
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 16 November 2022
Sec. Microbiome in Health and Disease
Volume 12 - 2022 | https://doi.org/10.3389/fcimb.2022.1055117
This article is part of the Research Topic The Salivary Microbiome in Health and Disease View all 7 articles
 Casper Sahl Poulsen1†
Casper Sahl Poulsen1† Nikoline Nygaard1,2†
Nikoline Nygaard1,2† Florentin Constancias3Evelina Stankevic1Timo Kern1Daniel R. Witte4,5Dorte Vistisen6,7
Florentin Constancias3Evelina Stankevic1Timo Kern1Daniel R. Witte4,5Dorte Vistisen6,7 Niels Grarup1Oluf Borbye Pedersen1,8
Niels Grarup1Oluf Borbye Pedersen1,8 Daniel Belstrøm2*Torben Hansen1*
Daniel Belstrøm2*Torben Hansen1*Introduction: Previous research indicates that the salivary microbiota may be a biomarker of oral as well as systemic disease. However, clarifying the potential bias from general health status and lifestyle-associated factors is a prerequisite of using the salivary microbiota for screening.
Materials & Methods: ADDDITION-PRO is a nationwide Danish cohort, nested within the Danish arm of the Anglo-Danish-Dutch Study of Intensive treatment in People with Screen-Detected Diabetes in Primary Care. Saliva samples from n=746 individuals from the ADDITION-PRO cohort were characterized using 16s rRNA sequencing. Alpha- and beta diversity as well as relative abundance of genera was examined in relation to general health and lifestyle-associated variables. Permutational multivariate analysis of variance (PERMANOVA) was performed on individual variables and all variables together. Classification models were created using sparse partial-least squares discriminant analysis (sPLSDA) for variables that showed statistically significant differences based on PERMANOVA analysis (p < 0.05).
Results: Glycemic status, hemoglobin-A1c (HbA1c) level, sex, smoking and weekly alcohol intake were found to be significantly associated with salivary microbial composition (individual variables PERMANOVA, p < 0.05). Collectively, these variables were associated with approximately 5.8% of the observed differences in the composition of the salivary microbiota. Smoking status was associated with 3.3% of observed difference, and smoking could be detected with good accuracy based on salivary microbial composition (AUC 0.95, correct classification rate 79.6%).
Conclusions: Glycemic status, HbA1c level, sex, smoking and weekly alcohol intake were significantly associated with the composition of the salivary microbiota. Despite smoking only being associated with 3.3% of the difference in overall salivary microbial composition, it was possible to create a model for detection of smoking status with a high correct classification rate. However, the lack of information on the oral health status of participants serves as a limitation in the present study. Further studies in other cohorts are needed to validate the external validity of these findings.
Saliva is the biological fluid of the oral cavity, which is critical to maintenance of oral health, as well as basic functions, such as speech, taste and swallowing (Pedersen et al., 2018; Lynge Pedersen and Belstrøm, 2019). In addition, saliva harbors a wealth of biological substances including DNA, RNA and proteins of both microbial and human origin, which is why saliva is referred to as the mirror of the body (Greabu et al., 2009; Tiwari, 2011). Therefore, due to the easy and non-invasive sampling, saliva is an attractive medium for biomarker analysis in both dentistry and general medicine (Yoshizawa et al., 2013).
Saliva is sterile when entering the oral cavity (Schrøder et al., 2017), and consequently the salivary microbiota is a conglomerate of bacteria shed from oral surfaces. Prime donor sites are the tongue, the throat and the palatine tonsils (Segata et al., 2012). There are also minor contributions from oral sites such as supragingival and subgingival plaques, which have direct relevance to the major oral diseases, periodontitis and dental caries (Segata et al., 2012). Several studies have shown that salivary levels of specific pathogens associated with periodontitis, such as Porphyromonas gingivalis, Tannerella forsythia, Treponema denticola and Prevotella intermedia correlate with that of subgingival plaque found in the periodontitis lesion (He et al., 2012; Haririan et al., 2014; Belstrøm et al., 2017; Nickles et al., 2017; Belstrøm et al., 2018). In addition, it has been documented that salivary abundance of specific pathogens, including P. gingivalis, associates with periodontal status (Morozumi et al., 2016; Kageyama et al., 2017; Damgaard et al., 2019). Consequently, salivary abundance of specific bacterial species has been suggested as a diagnostic biomarker of periodontitis, with the potential to be used in the dental office. However, a prerequisite of using the salivary microbiota for screening of oral diseases is to clarify the potential bias from general health status and life-style associated factors, which inevitably are different among patients.
Type 2 diabetes (T2D) and cardiovascular diseases are some of the most prevalent general medical disorders, and tobacco smoking is a common habit (World Health Organization, 2021a; World Health Organization, 2021b; World Health Organization, 2022). Cross-sectional data suggests that the salivary microbiota in patients with T2D is less diverse and harbors significantly higher levels of periodontal pathogens (Gogeneni et al., 2015; Ogawa et al., 2017; Saeb et al., 2019). Likewise, smoking has been described to have an impact on the salivary microbiota (Belstrøm et al., 2014; Wirth et al., 2020; Belstrøm et al., 2021). The salivary microbiota have even been found to predict the risk of cardiovascular disease with some success (Murugesan et al., 2021). In general, current evidence suggests that the salivary microbiota may not only be a diagnostic biomarker of oral diseases, but potentially also of general medical disorders. Whether these associations are causal or due to an indirect confounded association is currently not clear, and available data originates primarily from studies with small sample sizes in selected populations. Studies with a large sample size and substantial information on general medical health and lifestyle-associated factors are therefore needed, as they will allow for a more detailed assessment of associations and provide better options to account for confounding.
The ADDITION-PRO cohort is a prospective cohort nested within the Danish arm (ADDITION-DK) of the Anglo-Danish-Dutch Study of Intensive Treatment In People with Screen-Detected Diabetes in Primary Care study (ADDITION-Europe) (Johansen et al., 2012). ADDITION-PRO consists of 2082 Danes from across the country, at varying risk of developing T2D measured by their fasting blood glucose and glucose tolerance. Participants underwent a comprehensive health assessment at inclusion, resulting in an extensive dataset with measurements of anthropometry, biochemistry, physical fitness, cardiovascular risk factors and questionnaire data. In addition, blood, urine and saliva samples were collected from the participants. Given the considerable size of the population and its comprehensive characterization the ADDITION-PRO cohort makes for a highly relevant source of information in determining the association of general health status with lifestyle-associated factors on the composition of the salivary microbiota.
Therefore, the purpose of the present study was to evaluate the association of general medical disorders and lifestyle-associated factors with the composition of the salivary microbiota. To do this, we used 16S rRNA sequencing to characterize the salivary microbiota in N = 786 individuals from the ADDITION-PRO cohort, from which we also had access to comprehensive information on general medical health and lifestyle-associated factors. We tested the hypothesis that general medical health and lifestyle are significantly associated with the composition of the salivary microbiota, and that general health status and lifestyle can be predicted based on the salivary microbiota.
The ADDITION-PRO cohort has been described in detail elsewhere (Johansen et al., 2012). In brief, it is a cohort nested within the Danish arm (ADDITION-DK) of the ADDITION-Europe study, consisting of individuals at differing risks of T2D. Individuals were recruited for ADDITION-PRO between 2009 and 2011 during follow-up health examinations of a subset of the population from ADDITION-DK. Eligible for the follow-up health examination were those who were still alive, and still lived in the regions of the four participating research centers (Steno Diabetes Center, Aarhus University Hospital, Holstebro Hospital, and Hospital of South West Jutland). Lastly, eligibility required not having withdrawn consent to study participation.
Of the 16,136 participants in the follow-up health examination, all individuals with impaired glucose regulation at the time of screening, along with those diagnosed with diabetes during the follow-up period (n=1483), along with a 19% random sample of individuals from the low and high-risk groups (n=2705) were invited to participate in ADDITION-PRO. In total, 50% (n=2082) of invitees agreed to participate (Johansen et al., 2012). Saliva samples were collected only at the Copenhagen site, resulting in 786 saliva samples to be analyzed in the present study.
The ADDITION-PRO cohort was approved by the scientific ethics committee in the Central Denmark Region (H-20000183). Participants gave their written informed consent to participate in the study and for linkage of their data with national registers for the purposes of the ADDITION-PRO study. In addition, the present study was performed in compliance with the Helsinki Declaration.
The health assessment performed is described in detail elsewhere (Johansen et al., 2012). In brief, information on sex, age, and lifestyle-associated factors (smoking, alcohol consumption and physical activity) were collected via a general health questionnaire. Clinical measurements of height, weight, and waist circumference were performed, and BMI was calculated based on height and weight measurements. Blood pressure (systolic and diastolic) and heart rate was calculated as the average of three measures after a 10-minute rest (Omron M6 comfort, Omron Healthcare, Milton Keynes, UK).
The biochemical analyses have been described in detail elsewhere (Johansen et al., 2012). Briefly, biochemical measures were analyzed at the Clinical Chemistry Department at the Steno Diabetes Center in Gentofte, Denmark. HbA1c was measured by HPLC (TOSOH G7, Tokyo, Japan). Total cholesterol, HDL-cholesterol, LDL-cholesterol, triglycerides, were measured using the Hitachi 912 system (Roche Diagnostics, Mannheim, Germany).
Classification of the different variables of interest was done based on either internationally acknowledged classifications from the World Health Organization on BMI, HbA1c and physical activity (The Global Diabetes Community, 2019; World Health Organization, 2020; World Health Organization, 2021a), the American Heart Association on blood pressure (American Heart Association, 2022), or recommendations from the Danish Health Authority for weekly alcohol intake (The Danish Health Authority, 2022).
Upon further inspection, 40 individuals were missing significant amounts of data from biochemical analyses, questionnaires etc., and were therefore excluded, resulting in a final dataset of 746 individuals.
Venous blood samples were drawn after an overnight fast of more than 8 hours. Individuals without known diabetes diagnosis received a standard 75g oral glucose tolerance test with blood samples drawn at 0, 30 and 120 minutes. Further details on the storage and processing of blood samples can be found elsewhere (Johansen et al., 2012).
Paraffin-stimulated saliva samples were collected following a standardized protocol, as previously described (Johansen et al., 2012). Participants were instructed not to consume any food or beverages in the 8 hours prior to sample collection, and not to brush teeth on the morning of collection. All samples were stored at -80○C until further analysis (Johansen et al., 2012).
Core probes (200 mg) from the frozen samples were taken prior to bacterial DNA isolation. A CryoXtract CXT353 instrument operating below -120°C was used to preserve the sample integrity. Bacterial DNA was isolated, using the NucleoSpinSoil kit (Macherey-Nagel, Germany), following the manufacturer’s instruction. Bacterial cells were lysed using SL1 + Enhancer buffer SX. DNA yield and purity were assessed using a Qubit 2.0 flourometer, and a NanoDrop 2000 spectrometer (Thermo Fisher Scientific Inc., MA, USA). Genomic DNA in each sample was normalized to 30ng prior PCR amplification of the V4 hypervariable region using the 515F/806R primer set. The PCR products were purified with the AmpureXP 17 magnetic bead-based clean-up and size selection kit. The final library was quantified by determining the average molecule length using the 2100 bioanalyzer instrument (Agilent, DNA 1000 Reagents), and by library using real-time quantitative PCR (EvaGreenTM). All samples were processed consecutively at the same location and by the same equipment. Owing to the sample number, sequencing was carried out in two runs using paired-end 250 (PE251+8+8+251) sequencing chemistry on an Illumina HiSeq 2500 platform.
Raw sequencing data were processed with the dada2 (v.1.19.1) (Holmes et al., 2017) algorithm as implemented in metabaRpipe (Constancias and Mahé, 2022). Reads were first truncated after 170 for forward reads, and after 160 for reverse reads. Reads with expected error rates higher than 3 and 4 for forward and reverse reads were subsequently removed. After filtering, error rate learning was performed on all the reads (run_dada2_pipe (nbases = 10 11)), and ASV inference was run using pseudo-pooling strategy. Forward and reverse reads were then merged with a minimum overlap of 40 nucleotides (run_dada2_pipe (minover = 40)). As error rate profile was run dependent, the previous steps were performed independently on samples included in each of the 2 HiSeq runs. ASV tables were then merged and chimeric sequences were identified and removed. A final clustering step was performed to detect potential HiSeq run specific artefacts (run_dada2_pipe(collapseNoMis = TRUE)) and no ASV were clustered confirming the high sensitivity of the approach. Taxonomy was assigned to ASVs using dada2 against eHOMD database (V15.22) (eHOMD, 2020),dada2::assignTaxonomy() and dada2::assignSpecies() (Constancias, 2022) as implemented in the funci phyloseq_dada2_tax(), and a phylogenetic of ASV sequences was build using add_phylogeny_to_phyloseq().
All data analysis was performed using R statistical software (v4.2.1) (R Core Team, 2020). The script was created as an R notebook and made into a pdf file using Rmarkdown, where all function calls are available for the whole analysis (Supplementary file 1 and https://github.com/csapou/Additionpro_Poulsenetal2022). Sample variables were compared using chi-square tests and the -log10 transformed p-values with an upper limit of 3 were visualized using pheatmap 1.0.12 (Kolde, 2019) to investigate association patterns between the variables. Data were aggregated to genus level in the presented results. Study variables were not available for all individuals, therefore based on the variable of interest, filtering was done to remove these individuals separately for each analysis. Hence, a complete case analyses approach was used.
Alpha diversity including richness and Shannon diversity was performed using the diversity function in the package vegan 2.6.2 (Oksanen et al., 2022) and compared both using parametric tests (ANOVA and t-test if binary) and non-parametrically tests (Kruskal-Wallis and Mann-Whitney if binary). The beta diversity metric used were Bray-Curtis dissimilarities calculated from Hellinger transformed total sum scaled data and used to perform PERMANOVA and principal coordinate analysis (PCoA). The NA individuals were still visualized in the PCoAs
In order to choose a set of variables of interest, PERMANOVAs were run with individual variables as outcome, and significant variables (smoking, alcohol consumption, glycemic status, sex and HbA1c level) are presented in the present paper. In addition, we chose to present a variable that did not have a statistically significant association in the PERMANOVA in the present paper, using the variable physical activity, as it had the least associations to the other variables (Figure 1). PERMANOVA was also run with all variables (consequently on fewer individuals since complete variable availability is a prerequisite), and with just the 5 selected variables found to be statistically significant in the PERMANOVA run, with the addition of the variable physical activity.
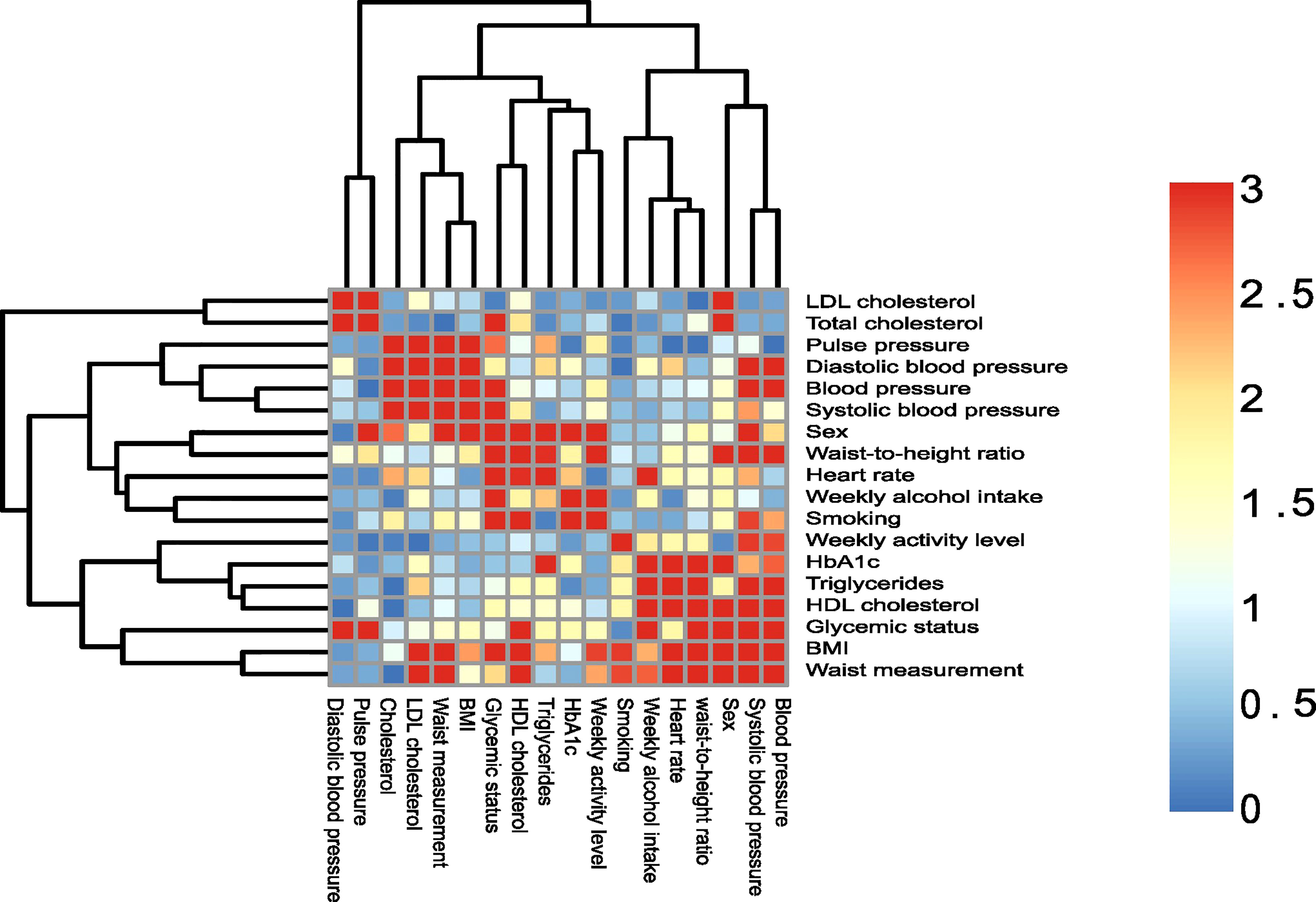
Figure 1 Association of sample variables. Heatmap of sample variables compared by chi-squared tests. The p-values are visualized on a log scale (-log10(pvalue)). A value of 1.30 represent a 0.05 significance cut-off, and values above 1.30 are considered statistically significant.
Performance of using different types of differential abundance tests was assessed with DAtest 2.8.0. Data were subset to only include taxa present in more than 20% of samples. Non-parametrical tests (Kruskal-Wallis and Mann-Whitney if binary) and DESeq2 (likelihood ratio test (LRT) and Wald test if binary) using relative log values are enclosed in the present study to be used as a reference (Supplementary files 2-3).
Stacked bar charts, horizontal relative abundance violin plots of individual genera, and horizontal bar charts were generated with rabuplot 0.0.1.04 (Stockholm, 2022). The p-values presented in the horizontal relative abundance violin plots were obtained from the non-parametric tests. Both the most abundant genera and significant differential abundant genera were presented. A p-value of less than 0.05 were considered statistically significant. All adjusted p-values presented are corrected by the Benjamini-Hochberg method.
To assess the classification potential of the salivary microbiota to differentiate the variable groups the mixOmics 6.20.0 (Rohart et al., 2017) implementation of Sparse partial least square discriminant analysis (sPLS-DA) with Hellinger transformed total sum scaled data were run on the selected variables from the PERMANOVA, but also important features for discrimination are made available (Supplementary file 4). Discrimination was measured by area under the curve (AUC) and correct classification rate.
Table 1 details the study population. Study participants were on average 68 years of age, and 55% were males. The majority of the study population (56%) was classified as normoglycemic. The majority of study participants (66%) were either overweight or obese, with the average blood pressure as well as the average HbA1c measure being within the normal range. About half of participants (47%) classified themselves as ex-smokers, while less than 20% reported being currents smokers. More than half (61%) reported spending more than one hour on moderate to vigorous activity daily and drinking on average 11 units of alcohol per week. To test whether the variables of interest correlated with each other, a heatmap of variable association was performed (Figure 1). As can be seen, many of the variables presented were correlated with each other.

Table 1 Population characteristics, N (%) and mean (SD).
Saliva samples were obtained from 786 individuals and sequenced in two runs (R1, n = 666 and R2, n = 120) resulting in a total of 33,448,846 reads (mean=42,556, sd=22,408). After removal of chimeras, a total of 20,669,468 reads (mean=26,972, sd=14,336) were left, resulting in 5,748 unique amplicon sequence variants (ASVs). Differences between sequencing runs were assessed by PCoA and PERMANOVA. Three samples were included on both runs and differed marginally (Bray-Curtis dissimilarity: 0.010, 0.011 and 0.014) as also visualized in the PCoA (Supplementary Figure 1). From the PERMANOVA, no difference was observed between sequencing runs (p=0.45). Thus, the two datasets were merged for analysis.
The core salivary microbiota consisted of 26 genera present in >95% of all samples, which accounted for 94.2% (range [45.4%; 99.8%]) of reads from each participant. The dominant genera were Veillonella, Streptococcus, Haemophilus and Prevotella, which represented 56.7% of the microbiota. In general, the composition of the predominant genera was not influenced by the variables tested (Figure 2, Supplementary Figure 2).
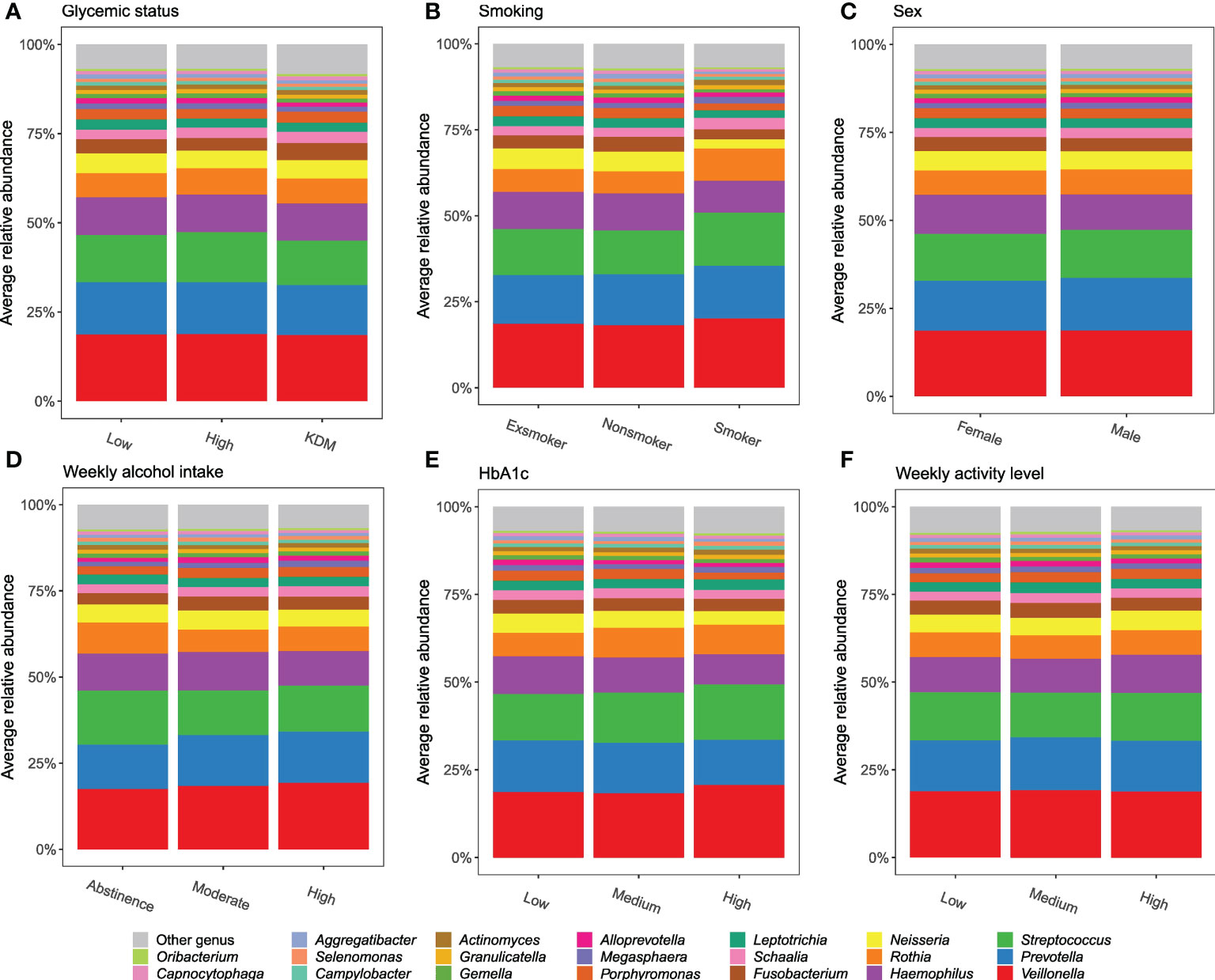
Figure 2 Predominant genera. Stacked bar charts of the 20 most abundant genera expressed as mean relative abundance. (A–F): glycemic status, smoking, sex, weekly alcohol intake, HbA1c, and weekly activity level.
Examining the alpha diversity between groups within the different variables in ADDITION-PRO, we found statistically significant differences in Richness for glycemic status, HbA1c, LDL cholesterol, total cholesterol and weekly alcohol intake, and in Shannon index for smoking, weekly alcohol intake and LDL cholesterol (Table 2, see Supplementary Figure 3.1, 3.2 for violin plots with ANOVA). To assess beta diversity, PCoA plots were used to visualize the data in a two-dimensional space representing most variation in the data, and plots were generated and colored according to each of the variables. No clear separation was observed, based on any of the variables included (Figure 3 and Supplementary Figure 4). Using PERMANOVA on each variable individually, the variables glycemic status, HbA1c, sex, smoking and weekly alcohol intake were found to have a significant association with salivary microbial composition (PERMANOVA unadjusted p < 0.05). In a PERMANOVA run with all variables included, only smoking and weekly alcohol intake remained statistically significant (Table 2) and accounted for R2 = 0.030 (3%) and R2 = 0.006 (0.6%) of the observed sample variance in salivary microbial composition, respectively (Table 2). Running the model with the variables in a continuous instead of categorical form yielded similar results, both in terms of variables included and the magnitude of their contribution to microbial composition, though with the addition of BMI and average waist measurement, and the exclusion of Glycemic status and HbA1c, as significant factors (Supplementary file 5).
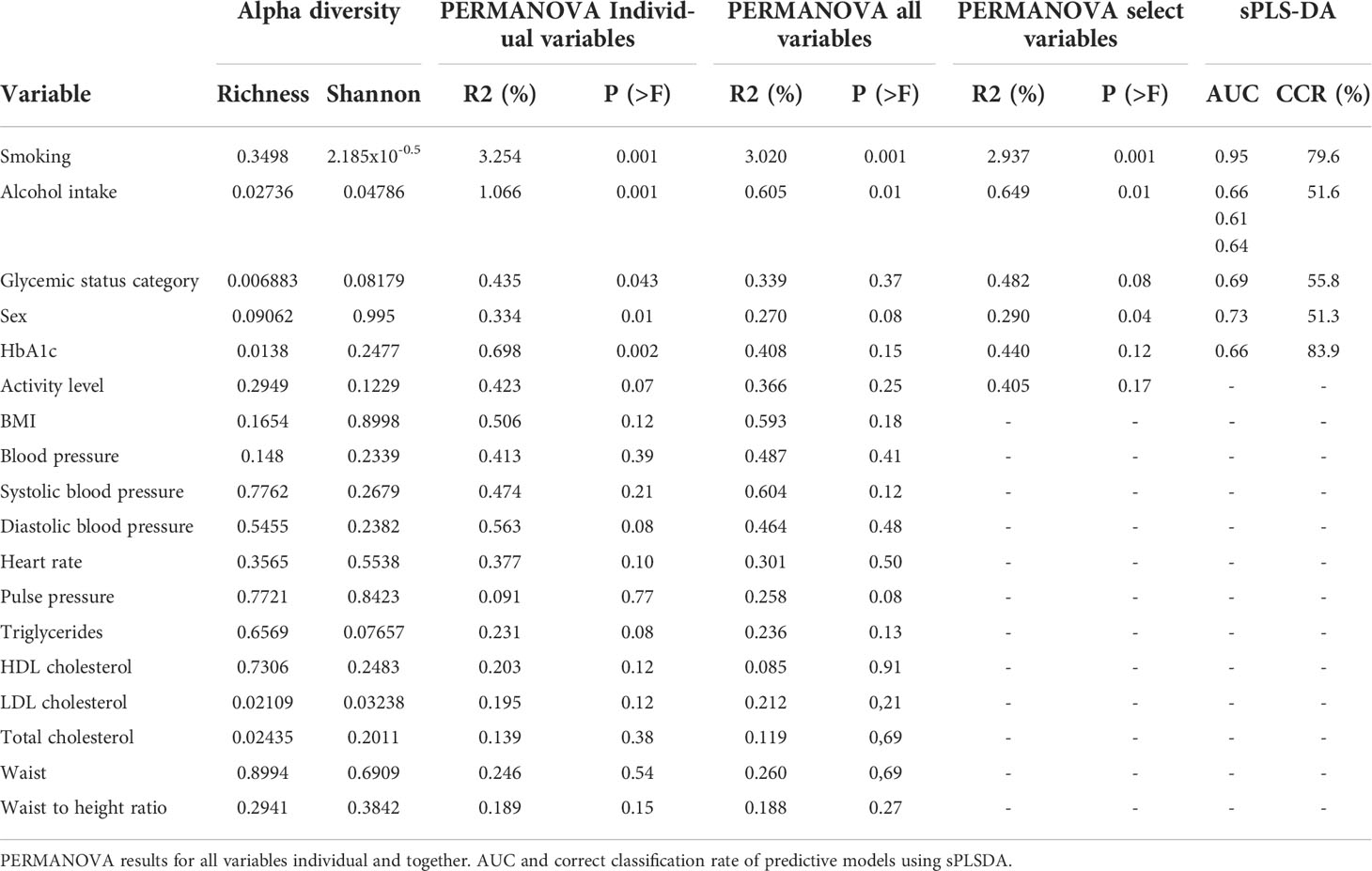
Table 2 Alpha diversity by Shannon index and Richness.
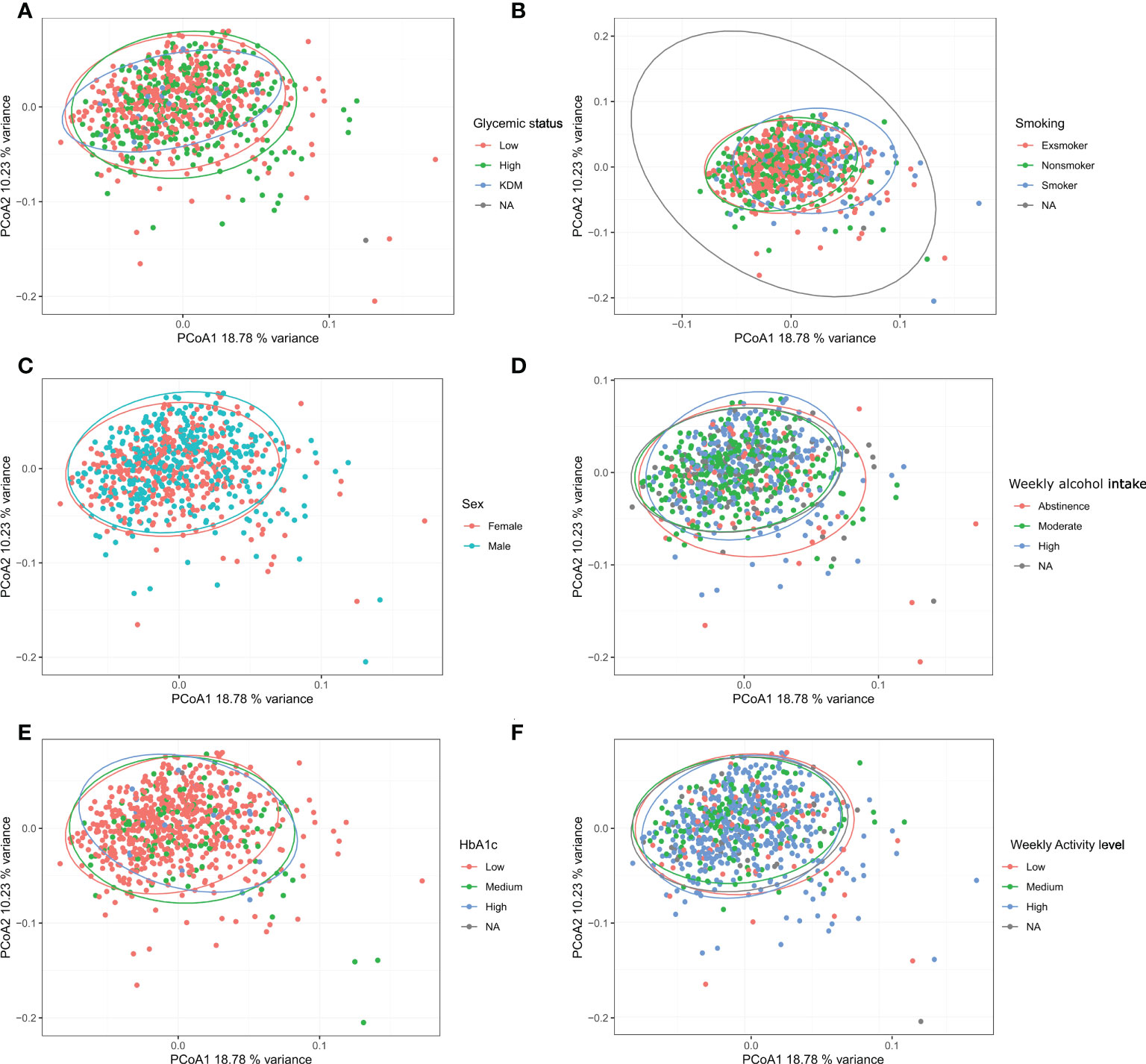
Figure 3 Beta diversity: Bray-Curtis dissimilarity calculated from Hellinger transformed total sum scaled data was used as beta-diversity measure and visualized with principal coordinate analysis (PCoA). (A–F): glycemic status, smoking, sex, weekly alcohol intake, HbA1c, and weekly activity level.
More than unique 40 genera were found to appear in significantly different abundances (p < 0.05), when examining differential abundance in groups within the variables found significant in the PERMANOVA runs (sex, glycemic status, HbA1c, weekly alcohol intake and smoking) (Figure 4, Supplementary Figure 5). Although significant, only minor differences in relative abundance were observed. Smoking status and sex were found to be associated with a substantially higher number of differentially abundant bacterial genera, as compared to the other variables. Genus Streptococcus was most frequently found to have a statistically significant differential abundance, and was associated with four out of the five variables of interest, namely smoking, weekly alcohol intake, HbA1c levels and glycemic status. Overall, Streptococcus was associated with high-risk groups, with the exception of variable alcohol intake, where the genus appeared in the highest abundance in those who abstained from alcohol. Current smoking was significantly associated with lower abundance of Neisseria, and higher abundance of Rothia species compared to former-smokers and those who had never smoked.
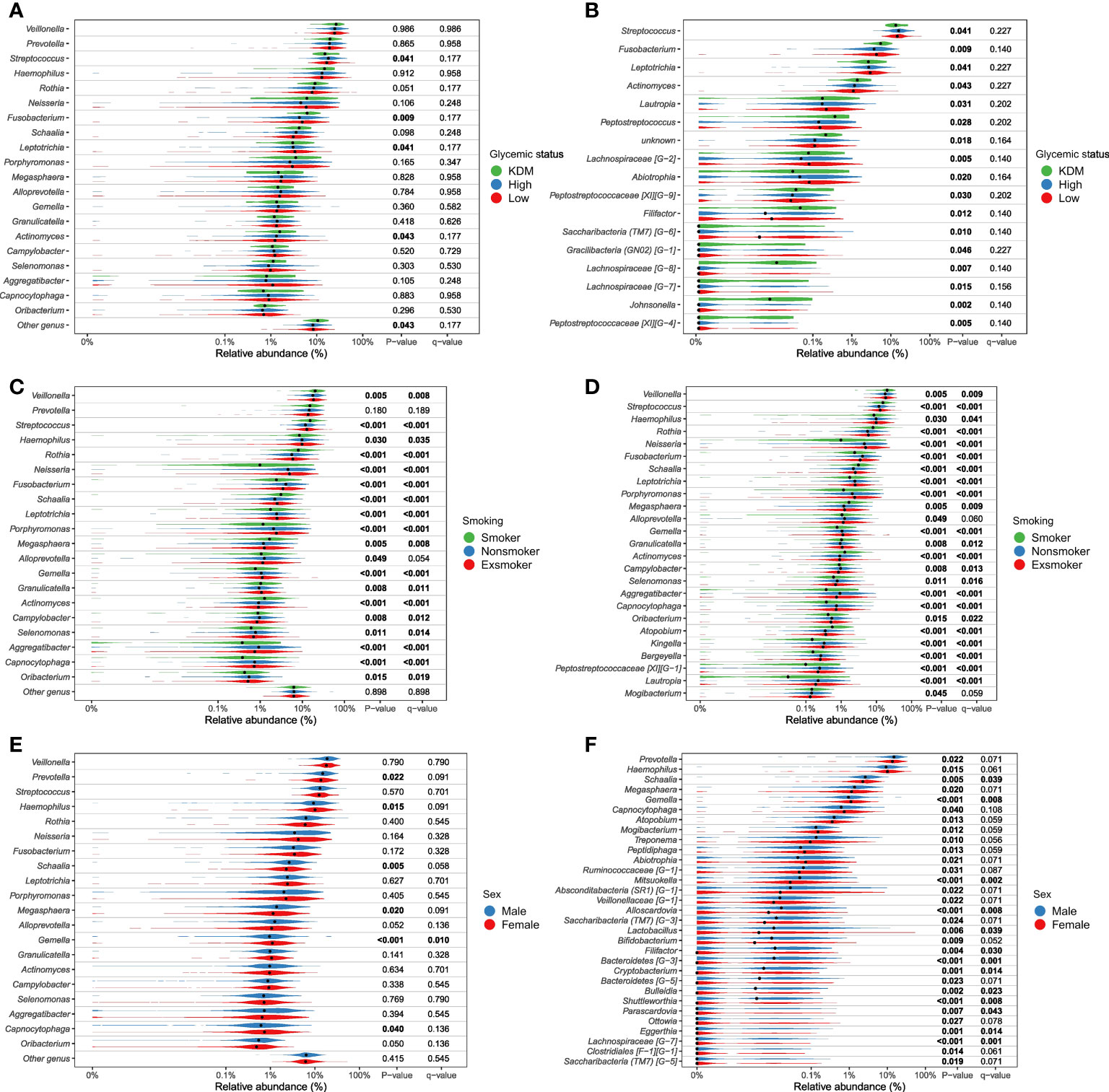
Figure 4.1 Genera significantly associated with glycemic status, smoking and sex. Relative abundance of most abundant genera (panels A, C, E) and differential abundant genera (panels B, D, F). Comparison of groups were performed with non-parametric tests (Kruskal-Wallis). Unadjusted p-values as well as Benjamini-Hochberg corrected q-values are included in the plot. (A, B), glycemic status, (C, D), smoking status, (E, F), sex. All tests are included in Supplementary File 2 as well as a parametric approach (DESeq as implemented in DAtest) in Supplementary file 3).
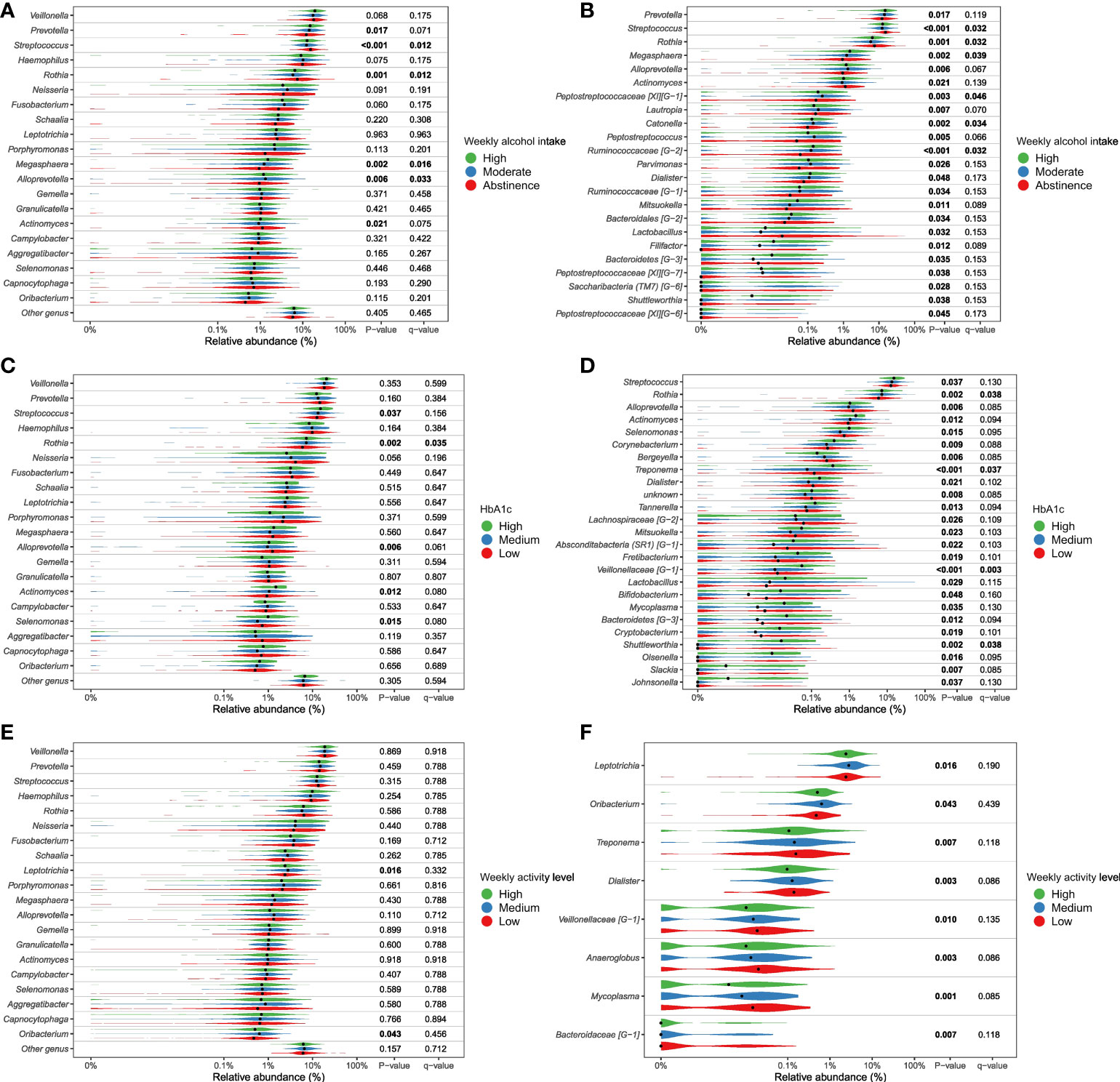
Figure 4.2 Genera significantly associated with alcohol intake, Hb1Ac and physical activity. Relative abundance of most abundant genera (panels A, C, E) and differential abundant genera (panels B, D, F). Comparison of groups were performed with non-parametric tests (Kruskal-Wallis). Unadjusted p-values as well as Benjamini-Hochberg corrected q-values are included in the plot. (A, B) weekly alcohol intake, (C, D) Hb1Ac, (E, F) weekly activity level. All tests are included in Supplementary File 2 as well as a parametric approach (DESeq as implemented in DAtest) in Supplementary File 3).
sPLSDA, was used both to investigate classification potential of the salivary microbiota and to investigate important features, and see if they correspond to the findings from the differential abundance analysis. sPLSDA was performed using the 5 variables found significant in the PERMANOVA (Figure 5 and Table 2). The classifier performed well in differentiating between smokers and non-smokers with an AUC=0.95. A relatively high correct classification rate of 79.6% was obtained, when validating the classifier with a test set. The model performed almost equally with guessing, obtaining correct classification rates of approx. 50-60%, with regards to sex, glycemic status and weekly alcohol intake. It was not possible to construct a model of HbA1c due to unbalanced group sizes and classifying almost all samples as a specific group.
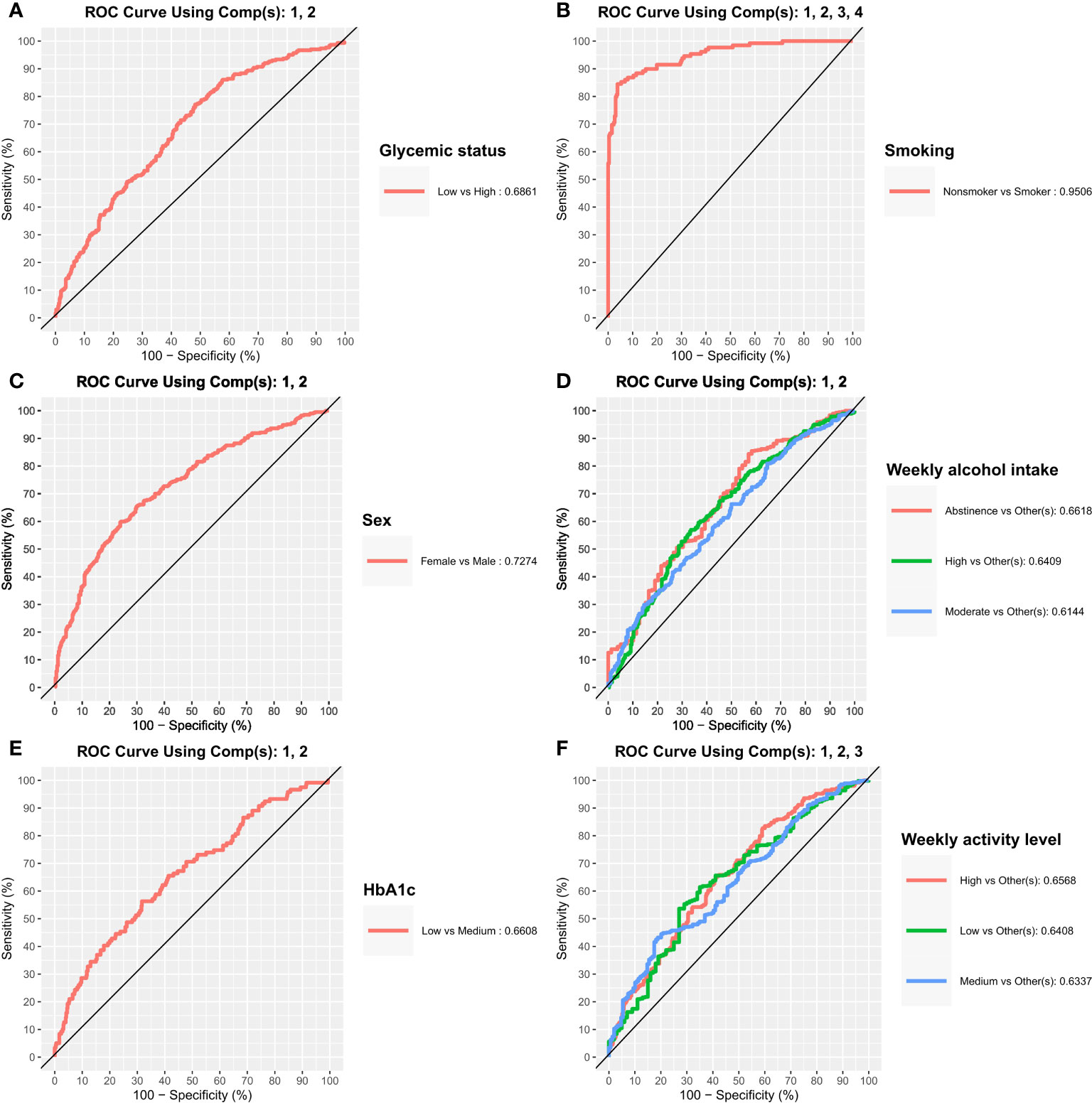
Figure 5 Classification potential of the salivary microbiota. Receiver Operating Characteristics (ROC) curves from sparse partial least squares discriminant analysis where used to assess classification of the different groups. Area under the curve (AUC) were used to quantify the performance of the classifier. (A–F) glycemic status, smoking, sex, weekly alcohol intake, HbA1c, and weekly activity level.
The purpose of the present study was to evaluate the association of general medical disorders and lifestyle-associated factors with the composition of the salivary microbiota. We tested the hypothesis that the composition of the salivary microbiota associates with both general health status and lifestyle.
The main finding was that the variables (sex, glycemic status, HbA1c levels, smoking and weekly alcohol intake), which in the present study were identified to have significant associations with the composition of the salivary microbiota, altogether accounted for only 5.8% of the observed differences in salivary microbial composition in the PERMANOVA run of individual variables (Table 2). In addition, none of the tested variables had a clear association with the variation of the data, as evaluated by clustering of samples in the PCoA plot (Figure 3). Thus, our data suggests that general health status and lifestyle-associated factors are not the main determinants of the composition of the salivary microbiota.
As expected, smoking was associated with characteristics of the salivary microbiota. Specifically, higher abundance of specific genera, such as Veillonella, Streptococcus and Rothia, and significantly lower abundance of Neisseria, Haemophilus, Pophyromonas and Actinomyces in smokers compared to ex-smokers and never-smokers, was observed (Figure 4.1C, D). In general, these findings are in line with previous studies, which have also identified taxonomic differences in the salivary microbiota in smokers versus non-smokers in different populations, with varying sample size (Wu et al., 2016; Rodríguez-Rabassa et al., 2018; Suzuki et al., 2022; Wang et al., 2022). Thus, our data confirms that smoking to some degree associates with characteristics of the salivary microbiota. However, the PERMANOVA analysis revealed that smoking status accounted for as little as 3.3% of the observed differences between individuals in the PERMANOVA run of individual variables, and 2.9% in the PERMANOVA run of select variables together (Table 2). Data from the present study suggest only a minor association of smoking and the composition of the salivary microbiota, which is different from the subgingival microbiota, which has been shown to strongly associate with smoking status. Nevertheless, it is noteworthy that it was still possible to build a good classifier based on the salivary microbiota to differentiate between current smokers and current non-smokers in the present study. Indeed, we are not the first to do so, as in 2022, another study used data on the salivary microbiota from 175 current smokers and 1070 ex- and non-smokers to create a model that achieved an AUC of 0.81 (Díez López et al., 2022), which is comparable with our findings (Table 2). Importantly, when considering that smoking, which accounted for only 3.2% of differences in our dataset, can deliver such a robust model, this reinforces the concept of using the salivary microbiota for identification of oral diseases, as periodontitis has been shown to have a much higher impact on the composition of the salivary microbiota, than smoking (Belstrøm et al., 2021).
Second to smoking, weekly alcohol intake was the variable with the greatest association with salivary microbial composition. Specifically, in the individual PERMANOVA analysis, weekly alcohol intake accounted for 1% of the observed difference in the composition of the salivary microbiota, which decreased to 0.6%, when including all variables in the same model (Table 2). The considerable decrease in contribution of weekly alcohol intake to salivary microbial composition between the two PERMANOVA runs was most likely the consequence of the high degree of correlation between smoking status and weekly alcohol intake, as seen in Figure 1. As such, confounding by smoking likely yields an artificially high estimate of the effect of weekly alcohol intake in unadjusted analyses. Likewise, it is likely that there are many other confounding pathways, such as socio-economic status and diet, which could be linked with both a higher alcohol intake and differences in the salivary microbiota. It should be noted that our measure of alcohol intake was self-reported and is therefore potentially subject to information bias (social desirability and recall bias). Nevertheless, we found genus Prevotella to significantly associate with alcohol intake, as evaluated by almost 2% higher relative abundance in individuals with high intake versus low intake of alcohol (Figure 4.2A, B). This result is in line with findings from previous reports (Fan et al., 2018; Liao et al., 2022). On the other hand, our finding of a lower abundance of Streptococcus in individuals with high alcohol intake is in conflict with previous studies, which finds Streptococcus to be higher in individuals with high alcohol intake (Fan et al., 2018; Barb et al., 2022). Taken together, our data suggest that alcohol intake may only cause relatively minor perturbations to the salivary microbiota. However, it should be noted that the majority of the study subjects (40%) display moderate drinking habits, with only a minority of the 35% in the category of heavy drinkers drinking more than 20 units of alcohol per week. As such, it is not unlikely that a high alcohol intake may associate with salivary microbial composition, but that our data does not show it, as our population is skewed towards drinking less.
Glycemic status and HbA1c were both found to have a significant association with salivary microbial composition, albeit a very small one (Table 2). Quite a few studies, most with a much smaller sample size than the present study, have previously examined the association of T2D with the oral microbiota. In general, previous data have been conflicting, where some studies find higher bacterial diversity (Casarin et al., 2013; Ganesan et al., 2017; Chen et al., 2020), and others lower in individuals with T2D (Ogawa et al., 2017; Longo et al., 2018; Saeb et al., 2019; Matsha et al., 2020; Yang et al., 2020; Sabharwal et al., 2021). In addition, several studies have reported no significant difference of the oral microbiota in people with diabetes and healthy controls (Kampoo et al., 2014; Janem et al., 2017; Sun et al., 2020; Almeida-Santos et al., 2021; Liu et al., 2021). It is therefore noteworthy that data from the present study clearly shows that diabetes, as evaluated by glycemic status and HbA1c level, only account for 0.4% and 0.3% of the observed sample differences, respectively (Table 2), and that these variables had a minimal influence on relative abundance of predominant genera (Figure 2) and variation of the data (Figure 3). Moreover, when running the PERMANOVA with all variables, no significant association between glycemic status and HbA1c, was observed (Table 2). We therefore speculate that the minimal effect of glycemic status and HbA1c on salivary microbial composition observed in the individual PERMANOVA runs, were merely the consequence that glycemic status and HbA1c were highly correlated with smoking status (Figure 1). Thus, data shows a minimal direct association of glycemic status and HbA1c level with the composition on the salivary microbiota, suggesting that the salivary microbiota is probably not a good biomarker candidate for classification of diabetes. This finding is in contrast to a recent study in 133 individuals with and without periodontitis and/or diabetes, using oral microbial composition to create models, which classified groups with AUCs between 96.3% and 100% (Sun et al., 2020). One possible explanation to this discrepancy may be that our model was only constructed to classify glycemic status groups or HbA1c levels, as compared to the other study, where the model distinguished individuals with different glycemic states combined with different periodontal status. Indeed, periodontitis is associated with significant changes in oral microbial composition (Sun et al., 2020; Ko et al., 2020), which may have contributed a stronger predictive ability, than we achieved in the present study using diabetic parameters alone. Taken together, there is a need for well-constructed studies with even larger sample sizes in order to determine, whether glycemic status is independently associated with the salivary microbiota, or merely the consequence of the association of periodontitis and T2D.
In addition to the aforementioned variables, sex showed a statistically significant association with salivary microbial composition, as evaluated by PERMANOVA (p < 0.05). However, sex accounted for just 0.3% of the observed differences in salivary microbial composition in this study sample (Table 2), and had minimal influence on relative abundance of predominant genera (Figure 2) and clustering of data (Figure 3). Moreover, by use of PERMANOVA we did not find any significant associations of anthropometric factors such as BMI, waist circumference, waist- to-hip ratio, blood pressure and dyslipidemia with the composition of the salivary microbiota. Previous studies have reported age (LaMonte et al., 2019; Chaudhari et al., 2020; Liu et al., 2020; Nearing et al., 2020), sex (Fischer et al., 2008; Nearing et al., 2020; Minty et al., 2021; Zhao et al., 2021) and anthropometric factors (Takeshita et al., 2016; Si et al., 2017; Tam et al., 2018; Fei et al., 2019; Yang et al., 2019; Nearing et al., 2020; LaMonte et al., 2022) to associate with characteristics of the oral microbiota. These findings are in line with differential abundance data from the present study, which identifies relative abundance of a number of bacterial genera to associate with each of the variables (Figures 4.1, 4.2; Supplementary Figure 5). However, the small magnitude of the differences observed, questions the clinical relevance of these findings.
The main limitation of the present study is that the oral health status was not recorded, as part of data collection in ADDITION-PRO. Indeed, periodontitis associates with both T2D and cardiovascular disease (Holmstrup et al., 2017), through systemic low-grade inflammation (Hajishengallis and Chavakis, 2021), and smoking is a risk factor of periodontitis (Rivera-Hidalgo, 2000). Moreover, based on epidemiological data from the US (Eke et al., 2020) we should expect at least 40% of the participants to have some degree of periodontitis, since the mean age in the ADDITION-PRO cohort is 68 years (Table 1). Likewise, epidemiological data from China suggest that as much as 60% of the cohort could have treatment requiring dental caries (Liu et al., 2013). Importantly, periodontitis and dental caries has been shown to associate with characteristics of the salivary microbiota (Belstrøm, 2020). In the same vein data on the salivary flow rate of participants were not collected, despite it being an important risk factor for oral disease (Dodds et al., 2015; Vallabhan et al., 2020). Thus, it is almost certain that the potentially heterogeneous oral health status of the ADDITION-PRO cohort may have confounded the observed associations. Another limitation was the selection process of ADDITION-PRO, which involved progressive diabetes screening steps, resulting in a population of largely middle-aged and predominantly sedentary individuals, which are not representative of the background population, and therefore hampers the external validity of the data presented. The main strength of the present study is the relatively large sample size, in combination with detailed information on a large number of highly relevant variables. Accordingly, this provided a hitherto unprecedented opportunity to characterize the association between multiple dependent variables and the composition of the salivary microbiota in one single cohort. In the Supplementary files 2-3, we offer an overview of all genera found to be significant for all variables examined, for comparison of data in future studies of salivary microbial composition.
In conclusion, data from the present study shows a modest association of general health and lifestyle-associated parameters with the composition of the salivary microbiota. However, it should be emphasized that our data on explained variance estimates are highly sample specific and not estimates of population explained variance. Consequently, data from the present study should be validated in independent cohorts, preferably with the inclusion of data on the oral health status of study participants, which was not available in the present study. Despite smoking accounting for only 3% of the difference observed, it was possible to create a model for classification of smoking status, with a correct classification rate of 79.6%. Given the fact that presence of manifest periodontitis and dental caries have much stronger associations with the composition of the salivary microbiota than smoking, further studies are warranted to determine, whether the salivary microbiota can be used as a diagnostic biomarker of periodontitis and dental caries at preclinical stages.
The data for this study have been deposited in the European Nucleotide Archive (ENA) at EMBL-EBI under accession number PRJEB57196 (https://www.ebi.ac.uk/ena/browser/view/PRJEB57196).
The studies involving human participants were reviewed and approved by The scientific ethics committee in the Central Denmark Region (case ID: H-20000183). The patients/participants provided their written informed consent to participate in this study.
Data collection and curation: DW, OP, and TH. Data pre-processing, statistics, and analysis: CS, TK, FC, and NN. Conceptualisation: TK, DB, NN, CS, and TH. Writing: NN, DB, and CS. Editing: NN, DB, CS, ES, DW, OP, TH, and FC. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2022.1055117/full#supplementary-material
Supplementary Figure 1 | Comparison of sequencing batches. Bray-Curtis dissimilarity calculated from Hellinger transformed total sum scaled data was used as beta-diversity measure and visualized with principal coordinate analysis (PCoA).
Supplementary Figure 2 | Predominant genera. Stacked bar charts of the 20 most abundant genera aggregated for variables that was not significant by PERMANOVA analysis. The variables include BMI, waist measurement, waist-to-height ratio, blood pressure, systolic blood pressure, diastolic blood pressure, heart rate, pulse pressure, triglyceride levels, HDL cholesterol, LDL cholesterol and total cholesterol.
Supplementary Figure 3.1 | Alpha diversity. Comparison of alpha diversity between selected samples variable groups by richness (panels A, C, E, G, I and K) and Shannon diversity index (panels B, D, F, H, J and L), with ANOVA. Selected variables include weekly alcohol intake, HbA1c, and weekly activity level depicted in panel (A–L) respectively.
Supplementary Figure 3.2 | Alpha diversity. Comparison of alpha diversity between additional samples variable groups by richness (panels A, C, E, G, I, K, M, O, Q, S, U and W) and Shannon diversity index (panels B, D, F, H, J, L, N, P, R, T, V and X), with ANOVA. The variables include BMI, waist measurement, waist-to-height ratio, blood pressure, systolic blood pressure, diastolic blood pressure, heart rate, pulse pressure, triglyceride levels, HDL cholesterol, LDL cholesterol and total cholesterol depicted in panel (A–X).
Supplementary Figure 4 | Beta diversity. Comparison of beta diversity exploring differences between additional sample variables. Bray-Curtis dissimilarity calculated from Hellinger transformed total sum scaled data was used as beta-diversity measure and visualized with principal coordinate analysis (PCoA). The variables include BMI, waist measurement, waist-to-height ratio, blood pressure, systolic blood pressure, diastolic blood pressure, heart rate, pulse pressure, triglyceride levels, HDL cholesterol, LDL cholesterol and total cholesterol depicted in panel (A–X), respectively.
Supplementary Figure 5 | Relative abundance plots. Relative abundance of most abundant genera (panels A, C, E, G, I, K, M, O, Q, S, U and W) and differential abundant genera (panels B, D, F, H, J, L, N, P, R, T, V and X). Variables include BMI, waist measurement, waist-to-height ratio, blood pressure, systolic blood pressure, diastolic blood pressure, heart rate, pulse pressure, triglyceride levels, HDL cholesterol, LDL cholesterol and total cholesterol depicted in panel (A–X), respectively. Comparison of groups were performed with non-parametric tests (Kruskal-Wallis). Unadjusted as well as Benjamini-Hochberg corrected q-values are included in the plot. All tests are included in supplementary file 2 as well as a parametric approach (DESeq as implemented in DAtest) in supplementary file 3).
Almeida-Santos, A., Martins-Mendes, D., Gayà-Vidal, M., Pérez-Pardal, L., Beja-Pereira, A. (2021). Characterization of the oral microbiome of medicated type-2 diabetes patients. Front. Microbiol. 12, 610370. doi: 10.3389/fmicb.2021.610370
American Heart Association. (2022). Healthy and unhealthy blood pressure ranges (Dallas, Texas, The United States of America: Heart Attack and Stroke Symptoms). Available at: https://www.heart.org/en/health-topics/high-blood-pressure/understanding-blood-pressure-readings.
Barb, J., Maki, K., Kazmi, N., Meeks, B., Krumlauf, M., Tuason, R., et al. (2022). The oral microbiome in alcohol use disorder: a longitudinal analysis during inpatient treatment. J. Oral. Microbiol. 14 (1), 2004790. doi: 10.1080/20002297.2021.2004790
Belstrøm, D. (2020). The salivary microbiota in health and disease. J. Oral. Microbiol. 12 (1), 1723975. doi: 10.1080/20002297.2020.1723975
Belstrøm, D., Constancias, F., Drautz-Moses, D. I., Schuster, S. C., Veleba, M., Mahé, F., et al. (2021). Periodontitis associates with species-specific gene expression of the oral microbiota. NPJ Biofilms Microbiomes 7 (1), 76. doi: 10.1038/s41522-021-00247-y
Belstrøm, D., Grande, M. A., Sembler-Møller, M. L., Kirkby, N., Cotton, S. L., Paster, B. J., et al. (2018). Influence of periodontal treatment on subgingival and salivary microbiotas. J. Periodontol. 89 (5), 531–539. doi: 10.1002/JPER.17-0377
Belstrøm, D., Holmstrup, P., Nielsen, C. H., Kirkby, N., Twetman, S., Heitmann, B. L., et al. (2014). Bacterial profiles of saliva in relation to diet, lifestyle factors, and socioeconomic status. J. Oral. Microbiol. 6 (1), 23609. doi: 10.3402/jom.v6.23609
Belstrøm, D., Sembler-Møller, M. L., Grande, M. A., Kirkby, N., Cotton, S. L., Paster, B. J., et al. (2017). Microbial profile comparisons of saliva, pooled and site-specific subgingival samples in periodontitis patients. PloS One 12 (8), e0182992. doi: 10.1371/journal.pone.0182992
Casarin, R. C. V., Barbagallo, A., Meulman, T., Santos, V. R., Sallum, E. A., Nociti, F. H., et al. (2013). Subgingival biodiversity in subjects with uncontrolled type-2 diabetes and chronic periodontitis: Subgingival biodiversity in diabetes mellitus. J. Periodontal Res. 48 (1), 30–36. doi: 10.1111/j.1600-0765.2012.01498.x
Chaudhari, D. S., Dhotre, D. P., Agarwal, D. M., Gaike, A. H., Bhalerao, D., Jadhav, P., et al. (2020). Gut, oral and skin microbiome of Indian patrilineal families reveal perceptible association with age. Sci. Rep. 10 (1), 5685. doi: 10.1038/s41598-020-62195-5
Chen, B., Wang, Z., Wang, J., Su, X., Yang, J., Zhang, Q., et al. (2020). The oral microbiome profile and biomarker in Chinese type 2 diabetes mellitus patients. Endocrine 68 (3), 564–572. doi: 10.1007/s12020-020-02269-6
Constancias, F. (2022) MetabaRpipe/databases at master · fconstancias/metabaRpipe ( Zürich, Switzerland: GitHub).). Available at: https://github.com/fconstancias/metabaRpipe.
Constancias, F., Mahé, F. (2022). fconstancias/metabaRpipe: v0.9 (Zürich Switzerland: Zenodo). Available at: https://zenodo.org/record/6423397.
Damgaard, C., Danielsen, A. K., Enevold, C., Massarenti, L., Nielsen, C. H., Holmstrup, P., et al. (2019). Porphyromonas gingivalis in saliva associates with chronic and aggressive periodontitis. J. Oral. Microbiol. 11 (1), 1653123. doi: 10.1080/20002297.2019.1653123
Díez López, C., Montiel González, D., Vidaki, A., Kayser, M. (2022). Prediction of smoking habits from class-imbalanced saliva microbiome data using data augmentation and machine learning. Front. Microbiol. 13, 886201. doi: 10.3389/fmicb.2022.886201
Dodds, M., Roland, S., Edgar, M., Thornhill, M. (2015). Saliva a review of its role in maintaining oral health and preventing dental disease. BDJ Team 2 (1–8), 15123. doi: 10.1038/bdjteam.2015.123
eHOMD (2020) eHOMD - expanded human oral microbiome database. Available at: http://www.homd.org/.
Eke, P. I., Borgnakke, W. S., Genco, R. J. (2020). Recent epidemiologic trends in periodontitis in the USA. Periodontol. 2000 82 (1), 257–267. doi: 10.1111/prd.12323
Fan, X., Peters, B. A., Jacobs, E. J., Gapstur, S. M., Purdue, M. P., Freedman, N. D., et al. (2018). Drinking alcohol is associated with variation in the human oral microbiome in a large study of American adults. Microbiome 6 (1), 59. doi: 10.1186/s40168-018-0448-x
Fei, N., Bernabé, B. P., Lie, L., Baghdan, D., Bedu-Addo, K., Plange-Rhule, J., et al. (2019). The human microbiota is associated with cardiometabolic risk across the epidemiologic transition. PloS One 14 (7), e0215262. doi: 10.1371/journal.pone.0215262
Fischer, C. C., Persson, R. E., Persson, G. R. (2008). Influence of the menstrual cycle on the oral microbial flora in women: A case-control study including men as control subjects. J. Periodontol. 79 (10), 1966–1973. doi: 10.1902/jop.2008.080057
Ganesan, S. M., Joshi, V., Fellows, M., Dabdoub, S. M., Nagaraja, H. N., O’Donnell, B., et al. (2017). A tale of two risks: smoking, diabetes and the subgingival microbiome. ISME J. 11 (9), 2075–2089. doi: 10.1038/ismej.2017.73
Gogeneni, H., Buduneli, N., Ceyhan-Öztürk, B., Gümüş, P., Akcali, A., Zeller, I., et al. (2015). Increased infection with key periodontal pathogens during gestational diabetes mellitus. J. Clin. Periodontol. 42 (6), 506–512. doi: 10.1111/jcpe.12418
Greabu, M., Battino, M., Mohora, M., Totan, A., Didilescu, A., Spinu, T., et al. (2009). Saliva–a diagnostic window to the body, both in health and in disease. J. Med. Life 2 (2), 124–132. PMC3018981/PMID: 20108531
Hajishengallis, G., Chavakis, T. (2021). Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 21 (7), 426–440. doi: 10.1038/s41577-020-00488-6
Haririan, H., Andrukhov, O., Bertl, K., Lettner, S., Kierstein, S., Moritz, A., et al. (2014). Microbial analysis of subgingival plaque samples compared to that of whole saliva in patients with periodontitis. J. Periodontol. 85 (6), 819–828. doi: 10.1902/jop.2013.130306
He, J., Huang, W., Pan, Z., Cui, H., Qi, G., Zhou, X., et al. (2012). Quantitative analysis of microbiota in saliva, supragingival, and subgingival plaque of Chinese adults with chronic periodontitis. Clin. Oral. Investig. 16 (6), 1579–1588. doi: 10.1007/s00784-011-0654-4
Holmes, B. C., Benjamin, J. C. C., McMurdie, P., Holmes, S. (2017). dada2 (Bioconductor). doi: 10.1038/nmeth.3869. Available at: https://bioconductor.org/packages/dada2.
Holmstrup, P., Damgaard, C., Olsen, I., Klinge, B., Flyvbjerg, A., Nielsen, C. H., et al. (2017). Comorbidity of periodontal disease: two sides of the same coin? an introduction for the clinician. J. Oral. Microbiol. 9 (1), 1332710. doi: 10.1080/20002297.2017.1332710
Janem, W. F., Scannapieco, F. A., Sabharwal, A., Tsompana, M., Berman, H. A., Haase, E. M., et al. (2017). Salivary inflammatory markers and microbiome in normoglycemic lean and obese children compared to obese children with type 2 diabetes. PloS One 12 (3), e0172647. doi: 10.1371/journal.pone.0172647
Johansen, N. B., Hansen, A. L. S., Jensen, T. M., Philipsen, A., Rasmussen, S. S., Jørgensen, M. E., et al. (2012). Protocol for ADDITION-PRO: a longitudinal cohort study of the cardiovascular experience of individuals at high risk for diabetes recruited from Danish primary care. BMC Public Health 12 (1), 1078. doi: 10.1186/1471-2458-12-1078
Kageyama, S., Takeshita, T., Asakawa, M., Shibata, Y., Takeuchi, K., Yamanaka, W., et al. (2017). Relative abundance of total subgingival plaque-specific bacteria in salivary microbiota reflects the overall periodontal condition in patients with periodontitis. PloS One 12 (4), e0174782. doi: 10.1371/journal.pone.0174782
Kampoo, K., Teanpaisan, R., Ledder, R. G., McBain, A. J. (2014). Oral bacterial communities in individuals with type 2 diabetes who live in southern Thailand. Appl. Environ. Microbiol. 80 (2), 662–671. doi: 10.1128/AEM.02821-13
Kolde, R.2019 Pheatmap: Pretty heatmaps. Available at: https://CRAN.R-project.org/package=pheatmap.
Ko, Y., Lee, E. M., Park, J. C., Gu, M. B., Bak, S., Ji, S. (2020). Salivary microbiota in periodontal health and disease and their changes following nonsurgical periodontal treatment. J. Periodontal Implant Sci. 50 (3), 171–182. doi: 10.5051/jpis.2020.50.3.171
LaMonte, M. J., Genco, R. J., Buck, M. J., McSkimming, D. I., Li, L., Hovey, K. M., et al. (2019). Composition and diversity of the subgingival microbiome and its relationship with age in postmenopausal women: an epidemiologic investigation. BMC Oral. Health 19 (1), 246. doi: 10.1186/s12903-019-0906-2
LaMonte, M. J., Gordon, J. H., Diaz-Moreno, P., Andrews, C. A., Shimbo, D., Hovey, K. M., et al. (2022). Oral microbiome is associated with incident hypertension among postmenopausal women. JAHA 11 (6), e021930. doi: 10.1161/JAHA.121.021930
Liao, Y., Tong, X. T., Jia, Y. J., Liu, Q. Y., Wu, Y. X., Xue, W. Q., et al. (2022). The effects of alcohol drinking on oral microbiota in the Chinese population. IJERPH 19 (9), 5729. doi: 10.3390/ijerph19095729
Liu, Y.K., Chen, V., He, J.z., Zheng, X., Xu, X., Zhou, X.d. (2021). A salivary microbiome-based auxiliary diagnostic model for type 2 diabetes mellitus. Arch. Oral. Biol. 126, 105118. doi: 10.1016/j.archoralbio.2021.105118
Liu, S., Wang, Y., Zhao, L., Sun, X., Feng, Q. (2020). Microbiome succession with increasing age in three oral sites. Aging (Albany NY) 12 (9), 7874–7907. doi: 10.18632/aging.103108
Liu, L., Zhang, Y., Wu, W., Cheng, M., Li, Y., Cheng, R. (2013). Prevalence and correlates of dental caries in an elderly population in northeast China. PloS One 8 (11), e78723. doi: 10.1371/journal.pone.0078723
Longo, P. L., Dabdoub, S., Kumar, P., Artese, H. P. C., Dib, S. A., Romito, G. A., et al. (2018). Glycaemic status affects the subgingival microbiome of diabetic patients. J. Clin. Periodontol. 45 (8), 932–940. doi: 10.1111/jcpe.12908
Lynge Pedersen, A. M., Belstrøm, D. (2019). The role of natural salivary defences in maintaining a healthy oral microbiota. J. Dentistry 80, S3–12. doi: 10.1016/j.jdent.2018.08.010
Matsha, T. E., Prince, Y., Davids, S., Chikte, U., Erasmus, R. T., Kengne, A. P., et al. (2020). Oral microbiome signatures in diabetes mellitus and periodontal disease. J. Dent. Res. 99 (6), 658–665. doi: 10.1177/0022034520913818
Minty, M., Loubières, P., Canceill, T., Azalbert, V., Burcelin, R., Tercé, F., et al. (2021). Gender-associated differences in oral microbiota and salivary biochemical parameters in response to feeding. J. Physiol. Biochem. 77 (1), 155–166. doi: 10.1007/s13105-020-00757-x
Morozumi, T., Nakagawa, T., Nomura, Y., Sugaya, T., Kawanami, M., Suzuki, F., et al. (2016). Salivary pathogen and serum antibody to assess the progression of chronic periodontitis: a 24-mo prospective multicenter cohort study. J. Periodont Res. 51 (6), 768–778. doi: 10.1111/jre.12353
Murugesan, S., Elanbari, M., Bangarusamy, D. K., Terranegra, A., Al Khodor, S. (2021). Can the salivary microbiome predict cardiovascular diseases? lessons learned from the Qatari population. Front. Microbiol. 12, 772736. doi: 10.3389/fmicb.2021.772736
Nearing, J. T., DeClercq, V., Van Limbergen, J., Langille, M. G. I. (2020). Assessing the variation within the oral microbiome of healthy adults. mSphere 5 (5), e00451–e00420. doi: 10.1128/mSphere.00451-20
Nickles, K., Scharf, S., Röllke, L., Dannewitz, B., Eickholz, P. (2017). Comparison of two different sampling methods for subgingival plaque: Subgingival paper points or mouthrinse sample? J. Periodontol. 88 (4), 399–406. doi: 10.1902/jop.2016.160249
Ogawa, T., Honda-Ogawa, M., Ikebe, K., Notomi, Y., Iwamoto, Y., Shirobayashi, I., et al. (2017). Characterizations of oral microbiota in elderly nursing home residents with diabetes. J. Oral. Sci. 59 (4), 549–555. doi: 10.2334/josnusd.16-0722
Oksanen, J., Simpson, G. L., Guillaume Blanchet, F., Kindt, R., Legendre, P. (2022) Vegan: Community ecology package. Available at: https://CRAN.R-project.org/package=vegan.
Pedersen, A., Sørensen, C., Proctor, G., Carpenter, G. (2018). Salivary functions in mastication, taste and textural perception, swallowing and initial digestion. Oral. Dis. 24 (8), 1399–1416. doi: 10.1111/odi.12867
R Core Team (2020). R: A language and environment for statistical computing (Vienna, Austria: R foundation for Statistical Computing). Available at: https://www.R-project.org/.
Rivera-Hidalgo, F. (2000). Smoking and periodontal disease. Periodontol 2003, 32:50–32:58. doi: 10.1046/j.0906-6713.2003.03205.x
Rodríguez-Rabassa, M., López, P., Rodríguez-Santiago, R., Cases, A., Felici, M., Sánchez, R., et al. (2018). Cigarette smoking modulation of saliva microbial composition and cytokine levels. IJERPH 15 (11), 2479. doi: 10.3390/ijerph15112479
Rohart, F., Gautier, B., Singh, A., Lê Cao, K. A. (2017). mixOmics: An r package for ‘omics feature selection and multiple data integration. PloS Comput. Biol. 13 (11), e1005752. doi: 10.1371/journal.pcbi.1005752
Sabharwal, A., Stellrecht, E., Scannapieco, F. A. (2021). Associations between dental caries and systemic diseases: a scoping review. BMC Oral. Health 21 (1), 472. doi: 10.1186/s12903-021-01803-w
Saeb, A. T. M., Al-Rubeaan, K. A., Aldosary, K., Udaya Raja, G. K., Mani, B., Abouelhoda, M., et al. (2019). Relative reduction of biological and phylogenetic diversity of the oral microbiota of diabetes and pre-diabetes patients. Microbial. Pathogen. 128, 215–229. doi: 10.1016/j.micpath.2019.01.009
Schrøder, S. A., Bardow, A., Eickhardt-Dalbøge, S., Johansen, H. K., Homøe, P. (2017). Is parotid saliva sterile on entry to the oral cavity? Acta Oto-Laryngol. 137 (7), 762–764. doi: 10.1080/00016489.2016.1272002
Segata, N., Haake, S., Mannon, P., Lemon, K. P., Waldron, L., Gevers, D., et al. (2012). Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome Biol. 13 (6), R42. doi: 10.1186/gb-2012-13-6-r42
Si, J., Lee, C., Ko, G. (2017). Oral microbiota: Microbial biomarkers of metabolic syndrome independent of host genetic factors. Front. Cell Infect. Microbiol. 7, 516. doi: 10.3389/fcimb.2017.00516
Stockholm, J. (2022). Rabuplot: Copenhagen, Denmark Relative abundance plot. https://github.com/jstokholm/rabuplot
Sun, X., Li, M., Xia, L., Fang, Z., Yu, S., Gao, J., et al. (2020). Alteration of salivary microbiome in periodontitis with or without type-2 diabetes mellitus and metformin treatment. Sci. Rep. 10 (1), 15363. doi: 10.1038/s41598-020-72035-1
Suzuki, N., Nakano, Y., Yoneda, M., Hirofuji, T., Hanioka, T. (2022). The effects of cigarette smoking on the salivary and tongue microbiome. Clin. Exp. Dental Res. 8 (1), 449–456. doi: 10.1002/cre2.489
Takeshita, T., Kageyama, S., Furuta, M., Tsuboi, H., Takeuchi, K., Shibata, Y., et al. (2016). Bacterial diversity in saliva and oral health-related conditions: the hisayama study. Sci. Rep. 6 (1), 22164. doi: 10.1038/srep22164
Tam, J., Hoffmann, T., Fischer, S., Bornstein, S., Gräßler, J., Noack, B. (2018). Obesity alters composition and diversity of the oral microbiota in patients with type 2 diabetes mellitus independently of glycemic control. PloS One 13 (10), e0204724. doi: 10.1371/journal.pone.0204724
The Danish Health Authority (2022). Sundhedsstyrelsens udmeldinger om indtag af alkohol (Copenhagen, Denmark: The Danish Health Authority). Available at: https://www.sst.dk/-/media/Udgivelser/2022/Alkohol/Notat-om-Sundhedsstyrelsens-alkoholudmeldinger.ashx.
The Global Diabetes Community (2019). HbA1c test for diabetes (University of Warwick, Coventry, United Kingdom: Diabetes.co.uk). Available at: https://www.diabetes.co.uk/hba1c-test.html.
Tiwari, M. (2011). Science behind human saliva. J. Nat. Sc Biol. Med. 2 (1), 53. doi: 10.4103/0976-9668.82322
Vallabhan, C. G., Sivarajan, S., Shivkumar, A. D., Narayanan, V., Vijayakumar, S., Indhuja, R. S. (2020). Assessment of salivary flow rate in patients with chronic periodontitis. J. Pharm. Bioallied. Sci. 12 (Suppl 1), S308–S312. doi: 10.4103/jpbs.JPBS_92_20
Wang, X., Mi, Q., Yang, J., Guan, Y., Zeng, W., Xiang, H., et al. (2022). Effect of electronic cigarette and tobacco smoking on the human saliva microbial community. Braz. J. Microbiol. 53 (2), 991–1000. doi: 10.1007/s42770-022-00721-5
Wirth, R., Maróti, G., Mihók, R., Simon-Fiala, D., Antal, M., Pap, B., et al. (2020). A case study of salivary microbiome in smokers and non-smokers in Hungary: analysis by shotgun metagenome sequencing. J. Oral. Microbiol. 12 (1), 1773067. doi: 10.1080/20002297.2020.1773067
World Health Organization (2020). Physical activity (Geneva, Switzerland: World Health Organisation). Available at: https://www.who.int/news-room/fact-sheets/detail/physical-activity.
World Health Organization (2021a). Diabetes (Geneva, Switzerland: World Health Organisation). Available at: https://www.who.int/news-room/fact-sheets/detail/diabetes.
World Health Organization (2021b). Cardiovascular diseases (CVDs) (Geneva, Switzerland: World Health Organisation). Available at: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds).
World Health Organization (2022). Tobacco (World Health Organisation). Available at: https://www.who.int/news-room/fact-sheets/detail/tobacco.
Wu, J., Peters, B. A., Dominianni, C., Zhang, Y., Pei, Z., Yang, L., et al. (2016). Cigarette smoking and the oral microbiome in a large study of American adults. ISME J. 10 (10), 2435–2446. doi: 10.1038/ismej.2016.37
Yang, Y., Cai, Q., Zheng, W., Steinwandel, M., Blot, W. J., Shu, X. O., et al. (2019). Oral microbiome and obesity in a large study of low-income and African-American populations. J. Oral. Microbiol. 11 (1), 1650597. doi: 10.1080/20002297.2019.1650597
Yang, Y., Liu, S., Wang, Y., Wang, Z., Ding, W., Sun, X., et al. (2020). Changes of saliva microbiota in the onset and after the treatment of diabetes in patients with periodontitis. Aging 12 (13), 13090–13114. doi: 10.18632/aging.103399
Yoshizawa, J. M., Schafer, C. A., Schafer, J. J., Farrell, J. J., Paster, B. J., Wong, D. T. W. (2013). Salivary biomarkers: Toward future clinical and diagnostic utilities. Clin. Microbiol. Rev. 26 (4), 781–791. doi: 10.1128/CMR.00021-13
Keywords: saliva, type 2 diabetes (T2D), smoking, microbiota, biomarker
Citation: Poulsen CS, Nygaard N, Constancias F, Stankevic E, Kern T, Witte DR, Vistisen D, Grarup N, Pedersen OB, Belstrøm D and Hansen T (2022) Association of general health and lifestyle factors with the salivary microbiota – Lessons learned from the ADDITION-PRO cohort. Front. Cell. Infect. Microbiol. 12:1055117. doi: 10.3389/fcimb.2022.1055117
Received: 27 September 2022; Accepted: 24 October 2022;
Published: 16 November 2022.
Edited by:
Gislane Lelis Vilela de Oliveira, São Paulo State University, BrazilReviewed by:
Divyashri Baraniya, Temple University, United StatesCopyright © 2022 Poulsen, Nygaard, Constancias, Stankevic, Kern, Witte, Vistisen, Grarup, Pedersen, Belstrøm and Hansen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel Belstrøm, ZGJlbEBzdW5kLmt1LmRr; Torben Hansen, dG9yYmVuLmhhbnNlbkBzdW5kLmt1LmRr
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.