastroenterology 152 (2), 327–339.e324. doi: 10.1053/j.gastro.2016.10.012
Sassone-Corsi, M., Nuccio, S. P., Liu, H., Hernandez, D., Vu, C. T., Takahashi, A. A., et al. (2016). Microcins Mediate Competition Among Enterobacteriaceae in the Inflamed Gut. Nature 540 (7632), 280–283. doi: 10.1038/nature20557
Scaldaferri, F., Gerardi, V., Mangiola, F., Lopetuso, L. R., Pizzoferrato, M., Petito, V., et al. (2016). Role and Mechanisms of Action of Escherichia coli Nissle 1917 in the Maintenance of Remission in Ulcerative Colitis Patients: An Update. World J. Gastroenterol. 22 (24), 5505–5511. doi: 10.3748/wjg.v22.i24.5505
Schaubeck, M., Clavel, T., Calasan, J., Lagkouvardos, I., Haange, S. B., Jehmlich, N., et al. (2016). Dysbiotic Gut Microbiota Causes Transmissible Crohn’s Disease-Like Ileitis Independent of Failure in Antimicrobial Defence. Gut 65 (2), 225–237. doi: 10.1136/gutjnl-2015-309333
Schirmer, M., Garner, A., Vlamakis, H., Xavier, R. J. (2019). Microbial Genes and Pathways in Inflammatory Bowel Disease. Nat. Rev. Microbiol. 17 (8), 497–511. doi: 10.1038/s41579-019-0213-6
Seregin, S. S., Golovchenko, N., Schaf, B., Chen, J., Pudlo, N. A., Mitchell, J., et al. (2017). NLRP6 Protects Il10(-/-) Mice From Colitis by Limiting Colonization of Akkermansia muciniphila. Cell Rep. 19 (10), 2174. doi: 10.1016/j.celrep.2017.05.074
Shapiro, J. M., de Zoete, M. R., Palm, N. W., Laenen, Y., Bright, R., Mallette, M., et al. (2021). Immunoglobulin A Targets a Unique Subset of the Microbiota in Inflammatory Bowel Disease. Cell Host Microbe 29 (1), 83–93.e83. doi: 10.1016/j.chom.2020.12.003
Shawki, A., McCole, D. F. (2017). Mechanisms of Intestinal Epithelial Barrier Dysfunction by Adherent-Invasive Escherichia coli. Cell Mol. Gastroenterol. Hepatol. 3 (1), 41–50. doi: 10.1016/j.jcmgh.2016.10.004
Song, X., Sun, X., Oh, S. F., Wu, M., Zhang, Y., Zheng, W., et al. (2020). Microbial Bile Acid Metabolites Modulate Gut RORgamma(+) Regulatory T Cell Homeostasis. Nature 577 (7790), 410–415. doi: 10.1038/s41586-019-1865-0
Stappenbeck, T. S., Virgin, H. W. (2016). Accounting for Reciprocal Host-Microbiome Interactions in Experimental Science. Nature 534 (7606), 191–199. doi: 10.1038/nature18285
Stevens, B. W., Borren, N. Z., Velonias, G., Conway, G., Cleland, T., Andrews, E., et al. (2017). Vedolizumab Therapy is Associated With an Improvement in Sleep Quality and Mood in Inflammatory Bowel Diseases. Dig. Dis. Sci. 62 (1), 197–206. doi: 10.1007/s10620-016-4356-2
Subramenium, G. A., Sabui, S., Marchant, J. S., Said, H. M., Subramanian, V. S. (2019). Enterotoxigenic Escherichia coli Heat Labile Enterotoxin Inhibits Intestinal Ascorbic Acid Uptake via a cAMP-Dependent NF-kappaB-Mediated Pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 316 (1), G55–G63. doi: 10.1152/ajpgi.00259.2018
Sun, X., He, S., Lv, C., Sun, X., Wang, J., Zheng, W., et al. (2017). Analysis of Murine and Human Treg Subsets in Inflammatory Bowel Disease. Mol. Med. Rep. 16 (3), 2893–2898. doi: 10.3892/mmr.2017.6912
Tito, R. Y., Chaffron, S., Caenepeel, C., Lima-Mendez, G., Wang, J., Vieira-Silva, S., et al. (2019). Population-Level Analysis of Blastocystis Subtype Prevalence and Variation in the Human Gut Microbiota. Gut 68 (7), 1180–1189. doi: 10.1136/gutjnl-2018-316106
van der Lelie, D., Oka, A., Taghavi, S., Umeno, J., Fan, T. J., Merrell, K. E., et al. (2021). Rationally Designed Bacterial Consortia to Treat Chronic Immune-Mediated Colitis and Restore Intestinal Homeostasis. Nat. Commun. 12 (1), 3105. doi: 10.1038/s41467-021-23460-x
Vich Vila, A., Imhann, F., Collij, V., Jankipersadsing, S. A., Gurry, T., Mujagic, Z., et al. (2018). Gut Microbiota Composition and Functional Changes in Inflammatory Bowel Disease and Irritable Bowel Syndrome. Sci. Transl. Med. 10 (472). doi: 10.1126/scitranslmed.aap8914
Wellman, A. S., Metukuri, M. R., Kazgan, N., Xu, X., Xu, Q., Ren, N. S. X., et al. (2017). Intestinal Epithelial Sirtuin 1 Regulates Intestinal Inflammation During Aging in Mice by Altering the Intestinal Microbiota. Gastroenterology 153 (3), 772–786. doi: 10.1053/j.gastro.2017.05.022
Wilson, I. D., Nicholson, J. K. (2017). Gut Microbiome Interactions With Drug Metabolism, Efficacy, and Toxicity. Transl. Res. 179, 204–222. doi: 10.1016/j.trsl.2016.08.002
Yilmaz, B., Spalinger, M. R., Biedermann, L., Franc, Y., Fournier, N., Rossel, J. B., et al. (2018). The Presence of Genetic Risk Variants Within PTPN2 and PTPN22 is Associated With Intestinal Microbiota Alterations in Swiss IBD Cohort Patients. PLoS One 13 (7), e0199664. doi: 10.1371/journal.pone.0199664
Zhang, M., Sun, K., Wu, Y., Yang, Y., Tso, P., Wu, Z. (2017). Interactions Between Intestinal Microbiota and Host Immune Response in Inflammatory Bowel Disease. Front. Immunol. 8. doi: 10.3389/fimmu.2017.00942
Zhou, L., Zhang, M., Wang, Y., Dorfman, R. G., Liu, H., Yu, T., et al. (2018). Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 24 (9), 1926–1940. doi: 10.1093/ibd/izy182
Zhuang, X., Tian, Z., Feng, R., Li, M., Li, T., Zhou, G., et al. (2020). Fecal Microbiota Alterations Associated With Clinical and Endoscopic Response to Infliximab Therapy in Crohn’s Disease. Inflammation Bowel Dis. 26 (11), 1636–1647. doi: 10.1093/ibd/izaa253
Keywords: gut microbiota, inflammatory bowel disease, treatment, metabolite, IBD
Citation: Qiu P, Ishimoto T, Fu L, Zhang J, Zhang Z and Liu Y (2022) The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell. Infect. Microbiol. 12:733992. doi: 10.3389/fcimb.2022.733992
Received: 02 July 2021; Accepted: 24 January 2022;
Published: 22 February 2022.
Edited by:
Veeranoot Nissapatorn, Walailak University, ThailandReviewed by:
Maria Manuela Rosado, Sapienza University of Rome, ItalyMaria De Lourdes Pereira, University of Aveiro, Portugal
Copyright © 2022 Qiu, Ishimoto, Fu, Zhang, Zhang and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yang Liu, MTM5OTg4NDM1ODNAMTYzLmNvbQ==; orcid.org/0000-0002-2129-9086