 Luiza F. O. Gervazoni†
Luiza F. O. Gervazoni† Gabrielle B. Barcellos
Gabrielle B. Barcellos
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Chem., 23 November 2020
Sec. Medicinal and Pharmaceutical Chemistry
Volume 8 - 2020 | https://doi.org/10.3389/fchem.2020.579891
This article is part of the Research TopicDiscovery and Development of Drugs for Neglected Diseases: Chagas Disease, Human African Trypanosomiasis, and Leishmaniasis.View all 11 articles
Leishmaniasis is an infectious parasitic disease that is caused by protozoa of the genus Leishmania, a member of the Trypanosomatidae family. Leishmaniasis is classified by the World Health Organization as a neglected tropical disease that is responsible for millions of deaths worldwide. Although there are many possible treatments for leishmaniasis, these treatments remain mostly ineffective, expensive, and long treatment, as well as causing side effects and leading to the development of resistance. For novel and effective treatments to combat leishmaniasis, many research groups have sought to utilize natural products. In addition to exhibiting potential as therapeutic compounds, natural products may also contribute to the development of new drugs based on their chemical structures. This review presents the most promising natural products, including crude extracts and isolated compounds, employed against Leishmania spp.
Caused by protozoa of the genus Leishmania, which is a member of the Trypanosomatidae family, leishmaniasis is an infectious parasitic disease. This disease has a wide variety of clinical manifestations, ranging from the cutaneous form to the visceral form. Visceral leishmaniasis, the form that can cause death, affects the organs and viscera of mammalian hosts; conversely, cutaneous leishmaniasis, which can be divided into different manifestations, affects the skin and mucous membranes of mammalian hosts. Classified as a neglected disease by the World Health Organization (WHO, 2016), leishmaniasis affects over 300 million people across all continents.
The current treatment for leishmaniasis is based on pentavalent antimonials, which are drugs that were developed over decades (Vianna, 1912; Burza et al., 2018) with a long-established administration profile in the hospital environment. These drugs are becoming increasingly ineffective due to resistance. Amphotericin B emerged as an alternative treatment; however, its long-standing treatment and dose-dependent side effects led to the development of a liposomal formulation to significantly reduce the side effects and duration of treatment; nevertheless, this formulation is expensive. Paromomycin is already registered in India, but the effectiveness of this treatment has not been determined to date in Africa. As the most promising treatment discovered in recent decades and the first oral drug for leishmaniasis, miltefosine is registered in India and a small number of other countries and has recently been registered by the FDA (IMPAVIDO) for the treatment of visceral and cutaneous leishmaniasis. Miltefosine is effective but expensive and teratogenic (DNDi, 2016).
Although there are many possibilities for leishmaniasis treatment, these treatments remain mostly ineffective, expensive and old, as well as causing side effects and leading to the development of resistance. For novel and effective treatments to combat leishmaniasis, many research groups have investigated natural products.
Natural products are secondary metabolites present in the roots, stalks, leaves, fruits, seeds, vegetables, and other parts of plants with a wide structural variety that mediates interactions between plants and their environment. These metabolites are usually observed around the world in diets as main foods or teas, spices, and sauces. A considerable number of metabolites have anti-protozoal activity (Winkel, 2006; Schmidt et al., 2012a).
Natural products are known in pharmacology for having potential applications as therapeutic drugs, which have been described since ancient times, in addition to contributing to the discovery and development of new drugs based on the chemical structures of these products with specific modifications (Viegas et al., 2006; Ioset, 2008).
In this review, we present the most promising crude extracts and isolated compounds derived from the four major plant metabolic pathways; these products were recently studied to determine their effectiveness as chemotherapy agents for treating leishmaniasis.
All crude extracts and compounds that have defined IC50 values are represented in tables at the end of each section.
This review aims to update and summarize information concerning the early drug discovery process based on crude extracts, fractions, and isolated compounds obtained from natural products, specifically herbal-derived compounds, to treat leishmaniasis. The keywords employed in this study included leishmaniasis, natural products, chemotherapy, in vivo, in vitro, and intracellular amastigote using the current databases: PubMed, Web of Science, Science Direct, and Google Scholar. Our search covered English-language articles published in international scientific journals, indexed over the period 2000-2020. The choice criteria were articles that investigated the leishmanicidal activity of natural products against promastigote, axenic amastigote, and intracellular amastigote forms, the mechanism of action and/or the use of advanced techniques to search for alternative treatments for leishmaniasis. Selected articles describing the use of novel natural products with leishmanicidal activity against promastigotes of Leishmania spp. were considered. All crude extracts and compounds that have defined IC50 values are represented in Tables 1–7.
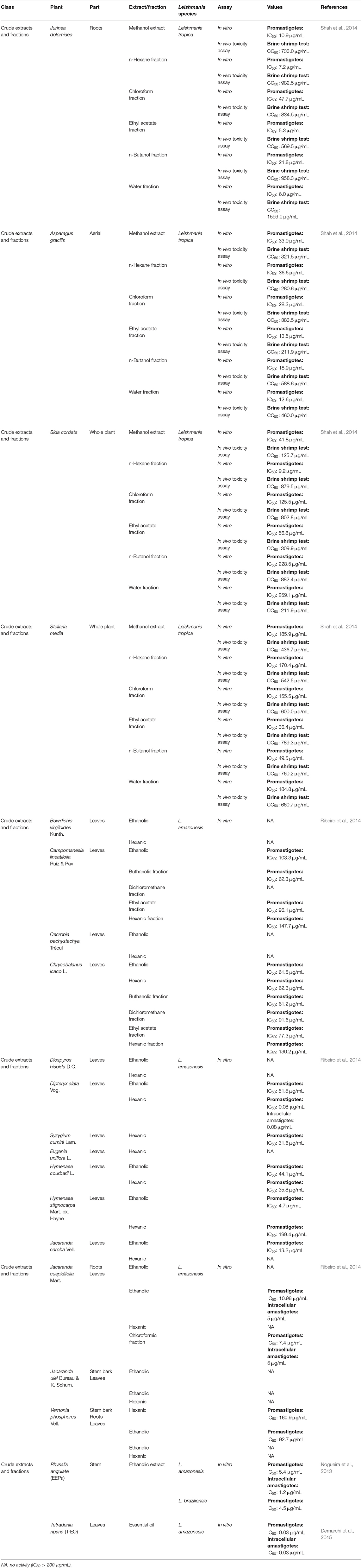
Table 1. Leishmanicidal activities of crude extracts and fractions.
It is well-known that crude extracts have been employed as medicinal drugs since the times of ancient civilizations. In those eras, the simple action of grinding the leaves of certain plants was considered to be medicine. With the growth of technology and knowledge, extracts from leaves, seeds, and other parts of the plant have been tested against several diseases, and some of these extracts have been highly successful (Viegas et al., 2006).
Four plants from different families, namely, Asparagus gracilis from the Asparagaceae family, Stellaria media from the Caryophyllaceae family, Sida cordata from the Malvaceae family and Jurinea dolomiaea (J. dolomiaea) from the Asteraceae family, were tested against a strain of Leishmania tropica isolated from a patient from Pakistan. All four plants were prepared as methanol extracts or n-hexane, chloroform, ethyl acetate, n-butanol and water fractions. The most potent methanol extract was from J. dolomiaea, which exhibited an IC50 value of 10.9 μg/mL, but the highest antileishmanial activity was obtained from the ethyl acetate fraction from J. dolomiaea with an IC50 value of 5.3 μg/mL. All of the extracts and fractions were not toxic, exhibiting IC50 values greater than 100 μg/mL and potent extracts with selectivity indices greater than 10 (Shah et al., 2014).
A series of 16 Brazilian medicinal plants were investigated in vitro to determine their efficacy against L. amazonensis. Among the 44 extracts and fractions, the most potent were the hexanic fraction of Dipteryx alata (D. alata) with an IC50 value of 0.08 μg/mL, the ethanolic fraction of Hymenaea stignocarpa with an IC50 value of 4.70 μg/mL, and both the chloroformic and ethanolic fractions of Jacaranda cuspidifolia (J. cuspidifolia), which exhibited IC50 values of 7.4 and 10.96 μg/mL, respectively (Ribeiro et al., 2014).
Physalis angulata, which is from the Solanaceae family, is a well-known medicinal plant (Mahalakshmi and Nidavani, 2014). For leishmaniasis, Nogueira et al. (2013) tested the ethanolic extract of this plant against two species of Leishmania. In an antipromastigote assay, EEPa (ethanolic extract of Physalis angulata) exhibited IC50 values of 5.35 and 4.50 g/mL for Leishmania amazonensis and Leishmania braziliensis, respectively. The antiamastigote assay using L. amazonensis demonstrated an IC50 value of 1.23 g/mL with a selectivity index of 5.
Tetradenia riparia, a plant from the Lamiaceae family, is commonly employed as a traditional medicine in Africa for infectious parasitic diseases, such as malaria, cryptococcosis, and candidiasis. Against an L. amazonensis promastigote, the essential oil of Tetradenia riparia (TrEO), which contains a mixture of terpenoids, exhibited an IC50 value of 0.03 μg/mL after 24 h. Cytotoxicity in human erythrocytes was tested, and at a concentration of 5 μg/mL, TrEO was determined not to be toxic (Demarchi et al., 2015). Alterations in promastigote morphology were observed. TrEO was able to induce cytoplasmic vacuolization, and membranous profiles and lipid vesicles appeared in the organelle along with membrane blebbing, nuclear fragmentation and chromatin condensation, which are events that are associated with autophagy. Against the intracellular amastigote form, TrEO showed an IC50 value of 0.03 μg/mL and a selectivity index of 5.6. TrEO was also capable of increasing the mRNA expression of iNOS in murine peritoneal macrophages; however, alterations in nitrite production were not observed.
Crude extracts and fraction presented in this section are summarized in Table 1.
Lignans and neolignans are metabolites that can be found in approximately 60 vascular plant families (Winkel, 2006). Lignans are dimeric phenylpropanoids, and neolignans are small molecules with two phenylpropanoid units. The diversity in this class consists of the distribution of aromatic rings and the nature of the propyl fragments (Rye and Barker, 2013). Several groups have chosen to investigate lignans and neolignans because they have properties favorable to drug development and for their anti-inflammatory and antioxidant activity, which may minimize the effects of the inflammatory response (Maia et al., 2020).
Notably, several studies have focused on the search for natural compounds with potential leishmanicidal activity directly using biological screening through phenotypic methods to assess the potential of lignans and neolignans against Leishmania species. The efficacy of a lignin found in garlic (Allium sativum) against L. amazonensis promastigotes was investigated. Dehydrodieuginol (DHDE), an ortho-biphenyl neolignan, showed an IC50 value of 42.2 μg/mL (Rodrigues et al., 2016).
The efficacy of 2,3-dihydrobenzofuran, a neolignan used to treat liver diseases and vascular diseases of the brain and found in propolis and other plants, against L. amazonensis was studied. This compound showed IC50 values of 1.04 and 1.4 μM for promastigotes and intracellular amastigotes, respectively. The intracellular amastigote activity may be mediated by the activation of macrophages, as L. amazonensis-infected BALB/c macrophages treated with 2,3-dihydrobenzofuran exhibited an increase in nitric oxide production, lysosomal volume, and macrophage phagocytic ability (De Castro Oliveira et al., 2017).
Saracoside and lyoniside, two lignans isolated from Saraca indica, were able to interact with L. donovani DNA, inducing apoptosis-like cell death. The IC50 values of lyoniside and saracoside against the intracellular amastigote were 0.79 and 0.82 μM, respectively. BALB/c mice infected with L. donovani were treated intraperitoneally with both lignans (lyoniside and saracoside) at doses of 2.5 and 5 mg/kg/day. Both doses of lyoniside and saracoside were capable of significantly decreasing the parasite loads in the spleen and liver (Saha et al., 2013).
Dyphylin, an arylnaphthalene lignin isolated from Haplophyllum bucharicum, is known to have activity against viruses and cancers. In promastigotes of L. infantum, dyphylin exhibited an IC50 value of 14.4 μM. Furthermore, dyphylin was not determined to be cytotoxic for macrophages exhibiting a CC50 value of 32.2 μM. In the intracellular amastigote of L. infantum, dyphylin exhibited an IC50 value of 0.2 μM, reaching a selectivity index of 178. As a possible mechanism of action, dyphylin may interferes with the cell cycle and protein synthesis and increases intracellular lipid accumulation. However, dyphylin did not increase nitric oxide (NO) production (Di Giorgio et al., 2005).
Niranthin, a lignan from Phyllanthus amarus, was evaluated against L. donovani. The compound was able to inhibit L. donovani promastigote proliferation and exhibited good activity against intracellular amastigote with an IC50 value of 1.26 μM; this lignan was not observed to be toxic to macrophages. The effects of niranthin were also tested against an antimony-resistant strain of L. donovani. Niranthin exhibited an IC50 value of 1.68 μM, indicating that the compound is able to inhibit resistant amastigotes. It is essential for the drug discovery process to determine how natural compounds interfere in the host-parasite relationship, since the ideal compound should not be as toxic to the host as it is to the parasite. Niranthin was determined to have the ability to induce apoptosis (Chowdhury et al., 2012).
To verify the effect of niranthin in vivo, BALB/c mice were infected intracardially with L. donovani and treated with niranthin for 3 weeks at intraperitoneal or intramuscular doses of 5 and 10 mg/kg/day. Splenic and hepatic parasitic loads were almost completely eliminated at the dose of 10 mg/kg/day. Immunological analyses were performed, indicating the ability of niranthin to increase NO levels and switch from a Th2 response to a Th1 response (Chowdhury et al., 2012).
To optimize the choice of lignans and neolignans with potential effects against Leishmnia species and to prevent possible failures from being detected only in preclinical tests, several computational tools can contribute strongly to database creation by predicting protein functions, modeling protein structures, simulating metabolic pathway kinetics, predicting biological activities, predicting toxicity, and predicting the affinities and flexibilities between receptors and ligands, which can facilitate the development and identification of drugs with the potential to treat various diseases and promote the development of efficacious drugs with reduced toxicity (Maia et al., 2020).
The design and synthesis of analogs of natural compounds is a strategy extensively used to identify effective new treatments against leishmaniasis. Design analogs enable the enhancement of many biological and chemical characteristics of compounds to afford new hits, such as bioactivity, selectivity, water solubility, and lipophilicity (Meanwell, 2011).
Along with the approach of advanced technologies for the design of new biologically more potent drugs, it is essential to understand the biology of the parasite to direct these studies.
Two neolignans, threo, threo-manassantin A and threo, erythro-manassantin A, isolated from Saururus cernuus exhibited activity against promastigotes (IC50 of 35.4 and 17.6 μM, respectively) and intracellular amastigotes (IC50 of 20.4 and 16.0 μM, respectively) of Leishmania amazonensis. Regardless of the mode of action, these compounds seem to act directly on parasites, since host cells did not show signs of cell activation. Both molecules were determined to be able to interact with the parasite plasmatic membrane and to interfere with the parasite nucleus (Brito et al., 2019).
Virtual screening and experimental validation have been utilized to identify lignans with leishmanicidal potential, low toxicity, and selective activity against several Leishmania targets. A set of 160 lignans (i.e., 14 furans, 10 furofurans, 14 dibenzylbutyrolactols, 22 dibenzylbutanes, 21 dibenzocycloocyadienes, 17 aryltetralins, 3 arylnaphthalenes, 8 neolignan alkyl aryl ethers, 16 neolignan benzofurans, and 9 neolignan benzodiones) were selected using predictive models that were built using data for L. major and L. braziliensis (Maia et al., 2020).
In brief, the workflow consisted of predicting the ADMET properties of these lignans. Through this tool, 42 compounds have good lipophilicity, water solubility, pharmacokinetic action and low or no predicted risk for the development of mutagenicity, tumorigenesis, negative effects on the reproductive system, or irritability. Next, ligand-based virtual screening was performed to evaluate the potential antileishmanial activity of these compounds using the random forest (RF) algorithm with the parameters of specificity, sensitivity, accuracy, positive predicted value (PPV), and negative predicted value (NPV) for performance and robustness. This model was able to select 11 compounds with active potential, with probabilities ranging from 50 to 75%, for L. major, and 21 potentially active compounds against L. braziliensis were selected and exhibited the same probabilities.
To choose the potential targets in both L. major and L. braziliensis, sequence alignment was employed to verify the similarities and identities of the enzymes selected in this study across different species. In addition, differences and structural similarities could be identified that might contribute to rational drug planning. After sequence alignment and homology modeling were performed, three enzymes were chosen for both species: GPDH (glycerol-3-phosphate dehydrogenase), PTR1 (pteridine reductase 1), and TR (trypanothione reductase).
After all these virtual screenings, four lignans (secoisolariciresinol, pinoresinol-4-O-β-D-glucopyranoside, epipinoresinol-4-O-β-D-glucopyranoside, and pinoresinol-4-O-β-d-apiofuranosyl-(1 → 2)-β-d-glucopyranoside) were selected, and their potential to inhibit the growth of promastigote forms of L. major and L. braziliensis was tested. Epipinoresinol-4-O-β-d-glucopyranoside was the only compound that exhibited activity against both species tested, presenting IC50 values for L. major and L. braziliensis of 36.5 and 5.4 μM, respectively (Maia et al., 2020).
Lignans that have a defined IC50 are summarized in Table 2.
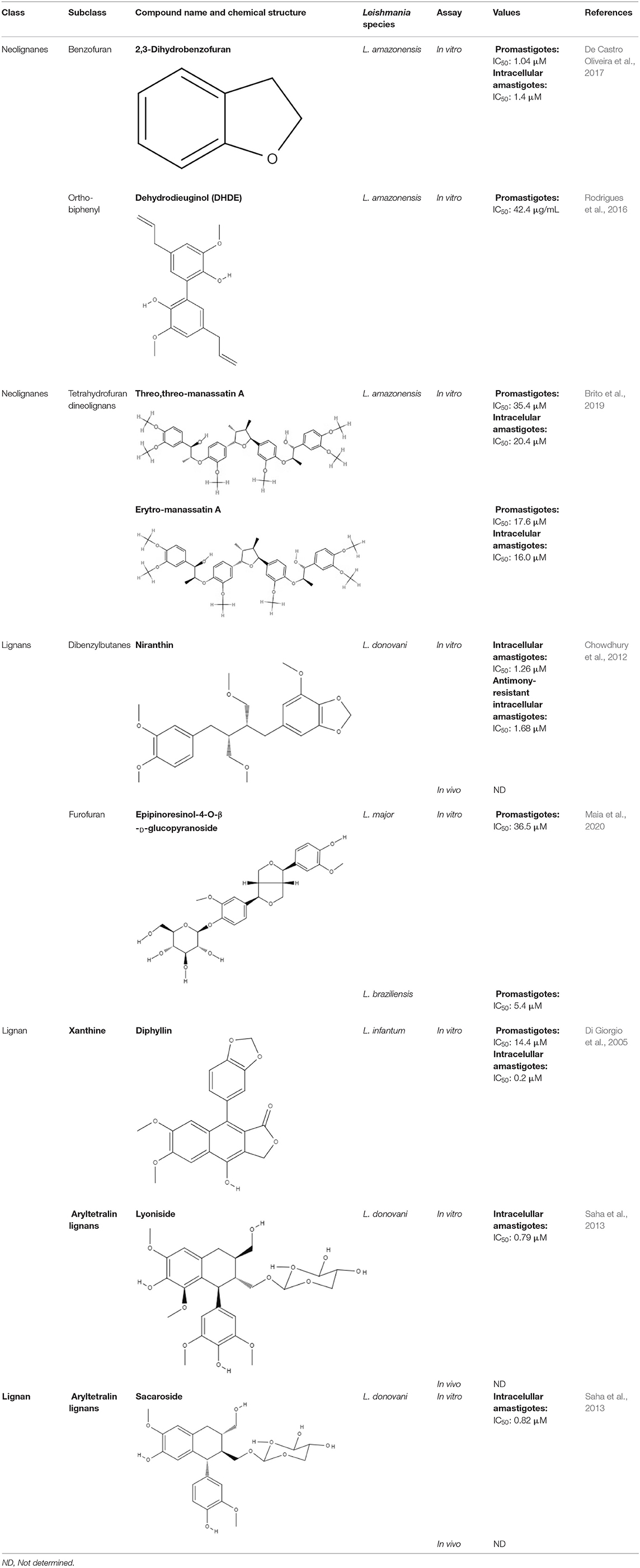
Table 2. Chemical structure and leishmanicidal activities of lignans and neoligans.
Coumarins are derivatives that have a hydroxyl group, which differs in their biological properties. Many enzymes are related to coumarin biosynthesis; therefore, this group has various classes, such as simple coumarin, dimeric coumarin, furanocoumarin, and pyranocoumarin (Jain and Joshi, 2012). The diversity of structures within the coumarin group enables them to exhibit many biological activities, including anti-Leishmania activity. Various studies have described not only the in vitro activity of these compounds against Leishmania but also their mechanism of action and performance in preclinical studies. Mammaea A/BB, which were extracted from Calophyllum brasiliense, showed IC50 values of 7.4 μM against promastigotes and 14.3 μM against intracellular amastigotes of Leishmania amazonensis (Brezan et al., 2008). Regarding the mechanism of action, mammaea A/BB was able to induce mitochondrial membrane damage and cause changes in ultrastructure in L. amazonensis promastigotes (Brenzan et al., 2012). However, only when topically and intramuscularly administered did mammaea A/BB reduce the lesion sizes in mice infected with L. amazonensis compared to mice treated with meglumine antimoniate (Tiuman et al., 2012).
Helietta apiculata Benth is a native plant of Paraguay, Brazil, and Argentina and is popularly known as “canela-de-veado” in Brazil. (+)-3-(1′-dimethylallyl)-Decursinol and (-)-heliettin, two coumarins extracted from Helietta apiculata Benth, were tested against L. amazonensis in vitro and in vivo. In L. amazonensis promastigotes, the IC50 values were 35.8 μM and 18.5 μM for (+)-3-(1′-dimethylallyl)-decursinol and (-)-heliettin, respectively (Table 3). In the in vivo study, BALB/c mice infected with L. amazonensis were injected intraperitoneally with 10 mg/kg/day (+)-3-(1′-dimethylallyl)-decursinol or (-)-heliettin for 14 days. Both coumarins were capable of decreasing parasite loads similar to those observed when the reference drug, meglumine antimoniate, was used (Ferreira et al., 2010).
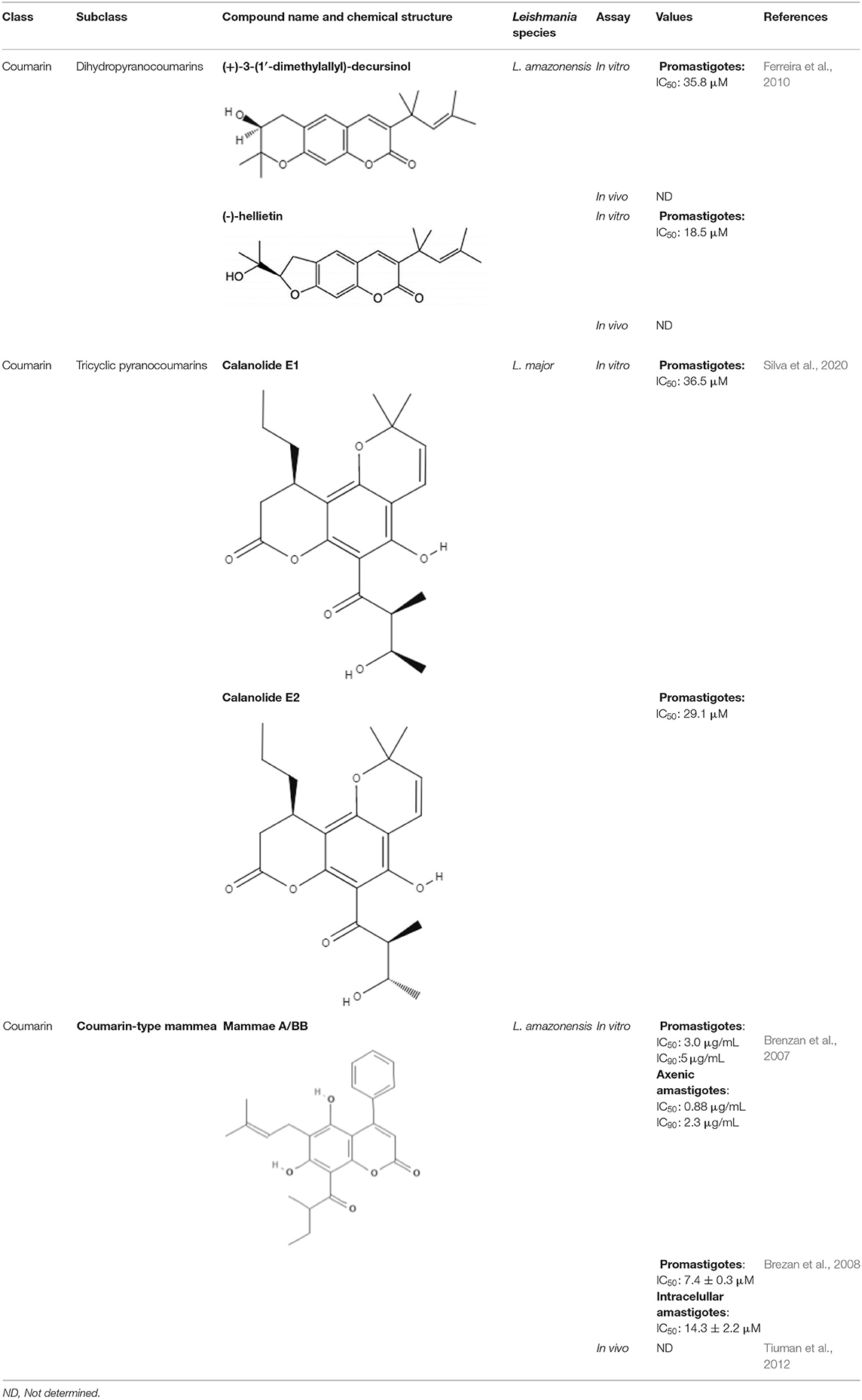
Table 3. Chemical structure and leishmanicidal activities of coumarins.
Part of their life cycle of Leishmania parasites occurs in the sand fly. Since these parasites develop entirely in the digestive system of the vector, interacting with digestive enzymes and other structures from the intestinal tract of the vector, little is known concerning the effect of plant-derived secondary metabolites during the interaction between parasites and vector or even on basic sand fly digestive physiology. Additionally, vector control is one of the key strategies for reducing the number of leishmaniasis cases, and it needs more research and development (Ferreira et al., 2018).
Based on this concept, Ferreira et al. (2010) tested the effect of two coumarins, esculin and esculetin, on sand flies infected with L. infantum and L. mexicana. These molecules were added to the sugar meal of Lutzomia longipalpis. Interestingly, esculetin significantly reduced the viability of L. infantum and L. mexicana in a concentration-dependent manner. Esculin also might block the transmission of leishmaniasis with no repellent effects or reduction in the amount of sugar ingested. In this way, these compounds may represent promising tools for starting the development of antiparasitic sugar baits with less selective pressure for resistance in vector populations (Ferreira et al., 2010). This work demonstrates that coumarin is a promising natural compound that can act on two fronts: as a treatment for leishmaniasis and as a tool to control leishmaniasis vectors.
Two coumarins obtained from stem bark of Calophyllum brasiliense demonstrated activity against amastigotes of Leishmania infantum. Calanolides E1 (1) and E2 (2) presented IC50 values of 37.1 and 29.1 μM, respectively (Table 3). The structure-activity relationship between compounds 1 and 2 was determined. Compound 2, corresponding to anti stereochemistry between carbons C-20 and C-30, showed higher activity against amastigote forms of L. infantum, suggesting that the configuration of C-30 plays an important role in the interaction of this derivative and the tested parasites (Silva et al., 2020).
Moreover, compounds 1 and 2 were subjected to substructure filtering to evaluate their PAINS characteristics. This analysis is crucial to the development of new lead compounds, since some physical/chemical proprieties of the studied compounds could be associated with their reactivity (non-covalent binding) or non-specific interactions with therapeutic targets of parasites (Silva et al., 2020). Both compounds did not contain any PAINS substructures; in other words, there is a reduced probability that their biological activities are artifacts caused by reactivity or colloidal aggregation. These data suggest that coumarins 1 and 2 may serve as scaffolds in the design of novel drug candidates for leishmaniasis (Silva et al., 2020).
Coumarins that have a defined IC50 are summarized in Table 3.
Generally observed in carbohydrate derivatives, such as glycosides, starches, esters and sugar esters, caffeic acids are the most representative hydroxycinnamic acids. Structural modifications, such as amides or esters, may increase the diversity of biological properties of new analogs. Radicals exhibiting 3,4-dihydroxy-substitution patterns have shown inhibitory properties and have attracted interest with respect to being used as drugs (Touaibia et al., 2012).
Computer tools have been used as a preview screening of compounds to evaluate whether the compounds have the chemical characteristics of an oral drug. Before bioguided assays of caffeic acid against Leishmania sp. were conducted, an in silico test of caffeic acid was performed to assess its potential as an oral drug. Molinspiration property calculation software (www.molinspiration.com) was used to calculate the parameters related to oral bioavailability according to Lipinski's rule of five. Lipinski's rule of five describes important molecular properties for a drug's pharmacokinetics in the human body with a high probability of human intestinal absorption and oral bioavailability. Caffeic acid satisfied Lipinski's rule of five with no violation, demonstrating that it is a good drug candidate for oral administration. After the in silico evaluation, bioguide assays were developed. Caffeic acid showed IC50 values of 12.5 μg/mL against promastigotes (Bortoleti et al., 2019), 16.0 μM against intracellular amastigotes of Leishmania amazonensis (Montrieux et al., 2014) and 21.9 μM for intracellular amastigotes of L. infantum (Garcia et al., 2019) (Table 4). Regarding these promising in vitro results, the effects of caffeic acid in an in vivo model of infection were examined. A preclinical trial of caffeic acid in BALB/c mice infected with L. amazonensis promastigotes was conducted. The caffeic acid treatment was administered by the intralesional route every 4 days for 30 days of the experiment. The treatment was able to reduce lesion sizes and parasitic loads in treated animals compared to untreated animals and animals treated with vehicle (Montrieux et al., 2014).
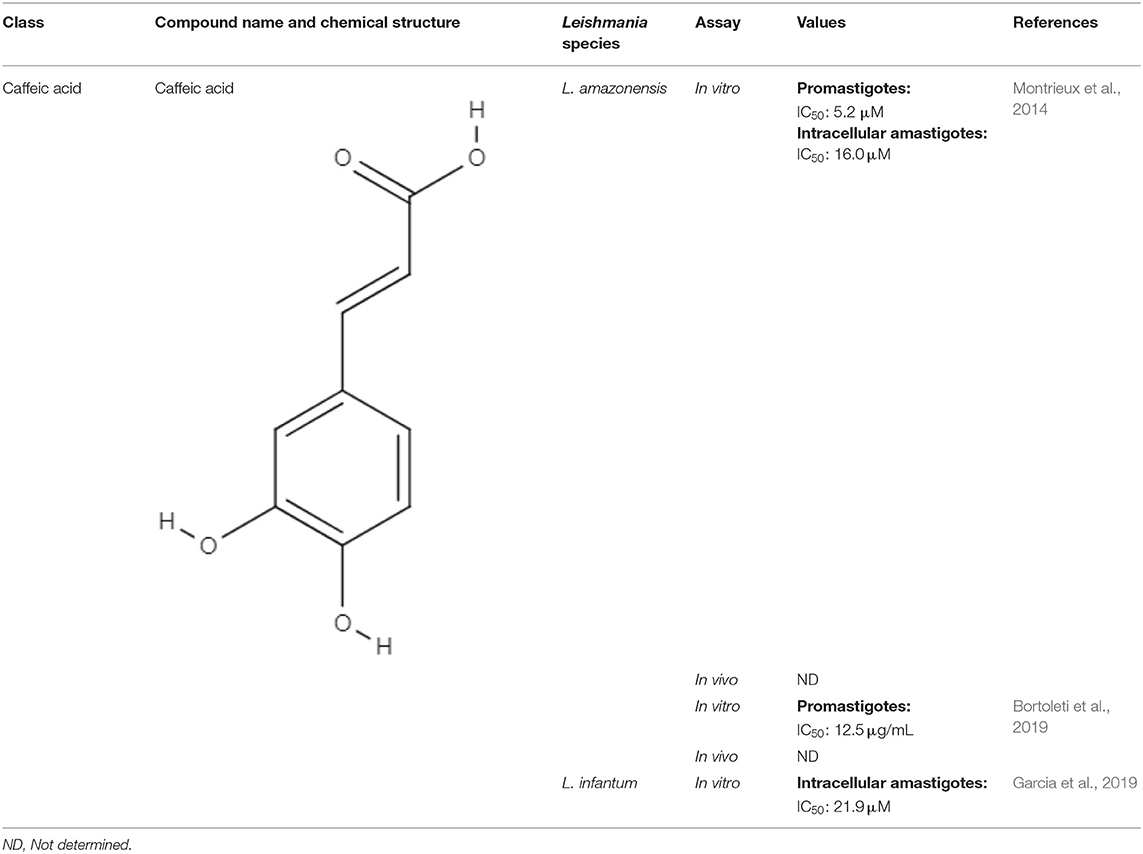
Table 4. Chemical structure and leishmanicidal activities of caffeic acid.
As a mechanism of action, caffeic acid was able to alter promastigote cell morphology and cell volume accompanied by loss of mitochondrial integrity, increase in reactive oxygen species (ROS) production, phosphatidylserine exposure, and loss of plasma membrane integrity, suggesting an apoptosis-like process. Caffeic acid also increased TNF-α, ROS, and NO and reduced IL-10 levels, as well as iron availability (Bortoleti et al., 2019; Garcia et al., 2019). Through these results, it is possible to conclude that caffeic acid has leishmanicidal effects with a mechanism of action that triggers multiple targets that affect the viability of the parasite.
Although caffeic acid has properties that make it a good candidate for oral drugs, its distribution in biological systems is limited due to its hydrophobic nature (Durak et al., 2020).
However, it is important to note that no studies were identified that employed advanced technologies in the investigation of caffeic acid or its derivatives in Leishmania spp. or evaluated the pharmacokinetics of this substance. One hypothesis for this scarcity of published research is that although caffeic acid has had its biological effects characterized, its limitations in relation to bioavailability, such as its hydrophobic nature, make the synthesis of this compound difficult (Durak et al., 2020).
Based on their aromatic carbon skeletons, quinones can be classified as benzoquinones, anthraquinones and naphthoquinones. The benzoquinones comprise ubiquinone and plastoquinone, which differ in their substitution patterns and exhibit different levels of unsaturation on their side chain. Ubiquinones are involved in respiratory chain reactions. Anthraquinones are the oldest known compounds that are used as colorants. Naphthalene is the most natural naphthoquinone and is important in medicinal chemistry because it exerts biological effects on various pathogens, such as Leishmania (Schmidt et al., 2012b).
Plumbagin, a naphthoquinone extracted from Pera benensis, was tested against L. donovani and exhibited an excellent IC50 value of 0.34 and 0.21 μM for promastigotes and axenic amastigotes, respectively. It has been shown that the possible mechanism of action of this compound involves the non-competitive inhibition of trypanothione reductase, a key enzyme in Leishmania redox homeostasis, leading to an increase in reactive oxygen species and changing the redox balance (Sharma et al., 2012).
Buparvaquone (BPQ) and its phosphate prodrugs (BPQ-3-PHOS and 3-POM-BPQ) are hydroxynaphthoquinone, which were tested against several species of Leishmania (L. major, L. amazonensis, L. aethiopica, L. mexicana, and L. panamensis) in vitro. BPQ, BPQ-3-PHOS, and 3-POM-BPQ demonstrated low IC50 values against promastigotes and amastigotes of all Leishmania species tested, exhibiting better activity than amphotericin B and pentostan (Mäntylä et al., 2004). The IC50 values of BPQ, BPQ-3-PHOS, and 3-POM-BPQ for each Leishmania species are described in Table 5.
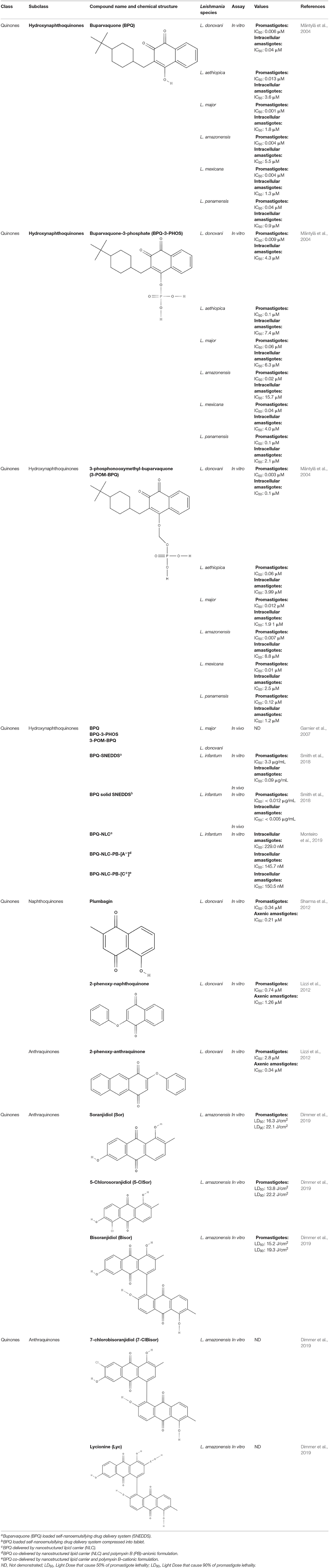
Table 5. Chemical structure and leishmanicidal activities of quinones.
BPQ, BPQ-3-PHOS, and 3-POM-BPQ were also tested in vivo with different topical formulations as a hydrous gel, an anhydrous gel and an emulsion, targeting Leishmania major-infected BALB/c mice (Garnier et al., 2007). The hydrous gel formulation produced the best results. This formulation inhibited the infiltration of infected cells and decreased parasitic load by approximately 50%. These formulations were additionally tested against L. donovani-infected BALB/c mice. BPQ-3-PHOS was demonstrated to be the most active compound, with a decrease of approximately two-thirds in the liver parasite burden compared to the untreated control (Garnier et al., 2007).
It has previously been described that BPQ is poorly soluble in water with a lower in vivo activity (Croft et al., 1992). To overcome this limitation, a novel BPQ-loaded self-nanoemulsifying drug delivery system (BPQ-SNEDDS and BPQ solid SNEDDS) was developed. These formulations demonstrated activity against L. infantum promastigote and intracellular amastigote forms that was superior to miltefosine (Smith et al., 2018). An oral pharmacokinetic assay in mice was performed, and the BPQ-SNEDDS showed good bioavailability, increasing the AUC0−24 by 55%. During in vivo infection, BPQ-SNEDDS and BPQ solid SNEDDS were able to inhibit parasite replication in the spleen and liver of infected mice. These formulations are promising and may be able to overcome the limitations found in the use of BPQ, and further studies are warranted to provide more information regarding their effects (Smith et al., 2018).
In a similar approach, three different nanostructured formulations (BPQ-NLC, BPQ-NLC-PB-[A−] and BPQ-NLC-PB-[C+]) were developed and employed against L. infantum. All formulations showed lower IC50 values (229.0 nM, 145.7 nM, 150.5 nM for BPQ-NLC, BPQ-NLC-PB-[A−] and BPQ-NLC-PB-[C+], respectively) than did free BPQ, thereby improving the antiamastigote activity of this compound (Monteiro et al., 2019).
Over the years, natural product libraries and collections have been successfully established, enabling investigators to link chemical classes to biological activities. In recent years, quinone activity has been exploited, and a library of quinone-polyamine conjugates has been constructed. These conjugates were tested against three species known to cause human parasitic diseases, including L. donovani. Some derivatives were determined to inhibit the activity of trypanothione reductase. All compounds presented good IC50 against L. donovani, with emphasis being placed on compounds 2-phenoxy-anthraquinone and 2-phenoxy-naphthoquinone, which demonstrated the best IC50 values against the axenic amastigotes (0.34 and 1.26 μM, respectively) and promastigotes (2.8 and 0.74 μM, respectively) of L. donovani (Lizzi et al., 2012).
Using a different approach to topical treatment of cutaneous leishmaniasis, Dimmer et al. (2019) tested the antiparasitic photodynamic inactivation of soranjidiol (Sor) and its derivatives 5-chlorosoranjidiol (5-ClSor), bisoranjidiol (Bisor), 7-chlorobisoranjidiol (7-ClBisor), and lycionine (Lyc). Sor and its derivatives are anthraquinones isolated from Heterophyllaea pustulata Hook f. Photodynamic inactivation (PDI) is a methodology that combines photosensitive drugs with light to kill parasites. Light excites photosensitive molecules that generate ROS in the presence of oxygen. Soranjidiol, 5-chlorosoranjidiol and bisoranjidiol combined with violet-blue light caused a decrease in parasite viability of L. amazonesis promastigotes. Bisoranjidiol-mediated PDI induced significant alterations in the size and shape of promastigotes. Furthermore, soranjidiol is the most efficient anthraquinone to combat leishmaniasis, causing fewer toxic effects in fibroblast cells (Dimmer et al., 2019).
The quinones that were presented along with their leishmanicidal activities (IC50) are summarized in Table 5.
Alkaloids are nitrogenous compounds with alkaline character. However, there are some exceptions, with certain compounds containing amino or amido atoms. Alkaloids are classified based on the presence and activity of specific amino acids, which form a fundamental component of the alkaloid skeleton. For example, the amino acid lysine produces piperidine, quinolizidine and indolizidine alkaloids, and the amino acid ornithine produces pyrrolidine and tropane alkaloids. The amino acid tyrosine gives rise to phenylethylamines and tetrahydroidoquinoline alkaloids. Tyrosine also produces other alkaloids in which phenolic oxidative links play a fundamental role (Kurek, 2019).
All alkaloids possessing amino acid precursors are true alkaloids or protoalkaloids. True alkaloids share a common heterocyclic ring with one nitrogen atom, while the main characteristic of protoalkaloids is a nitrogen atom that does not belong to the heterocyclic ring, such as cocaine. However, many alkaloids do not originate from amino acids but from the amination of other substrates, such as steroids, terpenoids, acetates and phenylalanine. Some authors have classified alkaloids that do not originate from amino acids as pseudoalkaloids. Finally, alkaloids that are produced via pathways resembling those by which purine nucleic acids are produced are classified as purine alkaloids (Dewick, 2009).
Many types of alkaloids have been described as having biological activities against trypanosomatids, such as Leishmania spp. Two heterocyclic steroids were isolated from Solanum lycocarpum, and their in vitro and in vivo activities were tested. Against L. mexicana, intracellular amastigote forms, solamargine, and solasonine showed IC50 values of 6.03 and 5.9 μM, respectively. These IC50 values were superior to the IC50 observed with sodium stibogluconate (Lezama-Dávila et al., 2016).
Interestingly, solamargine and solasonine induced different immunochemical pathways in macrophages and dendritic cells. L. mexicana was eliminated more efficiently by dendritic cells when incubated with solamargine and solasonine at a concentration of 10 μM. Additionally, both compounds were capable of enhancing the expression levels of transcription factors, such as NFκB/AP-1, also at a concentration of 10 μM. Nitric oxide levels decreased in both macrophages and dendritic cells only after treatment with solamargine, indicating that its mechanism of action is dependent on nitric oxide (Lezama-Dávila et al., 2016).
The in vivo study was performed using C57BL/6 mice infected with L. mexicana. Treatment with topical formulations of 10 μM solamargine and solasonine significantly reduced parasite loads and lesion sizes in the ear (Lezama-Dávila et al., 2016).
Combination therapy has been employed as a strategy for improving the treatment of leishmaniasis. The combination of piperine and its analog capsaicin with meglumine antimoniate has been tested. Against L. infantum, both alkaloids alone showed better antipromastigote activity than meglumine antimoniate with IC50 values of 5.01 μg/mL for capsaicin and 3.03 μg/mL for piperine. The combinations of piperine or capsaicin with meglumine antimoniate (50% + 50%) were the most effective against promastigotes, exhibiting IC50 values of 2.1 and 2.9 μg/mL, respectively. The best antiamastigote activity occurred in the combination of piperine with meglumine antimoniate (25% + 75%), presenting a synergistic effect with an IC50 value of 7.3 μg/mL (Vieira-Araújo et al., 2018).
It has already been described that piperine has a bioavailability enhancing effect (Randhawa et al., 2011). To explore this property and improve the bioavailability of amphotericin B, nanoformulations of piperine (PIP), and amphotericin B (AmB) coated with nanoparticles (HDGG-AmB-Pip-NPs and Eu-HDGG-AmB-Pip-NPs) were developed, and their leishmanicidal effects were evaluated. Both formulations showed good IC50 against promastigotes (21.9 and 24 ng/mL, respectively) and intracellular amastigotes (4.9 and 18.3 ng/mL, respectively) of L. donovani. When formulations were administered in L. donovani-infected golden hamsters by the intraperitoneal route, HDGG-AmB-Pip-NPs and Eu-HDGG-AmB-Pip-NPs reduced parasite load by 95 and 96%, respectively, compared to the untreated control and were more effective than amphotericin B treatment (Ray et al., 2020).
Pharmacokinetics analysis showed that Eu-HDGG-AmB-Pip-NPs improved the plasma concentration-time profile of amphotericin B compared to amphotericin B treatment. The tissue distribution was evaluated, and Eu-HDGG-AmB-Pip-NPs showed the highest amphotericin B accumulation in the liver and spleen. This compound was also detected in the kidney, but at the lowest concentration, it was detected in the liver and spleen. Furthermore, Eu-HDGG-AmB-Pip-NPs cause changes in serum levels. The use of piperine in association with reference drugs for the treatment of leishmaniasis in nanoformulations shows promising results that should be further explored (Ray et al., 2020).
Senna spectabilis is a tree of the family Fabaceae, and piperidine alkaloids, such as (-)-cassine, (-)-spectaline, (-)-3-o-acetylcassine and (-)-3-O-acetylspectaline, can be extracted from it. These alkaloids were tested against L. amazonensis promastigotes, and all of them presented leishmanicidal effects, with compound (-)-spectaline being more effective (IC50 = 15.8 μg/mL). However, the IC50 value of this compound was higher than that of amphotericin B. In addition, all piperidine alkaloids showed less toxicity than amphotericin B. In a more modern approach, in silico analysis using molecular docking was performed to evaluate how these piperidine alkaloids bound to the enzyme arginase. The alkaloid (-)-spectaline showed a stronger interaction with arginase than other alkaloids, suggesting arginase as a possible target for the (-)-spectaline (Lacerda et al., 2018).
Berberine is an isoquinoline alkaloid and is extracted from Berberis vulgaris. Previous studies have shown that berberine has leishmanicidal activity and can induce a redox imbalance following the enhanced generation of ROS (Saha et al., 2009). Since Leishmania has only one mitochondrion, which is the major ROS productor, the effects of berberine on the mitochondria of non-pathogenic Leishmania donovani UR6 were tested. This alkaloid showed a reduction in the viability of promastigotes in a concentration-dependent manner with an IC50 value of 4.8 μM and stimulated the generation of ROS in these cells. Berberine was also able to increase the levels of mitochondrial superoxide of promastigotes and induced depolarization of mitochondrial transmembrane potential. Concentration-dependent inhibition of complex I-III and II-III activities was observed in promastigotes, as well as a decrease in ATP levels. Although berberine has been tested on non-pathogenic parasites, these data provide support for future studies to search for a possible mechanism of action for this promising alkaloid (De Sarkar et al., 2018).
In the search for a new approach to improve the treatment of cutaneous leishmaniasis, the leishmanicidal effects of chitosan (CS)-polyethylene oxide (PEO) nanofibers containing berberine were tested. CS-PEO-Berberine showed an IC50 of 0.2 and 0.9 μg/mL against promastigotes and in intracellular amastigotes of L. major, respectively, indicating that this formulation can be provided as a good alternative topical treatment for cutaneous leishmaniasis (Rahimi et al., 2020).
Colchicine, demecolcine, and thiocolchicoside are tropolone alkaloids extracted from Colchicum kuurdicum (Bornm.) Stef., a perennial monocotyledon plant. Eight tropolones (colchicoside, 2-demethyl colchicine, 3-demethyl colchicine, demecolcine, colchifoline, N-deacetyl-N-formyl colchicine and cornigerine) were isolated from Colchicum kuurdicum and tested against L. major. All tropolones showed good IC50 and leishmanicidal effects. Colchicoside and colchicine were the most effective, exhibiting IC50 values of 4.0 and 8.7 μg/mL against intracellular amastigote forms. All tropolones presented in vitro iron chelating activity between 19 and 25%, with colchicine showing the highest activity. Tropolones also demonstrated significant anti-inflammatory effects, with anti-denaturation effects of between 50 and 80%. Additionally, these compounds caused only 5% hemolysis, demonstrating safety for systemic usages. To analyze toxicity, a brine shrimp toxicity test and an acute toxicity test in mice were performed. All tropolones showed higher LD50 and LD90, and the median lethal dose of these compounds was between 6 and 10 mg/kg. To evaluate a possible mechanism of action of these tropolones, molecular docking was performed targeting tubulin protein. Based on dock scores, colchicoside and demecolcine presented the highest and the lowest affinity to tubulin, respectively. The best isomers of antitubulin were colchicoside, colchicine and N-deacetyl-N-formyl colchicine. These findings demonstrate that these tropolone alkaloids are promising compounds, especially colchicoside alkaloids, which showed strong results in leishmanicidal effects and docking studies (Azadbakht et al., 2020).
Staurosporine (STS) is an indolocarbazole isolated from Streptomyces sanyensis. To evaluate the antileishmania activity and to elucidate a possible mechanism of induced cell death, natural staurosporines (STS, 7OSTS, 4′D4′OSTS, and SCZ B) and its commercial analogs rebeccamycin, K252a, K252b, K252c, and arcyriaflavin A were tested against L. amazonensis, L. donovani and T. cruzi. The compounds STS and 7OSTS showed the lowest IC50 values against promastigotes of L. amazonensis (0.08 and 3.6 μM, respectively) and L. donovani (2.1 and 0.1 μM, respectively), while 4′D4′OSTS and SCZ B were not active against L. donovani, and rebeccamycin K252c and arcyriaflavin were inactive in both parasites. Compound 7OSTS was more active against L. amazonensis amastigotes (IC50 of 0.1 μM) than was miltefosine. This indolocarbazole induced mitochondrial damage in L. amazonensis but not in L. donovani, as determined using the IC90. However, cytoplasmic membrane permeability in L. donovani was induced by 7OSTS but not in L. amazonensis. Despite the differences in activity observed during the tests against L. amazonesis and L. donovani, the natural indolocarbazole 7OSTS exhibited promise (Cartuche et al., 2020).
Aspidosperma spruceanum Benth. ex Müll. Arg is a tree of the Apocynaceae family that has medicinal properties and has been used for leishmaniasis treatment in Amazonian regions (Morales-Jadán et al., 2020). Using bioinformatic tools, a new approach to predict new compounds capable of fighting diseases, three indole alkaloids (from Aspidosperma spruceanum) were investigated against Leishmania targets. The chosen indole alkaloids were aspidocarpine (APC), aspidoalbine (APA) and tubotaiwine (TBT), and in silico tests showed that all of them fulfilled Lipinski's rule. Four Leishmania species cause leishmaniasis in the region where Aspidosperma spruceanum is used: L. braziliensis, L. panamensis, L. amazonensis, and L. mexicana. Five targets of Leishmania common to all these species were modeled, and 3D structures were determined for the targets, which were dihydrofolate reductase-thymidylate synthase (DHFR-TS), pteridine reductase 1 (PTR1), pyruvate kinase (PK), hypoxanthine-guanine phosphoribosyltransferase (HGPRT), and squalene synthase (SQS) (Morales-Jadán et al., 2020).
Docking simulations showed that all three indole alkaloids can interact strongly with the Leishmania targets. As pidoalbine had more affinity for the active site of PTR1 and against L. panamensis, it is able to inhibit some of the functional aspects. The alkaloids demonstrated more affinity to Leishmania proteins than to human homologs. These results may be useful for guiding future analyses of the leishmanicidal effects of these compounds in vivo and elucidating their possible mechanisms of action (Morales-Jadán et al., 2020).
Alkaloids that were presented along with their leishmanicidal activities (IC50) are summarized in (Table 6).
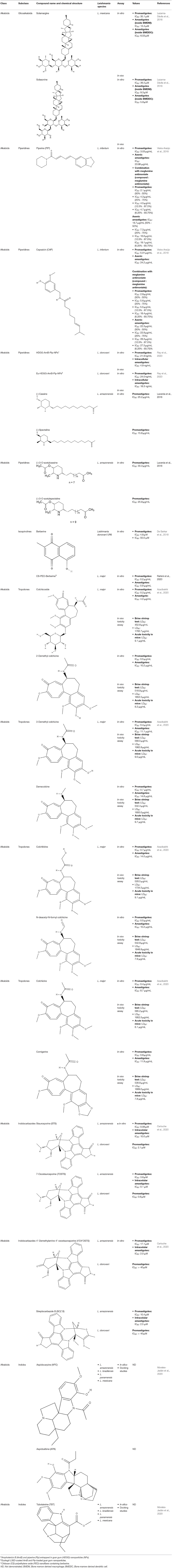
Table 6. Chemical structure and leishmanicidal activities of alkaloids.
Flavonoids can be defined as a group of metabolites originating from the combination of the skikimate and acetate pathways biosynthesized from cinnamic acid, which has a C6-C3-C6 basic structure and phenylbenzopyran functionality. The product of the first cyclization is chalcone, a precursor of most groups of flavonoids. These natural compounds are divided according to the linkage between the aromatic rings to the benzopyran portion in phenylbenzopyran flavonoids, isoflavonoids and neoflavonoids. Phenylbenzopyrans flavonoids (2-phenylbenzopyrans) are classified as flavan, flavanone, flavone, flavonol, dihydroflavonol, flavan-3-ol, flavan-4-ol and flavan-3,4-diol according to both oxidation and saturation of the heterocyclic C-ring. Isoflavonoids (3-benzopyrans) have a 3-phenylchroman structure with a wide range that can be classified as isoflavan, isoflavone, isoflavanone, isoflav-3-ene, isoflavanol, rotenoid, coumestane, 3-arylcoumarin, coumaronochromene, and pterocarpan. Neoflavonoids (4-benzopyrans) have a structure similar to flavonoids and isoflavonoids and are divided into 4-arylcoumarins, 3,4-dihydro-4-arylcoumarins and neoflavenes. These groups have a variety of biological activities, including against trypanosomes (Winkel, 2006; Schmidt et al., 2012b).
Quercetin, a flavonol extracted from Kalanchoe pinnata, demonstrated activity against promastigotes of L. amazonensis (IC50 value of 31.4 μM after 48 h). This compound increased reactive oxygen species levels, causing mitochondrial damage and leading to the death of the parasite (Fonseca-Silva et al., 2013). Against L. amazonensis intracellular amastigotes, quercetin exhibited an IC50 value of 3.4 μM and a selectivity index of 16.8 (Fonseca-Silva et al., 2013).
Quercetin was also capable of inhibiting arginase, an important enzyme in leishmanial infections, as a possible target for leishmaniasis chemotherapy (Manjolin et al., 2013). Despite these studies, the mechanism of action of quercetin is still unknown. This compound was also tested in vivo in L. amazonensis-infected mice, a murine model of cutaneous leishmaniasis. Quercetin was orally administered and reduced lesion size and parasite burden in the infected ear at a dose of 16 mg/kg/day (Muzitano et al., 2009).
Apigenin, an important flavone tested in the last decade (Kashyap et al., 2018), showed IC50 values of 23.7 μM and 4.3 μM for promastigotes and the intracellular amastigote form of L. amazonensis, respectively. The inhibition of intracellular amastigote growth reached 71% after 72 h at the highest dose tested (12 μM) (Fonseca-Silva et al., 2015).
Apigenin was also tested in vivo in the cutaneous form of leishmaniasis. This compound was able to reduce the lesion size and parasitic load compared to the control and the reference (meglumine antimoniate), presenting ED50 and ED90 values of 0.73 and 1.2 mg/kg, respectively (Fonseca-Silva et al., 2016).
As a possible mechanism of action, it was demonstrated that L. amazonensis-infected macrophages treated with apigenin showed an increase in the intracellular reactive oxygen species (ROS) and in the number of double-membrane vesicles and myelin-like membrane inclusions, which are characteristics of the autophagic pathway. Furthermore, fusion between autophagosome-like structures and parasitophorous vacuoles was observed (Fonseca-Silva et al., 2016).
Following new chemotherapy perspectives for leishmaniasis, Emiliano and Almeida-Amaral (2018) tested apigenin in a combination model with miltefosine, which is the first oral drug for leishmaniases. This association was first tested in vitro in THP-1-derived macrophages infected with L. amazonensis promastigotes. The ∑FIC (fractional inhibitory concentration sum) was 1.61, showing an additive effect. The in vivo efficacy of this combination was assessed in a cutaneous murine model with BALB/c mice infected with L. amazonensis. Apigenin and miltefosine were tested alone (2 and 8 mg/kg/day, respectively) or in combination, using half of the original doses (1 mg/kg/day + 4 mg/kg/day, respectively). Both compounds alone exhibited their expected effects in reducing lesion size and parasite load. The combination scheme was also able to significantly reduce the lesion size and parasite load compared to the control, achieving reductions of 75 and 95%, respectively. Serological toxicological markers were measured, indicating possible hepatoxicity of miltefosine alone (8 mg/kg/day). However, the combination of these compounds did not show any renal or hepatic toxicity, indicating the combination scheme as a new favorable model for leishmaniasis treatment since it is effective and less toxic (Emiliano and Almeida-Amaral, 2018).
The effects of (–)-epigallocatechin 3-O-gallate (EGCG), the most abundant flavanol constituent of green tea (Camellia sinensis (L.) Kuntze; Theaceae) has been tested in a murine model of cutaneous leishmaniasis against promastigotes and intracellular amastigotes of L. amazonensis and L. braziliensis. Against L. amazonensis intracellular amastigotes, EGCG demonstrated an IC50 value of 1.6 μM with a selectivity index of 129.4 (Inacio et al., 2013). When tested against L. braziliensis, EGCG demonstrated an IC50 value of 278.8 μM for promastigotes and 3.4 μM for amastigotes with a selectivity index of 149.5 (Inacio et al., 2014). As a possible mechanism of action, EGCG increased the ROS levels, which led to a decrease in the mitochondrial membrane potential and a decrease in the ATP levels. EGCG was also tested against L. infantum-infected macrophages that exhibited an EC50 of 2.6 μM (Inacio et al., 2019).
In vivo, EGCG was tested against a murine model of cutaneous leishmaniasis using L. amazonensis (Inacio et al., 2013) and L. braziliensis (Inacio et al., 2014) and a murine model of visceral leishmaniasis using L. infantum (Inacio et al., 2019). In cutaneous leishmaniasis, EGCG was able to reduce the lesion size and the parasitic load without serological toxicology. A similar effect was shown in visceral leishmaniasis; EGCG was capable of reducing the liver parasite load, presenting ED50 and ED90 values of 12.4 and 21.5 mg/kg/day, respectively.
It is well-known that antimonial resistance is a current problem in leishmaniasis chemotherapy. In an attempt to promote a new strategy for an old problem, flavonoids have been tested as a possible alternative for the treatment of antimonial-resistant leishmaniasis. 2′-Hydroxyflavanone (2HF), a flavanone commonly found in citric fruits, was able to reduce the infection index in BALB/c macrophages infected with wild-type or antimony-resistant L. amazonensis promastigotes with IC50 values of 3.09 and 3.36 μM, respectively. After in silico analysis suggested that 2HF was a safe oral compound, the in vivo assay was performed. BALB/c mice were infected with wild-type or antimony-resistant L. amazonensis promastigotes and treated with 2HF (50 mg/kg/day). 2HF was capable of reducing the lesion size and parasite load compared to untreated and meglumine antimoniate-treated groups in both wild-type and antimony-resistant infections with no hematological or toxicological alterations (Gervazoni et al., 2018).
Several compounds isolated from leaves of Piper rusbyi were tested against three species of Leishmania. Among all the compounds tested, Flavokavain B, a chalcone, demonstrated good results against Leishmania. The IC50 value was 11.2 μM against L. amazonensis, L. donovani, and L. braziliensis, which was more effective than pentamidine. Flavokavain B was also tested in vivo against L. amazonensis infection in the footpads of BALB/c mice. With a 5 mg/kg/day subcutaneous dose, flavokavain B exhibited the best results among those tested, reducing the lesion size and being effective in vivo (Flores et al., 2007).
The potential activity of two biflavonoids isolated from Selaginella sellowii, amentoflavone and robustoflavone, was investigated against the intracellular amastigote of L. amazonensis. The IC50 values of amentoflavone and robustoflavone were 0.2 and 5.3 μM, respectively. In addition, the production of NO decreased in the L. amazonensis-infected peritoneal macrophages treated with amentoflavone, while treatment with robustaflavone increased the production of NO (Rizk et al., 2014).
The genus Mimulus is native to California in North America. Four C-geranyl flavones (diplacone, 3′-O-methyldiplacone, yellow oil, and 3′-O-methyldiplacol) and one geranylated flavone (cannflavin A) were isolated from Mimulus bigelovii. The IC50 value was determined to be 7.5 μg/mL for both 3′-O-methyldiplacone and yellow oil against axenic amastigotes. 3′-O-Methyldiplacol obtained an IC50 value of 7.2 μg/mL, and cannflavin A obtained an IC50 value of 14.6 μg/mL, both of which were also tested against axenic amastigotes (Salem et al., 2011).
Maclura tiinctoria from the Moraceae family is a plant found in tropical countries worldwide, and its extracts are rich in flavonoids. 5,7,3′,4′-Tetrahydroxy-6,8-diprenylisoflavone (CMt), an isolated flavonoid from M. tiinctoria leaf extract, was tested against promastigotes and axenic amastigotes from L. amazonensis and L. infantum. CMt demonstrated IC50 values of 2.7, 6.4, 1.1, and 2.5 μg/mL for promastigote and axenic amastigotes of L. amazonensis and L. infantum, respectively. For both Leishmania species, 5,7,3′,4′-tetrahydroxy-6,8-diprenylisoflavone presented a selectivity index over 180. As a possible mechanism of action, CMt caused disturbances in membrane integrity and membrane potential, including increases in ROS production. In the in vivo model for visceral leishmaniasis, 5,7,3′,4′-tetrahydroxy-6,8-diprenylisoflavone and a new formulation, such as 5,7,3′,4′-tetrahydroxy-6,8-diprenylisoflavone-containing micelles (CMt/Mic), were able to reduce the parasite load in selected organs (liver, spleen, lymph node, and bone marrow) compared to the control groups and miltefosine-treated group. Cytokine analysis indicated a Th1-type response for 5,7,3′,4′-tetrahydroxy-6,8-diprenylisoflavone and CMt/Mic treatment as most promising (Pereira et al., 2020).
Brachydin A, brachydin B, and brachydin C, three dimeric flavonoids from Arrabidaea brachypoda, were evaluated against L. amazonensis, L. braziliensis, and L. infantum promastigotes. The most promising results were obtained for brachydin B and brachydin C, showing IC50 values of 7.05 and 8.8 μM for L. braziliensis and IC50 values of 9.16 and 10 μM for L. amazonensis. L. amazonensis was chosen for the antiamastigote assay, and only brachydin B and brachydin C were tested, exhibiting IC50 values of 2.2 and 6.25 μM, respectively It is interesting to highlight the only structural difference between brachydin A, brachydin B, and brachydin C as substitutes for the C-ring as a methoxyl group for brachydin B and a hydroxyl group for brachydin A. Methoxyl is known to improve membrane permeability, which can explain the best results observed with brachydin B (Rocha et al., 2018).
To investigate possible macrophage and amastigote alterations induced by brachydin B, transmission electron microscopy was performed. Brachydin B did not generate macrophage toxicity, even at higher concentrations (20 and 50 μM). However, for amastigotes, brachydin B induced enlargement of the Golgi apparatus, vesicle accumulation and cytoplasmic disorganization with consequent cell death (Rocha et al., 2018).
Brachydin B was administered in vivo in L. amazonensis-infected mice using two different routes: topical treatment (1% brachydin B) and oral treatment (25 and 50 mg/kg/day). The topical treatment, as well as oral treatment at 25 mg/kg/day, were not able to reduce lesion size compared to the control group. Oral treatment with 50 mg/kg/day reduced the lesion size 1 week after the start of treatment but showed no significant difference between the control group at the end of the treatment (Rocha et al., 2018).
Tryparedoxin peroxidase (Txnpx) and Trypanothione reductase (TryR) are two essential proteins for leishmania survival for their role in parasite redox metabolism. Therefore, these proteins have been considered good targets for the development of new leishmaniasis treatments (Kumar et al., 2017).
Molecular docking analysis was performed for several compounds, including such flavonoids as taxifolin, kaempeferol, quercetin, and epigalloctechin-3-gallate. Quercetin and taxifolin demonstrated the highest binding energy with Txnpx. The molecular docking study also indicated that the Lys136 residue is an essential ligand that is critical for the interactions (Gundampati et al., 2014; Kumar et al., 2017).
Molecular docking was also performed using gp63, a metalloprotease found on both promastigote and amastigote surfaces, which is essential for parasite virulence and pathogenesis. This enzyme is considered a good target for leishmaniasis new treatment development. Lanaroflavone, podocarpusflavone A, amentoflavone, and podocarpusflavone B, which are known biflavonoids, had the most significant interactions in a molecular docking study performed for L. major and L. panamensis gp63. These four flavonoids demonstrated the same pattern of interactions for both Leishmania species. Lanaroflavone was the most promising compound of all considering binding affinity (Mercado-Camargo et al., 2020).
The flavonoids present in this section with a defined IC50 are summarized in the Table 7.
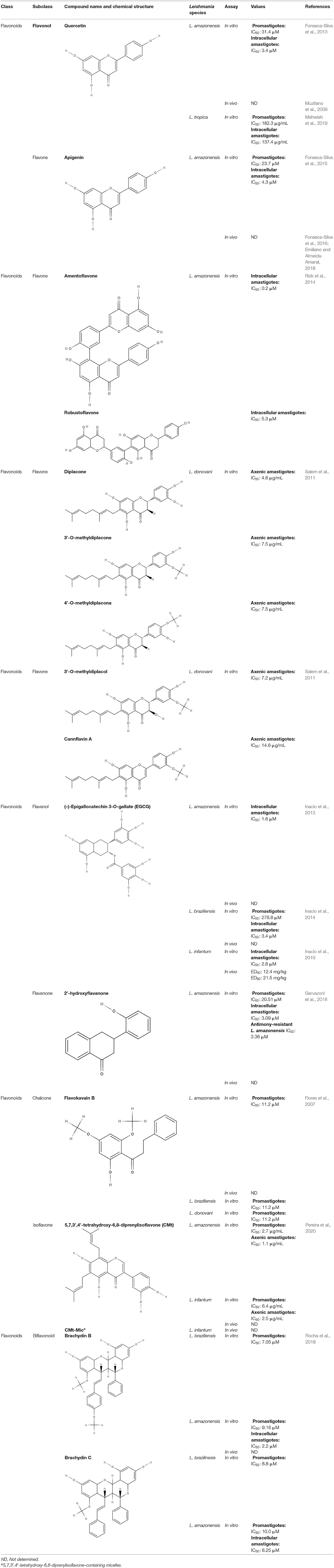
Table 7. Chemical structure and leishmanicidal activities of flavonoids.
Terpenoids are natural compounds derived from C5 isoprene (or isoprenoid) units. The characteristic chemical structure of these compounds contains a skeleton with 5 carbons in a head-to-tail linkage. These compounds are classified as hemiterpenes (C5), monoterpenes (C10), sesquiterpenes (C15), diterpenes (C20), sesterterpenes (C25), triterpenes (C30), and tetraterpenes (C40). The diversity of terpenoids increases their biological activity spectrum, including several species of Leishmania, such as L. major, L. donovani, L. infantum, L. amazonensis, L. braziliensis, L. Mexicana, and L. panamensis (Yamamoto et al., 2015).
Artemisinin extracted from Artemisia annua and its derivatives were tested against promastigotes of L. major, demonstrating an IC50 value of 0.75 μM. Against intracellular amastigotes, artemisinin presented an IC50 value of 3 μM and was not toxic to the macrophages (Yang and Liew, 1993).
Artemisinin was also evaluated against L. donovani. The IC50 value was 160 μM against promastigotes and 22 μM against intracellular amastigotes. Artemisinin induced apoptosis, depolarization of the mitochondrial membrane potential and DNA fragmentation. In vivo, using BALB/c mice infected with L. donovani, artemisinin was administered at 5 and 10 mg/kg/day to reduce the parasite burden in the spleen (Sen et al., 2007, 2010).
The activity of the monocyclic sesquiterpene alcohol (-)-α-bisabolol, utilized in fragrances and extracted from Matricaria chamomilla L., was tested against intracellular amastigotes of L. infantum and L. donovani, presenting IC50 values of 56.9 and 39.4 μM, respectively. In addition to the in vitro investigation, this compound was also evaluated in a visceral leishmaniasis model, and it was determined to be non-toxic when administered orally, showed no mutagenic activity, was equally distributed across the tissues and reduced the parasite load in the spleen (71.6%) and in the liver (89.2%) (Corpas-López et al., 2015).
The effect of (-)-α-bisabolol was also analyzed in L. tropica and L. major. Against intracellular amastigotes, the compound demonstrated IC50 values of 25.2 μM for L. tropica and 33.7 μM for L. major with selectivity indices of 46 and 34, respectively. As a mechanism of action, (-)-α-bisabolol was able to increase ROS levels and decrease the mitochondrial membrane potential and phosphatidylserine exposure. In addition, in an ultrastructural analysis, the compound was capable of inducing mitochondrial disruption and chromatin condensation, indicating apoptosis (Corpas-López et al., 2016a). This mechanism of action was also observed in L. amazonensis and L. infantum. (-)-α-Bisabolol induced phosphatidylserine externalization and caused plasmatic membrane damage, both of which are apoptosis indicators. The compound also decreased ATP levels and disrupted the mitochondrial membrane potential (Hajaji et al., 2018), supporting the hypothesis that the possible mechanism of action for (-)-α-bisabolol is inducing programmed cell death.
(-)-α-Bisabolol was also analyzed in vivo against a murine model of cutaneous leishmaniasis using L. tropica. Topical formulation was capable of reducing the lesion size and parasite burden (Corpas-López et al., 2016b). (-)-α-Bisabolol was employed in a different approach in a preclinical trial for a canine leishmaniasis model using naturally infected dogs. The dogs were divided into two groups, treated with meglumine antimoniate (100 mg/kg/day) subcutaneously or (-)-α-bisabolol (30 mg/kg/day) orally. The sesquiterpene reduced the parasite load in analyzed tissues, increasing INF-γ levels without any toxicity. An evaluation of cytokines and antibodies suggests a Th1 response induced by the compound, indicating an anti-inflammatory pathway (Corpas-López et al., 2018).
Oleanolic acid and its isomer, ursolic acid (triterpenoids), were studied in promastigotes and intracellular amastigotes of L. amazonensis. Oleanolic acid did not demonstrate activity against promastigotes, but when its isomer was tested against promastigotes, an IC50 value of 6.2 μg/mL was obtained. Ursolic acid in promastigotes of L. amazonensis induced programmed cell death independent of caspase 3/7 but dependent on mitochondria. When the in vivo assay was performed for cutaneous leishmaniasis, the compound reduced the lesion size and parasite load (Yamamoto et al., 2015).
To evaluate the effects of ursolic acid in the in vivo model of visceral leishmaniasis, female golden hamsters were infected with Leishmania infantum promastigotes. Two different doses of ursolic acid were employed, that is, 1 and 2 mg/kg/day, injected intraperitoneally for 15 days. Both doses were able to reduce the parasite load in the liver (over 96% reduction) and spleen (over 92% reduction). Histopathological analysis of the spleen indicated fewer parasites compared to the infected untreated control, and both white and red pulp were conserved by ursolic acid treatment, which was corroborated by INF-γ, IL-4, and IL-10 gene expression and splenic cell proliferation. Ursolic acid did not affect toxicological parameters (Jesus et al., 2017).
Toxicity, high cost, resistance, and reduced bioavailability are current challenges facing leishmaniasis chemotherapy. Nanotechnology has been reported as a promising alternative (Shah and Gupta, 2019). To improve ursolic acid (UA) use for leishmaniasis, a UA-loaded N-octyl-chitosan surface-decorated nanostructured lipid carrier system (UA-NLC) was tested against wild-type L. donovani and sodium stibogluconate (SSG-R) and paromomycin (PMM-R) L. donovani-resistant axenic amastigotes. UA-NLC exhibited IC50 values of 0.12, 1.07, and 3.51 μM for wild-type parasites, SSG-R and PMM-R, respectively, which were lower than those of regular ursolic acid (IC50 = 1.82, 16.15, and 36 μM, respectively). Against intracellular amastigotes, UA-NLC exhibited an IC50 of 0.09, 2.87, and 5.57 μM, and UA demonstrated an IC50 of 1.08, 11.54, and 24.46 μM for wild type, SSG-R and PMM-R, respectively (Das et al., 2017).
In the cytotoxicity evaluation, UA showed a selectivity index of 227.78, and UA-NLC showed an SI of 9111.11, almost 40 times higher. The effect of UA-NLC and regular UA in vivo, both administered orally at 10 mg/kg, was assessed. Spleen amastigote suppression was evaluated for wild-type, SSG-R- and PMM-R-infected mice. UA exhibited a percentage of suppression of 68.14, 64.69, and 59.55%, respectively, while UA-NLC achieved better results with 98.75, 88.4, and 90.37%, respectively. All these results suggest that UA-NLC and nanodelivery systems are a promising approach for leishmaniasis chemotherapy (Das et al., 2017).
The Antarctic sponge Dendrilla membranosa and other similar species have been a rich source for chemical studies. Dendrilla antarctica sponges, as named by Shilling et al. (2020), exhibit a variety of diterpenes in their composition, such as aplysulphurin (1), tetrahydroaplysulphurin-1 (2), membranolide (3), and darwinolide (4). These compounds were evaluated against Leishmania donovani-infected macrophages and J774 cells for cytotoxicity assays. Compounds 1 and 2 had the most promising IC50 values, 3.1 and 3.5 μM, respectively, while 3 and 4 exhibited values above 10 μM. However, the selective index for compound 1 was less than 10, while for compounds 2, 3, and 4, it was higher than 30 (Shilling et al., 2020).
Plumarella delicatissima is an octocoral specimen of the Southern Ocean known as a source of bioactive terpenoids. Seven terpenoids, (keikipukalide A-E, pukalide aldehyde, and ineleganolide), were isolated from Plumarella sp. and analyzed against L. donovani amastigote. Pukalide aldehyde was the most promising compound, exhibiting an IC50 value of 1.9 μM. X-ray crystallography of all isolated terpenoids indicates the differences between Pulmarella sp. terpenoid chemical structure, which is an important first step in structure-activity studies that should be conducted in further research (Thomas et al., 2018).
Using a novel and different approach, Ogungbe and Setzer (2013) studied potential in silico targets for terpenoids in Leishmania. Unlike in vitro and in vivo approaches, in silico approaches are able to predict different aspects of a potential bioactive compound, such as its possible targets, best radicals or chemical structure, for better antileishmanial activity. Molecular docking analysis was performed for several known antiparasitic plant-derived terpenoids. Each terpenoid class studied was docked with each chosen Leishmania species (L. infantum, L. Mexicana, L. major, and L. donovani). The most promising targets were nicotinamidase (L. infantum), uridine diphosphate-glucose pyrophosphorylase, methionyl t-RNA synthetase and dihydroorotate dehydrogenase (L. major), glycerol-3-phosphate dehydrogenase (L. mexicana) for the matching terpenoid classes. Furthermore, these results may help to guide new research on the development of new potent antileishmanial terpenoids (Ogungbe and Setzer, 2013).
The terpenoids present in this section with a defined IC50 are summarized in the Table 8.
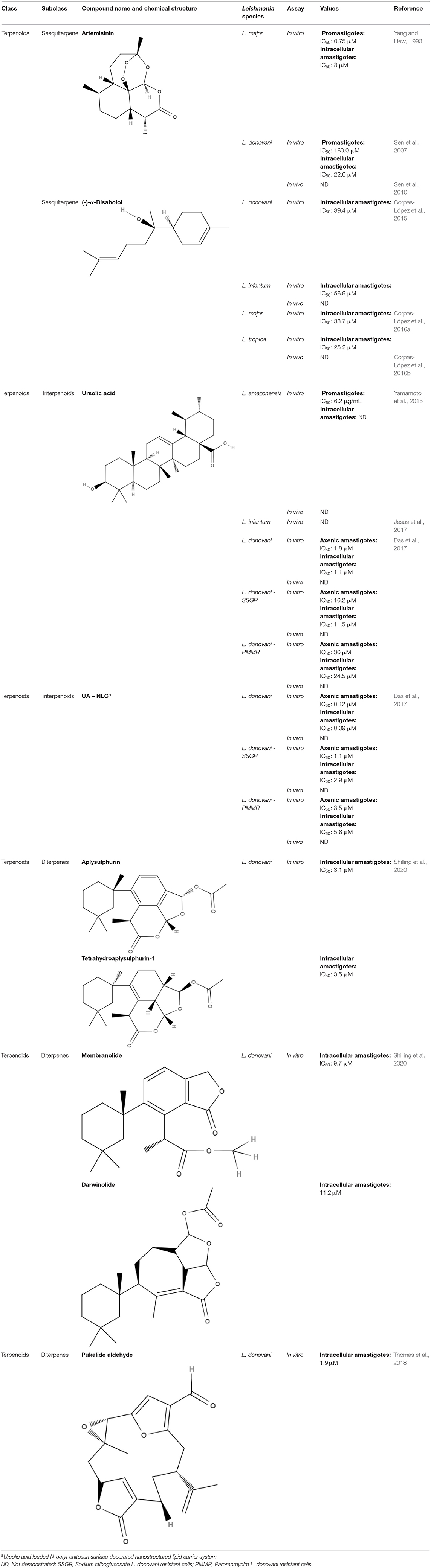
Table 8. Chemical structure and leishmanicidal activities of terpenoids.
Although leishmaniases are a group of diseases that have drug-based treatments, it remains a major challenger to research fields, since the currently available drug arsenal is reduced relative to the number of species that cause these diseases. Moreover, the chemotherapy that is utilized demands patient hospitalization and has been administered for many years, which can cause the occurrence of resistance and therapeutic failure. Therefore, research on new drugs for the treatment of these diseases is necessary.
The drug discovery process includes many steps to choose a new drug to treat a specific disease, and this process is expensive and time-consuming. Therefore, many strategies have been developed to optimize time and money.
There are 3 strategies more widely used in the drug discovery process:
(1) Fragment drug discovery based on molecules built for purpose. In this type of approach, automated techniques are used to trial compounds (i.e., high-throughput crystallography) to identify and optimize small molecules that bind to their target proteins with a variety of binding affinities (i.e., surface plasmon resonance). Hydrogen/deuterium exchange coupled with mass spectrometry (HDX-MS) and fragment libraries are techniques that play essential roles in this strategy. HDX-MS is a well-suited approach for investigating the alterations in protein conformation induced by small molecule ligand binding (Marciano et al., 2014). The fragment libraries identify smaller compounds, the “fragments,” which bind to different parts of a biological target. The primary rationale is that the identified hits provide access to a broader chemical space while screening a limited number of compounds (Schulz et al., 2011).
(2) Target direct screening is a biochemical approach based on repurposing or modifying existing molecules. In this approach, gene family platforms, compound libraries, computational models/informatics, structural biology and cellular and biochemical assays are extensively used to assess whether an existing molecule can be redirected as a treatment for the disease of interest (Lage et al., 2018). Some disadvantages of target direct screening, such as drug discovery and biochemical approach fragments for trypanosomatids, are the scarcity of fully validated drug targets and the need for additional screening to avoid off-target effects (Lage et al., 2018).
(3) Phenotypic drug discovery is a 'physiologically relevant' biological system or cellular signaling pathway that is directly investigated by chemical approaches to identify biologically active compounds. In contrast to the target-based strategies, these methods do not rely on knowledge of the identity of a specific drug target or a hypothesis about its role in disease. In this type of screening, advanced methodologies are able to answer many questions about a specific organic system, such as high content imaging, advanced informatics, advanced cellular assays, stem cells, SCORE, in vivo imaging, and the use of zebrafish models (Moffat et al., 2017).
The most widely employed methods for drug discovery against kinetoplastids are phenotypic, which is entirely justified, since the parasites have complex life cycles (the same parasite has different hosts and different forms in response to different temperatures and pH). The action of a compound on the parasite depends on its stage of life. As the drug discovery process can be developed at any stage of the parasite's life, the results may vary widely. Generally, in the case of trypanosomatids, the infective form of the parasite in the mammalian host is chosen to perform the drug discovery process; in the case of Leishmania sp., it is the intracellular amastigote (Lage et al., 2018).
In this review, the extensive models utilized for the drug discovery process employ dye-based indicators of parasite viability, such as resazourin, which is used for testing the drug susceptibilities of parasites. To evaluate the mechanism of action, methodologies are generally used to measure the levels of reactive oxygen species and the type of cell death triggered by the test compound.
Recent advances in automated microscopy have the capacity to increase throughput by replacing laborious manual microscopic observations with high-content imaging, and this technique is being successfully utilized in in vitro whole-organism screening against live kinetoplastid parasites (Siqueira-Neto et al., 2012). Bioinformatic tools that predict the potential of the compound as a drug, such as Linpinki's rules, along with other tools that predict the interaction of this compound with proteins (docking assays), provide robust data to assist in choosing cell-based assays to perform and enable the exclusion of certain compounds if they do not exhibit good results in silico.
Despite all the advantages of automated microscopy, it is important to note that there are some limitations that still need to be overcome; for example, the complex life cycles of trypanosomatids are challenging to reproduce in the laboratory, and effectiveness in one parasitic stage does not guarantee a strong in vivo effect (Lage et al., 2018).
Most drug discovery screenings for anti-kinetoplastid drugs are performed with synthetic compound libraries to search for active compounds, but such synthetic libraries are often limited in structural diversity and novelty (Fox et al., 2006). Natural products may be a solution to this problem, because as was extensively discussed throughout this review, in addition to having several biological activities, they have underexplored chemical entities that may be employed as templates for the synthesis of new drugs.
Research on natural products has been increasingly conducted over the years, as these products may represent an alternative treatment of leishmaniasis and may be considered potential chemotherapy agents, since they demonstrate promising results against Leishmania spp. All plant metabolic pathways described in this review showed strong activity against several species of Leishmania in vitro, and most importantly, some compounds showed activity in vivo using visceral and cutaneous leishmaniasis models of infection, which were often better than the results presented by the reference drugs.
Concerning the mechanism of action of these natural products, several compounds are capable of altering the mitochondrial membrane potential, causing an increase in intracellular ROS levels and a decrease in ATP concentration and leading to programmed cell death. Furthermore, using molecular docking approaches, some molecules were capable of interacting with important enzymes for the redox homeostasis of Leishmania, such as trypanothione reductase and trypanothione synthetase; however, it is important to demonstrate the inhibition of these activities by the selected molecules using recombinant enzymes. In vivo, some natural compounds were observed to reduce the parasite load and to act as immunomodulators.
In this scenario, it is possible that these compounds may be employed as a source of new treatments for leishmaniasis in the future, adding to the treatments already administered in the clinic.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This work was supported by Fundação Carlos Chagas Filho de Amparo a Pesquisa do Estado do Rio de Janeiro (FAPERJ), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Programa Estratégio de Apoio a Pesquisa em Saúde (PAPES/FIOCRUZ;), and the Fundação Oswaldo Cruz (FIOCRUZ). EA-A was the recipient of a research scholarship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
2HF, 2′-Hydroxyflavanone; ABC, ATP-binding cassette; AEPA, Aqueous extract of Physalis angulata root; AP-1, Activator protein 1; ATP, Adenosine triphosphate; BPQ, Buparvaquone; C, Carbon; CC50, Concentration that promotes 50% cytotoxicity; DHDE, Dehydrodieuginol; DNA, Deoxyribonucleic acid; DNDi, Drugs for Neglected Diseases Initiative; ED50, Median effective dose; EEPa, Ethanolic extract of Physalis angulate; EGCG, (–)-Epigallocatechin 3-O-gallate; FDA, Food and Drug Administration; GFP, Green fluorescent protein; HPLC, High-performance liquid chromatography; IC50, Half maximal inhibitory concentration; iNOS, Nitric oxide synthase inducible; LD50, Median lethal dose; MFI, Mean fluorescence intensity; mRNA, Messenger ribonucleic acid; MuEO, Myracrodruon urundeuva essential oil; NF-κB, Nuclear factor kappa-light-chain-enhancer of activated B cells; NO, Nitric oxide; PBS, Phosphate saline buffer; ROS, Reactive oxygen species; Sbv, Pentavalent antimony; SI, Selectivity index; ∑FIC, Fractional inhibitory concentration sum; Th1, Helper T cell 1; Th2, Helper T cell 2; TIC, Total ion chromatogram; TrEO, Tetradenia riparia essential oil; TrROY, 6,7-Dehydroroyleanone; WHO, World Health Organization.
Azadbakht, M., Davoodi, A., Hosseinimehr, S. J., Keighobadi, M., Fakhar, M., Valadan, R., et al. (2020). Tropolone alkaloids from Colchicum kurdicum (Bornm.) Stef. (Colchicaceae) as the potent novel antileishmanial compounds; purification, structure elucidation, antileishmanial activities and molecular docking studies. Exp. Parasitol. 213:107902. doi: 10.1016/j.exppara.2020.107902
Bortoleti, B. T. D. S., Tomiotto-Pellissiera, F., Gonçalves, M. D., Miranda-Sapla, M. M., Assolini, J. P., Carloto, A. C., et al. (2019). Caffeic acid has antipromastigote activity by apoptosis-like process; and anti-amastigote by TNF-α/ROS/NO production and decreased of iron availability. Phytomedicine 57, 262–270. doi: 10.1016/j.phymed.2018.12.035
Brenzan, M. A., Nakamura, C. V., Prado Dias Filho, B., Ueda-Nakamura, T., Young, M. C., and Aparício Garcia Cortez, D. (2007). Antileishmanial activity of crude extract and coumarin from Calophyllum brasiliense leaves against Leishmania amazonensis. Parasitol. Res. 101, 715–722. doi: 10.1007/s00436-007-0542-7
Brenzan, M. A., Santos, A. O., Nakamura, C. V., Filho, B. P., Ueda-Nakamura, T., Young, M. C., et al. (2012). Effects of (-) mammea A/BB isolated from Calophyllum brasiliense leaves and derivatives on mitochondrial membrane of Leishmania amazonensis. Phytomedicine 19, 223–230. doi: 10.1016/j.phymed.2011.10.008
Brezan, M. A., Nakamura, C. V., Filho, B. P. D., Ueda-Nakamura, T., Young, M. C., Córrea, A. G., et al. (2008). Structure-activity relationship of (-) mammea A/BB derivatives against Leishmania amazonensis. Biomed. Pharmacother. 62, 651–658. doi: 10.1016/j.biopha.2008.08.024
Brito, J. R., Passero, L. F. D., Bezerra-Souza, A., Laurenti, M. D., Romoff, P., Barbosa, H., et al. (2019). Antileishmanial activity and ultrastructural changes of related tetrahydrofuran dineolignans isolated from Saururus cernuus L. (Saururaceae). J. Pharm. Pharmacol. 12, 1871–1878. doi: 10.1111/jphp.13171
Burza, S., Croft, S. L., and Boelaert, M. (2018). Leishmaniasis. Lancet 392, 951–970. doi: 10.1016/S0140-6736(18)31204-2
Cartuche, L., Sifaoui, I., López-Arencibia, A., Bethencourt-Estrella, C. J., San Nicolás-Hernández, D., Lorenzo-Morales, J., et al. (2020). Antikinetoplastid activity of indolocarbazoles from Streptomyces sanyensis. Biomolecules 10:657. doi: 10.3390/biom10040657
Chowdhury, S., Mukherjee, T., Mukhopadhyay, R., Mukherjee, B., Sengupta, S., Chattopadhyay, S., et al. (2012). The lignan niranthin poisons Leishmania donovani topoisomerase IB and favours a Th1 immune response in mice. EMBO Mol. Med. 4, 1126–1143. doi: 10.1002/emmm.201201316
Corpas-López, V., Merino-Espinosa, G., Acedo-Sánchez, C., Díaz-Sáez, V., Navarro-Moll, M. C., Morillas-Márquez, F., et al. (2018). Effectiveness of the sesquiterpene (-)-α-bisabolol in dogs with naturally acquired canine leishmaniosis: an exploratory clinical trial. Vet. Res. Commun. 42, 121–130. doi: 10.1007/s11259-018-9714-4
Corpas-López, V., Merino-Espinosa, G., Díaz-Sáez, V., Morillas-Márquez, F., Navarro-Moll, M. C., and Martín-Sánchez, J. (2016a). The sesquiterpene (–)-α-bisabolol is active against the causative agents of Old World cutaneous leishmaniasis through the induction of mitochondrial-dependent apoptosis. Apoptosis 21, 1071–1081. doi: 10.1007/s10495-016-1282-x
Corpas-López, V., Merino-Espinosa, G., López-Viota, M., Gijón-Robles, P., Morillas-Mancilla, M. J., López-Viota, J., et al. (2016b). Topical treatment of Leishmania tropica infection using (-)-α-bisabolol ointment in a hamster model: effectiveness and safety assessment. J. Nat. Prod. 79, 2403–2407. doi: 10.1021/acs.jnatprod.6b00740
Corpas-López, V., Morillas-Márquez, F., Navarro-Moll, M. C., Merino-Espinosa, G., Díaz-Sáez, V., and Martín-Sánchez, J. (2015). (-)-α-bisabolol, a promising oral compound for the treatment of visceral leishmaniasis. J. Nat. Prod. 78, 1202–1207. doi: 10.1021/np5008697
Croft, S. L., Hogg, J., Gutteridge, W. E., Hudson, A. T., and Randall, A. W. (1992). The activity of hydroxynaphthoquinones against Leishmania donovani. J. Antimicrob. Chemother. 30, 827–832. doi: 10.1093/jac/30.6.827
Das, S., Ghosh, S., De, A. K., and Bera, T. (2017). Oral delivery of ursolic acid-loaded nanostructured lipid carrier coated with chitosan oligosaccharides: development, characterization, in vitro and in vivo assessment for the therapy of leishmaniasis. Int. J. Biol. Macromol. 102, 996–1008. doi: 10.1016/j.ijbiomac.2017.04.098
De Castro Oliveira, L. G., Brito, L. M., de Moraes Alves, M. M., Amorim, L. V., Sobrinho-Júnior, E. P., de Carvalho, C. E., et al. (2017). In vitro effects of the neolignan 2,3-dihydrobenzofuran against Leishmania amazonensis. Basic Clin. Pharmacol. Toxicol. 120, 52–58. doi: 10.1111/bcpt.12639
De Sarkar, S., Sarkar, D., Sarkar, A., Dighal, A., Staniek, K., Gille, L., et al. (2018). Berberine chloride mediates its antileishmanial activity by inhibiting Leishmania mitochondria. Parasitol. Res. 118, 335–345. doi: 10.1007/s00436-018-6157-3
Demarchi, I. G., Thomazella, M. V., de Souza Terron, M., Lopes, L., Gazim, Z. C., Cortez, D. A., et al. (2015). Antileishmanial activity of essential oil and 6,7-dehydroroyleanone isolated from Tetradenia riparia. Exp Parasitol. 157:128–137. doi: 10.1016/j.exppara.2015.06.014
Dewick, P. M. (ed.). (2009). “The shikimate pathway: aromatic amino acids and phenylpropanoids,” in Medicinal Natural Products. New Jersey: John Wiley & Sons, 137–186. doi: 10.1002/9780470742761.ch4
Di Giorgio, C., Delmas, F., Akhmedjanova, V., Ollivier, E., Bessonova, I., Riad, E., et al. (2005). In vitro antileishmanial activity of diphyllin isolated from Haplophyllum bucharicum. Planta Med. 71, 366–369. doi: 10.1055/s-2005-864106
Dimmer, J., Cabral, F. V., Sabino, C. P., Silva, C. R., Núñez-Montoya, S. C., Cabrera, J. L., et al. (2019). Natural anthraquinones as novel photosentizers for antiparasitic photodynamic inactivation. Phytomedicine 61:152894. doi: 10.1016/j.phymed.2019.152894
DNDi (2016). Available online at: https://www.dndi.org/diseases-projects/leishmaniasis/ (accessed December, 2016).
Durak, S., Arasoglu, T., Ates, S. C., and Derman, S. (2020). Enhanced antibacterial and antiparasitic activity of multifunctional 2 polymeric nanoparticles. Nanotechnology 31:5705. doi: 10.1088/1361-6528/ab6ab9
Emiliano, Y. S. S., and Almeida-Amaral, E. E. (2018). Efficacy of apigenin and miltefosine combination therapy against experimental cutaneous leishmaniasis. J. Nat. Prod. 81, 1910–1913. doi: 10.1021/acs.jnatprod.8b00356
Ferreira, M. E., Rojas de Arias, A., Yaluff, G., de Bilbao, N. V., Nakayama, H., Torres, S., et al. (2010). Antileishmanial activity of furoquinolines and coumarins from Helietta apiculata. Phytomedicine 17, 375–378. doi: 10.1016/j.phymed.2009.09.009
Ferreira, T. N., Pita-Pereira, D., Costa, S. G., Brazil, R. P., Moraes, C. S., Díaz-Albiter, H. M., et al. (2018). Transmission blocking sugar baits for the control of Leishmania development inside sand flies using environmentally friendly beta-glycosides and their aglycones. Parasit. Vectors 11:614. doi: 10.1186/s13071-018-3122-z
Flores, N., Cabrera, G., Jiménez, I. A., Piñero, J., Giménez, A., Bourdy, G., et al. (2007). Leishmanicidal constituents from the leaves of Piper rusbyi. Planta Med. 73, 206–211. doi: 10.1055/s-2007-967123
Fonseca-Silva, F., Canto-Cavalheiro, M. M., Menna-Barreto, R. F. S., and Almeida-Amaral, E. E. (2015). Effect of apigenin on Leishmania amazonensis is associated with reactive oxygen species production followed by mitochondrial dysfunction. J. Nat. Prod. 78, 880–884. doi: 10.1021/acs.jnatprod.5b00011
Fonseca-Silva, F., Inacio, J. D. F., Canto-Cavalheiro, M. M., and Almeida-Amaral, E. E. (2013). Reactive oxygen species production by quercetin causes the death of Leishmania amazonensis intracellular amastigotes. J. Nat. Prod. 76, 1505–1508. doi: 10.1021/np400193m
Fonseca-Silva, F., Inacio, J. D. F., Canto-Cavalheiro, M. M., Menna-Barreto, R. F., and Almeida-Amaral, E. E. (2016). Oral efficacy of apigenin against cutaneous leishmaniasis: involvement of reactive oxygen species and autophagy as a mechanism of action. PLoS Negl. Trop. Dis 10:e0004442. doi: 10.1371/journal.pntd.0004442
Fox, S., Farr-Jones, S., Sopchak, L., Boggs, A., Nicely, H. W., Khoury, R., et al. (2006). High-throughput screening: update on practices and success. J. Biomol. Screen 11, 864–869. doi: 10.1177/1087057106292473
Garcia, A. R., Oliveira, D. M. P., Amaral, A. C. F., Jesus, J. B., Rennó Sodero, A. C., Souza, A. M. T., et al. (2019). Leishmania infantum arginase: biochemical characterization and inhibition by naturally occurring phenolic substances. J. Enzyme Inhib. Med. Chem. 34, 1100–1109. doi: 10.1080/14756366.2019.1616182
Garnier, T., Mäntylä, A., Järvinen, T., Lawrence, J., Brown, M., and Croft, S. (2007). In vivo studies on the antileishmanial activity of buparvaquone and its prodrugs. J. Antimicrob. Chemother. 60, 802–810. doi: 10.1093/jac/dkm303
Gervazoni, L. F. O., Gonçalves-Ozório, G., and Almeida-Amaral, E. E. (2018). 2′-Hydroxyflavanone activity in vitro and in vivo against wild-type and antimony-resistant Leishmania amazonensis. PLoS Negl. Trop. Dis. 12: e0006930. doi: 10.1371/journal.pntd.0006930
Gundampati, R. K., Sahu, S., Shukla, A., Pandey, R. K., Patel, M., Banik, R. M., et al. (2014). Tryparedoxin peroxidase of Leishmania braziliensis: homology modeling and inhibitory effects of flavonoids for anti-leishmanial activity. Bioinformation 10, 353–357. doi: 10.6026/97320630010353
Hajaji, S., Sifaoui, I., López-Arencibia, A., Reyes-Batlle, M., Jiménez, I. A., Bazzocchi, I. L., et al. (2018). Leishmanicidal activity of α-bisabolol from Tunisian chamomile essential oil. Parasitol. Res. 117, 2855–2867. doi: 10.1007/s00436-018-5975-7
Inacio, J. D. F., Canto-Cavalheiro, M. M., and Almeida-Amaral, E. E. (2013). In vitro and in vivo effects of (-)-epigallocatechin 3- O -gallate on Leishmania amazonensis. J. Nat. Prod. 76, 1993–1996. doi: 10.1021/np400624d
Inacio, J. D. F., Fonseca, M. S., and Almeida-Amaral, E. E. (2019). (-)-Epigallocatechin 3-O-gallate as a new approach for the treatment of visceral leishmaniasis. J. Nat. Prod. 82, 2664–2667. doi: 10.1021/acs.jnatprod.9b00632
Inacio, J. D. F., Gervazoni, L., Canto-Cavalheiro, M. M., and Almeida-Amaral, E. E. (2014). The effect of (-)-Epigallocatechin 3-O - gallate in vitro and in vivo in Leishmania braziliensis: involvement of reactive oxygen species as a mechanism of action. PLoS Negl. Trop. Dis. 8:e3093. doi: 10.1371/journal.pntd.0003093
Ioset, J.-R. (2008). Natural products for neglected diseases: a review. Curr. Org. Chem. 12, 643–666. doi: 10.2174/138527208784577394
Jain, P. K., and Joshi, H. (2012). Coumarin: chemical and pharmacological profile. J. Appl. Pharm. Sci. 2, 236–240. doi: 10.7324/JAPS.2012.2643
Jesus, J. A., Fragoso, T. N., Yamamoto, E. S., Laurenti, M. D., Silva, M. S., Ferreira, A. F., et al. (2017). Therapeutic effect of ursolic acid in experimental visceral leishmaniasis. Int. J. Parasitol. Drugs Drug Resist. 7, 1–11. doi: 10.1016/j.ijpddr.2016.12.002
Kashyap, D., Sharma, A., Tuli, H. S., Sak, K., Garg, V. K., Buttar, H. S., et al. (2018). Apigenin: a natural bioactive flavone-type molecule with promising therapeutic function. J. Funct. Foods 48, 457–471 doi: 10.1016/j.jff.2018.07.037
Kumar, A., Saha, B., and Singh, S. (2017). Dataset generated for dissection of mechanisms of trypanothione reductase and tryparedoxin peroxidase through dynamic network analysis and simulations in leishmaniasis. Data Brief. 15, 757–769. doi: 10.1016/j.dib.2017.10.031
Kurek, J. (ed.). (2019). Introductory Chapter: Alkaloids - Their Importance in Nature and for Human Life, Alkaloids - Their Importance in Nature and Human Life. IntechOpen. Available online at: https://www.intechopen.com/books/alkaloids-their-importance-in-nature-and-human-life/introductory-chapter-alkaloids-their-importance-in-nature-and-for-human-life
Lacerda, R. B. M., Freitas, T. R., Martins, M. M., Teixeira, T. L., da Silva, C. V., Candido, P. A., et al. (2018). Isolation, leishmanicidal evaluation and molecular docking simulations of piperidine alkaloids from Senna spectabilis. Bioorg. Med. Chem. 26, 5816–5823. doi: 10.1016/j.bmc.2018.10.032
Lage, O. M., Ramos, M. C., Calisto, R., Almeida, E., Vasconcelos, V., and Vicente, F. (2018). Current screening methodologies in drug discovery for selected human diseases. Mar. Drugs 16:279. doi: 10.3390/md16080279
Lezama-Dávila, C. M., McChesney, J. D., Bastos, J. K., Miranda, M. A., Tiossi, R. F., da Costa J. C., et al. (2016). A new antileishmanial preparation of combined solamargine and solasonine heals cutaneous leishmaniasis through different immunochemical pathways. Antimicrob. Agents Chemother. 60, 2732–2738. doi: 10.1128/AAC.02804-15
Lizzi, F., Veronesi, G., Belluti, F., Bergamini, C., López-Sánchez, A., Kaiser, M., et al. (2012). Conjugation of quinones with natural polyamines: toward an expanded antitrypanosomatid profile. J. Med. Chem. 55, 10490–10500. doi: 10.1021/jm301112z
Mahalakshmi, A. M., and Nidavani, R. B. (2014). Physalis angulata L.: an ethanopharmacological review. Indo Am. J. Pharm. Res. 4, 1479–1486. doi: 10.1044/1980-iajpr.14337
Maia, M. S., Silva, J. P. R., Nunes, T. L., Sousa, J. M. S., Rodrigues, G. C. S., Monteiro, A. F. M., et al. (2020). Virtual screening and the in vitro assessment of the antileishmanial activity of lignans. Molecules 25:2281. doi: 10.3390/molecules25102281
Manjolin, L. C., dos Reis, M. B., do Maquiaveli, C. C., Santos-Filho, O. A., and da Silva, E. R. (2013). Dietary flavonoids fisetin, luteolin and their derived compounds inhibit arginase, a central enzyme in Leishmania (Leishmania) amazonensis infection. Food Chem. 141, 2253–2262. doi: 10.1016/j.foodchem.2013.05.025
Mäntylä, A., Garnier, T., Rautio, J., Nevalainen, T., Vepsälainen, J., Koskinen, A., et al. (2004). Synthesis, in vitro evaluation, and antileishmanial activity of water-soluble prodrugs of buparvaquone. J. Med. Chem. 47, 188–195. doi: 10.1021/jm030868a
Marciano, D. P., Dharmarajan, V., and Griffin, P. R. (2014). HDX-MS guided drug discovery: small molecules and biopharmaceuticals. Curr. Opin. Struct. Biol. 28, 105–111. doi: 10.1016/j.sbi.2014.08.007
Meanwell, N. A. (2011). Improving drug candidates by design: a focus on physicochemical properties as a means of improving compound disposition and safety. Chem. Res. Toxicol. 24, 1420–1456. doi: 10.1021/tx200211v
Mehwish, S., Khan, H., Rehman, A. U., Khan, A. U., Khan, M. A., Hayat, O., et al. (2019). Natural compounds from plants controlling leishmanial growth via DNA damage and inhibiting trypanothione reductase and trypanothione synthetase: an in vitro and in silico approach. 3 Biotech 9:303. doi: 10.1007/s13205-019-1826-1
Mercado-Camargo, J., Cervantes-Ceballos, L., Vivas-Reyes, R., Pedretti, A., Serrano-García, M. L., and Gómez-Estrada, H. (2020). Homology modeling of leishmanolysin (gp63) from Leishmania panamensis and molecular docking of flavonoids. ACS Omega 5, 14741–14749. doi: 10.1021/acsomega.0c01584
Moffat, J., Vincent, F., Lee, J., Eder, J., and Prunotto, M. (2017). Opportunities and challenges in phenotypic drug discovery: an industry perspective. Nat. Rev. Drug Discov. 16, 531–543. doi: 10.1038/nrd.2017.111
Monteiro, L. M., Löbenberg, R., Fotaki, N., de Araújo, G. L. B., Cotrim, P. C., and Bou-Chacra, N. (2019). Co-delivery of buparvaquone and polymyxin B in a nanostructured lipid carrier for leishmaniasis treatment. J. Glob. Antimicrob. Resist. 8, 279–283. doi: 10.1016/j.jgar.2019.06.006
Montrieux, E., Perera, W. H., García, M., Maes, L., Cos, P., and Monzote, L. (2014). In vitro and in vivo activity of major constituents from Pluchea carolinensis against Leishmania amazonensis. Parasitol. Res. 113, 2925–2932. doi: 10.1007/s00436-014-3954-1
Morales-Jadán, D., Blanco-Salas, J., Ruiz-Téllez, T., and Centeno, F. (2020). Three alkaloids from an apocynaceae species, Aspidosperma spruceanum as antileishmaniasis agents by in silico demo-case studies. Plants 9:E983. doi: 10.3390/plants9080983
Muzitano, M. F., Falcão, C. A. B., Cruz, E. A., Bergonzi, M. C., Bilia, A. R., Vincieri, F. F., et al. (2009). Oral metabolism and efficacy of Kalanchoe pinnata flavonoids in a murine model of cutaneous leishmaniasis. Planta Med. 75, 307–311. doi: 10.1055/s-0028-1088382
Nogueira, R. C., Rocha, V. P. C., Nonato, F. R., Tomassini, T. C., Ribeiro, I. M., dos Santos, R. R., et al. (2013). Genotoxicity and antileishmanial activity evaluation of Physalis angulata concentrated ethanolic extract. Environ. Toxicol. Pharmacol. 36, 1304–1311. doi: 10.1016/j.etap.2013.10.013
Ogungbe, I. V., and Setzer, W. N. (2013). In-silico Leishmania target selectivity of antiparasitic terpenoids. Molecules 18, 7761–7847. doi: 10.3390/molecules18077761
Pereira, I., Mendonça, D., Tavares, G., Lage, D. P., Ramos, F. F., and Oliveira-da-Silva, J. A., et al. (2020). Parasitological and immunological evaluation of a novel chemotherapeutic agent against visceral leishmaniasis. Parasite Immunol. e12784. doi: 10.1111/pim.12784. [Epub ahead of print].
Rahimi, M., Seyyed Tabaei, S. J., Ziai, S. A., and Sadri, M. (2020). Anti-leishmanial effects of chitosan-polyethylene oxide nanofibers containing berberine: an applied model for leishmania wound dressing. Iran. J. Med. Sci. 45, 286–297. doi: 10.30476/IJMS.2019.45784
Randhawa, G. K., and Kullar, J. S., and Rajkumar (2011). Bioenhancers from mother nature and their applicability in modern medicine. Int. J. Appl. Basic Med. Res. 1, 5–10. doi: 10.4103/2229-516X.81972
Ray, L., Karthik, R., Srivastava, V., Singh, S. P., Pant, A. B., Goyal, N., et al. (2020). Efficient antileishmanial activity of amphotericin B and piperine entrapped in enteric coated guar gum nanoparticles. Drug Deliv. Transl. Res. doi: 10.1007/s13346-020-00712-9
Ribeiro, T. G., Chávez-Fumagalli, M. A., Valadares, D. G., Franca, J. R., Lage, P. S., Duarte, M. C., et al. (2014). Antileishmanial activity and cytotoxicity of Brazilian plants. Exp Parasitol. 143, 60–68. doi: 10.1016/j.exppara.2014.05.004
Rizk, Y. S., Fischer, A., Cunha M de, C., Rodrigues, P. O., Marques, M. C., Matos, M. F., et al. (2014). In vitro activity of the hydroethanolic extract and biflavonoids isolated from Selaginella sellowii on Leishmania (Leishmania) amazonensis. Mem. Inst. Oswaldo Cruz 109, 1050–1056. doi: 10.1590/0074-0276140312
Rocha, V. P. C., Quintino da Rocha, C., Ferreira Queiroz, E., Marcourt, L., Vilegas, W., and Grimaldi, G. B., et al. (2018). Antileishmanial activity of dimeric flavonoids isolated from Arrabidaea brachypoda. Molecules. 24:1. doi: 10.3390/molecules24010001
Rodrigues, L. C., Barbosa-Filho, J. M., de Oliveira, M. R., do Nascimento Néris, P. L., Borges, F. V., and Mioso, R. (2016). Synthesis and antileishmanial activity of natural dehydrodieugenol and its mono- and dimethyl ethers. Chem. Biodivers. 13, 870–874. doi: 10.1002/cbdv.201500280
Rye, C. E., and Barker, D. (2013). Asymmetric synthesis and anti-protozoal activity of the 8,40-oxyneolignans virolin, surinamensin and analogues. Eur. J. Med. Chem. 60, 240–248. doi: 10.1016/j.ejmech.2012.12.013
Saha, P., Sen, R., Hariharan, C., Kumar, D., Das, P., and Chatterjee, M. (2009). Berberine chloride causes a caspase-independent, apoptotic-like death in Leishmania donovani promastigotes. Free Radic. Res. 43, 1101–1110. doi: 10.1080/10715760903186124
Saha, S., Mukherjee, T., Chowdhury, S., Mishra, A., Chowdhury, S. R., Jaisankar, P., et al. (2013). The lignan glycosides lyoniside and saracoside poison the unusual type IB topoisomerase of Leishmania donovani and kill the parasite both in vitro and in vivo. Biochem. Pharmacol. 86, 1673–1687. doi: 10.1016/j.bcp.2013.10.004
Salem, M. M., Capers, J., Rito, S., and Werbovetz, K. A. (2011). Antiparasitic activity of C-geranyl flavonoids from Mimulus bigelovii. Phyther. Res. 25, 1246–1249. doi: 10.1002/ptr.3404
Schmidt, T. J., Khalid, S. A., Romanha, A. J., Alves, T. M., Biavatti, M. W., Brun, R., et al. (2012a). The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases - Part I. Curr. Med. Chem. 19, 2128–2175. doi: 10.2174/092986712800229023
Schmidt, T. J., Khalid, S. A., Romanha, A. J., Alves, T. M., Biavatti, M. W., Brun, R., et al. (2012b). The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases - Part II. Curr. Med. Chem. 19, 2176–2228. doi: 10.2174/092986712800229087
Schulz, M. N., Landstro, J. M., Bright, B., and Hubbard, R. E. (2011). Design of a Fragment Library that maximally represents available chemical space. J. Comput. Aided Mol. Des. 25, 611–620. doi: 10.1007/s10822-011-9461-x
Sen, R., Bandyopadhyay, S., Dutta, A., Mandal, G., Ganguly, S., Saha, P., et al. (2007). Artemisinin triggers induction of cell-cycle arrest and apoptosis in Leishmania donovani promastigotes. J. Med. Microbiol. 56, 1213–1218. doi: 10.1099/jmm.0.47364-0
Sen, R., Ganguly, S., Saha, P., and Chatterjee, M. (2010). Efficacy of artemisinin in experimental visceral leishmaniasis. Int. J. Antimicrob. Agents 36, 43–49. doi: 10.1016/j.ijantimicag.2010.03.008
Shah, A., and Gupta, S. S. (2019). Anti-leishmanial nanotherapeutics: a current perspective. Curr. Drug Metab. 20, 473–482. doi: 10.2174/1389200219666181022163424
Shah, N. A., Khan, M. R., and Nadhman, A. (2014). Antileishmanial, toxicity, and phytochemical evaluation of medicinal plants collected from Pakistan. Biomed. Res. Int. 2014:384204. doi: 10.1155/2014/384204
Sharma, N., Shukla, A. K., Das, M., and Dubey, V. K. (2012). Evaluation of plumbagin and its derivative as potential modulators of redox thiol metabolism of Leishmania parasite. Parasitol. Res. 110, 341–348. doi: 10.1007/s00436-011-2498-x
Shilling, A. J., Witowski, C. G., Maschek, J. A., Azhari, A., Vesely, B. A., Kyle, D. E., et al. (2020). Spongian diterpenoids derived from the antarctic sponge dendrilla antarctica are potent inhibitors of the leishmania parasite. J. Nat. Prod. 83, 1553–1562. doi: 10.1021/acs.jnatprod.0c00025
Silva, L. G., Gomes, K. S., Costa-Silva, T. A., Romanelli, M. M., Tempone, A. G., Sartorelli, P., et al. (2020). Calanolides E1 and E2, two related coumarins from Calophyllum brasiliense Cambess. (Clusiaceae), displayed in vitro activity against amastigote forms of Trypanosoma cruzi and Leishmania infantum. Nat. Prod. Res. 1478–6419. doi: 10.1080/14786419.2020.1765347. [Epub ahead of print]
Siqueira-Neto, J. L., Moon, S., Jang, J., Yang, G., Lee, C., Moon, H. K., et al. (2012). An image-based high-content screening assay for compounds targeting intracellular Leishmania donovani amastigotes in human macrophages. PLoS Negl. Trop. Dis. 6:e1671. doi: 10.1371/journal.pntd.0001671
Smith, L., Serrano, D. R., Mauger, M., Bolás-Fernández, F., Dea-Ayuela, M. A., and Lalatsa, A. (2018). Orally bioavailable and effective buparvaquone lipid-based nanomedicines for visceral leishmaniasis. Mol. Pharm. 15, 2570–2583. doi: 10.1021/acs.molpharmaceut.8b00097
Thomas, S. A. L., von Salm, J. L., Clark, S., Ferlita, S., Nemani, P., and Azhari, A., et al. (2018). Keikipukalides, furanocembrane diterpenes from the antarctic deep sea octocoral Plumarella delicatissima. J. Nat. Prod. 81, 117–123. doi: 10.1021/acs.jnatprod.7b00732
Tiuman, T. S., Brenzan, M. A., Ueda-Nakamura, T., Filho, B. P., Cortez, D. A., and Nakamura, C. V. (2012). Intramuscular and topical treatment of cutaneous leishmaniasis lesions in mice infected with Leishmania amazonensis using coumarin (-) mammea A/BB. Phytomedicine 19, 1196–1199. doi: 10.1016/j.phymed.2012.08.001
Touaibia, M., Jean-Francois, J., and Doiron, J. (2012). Caffeic acid, a versatile pharmacophore: an overview. Mini Rev. Med Chem 11, 695–713. doi: 10.2174/138955711796268750
Vianna, G. (1912). Comunicação à Sessão de 24 de abril de 1912 da Sociedade Brasileira de Dermatologia. Arch. Bras. Med. 1, 36–38.
Viegas, C., Da Silva Bolzani, V., and Barreiro, E. J. (2006). OS produtos naturais e a química medicinal moderna. Quim. Nova 29, 326–337. doi: 10.1590/S0100-40422006000200025
Vieira-Araújo, F. M., Macedo Rondon, F. C., Pinto Vieira, Í. G., Pereira Mendes, F. N., Carneiro de Freitas, J. C., and Maia de Morais, S. (2018). Sinergism between alkaloids piperine and capsaicin with meglumine antimoniate against Leishmania infantum. Exp. Parasitol. 188, 79–82. doi: 10.1016/j.exppara.2018.04.001
WHO (2016). Global Health Observatory Data Repository. World Health Organization. [Internet]. Available online at: http://apps.who.int/ (accessed July 15, 2016).
Winkel, B. S. J. (2006). “The biosynthesis of flavonoids,” in The Science of Flavonoids, ed E. Grotewold (Columbus, Ohio: Springer), 71–96.
Yamamoto, E. S., Campos, B. L. S., Jesus, J. A., Laurenti, M. D., Ribeiro, S. P., and Kallás, E. G., et al. (2015). The effect of ursolic acid on leishmania (Leishmania) amazonensis is related to programed cell death and presents therapeutic potential in experimental cutaneous leishmaniasis. PLoS ONE 10:e0144946. doi: 10.1371/journal.pone.0144946
Keywords: natural product, Leishmaniasis, chemotherapy, in vivo, in vitro, intracellular amastigotes
Citation: Gervazoni LFO, Barcellos GB, Ferreira-Paes T and Almeida-Amaral EE (2020) Use of Natural Products in Leishmaniasis Chemotherapy: An Overview. Front. Chem. 8:579891. doi: 10.3389/fchem.2020.579891
Received: 03 July 2020; Accepted: 07 October 2020;
Published: 23 November 2020.
Edited by:
Gildardo Rivera, National Polytechnic Institute of Mexico (IPN), MexicoReviewed by:
Marcus Scotti, Federal University of Paraíba, BrazilCopyright © 2020 Gervazoni, Barcellos, Ferreira-Paes and Almeida-Amaral. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elmo E. Almeida-Amaral, ZWxtb0Bpb2MuZmlvY3J1ei5icg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.