
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 01 July 2021
Sec. Fungal Pathogenesis
Volume 11 - 2021 | https://doi.org/10.3389/fcimb.2021.642658
 Lana Sarita de Souza Oliveira1Luciana Magalhães Pinto1Mariana Araújo Paulo de Medeiros1Dena L. Toffaletti2
Lana Sarita de Souza Oliveira1Luciana Magalhães Pinto1Mariana Araújo Paulo de Medeiros1Dena L. Toffaletti2 Jennifer L. Tenor2Tânia Fraga Barros3
Jennifer L. Tenor2Tânia Fraga Barros3 Rejane Pereira Neves4
Rejane Pereira Neves4 Reginaldo Gonçalves de Lima Neto5Eveline Pipolo Milan6Ana Carolina Barbosa Padovan7Walicyranison Plinio da Silva Rocha8
Reginaldo Gonçalves de Lima Neto5Eveline Pipolo Milan6Ana Carolina Barbosa Padovan7Walicyranison Plinio da Silva Rocha8 John R. Perfect2
John R. Perfect2 Guilherme Maranhão Chaves1*
Guilherme Maranhão Chaves1*Cryptococcosis is an infectious disease of worldwide distribution, caused by encapsulated yeasts belonging to the phylum Basidiomycota. The genus Cryptococcus includes several species distributed around the world. The C. gattii/neoformans species complex is largely responsible for most cases of cryptococcosis. However, clinical series have been published of infections caused by Papiliotrema (Cryptococcus) laurentii and Naganishia albida (Cryptococcus albidus), among other related genera. Here, we examined the pathogenic potential and antifungal susceptibility of C. gattii/neoformans species complex (clades I and II) and related genera (Papiliotrema and Naganishia) isolated from environmental and clinical samples. P. laurentii (clade III), N. liquefasciens/N. albidosimilis (clade IV); and N. adeliensis/N. albida (clade V) strains produced higher levels of phospholipase and hemolysins, whereas the C. gattii/neoformans species complex strains (clades I and II) had markedly thicker capsules, produced more biofilm biomass and melanin, which are known virulence attributes. Interestingly, 40% of C. neoformans strains (clade II) had MICs above the ECV established for this species to amphotericin B. Several non-C. gattii/neoformans species complex (clades III to V) had MICs equal to or above the ECVs established for C. deuterogattii and C. neoformans for all the three antifungal drugs tested. Finally, all the non-C. gattii/neoformans clinical isolates (clades III to V) produced more melanin than the environmental isolates might reflect their particularly enhanced need for melanin during in vivo protection. It is very clear that C. gattii/neoformans species complex (clades I and II) strains, in general, show more similar virulence phenotypes between each other when compared to non-C. gattii/neoformans species complex (clades III to V) isolates. These observations together with the fact that P. laurentii and Naganishia spp. (clades III to V) strains were collected from the outside of a University Hospital, identify features of these yeasts important for environmental and patient colonization and furthermore, define mechanisms for infections with these uncommon pathogens.
Cryptococcosis is an infectious disease of worldwide distribution, caused by encapsulated yeasts belonging to the phylum Basidiomycota (Kwon-Chung and Bennett, 1992; Maziarz and Perfect, 2016). Cryptococcus spp., such as the Cryptococcus gattii/neoformans species complex, are able to enter mammalian hosts and cause damage to the central nervous system (CNS) (Kwon-Chung et al., 2014; Lakoh et al., 2020) and respiratory tract (Lakoh et al., 2020). In the last few decades, cryptococcal meningoencephalitis has become an infection of global relevance, with up to 1 million new infections annually and significant attributable morbidity and mortality. Even with widespread effective antiretroviral therapy, there is still approximately 200,000 deaths per year especially among patients with human immunodeficiency virus (HIV) infection and AIDS (Park et al., 2009; Park et al., 2011). Besides HIV/AIDS infection, other risk factors for cryptococcosis include corticosteroids and other immunosuppressive therapies, solid organ transplantation, malignant and rheumatological diseases (Maziarz and Perfect, 2016).
The genus Cryptococcus includes several species distributed around the world and found in different environments (Chee and Lee, 2005; Loperena-Alvarez et al., 2010), such as bird droppings (mainly pigeons) (Irokanulo et al., 1997; Soltani et al., 2013; Kassi et al., 2018), soil, rivers, decomposing vegetation and tree hollows (Lazera et al., 1998; Kidd et al., 2007; Alves et al., 2016; Santos Bentes et al., 2019). Recently, Hagen et al. (2015) recognized multiple species within the Cryptococcus gattii/neoformans species complex, including the existence of four C. neoformans genotypes: VNI and VNII (var. grubii, serotype A), VNIII (hybrid serotype AD), VNIV (var. neoformans, serotype D); and four genotypes of C. gattii: VGI, VGII, VGIII and VGIV (serotypes B and C). Others have also reported the presence of VNBI and VNBII genotypes within the taxon previously considered C. neoformans var. grubii (Desjardins et al., 2017). More recently, a new C. gattii molecular type has been discovered in the African environments of the Central Miombo Woodlands (VGV) (Farrer et al., 2019).
Cryptococcus gattii/neoformans species complex is primarily related to the vast majority of cases of cryptococcosis (Kwon-Chung et al., 2014; Maziarz and Perfect, 2016), but other species previously known as C. laurentii and C. albidus can cause disease (Cleveland et al., 2013; Choe et al., 2020). These species are responsible for 80% of the cases of infection caused by non-Cryptococcus gattii/neoformans species complex yeasts (Johnson et al., 1998; Khawcharoenporn et al., 2007; Smith et al., 2017; Castro-Lainez et al., 2019; Zhang et al., 2019). Based on DNA-sequencing of seven different genes, the class Tremellomycetes (Agaricomicotina lineage, Phylum Basidiomycota) including the genus Cryptococcus was recently reclassified. Therefore, the former species C. laurentii is now a synonym of Papiliotrema laurentii and a few other Cryptococcus species of medical interest have been currently included in the genus Naganishia as follows: Naganishia (Cryptococcus) diffluens, Naganishia (Cryptococcus) liquefaciens, Naganishia (Cryptococcus) albidosimilis, Naganishia albida (Cryptococcus albidus) and Naganishia (Cryptococcus) adeliensis (Liu et al., 2015).
Several virulence factors have classically been carefully characterized and validated for C. gattii/neoformans complex, such as the presence of a polysaccharide capsule that protects yeast cells against phagocytosis, the ability to grow at 37°C, biofilm formation, and the production of additional factors including melanin, phospholipase, metalloprotease, DNase, urease, superoxide dismutase and other antioxidant enzymes (Mancianti et al., 2002; Sanchez et al., 2008; Peng et al., 2018; Zaragoza, 2019). However, only a few studies have investigated the potential role of these and other virulence factors in the non-Cryptococcus gattii/neoformans species complex (Ikeda et al., 2002; Andrade-Silva et al., 2010; Araujo Gde et al., 2012; Ferreira-Paim et al., 2012; Araujo et al., 2017).
Because of the variations of virulence factors and clinical resistance to azole monotherapy, it is important to know and compare the relevant pathogenic potential and antifungal susceptibility profiling of Cryptococcus spp. collected from clinical and environmental sources. Therefore, the primary objectives of this study were to evaluate the pathogenic features and antifungal susceptibility of Cryptococcus-related genera isolated from pigeon droppings (environmental), compared to the Cryptococcus gattii/neoformans species complex strains obtained from clinical samples of patients belonging to 3 different Brazilian states (Bahia, Pernambuco and Rio Grande do Norte) in the Northeast region. This is the first study to demonstrate that non-Cryptococcus gattii/neoformans species complex may express a full range of known virulence factors in vitro and furthermore, some strains may exhibit increased minimal inhibitory concentrations (MICs) to antifungal drugs without previous known exposure to these compounds.
The study included samples of pigeon droppings collected outside the building of a tertiary care university hospital, located in Natal City, Rio Grande do Norte State, Brazil, from April 2012 to March 2014 and November 2017 to February 2018.
Approximately, 2g of dried and/or fresh pigeon guano was collected from several sites and diluted in saline solution (0.9% NaCl) in a ratio of 1:10, vortexed for 10 minutes and allowed to stand for 1h. Subsequently, 100 μL of the supernatant was inoculated on the surface of Sabouraud Dextrose Agar (SDA; Oxoid, Basingstoke, Hampshire, UK) and Rose Bengal (0.05 g/L) plates with the aid of a Drigalski loop. Each plate was incubated at 30°C for 48h. Yeast colonies with creamy to mucoid phenotypes were selected for subsequent identification.
After colony growth, yeast cells were streaked onto the surface of CHROMagar Candida ® (CHROMagar Microbiology, Paris, France) to check for purity and screening for different color colonies (Baumgartner et al., 1996). The initial identification (screening for the Cryptococcus genus) was based on the characteristics of the yeast cells observed microscopically after cultivation on corn meal agar with Tween 80, glucose fermentation (non-fermentative), urease test, Niger seed agar (NSA) for melanin production and capsule visualization with an India ink stain (Yarrow, 1998). After identification, all cryptococcal yeast isolates were cultured in YPD broth (Yeast Peptone Dextrose, yeast extract 10 g/L, dextrose 20g/L, peptone 20g/L) overnight at 30°C, and then transferred to cryotubes containing 20% glycerol and stored at -80°C. The isolates now belong to the fungal culture collection of the Medical and Molecular Mycology Laboratory, Department of Clinical and Toxicological Analyses, Federal University of Rio Grande do Norte.
The isolates were grown on YPD (BD Difco, NJ, USA) plates for 72 h and then, a single colony was inoculated into 3.5 mL of 2xYPD broth in a 14 ml culture tube and incubated at 30°C with shaking (225 rpm) for 48 h. DNA was extracted using the MasterPure Yeast DNA Purification Kit (Epicentre Biotechnologies, Madison, WI, USA). DNA concentrations were measured using the Qubit dsDNA HS assay according to instructions (Life Technologies).
The PCR Hot Start Taq 2X Master Mix (New England Biolabs) was used to amplify all fragments generated in this work. Samples were amplified in a Bio-rad thermocycler (Thermo Cycler T100) using the following cycling parameters for both regions: one initial cycle of 95°C for 30 sec; followed by 30 cycles of 30 sec at 95°C, 1 min at 47°C and 1 min at 68°C and a final cycle of 5 min at 68°C. PCR products were treated with ExoSAP-IT (Thermo Fisher Scientific) according to the manufacturer’s protocol. Purified PCR products were sequenced by the Sanger sequencing method on ABI 3730 xl DNA Sequencers by Eton Bioscience, Inc. For sequencing the ITS region, the internal primers ITS1 and ITS4 were used (White et al., 1990). Nucleotide sequences were submitted for BLAST analysis at the NCBI site (http://www.ncbi.nlm.nih.gov) for species identification. Only sequences deposited in GenBank showing high similarities with our query sequences and an E value of lower than 10−5 were used in this study.
GenBank accession numbers may be seen in Table 1.
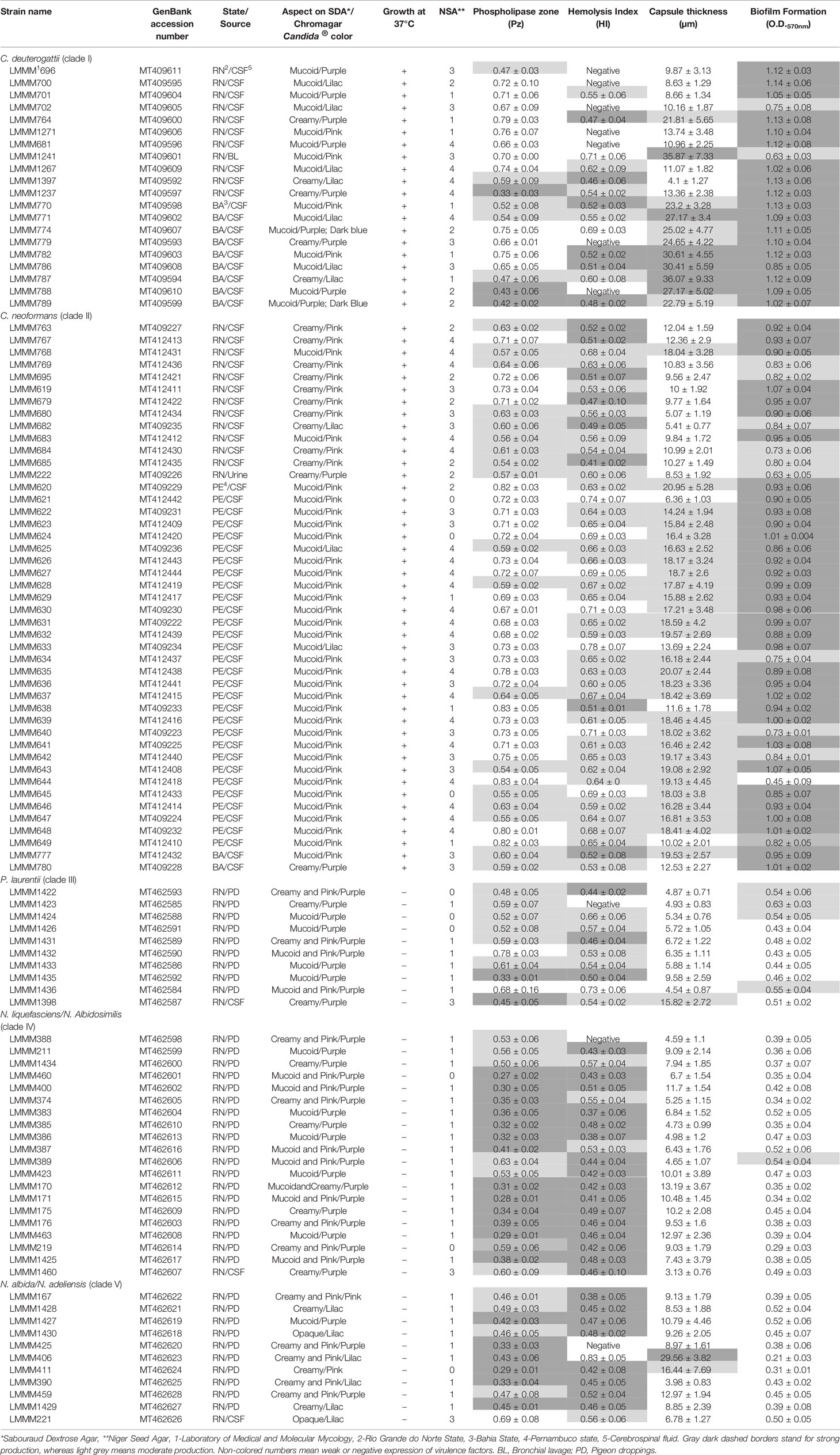
Table 1 Phenotypic properties and virulence attributes determined in vitro of clinical and environmental isolates of Cryptococcus spp. and related genera obtained from patients and PD in Northeast Brazil.
The ITS rDNA sequences generated from all the isolates were aligned with type strain sequences from Cryptococcus spp. and the other genera present as matches in the BLASTn searches (http://blast.ncbi.nlm.nih.gov/Blast.cgi), according to taxonomic reclassification of members within the Tremellomycetes (Liu et al., 2015). All sequences were aligned using the Muscle algorithm implemented by MEGA X (Kumar et al., 2018). The phylogeny was inferred using the Neighbor-Joining method (Saitou and Nei, 1987), including gap positions, and the evolutionary distances were corrected by the Kimura 2-parameter method (Kimura, 1980). Bootstrap analysis (bt) (Felsenstein, 1985) was conducted by evaluating 1,000 pseudoreplicates of the alignment. All analyses were computed using MEGA X (Kumar et al., 2018).
We have included 106 isolates of Cryptococcus spp. and related genera from our culture collection to perform the evaluation of these isolates virulence factor expressions and antifungal susceptibility testing in vitro. Thirty-eight of them were environmental: Papiliotrema laurentii (clade III; n=9), N. liquefasciens/N. albidosimilis (clade IV; n=19), N. adeliensis/N. albida (clade V; n=10). Sixty-eighth clinical isolates primarily from the Cryptococcus gattii/neoformans species complex were assessed including C. neoformans (clade II; n=45), C. gattii (clade I; n=20), P. laurenttii (clade III; n=1), N. liquefasciens/N. albidosimilis (clade IV; n=1) and N. adeliensis/N. albida (clade V; n=1). The clinical isolates were obtained from patients admitted to tertiary hospitals from three different Brazilian states (Rio Grande do Norte, Pernambuco and Bahia); all of them located in the Brazilian Northeast region (Table 1).
For the phenotypic characterizations of the different isolates, the yeasts were initially grown in NGY broth medium (Difco Neopeptone 1 g/L, Dextrose 4 g/L; Difco yeast extract 1 g/L) prior to the phospholipase and hemolysin enzymatic activity tests. Yeast cells were inoculated by “wet looping” in this medium with a ring loop loaded with a yeast suspension film, rapidly immersed in the medium and removed and incubated for 18-24h in a shaker incubator at 30°C at 200 rpm. An inoculum of approximately 2x108 cells/mL is produced (Chaves et al., 2007). Cultures were spectrophotometrically measured at a wavelength of 600nm ranging from 0.8 and 1.2 (Biochrom Libra S32). Subsequently, yeast cells were diluted to obtain the same inoculum for each condition evaluated in vitro.
For detection of the phospholipase activity, the method of Price et al. (1982) (Price et al., 1982) adapted for Cryptococcus spp. was used (Pini et al., 2017). Overnight NGY cultures were diluted and standardized to a concentration of 2x108 yeast cells/mL and the suspension of yeast cells was inoculated in triplicate onto the surface of phospholipase agar (10 g peptone, 40 g dextrose, 16 g agar, 80 mL Egg Yolk Emulsion [Fluka] was added to 1000 mL of distilled water). The plates were incubated at 30°C for 10 days. After the incubation period, the diameters of the colonies and the halo formed around them were measured. The Pz (phospholipase zone) was determined by dividing the colony diameter by the precipitation zone plus colony diameter. The strains were classified as follows, according to tertiles distribution: Pz = 1 as negative phospholipase activity; 0.65 ≤ Pz ≤ 0.83 as weak; 0.46 ≤ Pz ≤ 0.64 as moderate; 0.27 ≤ Pz ≤ 0.45 as strong phospholipase producers.
In order to evaluate hemolysin production, we followed the methodology proposed by Luo (Luo et al., 2001) with some adaptations. Yeast cells were initially cultured on SDA at 30°C for 24 h. Strains were next grown overnight in NGY broth. Ten microliters of the culture were streaked out in triplicate onto the surface of SDA containing 7% fresh sheep blood (Ebe-Farma) and 3% glucose, in Petri dishes of 155 mm of diameter. The plates were incubated for 48 h at 37°C in an atmosphere with 5% CO2. After the incubation period, the presence of a clear halo around the inoculum indicated positive hemolysis. The diameter of colonies and zones of hemolysis were measured in order to obtain the hemolysis index (HI) for each strain. HI was determined by dividing the colony diameter by the precipitation zone plus colony diameter, which allowed classification of isolates into strong, moderate and weak producers. The strains were classified as follows, according to tertiles distribution: HI= 1 as negative hemolytic index; 0.69 ≤ Pz ≤ 0.84 as weak; 0.53 ≤ HI ≤ 0.68 as moderate; 0.37 ≤ HI ≤ 0.52 as strong hemolysin producers.
To perform the capsule experiments, we used the protocol of Zaragoza et al., (Zaragoza et al., 2003) with adaptations. All the strains were grown in YPD broth for 24 h at 37°C with 200 rpm agitation. After washing with phosphate buffer saline (PBS), yeast cells were adjusted to 1×105 cells/mL using a hemocytometer to standardize the inoculum and inoculated in a solution containing 10% fetal bovine serum (FBS; Sigma-Aldrich; Missouri, Saint Louis, USA) diluted in PBS (5mL of final solution in conical tubes). The tubes were incubated for 24 h at 37°C in an atmosphere of 5% CO2. Subsequently, a drop of India ink stain was added to a small volume of yeast cells in suspension and placed on a microscope slide. The samples were imaged with a Nikon Eclipse E100 microscope. Pictures were taken using a Tsview 7 software. Total diameter (including capsule) (dt) and yeast cell diameter (dy) were measured for each strain. Capsule thickness (tc) was determined with the following equation: tc=1/2 (dt-dy) in micrometers (Fernandes et al., 2018).
To evaluate melanin production, yeast cells were cultured on SDA for 72 h at 30°C. Then, a single colony was streaked out thrice (parallel sides) on the surface of NSA medium at 30°C for 10 days for visualization of melanin production. Scores from 0 to 4 were attributed as follows: 0 (zero), no pigment; 1, light brown; 2,brown; 3, dark brown; and 4, for almost black colonies (Pedroso et al., 2009).
Biofilm formation assays were performed according to Palanco et al. (2017) with some modifications. The strains were previously cultivated in Sabouraud broth at 30°C, for 24 h with shaking at 200rpm. Then, yeast cells were washed twice with PBS and 100 μL aliquots of a standardized cell suspension (1 x 108 cells/mL)were transferred to flat bottom 96 well microtiter plates and incubated for 4 h at 37°C for the adhesion phase. As controls, eight wells of each microtiter plate were handled similarly, except that no yeast cell suspensions were added. Following the adhesion phase, cell suspensions were aspirated and each well was washed twice with 150 μL of PBS to remove loosely adherent cells. A total of 200 μL of BHI (Brain Heart Infusion) medium (BD Difco) was added to each of the washed wells and incubated at 37°C in a shaker incubator at 100 rpm. Biofilms were allowed to develop for 48 h and quantified by a crystal violet assay. Briefly, the biofilm-coated wells of the microtiter plates were washed twice with 150 μL of PBS and then air dried. Subsequently, each of the washed wells was stained with 100 μL of 0.4% aqueous crystal violet solution for 5 min. Subsequently, each well was washed once with 150 μL sterile distilled water and immediately distained with the addition of 100 μL of 95% ethanol for approximately 1 min until complete crystal violet solubilization. Subsequently, 100 μL of the destaining solution was transferred to a clean well and the amount of the crystal violet stain in the referred solution was measured with a microtiter plate reader (Spectra MAX 340 Tunable Microplate Reader; Molecular Devices Ltda.) at 570 nm. The absorbance values for the controls were subtracted from the values from the test wells to minimize background interference. The isolates were classified as follows, according to tertiles distribution: 0.21 ≤ OD570nm ≤ 0.52 as weak; 0.53≤ OD570nm ≤ 0.84 as moderate; 0.85 ≤ OD570nm ≤ 1.16 as strong biofilm producers.
Solutions of fluconazole (FLU), itraconazole (ITC) and amphotericin B (AMB) were prepared in accordance with guidelines M27-A3 (CLSI, 2008) by being diluted in RPMI 1640 (Roswell Park Memorial Institute; Angus buffers and Biochemical, Niagara Falls, NY, USA) buffered 3-(N-morpholino) propanesulfonic acid (MOPS) to pH 7.0. Antifungal drugs tested were diluted serially in 10 different concentrations, namely: FLU (Pfizer Incorporated, New York, NY, USA) 0.125–64 μg/mL; ITC (Pfizer Incorporated, New York, NY, USA), and AMB (Sigma Chemical Corporation, St. Louis, MO, USA) 0.015–8 μg/mL. The inocula of all strains tested were obtained from 48 h cultivation in Sabouraud broth at 30°C and an initial cellular suspension in saline solution equivalent to the 0.5 MacFarland standard, determined spectrophotometrically at 530 nm. Then, two serial dilutions were made, the first in saline solution (1:100) and the second in RPMI (1:20), in order to obtain a final concentration of 103 cells/mL. Susceptibility to antifungal agents was evaluated by broth microdilution, as recommended within the document CLSI M27-A3 (CLSI, 2008). Aliquots of 100 μL of the final inoculum solution were dispensed in microtiter plates of 96 wells containing 100 μL of various concentrations of the tested drugs. Finally, the plates were incubated at 37°C and test reading taken after 72 h incubation. All strains were tested in duplicate. MIC was defined for azoles to the lowest drug concentration which showed approximately 50% reduction in turbidity as compared to the positive control well. For AMB, the MIC was defined as the lowest concentration able to inhibit any growth visually perceptible (CLSI, 2008; CLSI, 2012). Epidemiological cutoff values (ECVs) were used for C. deuterogattii (clade I) and C. neoformans (clade II). The ECV determines the upper limit of wild-type distribution (without intrinsic or acquired resistance mechanisms), distinguishing from non-wild-type isolates harboring intrinsic or acquired resistance mechanisms (CLSI, 2020).
Data were analyzed using the statistical software “Graph Pad version 8.0”. Results were presented as mean ± standard deviation, and differences were analyzed by the Mann–Whitney test. For all the analyses, P was considered a default value of 0.05 and the confidence interval of 95%. In addition, the values obtained for some of the virulence attribute tests in vitro were divided onto tertile categories as weak, moderate or strong producers.
All the isolates produced encapsulated blastoconidia, did not ferment glucose and were able to hydrolyze urea. Some isolates, but not all of them, produced melanin, when grown on NSA. Therefore, they were presumptively identified as Cryptococcus spp. and related genera.
For molecular species identification, DNA fragments of the ITS region were amplified and DNA sequences obtained were sent to the GenBank genome database at the NCBI website (http://ncbi.nlm.nih.gov) using BLAST to compare gene sequences. All the Cryptococcus gattii/neoformans strains were unambiguously identified by sequencing, as follows: 45 as C. neoformans (formerly C. neoformans var. grubii) and 20 as C. deuterogattii (formerly C. gattii VGII genotype). It is important to recognize that the identification of these cryptococcal strains was further confirmed with the Multi-Locus Sequence Typing (MLST) consensus scheme of seven genetic loci (unpublished data). The same trend of accurate identification was also observed for P. laurentii (10 isolates). Nevertheless, for the other non-Cryptococcus gattii/neoformans species complex strains, a less precise identification was observed. For some of the strains (n=20), BLAST comparisons revealed 100% identity (E-value of 0) with three different Naganishia spp. (N. liquefasciens/N. albidosimilis/N. diffluens), while 10 of them were 99.8% identical to N. albida and N. adeliensis).
Phylogenetic analysis confirmed the species identification of 20 isolates as C. deuterogattii with bt of 54 (Clade I). The other isolates were confirmed as C. neoformans (45) and P. laurentii (10), respectively (Clades II and III), with high bt values (99). All the other yeast isolates were identified as Naganishia spp. due to low confidence values in the analysis to confirm BLASTn species identification. Twenty isolates belong to the N. liquefasciens/N. albidosimilis clade (bt=99), excluding N. diffluens from the most probable species (clade IV), while 11 isolates were identified within the N. adeliensis/N. albida clade, grouping closer to the first species (bt=62; clade V; Figure 1).
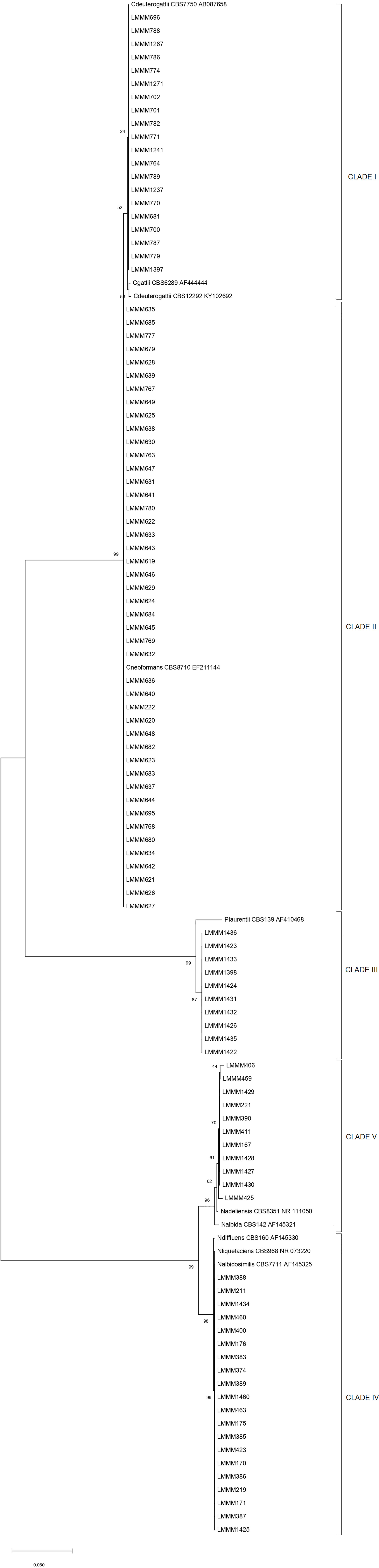
Figure 1 Neighbor-Joining phylogenetic tree of rDNA ITS sequences from 106 clinical and environmental isolates and 10 type strain sequences from Cryptococcus spp. and associated genera. Bootstrap values are depicted in all nodes (bt). The scale bar represents the number of base substitutions per site.
Cryptococcus gattii/neoformans species complex (clades I and II and N. adeliensis/N. albida (clade V) Chromagar Candida colony colors ranged from pale pink to purple (mainly pink for C. neoformans isolates), while all the isolates belonging to P. laurentii (clade II) and N. liquefasciens/N. albidosimilis (clade IV) were dark purple. All non-Cryptococcus gattii/neoformans species complex isolates (clades III to V) grew very poorly on Chromagar Candida (incubation at 37°C), whereas the opposite growth happened to clade I and II isolates (Cryptococcus gattii/neoformans species complex) All of the strains showed creamy to mucoid colony growth phenotypes when grown on YPD at 30°C, with all C. neoformans isolates being highly mucoid. Of note, strains belonging to clades III to V (non- Cryptococcus gattii/neoformans species complex) produced occasionally a pink pigment when grown on this culture medium (Figure 2). Approximately,67% of the isolates belonging to clades I and II produced dark colonies (scores 3 to 4) on NSA, while the other isolates (clades III to V) produced either light brown colonies (80.5%; score 1) or did not produce melanin at all (14.6%; score 0), except for isolates LMMM1398 (clade III), LMMM1460 (clade IV) and LMMM221 (clade V) colonies which were classified as score 3. Interestingly, these strains were non-Cryptococcus gattii/neoformans species complex strains obtained from patients (CSF samples) and not the environment (Table 1).
Figure 2 Phenotypic characteristics of Naganishia sp. after 48 h of incubation at 30° in YPD medium showing creamy colonies with an unusual pink pigment.
Phospholipase production was detected in all of the isolates (100%) evaluated. However, the levels of enzyme production varied among strains of each species. Phospholipase activity is inversely proportional to Pz, where values equal to 1 mean negative activity. The enzyme production ranged from 0.27 ± 0.02 (LMMM460; P. laurentii; clade III) to 0.83 ± 0.04 (LMMM644; C. neoformans clade II). Most of Cryptococcus gattii/neoformans species complex (clades I and II strains) were considered weak (60%) to moderate phospholipase producers (35.4%; Table 1 and Figure 3), while 51.2% of non- Cryptococcus gattii/neoformans species complex (clades III to V strains) were considered strong phospholipase producers and 41.5% showed moderate production of this enzyme. These data were reinforced with average value comparisons for the production of this enzyme, which showed significantly different results between C. gattii/neoformans species complex strains (clades I and II) and the other strains belonging to non- Cryptococcus gattii/neoformans species complex (clades III to V; Figure 3). Of note, the average value of P. laurentii (clade III) strains for phospholipase production was also considered lower than the results found for N. liquefasciens/N. albidosimilis (clade IV) and N. adeliensis/N. albida (clade V)strains (P<0.05; Figure 3). Our three clinical non-C. gattii/neoformans species complex isolates were considered strong (P. laurentii; clade III; LMMM1398; 0.45 ± 0.05 and N. adeliensis/N. albida;clade V; LMMM1429 0.45 ± 0.015) to moderate producers (N. liquefasciens/N. albidosimilis; clade IV; LMMM1460; 0.60 ± 0.09; Table 1 and Figure 3).

Figure 3 Virulence factors attributes of clinical and environmental strains of Cryptococcus spp. and related genera. (A) Phosphoplipase zone determined after incubating cells on egg yolk agar medium at 30°C, for 72 h. (B) Hemolytic index after incubation on SDA supplemented with sheep’s blood and glucose at 37°C for 48 h, 5% CO2. (C) Capsule thickness determined after cells incubation in 10% fetal bovine serum in PBS, incubated for 24 h at 37°C in an atmosphere of 5% CO2 and (D) Biofilm formation induced after cells incubation in 96-wells microtiter plates containing BHI medium at 37°C, for 48 h. Each bar represents mean ± standard deviation of the triplicate obtained for each isolate. *P < 0.05; ***P < 0.001.
Almost 91% of the isolates were able to produce hemolysis. HI is also inversely proportional to the diameter of the halo, where values equal to 1 mean negative activity. Most of the isolates in the present study were able to show medium (41.5%) to large halos (38.7%), meaning low to moderate HI. Interestingly, 7 C. deuterogattii (clade II) isolates did not produce halos suggesting that the enzyme(s) was not produced. The average values for non-C. gattii/neoformans species complex strains (clades III to V) of hemolysin production was significantly higher than those of Cryptococcus gattii/neoformans species complex (clades I and II) isolates (P<0.05; Figure 3). Most clade I and II strains were considered moderate hemolysin producers (53.8%) while larger halos (strong hemolysin production) were observed for 65.9% of clades III to V strains, being considered strong hemolysin producers, including all three clinical isolates within these clades. The average value for P. laurentii (clade III) strains hemolysin production was lower than the results found for N. liquefasciens/N. albidosimilis (clade IV) and N. adeliensis/N. albida (clade V)isolates (P<0.05; Figure 3). In addition, there was no statistically significant difference between P. laurentii (clade III) expression of this virulence factor in vitro and C. gattii/neoformans species complex (clades I and II)isolates. The isolates with the highest hemolytic activity was LMMM383, belonging to N. liquefasciens/N. albidosimilis (clade IV) (0.37 ± 0.06; Table 1).
There was a huge interspecies variation in average values of capsule thickness among certain isolates of different clades. It ranged from 4.1 ± 1.27 µm (LMMM1397) to 36.07 ± 9.33 µm (LMMM787). Interestingly, both of these isolates belonged to C. deuterogatti (clade I) showing wide strain variation. When this species was analyzed separately, it was observed that this species contained the isolates with greatest capsule thickness (LMMM1241; 35.87 ± 7.33 µm and LMMM782; 30.61 ± 4.55 µm), and several others with sizes above 20 µm were observed in this species. Furthermore, there was not a significant difference when the average values for the two clades (I and II) were compared, due to the enormous variation in measurements found within C. deuterogattii (clade I) strains, despite the observation that the greatest capsule thickness observed for C. neoformans (clade II) strains was 20.95 ± 5.28 µm (LMMM620; Table 1). All the other isolates belonging to the non- Cryptococcus gattii/neoformans species complex (clades III to V) had significantly smaller capsules when compared to Cryptococcus gattii/neoformans species complex strains (clades I and II) and no differences in average sizes were found among them (Figure 3).
All the isolates were able to form biofilms on polystyrene microtiter plates. However, there was a large variation in biofilm biomass among the isolates. The OD570nm readings ranged from 0.21 ± 0.03 (N. adeliensis/N. albida; clade V; LMMM406) to OD570nm of 1.14 ± 0.06, a C. deuterogattii (clade I) isolates (clade I; LMMM700 (P<0.05; Table 1 and Figure 3). In fact, this was the unique virulence factor expressed in vitro that showed a significant difference between C. deuterogatti (clade I) and C. neoformans (clade II) isolates. For instance, 90% of C. deuterogattii (clade I) isolates were considered strong biofilm producers, while 75.6% of C. neoformans (clade II) isolates were included in this category. Taken together, 80% of Cryptococcus gattii/neoformans species complex isolates (clades I and II) were considered strong biofilm producers. The average values for clades I and II OD570nm readings were greater than the results found for the within the non- Cryptococcus gattii/neoformans species complex (clades III to V; P<0.05; Figure 3). Furthermore, the vast majority of isolates belonging to clades I and II were not considered weak biofilm producers (except for LMMM644, a C. neoformans (clade II) strain; OD570nm of 0.45 ± 0.09). In contrast, the non-C. gattii/neoformans species complex strains (clades III to V) had no isolates identified as high biofilm producers. All N. liquefasciens/N. albidosimilis (clade IV) and 95% of N. adeliensis/N. albida (clade V strains were actually considered weak biofilm producers. In addition, 60% of P. laurentii strains (clade III) showed low biofilm formation, and the other 40% were considered moderate biofilm producers (Table 1). If we compare the average values of OD570nm readings among all three clades, P. laurentii (clade III) showed a trend for higher biofilm production compared to the other 2 clades (IV and V) and this difference was statistically significant between clades III and IV (Figure 3). Interestingly, the clinical isolates belonging to the non- Cryptococcus gattii/neoformans species complex (clades III to V) were also considered weak to moderate biofilm producers (Table 1 and Figure 3).
All MIC values obtained by the reference strains were compatible with the values expected by the CLSI methodology, assuring the reliability of the results obtained for the isolates tested. MIC ranges and geometric means (GMs) for all the antifungal drugs tested are displayed in Table 2. Because there are no breakpoints established to Cryptococcus spp. and related genera, we have included ECVs, according to the CLSI M59-Ed3 document for C. deuterogattii (clade I) and C. neoformans (clade II) strains (species that ECVs are available).
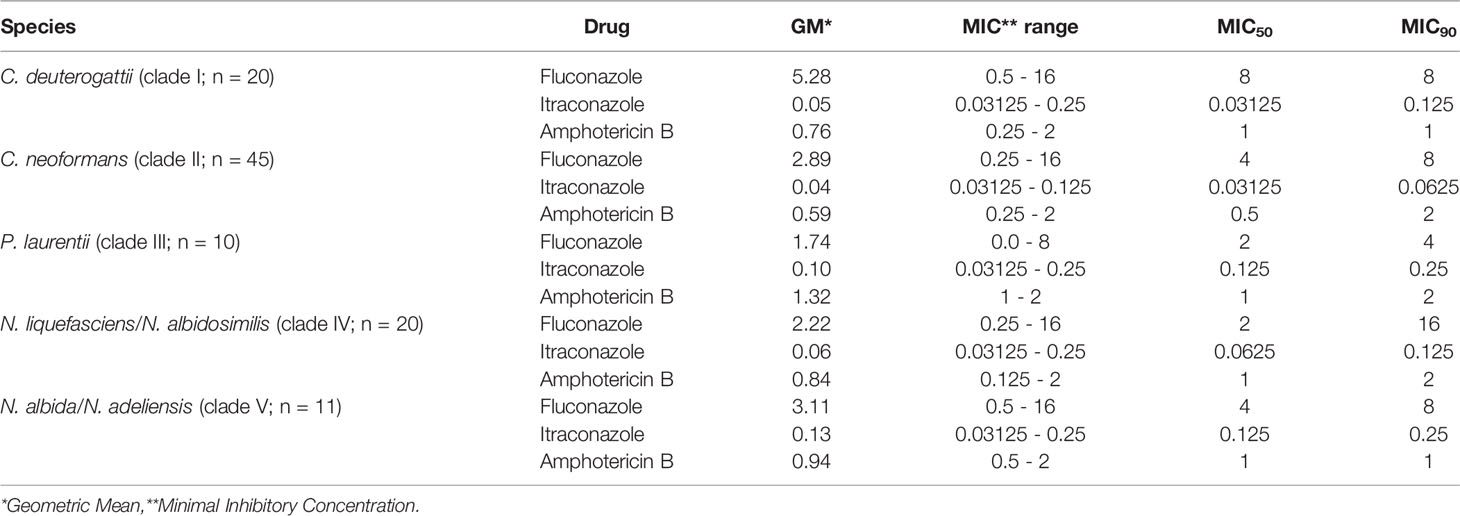
Table 2 Geometric means, MIC range, MIC50 and MIC90 of clinical and environmental isolates of Cryptococcus spp. and related genera obtained from patients and pigeon droppings in Northeast Brazil.
Most strains had low MICs to fluconazole, while a few of them showed increased MICs. Two out of 45 (4.4%) C. neoformans strains (LMMM629 and LMMM631) where considered non-wild-type. All P. laurentii isolates had low MICs to fluconazole. Four out of 20 (20%) clade IV strains and a single N. albida isolates (LMMM1430; clade V; 9.1%), had MICs ≥16 µg/mL (Table 2 and Supplementary Table 1).
All the strains tested had MICs below the ECVs established for C. deuterogattii (clade I) and C. neoformans (clade II) to itraconazole. Some strains belonging to the non-Cryptococcus gattii/neoformans species complex (clades III to V) had higher MICs, when compared to clade I and II strains MICs ≥ 0.25 µg/mL (Table 2 and Supplementary Table 1).
Several isolates in the present study were considered of non-wild-type phenotype to amphotericin B. For instance, in C. deuterogattii, 2 out of 20 strains (LMMM1271 and LMMM 774; 10%) had MICs above the ECV; in C. neoformans, this phenomenon happened with 18 out of 45 (40%) strains. All P. laurentii (clade III) and most of N. liquefasciens/N. albidosimilis (clade IV) and N. adeliensis/N. albida (clade V) strains showed MICs equal to or above the ECV established for C. deuterogattii (clade I; ECV=1 µg/mL) and C. neoformans (clade II; ECV=2 µg/mL; Table 2 and Supplementary Table 1).
Our molecular identification and phylogenetic analysis revealed five different Cryptococcus spp. and related genera clades. Regarding the C. gattii/neoformans species complex isolates (clades I and II), which comprised the vast majority of the clinical isolates, BLAST searches showed that our C. gattii isolates (clade I) belonged to the VGII genotype, named as C. deuterogatti (Hagen et al., 2015). This is the main species recovered in Brazil from both environmental and clinical samples (Dos Freire et al., 2012; Maruyama et al., 2019; Santos Bentes et al., 2019). Following the same trend of other studies performed in Brazil (Cogliati, 2013; Alves et al., 2016; Ferreira-Paim et al., 2017), our C. neoformans strains were all considered the former C. neoformans var. grubii, but because of the recent recognition as a separate species in the C.gattii/neoformans species complex (Hagen et al., 2015), we will use the nomenclature C. neoformans, as recommended by Hagen et al., (Hagen et al., 2017).
Regarding the non-C. gattii/C. neoformans species complex (clades III to V) isolates identification, we faced a limitation using ITS region sequencing, particularly among Naganishia spp. (clades IV and V) isolates. When our sequences were submitted to GenBank, the final identification was not precise, because they were considered identical to three different species as follows: N. liquefaciens, N.albidosimilis and N. diffluens (clade IV). However, our phylogenetic analysis excluded N. diffluens. The same level of indistinguishability was found for N. albida/adeliensis (clade V). Furthermore, all of them were previously considered Cryptococcus spp., but Liu et al. (Liu et al., 2015) reclassified several Tremellomycetes isolates, based on the results of phylogenetic analyses from a seven-genes dataset, including the large subunit ribosomal DNA (LSU rDNA locus). Because we have only used a single locus in our study (ITS region), we labeled our isolates as Naganishia spp. only and have submitted the sequences to the GenBank database as such. Further typing using DNA sequence of different loci will be needed for accurate species identification.
Similar to our findings, Brito et al., (Brito et al., 2019) reported the recovery of P. laurentii, N. albida and N. liquefaciens among other former Cryptococcus spp. from bird excreta, collected outside a Brazilian University hospital, but no C. neoformans strains were found. P. laurentii was also recovered from bird droppings samples in Brazil, together with C. neoformans in the same samples (Andrade-Silva et al., 2010; Ferreira-Paim et al., 2011). N. albida, N. uzbekistanensis, N. adeliensis and C. neoformans were isolated from pigeon nests. N. adeliensis was the only species isolated from Eucalyptus trees in Ilam, Iran (Kamari et al., 2017).
Although most human infections and specifically, those related to the CNS are caused by C. gattii/neoformans species complex isolates, less common former Cryptococcus spp. have been reported to cause invasive disease due to P. laurentii and N. albida (Cheng et al., 2001; Averbuch et al., 2002; McCurdy and Morrow, 2003; Shankar et al., 2006; Khawcharoenporn et al., 2007; Furman-Kuklinska et al., 2009; Gullo et al., 2013; Neves et al., 2015). A systematic review performed by Londero et al., (Londero et al., 2019) reported 35 cases of deep-seated infections due to P. laurentii. In this study, 16.1% of cases were catheter-related, 67.7% had positive blood (54.8%) or CSF (12.9%) cultures. Most patients were immunosuppressed and 48.4% of these patients had neoplasia as an underlying disease. Choe et al. (Choe et al., 2020) reported a case series of 20 patients with N. albida (or the former C. albidus infections), and these patients had high mortality rates (40%) despite antifungal treatment. The isolates came from the bloodstream in 9 cases, CNS in 6 cases, lower respiratory tract in 5 cases, and a single sample from peritoneal fluid. Seven cases of non-invasive infections were also reported, including colonization of the skin and fungal keratitis. Furthermore, a case of catheter-related fungemia due to N. liquefaciens has been described in a 71-year-old man with B-cell lymphoma (Takemura et al., 2015). In addition, a CSF mixed infection caused by N. liquequefaciens and Mycobacterium tuberculosis species complex has been reported in an HIV-positive female patient who died despite antifungal and antibacterial therapy. The final identification of this isolate was only possible with the sequencing of the D1/D2 domains of the large subunit of the rRNA region, since ITS rDNA sequencing was not able to differentiate between N. liquefaciens and N. albidosimilis (Conde-Pereira et al., 2015) and similar to our findings.
All C. gattii/C. neoformans species complex strains (clades I and II) isolates were able to grow at 37°C, but the high temperature growth phenotype was not observed for non-C. gattii/C. neoformans species complex strains (clades III to V), even if they were clinical isolates (LMMM1398, 1460 and 221). It is widely documented that the ability to grow at human physiological temperature is an important virulence factor for Cryptococcus spp. (Zaragoza, 2019). Nevertheless, other opportunistic fungi such as Rhodotorula spp. grow poorly in vitro in temperatures above 33°C, and yet, can cause human infections such as fungemia (Ioannou et al., 2019) and may also belong to the human gut microbiota (Borges et al., 2018).
It is very clear that C. gattii/neoformans species complex isolates (clades I and II), in general, show more similar virulence phenotypes among each other when compared to non-C. gattii/C. neoformans species complex isolates (clades III to V) and vice versa. However, occasionally, P. laurentii (clade III) strains do behave closer to C. gattii/neoformans species complex isolates (clades I and II) such as with phospholipase and hemolysins production. While clades I and II strains have thicker capsules, produced more melanin and formed greater biofilms, clades III to V strains were able to produce the enzymatic activities of phospholipase and hemolysins. However, it is important to emphasize that because we do not have a balanced number of clinical and environmental strains for each clade, comparisons were performed independently of the source of the strain isolation We recognize that if more clinical strains of P. laurentii and Naganishia spp. (clades III to V) or environmental strains of C. gattii/neoformans species complex isolates (clades I and II) were included, our results could have been different because of strain adaptations to different ecological niches. Therefore, we recognize this fact as a limitation of our study.
Ferreira-Paim et al. (2012) were the first to describe hemolysin production by the former C. laurentii isolates obtained from pigeon droppings. The fact that non-C. gattii/neoformans species complex strains are able to efficiently produce hemolysins may partially explain the reason why these species are more frequently described in cases of fungemia, rather than meningeal or deep-seated tissue infections (Londero et al., 2019). Conversely, phospholipase production has been largely reported for C. gattii/C. neoformans species complex strains (Pini et al., 2017). This enzyme in C. neoformans is thought to trigger capsule enlargement, reduce phagocytosis by macrophages (Chrisman et al., 2011) and secreted phospholipase B (PLB) activity favors the survival and replication of C. neoformans in macrophages in vitro, but it seems that this enzyme is not directly involved in the establishment of CNS infections (Santangelo et al., 2004). Interestingly, Pedroso et al. (2009) reported greater phospholipase activity in C. albidus than in C. laurenttii isolates recovered from several organs in a murine model of infection, consistent with our findings. However, it appears that these extracellular enzymes may be critical in breaking down the environmental milieu rather than participating in mammalian host tissue invasion.
Most of the clinical isolates when grown on NSA had colonies that were darkly pigmented (scores 3 to 4), including the non-C. gattii/C. neoformans species complex clinical isolates (LMMM1398, LMMM1460 and LMM221) and supports the importance of this as a mammalian virulence factor. Furthermore, melanin production is crucial for Cryptococcus spp. virulence, since mutants unable to produce this pigment have a significant attenuation in virulence (Salas et al., 1996). Melanin confers resistance to oxidative stress, heat and ionizing radiation (Zaragoza, 2019), besides impairing proper antifungal drug actions (Nosanchuk and Casadevall, 2006).
In our biofilm formation experiments, C. deuterogatti (clade I) was more efficiently able to form biofilm than C. neoformans (clade II) strains. Tavares et al. (2019) recently investigated C. gattii (VGI) kinetics of biofilm formation and described its structure as a dense network of yeast cells deeply encased in an extracellular polymeric matrix (Tavares et al., 2019). In terms of clinical significance, C. gattii species are frequently described in the immunocompetent host, cause more damage to the lungs and frequently produce cryptococcomas, which are considered biofilm-like structures (Chen et al., 2014; Aslanyan et al., 2017; Rodriguez-Goncer et al., 2018). In our study, there was also trend of greater biofilm formation in P. laurentii (clade III) than in Naganishia spp. (clades IV and V), but the three clades together were considered significantly less effective in biofilm formation than C. gattii/C. neoformans species complex strains (clades I and II). However, P. laurentii biofilm was recently described to have dense growth and colonization with extensive polymeric substances around the yeast cells (Ajesh and Sreejith, 2012).
C. deuterogatti (clade I) strains showed a large variation in capsule sizes, with some of the isolates possessing the largest capsules in the present study. Giant yeast cells (capsule thickness ≥ 15 µm) and variation in capsule thickness are more common in C. gattii complex than in the C. neoformans complex. However, both “large” and “small” capsular phenotypes are important in different stages of infection (early and late, respectively), reinforcing the advantage of strains with plasticity in capsule and cell sizes to cause cryptococcosis (Fernandes et al., 2018). The non-Cryptococcus gattii/neoformans species complex strains (clades III to V) had markedly thinner capsules. However, Araujo et al., (Araujo et al., 2017) described that polysaccharides of C. liquefaciens have strikingly similar ultrastructural and biological properties to those of C. neoformans, and found that this species led to mortality rates similar to C. neoformans in a virulence model with the larvae of Galleria melonella, reinforcing its pathogenic potential.
There are no antifungal breakpoints yet established for Cryptococcal species so discussions about drug susceptible and resistant isolates need to wait until breakpoints are determined. However, it is clear from our results most strains fit under the ECVs established for C. deuterogattii (clade I) and C. neoformans (clade II) and surely do not possess high-level resistance since azole MICs are within achievable drug levels in the blood. On the other hand, the isolates have some variability in their azole susceptibility. For the polyene in vitro activity, some of the MICs are relatively high and this may be more in vitro media importance rather than clinical importance. In a Chinese multicenter study almost 24% of 312 C. neoformans strains had non-wild-type phenotypes to fluconazole with 5 isolates possessing had MICs≥ 32 µg/mL (Ajesh and Sreejith, 2012) but Pfaller et al. with over 1,811 clinical isolates reported only 1% with very high MICs. Ferreira and Pahim (Ferreira-Paim et al., 2012) found that 71% of 38 environmental C. laurentii isolates had elevated azole MICs and Kantarcioglu et al. (2009) showed decreased susceptibility of 7/11 C. diffluens to both azoles and polyenes. Both our data and others suggest that azoles and polyenes will likely have clinical activity against both species complexes, the C. gattii/neoformans and non-C. gattii/neoformans species complex. However, some isolates push toward the higher antifungal MICs especially with the non-C. gattii/neoformans species complex strains which also possess less clinical treatment experience. Therefore, assessment of in vitro susceptibility for these isolates may be helpful to the clinician during clinical management.
In conclusion, we have shown different phenotypic traits between C. gattii/neoformans species complex (clades I and II) and other related genera such as Papiliotrema (clade III) and Naganishia (clades IV and V). In addition, P. laurentii (clade III) shows phenotypic properties more similar to C. gattii/C. neoformans species complex isolates (clades I and II), when compared to Naganishia spp (clades IV and V). But Naganishia spp. produced higher enzymatic levels of phospholipase and hemolysins. Furthermore, C. gattii/C. neoformans species complex (clades I and II) isolates had markedly larger capsules, produced more biofilm biomass and melanin. In addition, more isolates belonging to non- Cryptococcus gattii/neoformans species complex (clades III to V) possessed higher MICs to fluconazole, itraconazole and amphotericin B than those belonging to the C. gattii/neoformans species complex (clades I and II). However, 40% of C. neoformans strains had MICs equal to or above the ECV to amphotericin B. C. deuterogattii and C. neoformans showed very similar phenotypes among themselves. However, C. deuterogatti strains (clade I) produced statistically significant more biofilm biomass than C. neoformans (clade II). Another interesting observation is that all the non-C. gattii/neoformans clinical isolates (clades III to V) produced darker colonies, suggesting higher melanin production, when compared to their environmental counterparts. More isolates are needed to determine if there is an association of melanin production with non-C. gattii/C. neoformans clinical isolates. These findings, together with the fact that P. laurentii and Naganishia spp. strains were collected outside of a University Hospital, could be important for environmental to patient transmission that produces nosocomial infections especially in immunocompromised patients.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Conceptualization, GC. Methodology, GC, LO, LP, MM, DT, and JT. Software, AP and WS. Formal analysis, WS and GC. Investigation, EP, RN, RL, and TB. Resources, WS and AP. Data curation, LO, WS, and AP. Writing—original draft preparation, GC, JP, and JT. Writing review and editing, GC, JP, and JT. Supervision, EM, RN, RL and TB. Project administration, GC. Funding acquisition, GC and JP. All authors contributed to the article and approved the submitted version.
This research was funded by Public Service Grants (USA) from NIAID (AI-73896 and AI-93257) and from the “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)”– Finance Code 001, Brazil (Ministry of Education). L. S. S. O. received a scholarship from CAPES. GC, RL, and RN are research productivity fellows of the “Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)”, Brazilian Ministry of Science and Technology.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2021.642658/full#supplementary-material
Ajesh, K., Sreejith, K. (2012). Cryptococcus Laurentii Biofilms: Structure, Development and Antifungal Drug Resistance. Mycopathologia. doi: 10.1007/s11046-012-9575-2
Alves, G. S., Freire, A. K., Bentes Ados, S., Pinheiro, J. F., de Souza, J. V., Wanke, B., et al. (2016). Molecular Typing of Environmental Cryptococcus Neoformans/C. Gattii Species Complex Isolates From Manaus, Amazonas, Brazil. Mycoses 59 (8), 509–515. doi: 10.1111/myc.12499
Andrade-Silva, L., Ferreira-Paim, K., Silva-Vergara, M. L., Pedrosa, A. L. (2010). Molecular Characterization and Evaluation of Virulence Factors of Cryptococcus Laurentii and Cryptococcus Neoformans Strains Isolated From External Hospital Areas. Fungal Biol. 114 (5–6), 438–445. doi: 10.1016/j.funbio.2010.03.005
Araujo, G. R. S., Freitas, G. J. C., Fonseca, F. L., Leite, P. E. C., Rocha, G. M., de Souza, W., et al. (2017). The Environmental Yeast Cryptococcus Liquefaciens Produces Capsular and Secreted Polysaccharides With Similar Pathogenic Properties to Those of C. Neoformans. Sci. Rep. 7, 46768. doi: 10.1038/srep46768
Araujo Gde, S., Fonseca, F. L., Pontes, B., Torres, A., Cordero, R. J., Zancope-Oliveira, R. M., et al. (2012). Capsules From Pathogenic and non-Pathogenic Cryptococcus Spp. Manifest Significant Differences in Structure and Ability to Protect Against Phagocytic Cells. PloS One 7 (1), e29561. doi: 10.1371/journal.pone.0029561PONE-D-11-17932
Aslanyan, L., Sanchez, D. A., Valdebenito, S., Eugenin, E. A., Ramos, R. L., Martinez, L. R. (2017). The Crucial Role of Biofilms in Cryptococcus Neoformans Survival Within Macrophages and Colonization of the Central Nervous System. J. Fungi. 3 (1), 10 doi: 10.3390/jof3010010
Averbuch, D., Boekhoutt, T., Falk, R., Engelhard, D., Shapiro, M., Block, C., et al. (2002). Fungemia in a Cancer Patient Caused by Fluconazole-Resistant Cryptococcus Laurentii. Med. Mycol 40 (5), 479–484. doi: 10.1080/mmy.40.5.479.484
Baumgartner, C., Freydiere, A. M., Gille, Y. (1996). Direct Identification and Recognition of Yeast Species From Clinical Material by Using Albicans ID and Chromagar Candida Plates. J. Clin. Microbiol. 34 (2), 454–456. doi: 10.1128/JCM.34.2.454-456.1996
Borges, F. M., de Paula, T. O., Sarmiento, M. R. A., de Oliveira, M. G., Pereira, M. L. M., Toledo, I. V., et al. (2018). Fungal Diversity of Human Gut Microbiota Among Eutrophic, Overweight, and Obese Individuals Based on Aerobic Culture-Dependent Approach. Curr. Microbiol. 75 (6), 726–735. doi: 10.1007/s00284-018-1438-810.1007/s00284-018-1438-8
Brito, M. O., Bessa, M. A. S., Menezes, R. P., Roder, D. V. D. B., Penatti, M. P. A., Pimenta, J. P., et al. (2019). Isolation of Cryptococcus Species From the External Environments of Hospital and Academic Areas. J. Infect. Dev. Ctries 13 (6), 545–535. doi: 10.3855/jidc.10849
Castro-Lainez, M. T., Deliz-Aguirre, R., Antunez, D., Cruz-Codina, M., Cahuayme-Zuniga, L., Vitale, K., et al. (2019). Cryptococcus Laurentii Meningitis in a non-HIV Patient. IDCases 18, e00612. doi: 10.1016/j.idcr.2019.e00612S2214-2509(19)30097-6[pii]e00612
Chaves, G. M., Bates, S., Maccallum, D. M., Odds, F. C. (2007). Candida Albicans Grx2, Encoding a Putative Glutaredoxin, is Required for Virulence in a Murine Model. Genet. Mol. Res. 6 (4), 1051–1063.
Chee, H. Y., Lee, K. B. (2005). Isolation of Cryptococcus Neoformans Var. Grubii (Serotype a) From Pigeon Droppings in Seoul, Korea. J. Microbiol. 43 (5), 469–472.
Cheng, M. F., Chiou, C. C., Liu, Y. C., Wang, H. Z., Hsieh, K. S. (2001). Cryptococcus Laurentii Fungemia in a Premature Neonate. J. Clin. Microbiol. 39 (4), 1608–1611. doi: 10.1128/JCM.39.4.1608-1611.2001
Chen, S. C., Meyer, W., Sorrell, T. C. (2014). Cryptococcus Gattii Infections. Clin. Microbiol. Rev. 27 (4), 980–1024. doi: 10.1128/CMR.00126-13
Choe, Y. J., Blatt, D. B., Yalcindag, A., Geffert, S. F., Bobenchik, A. M., Michelow, I. C. (2020). Cryptococcus Albidus Fungemia in an Immunosuppressed Child: Case Report and Systematic Literature Review. J. Pediatr. Infect. Dis. Soc. 9 (1), 100–105. doi: 10.1093/jpids/piz039
Chrisman, C. J., Albuquerque, P., Guimaraes, A. J., Nieves, E., Casadevall, A. (2011). Phospholipids Trigger Cryptococcus Neoformans Capsular Enlargement During Interactions With Amoebae and Macrophages. PloS Pathog. 7 (5), e1002047. doi: 10.1371/journal.ppat.1002047PPATHOGENS-D-10-00314
Cleveland, K. O., Gelfand, M. S., Rao, V. (2013). Posaconazole as Successful Treatment for Fungemia Due to Cryptococcus Albidus in a Liver Transplant Recipient. QJM 106 (4), 361–362. doi: 10.1093/qjmed/hcs133
CLSI. (2008). “Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard—Third Edition (M27-A3)”. (Wayne, PA). Available at: https://clsi.org/standards/products/microbiology/documents/m27/.
CLSI. (2020). “Epidemiological Cutoff Values for Antifungal Susceptibility Testing. Clsi Supplement M59”. 3 rd editi, vol. 3rd edition. (Waine PA).
CLSI, Clinical and Laboratory Standards Institute. (2012). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Fourth Informational Supplement (Wayne: Clinical and Laboratory Standards Institute), (Document M27–S4).
Cogliati, M. (2013). Global Molecular Epidemiology of Cryptococcus Neoformans and Cryptococcus Gattii: An Atlas of the Molecular Types. Scientifica (Cairo) 2013, 75213. doi: 10.1155/2013/675213
Conde-Pereira, C., Rodas-Rodriguez, L., Diaz-Paz, M., Palacios-Rivera, H., Firacative, C., Meyer, W., et al. (2015). Fatal Case of Polymicrobial Meningitis Caused by Cryptococcus Liquefaciens and Mycobacterium Tuberculosis Complex in a Human Immunodeficiency Virus-Infected Patient. J. Clin. Microbiol. 53 (8), 2753–2755. doi: 10.1128/JCM.00840-15
Desjardins, C. A., Giamberardino, C., Sykes, S. M., Yu, C. H., Tenor, J. L., Chen, Y., et al. (2017). Population Genomics and the Evolution of Virulence in the Fungal Pathogen Cryptococcus Neoformans. Genome Res. 27 (7), 1207–1219. doi: 10.1101/gr.218727.116
Farrer, R. A., Chang, M., Davis, M. J., van Dorp, L., Yang, D. H., Shea, T., et al. (2019). A New Lineage of Cryptococcus Gattii (Vgv) Discovered in the Central Zambezian Miombo Woodlands. MBio 10 (6). doi: 10.1128/mBio.02306-19
Felsenstein, J. (1985). Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 39 (4), 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Fernandes, K. E., Brockway, A., Haverkamp, M., Cuomo, C. A., van Ogtrop, F., Perfect, J. R., et al. (2018). Phenotypic Variability Correlates With Clinical Outcome in Cryptococcus Isolates Obtained From Botswanan Hiv/Aids Patients. MBio 9 (5). doi: 10.1128/mBio.02016-18
Ferreira-Paim, K., Andrade-Silva, L., Fonseca, F. M., Ferreira, T. B., Mora, D. J., Andrade-Silva, J., et al. (2017). Mlst-Based Population Genetic Analysis in a Global Context Reveals Clonality Amongst Cryptococcus Neoformans Var. Grubii VNI Isolates From HIV Patients in Southeastern Brazil. PloS Negl. Trop. Dis. 11 (1), e0005223. doi: 10.1371/journal.pntd.0005223PNTD-D-16-00985
Ferreira-Paim, K., Andrade-Silva, L., Mora, D. J., Lages-Silva, E., Pedrosa, A. L., da Silva, P. R., et al. (2012). Antifungal Susceptibility, Enzymatic Activity, Pcr-Fingerprinting and its Sequencing of Environmental Cryptococcus Laurentii Isolates From Uberaba, Minas Gerais, Brazil. Mycopathologia 174 (1), 41–52. doi: 10.1007/s11046-011-9500-0
Ferreira-Paim, K., Andrade-Silva, L., Mora, D. J., Pedrosa, A. L., Rodrigues, V., Silva-Vergara, M. L. (2011). Genotyping of Cryptococcus Neoformans Isolated From Captive Birds in Uberaba, Minas Gerais, Brazil. Mycoses 54 (5), e294–e300. doi: 10.1111/j.1439-0507.2010.01901.x
Freire, A. K., dos Santos Bentes, A., de Lima Sampaio, I., Matsuura, A. B., Ogusku, M. M., Salem, J. I., et al. (2012). Molecular Characterisation of the Causative Agents of Cryptococcosis in Patients of a Tertiary Healthcare Facility in the State of Amazonas-Brazil. Mycoses 55 (3), e145–e150. doi: 10.1111/j.1439-0507.2012.02173.x
Furman-Kuklinska, K., Naumnik, B., Mysliwiec, M. (2009). Fungaemia Due to Cryptococcus Laurentii as a Complication of Immunosuppressive Therapy–a Case Report. Adv. Med. Sci. 54 (1), 116–119. doi: 10.2478/v10039-009-0014-7
Gullo, F. P., Rossi, S. A., Sardi Jde, C., Teodoro, V. L., Mendes-Giannini, M. J., Fusco-Almeida, A. M. (2013). Cryptococcosis: Epidemiology, Fungal Resistance, and New Alternatives for Treatment. Eur. J. Clin. Microbiol. Infect. Dis. 32 (11), 1377–1391. doi: 10.1007/s10096-013-1915-8
Hagen, F., Khayhan, K., Theelen, B., Kolecka, A., Polacheck, I., Sionov, E., et al. (2015). Recognition of Seven Species in the Cryptococcus Gattii/Cryptococcus Neoformans Species Complex. Fungal Genet. Biol. 78, 16–48. doi: 10.1016/j.fgb.2015.02.009
Hagen, F., Lumbsch, H. T., Arsic Arsenijevic, V., Badali, H., Bertout, S., Billmyre, R. B., et al. (2017). Importance of Resolving Fungal Nomenclature: The Case of Multiple Pathogenic Species in the Cryptococcus Genus. MSphere 2 (4). doi: 10.1128/mSphere.00238-17mSphere00238-17
Ikeda, R., Sugita, T., Jacobson, E. S., Shinoda, T. (2002). Laccase and Melanization in Clinically Important Cryptococcus Species Other Than Cryptococcus Neoformans. J. Clin. Microbiol. 40 (4), 1214–1218. doi: 10.1128/jcm.40.4.1214-1218.2002
Ioannou, P., Vamvoukaki, R., Samonis, G. (2019). Rhodotorula Species Infections in Humans: A Systematic Review. Mycoses 62 (2), 90–100. doi: 10.1111/myc.12856
Irokanulo, E. O., Makinde, A. A., Akuesgi, C. O., Ekwonu, M. (1997). Cryptococcus Neoformans Var Neoformans Isolated From Droppings of Captive Birds in Nigeria. J. Wildl. Dis. 33 (2), 343–345. doi: 10.7589/0090-3558-33.2.343
Johnson, L. B., Bradley, S. F., Kauffman, C. A. (1998). Fungaemia Due to Cryptococcus Laurentii and a Review of non-Neoformans Cryptococcaemia. Mycoses 41 (7–8), 277–280. doi: 10.1111/j.1439-0507.1998.tb00338.x
Kamari, A., Sepahvand, A., Mohammadi, R. (2017). Isolation and Molecular Characterization of Cryptococcus Species Isolated From Pigeon Nests and Eucalyptus Trees. Curr. Med. Mycol. 3 (2), 20–25. doi: 10.29252/cmm.3.2.20
Kantarcioglu, A. S., Boekhout, T., Yucel, A., Altas, K. (2009). Susceptibility Testing of Cryptococcus Diffluens Against Amphotericin B, Flucytosine, Fluconazole, Itraconazole, Voriconazole and Posaconazole. Med. Mycol. 47 (2), 169–176. doi: 10.1080/13693780802213407
Kassi, F. K., Bellet, V., Drakulovski, P., Krasteva, D., Roger, F., Valerie, B. A., et al. (2018). Comparative Typing Analyses of Clinical and Environmental Strains of the Cryptococcus Neoformans/Cryptococcus Gattii Species Complex From Ivory Coast. J. Med. Microbiol. 67 (1), 87–96. doi: 10.1099/jmm.0.000654
Khawcharoenporn, T., Apisarnthanarak, A., Mundy, L. M. (2007). Non-Neoformans Cryptococcal Infections: A Systematic Review. Infection 35 (2), 51–58. doi: 10.1007/s15010-007-6142-8
Kidd, S. E., Chow, Y., Mak, S., Bach, P. J., Chen, H., Hingston, A. O., et al. (2007). Characterization of Environmental Sources of the Human and Animal Pathogen Cryptococcus Gattii in British Columbia, Canada, and the Pacific Northwest of the United States. Appl. Environ. Microbiol. 73 (5), 1433–1443. doi: 10.1128/AEM.01330-06
Kimura, M. (1980). A Simple Method for Estimating Evolutionary Rate of Base Substitutions Through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 16, 111–120. doi: 10.1007/BF01731581
Kumar, S., Stecher, G., Li, M., Knyaz, C., Tamura, K. (2018). Mega X: Molecular Evolutionary Genetics Analysis Across Computing Platforms. Mol. Biol. Evol. 35 (6), 1547–1549. doi: 10.1093/molbev/msy096
Kwon-Chung, J. K., Bennett, J. E. (1992). “Cryptococcosis”. in Medical Mycology (Pennsylvania: Lea &Febiger), 397–446.
Kwon-Chung, K. J., Fraser, J. A., Doering, T. L., Wang, Z., Janbon, G., Idnurm, A., et al. (2014). Cryptococcus Neoformans and Cryptococcus Gattii, the Etiologic Agents of Cryptococcosis. Cold Spring Harb. Perspect. Med. 4 (7), a019760. doi: 10.1101/cshperspect.a019760
Lakoh, S., Rickman, H., Sesay, M., Kenneh, S., Burke, R., Baldeh, M., et al. (2020). Prevalence and Mortality of Cryptococcal Disease in Adults With Advanced HIV in an Urban Tertiary Hospital in Sierra Leone: A Prospective Study. BMC Infect. Dis. 20 (1), 141. doi: 10.1186/s12879-020-4862-x10.1186/s12879-020-4862-x
Lazera, M. S., Cavalcanti, M. A., Trilles, L., Nishikawa, M. M., Wanke, B. (1998). Cryptococcus Neoformans Var. Gattii–Evidence for a Natural Habitat Related to Decaying Wood in a Pottery Tree Hollow. Med. Mycol. 36 (2), 119–122. doi: 10.1080/02681219880000191
Liu, X. Z., Wang, Q. M., Goker, M., Groenewald, M., Kachalkin, A. V., Lumbsch, H. T., et al. (2015). Towards an Integrated Phylogenetic Classification of the Tremellomycetes. Stud. Mycol. 81, 85–147. doi: 10.1016/j.simyco.2015.12.001
Londero, M. R., Zanrosso, C. D., Corso, L. L., Michelin, L., Soldera, J. (2019). Catheter-Related Infection Due to Papiliotrema Laurentii in an Oncologic Patient: Case Report and Systematic Review. Braz. J. Infect. Dis. 23 (6), 451–461. doi: 10.1016/j.bjid.2019.10.005
Loperena-Alvarez, Y., Ren, P., Li, X., Schoonmaker-Bopp, D. J., Ruiz, A., Chaturvedi, V., et al. (2010). Genotypic Characterization of Environmental Isolates of Cryptococcus Gattii From Puerto Rico. Mycopathologia 170 (4), 279–285. doi: 10.1007/s11046-010-9296-3
Luo, G., Samaranayake, L. P., Yau, J. Y. (2001). Candida Species Exhibit Differential In Vitro Hemolytic Activities. J. Clin. Microbiol. 39 (8), 2971–2974. doi: 10.1128/JCM.39.8.2971-2974.2001
Mancianti, F., Nardoni, S., Ceccherelli, R. (2002). Occurrence of Yeasts in Psittacines Droppings From Captive Birds in Italy. Mycopathologia 153 (3), 121–124. doi: 10.1023/A:1014576304894
Maruyama, F. H., de Paula, D. A. J., Menezes, I. G., Favalessa, O. C., Hahn, R. C., Adbpf de Almeida, V. R. F., et al. (2019). Genetic Diversity of the Cryptococcus Gattii Species Complex in Mato Grosso State, Brazil. Mycopathologia 184 (1), 45–515. doi: 10.1007/s11046-018-0313-210.1007/s11046-018-0313-2
Maziarz, E. K., Perfect, J. R. (2016). Cryptococcosis. Infect. Dis. Clin. North Am. 30 (1), 179–206. doi: 10.1016/j.idc.2015.10.006
McCurdy, L. H., Morrow, J. D. (2003). Infections Due to non-Neoformans Cryptococcal Species. Compr. Ther. 29 (2–3), 95–101. doi: 10.1007/s12019-003-0012-9
Neves, R. P., Lima Neto, R. G., Leite, M. C., Silva, V. K., Santos Fde, A., Macedo, D. P. (2015). Cryptococcus Laurentii Fungaemia in a Cervical Cancer Patient. Braz. J. Infect. Dis. 19 (6), 660–663. doi: 10.1016/j.bjid.2015.06.014
Nosanchuk, J. D., Casadevall, A. (2006). Impact of Melanin on Microbial Virulence and Clinical Resistance to Antimicrobial Compounds. Antimicrob. Agents Chemother. 50 (11), 3519–3528. doi: 10.1128/AAC.00545-06
Palanco, A. C., Lacorte Singulani, J., Costa-Orlandi, C. B., Gullo, F. P., Strohmayer Lourencetti, N. M., Gomes, P. C., et al. (2017). Activity of 3’-Hydroxychalcone Against Cryptococcus Gattii and Toxicity, and Efficacy in Alternative Animal Models. Future Microbiol. 12, 1123–1134. doi: 10.2217/fmb-2017-0062
Park, B., Lockhart, S., Brandt, M., Chiller, T. (2011). “Public Health Importance of Cryptococcal Disease: Epidemiology, Burden, and Control. ” in Cryptococcus. Ed. Casadevall, A. (Washington: ASM Press), 585–593.
Park, B. J., Wannemuehler, K. A., Marston, B. J., Govender, N., Pappas, P. G., Chiller, T. M. (2009). Estimation of the Current Global Burden of Cryptococcal Meningitis Among Persons Living With HIV/AIDS. AIDS 23 (4), 525–530. doi: 10.1097/QAD.0b013e328322ffac
Pedroso, R. S., Ferreira, J. C., Lavrador, M. A., Maffei, C. M., Candido, R. C. (2009). Evaluation of the Experimental Inoculation of Cryptococcus Albidus and Cryptococcus Laurentii in Normal Mice: Virulence Factors and Molecular Profile Before and After Animal Passage. Mycopathologia 168 (2), 59–72. doi: 10.1007/s11046-009-9202-z
Peng, C. A., Gaertner, A. A. E., Henriquez, S. A., Fang, D., Colon-Reyes, R. J., Brumaghim, J. L., et al. (2018). Fluconazole Induces ROS in Cryptococcus Neoformans and Contributes to DNA Damage In Vitro. PloS One 13 (12), e0208471. doi: 10.1371/journal.pone.0208471PONE-D-18-24022
Pini, G., Faggi, E., Campisi, E. (2017). Enzymatic Characterization of Clinical and Environmental Cryptococcus Neoformans Strains Isolated in Italy. Rev. Iberoam Micol 34 (2), 77–82. doi: 10.1016/j.riam.2016.04.005
Price, M. F., Wilkinson, I. D., Gentry, L. O. (1982). Plate Method for Detection of Phospholipase Activity in Candida Albicans. Sabouraudia 20 (1), 7–14. doi: 10.1080/00362178285380031
Rodriguez-Goncer, I., Bongomin, F., Doran, H. M., Novak-Frazer, L., Masania, R., Moore, C. B., et al. (2018). A Case of Pulmonary Cryptococcoma Due to Cryptococcus Gattii in the United Kingdom. Med. Mycol. Case Rep. 21, 23–25. doi: 10.1016/j.mmcr.2018.03.007S2211-7539(18)30041-1
Saitou, N., Nei, M. (1987). The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 4 (4), 406–425. doi: 10.1093/oxfordjournals.molbev.a040454
Salas, S. D., Bennett, J. E., Kwon-Chung, K. J., Perfect, J. R., Williamson, P. R. (1996). Effect of the Laccase Gene CNLAC1, on Virulence of Cryptococcus Neoformans. J. Exp. Med. 184 (2), 377–386. doi: 10.1084/jem.184.2.377
Sanchez, A., Escandon, P., Castaneda, E. (2008). [In Vitro Determination of Virulence Factors Activity Associated With Several Cryptococcus Neoformans Clinical Isolates]. Rev. Iberoam Micol. 25 (3), 145–149. doi: 10.1016/s1130-1406(08)70034-2
Santangelo, R., Zoellner, H., Sorrell, T., Wilson, C., Donald, C., Djordjevic, J., et al. (2004). Role of Extracellular Phospholipases and Mononuclear Phagocytes in Dissemination of Cryptococcosis in a Murine Model. Infect. Immun. 72 (4), 2229–2239. doi: 10.1128/iai.72.4.2229-2239.2004
Santos Bentes, A.D., Wanke, B., Dos Santos Lazera, M., Freire, A. K. L., da Silva Junior, R. M., Rocha, D. F. S., et al. (2019). Cryptococcus Gattii Vgii Isolated From Native Forest and River in Northern Brazil. Braz. J. Microbiol. 50 (2), 495–500. doi: 10.1007/s42770-019-00066-610.1007/s42770-019-00066-6
Shankar, E. M., Kumarasamy, N., Bella, D., Renuka, S., Kownhar, H., Suniti, S., et al. (2006). Pneumonia and Pleural Effusion Due to Cryptococcus Laurentii in a Clinically Proven Case of AIDS. Can. Respir. J. 13 (5), 275–278. doi: 10.1155/2006/160451
Smith, N., Sehring, M., Chambers, J., Patel, P. (2017). Perspectives on non-Neoformans Cryptococcal Opportunistic Infections. J. Community Hosp Intern. Med. Perspect. 7 (4), 214–217. doi: 10.1080/20009666.2017.13500871350087
Soltani, M., Bayat, M., Hashemi, S. J., Zia, M., Pestechian, N. (2013). Isolation of Cryptococcus Neoformans and Other Opportunistic Fungi From Pigeon Droppings. J. Res. Med. Sci. 18 (1), 56–60.
Takemura, H., Ohno, H., Miura, I., Takagi, T., Ohyanagi, T., Kunishima, H., et al. (2015). The First Reported Case of Central Venous Catheter-Related Fungemia Caused by Cryptococcus Liquefaciens. J. Infect. Chemother. 21 (5), 392–394. doi: 10.1016/j.jiac.2014.11.007
Tavares, E. R., Gionco, B., Morguette, A. E. B., Andriani, G. M., Morey, A. T., do Carmo, A. O., et al. (2019). Phenotypic Characteristics and Transcriptome Profile of Cryptococcus Gattii Biofilm. Sci. Rep. 9 (1), 6438. doi: 10.1038/s41598-019-42896-210.1038/s41598-019-42896-2
White, T., Burns, T., Lee, S., Taylor, T. (1990). “Amplification and Direct Sequencing of Fungal Ribosomal RRNA Genes for Phylogenetics.” in Pcr Protocols. Eds. Innis, M. A., Gelfland, D. H., Sninsky, T. J. (San Diego: Academic Press), 315–322.
Yarrow, D. (1998). “Methods for the Isolation, Maintenance and Identification of Yeasts”. in The Yeasts, a Taxonomic Study, 4th ed. Eds. Kurtzman, C. P., Fell, J. W. (Amsterdam: Elsevier Science), 77–100.
Zaragoza, O. (2019). Basic Principles of the Virulence of Cryptococcus. Virulence 10 (1), 490–501. doi: 10.1080/21505594.2019.1614383
Zaragoza, O., Fries, B. C., Casadevall, A. (2003). Induction of Capsule Growth in Cryptococcus Neoformans by Mammalian Serum and CO(2). Infect. Immun. 71 (11), 6155–6164. doi: 10.1128/iai.71.11.6155-6164.2003
Keywords: Cryptococcus spp., Papiliotrema laurentii, Naganishia spp., virulence factors, antifungal drug susceptibility
Citation: Oliveira LSdS, Pinto LM, de Medeiros MAP, Toffaletti DL, Tenor JL, Barros TF, Neves RP, Neto RGdL, Milan EP, Padovan ACB, Rocha WPdS, Perfect JR and Chaves GM (2021) Comparison of Cryptococcus gattii/neoformans Species Complex to Related Genera (Papiliotrema and Naganishia) Reveal Variances in Virulence Associated Factors and Antifungal Susceptibility. Front. Cell. Infect. Microbiol. 11:642658. doi: 10.3389/fcimb.2021.642658
Received: 21 January 2021; Accepted: 10 June 2021;
Published: 01 July 2021.
Edited by:
Brian Wickes, The University of Texas Health Science Center at San Antonio, United StatesReviewed by:
Sudha Chaturvedi, Wadsworth Center, United StatesCopyright © 2021 Oliveira, Pinto, de Medeiros, Toffaletti, Tenor, Barros, Neves, Neto, Milan, Padovan, Rocha, Perfect and Chaves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guilherme Maranhão Chaves, Z3VpbGhlcm1lLmNoYXZlc0B1ZnJuZXQuYnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.