 Keri A. Lydon
Keri A. Lydon Thomas Kinsey
Thomas Kinsey Chinh Le2
Chinh Le2 Paul A. Gulig
Paul A. Gulig Jessica L. Jones
Jessica L. Jones
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 26 February 2021
Sec. Molecular Bacterial Pathogenesis
Volume 11 - 2021 | https://doi.org/10.3389/fcimb.2021.637019
This article is part of the Research Topic Vibrio Virulence Regulation and Host Interactions View all 13 articles
Vibrio vulnificus is a deadly human pathogen for which infections occur via seafood consumption (foodborne) or direct contact with wounds. Virulence is not fully characterized for this organism; however, there is evidence of biochemical and genotypic correlations with virulence potential. In this study, biochemical profiles and virulence genotype, based on 16S rRNA gene (rrn) and virulence correlated gene (vcg) types, were determined for 30 clinical and 39 oyster isolates. Oyster isolates were more biochemically diverse than the clinical isolates, with four of the 20 tests producing variable (defined as 20–80% of isolates) results. Whereas, for clinical isolates only mannitol fermentation, which has previously been associated with virulence potential, varied among the isolates. Nearly half (43%) of clinical isolates were the more virulent genotype (rrnB/vcgC); this trend was consistent when only looking at clinical isolates from blood. The majority (64%) of oyster isolates were the less virulent genotype (rrnA or AB/vcgE). These data were used to select a sub-set of 27 isolates for virulence testing with a subcutaneously inoculated, iron-dextran treated mouse model. Based on the mouse model data, 11 isolates were non-lethal, whereas 16 isolates were lethal, indicating a potential for human infection. Within the non-lethal group there were eight oyster and three clinical isolates. Six of the non-lethal isolates were the less virulent genotype (rrnA/vcgE or rrnAB/vcgE) and two were rrnB/vcgC with the remaining two of mixed genotype (rrnAB/vcgC and rrnB/vcgE). Of the lethal isolates, five were oysters and 11 were clinical. Eight of the lethal isolates were the less virulent genotype and seven the more virulent genotype, with the remaining isolate a mixed genotype (rrnA/vcgC). A discordance between virulence genotype and individual mouse virulence parameters (liver infection, skin infection, skin lesion score, and body temperature) was observed; the variable most strongly associated with mouse virulence parameters was season (warm or cold conditions at time of strain isolation), with more virulent strains isolated from cold conditions. These results indicate that biochemical profiles and genotype are not significantly associated with virulence potential, as determined by a mouse model. However, a relationship with virulence potential and seasonality was observed.
Vibrio vulnificus is a gram-negative opportunistic pathogen that is naturally found in shellfish and coastal brackish waters in warmer climates (Kaspar and Tamplin, 1993; Nilsson et al., 2003). It causes gastroenteritis, wound infections, or septicemia (Blake et al., 1979; Hlady and Klontz, 1996) through two primary routes: 1) consumption of raw shellfish, primarily oysters, and 2) exposure of open wounds to V. vulnificus (Shapiro et al., 1998; Oliver and Kaper, 2001). This bacterium is the deadliest foodborne pathogen with a case fatality rate greater than 30% (Hlady and Klontz, 1996; Shapiro et al., 1998; Scallan et al., 2011). Moreover, individuals who are immunocompromised have the greatest risk of mortality due to increased risk of sepsis (Strom and Paranjpye, 2000; Haq and Dayal, 2005). Although this pathogen has a low rate of infection (Strom and Paranjpye, 2000), it is important to investigate methods to evaluate the virulence potential of strains due to the severity of illness and high case fatality rate. Existing strategies to evaluate and mitigate the risks associated with V. vulnificus focus on the total population of the pathogen. However, there is evidence that not all strains have an equal potential to cause disease in humans, and identification of reliable markers of virulent populations would permit refinement of risk assessment models and mitigation efforts.
Biotyping and genetic markers are currently used to classify the virulence potential of V. vulnificus. There are three biotypes, defined by biochemical profiles, associated with virulence potential based on host specificity. Biotype 1 is associated with human infections (Warner and Oliver, 2008). Biotype 2 is associated with infections in eels and occasionally in humans (Veenstra et al., 1993; Amaro and Biosca, 1996). While biotype 3 has only been associated with wound infections of fish handlers (Bisharat et al., 1999). In addition, there are genetic markers used to subtype, primarily biotype 1, V. vulnificus isolates, which were developed based on source of strain isolation: clinical (i.e., from an ill individual) or environmental (e.g., shellfish, harvest water, etc.) (Nilsson et al., 2003). Rosche et al. (2005) identified two variants of the virulence correlated gene (vcg) which correlated with isolation source: clinical (vcgC) or environmental (vcgE). Additionally, an evaluation of the 16S rRNA gene (rrn) polymorphic variants identified two types, with rrnA primarily associated with environmental isolates and rrnB associated with clinical isolates (Nilsson et al., 2003; Vickery et al., 2007). The vcg and rrn genetic markers are often complementary to one another, with rrnB and vcgC most often identified in clinical isolates, and rrnA and vcgE genotypes appearing most often in environmental isolates (Han et al., 2009). Based on these strong associations between isolate source and gene variants, the genotype rrnA/vcgE is generally assumed to be less virulent, whereas rrnB/vcgC type strains are assumed to be more virulent (Jones et al., 2013). In addition, mannitol fermentation has been associated with the rrnB genotype (Drake et al., 2010), indicating this as a potential biochemical marker of virulence potential.
Subtyping and genotyping assays have served as a proxy for virulence potential based on the presumption that isolates from a clinical source are likely more virulent than environmental isolates, which has been largely supported in an animal model (Starks et al., 2000; DePaola et al., 2003). A subcutaneously (s.c.) inoculated iron-dextran treated mouse model has been used to evaluate virulence potential in V. vulnificus (Starks et al., 2000; DePaola et al., 2003; Thiaville et al., 2011). This model has revealed systemic infection and mortality presenting more often in mice injected with clinical strains (DePaola et al., 2003), while environmental strains appear to grow slower or are more easily attenuated by the mouse host (Starks et al., 2006). Thiaville et al. (2011) was one of the first studies that measured how virulence potential, as determined by the mouse model, relates to strain genotype on a large scale; however, the strain set selected for this study was somewhat limited, with oyster isolates from warmer months underrepresented. The study identified five virulence clusters associated with differing severity on type of mouse infection. Strains that caused systemic infection (liver) following skin infection were considered potentially lethal to humans, while less virulent strains (non-lethal) caused primarily skin infections. The study concluded that while vcgC was associated with virulence potential, it was not predictive of virulence in biotype 1 V. vulnificus strains (Thiaville et al., 2011). For example, some of the most virulent strains were of the vcgE genotype. Mouse models are not ideal assays due to resource and time requirements, as well as the ethical considerations; however, they remain the current gold standard for evaluating V. vulnificus virulence (Starks et al., 2000).
Previous studies have assumed that isolate source (i.e., clinical or environmental/food) is a reliable proxy for V. vulnificus virulence, but without specific testing they are intrinsically biased to that assumption. Investigations into the relationship of genotype and virulence potential, as measured in a mouse model, are limited by the number and diversity of the isolates examined. Most of the clinical isolates used in previous studies (Nilsson et al., 2003; DePaola et al., 2003; Vickery et al., 2007; Thiaville et al., 2011) were isolated in warm months (May–September), when most infections occur; however, the majority of environmental isolates for these studies were obtained from cooler months (October–April). Regardless, these studies indicate a relationship with (DePaola et al., 2003), but lack the predictive power to interpolate virulence potential from a common virulence genotypes (Thiaville et al., 2011). This raises questions about the utility of these existing typing schemes. Therefore, the current study aims to further investigate the relationship of genotype and virulence potential by utilizing a geographically and seasonally diverse set of V. vulnificus isolates. This isolate set was examined for partial biochemical profiles (API 20E), rrn and vcg genotypes, and virulence potential as determined through the s.c. inoculated iron-dextran treated mouse model, in order to determine if an association exists between virulence potential, biochemical phenotype, genotype, isolate source, and season of isolation.
A total of 69 V. vulnificus isolates were selected for this study. All V. vulnificus were isolated in 2006–2007 from various parts of the United States. Of these, 30 were isolated from ill patients as part of the Cholera and Other Vibrio Illness Surveillance (COVIS) program and were contributed by the Centers for Disease Control and Prevention (Table 1) and 39 were isolated from retail level raw oysters [Table 2; (DePaola et al., 2010)]. All isolates were purified and confirmed as V. vulnificus by real-time PCR as previously described (Kinsey et al., 2015). Isolates were stored in TSB + 30% glycerol at −80°C.
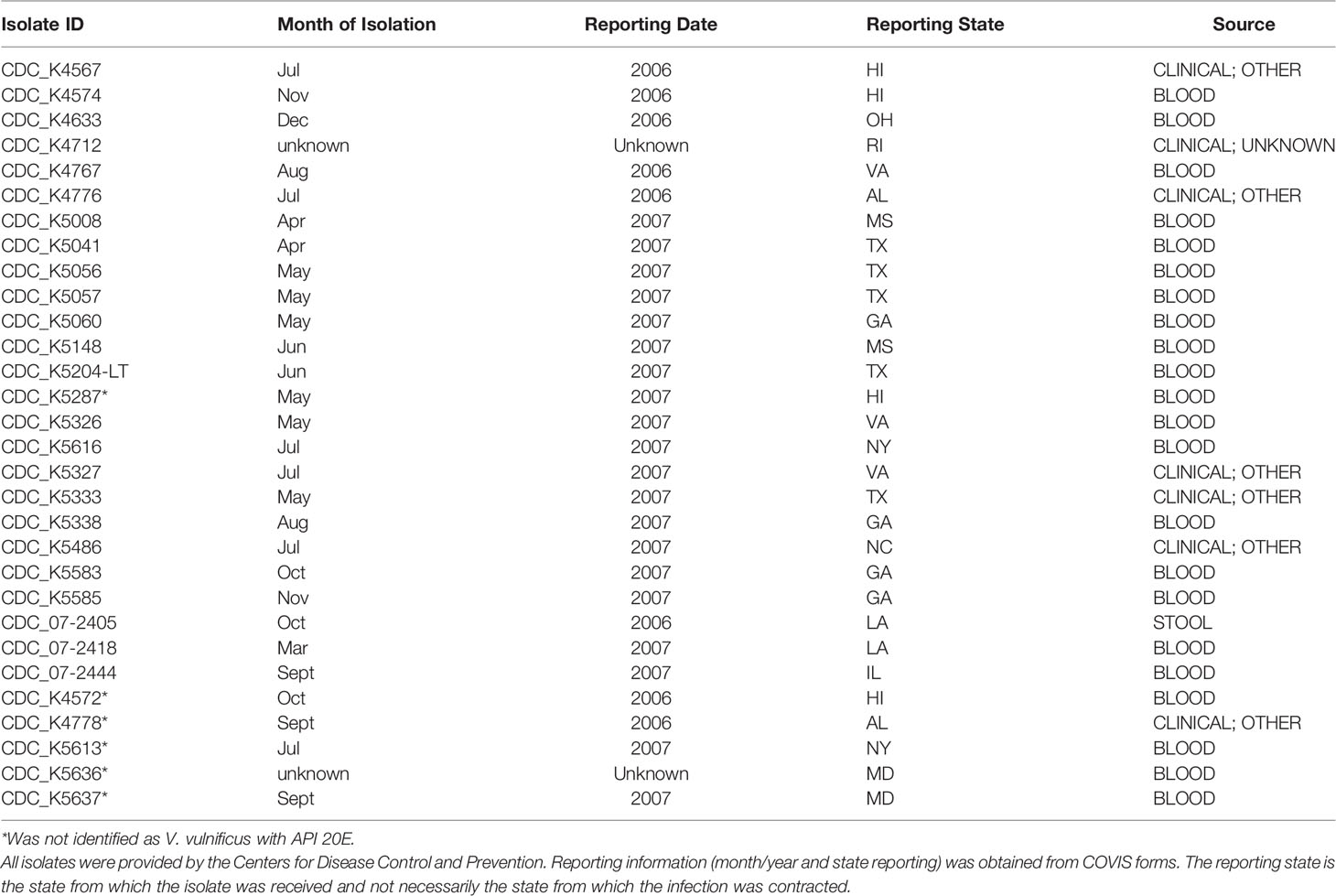
Table 1 Clinical Vibrio vulnificus isolates.
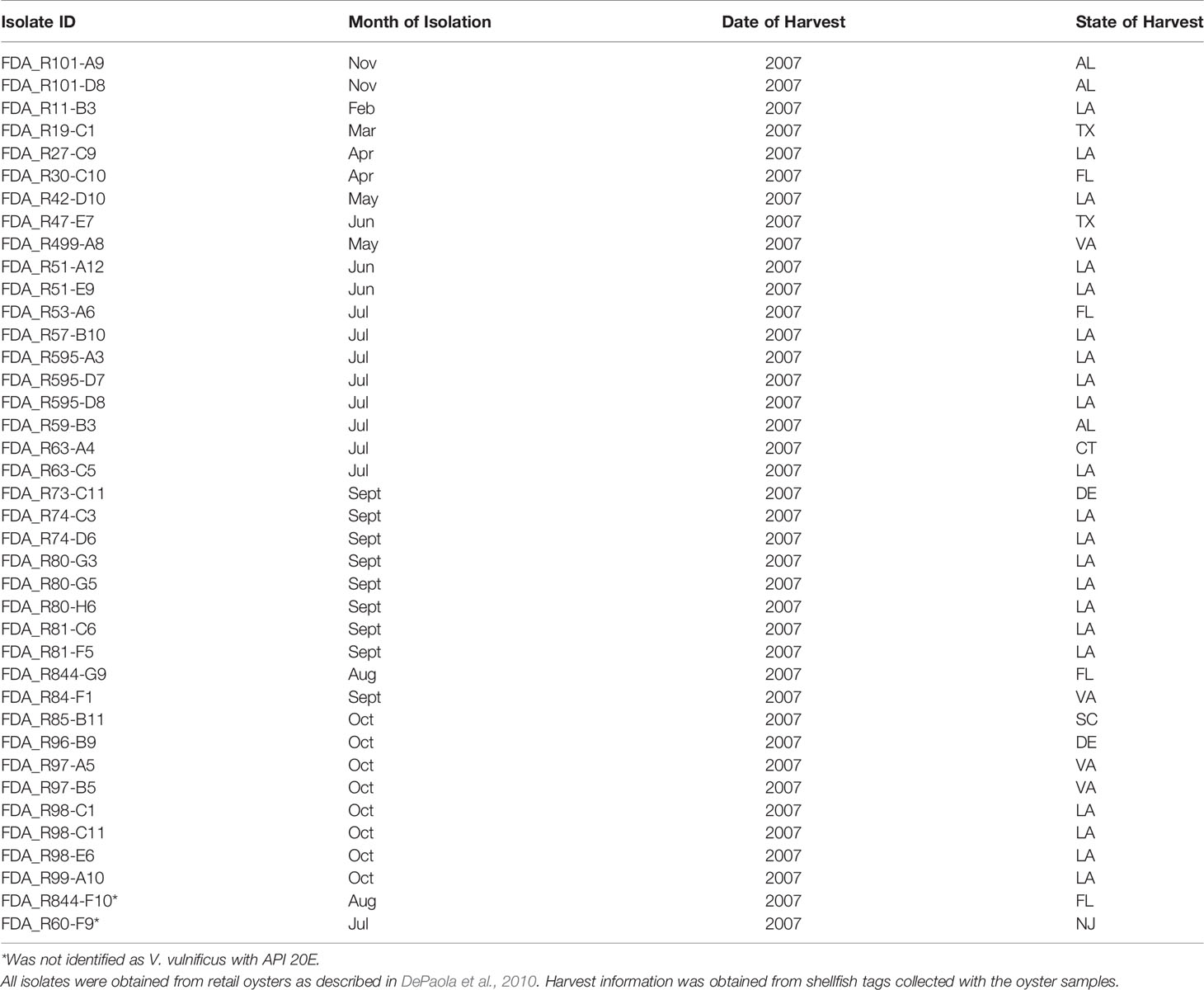
Table 2 Oyster Vibrio vulnificus isolates.
To evaluate partial biochemical profiles of the V. vulnificus isolates, API 20E (BioMerieux, Durham, NC) test strips were used according to the manufacturer’s protocol, except that 2% NaCl was used for cell suspensions (Kaysner et al., 2004; Martinez-Urtaza et al., 2005). Oxidase tests were completed using Dry Slides (BBL, Difco, Sparks, MD). API 20E results were entered into the manufacturer’s database for identification.
Isolates were streaked from frozen stocks to Tryptic Soy Agar (TSA; Difco) to confirm purity. A single colony was transferred to Tryptic Soy Broth (TSB; Remel, Atlanta, GA) and incubated at 35 ± 2°C for 18–24 h. One ml of the overnight culture was transferred to a microfuge tube and heated at 95–100°C for 10 min to produce a crude DNA lysate, which was used as template in subsequent real-time PCR assays. Isolate genotypes were determined using qualitative real-time PCR assays as previously described for 16S rRNA (rrn) gene type A or B (Vickery et al., 2007) and virulence correlated gene (vcg) type C or E (Drake et al., 2010) on a SmartCycler II System (Cepheid, Sunnyvale, CA). Results were used to define more (rrnB and vcgC) or less (rrnA or rrnAB and vcgE) virulent genotype categories.
For mouse virulence testing, 27 isolates were selected to be representative of source and genotype combinations. Approximately 1,000 CFU of each strain was inoculated into at least two groups of five mice, as previously described (Thiaville et al., 2011). Rectal temperature was used as an indicator of illness severity and as a surrogate for death (<33°C was determined to be dead) prior to sacrifice, when animals survived. Colony forming units (CFUs) were determined by standard plate count from the skin and liver following homogenization to determine local and systemic infection, respectively. Skin lesions were scored based on the size and nature of the lesion using a scale of 1–4. The skin and liver CFU data were used to cluster strains into virulence groups as previously described (Thiaville et al., 2011): Group 1 strains caused low skin and undetectable liver (systemic) infection, Group 2 strains caused moderate skin infection with little to no liver infection, Group 3 strains caused a high skin infection but low liver infection, Group 4 strains caused high skin and moderate liver infection, and Group 5 strains caused very high skin and very high liver infection. Assuming mouse virulence translates to human infection, Group 1, 2, and 3 strains would likely not be able to cause lethal infection in humans. Group 4 and 5 strains, because they cause high skin and moderate to high liver infection, have the potential to cause lethal infection in humans.
Data were analyzed by strain, using the mean results from all mice inoculated with that strain as the data point for each measured mouse virulence parameter to capture strain variability rather than individual mouse response. A three factor ANOVA (General Linear Model; GLM) was used to determine if interactions between source of isolation (clinical/oyster), genotype (more virulent/less virulent), or season of isolation (warm/cool) existed. As no significant interactions were found, GLM was used to evaluate quantitative data (log CFU/g skin, log CFU/g liver, body temperature, skin lesion score, and mortality) in comparison to the fixed variables. Fisher’s exact test was used to evaluate associations between isolate source, genotype, and season of isolation with biochemical reactions and lethal or non-lethal designation. Season of isolation was determined by grouping strains as either cool (October–April) or warm (May–September) seasons when V. vulnificus is historically less or more prevalent, respectively. Due to mixed genotype results, three isolates (FDA_R101-A9, CDC_K5148, and CDC_07-2444) were removed from statistical analyses in genotype comparisons. All statistical comparisons were conducted in JMP 13.
API 20E identified 61 isolates as V. vulnificus, with the eight remaining isolates identified as Vibrio spp. or Aeromonas spp. (Tables 1 and 2). As API 20E may incorrectly identify Vibrio spp. (O’Hara et al., 2003; Sanjuan et al., 2009), PCR targeting the vvh gene (Kinsey et al., 2015), was used to confirm all 69 isolates as V. vulnificus. All V. vulnificus isolates (n = 69) were positive for fermentation of glucose and amygdalin and the presence of oxidase and negative for arginine dihydrolase, tryptophan deaminase, inositol fermentation, sorbitol fermentation, rhamnose fermentation, and arabinose fermentation (Table 3).
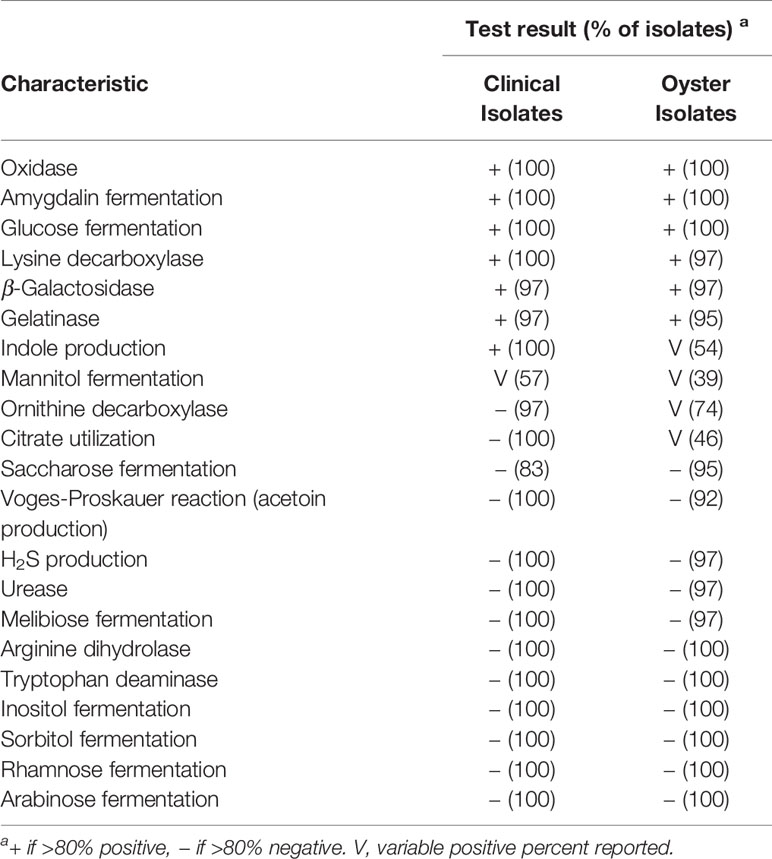
Table 3 Biochemical properties of V. vulnificus isolates.
Additionally, some of the tests were generally positive (β-Galactosidase, lysine decarboxylase, and gelatinase), while others were generally negative (H2S production, urease, Voges–Proskauer reaction, saccharose fermentation, and melibiose fermentation). This data may be useful in biochemical identification of V. vulnificus. Interestingly, there were three biochemical tests (ornithine decarboxylase, citrate utilization, and indole production) which were variable in oyster isolates, but generally present in clinical isolates, resulting in a statistically significant association between isolate source and these biochemical tests (p < 0.04). In this study, mannitol fermentation, which has been associated with virulent genotypes of V. vulnificus (Drake et al., 2010), was also significantly associated with the virulent genotypes (p < 0.001) here. Mannitol was the only biochemical reaction that was variable in both clinical and oyster isolates: 57% of clinical and 39% of oyster isolates were positive (Table 3).
For virulence genotyping of the clinical isolates (n = 30), 53% were the less virulent rrnA or AB/vcgE, and 47% were the more virulent (rrnB/vcgC) genotype (Table 4). Additionally, two clinical isolates had mixed genotypes: CDC_K5148 (rrnB/vcgE) and CDC_07-2444 (rrnA/vcgC). Of the strains isolated from blood infections (n = 22), half were the less virulent genotype and half the more virulent genotype. In the subset (n = 8) of clinical isolates not isolated from blood, half were the less virulent genotype, three were the more virulent genotype, and one was a mixed genotype (data not shown). Oyster isolate (n = 39) genotyping identified 64% of isolates as the less virulent genotype, rrnA or rrnAB/vcgE, and with a mixed genotype: FDA_R101-A9 (rrnAB/vcgC) and FDA_R63-C5 (rrnB/vcgE). The remaining oyster isolates (33%) were the more virulent (rrnB/vcgC) genotype.

Table 4 Genotype of V. vulnificus strains.
None of the V. vulnificus isolates in this study fell into mouse virulence Group 1, the least virulent group. Six isolates (22%) were classified as Group 2 (Table 5), with moderate skin infection (6.3–7.7 log CFU/g) and little to no liver infection (1.4–3.3 log CFU/g): five oyster and one clinical; five rrnA/vcgE and one mixed genotype (rrnAB/vcgE). Only one of these isolates, FDA_R51-A12 (rrnA/vcgE), caused mortality and all skin lesion scores were <3. Four isolates (15%) fell into Group 3: three oyster and one clinical; one rrnA/vcgE and three rrnB/vcgC. An additional clinical isolate, CDC_K5148 (rrnB/vcgE), was in its own novel classification, Group 6. However, based on its general characteristics, it is similar to Group 3, with high skin infection (7.8–8.5 log CFU/g) and low liver infection (2.8–3.7 log CFU/g for the Group 3 isolates and 1.4 log CFU/g for CDC_K5148). Only one of the Group 3 isolates, FDA_R73-C11 (rrnA/vcgE), caused mortality. All skin lesion scores were 3–4. Over half (56%) of the V. vulnificus isolates fell into Group 4: 5 oyster and 10 clinical; 8 rrnA or rrnAB/vcgE, 6 rrnB/vcgC, and 1 rrnA/vcgC. This group had high skin infection (7.2–8.3 log CFU/g) and moderate liver infection (4.4–6.0 log CFU/g). All strains caused mortality, ranging from 10% (FDA_R595-D7, rrnAB/vcgE) to 80% (CDC_07-2444, rrnA/vcgC). Skin lesion score for this group ranged from 2.8 to 4. One isolate, FDA_K5338 (rrnB/vcgC), was characterized as Group 5, with very high skin (8.5 log CFU/g) and very high liver (6.7 log CFU/g) infection.
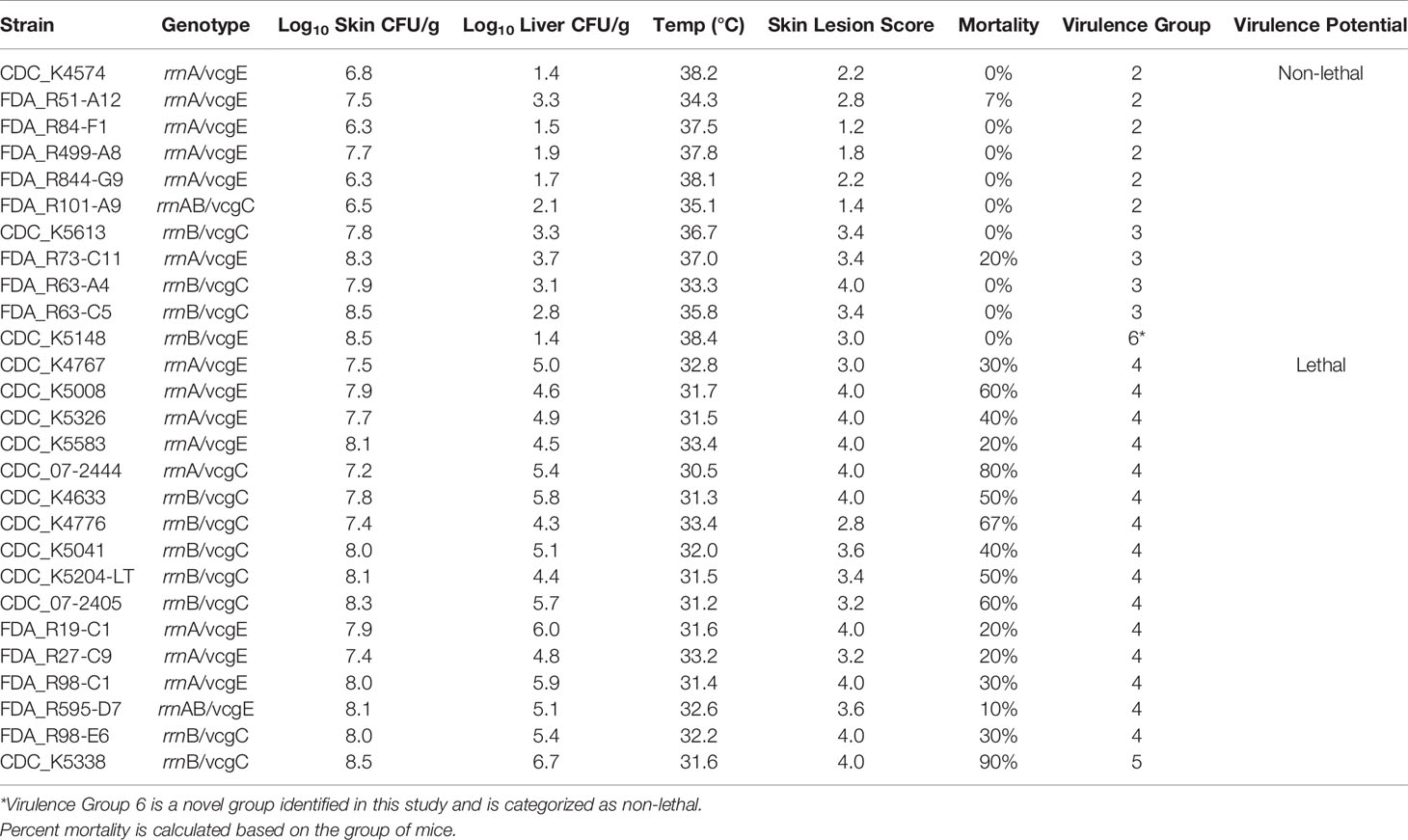
Table 5 Mouse virulence data. Skin infection, liver infection, body temperature, and skin lesion data are provided as means of the 5–10 animals tested with each strain.
Evaluation of the association between isolate source (clinical or oyster), virulence genotype (more virulent or less virulent), and season of strain isolation (cold or warm) with mouse virulence parameters including skin infection (log CFU/g skin), liver infection (log CFU/g liver), mouse temperature (as a proxy for illness severity), and mouse mortality (percentage of mice tested that died) were examined. The majority (6 of 11) of isolates with very high skin infection (≥8 log CFU/g) in the mouse model were the more virulent genotype or were of clinical origin. Similarly, isolates with very high liver (≥5 log CFU/g) infection were the more virulent genotype (five of 10) or were of clinical origin (six of 10).
However, there were no strong statistically significant relationships (p > 0.05) between the virulence genotype and skin infection, liver infection, mouse temperature, or mouse mortality (Figure 1). However, there was a weak association with virulence genotype and skin infection (p = 0.05). Similarly, no statistically significant (p > 0.05) relationships between isolate source and skin infection, liver infection, or mouse temperature were identified. There was a strong association between isolate source and mouse mortality, with clinical isolates causing significantly more mortality than oyster isolates (p = 0.003). Isolate source was also weakly associated with the lethal versus non-lethal categorization of mouse data, with clinical isolates being significantly more lethal (p = 0.05) than oyster isolates.
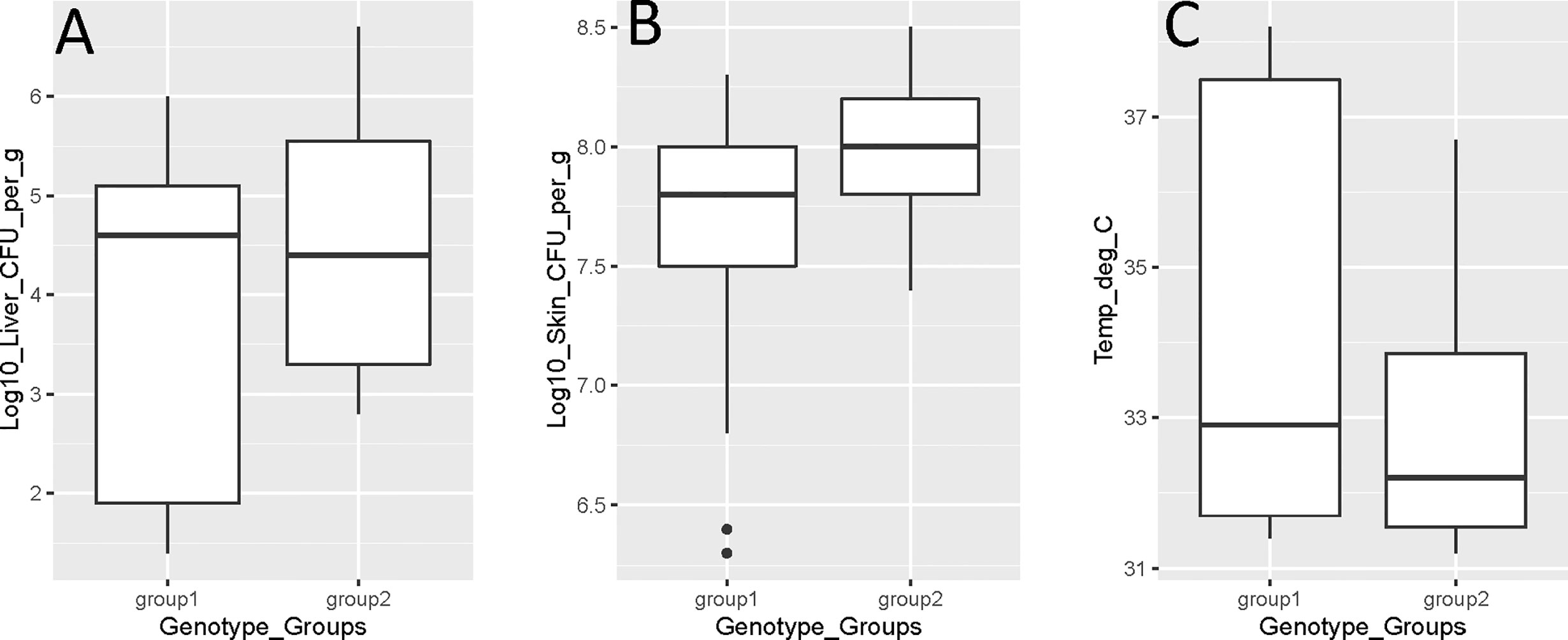
Figure 1 Vibrio vulnificus mouse model virulence parameter data [(A) liver infection; (B) skin infection; (C) mouse temperature] by genotype (group 1 is rrnA/vcgE and rrnAB/vcgE; group 2 is rrnB/vcgC). Each box displays the median with the upper (25%) and lower (75%) quartiles as hinges. Whiskers represent the highest or lowest observation + 1.5*Inter quartile range. Dots represent outliers.
The extrinsic factor with the strongest association to mouse virulence of V. vulnificus strains was season of isolation with significant associations between season of isolation and liver infection (p = 0.04) and mouse temperature (p = 0.04) identified (Figure 2). In both cases, isolates from the cooler season were more virulent, i.e., caused higher liver infection and lower mouse body temperatures. There were no statistically significant relationships (p > 0.05) between season of isolation and skin infection or mouse mortality.
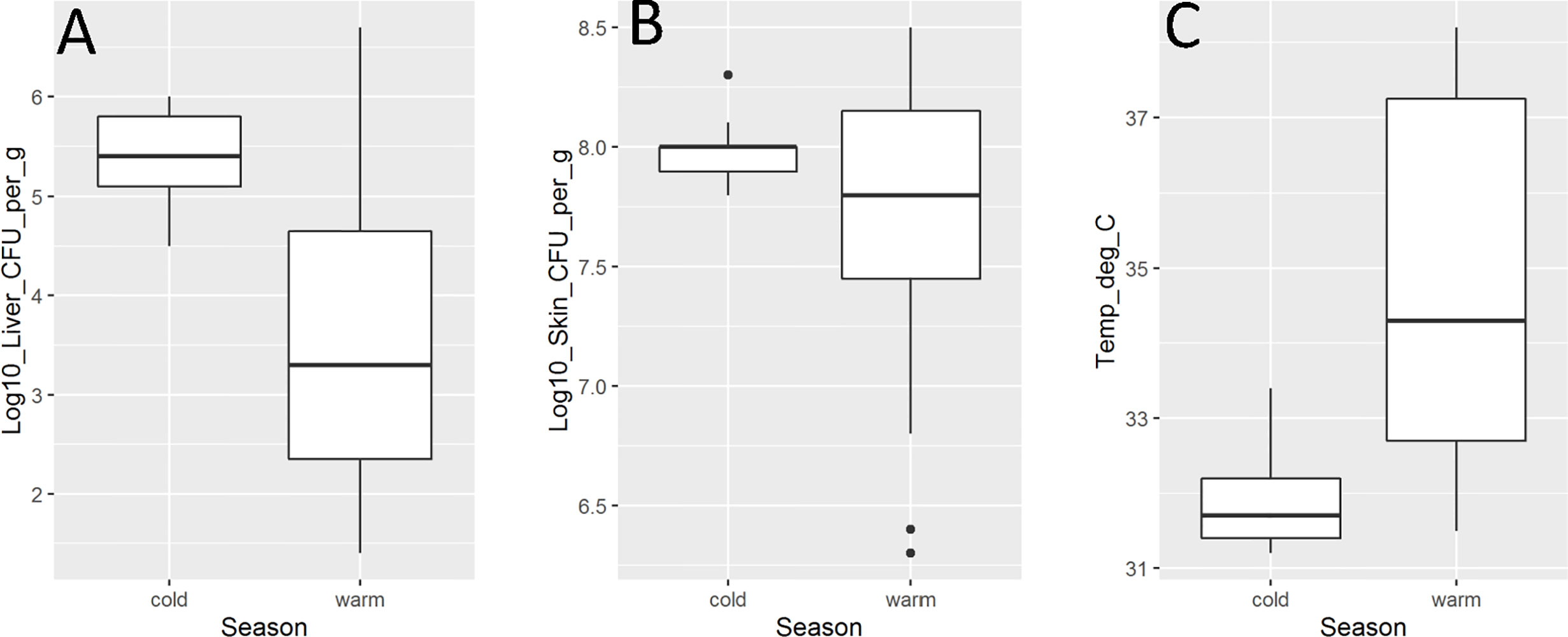
Figure 2 Vibrio vulnificus mouse model virulence parameter data [(A). liver infection; (B). skin infection; (C). mouse temperature] by season (cold is October–April; warm is May–September). Isolate from HI (CDC_K4574) is grouped with warm season due to lack of temperature variability in the state. Each box displays the median with the upper (25%) and lower (75%) quartiles as hinges. Whiskers represent the highest or lowest observation + 1.5*Inter quartile range. Dots represent outliers.
Biochemical profiles were determined by API 20E. Only 88% of the V. vulnificus isolates were correctly identified, which is slightly higher than previous reports (O’Hara et al., 2003). This is likely due to the use of 2% NaCl for inoculation of the biochemical test, rather than the manufacturer’s recommended 0.85%, as previously described as an improved identification method for Vibrio spp. from the environment (Martinez-Urtaza et al., 2005). Interestingly, a higher rate of misidentification was noted for the clinical isolates (20%), as compared to the oyster isolates (5%), which is contrary to previous findings (Martinez-Urtaza et al., 2005). Another noteworthy observation was the significantly (p = 0.02) lower presence of indole production in isolates from the cooler season. A higher variability in biochemical profiles was observed for the oyster isolates as compared to the clinical isolates. This difference in variability has not been noted previously, but is logical assuming environmental (oyster) isolates are under less selective pressure than clinical isolates, which need specific traits to survive in the human host.
In studies establishing and investigating the utility of V. vulnificus genotypes for association with virulence potential (Nilsson et al., 2003; DePaola et al., 2003; Vickery et al., 2007; Thiaville et al., 2011) a similar set of isolates was used. Repeated use of this set of V. vulnificus isolates provided good reference for repeatability and is valuable for assay development. However, the isolate set remains limited in that clinical isolates were from the warm months (May–September) and environmental isolates were from cooler months (October–April). In contrast, this study utilized a more balanced set of isolates collected from a range of seasonal and geographical sources. As a result, well-defined relationships between isolate origin and genotype were not clearly identified.
While the majority (64%) of oyster isolates were rrnA/vcgE, the prevalence of the rrnB/vcgC genotype (33%) was higher than a previous observation where virulent genotypes represented ~6% of isolates from the environment (Rosche et al., 2005). However, recent studies found results similar to the current work, with reports of up to 40% of V. vulnificus isolated from oysters having the virulent genotype (Han et al., 2009; Drake et al., 2010; Jones et al., 2013). As V. vulnificus that causes infection originates from the environment (where exposure occurs), one would expect a mix of genotypes and virulence potential such as observed in the current work.
Genotypes of clinical V. vulnificus isolates in this study were surprising, with a lower than expected prevalence of the rrnB/vcgC genotype. This holds true even when looking at the subset of clinical isolates from blood cultures and deviates from previous findings where isolates of clinical origin are nearly all rrnB/vcgC (Nilsson et al., 2003; Warner and Oliver, 2008; Drake et al., 2010). We hypothesized this discrepancy is due to seasonal and geographic variability, as our study focused on a more diverse panel compared to previous studies. This theory is supported by previous studies that observed differences in distribution of genotypes in the environment based on season and/or region (Lin and Schwarz, 2003; Warner and Oliver, 2008; Jones et al., 2013; Williams et al., 2017).
Mouse virulence testing has previously identified non-lethal (Groups 1–3) and lethal (Groups 4–5) clusters (Thiaville et al., 2011). When using this model, with a greater sensitivity for liver infection, a novel, non-lethal cluster (Group 6) was identified. This Group caused high rates of skin infection, but low liver infection, so was classified as non-lethal. One surprising finding was that no isolates fell into virulence Group 1 (the least virulent group), even with a diversity of oyster isolates tested. Although this is different from the findings of Thiaville et al. (2011), where 16% of isolates were Group 1, the two studies are similar in that the majority of isolates were identified as Group 4 in both. Taken together, these findings suggest that the majority of V. vulnificus isolates can cause high skin and moderate liver infection, regardless of genotype or isolation source.
We found that lethal and non-lethal strains were only weakly correlated to clinical or oyster origin of the isolates, but strongly correlated to mouse mortality. Regardless of statistical associations, the non-lethal isolates were generally from oysters and have the less virulent genotype, as expected. Additionally, all strains that resulted in >40% mouse mortality were isolated from a clinical source, and all but one had the more virulent genotype. These observations, along with the previous data used in establishing the iron-dextran mouse model (Starks et al., 2000; DePaola et al., 2003), support the utility of this model and validity of resultant data. Interestingly, no correlation was identified between the virulence genotype of isolates and the observed mouse virulence parameters, other than skin infection. These genotypes have been reported as an indicator of severe illness potential in humans; however, the lack of correlation with mouse virulence (as a proxy for potential human infection) questions their suitability as predictors. These results are consistent with previous research demonstrating that genotypes are associated with, but do not predict, virulence in a mouse model (Thiaville et al., 2011).
Season of isolation was the factor which most correlated with virulence potential in V. vulnificus. Isolates from the cooler months (except the isolate from Hawaii) were lethal and were associated with higher levels of liver infection and lower body temperature. Isolates from the warmer season had mixed virulence potential; this is reflected by the wide range in liver and skin infection, as well as body temperature. Environmental drivers of Vibrio spp. are often attributed to two main factors, temperature and salinity, the primary differences across seasons, with higher levels generally found during the warmer season (Takemura et al., 2014; Johnson, 2015). Our results, therefore, may appear counter-intuitive, especially combined with knowledge that the majority of V. vulnificus infections occur during warmer months (Oliver, 2015; Centers for Disease Control and Prevention, 2017). However, this apparent discrepancy may be explained by a higher proportion of virulent V. vulnificus in the environment (and oysters) during the cooler season when total populations are lowest, similar to what has been observed for V. parahaemolyticus (Johnson et al., 2010; DePaola et al., 2010). The hypothesis that a greater proportion of the V. vulnificus population is virulent during the cooler season is supported by objective evidence of V. vulnificus infections from oyster consumption in the US (as reported to the FDA). During the cooler months (November–April), 10% of oysters have >3 log V. vulnificus/g compared to 34% of oysters with these high levels of V. vulnificus during the warm months (DePaola et al., 2010). Assuming equally virulent populations, it would be expected that there be three times more cases in the warmer months than the colder months; however, the difference is not always that large (Personal Communication (2020)), suggesting a higher proportion of disease-causing strains in the cooler season. Taken together, these data indicate an association between virulence and season; additionally, this is the first report to define this association using mouse model data.
Due to the lack of reliable markers for virulence potential, current risk evaluation and management strategies are based on total V. vulnificus populations. This study indicates that vcgC and rrnB, two gene variant candidates for differentiating isolate virulence, are not reliable markers of systemic virulence potential in V. vulnificus. It is likely that the different genotypes are reflective of a bifurcation of phylogenetic lineage (Lopez-Perez et al., 2019), rather than functional differences. This, and previous studies, demonstrated that although no reliable indicator, or set of indicators, has yet been identified, not all V. vulnificus isolates have the same virulence potential. In addition, we have identified a relationship between isolates from the cooler season and systemic virulence potential in V. vulnificus.
Using the findings of this work as a basis, future research may be directed towards identifying markers to differentiate virulence potential and to identify the driving factor(s) behind the association between season and virulence. Identification of those factors may allow focus on regulatory pathways such as long-term cold adaptation of V. vulnificus. Next-generation sequencing makes it is possible to discern new potential markers and pathways important to V. vulnificus virulence with isolate virulence known a priori. Previous SOLiD sequencing of four V. vulnificus isolates (Gulig et al., 2010), resulted in an extensive list of potential markers, likely due a lack of robust coverage of isolate genomes and small sample size. By sequencing additional well-characterized isolates on platforms with increased genome coverage, we can more readily narrow down these lists of candidate virulence genes. Identification of reliable virulence markers would allow for risk assessments and risk management approaches to be refined in order to better protect public health.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
The animal study was reviewed and approved by the Institutional Animal Care & Use Committee, PO Box 100142 Gainesville, Florida 32610-0142, Chair, Michael Katovich, University of Florida.
Designed experiment: JJ. Conducted experiments: TK, JJ, CL, and PG. Wrote paper: KL and JJ. Edited paper: TK, CL, and PG. Analysis: JJ, KL, TK, CL, and PG. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank Dr. Cheryl Tarr, CDC for providing the clinical isolates. Huge thanks to Stuart Chirtel and John Bowers for advice on appropriate selection of statistical methods. Thank you to Ms. Lizzie Farrell, Mrs. Whitney Neil, Dr. Carolyn Simmons, and Dr. Stacey Wiggins for their comments and review of this manuscript.
Amaro C., Biosca E. G. (1996). Vibrio vulnificus biotype 2, pathogenic for eels, is also an opportunistic pathogen for humans. Appl. Environ. Microbiol. 62, 1454–1457. doi: 10.1128/AEM.62.4.1454-1457.1996
Bisharat N., Agmon V., Finkelstein R., Raz R., Ben-Dror G., Lerner L., et al. (1999). Clinical, epidemiological, and microbiological features of Vibrio vulnificus biogroup 3 causing outbreaks of wound infection and bacteraemia in Israel. Israel Vibrio Study Group. Lancet 354, 1421–1424. doi: 10.1016/S0140-6736(99)02471-X
Blake P. A., Merson M. H., Weaver R. E., Hollis D. G., Heublein P. C. (1979). Disease caused by a marine Vibrio. Clinical characteristics and epidemiology. N. Engl. J. Med. 300, 1–5. doi: 10.1056/NEJM197901043000101
Centers for Disease Control and Prevention (2017). Vibrio species causing vibriosis. Available at: https://www.cdc.gov/vibrio/index.html (Accessed October 28, 2020).
DePaola A., Nordstrom J. L., Dalsgaard A., Forslund A., Oliver J., Bates T., et al. (2003). Analysis of Vibrio vulnificus from market oysters and septicemia cases for virulence markers. Appl. Environ. Microbiol. 69, 4006–4011. doi: 10.1128/AEM.69.7.4006-4011.2003
DePaola A., Jones J. L., Woods J., Burkhardt W. 3rd, Calci K. R., Krantz J. A., et al. (2010). Bacterial and viral pathogens in live oysters: 2007 United States market survey. Appl. Environ. Microbiol. 76, 2754–2768. doi: 10.1128/AEM.02590-09
Drake S. L., Whitney B., Levine J. F., Depaola A., Jaykus L. A. (2010). Correlation of mannitol fermentation with virulence-associated genotypic characteristics in Vibrio vulnificus isolates from oysters and water samples in the Gulf of Mexico. Foodborne Pathog. Dis. 7, 97–101. doi: 10.1089/fpd.2009.0362
Gulig P. A., De Crecy-Lagard V., Wright A. C., Walts B., Telonis-Scott M., Mcintyre L. M. (2010). SOLiD sequencing of four Vibrio vulnificus genomes enables comparative genomic analysis and identification of candidate clade-specific virulence genes. BMC Genomics 11, 512. doi: 10.1186/1471-2164-11-512
Han F., Pu S., Hou A., Ge B. (2009). Characterization of clinical and environmental types of Vibrio vulnificus isolates from Louisiana oysters. Foodborne Pathog. Dis. 6, 1251–1258. doi: 10.1089/fpd.2009.0343
Haq S. M., Dayal H. H. (2005). Chronic liver disease and consumption of raw oysters: a potentially lethal combination–a review of Vibrio vulnificus septicemia. Am. J. Gastroenterol. 100, 1195–1199. doi: 10.1111/j.1572-0241.2005.40814.x
Hlady W. G., Klontz K. C. (1996). The epidemiology of Vibrio infections in Florida 1981-1993. J. Infect. Dis. 173, 1176–1183. doi: 10.1093/infdis/173.5.1176
Johnson C. N., Flowers A. R., Noriea N.F. 3rd, Zimmerman A. M., Bowers J. C., Depaola A., et al. (2010). Relationships between environmental factors and pathogenic Vibrios in the Northern Gulf of Mexico. Appl. Environ. Microbiol. 76, 7076–7084. doi: 10.1128/AEM.00697-10
Johnson C. N. (2015). Influence of Environmental Factors on Vibrio spp. in Coastal Ecosystems. Microbiol. Spectr. 3, VE-0008-2014. doi: 10.1128/microbiolspec.VE-0008-2014
Jones J. L., Ludeke C. H., Bowers J. C., Depaola A. (2013). Comparison of plating media for recovery of total and virulent genotypes of Vibrio vulnificus in U.S. market oysters. Int. J. Food Microbiol. 167, 322–327. doi: 10.1016/j.ijfoodmicro.2013.09.017
Kaspar C. W., Tamplin M. L. (1993). Effects of temperature and salinity on the survival of Vibrio vulnificus in seawater and shellfish. Appl. Environ. Microbiol. 59, 2425–2429. doi: 10.1128/AEM.59.8.2425-2429.1993
Kaysner C., Depaola A., Jones J. (2004). Vibrio. Bacteriological Analytical Manual. Available at: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-9-vibrio (Accessed October 28, 2020).
Kinsey T. P., Lydon K. A., Bowers J. C., Jones J. L. (2015). Effects of Dry Storage and Resubmersion of Oysters on Total Vibrio vulnificus and Total and Pathogenic (tdh+/trh+) Vibrio parahaemolyticus Levels. J. Food Prot. 78, 1574–1580. doi: 10.4315/0362-028X.JFP-15-017
Lin M., Schwarz J. R. (2003). Seasonal shifts in population structure of Vibrio vulnificus in an estuarine environment as revealed by partial 16S ribosomal DNA sequencing. FEMS Microbiol. Ecol. 45, 23–27. doi: 10.1016/S0168-6496(03)00091-6
Lopez-Perez M., Jayakumar J. M., Haro-Moreno J. M., Zaragoza-Solas A., Reddi G., Rodriguez-Valera F., et al. (2019). Evolutionary Model of Cluster Divergence of the Emergent Marine Pathogen Vibrio vulnificus: From Genotype to Ecotype. mBio 10, e02852–18. doi: 10.1128/mBio.02852-18
Martinez-Urtaza J., Simental L., Velasco D., Depaola A., Ishibashi M., Nakaguchi Y., et al. (2005). Pandemic Vibrio parahaemolyticus O3:K6, Europe. Emerg. Infect. Dis. 11, 1319–1320. doi: 10.3201/eid1108.050322
National Shellfish Sanitation Program (Us) (2019).Guide for the control of molluscan shellfish. In: Interstate Shellfish Sanitation Conference. Available at: https://www.fda.gov/food/federalstate-food-programs/national-shellfish-sanitation-program-nssp (Accessed January 5, 2021).
Nilsson W. B., Paranjype R. N., Depaola A., Strom M. S. (2003). Sequence polymorphism of the 16S rRNA gene of Vibrio vulnificus is a possible indicator of strain virulence. J. Clin. Microbiol. 41, 442–446. doi: 10.1128/JCM.41.1.442-446.2003
Oliver J. D., Kaper J. B. (2001). “Vibrio species,” in Food microbiology: fundamentals and frontiers, 2nd ed. Ed. Doyle M. P. (Washington, D.C: American Society for Microbiology).
Oliver J. D. (2015). The Biology of Vibrio vulnificus. Microbiol. Spectr. 3, VE-0001-2014 doi: 10.1128/microbiolspec.VE-0001-2014
O’Hara C. M., Sowers E. G., Bopp C. A., Duda S. B., Strockbine N. A. (2003). Accuracy of six commercially available systems for identification of members of the family vibrionaceae. J. Clin. Microbiol. 41, 5654–5659. doi: 10.1128/JCM.41.12.5654-5659.2003
Rosche T. M., Yano Y., Oliver J. D. (2005). A rapid and simple PCR analysis indicates there are two subgroups of Vibrio vulnificus which correlate with clinical or environmental isolation. Microbiol. Immunol. 49, 381–389. doi: 10.1111/j.1348-0421.2005.tb03731.x
Sanjuan E., Fouz B., Oliver J. D., Amaro C. (2009). Evaluation of genotypic and phenotypic methods to distinguish clinical from environmental Vibrio vulnificus strains. Appl. Environ. Microbiol. 75, 1604–1613. doi: 10.1128/AEM.01594-08
Scallan E., Hoekstra R. M., Angulo F. J., Tauxe R. V., Widdowson M. A., Roy S. L., et al. (2011). Foodborne illness acquired in the United States–major pathogens. Emerg. Infect. Dis. 17, 7–15. doi: 10.3201/eid1701.P11101
Shapiro R. L., Altekruse S., Hutwagner L., Bishop R., Hammond R., Wilson S., et al. (1998). The role of Gulf Coast oysters harvested in warmer months in Vibrio vulnificus infections in the United States 1988-1996. Vibrio Working Group. J. Infect. Dis. 178, 752–759. doi: 10.1086/515367
Starks A. M., Schoeb T. R., Tamplin M. L., Parveen S., Doyle T. J., Bomeisl P. E., et al. (2000). Pathogenesis of infection by clinical and environmental strains of Vibrio vulnificus in iron-dextran-treated mice. Infect. Immun. 68, 5785–5793. doi: 10.1128/IAI.68.10.5785-5793.2000
Starks A. M., Bourdage K. L., Thiaville P. C., Gulig P. A. (2006). Use of a marker plasmid to examine differential rates of growth and death between clinical and environmental strains of Vibrio vulnificus in experimentally infected mice. Mol. Microbiol. 61, 310–323. doi: 10.1111/j.1365-2958.2006.05227.x
Strom M. S., Paranjpye R. N. (2000). Epidemiology and pathogenesis of Vibrio vulnificus. Microbes Infect. 2, 177–188. doi: 10.1016/S1286-4579(00)00270-7
Takemura A. F., Chien D. M., Polz M. F. (2014). Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front. Microbiol. 5, 38. doi: 10.3389/fmicb.2014.00038
Thiaville P. C., Bourdage K. L., Wright A. C., Farrell-Evans M., Garvan C. W., Gulig P. A. (2011). Genotype is correlated with but does not predict virulence of Vibrio vulnificus biotype 1 in subcutaneously inoculated, iron dextran-treated mice. Infect. Immun. 79, 1194–1207. doi: 10.1128/IAI.01031-10
Veenstra J., Rietra P. J., Goudswaard J., Kaan J. A., Van Keulen P. H., Stoutenbeek C. P. (1993). Extra-intestinal infections caused by Vibrio spp. in The Netherlands. Ned. Tijdschr. Geneeskd. 137, 654–657.PIMD 8469298
Vickery M. C., Nilsson W. B., Strom M. S., Nordstrom J. L., Depaola A. (2007). A real-time PCR assay for the rapid determination of 16S rRNA genotype in Vibrio vulnificus. J. Microbiol. Methods 68, 376–384. doi: 10.1016/j.mimet.2006.02.018
Warner E., Oliver J. D. (2008). Population structures of two genotypes of Vibrio vulnificus in oysters (Crassostrea virginica) and seawater. Appl. Environ. Microbiol. 74, 80–85. doi: 10.1128/AEM.01434-07
Williams T. C., Froelich B. A., Phippen B., Fowler P., Noble R. T., Oliver J. D. (2017). Different abundance and correlational patterns exist between total and presumed pathogenic Vibrio vulnificus and V. parahaemolyticus in shellfish and waters along the North Carolina coast. FEMS Microbiol. Ecol. 93, fix071. doi: 10.1093/femsec/fix071
Keywords: Vibrio, mouse model, mannitol, virulence, 16S rDNA, season
Citation: Lydon KA, Kinsey T, Le C, Gulig PA and Jones JL (2021) Biochemical and Virulence Characterization of Vibrio vulnificus Isolates From Clinical and Environmental Sources. Front. Cell. Infect. Microbiol. 11:637019. doi: 10.3389/fcimb.2021.637019
Received: 02 December 2020; Accepted: 19 January 2021;
Published: 26 February 2021.
Edited by:
Lixing Huang, Jimei University, ChinaCopyright © 2021 Lydon, Kinsey, Le, Gulig and Jones. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jessica L. Jones, amVzc2ljYS5qb25lc0BmZGEuaGhzLmdvdg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.