
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 19 June 2020
Sec. Clinical Microbiology
Volume 10 - 2020 | https://doi.org/10.3389/fcimb.2020.00303
This article is part of the Research Topic Bacterial Vaginosis, a Model of True Polymicrobial Infections: Genetics, Evolution, Clinical and Socio-Clinical Implications View all 14 articles
 David Pacha-Herrera1
David Pacha-Herrera1 Gabriela Vasco1,2
Gabriela Vasco1,2 Cecilia Cruz-Betancourt3
Cecilia Cruz-Betancourt3 Juan Miguel Galarza3
Juan Miguel Galarza3 Verónica Barragán1
Verónica Barragán1 António Machado1*
António Machado1*Pregnancy outcomes and women's health are directly affected by vaginal microbiota. This microbiota consists of a dynamic ecosystem of various microbes in different ratios, which in healthy conditions protect the vaginal epithelium from infections. However, cases of vaginal infection are regularly diagnosed in women of reproductive age, contributing to more severe outcomes. Therefore, our main goal was to determine the prevalence of bacterial vaginosis (BV), aerobic vaginitis (AV), and vulvovaginal candidiasis (VVC) among Ecuadorian pregnant and non-pregnant women. A cross-sectional study was conducted among 217 women between 13 and 40 years old seeking primary healthcare in Carlos Andrade Marin Hospital (HCAM), Gynecological-Obstetric Hospital Isidro Ayora (HGOIA) and Center for Teaching Health Cipriana Dueñas during October 2018 to February 2019. The classical characterization of the vaginal microbiota was performed through microscopy by the Nugent criteria to evaluate the presence of BV, healthy and intermediate microbiota, by the criteria of Donders to determine the presence of AV and by the Marot-Leblond criteria to diagnose VVC. DNA extraction from vaginal samples and Polymerase Chain Reaction (PCR) analysis was performed to characterize the presence of Gardnerella spp., Mobiluncus mulieris, Escherichia coli, Enterococcus spp., and Lactobacillus spp. Finally, quantification of the lactobacilli was performed by quantitative real-time PCR (qPCR) for samples from women with normal vaginal microbiota and women with AV. Our results showed 52% of women with healthy microbiota, 7% with intermediate microbiota, and 41% with vaginal dysbiosis, comprising 27% with AV, 8% with BV and 4% with VVC and 2% with co-infections or co-dysbiosis. Additionally, a higher amount of lactobacilli were found in pregnant women when compared to non-pregnant women, while AV cases were characterized by a significant drop of Lactobacillus spp., more precisely, between 1E3 and 1E5 colony forming units (CFU)/ml. Finally, women with normal vaginal microbiota showed an average load of lactobacilli between 1E6 and 1E7 CFU/ml. This pilot study showed no statistically significant differences between pregnant and non-pregnant women, pointing to the possibility to use lactobacilli quantification for the prevention of future vaginal infections.
The normal vaginal microbiota plays a crucial role for the health of pregnant and non-pregnant women (Vaneechoutte, 2017b), preventing several urogenital diseases (Ling et al., 2013), including bacterial vaginosis (BV) (Dai et al., 2010; Ling et al., 2010, 2013; Gondo et al., 2011; Van De Wijgert et al., 2014), aerobic vaginitis (AV) (Donders et al., 2005, 2011; Fan et al., 2013; Jahic et al., 2013; Tansarli et al., 2013), urinary tract infections (UTI) (Cauci et al., 2002; Zhou et al., 2004; Borges et al., 2014), yeast vaginitis (Ringdahl, 2006; Dai et al., 2010; Xu et al., 2010), and sexually transmitted diseases (such as HIV) (Bolton et al., 2008; Srinivasan and Fredricks, 2008; Petrova et al., 2013; Van De Wijgert et al., 2014). In the context of this study, it is also important to mention that women and teenagers in Ecuador have a wide range of health care needs, in particular, related to sexual and reproductive health (Svanemyr et al., 2017). In Ecuador, a major concern is the high rate of adolescent pregnancy, i.e., pregnancy between ages 10 to 19. Several studies worldwide also demonstrated a higher risk of acquiring HIV, herpes simplex virus type 2 and other sexually transmitted infections in non-pregnant women with vaginal infections or intermediate vaginal microbiota (Li et al., 2012; Petrova et al., 2013; Datcu et al., 2014; Van De Wijgert et al., 2014). Thus, lactic acid-producing bacteria (such as Lactobacillus spp.) metabolize glycogen, increasing lactic acid and a normal acidic vaginal pH of 3.8–4.4 (Farage et al., 2010; Borges et al., 2014; Mendling, 2016; Vaneechoutte, 2017b).
The vaginal microbial community is a variable econiche that fluctuates between normal and dysbiotic microbiota (Vaneechoutte, 2017b), which could be influenced by several intrinsic and extrinsic factors (Mendling, 2016) and eventually leading to an increment of both aerobic and anaerobic microorganisms (Larsen and Monif, 2001; Ling et al., 2010; Fredricks, 2011; Ravel et al., 2011). However, the most predominant genus in a healthy vaginal microbiota is Lactobacillus (Borges et al., 2014; Vaneechoutte, 2017b). Lactobacillus genus is known to inhibit the adhesion and proliferation of opportunistic and primary pathogens (Bolton et al., 2008). The mechanisms by which vaginal lactobacilli provide colonization resistance is generally considered to be through production of several antimicrobial compounds such as hydrogen peroxide, lactic acid and/or bacteriocins (Aroutcheva et al., 2001; Alpay et al., 2003; Vaneechoutte, 2017b; Collins et al., 2018), as well as acting as biosurfactant on the vaginal epithelium (Boris and Barbés, 2000; Borges et al., 2014).
Although several species of Lactobacillus were already identified in vaginal microbiota, the most predominant species found in normal vaginal microbiota are L. crispatus, L. gasseri, L. jensenii, and L. iners (Farage et al., 2010; Borges et al., 2014; Oliveira et al., 2018). Also, other species could be detected in low amount among healthy vaginal microbiota such as Atopobium, Enterobacter, Escherichia, Gardnerella, Mobiluncus, Prevotella, Staphylococcus, Shigella (Hernández-Rodríguez et al., 2011; Gajer et al., 2012; Romero et al., 2014; Oliveira et al., 2018). These species can also behave as opportunistic pathogens (Gajer et al., 2012; Vaneechoutte, 2017a). Several factors can induce disruptions of the healthy microbiota equilibrium, establishing a microbial dysbiosis and, thus, future vaginal infections (Gajer et al., 2012; Johnson and Versalovic, 2012; Petrova et al., 2013; Vaneechoutte, 2017b).
According to previous studies, bacterial vaginosis (BV) is the most common vaginal dysbiosis among women of reproductive age (Cristiano et al., 1996; Nelson et al., 2009; Dai et al., 2010; Gondo et al., 2011), being characterized by lactobacilli replacement by anaerobes (Donders et al., 2011). Gardnerella spp., Atopobium vaginae, Bacteroides spp. and Mobiluncus spp. are the main pathogenic anaerobes associated with BV (Mendling, 2016), which is usually diagnosed by Nugent criteria (Nugent et al., 1991) or the Amsel criteria (Van De Wijgert et al., 2014). Besides BV, a condition designated aerobic vaginitis (AV) has also been recognized, characterized by the presence of aerobic bacteria in detriment of lactobacilli and by inflammation diagnosed a yellow-green discharge (Donders, 2007; Mendling, 2016). This vaginal infection is usually dominated by Streptococcus sp., Enterococcus sp., and/or Gram-negative bacteria of enteric origin (mainly, Escherichia coli). Finally, vulvovaginal candidiasis (VVC) is the most prevalent cause of vaginal infection by fungi, with at least 75% of healthy women suffering one episode of VVC during lifetime (Ringdahl, 2006) and whereby Candida albicans is the most important species (Marot-Leblond et al., 2009).
Our main goal of the present study was to evaluate the presence of vaginal infection among Ecuadorian women by classical and standard microbiological techniques or criteria (Fredricks, 2011) and to determine the dominance of different types of vaginal infection among pregnant and non-pregnant women. Also, the present study aimed to detect the presence of specific opportunistic pathogens (E. coli, Enterococcus faecalis, Gardnerella spp., and Mobiluncus mulieris) by Polymerase Chain Reaction (PCR) and quantified the number of lactobacilli through quantitative real-time PCR (qPCR). The analysis of the normal amount of lactobacilli in pregnant and non-pregnant women might enable to determine the lactobacilli threshold associated with the establishment of vaginal infection.
The study was conducted in the Microbiology Institute at USFQ in collaboration with Hospital Carlos Andrade Marín (HCAM) and Universidad Central del Ecuador (UCE) from October 2018 to February 2019. The research team recruited 217 Ecuadorian female volunteers of Hispanic ethnicity but in reproductive age (13 and 40 years old), of which 111 were pregnant. Applicants were excluded from the study if they reported antimicrobial treatment in the last 3 months or any evidence of bleeding, and also if they had sexual intercourse within the previous 48 h. Also, a questionnaire was taken regarding demographic characteristics, sexual and health behavior of each patient, and each enrolled woman provided a usable vulvovaginal swab sample.
This study was approved by the Ethics Committee of Universidad San Francisco de Quito (USFQ) and the Ministry of Health of Ecuador (Protocol code: 2016-140M by MSP-SDM-10-2013-2019-O review board). The female participants were recruited to our study set, after having read and signed the informed consent or, in the case of underaged participants, from their parents or legal representatives.
Samples were taken by a gynecologist using a sterile disposable vaginal speculum. The lateral vaginal walls were swabbed with a sterile swab to collect the cervical fluid, to prepare a smear on a microscope slide. Briefly, each vaginal smear was obtained by rolling the previous swab onto a glass slide, then heat-fixed and Gram-stained by using safranin as the counterstain. Following the Gram smear procedure, the swab was placed in 1 ml of phosphate buffer saline (PBS) and vortexed vigorously for ~3 min. The remaining vaginal material was collected by centrifugation at 16,000 g for 5 min. The obtained pellet was suspended into an aliquot of 1 ml of saline (0.9% NaCl) which was used for culture of Candida spp. in different media (see section Culture of Candida spp.) and for wet mount microscopy for a better diagnosis of AV and VVC (see section Microbiological Classification of Vaginal Infections).
A second sample was taken by a cervical brush (Rovers Cervex Brush®) through endo and exo-cervical brushing, placed immediately in Cobas® Preservative Fluid, stored at 4°C until processing in the clinical laboratory of HCAM, and used for DNA extraction (see section DNA Extraction of Vaginal Swabs). Each sample was further used to culture of Candida spp.
Candida spp. was cultured on different media from the saline aliquot (see section Sample Collection). Briefly, 100 μl of saline solution was plated onto Petri dishes containing 5% human blood agar (HBA), chocolate agar (heated human blood agar) or Sabouraud dextrose agar (SDA). The plates were incubated at 37°C for 48 h, under aerobic conditions, and colonies were analyzed and identified by gram staining, biochemical properties (catalase, oxidase, and hemolysis) and PCR (data not shown).
The Gram-stained vaginal smears were classified according to Nugent criteria for bacterial vaginosis (BV) (Nugent et al., 1991), the criteria of Donders et al. for aerobic vaginitis (AV) (Donders, 1999) and those of Marot-Leblond et al. for vulvovaginal candidiasis (VVC) (Marot-Leblond et al., 2009). The evaluation of several cell types present in each smear was performed for 10 to 15 microscopic fields under oil immersion at 1000 X magnification (Donders, 2007).
After an initial evaluation of the Gram-stained smears by the Nugent criteria, all samples were evaluated by means of phase-contrast microscopy (X400 magnification) of wet smears, according to Schröders classification (Donders et al., 2005) and the Marot-Leblond et al. (2009) criteria (see Table 1). The absence of Lactobacillus spp., presence of cocci or coarse bacilli in high numbers, presence of parabasal epithelial cells representing >10% of the epithelial cells, and/or presence of leucocytes were considered as indicative for AV (Donders et al., 2005). In addition, aggravated AV diagnosis was defined as the most extreme form of aerobic vaginitis under Donders evaluation from Schröders classification (Donders et al., 2005), where AV samples showed lactobacilli severely depressed or absent because of overgrowth of other bacteria (Cocci or chains), more than 10 leukocytes per epithelial cell present in the samples and more than 50% of the leukocytes had a toxic appearance. It is important to mention that leukocytes were also evaluated on their granular appearance due to abundant lysozyme activity (“toxic leukocytes”) (Donders et al., 2005). Finally, VVC was assessed accordingly to Marot-Leblond and colleagues through at least one of the following criteria: positive Gram-stain preparation with budding yeasts in high numbers (five or more) in more than two microscopic fields, pseudohyphae, and/or hyphal forms in wet smears observation; and positive culture in Chocolate agar, HBA and/or SDA, along with negative microscopic examination results associated with eventual symptoms (thick, white vaginal discharge with no odor, vulvar and vaginal pruritus, burning, or dyspareunia) or clinical history (previous infection) obtained from the medical survey with the professional gynecologist. Absence of Candida cells in more than two microscopic fields and/or a low number of Candida spp. result on wet smears observation and culture growth was considered as normal Candida colonization rather than VVC (Marot-Leblond et al., 2009).
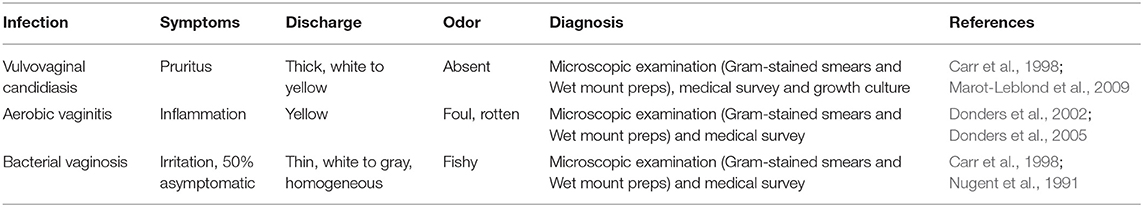
Table 1. Parameters used for the diagnosis of vaginal infections.
The Cobas® 4800 system (Roche Molecular Systems Inc., Pleasanton, CA) was used to extract the DNA of vaginal brushes, according to the manufacturer's instructions. DNA was quantified with a Nanovue spectrophotometer (GE Healthcare Life Science). DNA was eluted at 20 ng/μl with molecular grade water and stored at −20°C until the Polymerase Chain Reaction (PCR) analysis was performed. The quality of DNA was evaluated by measuring the concentration of phenolic compounds or the presence of salts (260/230) and protein contaminants (260/280).
From 217 vaginal samples previously diagnosed by classical criteria through microscopy analysis, 97 were selected for molecular characterization by PCR in a Bio-Rad Thermocycler (Bio-Rad, Hercules, CA). Samples with scores between 0 and 1 of Nugent criteria were selected as healthy microbiota, while samples with scores between 9 and 10 of Nugent criteria (BV) and diagnosed as representing aggravated AV (see section Microbiological Classification of Vaginal Infections) were used as dysbiotic microbiota. Thus, sixty samples with healthy microbiota (38 pregnant and 22 non-pregnant women), 23 samples with AV (14 pregnant and 9 non-pregnant women), and 14 samples with BV microbiota (6 pregnant and 8 non-pregnant women) were included. All samples were analyzed with a total of five primer pairs, targeting two anaerobes (Gardnerella species, and M. mulieris), two aerobes (E. coli and E. faecalis) and for the genus Lactobacillus. Single-template PCR assays were performed for each primer set. The sequence, amplicon size, target gene, and temperature of annealing for each primer pair are described in Table 2.
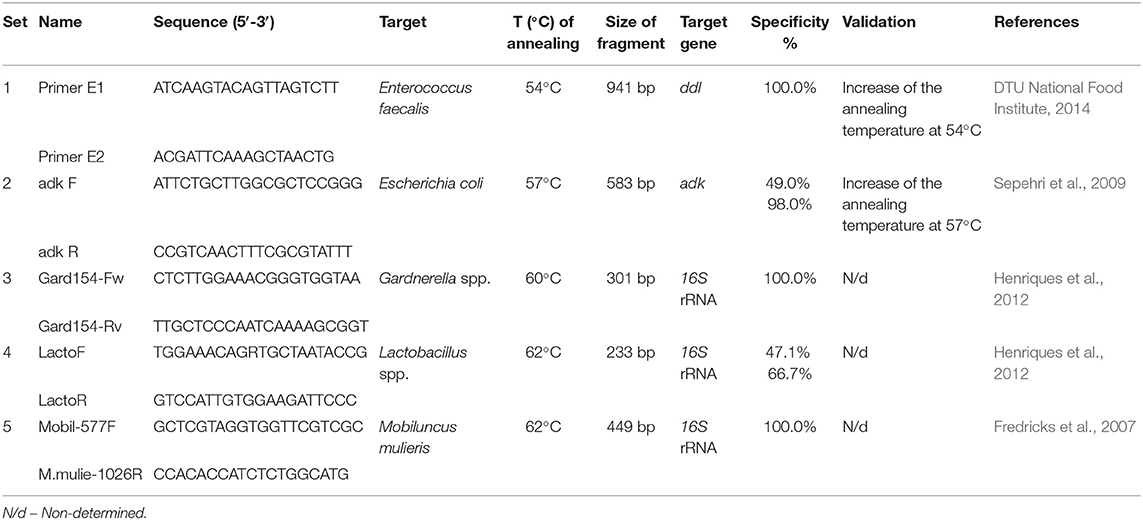
Table 2. PCR primers used in this study.
For PCR, a final volume of 20 μl was used according to the reference protocols (Galán et al., 2006; Fredricks et al., 2007; Sepehri et al., 2009; Henriques et al., 2012; DTU National Food Institute, 2014); which included 0.5 U of Go Taq® DNA Polymerase (Promega, Madison, WI), 1X of Green GoTaq® Flexi Buffer (Promega), 0.25 mM of MgCL2 (Promega), 200 μM of dNTP mix (Promega), 0.5 μM of each primer and target template DNA concentration of ~4 ng/μL, and the remaining volume with molecular grade H2O. The PCR thermal cycling consisted of initial denaturation at 94°C for 2 min; followed by 29 cycles of denaturation at 94°C for 30 s, annealing at each primer pair temperature (Table 2) for 30 s and extension at 72°C for 1 min, and final extension of 5 min at 72°C. The respective use of negative (without DNA sample and samples with other related bacteria) and positive (collection of identified strains of each species through DNA sequencing) controls were used in each PCR assay. These positive controls were provided by the Microbiology Institute at USFQ. All samples were randomly performed in duplicate or triplicate with different negative and positive controls.
After PCR amplification, a volume of 4 μL from each PCR product was visualized in 1.5% (w/w) agarose (Promega) gel electrophoresis using 0.1% ethidium bromide staining. The DNA analysis was performed under permit No. MAE-DNB-CM-2016-0046.
To create positive controls and standard quantification solutions with a well-known Lactobacillus sp. concentration (CFU/mL), a sample of known concentration (also known as a calibrator) (e.g., number of CFU per mL) was obtained through a validated calibration curve (CFU/OD) (Begot et al., 1996). This calibrator was serially diluted tenfold and used to construct a standard curve for qPCR assays. Accordingly, Lactobacillus gasseri strain JCM1131 was cultured during 24 h in Mann Rogosa Sharp Agar at 37°C under microaerophilic conditions (Begot et al., 1996; Mytilinaios et al., 2012; Machado et al., 2013). The calibrator concentration was previously proved by media growth culture counting as previously described (Naghili et al., 2013). The DNA extraction was performed from the highest CFU/ml concentration, and serial dilutions from 1E9 to 1E0 CFU/mL were used as qPCR standards. The DNA extraction of this solution with the highest concentration was performed under the same procedure already described in section DNA Extraction of Vaginal Swabs. In each qPCR assay, two random controls were also used as blind samples in triplicate.
Each reaction was performed with GoTaq® Master Mix qPCR (Promega, Madison, WI, USA) in a final volume of 20 μl, 0.5 μM of each primer (LactoF-TGGAAACAGRTGCTAATACCG and LactoR-GTCCATTGTGGAAGATTCCC) and 2 μl of DNA template. Each qPCR assay was performed in a quantitative real-time PCR Thermocycler (Bio-Rad CA, USA) under the following conditions: initial denaturation at 94°C for 2 min followed by 40 cycles of denaturation at 94°C for 30 s, annealing at 62°C for 30 s, extension at 72°C for 1 min, and a final extension for 5 min at 72°C. Each qPCR assay was followed by this melt curve analysis, allowing amplicon validation and identification of false positives through its profile and the specific temperature of melting (Tm). Each sample was analyzed by triplicate, and qPCR assays were realized in different days. Negative target controls and no template controls were included in all plates.
Primers used for these qPCR assays were previously described to amplify Lactobacillus spp. through classical PCR, but not for quantitative real-time PCR. Therefore assay metrics were determined by testing their performance across the limit of quantification (LoQ) and limit of detection (LoD), as well as linearity as previously described (Price et al., 2012). The optimized assay exhibited the LoQ and LoD to be 1E2 CFU per ml, while the range of linearity of the assay was from 1E9 to 1E3 CFU/mL. The load of lactobacilli in each sample was determined by running six or five standard dilutions (1E9-1E3 CFU/mL), both in duplicate or triplicate on each qPCR assay.
For the quantification of Lactobacillus spp., 83 samples were selected for qPCR analysis from the initial subset of 97 vaginal samples previously characterized by PCR assays to molecular characterization of the main bacteria (see section Identification of the Major Bacterial Species by PCR), i.e., 60 from healthy microbiota samples and 23 from AV samples.
Statistically significant differences in Lactobacillus spp. quantity among women with healthy and dysbiotic microbiota were evaluated using Kruskal Wallis one-way ANOVA and Mann Whitney tests. In addition, the same statistical analysis was carried out among pregnant and non-pregnant women. Finally, multivariable analysis was performed for sociodemographic and behavioral factors by using Minitab 2017 (Version 17, Minitab, State College, PA).
The sociodemographic characteristics for 217 women were included in the statistical analysis and presented in Table 3. Half of the women in the study were pregnant (51.2%) and approximately half (47.4%) were non-pregnant. They were between 21 and 30 years of age. Only 11 women (5.1%) identified themselves as White, Afro-Ecuadorian, or Indigenous women. So, the majority of the women in our study set (94.9%) were categorized as “Half-blood,” being of Hispanic ethnicity mixed with another background ethnicity (Caucasian, African, or Indigenous women). When performing an overall statistical analysis of age, the results do not show a significant relationship between age and the probability of having a specific diagnosis. Hence, there is no statistical evidence to determine that a woman's age is directly related to a specific vaginal disruption or having a healthy microbiota. From all sociodemographic factors analyzed, only the occupation category had a statistical significance over the diagnostic classification of vaginal infection with a P-value of 0.003 through the Chi-square test (see Table 3). Similarly, the use of contraceptive methods, having different sexual partners, vaginal douches, or cigarette smoking did not show any relation to the development of any vaginal infection type during the study (see Table 4).
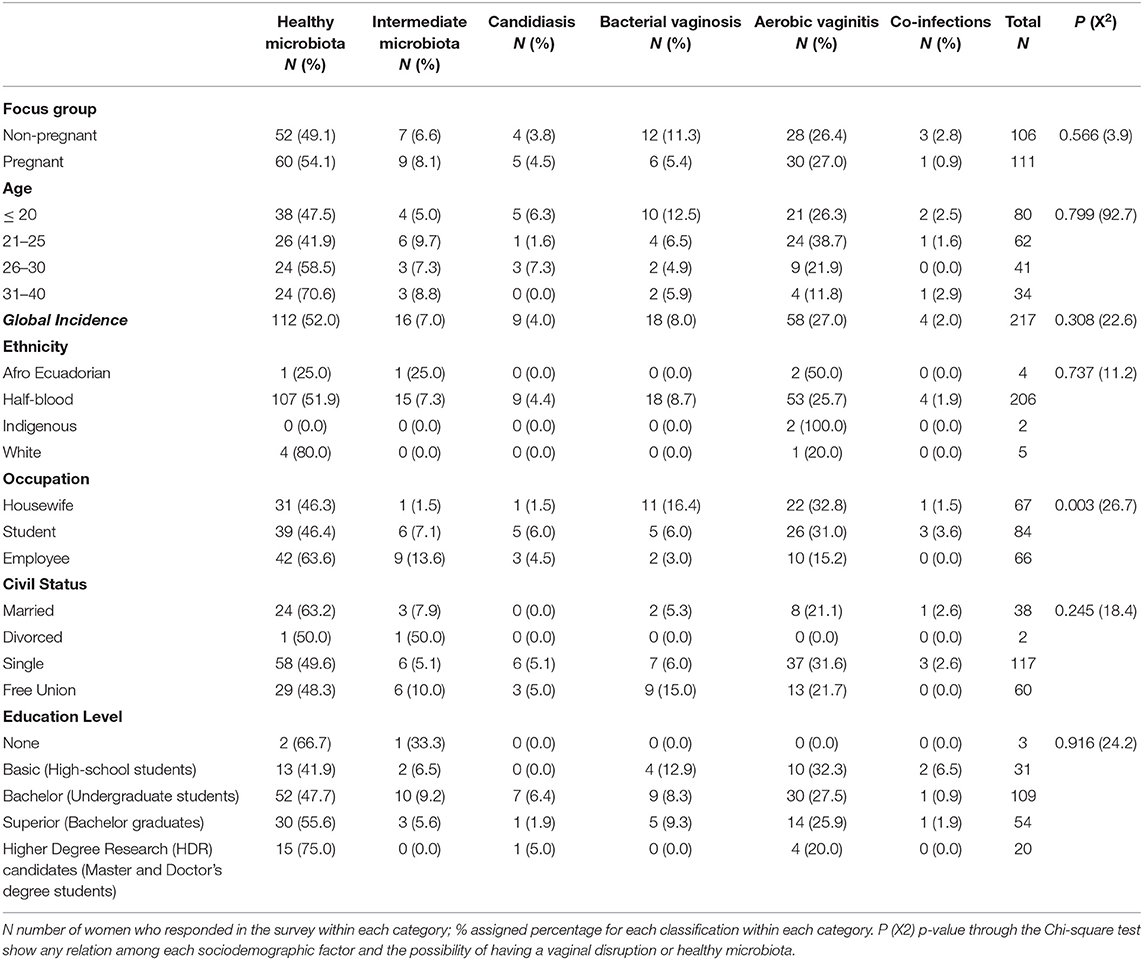
Table 3. Sociodemographic among women in this study with healthy vaginal microbiota, intermediate vaginal microbiota, and vaginal infections (bacterial vaginosis, aerobic vaginitis, candidiasis, and co-infections).
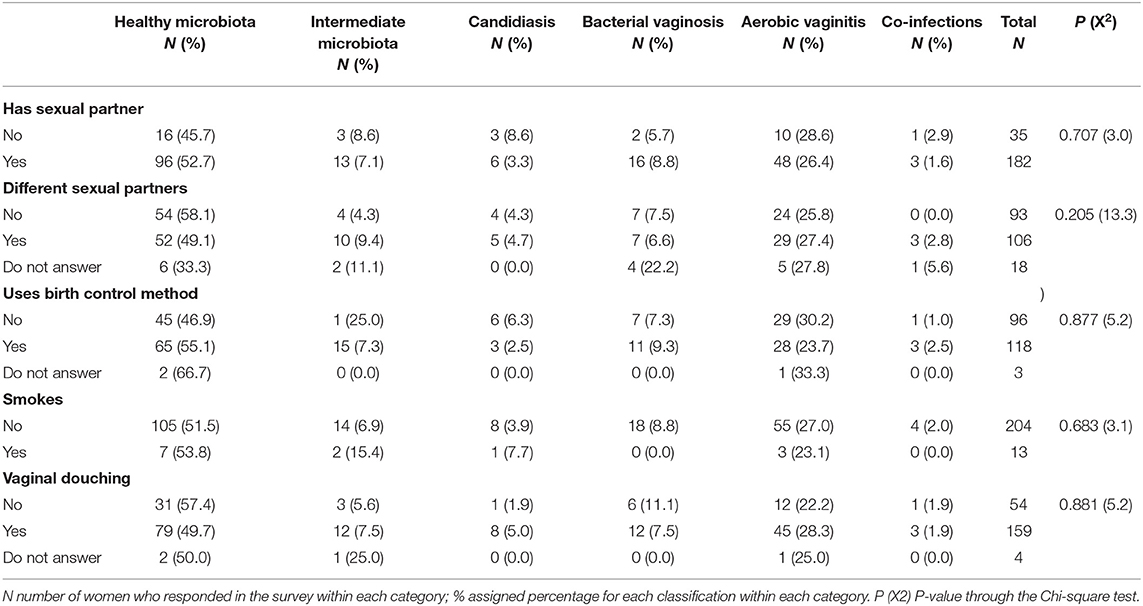
Table 4. Behavioral variables among women in this study with healthy vaginal microbiota, intermediate vaginal microbiota, and vaginal infections (bacterial vaginosis, aerobic vaginitis, candidiasis, and co-infections).
The vaginal samples were evaluated in the Microbiology Institute of USFQ, according to microbiological criteria of Nugent et al. (1991) to identify healthy microbiota, BV, and intermediate microbiota (Nugent et al., 1991); Schröders classification under criteria of Donders et al. (2005) to characterize AV (Donders et al., 2005), and the criteria of Marot-Leblond et al. (2009) to determine VVC (Marot-Leblond et al., 2009). As shown in Table 3, 112 (52.0%) vaginal samples were classified as healthy microbiota, 16 (7.0%) were identified as intermediate microbiota, and 89 (41.0%) were diagnosed as dysbiotic (41.0%), which includes single cases of BV, AV, and VVC but also co-infections. The presence of a unique type of vaginal infection was identified in 85 vaginal samples (39.2%), whereby AV was the most prevalent infection with 26.7% of the vaginal samples, followed by BV (8.3%) and 4.1% with VVC. Furthermore, four vaginal samples were diagnosed with co-infections (1.8%), more precisely two of them with AV and BV, one with AV and VVC, and one with BV and VVC. None of the co-infection samples was further evaluated during qPCR analysis.
Each focus group (pregnant and non-pregnant women) was analyzed to identify any relation between vaginal infection and pregnancy (see Table 5). Although pregnant and non-pregnant women have similar prevalence values in the healthy microbiota, most cases of BV and co-infection were found in non-pregnant women with 67% (12/18) and 75% (3/4) of the cases, respectively, as shown in Table 5. However, these differences were not significant.
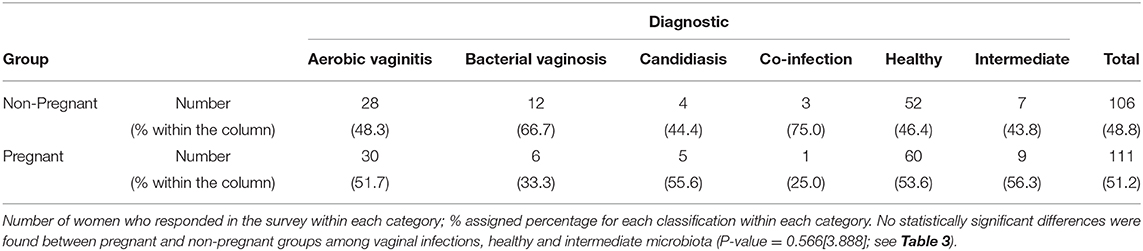
Table 5. Contingency table of vaginal samples between Focus Group and the diagnosis of vaginal infections, healthy and intermediate vaginal microbiota.
The presence of G. vaginalis and M. mulieris (as BV biomarkers), E. coli and E. faecalis (as AV biomarkers), and Lactobacillus spp. (as healthy biomarker) were analyzed by PCR assays from the selected 97 samples (see section Identification of the Major Bacterial Species by PCR). As previously mentioned, almost a half of population set was chosen by classical criteria through microscopy analysis (data not shown), more exactly, healthy microbiota samples with 0–1 and BV samples with 9–10 according to Nugent criteria (Nugent et al., 1991), and the most aggravated AV samples (Donders, 1999).
The results still evidenced the presence of Lactobacillus spp. in both types of vaginal dysbiosis, although their presence decreased to 21% in BV (P = 0.006) and 13% in AV (P = 0.019) when compared to healthy microbiota samples (see Figure S1). Regarding the presence of Gardnerella species, it was present less frequently in healthy microbiota (37%) while in BV and AV prevalence was 71% (P = 0.001) and 78% (P = 0.033), respectively. On the other hand, M. mulieris and E. coli were found in BV at 79% and 36%, respectively; while being detected in low frequency in healthy microbiota and AV cases, as shown in Figure S1. The presence of M. mulieris was low in frequency on AV and normal microbiota, when compared to BV cases. However, presence of M. mulieris was statistically different among healthy microbiota against BV (P < 0.001) and AV cases (P = 0.002), being less recurrent in healthy samples. Finally, E. coli did not show statistical differences among healthy microbiota and BV cases (P = 0.062). Also, it is important to mention that E. faecalis was found to be absent in the population set of the present study.
Among pregnant and non-pregnant women with healthy microbiota, we found that prevalence of Lactobacillus spp. was similar, as shown in Figure 1. On the other hand, pregnant women evidenced higher presence of Gardnerella species (39%), and M. mulieris (16%) when compared to non-pregnant women (32% Gardnerella spp., and 9% of M. mulieris).
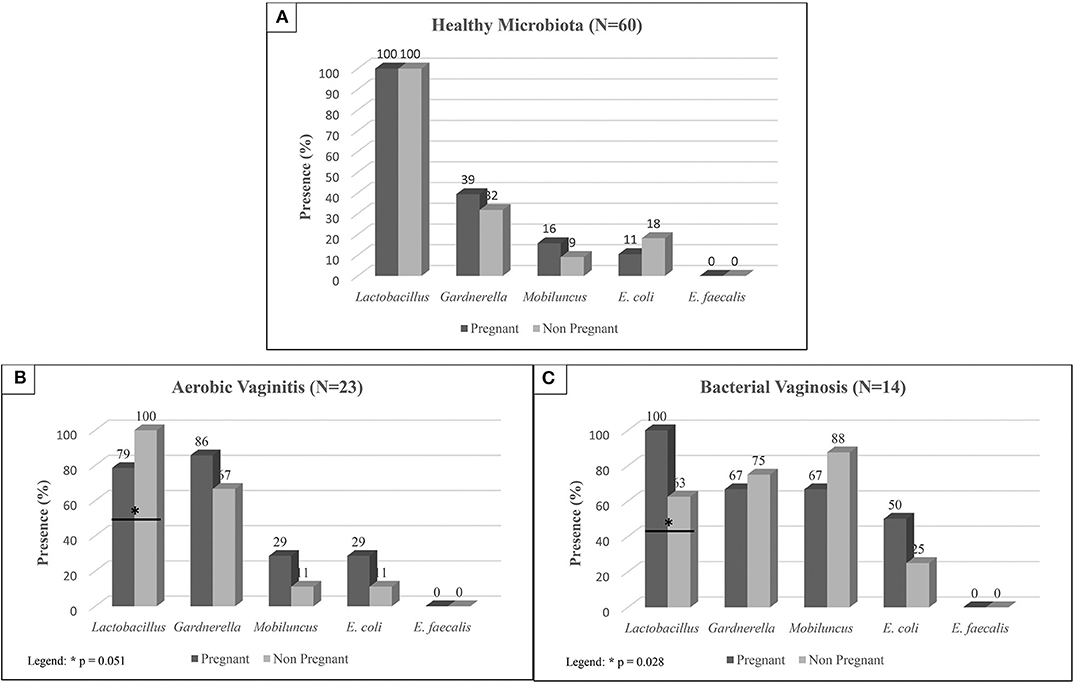
Figure 1. Prevalence of each bacterium in pregnant and non-pregnant women diagnosed as: (A) Healthy Microbiota, (B) Aerobic Vaginitis, and (C) Bacterial Vaginosis according to the microbiological diagnosis. Statistically significant differences were evaluated by Chi-square tests.
In the presence of vaginal dysbiosis and infection, no statistically significant differences were found around opportunistic species between pregnant and non-pregnant women, as shown in Figure 1. However, Lactobacillus spp. showed statistically significant differences between pregnant and non-pregnant women in both BV and AV cases, as shown in Figure 1. In the case of BV, a higher prevalence of Lactobacillus spp. is shown in pregnant women (100%) compared to non-pregnant women (63%; P = 0.028). No significant statistically differences were found on the frequency of Gardnerella spp. and M. mulieris in these samples. In opposition, a drop of Lactobacillus spp. prevalence is shown in pregnant women with AV (79%; P = 0.051) when compared to non-pregnant women (100%).
Due to the small number of samples with BV, we restricted comparison of the lactobacilli quantification to healthy (60) vs. AV (23) cases.
Due to the low number of data, a non-parametrical statistical analysis was performed by means of a Mann-Whitney. Significant differences were shown between healthy and AV groups (P < 0.001; see Figure 2), whereby Lactobacillus spp. varied between 1E6 and 1E7 CFU/ml in healthy microbiota decreased to between 1E3 and 1E5 CFU/ml in AV cases. This was confirmed by Kruskal–Wallis one-way ANOVA testing (P < 0.001; see Figure 2).
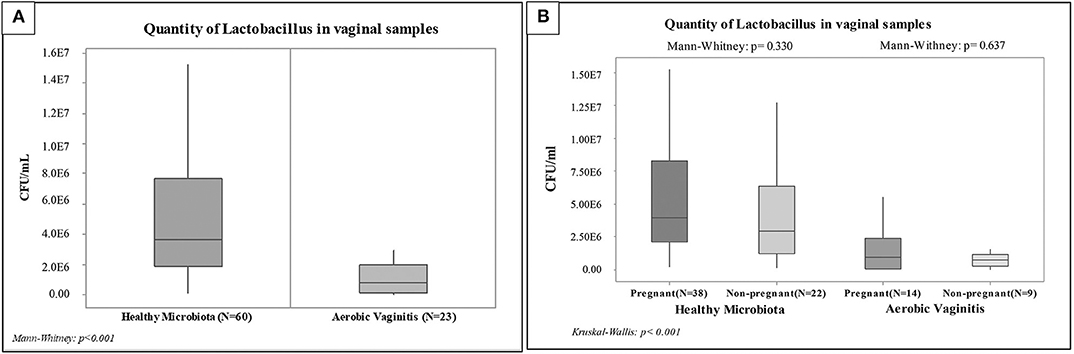
Figure 2. Box plot of the quantification by qPCR of Lactobacillus spp. among vaginal samples: (A) Non-parametric. Statistical analysis among the overall groups (Healthy Microbiota and Aerobic Vaginitis), (B) Non-parametric. Statistical analysis among pregnant and non-pregnant women of each overall group.
Mann-Whitney testing indicated no statistically significant differences between pregnant and non-pregnant women with healthy vaginal microbiota and with AV (P = 0.330 and P = 0.637), as shown in Figure 2. However, the analysis showed statistically significant differences (P < 0.001) when comparing pregnant women with healthy microbiota against AV. Likewise, we found slight differences (P = 0.006) when comparing non-pregnant women with healthy microbiota against AV. Finally, it is worth noting that the same significance levels were also observed between healthy pregnant women against AV non-pregnant women (P < 0.001) and between healthy non-pregnant women against AV pregnant women (P = 0.0041). These preliminary results showed similar ranges of lactobacilli load in pregnant and non-pregnant women from each group set (AV and healthy vaginal microbiota).
This study evaluated a possible relationship between vaginal infection, vaginal dysbiosis and sociodemographic or behavioral variables among pregnant and non-pregnant women. A disruption of the vaginal microbiota usually occurs when any cause promotes a diminution in lactobacilli levels, leading to other microorganisms' augmentation (primary or opportunistic pathogens). These causes of imbalance can be due to several intrinsic and extrinsic factors (Bolton et al., 2008; Borges et al., 2014). As intrinsic factors, the vaginal microbiota of women is driven mainly by hormonal changes during their reproductive life (Farage et al., 2010). These intrinsic factors were distinctively different in the two focus groups (pregnant and non-pregnant women). However, no statistically significant differences regarding composition of vaginal microbiota were detectable (see Table 3). Likewise, in this study, there was no statistically significant relationship among any extrinsic factor (behavioral variables) obtained in the questionnaire by multivariate analysis (see Table 4). This differs from other studies that established statistically significant association with some of the extrinsic factors analyzed by this study, such as the number of sexual partners (Schwebke et al., 1999), and ethnicity (Zhou et al., 2004). Others could not establish associations with the use of contraception, lubricant or spermicide, as well as personal hygiene habits (Keane et al., 1997). Similarly, others did not find any effect of oral contraceptives on the vaginal microbiota of 36 women (Eschenbach et al., 2000). As such, several studies reported contradicting results regarding sociodemographic and behavioral variables, making conclusive comparisons difficult to achieve.
In our study set, 52% of women were characterized by a healthy vaginal microbiota, 7% were diagnosed with intermediate vaginal microbiota and 41% with some vaginal infection or vaginal dysbiosis (BV). Similar results were reported in the United Kingdom (Keane et al., 1997), identifying 48% of female participants with healthy vaginal microbiota and 19% with an abnormal microbiota. Similarly, Gondo et al. (2011) reported that 47.5% of the women showed infection in a study enrolling 245 Brazilian women (Gondo et al., 2011). Bacterial vaginosis (BV) is usually reported as the most prevalent vaginal infection around the world (Nelson et al., 2009; Ling et al., 2013; Machado et al., 2013), followed by vulvovaginal candidiasis (VVC) (Ringdahl, 2006). Another condition, aerobic vaginitis (AV), has been recently characterized by Donders and colleagues in 1999, and has been shown to play an important role for vaginal health (Donders, 1999; Datcu et al., 2014; Donders et al., 2017). Furthermore, Donders (2007) showed that this type of vaginal infection could easily be confused with an intermediate microbiota or even bacterial vaginosis (Donders, 2007), which may be a major reason why reliable data on the prevalence of AV in the general population are not very abundant (Donders et al., 2017).
AV can also be associated with the increased risk of preterm pre-labor rupture of membranes, chorioamnionitis, and preterm delivery (Donders et al., 2017). Contrary to previous studies (Schwebke et al., 1996; Cauci et al., 2002; Donders et al., 2005; Vieira-Baptista et al., 2017), in the present study AV was the most prevalent vaginal infection with a similar percentage of AV among pregnant (51.7%) and non-pregnant (48.3%) women. Again, the latter is in contradiction with other studies that reported low AV prevalence among pregnant women. Although Donders et al. (2009) postulated that AV was not common in pregnancy, a more recently publication by Donders et al. (2017) reported that AV could easily be confused with an intermediate microbiota and bacterial vaginosis and so reliable data on the prevalence of AV could be available in few amounts. In 2013, Jahic and colleagues diagnosed AV in 51% of the enrolled female participants, where E. coli and E. faecalis were the most prevalent bacteria (Jahic et al., 2013). In agreement, Fan et al. (2013) reported the same main bacteria and S. epidermidis in their AV cases (Fan et al., 2013).
In non-pregnant women, several studies reported a prevalence of AV between 5 and 10.5% in symptomatic women (Bologno et al., 2011; Marconi et al., 2012; Donders et al., 2017), whereby the most frequently identified bacteria were E. coli (4–23%) (Tansarli et al., 2013), Staphylococcus (around 27%), Streptococcus (0.7–58.7%) and Enterococcus spp. (0.3–2.4%) (Von Gruenigen et al., 2000; Iavazzo et al., 2008; Tansarli et al., 2013). These previous studies could partially explain the absence of E. faecalis in our study due to the low rate of detection. Finally, E. coli prevalence in pregnant (28.57%) and non-pregnant (11.11%), as established in our study, were within the range described by Tansarli et al. (2013) and in agreement with postulations made by Donders (2007).
The vaginal microbiota complexity in healthy and dysbiosis samples had already been described by several authors in women with AV and BV (Tempera and Furneri, 2010; Zozaya-Hinchliffe et al., 2010; Rumyantseva et al., 2016). Similar to Zozaya-Hinchliffe et al. (2010), we believed that the PCR characterization of the major bacterial species by PCR and the development of qPCR assays would be facilitated by first working with specimens whose microbiota would be most likely to differ significantly (Zozaya-Hinchliffe et al., 2010). So, we only selected vaginal samples from sixty women with normal vaginal microbiota who had Nugent scores of 0 and 1, twenty tree women with aggravated AV diagnosis (see section Microbiological Classification of Vaginal Infections), and fourteen women with BV who had Nugent scores of 9 and 10. These 97 women were selected to identify the major bacterial species by PCR, and then healthy and AV women (83 samples) were evaluated by qPCR (see section Amount of Lactobacillus spp. Among Healthy Women and Women With Vaginal Infections). However, this selection of samples could be considered a limitation of the present study.
The presence of Gardnerella species in a low number in the vaginal microbiota is not an indicator of BV (De Backer et al., 2007; de Vos et al., 2012; Mendling, 2016), being considered as part of the healthy vaginal microbiota. Meanwhile, several studies have shown that there are almost four different groups of Gardnerella species (A, B, C, and D), previously all considered as Gardnerella vaginalis (Vaneechoutte et al., 2019), which not all of them are related to the development of BV (Santiago et al., 2011; Hardy et al., 2017; Hill et al., 2019). In 2019, Vaneechoutte and colleagues amended several species of G. vaginalis, through Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS), and described then as Gardnerella leopoldii, Gardnerella piotii and Gardnerella swidsinskii, Therefore, not all Gardnerella species detected in several studies constituted Gardnerella vaginalis and could explain virulence differences between Gardnerella species (Iavazzo et al., 2008; Leite et al., 2010; Muzny and Schwebke, 2013). Since these species could not be delineated using full-length 16S rRNA gene sequences, Hill and colleagues applied partial chaperonin 60 (cpn60) sequences to resolve these four group species (Hill et al., 2019). Both studies showed that G. swidsinskii and G. leopoldii constituted group A, G. piotii corresponded to group B, G. vaginalis belonged to group C, and finally, group D was the most diverse subgroup with several Gardnerella sp. (such as strains 101, 1500E, 6119V5, and 00703Dmash). However, this last group will require an analysis of additional isolates to establish a species differentiation (Hill et al., 2019; Vaneechoutte et al., 2019). Nonetheless, an abundance of G. vaginalis and G. swidsinskii was associated with vaginal symptoms of abnormal odor and discharge in their study set (Hill et al., 2019). This heterogeneity and diversity within the genus Gardnerella may distinguish clades and how these features may impact BV development (Castro et al., 2020). So, future studies should isolate all Gardnerella species of the vaginal samples and further analysis could allow the qPCR methodology to quantify different species of Gardnerella.
As previously mentioned in results, statistically significant differences were found among the amount of Lactobacillus spp. between healthy and AV women (P < 0.001). Moreover, lactobacilli load among healthy women was established between 1E6 and 1E7 CFU/ml; meanwhile, the amount of Lactobacillus spp. in altered microbiota (AV) was defined between 1E3 and 1E5 CFU/ml. These results are comparable to previous studies with BV (Sha et al., 2005; De Backer et al., 2007; Ling et al., 2011). However, it is important to mention that the specificity of the lactobacilli primers (LactoF: 47.1%%; LactoR: 66.7%) was a limitation of the present study.
Furthermore, the results of the present study showed that both healthy and AV pregnant women have a higher concentration of Lactobacillus spp. when compared to non-pregnant women of the same categories. These results agree with Walther-António et al. (2014). These authors reported that lactobacilli augmentation during pregnancy and preterm birth help to prevent vaginal infection and counteract higher immune tolerance (Walther-António et al., 2014; Kim et al., 2017). However, there were no statistically significant differences between the amount of Lactobacillus spp. of pregnant and non-pregnant women per category. Although the present study is a preliminary analysis of lactobacilli load between pregnant and non-pregnant women, these results point to the possibility to use the same lactobacilli load range to evaluate AV and healthy vaginal microbiota (whether pregnant or non-pregnant) and thus to avoid future vaginal infection establishment in women by monitoring lactobacilli load through qPCR.
These results could corroborate with several studies, which postulated an increment of lactobacilli load in pregnant women (Aagaard et al., 2012; Walther-António et al., 2014). However, there are some major limitations of this study: (1) with 97 participants in PCR assays and 83 participants in qPCR assays, small numbers of particular cases were retained in each subgroup, (2) in PCR assays not all possible species of aerobic bacteria could be targeted in AV samples and, (3) in qPCR assays, normalized concentrations of lactobacilli were realized through low specificity primers for Lactobacillus spp. Therefore, future studies must optimize lactobacilli quantification, also quantify certain Lactobacillus species and other aerobic bacteria among pregnant and non-pregnant women. Previous studies showed that the presence of different Lactobacillus species is a major determinant to the stability of the vaginal microbial community in pregnancy (Verstraelen et al., 2009; Ling et al., 2010). Furthermore, Verstraelen and colleagues demonstrated L. crispatus ability to promote and stabilize the normal microbiota while L. gasseri and L. iners predisposed to some extent to the occurrence of abnormal microbiota (Verstraelen et al., 2009). Future studies should be realized with a bigger and more diverse population set as well as quantification of specific Lactobacillus species (such as L. crispatus, L. gasseri, and L. iners) as postulated by others authors (Verstraelen et al., 2009; Walther-António et al., 2014; Vaneechoutte, 2017a).
All datasets generated for this study are included in the article/Supplementary Material.
This study was approved by the Ethics Committee of Universidad San Francisco de Quito (USFQ) and the Ministry of Health of Ecuador (Protocol code: 2016-140M by MSP-SDM-10-2013-2019-O review board). The volunteers were recruited to our study set, after having read and signed the informed consent or, in the case of the underage volunteers, from their parents or legal representatives.
AM, GV, and DP-H were responsible for modeling and experimental design. Samples collection oversaw doctors of Gynecology and Obstetrics Service of Carlos Andrade Marin Hospital (HCAM), Gynecological-Obstetric Hospital Isidro Ayora (HGOIA), and Center for Teaching Health Cipriana Dueñas. The DNA extraction was performed at Hospital Carlos Andrade Marín by CC-B. Molecular characterization was conducted by DP-H in the Microbiology Institute at USFQ. Biostatistics analysis was developed by VB and DP-H at USFQ. All authors contributed to the article and approved the submitted version.
This work was supported by The Chancellor Grant 2018 and COCIBA research budget, under the Project ID: 5456 entitled (Caracterización de la microbiota vaginal y sus factores de riesgos en mujeres ecuatorianas).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
A special recognition deserves all the staff of the Microbiology Institute of USFQ, doctors involved in the project, and volunteer researchers, more exactly, Sully Márquez, Erika Montenegro, and Sebastián Almagro.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2020.00303/full#supplementary-material
Aagaard, K., Riehle, K., Ma, J., Segata, N., Mistretta, T.-A. A., Coarfa, C., et al. (2012). A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS ONE 7:e36466. doi: 10.1371/journal.pone.0036466
Alpay, S., Aydin, F., Kiliç, S. S., and Kiliç, A. O. (2003). Antimicrobial activity and characteristics of bacteriocins produced by vaginal lactobacilli received: turkish J. Med. Sci. 33, 7–13. Retrieved from http://journals.tubitak.gov.tr/medical/issues/sag-03-33-1/sag-33-1-2-0208-4.pdf
Aroutcheva, A., Gariti, D., Simon, M., Shott, S., Faro, J., Simoes, J. A., et al. (2001). Defense factors of vaginal lactobacilli. Am. J. Obstet. Gynecol. 185, 375–379. doi: 10.1067/mob.2001.115867
Begot, C., Desnier, I., Daudin, J. D., Labadie, J. C., and Lebert, A. (1996). Recommendations for calculating growth parameters by optical density measurements. J. Microbiol. Methods 25, 225–232. doi: 10.1016/0167-7012(95)00090-9
Bologno, R., Díaz, Y. M., Giraudo, M. C., Fernández, R., Menéndez, V., Brizuela, J. C., et al. (2011). Importance of studying the balance of vaginal content (BAVACO) in the preventive control of sex workers. Rev. Argent. Microbiol. 43, 246–250. doi: 10.1590/S0325-75412011000400002
Bolton, M., van der Straten, A., and Cohen, C. R. (2008). Probiotics: potential to prevent HIV and sexually transmitted infections in women. Sex. Transm. Dis. 35, 214–225. doi: 10.1097/OLQ.0b013e31815b017a
Borges, S., Silva, J., and Teixeira, P. (2014). The role of lactobacilli and probiotics in maintaining vaginal health. Arch. Gynecol. Obstet. 289, 479–489. doi: 10.1007/s00404-013-3064-9
Boris, S., and Barbés, C. (2000). Role played by lactobacilli in controlling the. Microbes Infect. 2, 543–546. doi: 10.1016/S1286-4579(00)00313-0
Carr, P. L., Felsenstein, D., and Friedman, R. H. (1998). Evaluation and management of vaginitis. J. Gen. Intern. Med. 13, 335–346. doi: 10.1046/j.1525-1497.1998.00101.x
Castro, J., Jefferson, K. K., and Cerca, N. (2020). Genetic heterogeneity and taxonomic diversity among gardnerella species. Trends Microbiol. 28, 202–211. doi: 10.1016/j.tim.2019.10.002
Cauci, S., Driussi, S., De Santo, D., Penacchioni, P., Iannicelli, T., Lanzafame, P., et al. (2002). Prevalence of bacterial vaginosis and vaginal flora changes in peri-and postmenopausal women downloaded. J. Clin. Microbiol. 40, 2147–2152. doi: 10.1128/JCM.40.6.2147-2152.2002
Collins, S. L., McMillan, A., Seney, S., van der Veer, C., Kort, R., Sumarah, M. W., et al. (2018). Promising prebiotic candidate established by evaluation of lactitol, lactulose, raffinose, and oligofructose for maintenance of a Lactobacillus-dominated vaginal microbiota. Appl. Environ. Microbiol. 84, e02200–17. doi: 10.1128/AEM.02200-17
Cristiano, L., Rampello, S., Noris, C., and Valota, V. (1996). Bacterial vaginosis: prevalence in an italian population of asymptomatic pregnant women and diagnostic aspects. Eur. J. Epidemiol. 12, 383–390. doi: 10.1007/BF00145302
Dai, Q., Hu, L., Jiang, Y., Shi, H., Liu, J., Zhou, W., et al. (2010). An epidemiological survey of bacterial vaginosis, vulvovaginal candidiasis and trichomoniasis in the Tibetan area of Sichuan Province, China. Eur. J. Obstet. Gynecol. Reprod. Biol. 150, 207–209. doi: 10.1016/j.ejogrb.2010.02.027
Datcu, R., Gesink, D., Mulvad, G., Montgomery-Andersen, R., Rink, E., Koch, A., et al. (2014). Bacterial vaginosis diagnosed by analysis of first-void-urine specimens. J Clin Microbiol. 52, 218–225. doi: 10.1128/JCM.02347-13
De Backer, E., Verhelst, R., Verstraelen, H., Alqumber, M. A., Burton, J. P., Tagg, J. R., et al. (2007). Quantitative determination by real-time PCR of four vaginal Lactobacillus species, gardnerella vaginalis and atopobium vaginae indicates an inverse relationship between L. gasseri and L. iners. BMC Microbiol. 7:115. doi: 10.1186/1471-2180-7-115
de Vos, W., Engstrand, L., Drago, L., Reid, G., Schauber, J., Hay, R., et al. (2012). Human Microbiota in Health and Disease. (SelfCare) 3, 1–68.
Donders, G., Bellen, G., and Rezeberga, D. (2011). Aerobic vaginitis in pregnancy. BJOG 118, 1163–1170. doi: 10.1111/j.1471-0528.2011.03020.x
Donders, G. G. G. (1999). Erratum: microscopy of the bacterial flora on fresh vaginal smears. Infect. Dis. Obstet. Gynecol. 7, 177–179. doi: 10.1155/S1064744999000290
Donders, G. G. G. (2007). Definition and classification of abnormal vaginal flora. Best Pract. Res. Clin. Obstet. Gynaecol. 21, 355–373. doi: 10.1016/j.bpobgyn.2007.01.002
Donders, G. G. G., Bellen, G., Grinceviciene, S., Ruban, K., and Vieira-Baptista, P. (2017). Aerobic vaginitis: no longer a stranger. Res. Microbiol. 168, 845–858. doi: 10.1016/j.resmic.2017.04.004
Donders, G. G. G., Van Calsteren, K., Bellen, G., Reybrouck, R., Van den Bosch, T., Riphagen, I., et al. (2009). Predictive value for preterm birth of abnormal vaginal flora, bacterial vaginosis and aerobic vaginitis during the first trimester of pregnancy. BJOG. 116, 1315–24. doi: 10.1111/j.1471-0528.2009.02237.x
Donders, G. G. G., Vereecken, A., Bosmans, E., Dekeersmaecker, A., Salembier, G., and Spitz, B. (2002). Definition of a type of abnormal vaginal flora that is distinct from bacterial vaginosis: aerobic vaginitis. Int. Congr. Ser. 1279, 118–129. doi: 10.1016/j.ics.2005.02.064
Donders, G. G. G., Vereecken, A., Bosmans, E., Dekeersmaecker, A., Salembier, G., and Spitz, B. (2005). Aerobic vaginitis: Abnormal vaginal flora entity that is distinct from bacterial vaginosis. Int. Congr. Ser. 1279, 118–129. doi: 10.1016/j.ics,.2005.02.064
DTU National Food Institute (2014). Protocol for PCR amplification of E. faecium and E. faecalis Recommended by the EURL-AR. 5–8. DTU National Food Institute.
Eschenbach, D., Patton, D., Meier, A., Thwin, S., Aura, J., Stapleton, A., et al. (2000). Effects of intrauterine device and oral contraceptive on vaginal flora and epithelium. Saudi Med. J. 62, 107–112. doi: 10.1016/S0010-7824(00)00155-4
Fan, A., Yue, Y., Geng, N., Zhang, H., Wang, Y., and Xue, F. (2013). Aerobic vaginitis and mixed infections: comparison of clinical and laboratory findings. Arch. Gynecol. Obstet. 287, 329–335. doi: 10.1007/s00404-012-2571-4
Farage, M. A., Miller, K. W., and Sobel, J. D. (2010). Dynamics of the Vaginal Ecosystem-Hormonal Influences. Res. Treat. 3, 1–15. doi: 10.4137/IDRT.S3903
Fredricks, D. N. (2011). Molecular methods to describe the spectrum and dynamics of the vaginal microbiota. Anaerobe 17, 191–195. doi: 10.1016/j.anaerobe.2011.01.001
Fredricks, D. N., Fiedler, T. L., Thomas, K. K., Oakley, B. B., and Marrazzo, J. M. (2007). Targeted PCR for detection of vaginal bacteria associated with bacterial vaginosis. J. Clin. Microbiol. 45, 3270–3276. doi: 10.1128/JCM.01272-07
Gajer, P., Brotman, R. M., Bai, G., Sakamoto, J., Schütte, U. M. E., Zhong, X., et al. (2012). Temporal dynamics of the human vaginal microbiota. Sci. Transl. Med. 4:132ra52. doi: 10.1126/scitranslmed.3003605
Galán, A., Veses, V., Murgui, A., Casanova, M., and Martínez, J. P. (2006). Rapid PCR-based test for identifying Candida albicans by using primers derived from the pH-regulated KER1 gene. FEMS Yeast Res. 6, 1094–1100. doi: 10.1111/j.1567-1364.2006.00114.x
Gondo, F., Da Silva, M. G., Polettini, J., Tristao, A. D. R., Peracoli, J. C., Witkin, S. S., et al. (2011). Vaginal flora alterations and clinical symptoms in low-risk pregnant women. Gynecol. Obstet. Invest. 71, 158–162. doi: 10.1159/000316051
Hardy, L., Jespers, V., Van Bulck, M., Den, B.uyze, J., Mwambarangwe, L., Musengamana, V., et al. (2017). The presence of the putative Gardnerella vaginalis sialidase A gene in vaginal specimens is associated with bacterial vaginosis biofilm. PLoS ONE 12:e0172522. doi: 10.1371/journal.pone.0172522
Henriques, A., Cereija, T., Machado, A., and Cerca, N. (2012). In silico vs in vitro analysis of primer specificity for the detection of Gardnerella vaginalis, Atopobium vaginae and Lactobacillus spp. BMC Res. Notes 5:637. doi: 10.1186/1756-0500-5-637
Hernández-Rodríguez, C., Romero-González, R., Albani-Campanario, M., Figueroa-Damián, R., Meraz-Cruz, N., and Hernández-Guerrero, C. (2011). Vaginal microbiota of healthy pregnant mexican women is constituted by four lactobacillus species and several vaginosis-associated bacteria. Infect. Dis. Obstet. Gynecol. 2011:851485. doi: 10.1155/2011/851485
Hill, J. E., Albert, A., and VOGUE Research Group (2019). Resolution and cooccurrence patterns of Gardnerella leopoldii, G. swidsinskii, G. piotii, and G. vaginalis within the Vaginal Microbiome. Infect. Immun. Immun 87:e00532–19. doi: 10.1128/IAI.00532-19
Iavazzo, C., Vogiatzi, C., and Falagas, M. E. (2008). A retrospective analysis of isolates from patients with vaginitis in a private Greek obstetric/gynecological hospital (2003-2006). Med. Sci. Monit. 14:CR228–31.
Jahic, M., Mulavdic, M., Nurkic, J., Jahic, E., Nurkic, M., and Clinic, G. (2013). Clinical characteristics of aerobic vaginitis and its association to vaginal candidiasis, trichomonas vaginitis and bacterial vaginosis. Med. Arhit. 67, 428–430. doi: 10.5455/medarh.2013.67.428-430
Johnson, C. L., and Versalovic, J. (2012). The human microbiome and its potential importance to pediatrics. Pediatrics 129, 950–960. doi: 10.1542/peds.2011-2736
Keane, F. E. A., Ison, C. A., and Taylor-Robinson, D. (1997). A longitudinal study of the vaginal flora over a menstrual cycle. Int. J. STD AIDS 8, 489–494. doi: 10.1258/0956462971920631
Kim, J. H., Yoo, S. M., Sohn, Y. H., Jin, C. H., Yang, Y. S., Hwang, I. T., et al. (2017). Predominant Lactobacillus species types of vaginal microbiota in pregnant Korean women: quantification of the five Lactobacillus species and two anaerobes. J. Matern. Neonatal Med. 30, 2329–2333. doi: 10.1080/14767058.2016.1247799
Larsen, B., and Monif, G. R. G. (2001). Understanding the bacterial flora of the female genital tract. Clin. Infect. Dis. 32, e69–e77. doi: 10.1086/318710
Leite, S. R., de, F., Amorim, M. M. R., Calabria, W. B., Leite, T. N. de, F., Oliveira, V. S., et al. (2010). Clinical and microbiological profile of women with bacterial vaginosis. Rev. Bras. Ginecol. Obs. 32, 82–87. doi: 10.1590/S0100-72032010000200006
Li, J., McCormick, J., Bocking, A., and Reid, G. (2012). Importance of vaginal microbes in reproductive health. Reprod. Sci. 19, 235–242. doi: 10.1177/1933719111418379
Ling, Z., Kong, J., Liu, F., Zhu, H., Chen, X., Wang, Y., et al. (2010). Molecular analysis of the diversity of vaginal microbiota associated with bacterial vaginosis. BMC Genomics 11:488. doi: 10.1186/1471-2164-11-488
Ling, Z., Liu, X., Chen, X., Zhu, H., Nelson, K. E., Xia, Y., et al. (2011). Diversity of cervicovaginal microbiota associated with female lower genital tract infections. Microbiol. Ecol. 61, 704–714. doi: 10.1007/s00248-011-9813-z
Ling, Z., Liu, X., Luo, Y., Wu, X., Yuan, L., Tong, X., et al. (2013). Associations between vaginal pathogenic community and bacterial vaginosis in chinese reproductive-age women. PLoS ONE 8:e76589. doi: 10.1371/journal.pone.0076589
Machado, A., Jefferson, K. K., and Cerca, N. (2013). Interactions between Lactobacillus crispatus and bacterial vaginosis (BV)-associated bacterial species in initial attachment and biofilm formation. Int. J. Mol. Sci. 14, 12004–12012. doi: 10.3390/ijms140612004
Marconi, C., Donders, G. G. G., Martin, L. F., Ramos, B. R. A., Duarte, M. T. C., Parada, C. M. G. L., et al. (2012). Chlamydial infection in a high risk population: association with vaginal flora patterns. Arch. Gynecol. Obstet. 285, 1013–1018. doi: 10.1007/s00404-011-2085-5
Marot-Leblond, A., Nail-Billaud, S., Pilon, F., Beucher, B., Poulain, D., and Robert, R. (2009). Efficient diagnosis of vulvovaginal candidiasis by use of a new rapid immunochromatography test. J. Clin. Microbiol. 47, 3821–3825. doi: 10.1128/JCM.01168-09
Mendling, W. (2016). Microbiota of the human body. Adv. Exp. Med. Biol. 902, 83–93. doi: 10.1007/978-3-319-31248-4_6
Muzny, C. A., and Schwebke, J. R. (2013). Gardnerella vaginalis: still a prime suspect in the pathogenesis of bacterial vaginosis. Curr. Infect. Dis. Rep. 15, 130–135. doi: 10.1007/s11908-013-0318-4
Mytilinaios, I., Salih, M., Schofield, H. K., and Lambert, R. J. W. (2012). Growth curve prediction from optical density data. Int. J. Food Microbiol. 154, 169–176. doi: 10.1016/j.ijfoodmicro.2011.12.035
Naghili, H., Tajik, H., Mardani, K., Mehdi, S., Rouhani, R., Ehsani, A., et al. (2013). Validation of drop plate technique for bacterial enumeration by parametric and nonparametric tests. Vet. Res. Forum. 4, 179–183.
Nelson, D. B., Hanlon, A., Hassan, S., Britto, J., Geifman-Holtzman, O., Haggerty, C., et al. (2009). Preterm labor and bacterial vaginosis-associated bacteria among urban women. J. Perinat. Med. 37, 130–134. doi: 10.1515/JPM.2009.026
Nugent, R. P., Krohn, M. A., and Hillier, S. L. (1991). Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. J. Clin. Microbiol. 29, 297–301. doi: 10.1128/JCM.29.2.297-301.1991
Oliveira, L. M. A., Diniz, C. G., Fernandes, A. A. S., De Souza-Sotte, D. M. K., De Freitas, M. C. R., Machado, A. B. F., et al. (2018). Assessment of vaginal microbiota in Brazilian women with and without bacterial vaginosis and comparison with Nugent score. J. Infect. Dev. Ctries. 12, 127–136. doi: 10.3855/jidc.9532
Petrova, M. I., van den Broek, M., Balzarini, J., Vanderleyden, J., and Lebeer, S. (2013). Vaginal microbiota and its role in HIV transmission and infection. FEMS Microbiol. Rev. 37, 762–792. doi: 10.1111/1574-6976.12029
Price, E. P., Dale, J. L., Cook, J. M., Sarovich, D. S., Seymour, M. L., Ginther, J. L., et al. (2012). Development and validation of burkholderia pseudomallei-specific real-time pcr assays for clinical, environmental or forensic detection applications. PLoS ONE 7:e37723. doi: 10.1371/journal.pone.0037723
Ravel, J., Gajer, P., Abdo, Z., Schneider, G. M., Koenig, S. S. K., Mcculle, S. L., et al. (2011). Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. U.S.A. 108, 4680–4687. doi: 10.1073/pnas.1002611107
Romero, R., Hassan, S. S., Gajer, P., Tarca, A. L., Fadrosh, D. W., Nikita, L., et al. (2014). The composition and stability of the vaginal microbiota of normal pregnant women is different from that of non-pregnant women. Microbiome 2:4. doi: 10.1186/2049-2618-2-10
Rumyantseva, T. A., Bellen, G., Savochkina, Y. A., Guschin, A. E., and Donders, G. G. G. (2016). Diagnosis of aerobic vaginitis by quantitative real-time PCR. Arch. Gynecol. Obstet. 294, 109–114. doi: 10.1007/s00404-015-4007-4
Santiago, G., Deschaght, P., El Aila, N., Kiama, T., Verstraelen, H., Jefferson, K., et al. (2011). Gardnerella vaginalis comprises three distinct genotypes of which only two produce sialidase. Am. J. Obs. Gynecol. 204, 450.e1–e7. doi: 10.1016/j.ajog.2010.12.061
Schwebke, J. R., Hillier, S. L., Sobel, J. D., McGregor, J. A., and Sweet, R. L. (1996). Validity of the vaginal gram stain for the diagnosis of bacterial vaginosis. Obstet. Gynecol. 88, 573–576. doi: 10.1016/0029-7844(96)00233-5
Schwebke, J. R., Richey, C. M., and Weiss, H. L. (1999). Correlation of behaviors with microbiological changes in vaginal flora. J. Infect. Dis. 180, 1632–1636. doi: 10.1086/315065
Sepehri, S., Kotlowski, R., Bernstein, C. N., and Krause, D. O. (2009). Phylogenetic analysis of inflammatory bowel disease associated Escherichia coli and the FimH virulence determinant. Inflamm. Bowel Dis. 15, 1737–1745. doi: 10.1002/ibd.20966
Sha, B. E., Chen, H. Y., Wang, Q. J., Zariffard, M. R., Cohen, M. H., and Spear, G. T. (2005). Utility of amsel criteria, nugent score, and quantitative pcr for for diagnosis of bacterial vaginosis in human immunodeficiency virus-infected women. J. Clin. Mircobiol. 43, 4607–4612. doi: 10.1128/JCM.43.9.4607-4612.2005
Srinivasan, S., and Fredricks, D. N. (2008). The human vaginal bacterial biota and bacterial vaginosis. Interdiscip. Perspect. Infect. Dis. 2008:750479. doi: 10.1155/2008/750479
Svanemyr, J., Guijarro, S., Riveros, B. B., and Chandra-Mouli, V. (2017). The health status of adolescents in Ecuador and the country's response to the need for differentiated healthcare for adolescents. Reprod. Health 14, 1–11. doi: 10.1186/s12978-017-0294-5
Tansarli, G. S., Kostaras, E. K., Athanasiou, S., and Falagas, M. E. (2013). Prevalence and treatment of aerobic vaginitis among non-pregnant women: evaluation of the evidence for an underestimated clinical entity. Eur. J. Clin. Microbiol. Infect. Dis. 32, 977–984. doi: 10.1007/s10096-013-1846-4
Tempera, G., and Furneri, P. M. (2010). Management of aerobic vaginitis. Gynecol. Obstet. Invest. 70, 244–249. doi: 10.1159/000314013
Van De Wijgert, J. H. H. M., Borgdorff, H., Verhelst, R., Crucitti, T., Francis, S., Verstraelen, H., et al. (2014). The vaginal microbiota: What have we learned after a decade of molecular characterization? PLoS ONE 9:e105998. doi: 10.1371/journal.pone.0105998
Vaneechoutte, M. (2017a). Lactobacillus iners, the unusual suspect. Res. Microbiol. 168, 826–836. doi: 10.1016/j.resmic.2017.09.003
Vaneechoutte, M. (2017b). The human vaginal microbial community. Res. Microbiol. 168, 811–825. doi: 10.1016/j.resmic.2017.08.001
Vaneechoutte, M., Guschin, A., Van Simaey, L., Gansemans, Y., Van Nieuwerburgh, F., and Cools, P. (2019). Emended description of Gardnerella vaginalis and description of Gardnerella leopoldii sp. Nov., Gardnerella piotii sp. nov. and Gardnerella swidsinskii sp. nov., with delineation of 13 genomic species within the genus Gardnerella. Int. J. Syst. Evol. Microbiol. 69, 679–687. doi: 10.1099/ijsem.0.003200
Verstraelen, H., Verhelst, R., Claeys, G., De Backer, E., Temmerman, M., and Vaneechoutte, M. (2009). Longitudinal analysis of the vaginal microflora in pregnancy suggests that L. crispatus promotes the stability of the normal vaginal microflora and that L. gasseri and/or L. iners are more conducive to the occurrence of abnormal vaginal microflora. BMC Microbiol. 9:116. doi: 10.1186/1471-2180-9-116
Vieira-Baptista, P., Grinceviciene, S., Bellen, G., Sousa, C., Saldanha, C., Broeck, D. V., et al. (2017). Genital tract infections in an isolated community: 100 women of the Pr?ncipe Island. Infect Dis Obstet Gynecol. 2017, 1–6. doi: 10.1155/2017/3058569
Von Gruenigen, V. E., Coleman, R. L., Li, A. J., Heard, M. C., Miller, D. S., and Hemsell, D. L. (2000). Bacteriology and treatment of malodorous lower reproductive tract in gynecologic cancer patients. Obstet. Gynecol. 96, 23–27. doi: 10.1016/S0029-7844(00)00850-4
Walther-António, M. R. S., Jeraldo, P., Berg Miller, M. E., Yeoman, C. J., Nelson, K. E., Wilson, B. A., et al. (2014). Pregnancy's stronghold on the vaginal microbiome. PLoS ONE 9:e98514. doi: 10.1371/journal.pone.0098514
Xu, C., Zhang, W., Wu, M., and Zhang, S. (2010). Prevalence and determinants of lower reproductive tract infections among women aged 25 - 54 years in Beijing. Zhonghua Liu Xing Bing Xue Za Zhi 31, 138–141.
Zhou, X., Bent, S., Schneider, M., David, C., Islam, M., and Forney, L. (2004). Characterization of vaginal microbial communities in adult healthy women using cultivation-independent methods. Microbiology 150, 2565–2573. doi: 10.1099/mic.0.26905-0
Keywords: vaginal microbiota, vaginal infection, bacterial vaginosis, aerobic vaginitis, pregnant, opportunistic pathogen, Lactobacillus spp.
Citation: Pacha-Herrera D, Vasco G, Cruz-Betancourt C, Galarza JM, Barragán V and Machado A (2020) Vaginal Microbiota Evaluation and Lactobacilli Quantification by qPCR in Pregnant and Non-pregnant Women: A Pilot Study. Front. Cell. Infect. Microbiol. 10:303. doi: 10.3389/fcimb.2020.00303
Received: 20 September 2019; Accepted: 20 May 2020;
Published: 19 June 2020.
Edited by:
Mario Vaneechoutte, Ghent University, BelgiumCopyright © 2020 Pacha-Herrera, Vasco, Cruz-Betancourt, Galarza, Barragán and Machado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: António Machado, YW1hY2hhZG9AdXNmcS5lZHUuZWM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.