 Kelly Cristina Oliveira de Albuquerque1Andreza do Socorro Silva da Veiga2Fernando Tobias Silveira3Marliane Batista Campos3Ana Paula Lima da Costa4Ananda Karolyne Martins Brito5Paulo Ricardo de Souza Melo6
Kelly Cristina Oliveira de Albuquerque1Andreza do Socorro Silva da Veiga2Fernando Tobias Silveira3Marliane Batista Campos3Ana Paula Lima da Costa4Ananda Karolyne Martins Brito5Paulo Ricardo de Souza Melo6 Sandro Percario1
Sandro Percario1 Fábio Alberto de Molfetta4
Fábio Alberto de Molfetta4 Maria Fâni Dolabela1,2,5,6*
Maria Fâni Dolabela1,2,5,6*- 1Biotechnology and Biodiversity Postgraduate Program (BIONORTE), Federal University of Pará, Belém, PA, Brazil
- 2Pharmaceutical Innovation Postgraduate Program, Federal University of Pará, Belém, PA, Brazil
- 3Leishmaniasis Laboratory, Evandro Chagas Institute, Ananindeua, PA, Brazil
- 4Laboratory of Molecular Modeling, Institute of Exact and Natural Sciences, Federal University of Pará, Belém, PA, Brazil
- 5Faculty of Pharmacy, Federal University of Pará, Belém, PA, Brazil
- 6Pharmaceutical Sciences Postgraduate Program, Federal University of Pará, Belém, PA, Brazil
Introduction: Leishmaniasis is caused by protozoa of the genus Leishmania, classified as tegumentary and visceral. The disease treatment is still a serious problem, due to the toxic effects of available drugs, the costly treatment and reports of parasitic resistance, making the search for therapeutic alternatives urgent. This study assessed the in vitro anti-leishmanial potential of the extract, fractions, and isoeleutherin from Eleutherine plicata, as well as the in silico interactions of isoeleutherin and its analogs with Trypanothione Reductase (TR), in addition to predicting pharmacokinetic parameters.
Methods: From the ethanolic extract of E. plicata (EEEp) the dichloromethane fraction (FDEp) was obtained, and isoeleutherin isolated. All samples were tested against promastigotes, and parasite viability was evaluated. Isoeleutherin analogues were selected based on similarity in databases (ZINK and eMolecules) to verify the impact on structural change.
Results and Discussion: The extract and its fractions were not active against the promastigote form (IC50 > 200 μg/mL), while isoeleutherin was active (IC50 = 25 μg/mL). All analogues have high intestinal absorption (HIA), cell permeability was moderate in Caco2 and low to moderate in MDCK. Structural changes interfered with plasma protein binding and blood-brain barrier permeability. Regarding metabolism, all molecules appear to be CYP3A4 metabolized and inhibited 2–3 CYPs. Molecular docking and molecular dynamics assessed the interactions between the most stable configurations of isoeleutherin, analogue compound 17, and quinacrine (control drug). Molecular dynamics simulations demonstrated stability and favorable interactions with TR. In summary, fractionation contributed to antileishmanial activity and isoleutherin seems to be promising. Structural alterations did not contribute to improve pharmacokinetic aspects and analogue 17 proved to be more promising than isoeleutherin, presenting better stabilization in TR.
1 Introduction
American tegumentary leishmaniasis (ATL), an infectious and non-contagious disease, is caused by protozoa of the genus Leishmania, with 7 species of parasites responsible for the disease in Brazil (WHO, 2020). Since 2015, a downward trend in ATL cases has been reported in 17 endemic countries in the Americas, and Brazil reported the highest number of cases registered in 2019 (OPAS, 2020).
For the treatment of ATL, pentavalent antimonials and amphotericin B are mainly used, drugs that have high toxicity, are parenterally administered, high-cost, and there are reports of lack of therapeutic response and parasite resistance to antimonials (Ponte-Sucre et al., 2017; Mann et al., 2021). Such resistance has been associated with an increase in the production levels of trypanothione proteins by the resistant parasites (Mukhopadhyay et al., 1996).
Trypanothione Reductase (TR) is an enzyme found in flagellated protozoa of the Leishmania genus and plays a crucial role in regulating the oxidative stress in these parasites. This NADPH-dependent flavoenzyme functions to control the concentration of reactive oxygen species and is, therefore, a potential target for research in the development of selective inhibitors (Mukherjee et al., 2020). Given the above, it is urgent to search for therapeutic alternatives that act on parasites resistant to antimonials, which can be administered orally and with less toxic potential.
Medicinal plants from the Amazon can be a promising source of leishmanicidal drugs, with some species being used to treat difficult-to-heal wounds (Silva et al., 2018). Eleutherine plicata Herb. is widely used in Amazonian folk medicine for the treatment of amoebiasis, liver diseases, parasitic infections, hemorrhages, anemia (Couto et al., 2016), as well as for the healing of superficial wounds and gastric ulcers (Villegas et al., 1997). Its main chemical constituents are naphthoquinones, including isoeleutherin (Figure 1A), eleutherin (Figure 1B), and eleutherol (Figure 1C), isolated from the bulb extract of this species (Figure 1) (Malheiros et al., 2015; Vale et al., 2020). Naphthoquinones eleutherin and isoeleutherin have been associated with the biological activities of the species (Paramapojn et al., 2008), and other studies have suggested their potential as potent trypanocidal and anticancer agents (Silva-Junior et al., 2019; Almeida et al., 2020; Castro et al., 2021b).
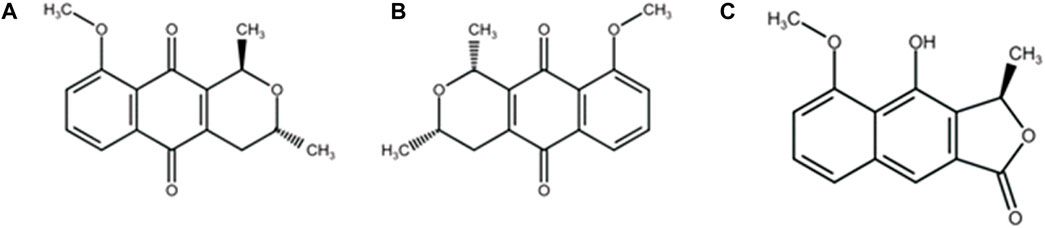
FIGURE 1. Compounds isolated from Eleutherine plicata Herb: (A) isoeleutherin; (B) eleutherin; (C) eleutherol.
Toxicity assays in the Allium cepa model demonstrated eleutherin caused a higher percentage of chromosomal aberrations than isoeleutherin (12.5 μg/mL in 72 h). In the micronucleus assay, isoeleutherin showed low genotoxic potential, with a low frequency of micronuclei (Castro et al., 2021a).
Other study evaluated the genotoxicity of ethanol Extract (EEEp), Dichloromethane Fraction (FDCMEp) and isoeleutherin isolated from Eleutherine plicata, using the micronucleus test, is eleutherin was less genotoxic. Isoeleutherin and analogues were subjected to in silico toxicity prediction, and compounds free of toxicological risks (CP13, CP14, CP17, and isoeleutherin) were selected for molecular docking in Topoisomerase II. The structural changes suggest an increase in affinity with the TOPO II enzyme, observed in the increase in the amount of hydrogen bond interactions performed with amino acid residues of the active site (Albuquerque et al., 2023). The present study evaluated the leishmanicidal activity of isoeleutherin against the species of Leishmania amazonensis, associating in silico assays to search for new bioactive molecules.
2 Methodology
2.1 Plant material and isolation of isoeleutherin
Bulbs of E. plicata were collected in Tracuateua, Pará, Brazil (Lat. 1.1436°, Long. 46.9551°), a specimen was deposited at Museu Paraense Emílio Goeldi (MG 202631). The research project complies with national guidelines and international legislation, registered on the platform of the National System of Management and Genetic Heritage and Associated Traditional Knowledge (SISGEN), under registration number A49DEEE that grants license for collecting the species. The EEEp was obtained by macerating the dry powder of the bulbs (924 g) in ethanol (2 L for 7 days), subjected to fractionation in an open chromatographic column, using silica gel mesh (63–200 mm) as stationary phase and mobile solvents of increasing polarity (hexane, dichloromethane, ethyl acetate and methanol), obtaining the fractions: FHEp, FDEp, FAEEp, and FMEp, concentrated in a rotary evaporator. The FDEp was subjected to fractionation in thin layer chromatography on a preparative scale and showed four yellow spots with different retention factors, which were removed separately, being named Subfraction FA1, FA2, FA3, and FA4. From FA3, isoeleutherin was isolated, and identified by nuclear magnetic resonance (NMR) spectra, using a Bruker Advance DPX 400 MHz NMR spectrometer (Bruker Ascend).
2.2 Leishmanial activity against promastigotes and amastigotes of L. amazonensis
Inhibition of Leishmania growth was evaluated in vitro by cultivating promastigotes of L. amazonensis in stationary phase (5×106 parasites) in the presence of the extract, fractions and isoeleutherin (200–3.125 μg/mL) in 96-well culture plates (Nunc, Nunclon®, Roskilde, Denmark), for 72 h at 26°C. Viability was assessed by measuring the cleavage of MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide] (Sigma) (Ngure et al., 2009). Absorbances were measured using a multiwell scanning spectrophotometer (Molecular Devices, Spectra Max Plus, Canada) at 490 nm. Amphotericin B (AmpB) was used as a positive control (25–0.3906 μg/mL). The concentration of products required to inhibit 50% of the viability of L. amazonensis (IC50) was determined by applying a sigmoidal regression of the individual concentration response curves of the compounds. The data are representative of independent experiments, carried out in triplicate, which presented similar results (Mota et al., 2011).
Murine macrophages (4 × 105 cells) were seeded on round glass coverslips into 24-well culture plates (Nunc) in RPMI 1640 medium (Sigma and Aldrich), supplemented with 20% fetal bovine serum (FBS), 2 mM L-glutamine, penicillin 50 IU/mL, and streptomycin 50 μg/mL, pH 7.4. After 24 h of incubation at 35°C in 5% CO2, promastigotes of L. amazonensis in stationary phase were added to the wells (4 × 106 parasites) to promote infection of macrophages, and the cultures were incubated for 4 h at 35°C in 5% CO2. Next, the free parasites were removed by extensive washing with RPMI 1640 medium, and the infected macrophages were quantified and treated with the extract, FA2, FA3, fraction FA4, and isoeleutherin (500–125 μg/mL, each) for 72 h at 35°C in 5% CO2. The negative control consisted of infected macrophages and culture medium. The positive control used AmpB (100–25 μg/mL). Then, coverslips were stained with Giemsa, and the percentage of inhibition of intramacrophage viability of Leishmania was determined by counting the number of amastigotes per 100 macrophages on each coverslip, under a light microscope (×100 magnification). The presented data were performed in triplicate, and the IC50 was determined using GraphPad Prism version 5.04 (Silva, 2005).
2.3 Prediction studies of physical-chemical and pharmacokinetic aspects
Free online platform PreADMET (https://preadmet.webservice.bmdrc.org/) was used to predict absorption, distribution, metabolism, and elimination properties. Molecular descriptors related to Lipinski’s rule of five (Lipinski et al., 2001) and Veber extensions (Veber et al., 2002), such as molar mass (MW), octanol/water partition coefficient (cLog P), number of hydrogen bond acceptor (HBA) and hydrogen bond donor (HBD), number of rotatable bonds (nRotb) and topological polar surface area (TPSA) were predicted in Molinspiration (https://www.molinspiration.com) (Molinspiration, 2023).
Bioavailability prediction considered Lipinski’s “Rule of Five”, where a good drug candidate will have molecular weight <500, partition coefficient (log P) < 5, no more than five hydrogen bond donors and ten acceptors of hydrogen bonding (Lipinski et al., 2001). In the pharmacokinetic studies, intestinal absorption (Human Intestinal Absorption = HIA) was evaluated, considering parameters in the range of 0%–20% (low absorption), 20%–70% (moderate absorption), >70% (high absorption; (Hou et al., 2007). Molecule permeability in Caco-2 and MDCK cells were considered high when presented values >70 nm/s, average of 4–70 nm/s and low <4 nm/s (Yee, 1997; Yazdanian et al., 1998; Balimane et al., 2000). For distribution analysis, values >90% indicate strong binding to albumin, while <90% binding will be moderate to weak (Sun et al., 2018). As for the ability to cross the blood-brain barrier, the following criteria were used: freely crosses BBB >2.0, moderately crosses BBB values between 2.0 and 0.1 and reduced crossing or not crosses <0.1 (Ajay and Murcko, 1999).
2.4 Molecular docking simulations
Molecular docking was used to explore the possible conformations of the ligand with the binding receptor, estimating the intensity of enzyme-ligand interaction (Meng et al., 2011). Isoeleutherin and the subset derived from isoeleutherin analogue compounds were prepared from SMILES files downloaded from the ZINC (Irwin et al., 2012) and eMolecules (www.emolecules.com) databases (eMolecules, 2023). The crystallographic structure of the TR enzyme was retrieved from the Protein Data Bank (PDB) under the code 2JK6 (Baiocco et al., 2009) with a resolution of 2.95Å, prepared using the Chimera program (Lang et al., 2009), removing water molecules, ligands and adding hydrogen atoms. Molecular docking simulations were performed using the GOLD 2020.1 program (Cambridge Crystallo-graphic Data Center—CCDC, Cambridge, United Kingdom), which uses a genetic algorithm to generate and select conformations of flexible compounds that bind to the receptor site of a protein (Jones et al., 1995).
Compounds were scored by applying the GoldScore scoring function with a 100% efficient search. The binding site was defined based on studies by Padey et al. (2016), at a 10Å sphere centered on the flavin-adenine dinucleotide ligand (FAD); (Pandey et al., 2017a; Pandey et al., 2017b). The methodology was validated through redocking, evaluated using the fconv 1.24 program (Neudert and Klebe, 2011). The procedure was carried out to evaluate the convergence of the results and to determine the smallest value of the root mean square deviation (RMSD), making it possible to select the best interactions of the crystallographic ligand in the complex formed. To analyze the hydrogen bonds and hydrophobic interactions between the selected ligands and the enzyme’s amino acids, the PoseView online server was used (Stierand et al., 2006), a tool that displays molecular complexes that incorporate a simple and easy-to-perceive arrangement of the interactions formed between ligands and amino acids (Stierand and Rarey, 2007). Quinacrine was used as a control drug, which was chosen because it is a small molecule like the ones studied and because it has activity on TR (Saravanamuthu et al., 2004).
2.5 Molecular dynamics simulations
Firstly, the calculation of electrostatic potential charge was performed for the structure obtained during docking using the Gaussian 03 program (Frisch et al., 2003), applying the Restricted Electrostatic Potential (RESP) (Bayly et al., 1993) associated with the Hartree-Fock methods (Fock, 1930) and HF/6-31G (d,p) base function (Hariharan and Pople, 1973). The protonation states of all amino acid residues present in the enzyme were determined at pH 7.0 using the Propka server (Olsson et al., 2011). Molecular dynamics simulations were conducted using the AMBER 18 program, implemented by the pmemd. CUDA module (Li et al., 2013). The general AMBER force field (GAFF) (Wang et al., 2004) was employed to describe ligands, and the MMFF99SB force field (Hornak et al., 2006) was used for enzyme amino acid residues. The enzyme-ligand complex was solvated with the TIP3P explicit solvent model in a cubic box with an edge length of 12 Å, with the inclusion of Clˉ counter-ions to achieve electrical neutrality in the system, using the tleap module included in the AMBER program (Jorgensen et al., 1983).
Minimizations and heating were carried out with a SANDER module (Case et al., 2005), the systems were fragmented into four stages of energy minimization. In the first phase, 25,000 minimization steps were performed, divided into 10,000 steps, performed with the steepest descent method and 15,000 with the conjugate gradient. The remaining three phases were performed in 10,000 minimization steps, using the same methodology for each step. Subsequently, the systems were gradually heated using the Langevin algorithm, with protein atoms subjected to a restriction constant of 25 kcal/mol. Å2, considering the NVT set from 0 K to 298 K (Loncharich et al., 1992).
Periodic boundary conditions were simulated using the Particle Mesh Ewald (PME) method, employed for long-range electrostatic interactions (Darden et al., 1993). Cutoff distances for the long-range and van der Waals interactions were set at 9 Å. After minimization and equilibrium of the system, during the period of 50 ns of DM simulation, an integration time of 2.0 fs was produced using the Verlet algorithm (Verlet, 1968), considering the NPT set adjusted to a temperature of 298 K and pressure of 1 atm for each enzymatic binding complexes, all bonds with hydrogen atoms were restricted using the SHAKE algorithm (Ryckaert et al., 1977). The CPPTRAJ module of AMBERTOOLS 18 (Roe and Cheatham, 2013) was used to carry out the structural analyzes of RMSD and B-factor. The RMSD calculation verified the stability of the systems in relation to the initial structure, and the application of the B-factor identified which amino acid residues were most flexible in the enzyme. The binding free energy calculation was performed in the last 10 ns, using the AMBERTOOLS 18 modules CPPTRAJ and MMPBSA. py (Miller et al., 2012). The single-path MM-PB(GB)/SA protocol considers identical conformation states of the protein-ligand complex, unbound protein and free ligand (Kollman et al., 2000; Massova and Kollman, 2000).
The binding free energy (ΔGbind) of the complexes was determined according to Eq. 1, where ΔH is the enthalpy term, TΔS represents the product between absolute temperature and entropy resulting from the conformations obtained in the DM simulation, ΔEMM consists of the energy obtained by molecular mechanics and ΔGbind, solv is the free energy of solvation. Entropy contributions to energy result from changes in translation, rotation, and vibration.
The mechanical energies, represented by Eq. 2, are calculated involving the individual contributions of internal energy (ΔEint), electrostatic (ΔEele) and van der Waals (ΔEvdW).
The sum to compose the internal energy, described in Eq. 3, contains the contributions of bond length (ΔEbond), bond angles (ΔEangle) and torsion angles (ΔEtorsion).
Solvation free energy (ΔGbonding, solv) results from the sum of polar (ΔGPB/GB) and non-polar (ΔGn-polar) contributions. According to Eq. 4, the polar electrostatic contribution to the solvation free energy can be calculated by the Poisson-Boltzmann method (PB) or by the generalized methods of Born approximation (GB). ΔGPB and ΔGGB were calculated using the generalized Born model (igb = 2) (Onufriev et al., 2000) and in the MMPBSA method, considering the dielectric constants of solute (Ajay and Murcko, 1999) and solvent (Yazdanian et al., 1998; Sun et al., 2014).
The nonpolar energy is estimated by the product of the surface tension γ with a value equal to 0.0072 kcal/mol Å2 and the surface accessible solvent area (SASA) according to Eq. 5.
The interaction free energy that allows determining the individual contribution of all residues to the free energy of the complexes, obtained by the MMGBSA method, is described in Eq. 6, elucidating its importance in the active site of the enzyme (Alves et al., 2020).
The main residues that contribute to the total energy were visualized using the CHEWD plugin (Raza et al., 2019) of the Chimera program (Pettersen et al., 2004). The van der Waals (ΔGvdW) and electrostatic (ΔGele) interactions between amino acid residues of the TR enzyme were determined by the SANDER module, implemented in the AMBER 18 program.
3 Results
3.1 Phytochemistry, leishmanicidal activity, and cytotoxicity
From the ethanolic extract (EEEp; yield 2.84%) the following fractions were obtained: hexane (FHEp; yield 3.1%), dichloromethane (FDEp; yield 19.6%), ethyl acetate (FAEEp; yield 10.2%) and methanol (FMEp; 61.9% yield). Isoeleutherin was isolated from FDEp and its identification is described by Borges et al. (2020).
In vitro anti-leishmanial assays were conducted to evaluate the effect of the samples (EEEp, FDEp, FAEEp, FMEp, and isoeleutherin) on promastigote and intracellular amastigote forms of L. amazonensis. The results indicated no activity for the extract and its fractions, with an inhibitory concentration of 50% (IC50) exceeding 200 μg/mL, while isoeleutherin demonstrated activity (IC50 = 25 μg/mL). Additionally, it is important to note that the concentrations of the samples used in this study were not cytotoxic to macrophages (CC50 > 500 μg/mL), similar to the control drug (amphotericin B; CC50 > 100 μg/mL). The EEEp, its fractions, and isoeleutherin showed CC50 greater than 500 μg/mL and IC50 greater than 200 μg/mL against L. amazonensis, which is why the selectivity index calculation was not possible.
Then, macrophages infected with promastigote form were submitted to treatment with EEEp, FDEp, FAEEp, FMEp, and isoeleutherin (concentration of 500, 250 e 125 μg/mL). In Figure 2, amastigotes forms can be observed around the destroyed cells, exposed to EEEp (Figure 2A) and its fractions FDEp (Figure 2B), FAEEp (Figure 2C), FMEp (Figure 2D) and isoeleutherin (Figure 2E). The destruction of macrophages observed, likely caused by intracellular forms of L. amazonensis, since EEEp, its fractions, and isoeleutherin did not show cytotoxicity on macrophages. No reductions in the number of amastigotes were observed in infected cells when compared to the negative control (Figure 3).
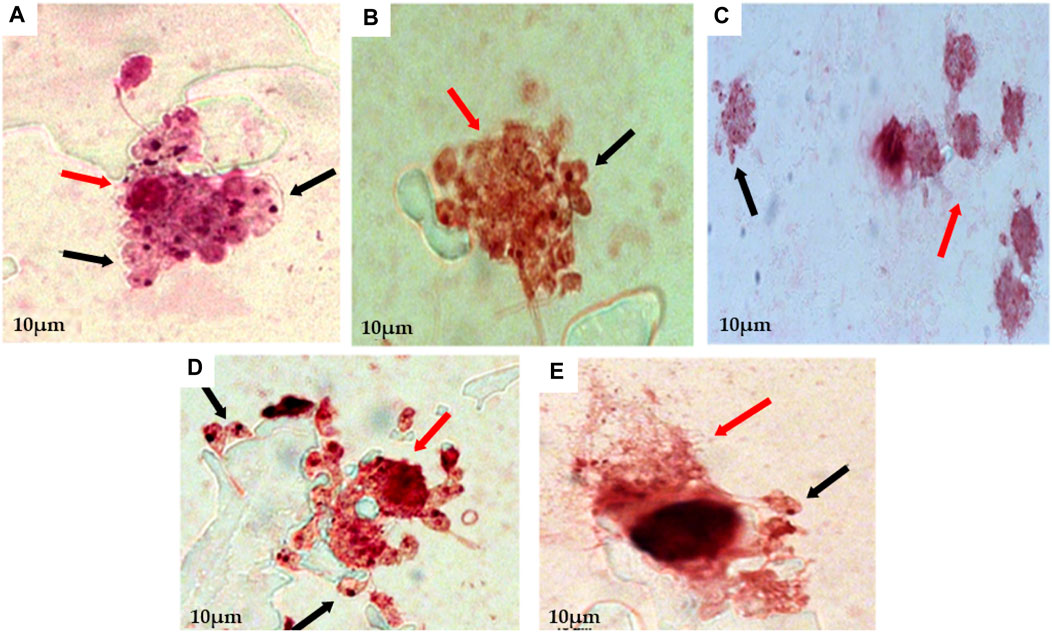
FIGURE 2. Assay for anti-amastigote activity, showing macrophages infected with amastigotes of L. amazonensis and then treated with 500 μg/mL of EEEp (A), FDEp (B), FAEp (C), FMEp (D) and Isoeleutherin (E). Red arrows indicate cell destruction (macrophages) and black arrows indicate the presence of amastigotes around the destroyed cell (×100 magnification).
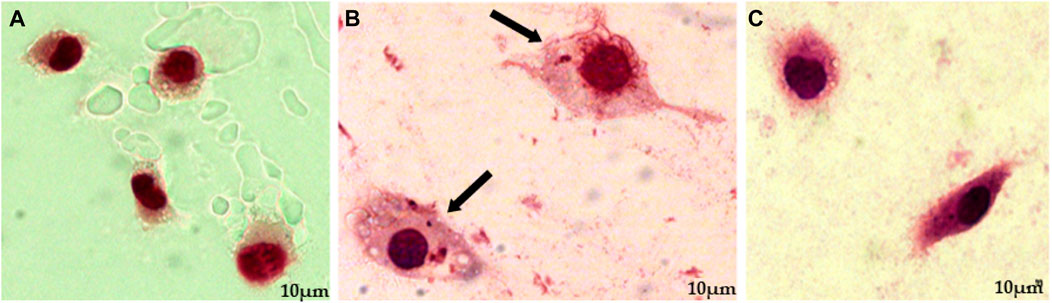
FIGURE 3. Anti-amastigote activity assay, showing uninfected macrophages, macrophages infected with L. amazonensis amastigotes, and subsequently treated with Amphotericin (B). Caption: (A). Uninfected macrophage control (×100 magnification); (B). Macrophage control infected with L. amazonensis amastigotes (Negative control; ×100 magnification); (C). Anti-amastigote action of Amphotericin B at a concentration of 100 μg/mL (Positive control; ×100 magnification). Arrows indicate the presence of amastigotes inside macrophages.
Amastigote forms can be observed around the destroyed cells exposed to treatment with EEEp, FDEp, FAEp, FMEp, and isoeleutherin, highlighting that fractionation did not enhance the activity against amastigote forms of L. amazonensis, as the fractions and isoeleutherin remained inactive. The large number of promastigote forms around the destroyed macrophages indicates that they were unable to invade the cells, considering that these forms can survive in unfavorable conditions. Some of them assumed a more rounded shape, resembling amastigotes (Figure 2).
3.2 Prediction studies of pharmacokinetic and physicochemical aspects of isoeleutherin analogues
Isoeleutherin (ISO; Figure 4) was employed as the starting molecule in the search for analogs through structural similarity in databases, with the aim of examining how structural modifications would impact physicochemical and pharmacokinetic properties. As a result of this process, 20 analogs (Figure 4) were identified and subsequently subjected to in silico prediction studies.
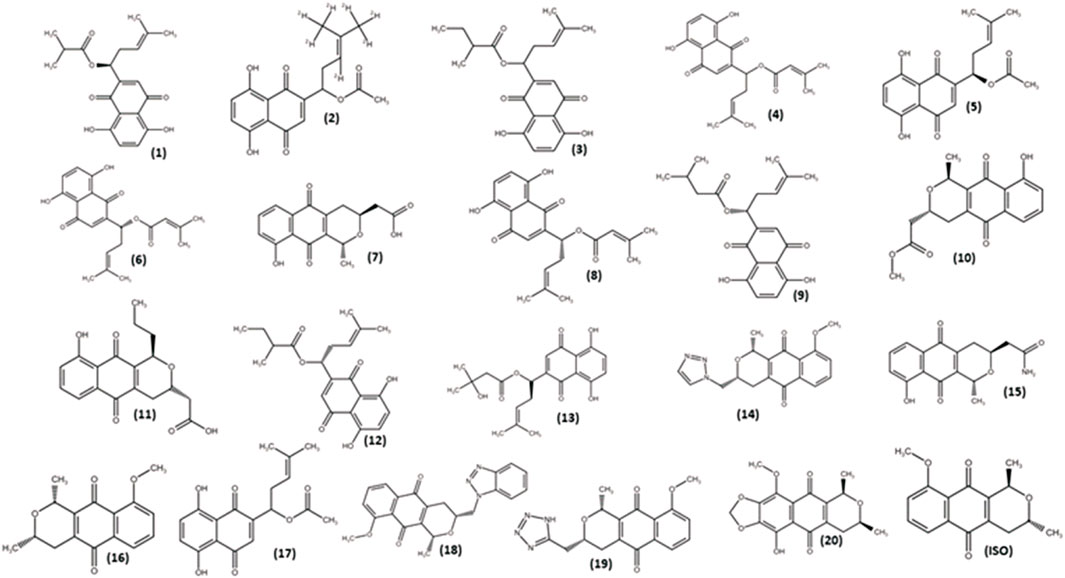
FIGURE 4. Isoeleutherin and its analogues.
Isoeleutherin and its 20 analogues did not violate Lipinski’s rule, showing octanol-water partition coefficient (miLog p ≤ 5); Molecular Mass (MM) ≤ 500 g/mol, number of hydrogen bond acceptor groups (nHAG) ≤ 10 and number of hydrogen bond donor groups (nHDG ≤5) (Lipinski et al., 1997). The values obtained for the topological polar surface area (TPSA) parameters ≤140Å2; number of rotational bonds (Nrotb) ≤ 10 and molecular volume are within the limits established by Veber’s descriptors (Table 1) (Veber et al., 2002).
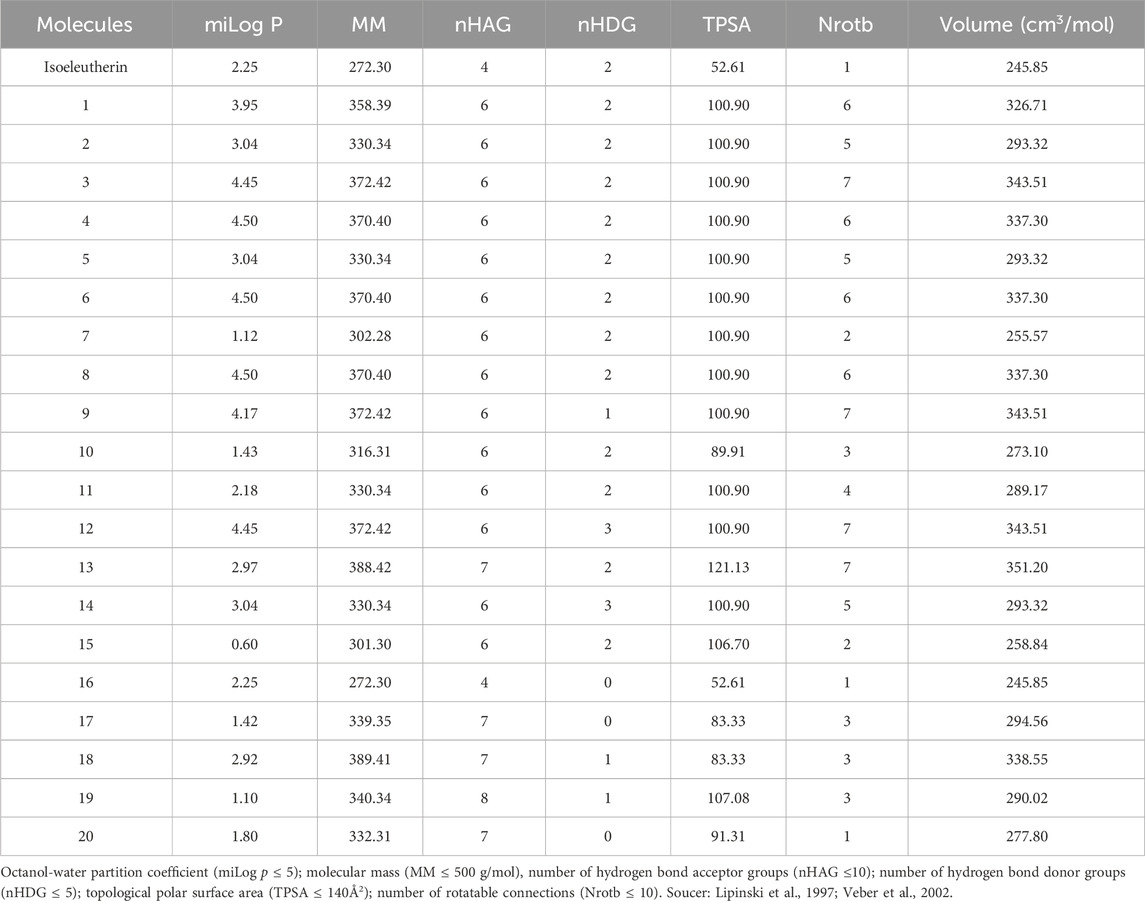
TABLE 1. Physicochemical aspects of isoeleutherin analogues.
The results suggest isoeleutherin and its analogues have moderate permeability in Caco2 and low to moderate in MDCK, whereas the permeability of isoeleutherin in MDCK is moderate. However, it is possible to observe that these compounds have high intestinal absorption (AIH). Isoeleutherine appears to bind to albumin moderately, with some structural alterations increasing the affinity for albumin. Only isoeleutherin and compound 18 freely crossed the BBB, structural alterations reduced the ability to cross the barrier. All compounds appear to be metabolized by CYP3A4, with variations in the magnitude of metabolism. Structural changes did not significantly impact the CYP inhibitory potential (Table 2).
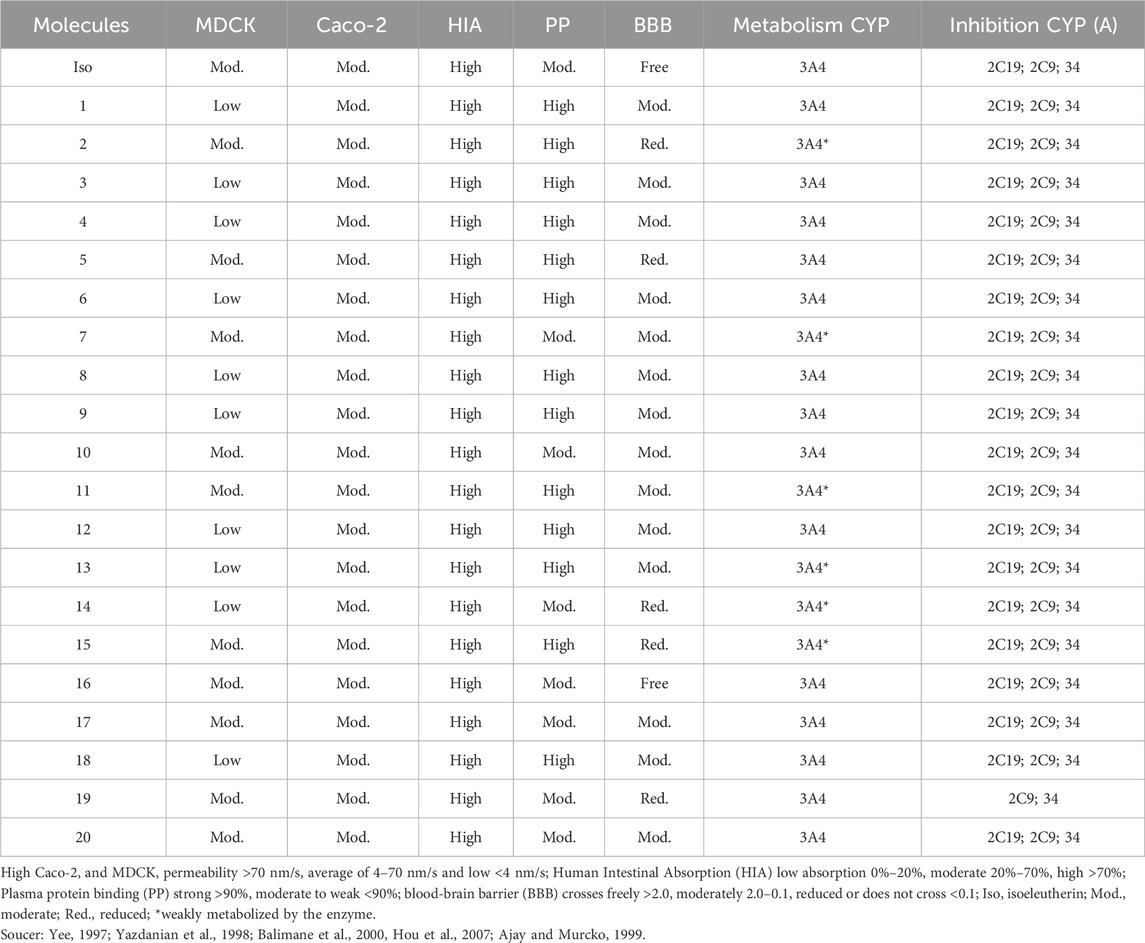
TABLE 2. Pharmacokinetic aspects of isoeleutherin analogues.
Therefore, it is important to evaluate the impact of structural alterations in different aspects, such as: physicochemical, pharmacokinetic and receptor binding. Among the isoeleutherin analogues, there were no significant alterations in physicochemical and pharmacokinetic aspects, except in distribution, where alterations in plasma protein binding and distribution to the CNS were observed.
3.4 Molecular docking and molecular dynamics
Molecular docking of the isoeleutherin and its 20 analogues were evaluated at the catalytic site of the TR enzyme (PDB 2JK6), performed at a distance of 10Å from the Flavin-Adenine Dinucleotide (FAD) cofactor, with RMSD values below 2Å, demonstrating that the redocking was successful according to literature data (Hevener et al., 2009). Connections that occurred in the active site of the enzyme were analyzed using the PoseView online server (Stierand et al., 2006), taking into account the interactions performed with residues Cys52, Cys57, His461’, and Glu466’, involved in the redox metabolism of Leishmania (Baiocco et al., 2009). The GoldScore scoring function was employed to predict the binding affinity of the most stable configuration, and the highest scores were considered to select the top 10 compounds with potential anti-leishmanial activity, as shown in Table 3.
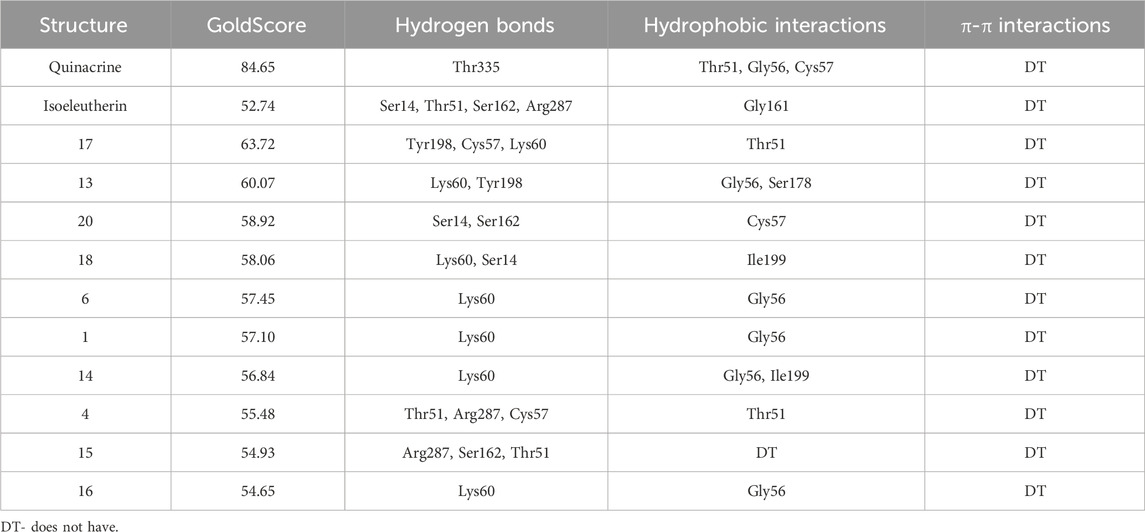
TABLE 3. GoldScore values, hydrogen bonds, hydrophobic and π-π interactions of isoeleutherin and analogous obtained through Poseview.
Quinacrine is an effective and widely used antiparasitic drug with potential adverse effect. In this experiment, it was employed as the control drug. Compound 17 (Zinc317780204) was selected among the isoeleutherin analogues, based on the results found in molecular docking, showing that the molecule’s conformation interacts with the enzyme, through hydrophobic interactions with the amino acid residues Thr51 and hydrogen bonds with the Tyr198, Cys57, Lys60 residue. Isoeleuterin presented conformations with the enzyme by hydrophobic interactions with the residue Gly161 and hydrogen bonds with residues Ser14, Thr51, Ser162, and Arg287 (Figure 5).
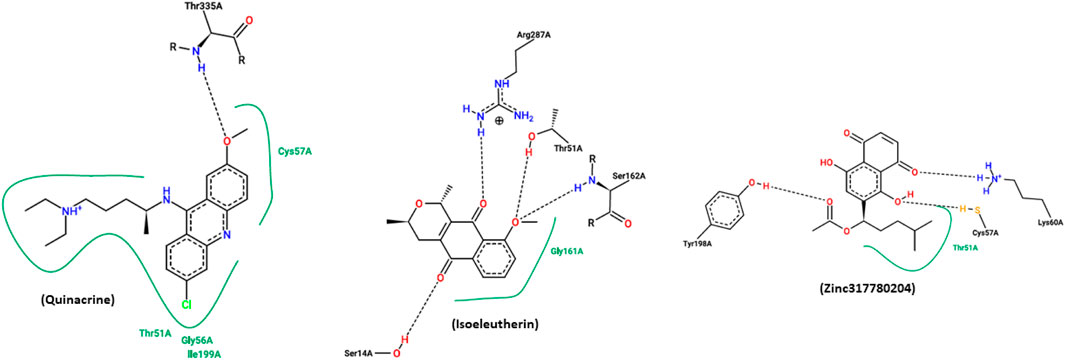
FIGURE 5. A figure generated by the online PoseView server, presenting the interactions resulting from docking simulations between the most stable configurations of quinacrine, isoeleutherin, and analogue compound 17 (Zinc317780204) with the enzyme TR. Hydrophobic interactions are indicated by continuous green lines, while hydrogen bonds are represented by dashed black lines.
The coordinates obtained from the molecular docking simulation procedures, representing the most stable configurations of quinacrine, isoeleutherin, and the analogue compound (Zinc317780204), were subjected to 50 ns of Molecular Dynamics (MD) to assess the structural stability of the enzyme-ligand complexes through the analysis of RMSD values over time, as depicted in Figure 6. After the process was initiated, the RMSD values of the TR-ligand complexes fluctuated (1–4 Å) until reaching equilibrium (10 ns) and then remained stable throughout the simulation, with average values of 2.50 and a standard deviation of ±0.31 for quinacrine, 2.70 Å and a standard deviation of ±0.39 for isoeleutherin, and 2.33 Å and a standard deviation of ±0.23 for Zinc317780204. This indicates that the compounds remained within the enzyme cavity, showing minimal conformational changes in the complex structure.
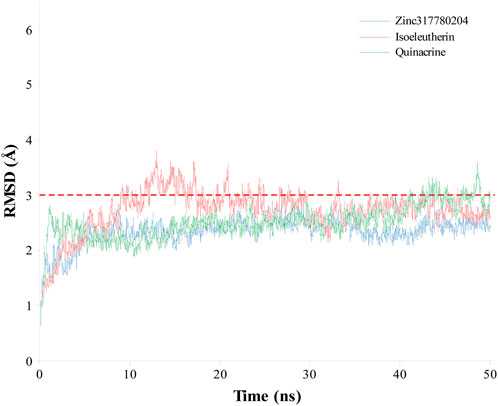
FIGURE 6. Graphical representation of RMSD (Å) values over time in the molecular dynamics simulation for isoeleutherin (red line), Zinc317780204 compound (blue line), and quinacrine (green line).
The flexibility of the protein regions in relation to each ligand was evaluated through the graph of B-factor (Figure 7). The greatest fluctuations occurred in regions corresponding to the bands of amino acid residues Asn91-Gly80 (highlighted in red), Glu410-Thr397 (highlighted in yellow) and Ser489 fragment region, highlighted in green, considered the most flexible regions of the enzyme. These residues are found close to the Cys52 and Cys57 active sites and play an important functional role in enzyme inhibition (Verma et al., 2012; Pandey et al., 2016).
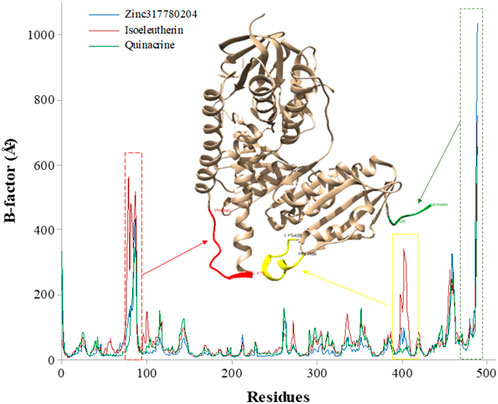
FIGURE 7. Graph of B-factor (Å) of the enzyme Trypanothione Reductase with the amino acid residues during the MD simulation time (50 ns): isoeleutherin (red line), Zinc317780204 compound (blue line), and quinacrine (green line).
Comparing the binding free energy values of the two methods, described in Table 4, both are favorable and stable for complex formation. The calculation of binding free energy allowed for quantifying the affinity between the enzyme-ligand system. When comparing the free energy values obtained through the MM-GB(PB)SA method, it was observed that quinacrine, used as the reference drug in the present study, displayed the lowest value. The compound Zinc317780204 followed, with values lower than those of isoeleutherin, indicating that these compounds may be useful in situations where the reference drug has limitations.
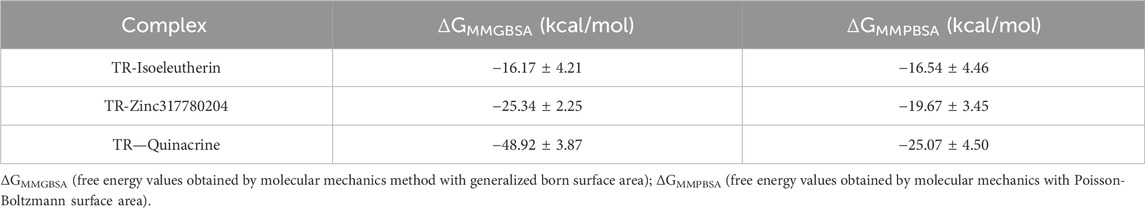
TABLE 4. Binding free energies with standard deviation calculated by the MM-GB(PB)SA method.
Energy decomposition by residue was employed to identify the TR enzyme residues involved in interactions with Zinc317780204, isoeleutherin, and quinacrine, based on the energy contribution calculated by the MMGBSA method for the last 10 ns of MD simulations. Free binding energy values below −1 kcal/mol were set as the criteria for selecting amino acid residues that participated in the most favorable interactions, contributing to ligand stabilization within the complex.
The analysis of residue decomposition values reveals those that played a significant role in the total interaction energy and stabilization of the TR-Zinc317780204 (Cys52, Tyr198, Val332, and Thr335), TR-isoeleutherin (Cys52, 57, and Thr335), and TR-Quinacrine (Cys52, Tyr51, Cys57, Arg287, Asp327, Met333, and Ala338) complexes, contributing significantly to the overall free energy of the complex (Figure 8).
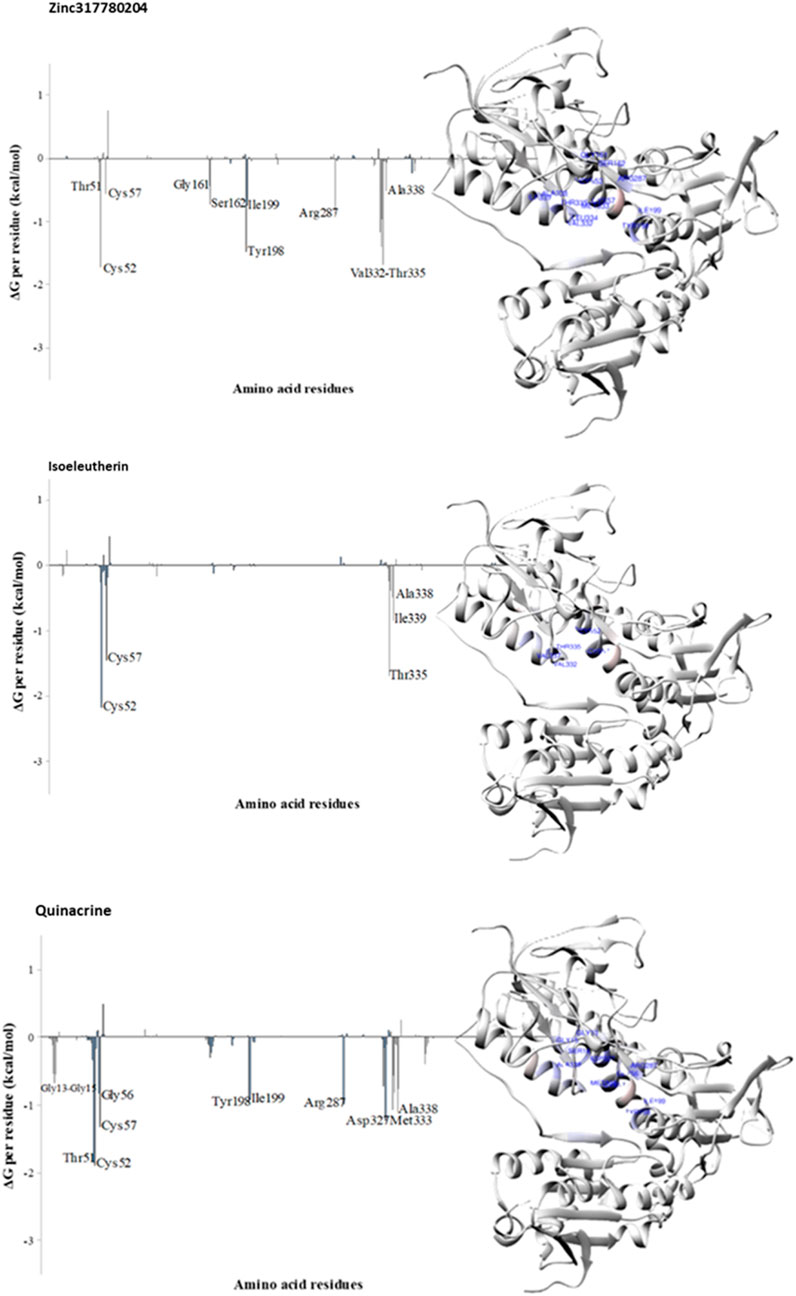
FIGURE 8. Decomposition of free binding energy per residue (in kcal/mol) obtained with the MMBGSA method for TR-Zinc317780204, TR-Isoeleutherin and TR-Quinacrine. On the right, residues that most contributed to the binding free energy, visualized by CHEWD plugin in the Chimera program, are shown in blue.
4 Discussion
The identified chemical constituents of E. plicata bulbs, involve several classes of phy-tochemicals, condensed tannins, coumarins, steroids, triterpenoids, anthraquinones and naphthoquinones, being isolated, eleutherol, eleutherin and their respective isomers, isoeleutherol and isoeleutherin, indicative of being the major constituents and markers of the species (Malheiros et al., 2015; Vale et al., 2020).
One study evaluated the EEEp, PDEp and isoeleutherin that were active in another parasite, Plasmodium falciparum sensitive to chloroquine (Vale et al., 2020), reinforcing the premise that the antiparasitic activity of E. plicata is related to naphthoquinone isoeleutherin. However, it is important to investigate the possible mechanisms involved in these antiparasitic activities.
Regarding the physicochemical properties, all compounds met the criteria of Lipinski’s rule of five, The octanol-water partition coefficient allows to determine the degree of hydrophobicity of the molecules and directly influences the absorption and bioavailability of drugs, substances that present (miLog p ≤ 5), are more soluble in organic medium, in this study, all molecules present miLogP within the established parameter, demonstrating that they have a more polar character, capable of dissolving in aqueous and crossing cellular barriers (Lipinski et al., 1997). The compounds had a molecular weight >500 Da, and this may be directly related to the permeability of the substances, since the higher the molecular weight, the more difficult it will be for the compound to permeate biological membranes (Lipinski et al., 1997). The topological polar surface area (TPSA) of each molecule was less than 140 Å, which is justified by the lower bond between hydrogen acceptors and donors, respectively ≤10 and ≤5. Thus, isoeleuterin would be the most promising compound, due to its lower molecular weight and better physicochemical profile for permeability between cell membranes.
In the pharmacokinetic prediction, due to the physicochemical characteristics of the polar compounds, the permeability in MDCK cells (Madin-Darby canine kidney) was low to moderate, when evaluating the permeability in MDCK cells, the active permeability rate of each molecule was analyzed. All molecules have moderate permeability to Caco-2 cells (human colon carcinoma epithelial cells), suggesting that the rate of intestinal absorption is moderate through the passive diffusion mechanism (Chen et al., 2018; Panse and Gerk, 2022). However, isoeleutherin and its studied analogues have been shown to have high absorption in the small intestine through human intestinal absorption analysis. Regarding the permeability of the blood-brain barrier, only isoeleutherin and compound 18 freely cross the blood-brain barrier, probably due to their lower molecular weight and better plasma distribution, due to their lower binding to plasma proteins compared to the other compounds that showed high binding to plasma proteins, so isoeleutherin has a better pharmacokinetic profile of absorption and distribution.
In the present study, the involvement of RT in the leishmanicidal activity of isoeleutherin and analogues was evaluated. TR is an NADPH-dependent flavoprotein disulfide reductase, found in several parasites, including Leishmania, which participates in its redox system, being a substitute for the glutathione/glutathione reductase and thioredoxin/thioredoxin reductase systems (Baiocco et al., 2009; Fairlamb and Cerami, 1992]. Naphthoquinones, in the presence of oxygen, are reduced and reoxidized, generating reactive oxygen species that affect the capacity of TR, which is why this target was selected (Ferreira, 2008).
In Leishmania spp. and trypanosomatids, the redox balance is carried out by the enzyme TR, which functions as a FAD-dependent disulfide oxidoreductase that catalyzes the reduction of trypanothione [N1, N8-bis-glutathionylspermidine or T (SH)2] as a function of NADPH (Leroux and Krauth-Siegel, 2016). Trypanothione is a dithiol formed by two glutathione molecules linked by a spermidine bridge, it acts as an electron donor in biological reactions, including the elimination of hydroperoxides, being the main thiol in trypanosomatids, assuming the functions of glutathione in other organisms. The return of trypanothione to its reduced active form is carried out by TR, an essential process for the redox balance of the parasite and cell viability, demonstrating the essential character of TR (Tovar et al., 1998). The closest mammalian homologue of TR is the enzyme glutathione reductase (GR), despite their general similarity, TR and GR differ significantly in their thiol binding sites, making it feasible to target TR with chemical entities that do not compromise GR activity (Castro et al., 2018). Over-all, the essential role played by TR in Leishmania spp. and its absence in the human host, makes this molecule an attractive target for the development of potential new drugs.
In this work, the leishmanicidal activity was demonstrated, against promastigotes forms of L. amazonensis, where FDEp showed moderate activity and isoeleutherin was more promise. We also observed the presence of parasites around infected macrophages, requiring further investigation, whether there was interference in the mechanism of phagocytosis or destruction of these cells. Everything indicates that derivatives of E. plicata, especially isoeleutherin, have activity on L. amazonensis.
The synthetic naphthoquinone, LQB-118 caused a concentration-dependent reduction in the number of amastigotes of L. amazonensis (Cunha-Junior et al., 2011). The hydroxynaphthoquinone Buparvaquone showed excellent in vitro activity against amastigotes of L. donovani (Effective dose 50%—ED50 between 0.12 and 0.005 µM) (Croft et al., 1992). The dimeric naphthoquinones 3,3-Bijuglone and 6,6-Dibenzyloxy-3,3-bi-plumbagin, exhibited high toxicity for amastigotes of L. donovani (IC50 of 15 and 14.2 μg/mL, respectively) (Kaiser et al., 2000).
It is known that structural alterations can interfere with the biological activities of quinones, and eleutherin, isoeleutherin and eleutherol presented different immune responses mediated by helper T-cells (Castro et al., 2021b). Isoeleuterin has a 1,4-naphthoquinone ring with an α-methyl group, selectively stimulating IFNc production by activating transcription of the T-bet gene, thus enhancing Th1-mediated immune responses. While, naphthopy-ran-4-one, eleutherinol, inhibited the production of IFNc and IL-2 during the activation of Th cells, suppressing the transcripts of the cytokine gene. Therefore, chemical modification and chirality of the naphthopyran moiety in isoeleutherin and eleutherinol may be critical for the selective modulation of immune responses mediated by helper T-cells (Hong et al., 2008).
In this study, using peritoneal macrophages from BALB/c mice, the cytotoxic activity of EEEp, its fractions (FDEp, FAEp and FMEp) and isoeleutherin were evaluated, with no cytotoxicity being observed. Gomes et al. (2021), evaluated the cytotoxicity of EEEp, FDMEp and isoeleutherin from E. plicata in human hepatoma cells (HepG2), after exposure for 24 h, and observed that fractionation reduced cytotoxicity, with isoeleutherin being the least toxic sample. These results suggest the cytotoxicity of EEEp and FDMEp are related to the synergism between eleutherin and isoeleutherin compounds. When administered together they are more active, which may increase toxicity (Vale et al., 2020; Gomes et al., 2021).
Another study using an integrative approach of in vitro and in silico methodologies, found isoeleutherin was the compound with lowest cytotoxicity in the micronucleus assay and structural changes. Also, there was an increase in the number of important interactions with amino acid residues of the active site, suggesting increased affinity with the Topoisomerase II enzyme, representing a good starting point in the search for new drugs for anticancer therapy (Albuquerque et al., 2023).
5 Conclusion
In summary, structural alterations made to isoeleutherin resulted in obtaining a less toxic analogue compound (CP13), as well as obtaining a more promising molecule as a leishmanicidal agent (CP17). Thus, naphthoquinones may represent an important class of molecules for discovering new leishmanicidal therapies.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statement
Approval was obtained from the Ethics Council regarding the use of macrophages, which were retrieved from murine rats. The experimental protocol was submitted to the Ethics and Research Committee on the Use of Animals of the Evandro Chagas Institute (CEUA) and approved by it with approval opinion no.: 0022/2011/CEPAN/IEC/SVS/MS.
Author contributions
KA: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing–original draft, Writing–review and editing. AV: Data curation, Formal Analysis, Investigation, Methodology, Writing–original draft, Writing–review and editing. FS: Formal Analysis, Investigation, Methodology, Writing–review and editing. MC: Data curation, Formal Analysis, Investigation, Writing–review and editing. AC: Formal Analysis, Investigation, Methodology, Writing–review and editing. AB: Formal Analysis, Investigation, Methodology, Writing–review and editing. PM: Formal Analysis, Investigation, Methodology, Writing–review and editing. SP: Data curation, Investigation, Methodology, Writing–review and editing. FM: Writing–review and editing. MD: Conceptualization, Supervision, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The authors acknowledge the financial support of the Universal CNPQ project through the process 432458/2018.2. PROPESP/UFPA support the publication of this article.
Acknowledgments
The authors thank the Dra Márlia Regina Coelho-Ferreira of the Emílio Goeldi Paraense Museum, Brazil for the botanical identification of the species.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ajay, B. G. W., and Murcko, M. A. (1999). Designing libraries with CNS activity. J. Med. Chemmistry 42 (24), 4942–4951. doi:10.1021/jm990017w
Albuquerque, K. C. D.O., Galucio, N. C. D. R., Ferreira, G. G., Quaresma, A. C. S., Vale, V. V., Bahia, M. D. O., et al. (2023). Study of genotoxicity, activities on caspase 8 and on the stabilization of the Topoisomerase complex of isoeleutherin and analogues. Molecules 28 (4), 1630–1712. doi:10.3390/molecules28041630
Almeida, R. G., Valença, W. O., Rosa, L. G., de Simone, C. A., de Castro, S. L., Barbosa, J. M. C., et al. (2020). Synthesis of quinone imine and sulphur-containing compounds with antitumor and trypanocidal activities: redox and biological implications. RSC Quím. Med. 11 (10), 1145–1160. doi:10.1039/d0md00072h
Alves, K. M. A., Cardoso, F. J. B., Honorio, K. M., and Molfetta, F. A. (2020). Design of inhibitors for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) enzyme of Leishmania Mexicana. Med. Chem. 16 (6), 784–795. doi:10.2174/1573406415666190712111139
Baiocco, P., Colotti, G., Franceschini, S., and Ilari, A. (2009). Molecular basis of antimony treatment in leishmaniasis. J. Med. Chem. 52 (8), 2603–2612. doi:10.1021/jm900185q
Balimane, P. V., Chong, S., and Morrison, R. A. (2000). Current methodologies used for evaluation of intestinal permeability and absorption. J. Pharmacol. Toxicol. Methods 44 (1), 301–312. doi:10.1016/S1056-8719(00)00113-1
Bayly, C. I., Cieplak, P., Cornell, W., and Kollman, P. A. (1993). A well-behaved electrostatic potential based method using charge restraints for deriving atomic charges - the RESP model. J. Phys. Chem. 97 (40), 10269–10280. doi:10.1021/j100142a004
Borges, E. S., Galucio, N. C. D. R., Veiga, A. S. S., Busman, D. V., Lins, A. L. F. A., Bahia, M. O., et al. (2020). Botanical studies, antimicrobial activity and cytotoxity of Eleutherine bulbosa (Mill). Urb. Res. Soc. Dev. 9, 1–21. doi:10.33448/rsd-v9i11.9992
Case, D. A., Cheatham, T. E., Darden, T., Gohlke, H., Luo, R., Merz, K. M., et al. (2005). The Amber biomolecular simulation programs. J. Comput. Chem. 26 (16), 1668–1688. doi:10.1002/jcc.20290
Castro, A. L. G., Correa-Barbosa, J., De Campos, P. S., Matte, B. F., Lamers, M. L., and Siqueira, J. E. S. (2021b). Antitumoral activity of Eleutherine plicata Herb. and its compounds. Int. J. Dev. Res. 11 (2), 44673–44678. doi:10.37118/ijdr.21154.02.2021
Castro, A. L. G., Cruz, J. N., Sodré, D. F., Correa-Barbosa, J., Azonsivo, R., De Oliveira, M. S., et al. (2021a). Evaluation of the genotoxicity and mutagenicity of isoeleutherin and eleutherin isolated from Eleutherine plicata Herb. using bioassays and in silico approaches. Arabian J. Chem. 14 (4), 103084–103111. doi:10.1016/j.arabjc.2021.103084
Castro, H., Duarte, M., and Tomás, A. M. (2018). “The redox metabolism and oxidative stress in leishmania as a crossroads for the lethal effect of drugs,” in Drug discovery for leishmaniasis (Croydon, England: The Royal Society of Chemistry), 316–347. doi:10.1039/9781788010177-00316
Chen, E. C., Broccatelli, F., Plise, E., Chen, B., Liu, L., Cheong, J., et al. (2018). Evaluating the utility of canine Mdr1 knockout Madin-Darby canine kidney I cells in permeability screening and efflux substrate determination. Mol. Pharm. 15 (11), 5103–5113. doi:10.1021/acs.molpharmaceut.8b00688
Couto, C. L. L., Moraes, D. F. C., Cartágenes, M. d. S. S., Amaral, F. M. M. d., and Guerra, R. N. (2016). Eleutherine bulbousa (Mill.) Urb.: a review study. J. Med. Plants Res. 10, 286–297. doi:10.5897/JMPR2016.6106
Croft, S. L., Hogg, J., Gutteridge, W. E., Hudson, A. T., and Randall, A. W. (1992). The activity of hydroxynaphthoquinones against Leishmania donovani. J. Antimicrob. Chemother. 30 (6), 827–832. doi:10.1093/jac/30.6.827
Cunha-Junior, E. F., Pacienza-Lima, W., Ribeiro, G. A., Daher Neto, C., Canto-Cavalheiro, M. M., Silva, A. J. M., et al. (2011). Effectiveness of the local or oral delivery of the novel naphthopterocarpanquinone LQB-118 against cutaneous leishmaniasis. J. Antimicrob. Chemother. 66 (7), 1555–1559. doi:10.1093/jac/dkr158
Darden, T., York, D., and Pedersen, L. (1993). Particle mesh Ewald: an N-log(N) method for Ewald sums in large systems. J. Chem. Phys. 98 (12), 10089–10092. doi:10.1063/1.464397
Fairlamb, A. H., and Cerami, A. (1992). Metabolism and functions of trypanothione in the Kinetoplastida. Annu. Rev. Microbiol. 46, 695–729. doi:10.1146/annurev.mi.46.100192.003403
Ferreira, J. G. (2008). Study of quinone compounds with activity against Chagas disease. These (PhD in Physical Chemistry). São Carlos: University of São Paulo. doi:10.11606/T.75.2008.tde-23062008-163355
Fock, V. (1930). Nӓherungsmethode zur Lӧsung des quantenmechanischen Mehrkӧrperproblems. Z. für Phys. 61 (1-2), 126–148. doi:10.1007/BF01340294
Frisch, M. J., Trucks, G. W., Schlegel, H. B., Scuseria, G. E., Robb, M. A., Cheeseman, J. R., et al. (2003). Gaussian 03, in (version 2003). Wallingford, CT: Gaussian.
Gomes, A. R. Q., Galucio, N. C. D. R., Albuquerque, K. C. O., Brígido, H. P. C., Varela, E. L. P., Castro, A. L. G., et al. (2021). Toxicity evaluation of Eleutherine plicata Herb. extracts and possible cell death mechanism. Toxicol. Rep. 8, 1480–1487. doi:10.1016/j.toxrep.2021.07.015
Hariharan, P. C., and Pople, J. A. (1973). The influence of polarization functions on molecular orbital hydrogenation energies. Theor. Chim. Acta 28 (3), 213–222. doi:10.1007/BF00533485
Hevener, K. E., Zhao, W., Ball, D. M., Babaoglu, K., Qi, J., White, S. W., et al. (2009). Validation of molecular docking programs for virtual screening against dihydropteroate synthase. J. Chem. Inf. Model. 49 (2), 444–460. doi:10.1021/ci800293n
Hong, J. H., Yu, E. S., Han, A. R., Nam, J. W., Seo, E. K., and Hwang, E. S. (2008). Isoeleutherin and eleutherinol, naturally occurring selective modulators of Th cell-mediated immune responses. Biochem. Biophysical Res. Commun. 371 (2), 278–282. doi:10.1016/j.bbrc.2008.04.060
Hornak, V., Abel, R., Okur, A., Strockbine, B., Roitberg, A., and Simmerling, C. (2006). Comparison of multiple Amber force fields and development of improved protein backbone parameters. Proteins 65 (3), 712–725. doi:10.1002/prot.21123
Hou, T., Wang, J., Zhang, W., and Xu, X. (2007). ADME evaluation in drug discovery. 7. Prediction of oral absorption by correlation and classification. J. Chem. Inf. Model. 47 (1), 208–218. doi:10.1021/ci600343x
Irwin, J. J., Sterling, T., Mysinger, M. M., Bolstad, E. S., and Coleman, R. G. (2012). Zinc: a free tool to discover chemistry for biology. J. Chem. Inf. Model. 52 (7), 1757–1768. doi:10.1021/ci3001277
Jones, G., Willett, P., and Glen, R. C. (1995). Molecular recognition of receptor sites using a genetic algorithm with a description of desolvation. J. Mol. Biol. 245 (1), 43–53. doi:10.1016/S0022-2836(95)80037-9
Jorgensen, W. L., Chandrasekhar, J., Madura, J. D., Impey, R. W., and Klein, M. L. (1983). Comparison of simple potential functions for simulating liquid water. J. Phys. Chem. 79 (2), 926–935. doi:10.1063/1.445869
Kaiser, O., Kiderlen, A. F., Laatsch, H., and Croft, S. L. (2000). In vitro leishmanicidal activity of monomeric and dimeric naphthoquinones. Acta Trop. 77 (3), 307–314. doi:10.1016/s0001-706x(00)00161-3
Kollman, P. A., Massova, I., Reyes, C., Kuhn, B., Huo, S., Chong, L., et al. (2000). Calculating structures and free energies of complex molecules: combining molecular mechanics and continuum models. Accounts Chem. Res. 33 (12), 889–897. doi:10.1021/ar000033j
Lang, P. T., Brozell, S. R., Mukherjee, S., Pettersen, E. F., Meng, E. C., Thomas, V., et al. (2009). DOCK 6: combining techniques to model RNA-small molecule complexes. RNA Soc. 15 (6), 1219–1230. doi:10.1261/rna.1563609
Leroux, A. E., and Krauth-Siegel, R. L. (2016). Thiol redox biology of trypanosomatids and potential targets for chemotherapy. Mol. Biochem. Parasitol. 206, 67–74. doi:10.1016/j.molbiopara.2015.11.003
Li, P., Roberts, B. P., Chakravorty, D. K., and Merz, K. M. (2013). Rational design of particle mesh Ewald compatible Lennard-Jones parameters for +2 metal cations in explicit solvent. J. Chem. Theory Comput. 9 (6), 2733–2748. doi:10.1021/ct400146w
Lipinski, C. A., Lombardo, F., Dominy, B. W., and Feeney, P. J. (1997). Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 23 (1-3), 3–25. doi:10.1016/S0169-409X(96)00423-1
Lipinski, C. A., Lombardo, F., Dominy, B. W., and Feeney, P. J. (2001). Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings 1PII of original article: S0169-409X(96)00423-1. The article was originally published in Advanced Drug Delivery Reviews 23 (1997) 3–25. 1. Adv. Drug Deliv. Rev. 46 (1-3), 3–26. doi:10.1016/S0169-409X(00)00129-0
Loncharich, R. J., Brooks, B. R., and Pastor, R. W. (1992). Langevin dynamics of peptides: the frictional dependence of isomerization rates of N-acetylalanyl-N’-methylamide. Biopolymers 32 (5), 523–535. doi:10.1002/bip.360320508
Malheiros, L. C. S., Mello, J. C. P., and Barbosa, W. L. R. (2015). “Eleutherine plicata – quinones and antioxidante activity,” in Phytochemicals – isolation, characterisation and role in human Health. Editors A. V. Rao, and L. G. Rao (Croacia: Intechopen), 323–338. doi:10.5772/59865
Mann, S., Frasca, K., Scherrer, S., Henao-Martínez, A. F., Newman, S., Ramanan, P., et al. (2021). A review of leishmaniasis: current Knowledge and future directions. Curr. Trop. Med. Rep. 8, 121–132. doi:10.1007/s40475-021-00232-7
Massova, I., and Kollman, P. A. (2000). Combined molecular mechanical and continuum solvent approach (MM-PBSA/GBSA) to predict ligand binding. Perspect. Drug Discov. Des. 18 (1), 113–135. doi:10.1023/A:1008763014207
Meng, X. Y., Zhang, H. X., Mezei, M., and Cui, M. (2011). Molecular docking: a powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 7 (2), 146–157. doi:10.2174/157340911795677602
Miller, B. R., Mcgee, T. D., Swails, J. M., Homeyer, N., Gohlke, H., and Roitberg, A. E. (2012). MMPBSA.py: an efficient program for end-state free energy calculations. J. Chem. Theory Comput. 8 (9), 3314–3321. doi:10.1021/ct300418h
Molinspiration (2003). Molinspiration. Available at: https://www.molinspiration.com.
Mota, T. C., Cardoso, P. C. S., Gomes, L. M., Vieira, P. C. M., Corrêa, R. M. S., Santana, P. D. P. B., et al. (2011). In vitro evaluation of the genotoxic and cytotoxic effects of artesunate, an antimalarial drug, in human lymphocytes. Environ. Mol. Mutagen. 52 (7), 590–594. doi:10.1002/em.20659
Mukherjee, D., Yousuf, M., Dey, S., Chakraborty, S., Chaudhuri, A., Kumar, V., et al. (2020). Targeting the trypanothione reductase of tissue-residing Leishmania in hosts’ reticuloendothelial system: a flexible water-soluble ferrocenylquinoline-based preclinical drug candidate. J. Med. Chem. 63 (24), 15621–15638. doi:10.1021/acs.jmedchem.0c00690
Mukhopadhyay, R., Dey, S., Xu, N., Gage, D., Lightbody, J., Ouellette, M., et al. (1996). Trypanothione overproduction and resistance to antimonials and arsenicals in Leishmania. Proc. Natl. Acad. Sci. 93 (19), 10383–10387. doi:10.1073/pnas.93.19.10383
Neudert, G., and Klebe, G. (2011). Fconv: format conversion, manipulation and feature computation of molecular data. Bioinformatics 27 (7), 1021–1022. doi:10.1093/bioinformatics/btr055
Ngure, P. K., Tonui, W. K., Ingonga, J., Mutai, C., Kigondu, E., Ng’ang’a, Z., et al. (2009). In vitro antileishmanial activity of extracts of Warburgia ugandensis (Canellaceae), a Kenyan medicinal plant. J. Med. Plants Res. 3 (2), 61–66. doi:10.5897/JMPR.9000739
Olsson, M. H. M., Søndergaard, C. R., Rostkowski, M., and Jensen, J. H. (2011). PROPKA3: consistent treatment of internal and surface residues in empirical pKa predictions. J. Chem. Theory Comput. 7 (2), 525–537. doi:10.1021/ct100578z
Onufriev, A., Bashford, D., and Case, D. A. (2000). Modification of the generalized Born model suitable for macromolecules. J. Phys. Chem. B 104 (15), 3712–3720. doi:10.1021/jp994072s
Organização Pan-Americana da Saúde (OPAS) (2020). Leishmaniases: epidemiological report of the Americas. Washington, D.C.: Organização Pan-Americana da Saúde. Avaialable at: https://iris.paho.org/handle/10665.2/53091 (Accessed March 18, 2023).
Pandey, R. K., Kumbhar, B. V., Srivastava, S., Malik, R., Sundar, S., Kunwar, A., et al. (2017a). Febrifugine analogues as Leishmania donovani trypanothione reductase inhibitors: binding energy analysis assisted by molecular docking, ADMET and molecular dynamics simulation. J. Biomol. Struct. Dyn. 35 (1), 141–158. doi:10.1080/07391102.2015.1135298
Pandey, R. K., Narula, A., Naskar, M., Srivastava, S., Verma, P., Malik, R., et al. (2017b). Exploring dual inhibitory role of febrifugine analogues against Plasmodium utilizing structure-based virtual screening and molecular dynamic simulation. J. Biomol. Struct. Dyn. 35 (4), 791–804. doi:10.1080/07391102.2016.1161560
Pandey, R. K., Verma, P., Sharma, D., Bhatt, T. K., Sundar, S., and Prajapati, V. K. (2016). High-throughput virtual screening and quantum mechanics approach to develop imipramine analogues as leads against trypanothione reductase of leishmania. Biomed. Pharmacother. 83, 141–152. doi:10.1016/j.biopha.2016.06.010
Panse, N., and Gerk, P. M. (2022). The Caco-2 Model: modifications and enhancements to improve efficiency and predictive performance. Int. J. Pharm. 624, 122004. doi:10.1016/j.ijpharm.2022.122004
Paramapojn, S., Ganzera, M., Gritsanapan, W., and Stuppner, H. (2008). Analysis of naphthoquinone derivatives in the Asian medicinal plant Eleutherine americana by RP-HPLC and LC–MS. J. Pharm. Biomed. Analysis 47 (4-5), 990–993. doi:10.1016/j.jpba.2008.04.005
Pettersen, E. F., Goddard, T. D., Huang, C. C., Couch, G. S., Greenblatt, D. M., Meng, E. C., et al. (2004). UCSF Chimera - a visualization system for exploratory research and analysis. J. Comput. Chem. 25 (13), 1605–1612. doi:10.1002/jcc.20084
Ponte-Sucre, A., Gamarro, F., Dujardin, J. C., Barrett, M. P., López-Vélez, R., García-Hernández, R., et al. (2017). Drug resistance and treatment failure in leishmaniasis: a 21st century challenge. PLoS Neglected Trop. Dis. 11, e0006052. doi:10.1371/journal.pntd.0006052
Raza, S., Ranaghan, K. E., Van Der Kamp, M. W., Woods, C. J., Mulholland, A. J., and Azam, S. S. (2019). Visualizing protein-ligand binding with chemical energy-wise decomposition (CHEWD): application to ligand binding in the kallikrein-8 S1 Site. J. Computer-Aided Mol. Des. 33 (5), 461–475. doi:10.1007/s10822-019-00200-4
Roe, D. R., and Cheatham, T. E. (2013). PTRAJ and CPPTRAJ: software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 9, 3084–3095. doi:10.1021/ct400341p
Ryckaert, J. P., Ciccotti, G., and Berendsen, H. J. C. (1977). Numerical integration of the cartesian equations of motion of a system with constraints: molecular dynamics of n-alkanes. J. Comput. Phys. 23 (3), 327–341. doi:10.1016/0021-9991(77)90098-5
Saravanamuthu, A., Vickers, T. J., Bond, C. S., Peterson, M. R., Hunter, W. N., and Fairlamb, A. H. (2004). Two interacting binding sites for quinacrine derivatives in the active site of trypanothione reductase: a template for drug design. J. Biol. Chem. 279 (28), 29493–29500. doi:10.1074/jbc.M403187200
Silva, B. J. M., Hage, A. A. P., Silva, E. O., and Rodrigues, A. P. D. (2018). Medicinal plants from the Brazilian Amazonian region and their antileishmanial activity: a review. J. Integr. Med. 16 (4), 211–222. doi:10.1016/j.joim.2018.04.004
Silva, F. d.O. (2005). Avaliação in vitro da azitromicina nas espécies Leishmania (Leishmania) amazonensis, Leishmania (Viannia) braziliensis e Leishmania (Leishmania) chagasi. Theses (Master of Science) - Centro de Pesquisas René Rachou, Fundação Oswaldo Cruz. Belo Horizonte: Brazilian Institute of Information in Science and Technology.
Silva-Júnior, E. N. d., Jardim, G. A. M., Jacob, C., Dhawa, U., Ackermann, L., and Castro, S. L. d. (2019). Synthesis of quinones with highlighted biological applications: a critical update on the strategies towards bioactive compounds with emphasis on lapachones. Eur. J. Med. Chem. 179, 863–915. doi:10.1016/j.ejmech.2019.06.056
Stierand, K., Maaß, P. C., and Rarey, M. (2006). Molecular complexes at a glance: automated generation of two-dimensional complex diagrams. Bioinformatics 22 (14), 1710–1716. doi:10.1093/bioinformatics/btl150
Stierand, K., and Rarey, M. (2007). From modeling to medicinal chemistry: automatic generation of two-dimensional complex diagrams. ChemMedChem 2 (6), 853–860. doi:10.1002/cmdc.200700010
Sun, H., Li, Y., Shen, M., Tian, S., Xu, L., Pan, P., et al. (2014). Assessing the performance of MM/PBSA and MM/GBSA methods. 5. Improved docking performance using high solute dielectric constant MM/GBSA and MM/PBSA rescoring. Phys. Chem. Chem. Phys. 16, 22035–22045. doi:10.1039/c4cp03179b
Sun, L., Yang, H., Li, J., Wang, T., Li, W., Liu, G., et al. (2018). In silico prediction of compounds binding to human plasma proteins by QSAR models. ChemMedChem 13 (6), 572–581. doi:10.1002/cmdc.201700582
Tovar, J., Cunningham, M. L., Smith, A. C., Croft, S. L., and Fairlamb, A. H. (1998). Downregulation of Leishmania donovani trypanothione reductase by heterologous expression of a trans-dominant mutant homologue: effect on parasite intracellular survival. Proc. Natl. Acad. Sci. U. S. A. 95 (9), 5311–5316. doi:10.1073/pnas.95.9.5311
Vale, V. V., Cruz, J. N., Viana, G. M. R., Póvoa, M. M., Brasil, D. S. B., and Dolabela, M. F. (2020). Naphthoquinones isolated from Eleutherine plicata Herb: in vitro antimalarial activity and molecular modeling to investigate their binding modes. Med. Chem. Res. 29, 487–494. doi:10.1007/s00044-019-02498-z
Veber, D. F., Johnson, S. R., Cheng, H.-Y., Smith, B. R., Ward, K. W., and Kopple, K. D. (2002). Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 45 (12), 2615–2623. doi:10.1021/jm020017n
Verlet, L. (1968). Computer “experiments” on classical fluids. II. Equilibrium correlation functions. Phys. Rev. Journals Arch. 165 (1), 201–214. doi:10.1103/PhysRev.165.201
Verma, R. K., Prajapati, V. K., Verma, G. K., Chakraborty, D., Sundar, S., Rai, M., et al. (2012). Molecular docking and in vitro antileishmanial evaluation of chromene-2-thione analogues. ACS Med. Chem. Lett. 3 (3), 243–247. doi:10.1021/ml200280r
Villegas, L. F., Fernández, I. D., Maldonado, H., Torres, R., Zavaleta, A., Vaisberg, A. J., et al. (1997). Evaluation of the wound-healing activity of selected traditional medicinal plants from Perú. J. Ethnofarmacology 55 (3), 193–200. doi:10.1016/s0378-8741(96)01500-0
Wang, J. M., Wolf, R. M., Caldwell, J. W., Kollman, P. A., and Case, D. A. (2004). Development and testing of a general amber force field. J. Comput. Chem. 25 (9), 1157–1174. doi:10.1002/jcc.20035
World Health Organization (WHO) (2020). Ending the neglect to attain the Sustainable Development Goals: a road map for neglected tropical diseases 2021-2030. Geneva: World Health Organization. Avaialable at: https://www.who.int/teams/control-of-neglected-tropical-diseases/ending-ntds-together-towards-2030 (accessed on July 16, 2023).
Yazdanian, M., Glynn, S. L., Wright, J. L., and Hawi, A. (1998). Correlating partitioning and Caco-2 cell permeability of structurally diverse small molecular weight compounds. Pharm. Res. 15 (9), 1490–1494. doi:10.1023/a:1011930411574
Keywords: isoeleutherin, trypanothione reductase, antiamastigote activity, naftoquinones, medical plant
Citation: Albuquerque KCOd, Veiga AdSSd, Silveira FT, Campos MB, Costa APLd, Brito AKM, Melo PRdS, Percario S, Molfetta FAd and Dolabela MF (2024) Anti-leishmanial activity of Eleutherine plicata Herb. and predictions of isoeleutherin and its analogues. Front. Chem. 12:1341172. doi: 10.3389/fchem.2024.1341172
Received: 19 November 2023; Accepted: 16 February 2024;
Published: 06 March 2024.
Edited by:
Suraj Narayan Mali, Institute of Chemical Technology, IndiaReviewed by:
Mikel Etxebeste-Mitxeltorena, Health Research Institute Foundation Jimenez Diaz (IIS-FJD), SpainSusmita Yadav, Birla Institute of Technology, Mesra, India
Copyright © 2024 Albuquerque, Veiga, Silveira, Campos, Costa, Brito, Melo, Percario, Molfetta and Dolabela. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Fâni Dolabela, ZmFuaUB1ZnBhLmJy, ZmFuaWRvbGFiZWxhMjBAZ21haWwuY29t