
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Chem. , 24 August 2016
Sec. Cellular Biochemistry
Volume 4 - 2016 | https://doi.org/10.3389/fchem.2016.00037
 Ying Sun1,2*
Ying Sun1,2* Pingbo Huang3,4,5*
Pingbo Huang3,4,5*Extracellular adenosine is a ubiquitous signaling molecule that modulates a wide array of biological processes. Recently, significant advances have been made in our understanding of A2B adenosine receptor (A2BAR). In this review, we first summarize some of the general characteristics of A2BAR, and then we describe the multiple binding partners of the receptor, such as newly identified α-actinin-1 and p105, and discuss how these associated proteins could modulate A2BAR's functions, including certain seemingly paradoxical functions of the receptor. Growing evidence indicates a critical role of A2BAR in cancer, renal disease, and diabetes, in addition to its importance in the regulation of vascular diseases, and lung disease. Here, we also discuss the role of A2BAR in cancer, renal disease, and diabetes and the potential of the receptor as a target for treating these three diseases.
Extracellular adenosine is a ubiquitous signaling molecule that modulates a wide array of biological processes. Most of the extracellular adenosine is derived from the release and metabolism of adenine nucleotides such as ATP following diverse stimuli, including mechanical stress, osmotic challenge, inflammation, and tissue injury (Dunwiddie et al., 1997; Fredholm et al., 2001a; Picher et al., 2003, 2004; Eckle et al., 2007; Grenz et al., 2007; Ohta and Sitkovsky, 2014; Ross et al., 2014; Fuentes and Palomo, 2015; Kowal et al., 2015; Borea et al., 2016; Covarrubias et al., 2016; Hamidzadeh and Mosser, 2016). Conversely, extracellular adenosine is eliminated mainly through two mechanisms: one, transport of adenosine back into the cell by nucleoside transporters; and two, deamination of adenosine to inosine by adenosine deaminase (ADA; Blackburn and Kellems, 1996) or phosphorylation of adenosine to AMP by adenosine kinase (Lloyd and Fredholm, 1995; Spychala et al., 1996). The combined actions of these adenosine generation and elimination mechanisms regulate extracellular adenosine levels, which range from 10 to 200 nM under homeostatic conditions but can be elevated to 10–100 μM in hypoxic or stressed environments (Fredholm, 2007).
The biological functions of extracellular adenosine are mediated by four subtypes of adenosine receptors (ARs), A1, A2A, A2B, and A3, each of which presents a unique pharmacological profile, tissue distribution, and effector coupling (Fredholm et al., 2001b). Among human ARs, A1AR, and A3AR share 49% sequence similarity and A2AAR and A2BAR share 59% similarity (Jacobson and Gao, 2006; Goblyos and Ijzerman, 2009).
Perhaps because A2BAR binds to adenosine with low affinity (EC50 = 24 μM; Beukers et al., 2000; Fredholm et al., 2001b, 2011a), A2BAR is frequently considered to represent a low-affinity version of A2AAR and to be of comparatively lesser physiological relevance. However, recent advances in pharmacological and molecular tools have allowed researchers to determine that A2BAR can be coupled to distinct intracellular signaling pathways and play physiological roles that differ from those of A2AAR (Yang et al., 2006, 2010a; Grenz et al., 2012a; Johnston-Cox et al., 2012; Koupenova et al., 2012; Eckle et al., 2013; Morello and Miele, 2014; Patel et al., 2014; Tak et al., 2014; Eisenstein et al., 2015; Tang et al., 2015; Vecchio et al., 2016). In this review, we discuss our current understanding of the cellular functions of A2BAR and their implications for the pathogenesis of several human diseases.
A2BAR was first identified and cloned in 1992 by Rivkees and Reppert and by Pierce et al. from the rat hypothalamus (Rivkees and Reppert, 1992) and human hippocampus (Pierce et al., 1992). The proposed structure of A2BAR is the typical G-protein-coupled receptor (GPCR) structure, and the predicted molecular mass of A2BAR is 36–37 kDa (Feoktistov and Biaggioni, 1997).
The major signaling pathway of A2BAR is suggested to be the pathway involving adenylyl cyclase (AC) that leads to an increase in intracellular cAMP levels and results in the subsequent activation of PKA and other cAMP effectors such as Epac (Peakman and Hill, 1994; Murakami et al., 2000; Sitaraman et al., 2001; Lynge et al., 2003; Fang and Olah, 2007; Darashchonak et al., 2014; He et al., 2014). However, the A2BAR-Gq-PLC pathway also mediates several crucial functions of A2BAR (Gao et al., 1999; Linden et al., 1999; Panjehpour et al., 2005), and A2BAR further couples to the MAPK and arachidonic acid signaling pathways and regulates membrane ion channels probably through G-protein βγ subunits (Feoktistov et al., 1999; Jimenez et al., 1999; Schulte and Fredholm, 2003a,b; Donoso et al., 2005).
The recent development of A2BAR-knockout/lacZ-knockin mice has enabled the determination of A2BAR distribution in vivo (Yang et al., 2006); A2BAR is widely expressed in numerous tissues and organs, including the vasculature, aortic vascular smooth muscle, cecum, large intestine, brain, and urinary bladder (Yaar et al., 2005; Wang and Huxley, 2006; Yang et al., 2006). Furthermore, a high level of A2BAR expression has been detected in diverse types of cells, including various immune cells such as mast cells (Hua et al., 2007; Ryzhov et al., 2008b), neutrophils (Eckle et al., 2008a), dendritic cells (Pacheco et al., 2005; Ben Addi et al., 2008; Novitskiy et al., 2008), macrophages (Yang et al., 2006), and lymphocytes (Mirabet et al., 1999; Eckle et al., 2008a), as well as other cell types such as type II alveolar epithelial cells (Cagnina et al., 2009), endothelial cells (Yang et al., 2006), chromaffin cells (Casado et al., 1992), astrocytes (Peakman and Hill, 1994; Jimenez et al., 1999), neurons (Corset et al., 2000; Christofi et al., 2001; Stein et al., 2001), and taste cells (Nishida et al., 2014). Moreover, A2BAR expression is influenced by diverse environmental cues such as inflammation, cell stress, injury, and hypoxia (Xaus et al., 1999; Fredholm et al., 2001a; Kolachala et al., 2005; Kong et al., 2006; Hart et al., 2009; Hasko et al., 2009). For example, previous studies have shown that interferon-γ, a proinflammatory cytokine, increases the A2BAR transcriptional level in mouse macrophage cells (Xaus et al., 1999); TNF-α upregulates A2BAR mRNA and protein levels in human colonic epithelial cells (Kolachala et al., 2005); and other mediators such as LPS (Nemeth et al., 2003), IL-1β (Nguyen et al., 2003), free radicals (St Hilaire et al., 2008), and endogenous adenosine (Sitaraman et al., 2002) also enhance A2BAR expression.
Identifying the binding partners of A2BAR is crucial for understanding the receptor's function and regulation. As in other GPCRs, the intracellular portions of A2BAR serve as signal integrators by providing binding sites for effectors or regulatory proteins, although other parts of A2BAR might also be involved in protein interaction. Besides trimeric G proteins and β-arrestin (Feoktistov and Biaggioni, 1997; Mundell et al., 2000; Klinger et al., 2002), the two universal binding partners of GPCRs, numerous other proteins interact with A2BAR. Here, we list these A2BAR binding partners in the order of interaction discovery, and discuss how these proteins modulate or mediate A2BAR functions (Figure 1).
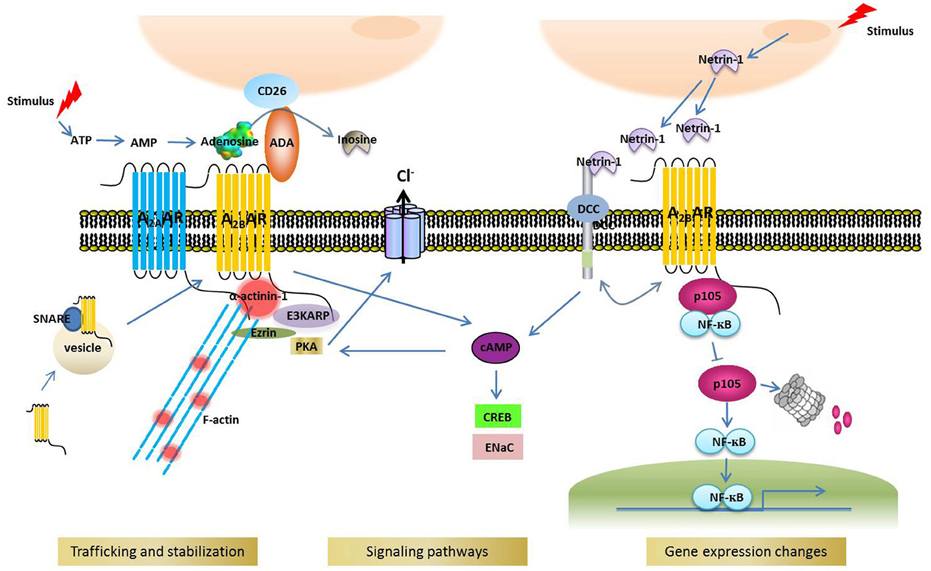
Figure 1. A2BAR binding partners and their cellular functions.
ADA is an enzyme that catalyzes the hydrolytic deamination of adenosine to inosine. Apart from being present in the cytosol and the nucleus, ADA is anchored to the cell surface by other membrane proteins, including CD26 (Pacheco et al., 2005) and A1AR (Saura et al., 1998) in various cell/tissue types such as cultured cortical neurons (Ruiz et al., 2000), DDT1MF-2 cells (Ciruela et al., 1996), and pig brain cortical membrane (Saura et al., 1996). In addition to A1AR and CD26, A2BAR was reported to mediate ADA docking—in CHO and Jurkat cells—onto the extracellular surface (Herrera et al., 2001); counterintuitively, the binding of ADA, even when ADA lacked enzymatic activity, increased the binding affinity of NECA (a nonselective A2AR agonist) for A2BAR and the subsequent production of cAMP. The interaction between ADA and A2BAR was also confirmed in dendritic cells (Pacheco et al., 2005) and gastric mucosa parietal cells (Arin et al., 2015). In dendritic cells, the ADA-A2BAR complex triggers a cell adhesion-costimulatory signal that promotes an immune response, and this is also independent of ADA enzymatic activity (Pacheco et al., 2005). Thus, the ADA-A2BAR complex appears to perform multiple functions, including modulating agonist binding, promoting cell adhesion/costimulation, and degrading extracellular adenosine.
DCC has been proposed to function as a netrin-1 receptor and thus mediate netrin-1-induced axon outgrowth. Corset and collaborators identified A2BAR as one of the proteins that directly binds to DCC and functions as a netrin-1 coreceptor, because netrin-1 activated A2BAR and induced cAMP production, and further suggested that A2BAR is the central mediator of netrin signaling in the regulation of the outgrowth of dorsal spinal cord axons (Corset et al., 2000). However, a subsequent study argued against this view (Stein et al., 2001): the DCC ectodomain was found to interact directly with netrin-1 and mediate netrin signaling to regulate axon growth, and the results of pharmacological analyses suggested that A2BAR function was not required for netrin-1-induced axon growth and guidance. Thus, DCC was proposed to mediate netrin signaling in axon growth and guidance independently of A2BAR activation (Stein et al., 2001). Intriguingly, more recent studies have reported that netrin-1 attenuates neutrophil transmigration and hypoxia-induced inflammation (Rosenberger et al., 2009), alveolar fluid clearance (He et al., 2014), and diabetic nephropathy (Tak et al., 2013) and induces cancer-cell invasion (Rodrigues et al., 2007) in an A2BAR-dependent manner. These results appear to support the general notion that A2BAR mediates the function of netrin-1 at least in certain tissues. Further investigation is required to clarify the discrepancy between the aforementioned studies.
Sitaraman and colleagues demonstrated that the majority of A2BAR localizes intracellularly in quiescent cells and is recruited to the plasma membrane upon agonist stimulation (Sitaraman et al., 2002). The SNARE protein SNAP-23 directly interacts with human A2BAR and participates in A2BAR recruitment to the plasma membrane (Wang et al., 2004), and following SNARE-dependent translocation to the plasma membrane, human A2BAR directly associates with E3KARP (NHERF2) and ezrin and forms a multiprotein complex (Sitaraman et al., 2002). Ezrin is a PKA-anchoring protein, or AKAP, that associates with the actin cytoskeleton (Sun et al., 2000), and this multiprotein complex not only anchors A2BAR to the plasma membrane, but also stabilizes A2BAR expression in the plasma membrane. Furthermore, compartmentalized PKA is effectively activated by A2BAR-induced cAMP production, and the PKA thus activated stimulates CFTR-mediated chloride secretion; this model is consistent with the functional evidence obtained in an early study (Huang et al., 2001).
Interestingly, at its C-terminal end, human A2BAR contains a type 2 PDZ-binding motif (XΦXΦ), GVGL, but not a type 1 PDZ-binding motif (XS/TXV/L). Sitaraman et al. speculated that a PDZ-binding-motif-like sequence in the 3rd intracellular loop in A2BAR might mediate the interaction with E3KARP, a PDZ-domain-containing protein (Sitaraman et al., 2002). However, recent studies indicate that the GVGL sequence of A2BAR participates in the trafficking and surface expression of A2BAR (Watson et al., 2011, 2016), possibly by binding to a PDZ-domain-containing protein. Further investigation is required to determine whether GVGL binds to E3KARP or another PDZ-domain-containing protein.
The function and trafficking of several GPCRs are affected by the heterooligomerization of these receptors. Moriyama and Sitkovsky reported that A2AAR coexpression with A2BAR improves the cell-surface expression of A2BAR, which is normally poor because A2BAR lacks a dominant forward-transport signal for export from the ER to the cell surface (Moriyama and Sitkovsky, 2010). The study further suggested that the functional interaction between A2AAR and A2BAR might be a consequence of their physical association (Moriyama and Sitkovsky, 2010), but how these two receptors interact was not explored. Because both A2AAR and A2BAR were shown to interact with actinins in one previous study (in which the specific actinin isoform was not identified; Burgueno et al., 2003) or with α-actinin-1 in another study (Sun et al., 2016), the α-actinin-1 homodimer or a heterodimer of α-actinin-1 with another actinin isoform might mediate the dimerization of A2AAR and A2BAR and thus promote the surface expression of A2BAR. This mechanism is clearly not mutually exclusive with the mechanism by which α-actinin-1 mediates A2BAR interaction with actin filaments and thereby modulates the trafficking and surface expression of A2BAR (Sun et al., 2016).
NFκB1/p105 is a member of the NFκB family of proteins that perform regulatory functions in diverse biological processes such as inflammation and cell survival and differentiation, as well as in various diseases, including cancer (Barkett and Gilmore, 1999; Hatada et al., 2000; Perkins and Gilmore, 2006). Sun et al. reported that the C-terminal tail of A2BAR binds to NFκB1/p105 independently of ligand activation (Sun et al., 2012). Intriguingly, A2BAR binding to specific sites on p105 prevents the polyubiquitination and degradation of p105 protein and thereby inhibits NFκB activation and reduces inflammation (Sun et al., 2012). In previous studies, both pro- and anti-inflammatory activities have been associated with A2BAR (Blackburn et al., 2009), and the work by Sun et al. potentially sheds light on this paradox: although A2BAR activation by adenosine produces proinflammatory effects, A2BAR can also induce adenosine-independent downregulation of the proinflammatory response by associating with p105. Such receptor bifunctionality displayed by A2BAR—mediation of diametrically opposite effects in the presence and absence of ligand—is reminiscent of dependence receptors (Thibert and Fombonne, 2010). GPCRs other than A2BAR have previously been shown to signal through G-protein-independent pathways, including pathways involving transcription factors (Nehring et al., 2000; White et al., 2000). The study of Sun et al. further suggests that the C-terminus of A2BAR potentially provides a target for developing peptidemimetic drugs that block NFκB signaling, which could be used for treating NFκB-related diseases such as inflammation and cancer (Sun et al., 2012).
Actinins, or α-actinins, represent a family of ubiquitously expressed actin-filament-crosslinking proteins. In addition to performing their critical function of actin-filament crosslinking, actinins link membrane receptors, and cell adhesion proteins to actin filaments and thereby modulate the function and trafficking of these membrane proteins (Oikonomou et al., 2011; Foley and Young, 2014). A recent study by Sun and colleagues suggested that α-actinin-1 binds to the A2BAR C-terminus and stabilizes the receptor's global and cell-surface expression (Sun et al., 2016), which revealed a previously unidentified molecular mechanism for controlling the cellular levels of A2BAR. Because the actinin-1 isoform investigated in the study was the Ca2+-sensitive exon19a splice variant, an intriguing question is whether actinin-1-dependent regulation of A2BAR is also Ca2+ sensitive under physiological conditions.
In contrast to α-actinin-1, actinin-4, another highly homologous non-muscle actinin isoform, did not interact with A2BAR (Sun et al., 2016). Interestingly, actinin-4 has been suggested to interact with the NFκB subunits p65 and p50 and function as a coactivator of the transcription factor NFκB (Zhao et al., 2015). Thus, future studies could investigate whether actinin-1 also associates with NFκB proteins, including p105, and how this association affects the interaction between p105 and A2BAR.
Numerous studies have demonstrated a critical role of A2BAR in the regulation of vascular diseases (Martin, 1992; Dubey et al., 1996; Yang et al., 2008, 2010a), chronic lung disease (Sun et al., 2006; Wilson et al., 2009; Zhou et al., 2009; Zaynagetdinov et al., 2010), and acute lung injury (Eckle et al., 2008a,b; Schingnitz et al., 2010), and several excellent reviews have summarized these studies (Spicuzza et al., 2006; Hasko et al., 2009; Aherne et al., 2011; Headrick et al., 2013). Therefore, in this review, we discuss only the potential functions of A2BAR in three other common human diseases, cancer, renal disease, and diabetes (Figure 2).
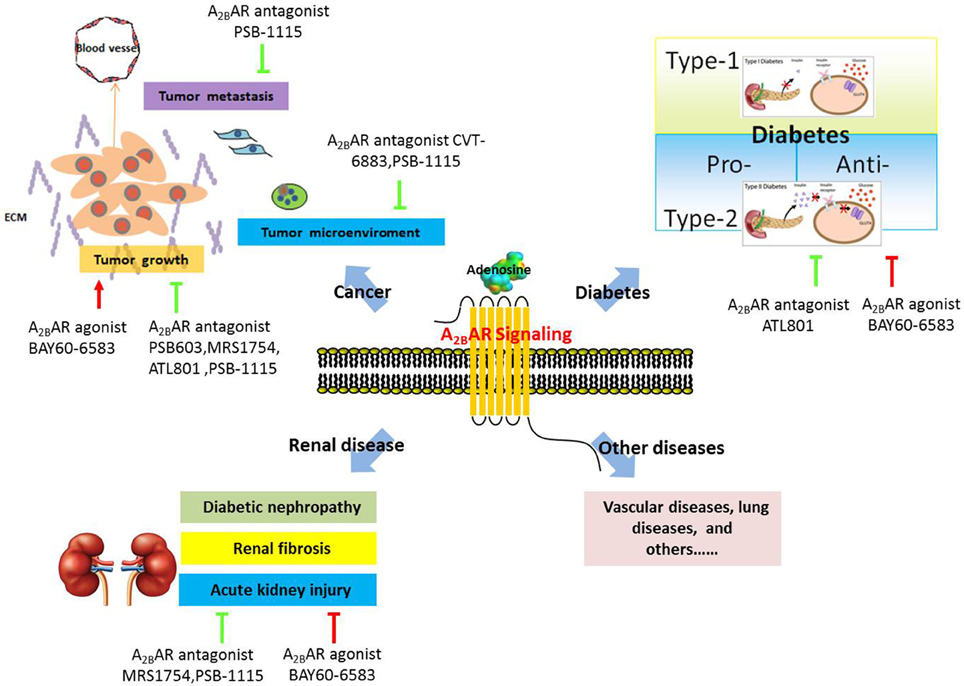
Figure 2. Schematic presentation of the role of A2BAR in various human diseases.
Growing evidence indicates that A2BAR potentially plays a pathophysiological role in human cancer and might serve as a target for novel therapies or cotherapies for cancer. The possible functions of A2BAR in tumor progression and metastasis are discussed here.
First, A2BAR is highly expressed in various types of tumor cells or tissues and promotes tumor-cell proliferation. For instance, A2BAR was found to be overexpressed in colorectal carcinoma cells and tissues, and inhibition of A2BAR blocked the proliferation of colon cancer cells (Ma et al., 2010). In prostate cancer, A2BAR increased cancer-cell proliferation in both ligand-dependent, and ligand-independent manners (Wei et al., 2013; Vecchio et al., 2016). In human oral cancer, A2BAR was shown to be upregulated in oral squamous carcinoma cells, and A2BAR knockdown reduced the proliferation of oral cancer cells through HIF-1α activation (Kasama et al., 2015). Moreover, A2BAR was reported to foster bladder and breast tumor growth in syngeneic mice (Cekic et al., 2012).
Second, A2BAR modulates tumor-cell metastasis. A2BAR was implicated in promoting breast cancer cell migration in vitro and lung metastasis in vivo (Stagg et al., 2010; Desmet et al., 2013), although the underlying molecular mechanism was not fully elucidated. However, the results of a subsequent study suggested a possible explanation: A2BAR activation suppressed the prenylation of the small GTPase Rap1B and diminished Rap1B-mediated cell adhesion, which promoted cell migration (Ntantie et al., 2013).
Third, A2BAR might regulate the tumor microenvironment, including the surrounding blood vessels, immune cells, fibroblasts, and the extracellular matrix. Ryzhov and colleagues provided the first genetic evidence indicating that A2BAR regulates vascular endothelial growth factor (VEGF) production from tumor-infiltrating host immune cells and thereby promotes tumor growth (Ryzhov et al., 2008a). Concomitantly, other groups suggested that A2BAR alters angiogenesis by regulating the production of a wide array of pro- or anti-angiogenic factors such as basic fibroblast growth factor (bFGF), angiopoietin2, and a subset of cytokines (Feoktistov et al., 2002, 2003; Merighi et al., 2009). In addition to affecting angiogenesis, A2BAR regulates dendritic-cell differentiation and function (Novitskiy et al., 2008; Yang et al., 2010b) and alternative macrophage activation (Csoka et al., 2012) and thus contributes to cancer progression.
Thus, A2BAR exerts various effects on tumor progression and metastasis. Notably, most of the aforementioned evidence was collected using in vitro systems, and it is critical to further confirm the role of A2BAR in cancer by using in vivo models before A2BAR is used as a potential cancer therapeutic target.
Renal diseases are estimated to affect millions of people worldwide, whose numbers are growing at a rate of approximately 5–8% annually (Hamer and El Nahas, 2006). Several studies have indicated a critical role of A2BAR in mediating the progression of diabetic nephropathy. Patel et al. and Valladares et al. observed that inhibition of A2BAR activation suppressed VEGF production in glomeruli and further attenuated renal dysfunction in diabetic nephropathy; these data suggested a protective role of A2BAR antagonists in VEGF-induced diabetic nephropathy (Valladares et al., 2008; Patel and Thaker, 2014). However, this view was challenged by Tak et al., who reported elevated VEGF levels in diabetic A2BAR-knockout mice (Tak et al., 2014); concordantly, diabetic nephropathy was highly severe in mice with global or vascular endothelial tissue-specific A2BAR deletion, but not in mice with tubular-epithelial A2BAR deletion. Therefore, Tak et al. suggested that vascular A2BAR signaling is the key mediator of kidney protection during diabetic nephropathy (Tak et al., 2014). The methods used and the specific tissues studied by the aforementioned groups were distinct, which might explain their conflicting observations on the role of A2BAR during diabetic nephropathy. Moreover, the different time windows in which A2BAR inhibition was induced pharmacologically and genetically might also contribute to the discrepancy in the results (Eisenstein et al., 2015).
In addition to playing a role in diabetic nephropathy, A2BAR has been suggested, based on studies on several mouse models, to protect against renal fibrosis. In ADA-deficient mice, a high level of adenosine in kidney tissues resulted in proteinuria and renal fibrosis, and treatment with A2BAR antagonists attenuated renal dysfunction and fibrosis (Dai et al., 2011). Moreover, genetic deletion of A2BAR protected against renal fibrosis in both mice infused with angiotensin II and mice subjected to unilateral ureteral obstruction (Dai et al., 2011). Furthermore, renal biopsy samples from patients with chronic kidney disease (CKD) showed higher levels of A2BAR expression than did samples from patients without CKD (Zhang et al., 2013). All of these data suggest that A2BAR could serve as a potential therapeutic target in the treatment of CKD.
Acute kidney injury, a devastating kidney disease, is often caused by renal ischemia. Rigorous studies from different laboratories have suggested a pivotal role of A2BAR in acute kidney injury. For example, Grenz et al. used genetic and pharmacological approaches to reveal a role of A2BAR in protecting against renal injury resulting from ischemia, although the underlying molecular mechanism was not fully clarified (Grenz et al., 2008). Subsequently, the same group proposed two possible explanations for how A2BAR might provide renal protection: one, A2BAR reduces neutrophil-dependent TNF-α production and suppresses inflammation (Grenz et al., 2012b); and two, A2BAR promotes optimal postischemic blood flow within the kidney and thereby ensures the maximal return of blood flow, tissue oxygenation, and removal of waste products from the ischemic kidney through the A2BAR-ENT1 (equilibrative nucleoside transporter) pathway (Grenz et al., 2012a).
Diabetes mellitus (DM) is the most common endocrine disorder; in 2014, 9% of all adults aged 18+ years were estimated to have diabetes (WHO, 2014), and by 2025, 300 million people worldwide will have the disease (Mane et al., 2012). Adenosine has long been recognized to affect insulin secretion and glucose homeostasis by activating the four AR subtypes (Dong et al., 2001; Nemeth et al., 2007; Fredholm et al., 2011b; Koupenova and Ravid, 2013; Andersson, 2014; Antonioli et al., 2015). Recently, A2BAR in particular has been suggested to function as a critical regulator in DM (Rusing et al., 2006; Johnston-Cox et al., 2012, 2014; Eisenstein et al., 2015; Merighi et al., 2015; Wen et al., 2015).
In a type I DM model, the nonselective receptor agonist NECA blocked diabetes development, and this appeared to be mediated by A2BAR-dependent suppression of proinflammatory cytokine production (Nemeth et al., 2007). These data suggest that A2BAR represents a potential target for the treatment of type I diabetes.
Conversely, some of the evidence obtained using a type II DM model indicated that A2BAR plays a pro-diabetic role. Figler et al. suggested that A2BAR activation increases insulin resistance by elevating the production of proinflammatory mediators such as IL-6 and C-reactive protein (Figler et al., 2011). Deletion of the A2BAR gene and selective blockade of A2BAR in mice reduced hepatic glucose production and enhanced glucose disposal into skeletal muscle and brown adipose tissue (Figler et al., 2011). By contrast, other studies suggested an anti-diabetic role of A2BAR. Johnston-Cox and colleagues showed that A2BAR plays an essential role in high fat diet (HFD)-induced insulin resistance in mice, and mice lacking A2BAR displayed diminished glucose clearance and elevated insulin resistance and inflammatory cytokine production (Johnston-Cox et al., 2012). The underlying cellular mechanism here is mediated by A2BAR expressed in macrophages: reinstatement of macrophage A2BAR expression in A2BAR-null mice restored HFD-induced insulin tolerance and tissue insulin signaling to the level in control mice. The molecular mechanism involves A2BAR altering cAMP signaling and the levels of macrophage cytokine expression and secretion, and this regulates the levels of insulin receptor-2 and downstream insulin signaling (Johnston-Cox et al., 2014). Similar results were obtained by Csoka et al. (2014), who suggested that A2BAR plays a crucial role in sustaining glucose homeostasis and preventing insulin resistance under normal dietary conditions by regulating alternative macrophage activation. Insulin- and glucose-induced glucose clearance was impaired in A2BAR-knockout mice that were fed chow diet, and these knockout mice also exhibited a low level of physical activity, which might contribute to decreased insulin sensitivity in skeletal muscles. Csoka et al. also highlighted the complex role of A2BAR in regulating liver metabolism (Csoka et al., 2014).
In this review, we have discussed certain general characteristics of A2BAR and have described multiple binding partners of the receptor, including α-actinin-1 and p105, whose interactions with the receptor were discovered recently. This identification of A2BAR-binding proteins will undoubtedly help enhance our understanding of the molecular and cellular functions of A2BAR; however, to date, fewer binding partners have been reported for A2BAR than for other AR subtypes. Several reasons might account for this: (1) Little attention was previously devoted to A2BAR because the receptor was long assumed, inaccurately, to be of lesser physiological relevance as compared with other ARs; (2) studies on A2BAR were hampered by a lack of useful biological tools such as specific agonists; and (3) novel experimental approaches such as mass spectrometry were not used to identify A2BAR binding partners.
Recent studies have considerably advanced our understanding of the critical role of A2BAR in the pathogenesis of human diseases, and this raises the possibility that A2BAR could be used as a potential target in the treatment of cancer, diabetes, or other diseases. However, opposing functions of A2BAR have been identified in several diseases. For example, A2BAR activation produces pro- and anti-tumoral effects and the receptor performs pro- and anti-inflammatory functions. These paradoxical effects are least partly contributed by the incompletely explored, agonist-independent activities of A2BAR, including its interactions with p105 (Sun et al., 2012), netrin-1 (Corset et al., 2000), ADA (Herrera et al., 2001; Pacheco et al., 2005), or other effector proteins in specific contexts. Moreover, the discrepant effects might be ascribed to different systems and conditions used for studying them, including cell types, animal models, time window of modulation of A2BAR activity, and the potential side effects of given agonists or antagonists. From a clinical perspective, these opposite effects of A2BAR make it highly challenging to decide whether agonists or antagonists should be used in pharmacological interventions for a given disease. Therefore, to effectively use A2BAR as a therapeutic target, studies must be conducted to elucidate precisely how A2BAR agonist-dependent and -independent functions modulate a particular pathological condition in a specific cellular setting and time window.
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The work was supported by the National Natural Science Foundation of China [Grant No. 81402316], the Shenzhen Innovation Committee of Science and Technology, China [Grant Nos. JCYJ20130401144532136 and JCYJ20160226192238361], and Shenzhen Key Laboratory of Cell Microenvironment [Grant No. ZDSYS20140509142721429] (to YS); and the Hong Kong Research Grants Council grants GRF660913 and GRF16102415 (to PH).
Aherne, C. M., Kewley, E. M., and Eltzschig, H. K. (2011). The resurgence of A2B adenosine receptor signaling. Biochim. Biophys. Acta 1808, 1329–1339. doi: 10.1016/j.bbamem.2010.05.016
Andersson, O. (2014). Role of adenosine signalling and metabolism in beta-cell regeneration. Exp. Cell Res. 321, 3–10. doi: 10.1016/j.yexcr.2013.11.019
Antonioli, L., Blandizzi, C., Csoka, B., Pacher, P., and Hasko, G. (2015). Adenosine signalling in diabetes mellitus–pathophysiology and therapeutic considerations. Nat. Rev. Endocrinol. 11, 228–241. doi: 10.1038/nrendo.2015.10
Arin, R. M., Vallejo, A. I., Rueda, Y., Fresnedo, O., and Ochoa, B. (2015). The A2B adenosine receptor colocalizes with adenosine deaminase in resting parietal cells from gastric mucosa. Biochem. Mosc. 80, 120–125. doi: 10.1134/S0006297915010149
Barkett, M., and Gilmore, T. D. (1999). Control of apoptosis by Rel/NF-kappaB transcription factors. Oncogene 18, 6910–6924. doi: 10.1038/sj.onc.1203238
Ben Addi, A., Lefort, A., Hua, X., Libert, F., Communi, D., Ledent, C., et al. (2008). Modulation of murine dendritic cell function by adenine nucleotides and adenosine: involvement of the A(2B) receptor. Eur. J. Immunol. 38, 1610–1620. doi: 10.1002/eji.200737781
Beukers, M. W., den Dulk, H., van Tilburg, E. W., Brouwer, J., and Ijzerman, A. P. (2000). Why are A(2B) receptors low-affinity adenosine receptors? Mutation of Asn273 to Tyr increases affinity of human A(2B) receptor for 2-(1-Hexynyl)adenosine. Mol. Pharmacol. 58, 1349–1356. doi: 10.1124/mol.58.6.1349
Blackburn, M. R., and Kellems, R. E. (1996). Regulation and function of adenosine deaminase in mice. Prog. Nucleic Acid Res. Mol. Biol. 55, 195–226. doi: 10.1016/S0079-6603(08)60194-4
Blackburn, M. R., Vance, C. O., Morschl, E., and Wilson, C. N. (2009). Adenosine receptors and inflammation. Handb. Exp. Pharmacol. 193, 215–269. doi: 10.1007/978-3-540-89615-9_8
Borea, P. A., Gessi, S., Merighi, S., and Varani, K. (2016). Adenosine as a multi-signalling guardian angel in human diseases: when, where and how does it exert its protective effects? Trends Pharmacol. Sci. 37, 419–434. doi: 10.1016/j.tips.2016.02.006
Burgueno, J., Blake, D. J., Benson, M. A., Tinsley, C. L., Esapa, C. T., Canela, E. I., et al. (2003). The adenosine A2A receptor interacts with the actin-binding protein alpha-actinin. J. Biol. Chem. 278, 37545–37552. doi: 10.1074/jbc.M302809200
Cagnina, R. E., Ramos, S. I., Marshall, M. A., Wang, G., Frazier, C. R., and Linden, J. (2009). Adenosine A2B receptors are highly expressed on murine type II alveolar epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 297, L467–L474. doi: 10.1152/ajplung.90553.2008
Casado, V., Casillas, T., Mallol, J., Canela, E. I., Lluis, C., and Franco, R. (1992). The adenosine receptors present on the plasma membrane of chromaffin cells are of the A2B subtype. J. Neurochem. 59, 425–431. doi: 10.1111/j.1471-4159.1992.tb09388.x
Cekic, C., Sag, D., Li, Y., Theodorescu, D., Strieter, R. M., and Linden, J. (2012). Adenosine A2B receptor blockade slows growth of bladder and breast tumors. J. Immunol. 188, 198–205. doi: 10.4049/jimmunol.1101845
Christofi, F. L., Zhang, H., Yu, J. G., Guzman, J., Xue, J., Kim, M., et al. (2001). Differential gene expression of adenosine A1, A2A, A2B, and A3 receptors in the human enteric nervous system. J. Comp. Neurol. 439, 46–64. doi: 10.1002/cne.1334
Ciruela, F., Saura, C., Canela, E. I., Mallol, J., Lluis, C., and Franco, R. (1996). Adenosine deaminase affects ligand-induced signalling by interacting with cell surface adenosine receptors. FEBS Lett. 380, 219–223. doi: 10.1016/0014-5793(96)00023-3
Corset, V., Nguyen-Ba-Charvet, K. T., Forcet, C., Moyse, E., Chedotal, A., and Mehlen, P. (2000). Netrin-1-mediated axon outgrowth and cAMP production requires interaction with adenosine A2B receptor. Nature 407, 747–750. doi: 10.1038/35037600
Covarrubias, R., Chepurko, E., Reynolds, A., Huttinger, Z. M., Huttinger, R., Stanfill, K., et al. (2016). Role of the CD39/CD73 Purinergic Pathway in Modulating Arterial Thrombosis in Mice. Arterioscler. Thromb. Vasc. Biol. doi: 10.1161/ATVBAHA.116.307374. [Epub ahead of print].
Csoka, B., Koscso, B., Toro, G., Kokai, E., Virag, L., Nemeth, Z. H., et al. (2014). A2B adenosine receptors prevent insulin resistance by inhibiting adipose tissue inflammation via maintaining alternative macrophage activation. Diabetes 63, 850–866. doi: 10.2337/db13-0573
Csoka, B., Selmeczy, Z., Koscso, B., Nemeth, Z. H., Pacher, P., Murray, P. J., et al. (2012). Adenosine promotes alternative macrophage activation via A2A and A2B receptors. FASEB J. 26, 376–386. doi: 10.1096/fj.11-190934
Dai, Y., Zhang, W., Wen, J., Zhang, Y., Kellems, R. E., and Xia, Y. (2011). A2B adenosine receptor-mediated induction of IL-6 promotes CKD. J. Am. Soc. Nephrol. 22, 890–901. doi: 10.1681/ASN.2010080890
Darashchonak, N., Koepsell, B., Bogdanova, N., and von Versen-Hoynck, F. (2014). Adenosine A2B receptors induce proliferation, invasion and activation of cAMP response element binding protein (CREB) in trophoblast cells. BMC Pregnancy Childbirth 14:2. doi: 10.1186/1471-2393-14-2
Desmet, C. J., Gallenne, T., Prieur, A., Reyal, F., Visser, N. L., Wittner, B. S., et al. (2013). Identification of a pharmacologically tractable Fra-1/ADORA2B axis promoting breast cancer metastasis. Proc. Natl. Acad. Sci. U.S.A. 110, 5139–5144. doi: 10.1073/pnas.1222085110
Dong, Q., Ginsberg, H. N., and Erlanger, B. F. (2001). Overexpression of the A1 adenosine receptor in adipose tissue protects mice from obesity-related insulin resistance. Diabetes Obes. Metab. 3, 360–366. doi: 10.1046/j.1463-1326.2001.00158.x
Donoso, M. V., Lopez, R., Miranda, R., Briones, R., and Huidobro-Toro, J. P. (2005). A2B adenosine receptor mediates human chorionic vasoconstriction and signals through arachidonic acid cascade. Am. J. Physiol. Heart Circ. Physiol. 288, H2439–H2449. doi: 10.1152/ajpheart.00548.2004
Dubey, R. K., Gillespie, D. G., Osaka, K., Suzuki, F., and Jackson, E. K. (1996). Adenosine inhibits growth of rat aortic smooth muscle cells. Possible role of A2B receptor. Hypertension 27, 786–793. doi: 10.1161/01.HYP.27.3.786
Dunwiddie, T. V., Diao, L., and Proctor, W. R. (1997). Adenine nucleotides undergo rapid, quantitative conversion to adenosine in the extracellular space in rat hippocampus. J. Neurosci. 17, 7673–7682.
Eckle, T., Faigle, M., Grenz, A., Laucher, S., Thompson, L. F., and Eltzschig, H. K. (2008a). A2B adenosine receptor dampens hypoxia-induced vascular leak. Blood 111, 2024–2035. doi: 10.1182/blood-2007-10-117044
Eckle, T., Grenz, A., Laucher, S., and Eltzschig, H. K. (2008b). A2B adenosine receptor signaling attenuates acute lung injury by enhancing alveolar fluid clearance in mice. J. Clin. Invest. 118, 3301–3315. doi: 10.1172/jci34203
Eckle, T., Hughes, K., Ehrentraut, H., Brodsky, K. S., Rosenberger, P., Choi, D. S., et al. (2013). Crosstalk between the equilibrative nucleoside transporter ENT2 and alveolar Adora2b adenosine receptors dampens acute lung injury. FASEB J. 27, 3078–3089. doi: 10.1096/fj.13-228551
Eckle, T., Krahn, T., Grenz, A., Kohler, D., Mittelbronn, M., Ledent, C., et al. (2007). Cardioprotection by ecto-5′-nucleotidase (CD73) and A2B adenosine receptors. Circulation 115, 1581–1590. doi: 10.1161/CIRCULATIONAHA.106.669697
Eisenstein, A., Patterson, S., and Ravid, K. (2015). The many faces of the A2B adenosine receptor in cardiovascular and metabolic diseases. J. Cell. Physiol. 230, 2891–2897. doi: 10.1002/jcp.25043
Fang, Y., and Olah, M. E. (2007). Cyclic AMP-dependent, protein kinase A-independent activation of extracellular signal-regulated kinase 1/2 following adenosine receptor stimulation in human umbilical vein endothelial cells: role of exchange protein activated by cAMP 1 (Epac1). J. Pharmacol. Exp. Ther. 322, 1189–1200. doi: 10.1124/jpet.107.119933
Feoktistov, I., Goldstein, A. E., and Biaggioni, I. (1999). Role of p38 mitogen-activated protein kinase and extracellular signal-regulated protein kinase kinase in adenosine A2B receptor-mediated interleukin-8 production in human mast cells. Mol. Pharmacol. 55, 726–734.
Feoktistov, I., Goldstein, A. E., Ryzhov, S., Zeng, D., Belardinelli, L., Voyno-Yasenetskaya, T., et al. (2002). Differential expression of adenosine receptors in human endothelial cells: role of A2B receptors in angiogenic factor regulation. Circ. Res. 90, 531–538. doi: 10.1161/01.RES.0000012203.21416.14
Feoktistov, I., Ryzhov, S., Goldstein, A. E., and Biaggioni, I. (2003). Mast cell-mediated stimulation of angiogenesis: cooperative interaction between A2B and A3 adenosine receptors. Circ. Res. 92, 485–492. doi: 10.1161/01.RES.0000061572.10929.2D
Figler, R. A., Wang, G., Srinivasan, S., Jung, D. Y., Zhang, Z., Pankow, J. S., et al. (2011). Links between insulin resistance, adenosine A2B receptors, and inflammatory markers in mice and humans. Diabetes 60, 669–679. doi: 10.2337/db10-1070
Foley, K. S., and Young, P. W. (2014). The non-muscle functions of actinins: an update. Biochem. J. 459, 1–13. doi: 10.1042/BJ20131511
Fredholm, B. B. (2007). Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ. 14, 1315–1323. doi: 10.1038/sj.cdd.4402132
Fredholm, B. B., IJzerman, A. P., Jacobson, K. A., Linden, J., and Muller, C. E. (2011a). International union of basic and clinical pharmacology. LXXXNomenclature, I., and classification of adenosine receptors–an update. Pharmacol. Rev. 63, 1–34. doi: 10.1124/pr.110.003285
Fredholm, B. B., IJzerman, A. P., Jacobson, K. A., Klotz, K. N., and Linden, J. (2001a). International union of pharmacology. XXNomenclature, V., and classification of adenosine receptors. Pharmacolog. Rev. 53, 527–552. doi: 10.1124/pr.110.003285
Fredholm, B. B., Irenius, E., Kull, B., and Schulte, G. (2001b). Comparison of the potency of adenosine as an agonist at human adenosine receptors expressed in Chinese hamster ovary cells. Biochem. Pharmacol. 61, 443–448. doi: 10.1016/S0006-2952(00)00570-0
Fredholm, B. B., Johansson, S., and Wang, Y. Q. (2011b). Adenosine and the regulation of metabolism and body temperature. Adv. Pharmacol. 61, 77–94. doi: 10.1016/B978-0-12-385526-8.00003-5
Fuentes, E., and Palomo, I. (2015). Extracellular ATP metabolism on vascular endothelial cells: a pathway with pro-thrombotic and anti-thrombotic molecules. Vascul. Pharmacol. 75, 1–6. doi: 10.1016/j.vph.2015.05.002
Gao, Z., Chen, T., Weber, M. J., and Linden, J. (1999). A2B adenosine and P2Y2 receptors stimulate mitogen-activated protein kinase in human embryonic kidney-293 cells. cross-talk between cyclic AMP and protein kinase c pathways. J. Biol. Chem. 274, 5972–5980. doi: 10.1074/jbc.274.9.5972
Goblyos, A., and Ijzerman, A. P. (2009). Allosteric modulation of adenosine receptors. Purinergic Signal. 5, 51–61. doi: 10.1007/s11302-008-9105-3
Grenz, A., Bauerle, J. D., Dalton, J. H., Ridyard, D., Badulak, A., Tak, E., et al. (2012a). Equilibrative nucleoside transporter 1 (ENT1) regulates postischemic blood flow during acute kidney injury in mice. J. Clin. Invest. 122, 693–710. doi: 10.1172/JCI60214
Grenz, A., Kim, J. H., Bauerle, J. D., Tak, E., Eltzschig, H. K., and Clambey, E. T. (2012b). Adora2b adenosine receptor signaling protects during acute kidney injury via inhibition of neutrophil-dependent TNF-alpha release. J. Immunol. 189, 4566–4573. doi: 10.4049/jimmunol.1201651
Grenz, A., Osswald, H., Eckle, T., Yang, D., Zhang, H., Tran, Z. V., et al. (2008). The reno-vascular A2B adenosine receptor protects the kidney from ischemia. PLoS Med. 5:e137. doi: 10.1371/journal.pmed.0050137
Grenz, A., Zhang, H., Eckle, T., Mittelbronn, M., Wehrmann, M., Kohle, C., et al. (2007). Protective role of ecto-5′-nucleotidase (CD73) in renal ischemia. J. Am. Soc. Nephrol. 18, 833–845. doi: 10.1681/ASN.2006101141
Hamer, R. A., and El Nahas, M. A. (2006). The burden of chronic kidney disease. BMJ 332, 563–564. doi: 10.1136/bmj.332.7541.563
Hamidzadeh, K., and Mosser, D. M. (2016). Purinergic signaling to terminate TLR responses in macrophages. Front. Immunol. 7:74. doi: 10.3389/fimmu.2016.00074
Hart, M. L., Jacobi, B., Schittenhelm, J., Henn, M., and Eltzschig, H. K. (2009). Cutting Edge: A2B Adenosine receptor signaling provides potent protection during intestinal ischemia/reperfusion injury. J. Immunol. 182, 3965–3968. doi: 10.4049/jimmunol.0802193
Hasko, G., Csoka, B., Nemeth, Z. H., Vizi, E. S., and Pacher, P. (2009). A(2B) adenosine receptors in immunity and inflammation. Trends Immunol. 30, 263–270. doi: 10.1016/j.it.2009.04.001
Hatada, E. N., Krappmann, D., and Scheidereit, C. (2000). NF-kappaB and the innate immune response. Curr. Opin. Immunol. 12, 52–58. doi: 10.1016/S0952-7915(99)00050-3
He, J., Zhao, Y., Deng, W., and Wang, D. X. (2014). Netrin-1 promotes epithelial sodium channel-mediated alveolar fluid clearance via activation of the adenosine 2B receptor in lipopolysaccharide-induced acute lung injury. Respiration 87, 394–407. doi: 10.1159/000358066
Headrick, J. P., Ashton, K. J., Rose'meyer, R. B., and Peart, J. N. (2013). Cardiovascular adenosine receptors: expression, actions and interactions. Pharmacol. Ther. 140, 92–111. doi: 10.1016/j.pharmthera.2013.06.002
Herrera, C., Casado, V., Ciruela, F., Schofield, P., Mallol, J., Lluis, C., et al. (2001). Adenosine A2B receptors behave as an alternative anchoring protein for cell surface adenosine deaminase in lymphocytes and cultured cells. Mol. Pharmacol. 59, 127–134. doi: 10.1124/mol.59.1.127
Hua, X., Kovarova, M., Chason, K. D., Nguyen, M., Koller, B. H., and Tilley, S. L. (2007). Enhanced mast cell activation in mice deficient in the A2B adenosine receptor. J. Exp. Med. 204, 117–128. doi: 10.1084/jem.20061372
Huang, P., Lazarowski, E. R., Tarran, R., Milgram, S. L., Boucher, R. C., and Stutts, M. J. (2001). Compartmentalized autocrine signaling to cystic fibrosis transmembrane conductance regulator at the apical membrane of airway epithelial cells. Proc. Natl. Acad. Sci. U.S.A. 98, 14120–14125. doi: 10.1073/pnas.241318498
Jacobson, K. A., and Gao, Z. G. (2006). Adenosine receptors as therapeutic targets. Nat. Rev. Drug Discov. 5, 247–264. doi: 10.1038/nrd1983
Jimenez, A. I., Castro, E., Mirabet, M., Franco, R., Delicado, E. G., and Miras-Portugal, M. T. (1999). Potentiation of ATP calcium responses by A2B receptor stimulation and other signals coupled to Gs proteins in type-1 cerebellar astrocytes. Glia 26, 119–128.
Johnston-Cox, H., Eisenstein, A. S., Koupenova, M., Carroll, S., and Ravid, K. (2014). The macrophage A2B adenosine receptor regulates tissue insulin sensitivity. PLoS ONE 9:e98775. doi: 10.1371/journal.pone.0098775
Johnston-Cox, H., Koupenova, M., Yang, D., Corkey, B., Gokce, N., Farb, M. G., et al. (2012). The A2B adenosine receptor modulates glucose homeostasis and obesity. PLoS ONE 7:e40584. doi: 10.1371/journal.pone.0040584
Kasama, H., Sakamoto, Y., Kasamatsu, A., Okamoto, A., Koyama, T., Minakawa, Y., et al. (2015). Adenosine A2B receptor promotes progression of human oral cancer. BMC Cancer 15:563. doi: 10.1186/s12885-015-1577-2
Klinger, M., Freissmuth, M., and Nanoff, C. (2002). Adenosine receptors: G protein-mediated signalling and the role of accessory proteins. Cell. Signal. 14, 99–108. doi: 10.1016/S0898-6568(01)00235-2
Kolachala, V., Asamoah, V., Wang, L., Obertone, T. S., Ziegler, T. R., Merlin, D., et al. (2005). TNF-alpha upregulates adenosine 2b (A2B) receptor expression and signaling in intestinal epithelial cells: a basis for A2BR overexpression in colitis. Cell. Mol. Life Sci. 62, 2647–2657. doi: 10.1007/s00018-005-5328-4
Kong, T., Westerman, K. A., Faigle, M., Eltzschig, H. K., and Colgan, S. P. (2006). HIF-dependent induction of adenosine A2B receptor in hypoxia. FASEB J. 20, 2242–2250. doi: 10.1096/fj.06-6419com
Koupenova, M., Johnston-Cox, H., Vezeridis, A., Gavras, H., Yang, D., Zannis, V., et al. (2012). A2B adenosine receptor regulates hyperlipidemia and atherosclerosis. Circulation 125, 354–363. doi: 10.1161/CIRCULATIONAHA.111.057596
Koupenova, M., and Ravid, K. (2013). Adenosine, adenosine receptors and their role in glucose homeostasis and lipid metabolism. J. Cell. Physiol. doi: 10.1002/jcp.24352. [Epub ahead of print].
Kowal, J. M., Yegutkin, G. G., and Novak, I. (2015). ATP release, generation and hydrolysis in exocrine pancreatic duct cells. Purinergic Signal. 11, 533–550. doi: 10.1007/s11302-015-9472-5
Linden, J., Thai, T., Figler, H., Jin, X., and Robeva, A. S. (1999). Characterization of human A(2B) adenosine receptors: radioligand binding, western blotting, and coupling to G(q) in human embryonic kidney 293 cells and HMC-1 mast cells. Mol. Pharmacol. 56, 705–713.
Lloyd, H. G., and Fredholm, B. B. (1995). Involvement of adenosine deaminase and adenosine kinase in regulating extracellular adenosine concentration in rat hippocampal slices. Neurochem. Int. 26, 387–395. doi: 10.1016/0197-0186(94)00144-J
Lynge, J., Schulte, G., Nordsborg, N., Fredholm, B. B., and Hellsten, Y. (2003). Adenosine A 2B receptors modulate cAMP levels and induce CREB but not ERK1/2 and p38 phosphorylation in rat skeletal muscle cells. Biochem. Biophys. Res. Commun. 307, 180–187. doi: 10.1016/S0006-291X(03)01125-2
Ma, D. F., Kondo, T., Nakazawa, T., Niu, D. F., Mochizuki, K., Kawasaki, T., et al. (2010). Hypoxia-inducible adenosine A2B receptor modulates proliferation of colon carcinoma cells. Hum. Pathol. 41, 1550–1557. doi: 10.1016/j.humpath.2010.04.008
Mane, K., Chaluvaraju, K., Niranjan, M., Zaranappa, T., and Manjuthej, T. (2012). Review of insulin and its analogues in diabetes mellitus. J. Basic Clin. Pharm. 3, 283–293. doi: 10.4103/0976-0105.103822
Martin, P. L. (1992). Relative agonist potencies of C2-substituted analogues of adenosine: evidence for adenosine A2B receptors in the guinea pig aorta. Eur. J. Pharmacol. 216, 235–242. doi: 10.1016/0014-2999(92)90365-B
Merighi, S., Borea, P. A., and Gessi, S. (2015). Adenosine receptors and diabetes: focus on the A(2B) adenosine receptor subtype. Pharmacol. Res. 99, 229–236. doi: 10.1016/j.phrs.2015.06.015
Merighi, S., Simioni, C., Gessi, S., Varani, K., Mirandola, P., Tabrizi, M. A., et al. (2009). A(2B) and A(3) adenosine receptors modulate vascular endothelial growth factor and interleukin-8 expression in human melanoma cells treated with etoposide and doxorubicin. Neoplasia 11, 1064–1073. doi: 10.1593/neo.09768
Mirabet, M., Herrera, C., Cordero, O. J., Mallol, J., Lluis, C., and Franco, R. (1999). Expression of A2B adenosine receptors in human lymphocytes: their role in T cell activation. J. Cell Sci. 112(Pt 4), 491–502.
Morello, S., and Miele, L. (2014). Targeting the adenosine A2B receptor in the tumor microenvironment overcomes local immunosuppression by myeloid-derived suppressor cells. Oncoimmunology 3:e27989. doi: 10.4161/onci.27989
Moriyama, K., and Sitkovsky, M. V. (2010). Adenosine A2A receptor is involved in cell surface expression of A2B receptor. J. Biol. Chem. 285, 39271–39288. doi: 10.1074/jbc.M109.098293
Mundell, S. J., Matharu, A. L., Kelly, E., and Benovic, J. L. (2000). Arrestin isoforms dictate differential kinetics of A2B adenosine receptor trafficking. Biochemistry 39, 12828–12836. doi: 10.1021/bi0010928
Murakami, S., Terakura, M., Kamatani, T., Hashikawa, T., Saho, T., Shimabukuro, Y., et al. (2000). Adenosine regulates the production of interleukin-6 by human gingival fibroblasts via cyclic AMP/protein kinase A pathway. J. Periodont. Res. 35, 93–101. doi: 10.1034/j.1600-0765.2000.035002093.x
Nehring, R. B., Horikawa, H. P., El Far, O., Kneussel, M., Brandstatter, J. H., Stamm, S., et al. (2000). The metabotropic GABAB receptor directly interacts with the activating transcription factor 4. J. Biol. Chem. 275, 35185–35191. doi: 10.1074/jbc.M002727200
Nemeth, Z. H., Bleich, D., Csoka, B., Pacher, P., Mabley, J. G., Himer, L., et al. (2007). Adenosine receptor activation ameliorates type 1 diabetes. FASEB J. 21, 2379–2388. doi: 10.1096/fj.07-8213com
Nemeth, Z. H., Leibovich, S. J., Deitch, E. A., Vizi, E. S., Szabo, C., and Hasko, G. (2003). cDNA microarray analysis reveals a nuclear factor-kappaB-independent regulation of macrophage function by adenosine. J. Pharmacol. Exp. Ther. 306, 1042–1049. doi: 10.1124/jpet.103.052944
Nguyen, D. K., Montesinos, M. C., Williams, A. J., Kelly, M., and Cronstein, B. N. (2003). Th1 cytokines regulate adenosine receptors and their downstream signaling elements in human microvascular endothelial cells. J. Immunol. 171, 3991–3998. doi: 10.4049/jimmunol.171.8.3991
Nishida, K., Dohi, Y., Yamanaka, Y., Miyata, A., Tsukamoto, K., Yabu, M., et al. (2014). Expression of adenosine A2B receptor in rat type II and III taste cells. Histochem. Cell Biol. 141, 499–506. doi: 10.1007/s00418-013-1171-0
Novitskiy, S. V., Ryzhov, S., Zaynagetdinov, R., Goldstein, A. E., Huang, Y., Tikhomirov, O. Y., et al. (2008). Adenosine receptors in regulation of dendritic cell differentiation and function. Blood 112, 1822–1831. doi: 10.1182/blood-2008-02-136325
Ntantie, E., Gonyo, P., Lorimer, E. L., Hauser, A. D., Schuld, N., McAllister, D., et al. (2013). An adenosine-mediated signaling pathway suppresses prenylation of the GTPase Rap1B and promotes cell scattering. Sci. Signal. 6, ra39. doi: 10.1126/scisignal.2003374
Ohta, A., and Sitkovsky, M. (2014). Extracellular adenosine-mediated modulation of regulatory T cells. Front. Immunol. 5:304. doi: 10.3389/fimmu.2014.00304
Oikonomou, K. G., Zachou, K., and Dalekos, G. N. (2011). Alpha-actinin: a multidisciplinary protein with important role in B-cell driven autoimmunity. Autoimmun. Rev. 10, 389–396. doi: 10.1016/j.autrev.2010.12.009
Pacheco, R., Martinez-Navio, J. M., Lejeune, M., Climent, N., Oliva, H., Gatell, J. M., et al. (2005). CD26, adenosine deaminase, and adenosine receptors mediate costimulatory signals in the immunological synapse. Proc. Natl. Acad. Sci. U.S.A. 102, 9583–9588. doi: 10.1073/pnas.0501050102
Panjehpour, M., Castro, M., and Klotz, K. N. (2005). Human breast cancer cell line MDA-MB-231 expresses endogenous A2B adenosine receptors mediating a Ca2+ signal. Br. J. Pharmacol. 145, 211–218. doi: 10.1038/sj.bjp.0706180
Patel, L., and Thaker, A. (2014). The effects of adenosine A2B receptor inhibition on VEGF and nitric oxide axis-mediated renal function in diabetic nephropathy. Ren. Fail. 36, 916–924. doi: 10.3109/0886022X.2014.900404
Patel, N., Wu, W., Mishra, P. K., Chen, F., Millman, A., Csoka, B., et al. (2014). A2B adenosine receptor induces protective antihelminth type 2 immune responses. Cell Host Microbe 15, 339–350. doi: 10.1016/j.chom.2014.02.001
Peakman, M. C., and Hill, S. J. (1994). Adenosine A2B-receptor-mediated cyclic AMP accumulation in primary rat astrocytes. Br. J. Pharmacol. 111, 191–198. doi: 10.1111/j.1476-5381.1994.tb14043.x
Perkins, N. D., and Gilmore, T. D. (2006). Good cop, bad cop: the different faces of NF-kappaB. Cell Death Differ. 13, 759–772. doi: 10.1038/sj.cdd.4401838
Picher, M., Burch, L. H., and Boucher, R. C. (2004). Metabolism of P2 receptor agonists in human airways: implications for mucociliary clearance and cystic fibrosis. J. Biol. Chem. 279, 20234–20241. doi: 10.1074/jbc.M400305200
Picher, M., Burch, L. H., Hirsh, A. J., Spychala, J., and Boucher, R. C. (2003). Ecto 5′-nucleotidase and nonspecific alkaline phosphatase. Two AMP-hydrolyzing ectoenzymes with distinct roles in human airways. J. Biol. Chem. 278, 13468–13479. doi: 10.1074/jbc.M300569200
Pierce, K. D., Furlong, T. J., Selbie, L. A., and Shine, J. (1992). Molecular cloning and expression of an adenosine A2B receptor from human brain. Biochem. Biophys. Res. Commun. 187, 86–93. doi: 10.1016/S0006-291X(05)81462-7
Rivkees, S. A., and Reppert, S. M. (1992). RFL9 encodes an A2B-adenosine receptor. Mol. Endocrinol. 6, 1598–1604.
Rodrigues, S., De Wever, O., Bruyneel, E., Rooney, R. J., and Gespach, C. (2007). Opposing roles of netrin-1 and the dependence receptor DCC in cancer cell invasion, tumor growth and metastasis. Oncogene 26, 5615–5625. doi: 10.1038/sj.onc.1210347
Rosenberger, P., Schwab, J. M., Mirakaj, V., Masekowsky, E., Mager, A., Morote-Garcia, J. C., et al. (2009). Hypoxia-inducible factor-dependent induction of netrin-1 dampens inflammation caused by hypoxia. Nat. Immunol. 10, 195–202. doi: 10.1038/ni.1683
Ross, A. E., Nguyen, M. D., Privman, E., and Venton, B. J. (2014). Mechanical stimulation evokes rapid increases in extracellular adenosine concentration in the prefrontal cortex. J. Neurochem. 130, 50–60. doi: 10.1111/jnc.12711
Ruiz, M. A., Escriche, M., Lluis, C., Franco, R., Martin, M., Andres, A., et al. (2000). Adenosine A(1) receptor in cultured neurons from rat cerebral cortex: colocalization with adenosine deaminase. J. Neurochem. 75, 656–664. doi: 10.1046/j.1471-4159.2000.0750656.x
Rusing, D., Muller, C. E., and Verspohl, E. J. (2006). The impact of adenosine and A(2B) receptors on glucose homoeostasis. J. Pharm. Pharmacol. 58, 1639–1645. doi: 10.1211/jpp.58.12.0011
Ryzhov, S., Novitskiy, S. V., Zaynagetdinov, R., Goldstein, A. E., Carbone, D. P., Biaggioni, I., et al. (2008a). Host A(2B) adenosine receptors promote carcinoma growth. Neoplasia 10, 987–995. doi: 10.1593/neo.08478
Ryzhov, S., Zaynagetdinov, R., Goldstein, A. E., Novitskiy, S. V., Dikov, M. M., Blackburn, M. R., et al. (2008b). Effect of A2B adenosine receptor gene ablation on proinflammatory adenosine signaling in mast cells. J. Immunol. 180, 7212–7220. doi: 10.4049/jimmunol.180.11.7212
Saura, C. A., Mallol, J., Canela, E. I., Lluis, C., and Franco, R. (1998). Adenosine deaminase and A1 adenosine receptors internalize together following agonist-induced receptor desensitization. J. Biol. Chem. 273, 17610–17617. doi: 10.1074/jbc.273.28.17610
Saura, C., Ciruela, F., Casado, V., Canela, E. I., Mallol, J., Lluis, C., et al. (1996). Adenosine deaminase interacts with A1 adenosine receptors in pig brain cortical membranes. J. Neurochem. 66, 1675–1682. doi: 10.1046/j.1471-4159.1996.66041675.x
Schingnitz, U., Hartmann, K., Macmanus, C. F., Eckle, T., Zug, S., Colgan, S. P., et al. (2010). Signaling through the A2B adenosine receptor dampens endotoxin-induced acute lung injury. J. Immunol. 184, 5271–5279. doi: 10.4049/jimmunol.0903035
Schulte, G., and Fredholm, B. B. (2003a). The G(s)-coupled adenosine A(2B) receptor recruits divergent pathways to regulate ERK1/2 and p38. Exp. Cell Res. 290, 168–176. doi: 10.1016/S0014-4827(03)00324-0
Schulte, G., and Fredholm, B. B. (2003b). Signalling from adenosine receptors to mitogen-activated protein kinases. Cell. Signal. 15, 813–827. doi: 10.1016/S0898-6568(03)00058-5
Sitaraman, S. V., Merlin, D., Wang, L., Wong, M., Gewirtz, A. T., Si-Tahar, M., et al. (2001). Neutrophil-epithelial crosstalk at the intestinal lumenal surface mediated by reciprocal secretion of adenosine and IL-6. J. Clin. Invest. 107, 861–869. doi: 10.1172/JCI11783
Sitaraman, S. V., Wang, L., Wong, M., Bruewer, M., Hobert, M., Yun, C. H., et al. (2002). The adenosine 2b receptor is recruited to the plasma membrane and associates with E3KARP and Ezrin upon agonist stimulation. J. Biol. Chem. 277, 33188–33195. doi: 10.1074/jbc.M202522200
Spicuzza, L., Di Maria, G., and Polosa, R. (2006). Adenosine in the airways: implications and applications. Eur. J. Pharmacol. 533, 77–88. doi: 10.1016/j.ejphar.2005.12.056
Spychala, J., Datta, N. S., Takabayashi, K., Datta, M., Fox, I. H., Gribbin, T., et al. (1996). Cloning of human adenosine kinase cDNA: sequence similarity to microbial ribokinases and fructokinases. Proc. Natl. Acad. Sci. U.S.A. 93, 1232–1237. doi: 10.1073/pnas.93.3.1232
Stagg, J., Divisekera, U., McLaughlin, N., Sharkey, J., Pommey, S., Denoyer, D., et al. (2010). Anti-CD73 antibody therapy inhibits breast tumor growth and metastasis. Proc. Natl. Acad. Sci. U.S.A. 107, 1547–1552. doi: 10.1073/pnas.0908801107
Stein, E., Zou, Y., Poo, M., and Tessier-Lavigne, M. (2001). Binding of DCC by netrin-1 to mediate axon guidance independent of adenosine A2B receptor activation. Science 291, 1976–1982. doi: 10.1126/science.1059391
St Hilaire, C., Koupenova, M., Carroll, S. H., Smith, B. D., and Ravid, K. (2008). TNF-alpha upregulates the A2B adenosine receptor gene: the role of NAD(P)H oxidase 4. Biochem. Biophys. Res. Commun. 375, 292–296. doi: 10.1016/j.bbrc.2008.07.059
Sun, C. X., Zhong, H., Mohsenin, A., Morschl, E., Chunn, J. L., Molina, J. G., et al. (2006). Role of A2B adenosine receptor signaling in adenosine-dependent pulmonary inflammation and injury. J. Clin. Invest. 116, 2173–2182. doi: 10.1172/JCI27303
Sun, F., Hug, M. J., Lewarchik, C. M., Yun, C. H., Bradbury, N. A., and Frizzell, R. A. (2000). E3KARP mediates the association of ezrin and protein kinase A with the cystic fibrosis transmembrane conductance regulator in airway cells. J. Biol. Chem. 275, 29539–29546. doi: 10.1074/jbc.M004961200
Sun, Y., Duan, Y., Eisenstein, A. S., Hu, W., Quintana, A., Lam, W. K., et al. (2012). A novel mechanism of control of NFkappaB activation and inflammation involving A2B adenosine receptors. J. Cell Sci. 125, 4507–4517. doi: 10.1242/jcs.105023
Sun, Y., Hu, W., Yu, X., Liu, Z., Tarran, R., Ravid, K., et al. (2016). Actinin-1 binds to the C-terminus of A2B adenosine receptor (A2BAR) and enhances A2BAR cell-surface expression. Biochem. J. 473, 2179-2186. doi: 10.1042/BCJ20160272
Tak, E., Ridyard, D., Badulak, A., Giebler, A., Shabeka, U., Werner, T., et al. (2013). Protective role for netrin-1 during diabetic nephropathy. J. Mol. Med. 91, 1071–1080. doi: 10.1007/s00109-013-1041-1
Tak, E., Ridyard, D., Kim, J. H., Zimmerman, M., Werner, T., Wang, X. X., et al. (2014). CD73-dependent generation of adenosine and endothelial Adora2b signaling attenuate diabetic nephropathy. J. Am. Soc. Nephrol. 25, 547–563. doi: 10.1681/ASN.2012101014
Tang, J., Jiang, X., Zhou, Y., and Dai, Y. (2015). Effects of A2BR on the biological behavior of mouse renal fibroblasts during hypoxia. Mol. Med. Rep. 11, 4397–4402. doi: 10.3892/mmr.2015.3320
Thibert, C., and Fombonne, J. (2010). Dependence receptors: mechanisms of an announced death. Cell Cycle 9, 2085–2091. doi: 10.4161/cc.9.11.11809
Valladares, D., Quezada, C., Montecinos, P., Concha, I. I., Yanez, A. J., Sobrevia, L., et al. (2008). Adenosine A(2B) receptor mediates an increase on VEGF-A production in rat kidney glomeruli. Biochem. Biophys. Res. Commun. 366, 180–185. doi: 10.1016/j.bbrc.2007.11.113
Vecchio, E. A., Tan, C. Y., Gregory, K. J., Christopoulos, A., White, P. J., and May, L. (2016). Ligand-independent adenosine A2B receptor constitutive activity as a promoter of prostate cancer cell proliferation. J. Pharmacol. Exp. Ther. 357, 36–44. doi: 10.1124/jpet.115.230003
Wang, J., and Huxley, V. H. (2006). Adenosine A2A receptor modulation of juvenile female rat skeletal muscle microvessel permeability. Am. J. Physiol. Heart Circ. Physiol. 291, H3094–H3105. doi: 10.1152/ajpheart.00526.2006
Wang, L., Kolachala, V., Walia, B., Balasubramanian, S., Hall, R. A., Merlin, D., et al. (2004). Agonist-induced polarized trafficking and surface expression of the adenosine 2b receptor in intestinal epithelial cells: role of SNARE proteins. Am. J. Physiol. Gastrointest. Liver Physiol. 287, G1100–G1107. doi: 10.1152/ajpgi.00164.2004
Watson, M. J., Lee, S. L., Marklew, A. J., Gilmore, R. C., Gentzsch, M., Sassano, M. F., et al. (2016). The Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Uses its C-Terminus to Regulate the A2B Adenosine Receptor. Sci. Rep. 6:27390. doi: 10.1038/srep27390
Watson, M. J., Worthington, E. N., Clunes, L. A., Rasmussen, J. E., Jones, L., and Tarran, R. (2011). Defective adenosine-stimulated cAMP production in cystic fibrosis airway epithelia: a novel role for CFTR in cell signaling. FASEB J. 25, 2996–3003. doi: 10.1096/fj.11-186080
Wei, Q., Costanzi, S., Balasubramanian, R., Gao, Z. G., and Jacobson, K. A. (2013). A2B adenosine receptor blockade inhibits growth of prostate cancer cells. Purinergic Signal. 9, 271–280. doi: 10.1007/s11302-012-9350-3
Wen, J., Wang, B., Du, C., Xu, G., Zhang, Z., Li, Y., et al. (2015). A2B Adenosine receptor agonist improves erectile function in diabetic rats. Tohoku J. Exp. Med. 237, 141–148. doi: 10.1620/tjem.237.141
White, J. H., McIllhinney, R. A., Wise, A., Ciruela, F., Chan, W. Y., Emson, P. C., et al. (2000). The GABAB receptor interacts directly with the related transcription factors CREB2 and ATFx. Proc. Natl. Acad. Sci. U.S.A. 97, 13967–13972. doi: 10.1073/pnas.240452197
Wilson, C. N., Nadeem, A., Spina, D., Brown, R., Page, C. P., and Mustafa, S. J. (2009). Adenosine receptors and asthma. Handb. Exp. Pharmacol. 193, 329–362. doi: 10.1007/978-3-540-89615-9_11
Xaus, J., Mirabet, M., Lloberas, J., Soler, C., Lluis, C., Franco, R., et al. (1999). IFN-gamma up-regulates the A2B adenosine receptor expression in macrophages: a mechanism of macrophage deactivation. J. Immunol. 162, 3607–3614.
Yaar, R., Jones, M. R., Chen, J. F., and Ravid, K. (2005). Animal models for the study of adenosine receptor function. J. Cell. Physiol. 202, 9–20. doi: 10.1002/jcp.20138
Yang, D., Chen, H., Koupenova, M., Carroll, S. H., Eliades, A., Freedman, J. E., et al. (2010a). A new role for the A2B adenosine receptor in regulating platelet function. J. Thromb. Haemost. 8, 817–827. doi: 10.1111/j.1538-7836.2010.03769.x
Yang, D., Koupenova, M., McCrann, D. J., Kopeikina, K. J., Kagan, H. M., Schreiber, B. M., et al. (2008). The A2B adenosine receptor protects against vascular injury. Proc. Natl. Acad. Sci. U.S.A. 105, 792–796. doi: 10.1073/pnas.0705563105
Yang, D., Zhang, Y., Nguyen, H. G., Koupenova, M., Chauhan, A. K., Makitalo, M., et al. (2006). The A2B adenosine receptor protects against inflammation and excessive vascular adhesion. J. Clin. Invest. 116, 1913–1923. doi: 10.1172/JCI27933
Yang, M., Ma, C., Liu, S., Shao, Q., Gao, W., Song, B., et al. (2010b). HIF-dependent induction of adenosine receptor A2B skews human dendritic cells to a Th2-stimulating phenotype under hypoxia. Immunol. Cell Biol. 88, 165–171. doi: 10.1038/icb.2009.77
Zaynagetdinov, R., Ryzhov, S., Goldstein, A. E., Yin, H., Novitskiy, S. V., Goleniewska, K., et al. (2010). Attenuation of chronic pulmonary inflammation in A2B adenosine receptor knockout mice. Am. J. Respir. Cell Mol. Biol. 42, 564–571. doi: 10.1165/rcmb.2008-0391OC
Zhang, W., Zhang, Y., Wang, W., Dai, Y., Ning, C., Luo, R., et al. (2013). Elevated ecto-5′-nucleotidase-mediated increased renal adenosine signaling via A2B adenosine receptor contributes to chronic hypertension. Circ. Res. 112, 1466–1478. doi: 10.1161/CIRCRESAHA.111.300166
Zhao, X., Hsu, K. S., Lim, J. H., Bruggeman, L. A., and Kao, H. Y. (2015). alpha-Actinin 4 potentiates nuclear factor kappa-light-chain-enhancer of activated B-cell (NF-kappaB) activity in podocytes independent of its cytoplasmic actin binding function. J. Biol. Chem. 290, 338–349. doi: 10.1074/jbc.M114.597260
Keywords: A2B adenosine receptor, binding proteins, cancer, renal disease, diabetes
Citation: Sun Y and Huang P (2016) Adenosine A2B Receptor: From Cell Biology to Human Diseases. Front. Chem. 4:37. doi: 10.3389/fchem.2016.00037
Received: 23 June 2016; Accepted: 11 August 2016;
Published: 24 August 2016.
Edited by:
Cesare Indiveri, University of Calabria, ItalyReviewed by:
Rafael Franco, University of Barcelona, SpainCopyright © 2016 Sun and Huang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ying Sun, c3VueUBzdXN0Yy5lZHUuY24=
Pingbo Huang, Ym9odWFuZ3BAdXN0Lmhr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.