
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Chem. Eng., 27 September 2022
Sec. Sustainable Process Engineering
Volume 4 - 2022 | https://doi.org/10.3389/fceng.2022.1013873
This article is part of the Research TopicThe Role of Agave as Feedstock within a Sustainable Circular BioeconomyView all 6 articles
 Diego Díaz-Vázquez1
Diego Díaz-Vázquez1 Marycarmen Verduzco Garibay1
Marycarmen Verduzco Garibay1 Alberto Fernández del Castillo1
Alberto Fernández del Castillo1 Danielle A. Orozco-Nunnelly2
Danielle A. Orozco-Nunnelly2 Carolina Senés-Guerrero1
Carolina Senés-Guerrero1 Misael Sebastián Gradilla-Hernández1*
Misael Sebastián Gradilla-Hernández1*The Tequila industry is a major producer of wastewater in the state of Jalisco, Mexico. Tequila vinasses (TV) are a residue from the distillation of fermented agave wort during tequila production. TV are difficult to treat due their high organic content, high nutrient loads, acidic pH and high discharge temperature. TV are frequently released into waterbodies or soil without any treatment, leading to environmental degradation of soil and water sources. To reduce the environmental impact of the tequila industry, cost-effective TV revalorization approaches must be developed. The goals of the present study were to assess the treatment and revalorization potential of TV using mono and mixed yeast cultures to produce single-cell protein (SCP) and to analyze yeast community composition using high-throughput sequencing during the mixed-culture fermentation of TV. The fermentation process was performed using a mixed culture of three fodder yeast species (Candida utilis, Rhodotorula mucilaginosa and Kluyveromyces marxianus) during 48 h at benchtop-scale. High-throughput sequencing was performed to assess the relative abundance of the yeast communities. Additionally, a redundancy analysis was performed to analyze the bidirectional influence between yeast communities and pollutant removal (COD, nitrogen, phosphorus, proteins, and sugars). Mixed yeast cultures displayed overall higher pollutant removal rates than monocultures, where C. utilis and K. marxianus contributed the most to pollutant removal and protein accumulation. The R. mucilaginosa population declined rapidly in mixed culture, presumably due to TV acidity and phenolic composition. However, the presence of The R. mucilaginosa in the mixed culture enhanced pollutant removal and amino acid contents. Accordingly, the protein and amino acid content within mixed cultures were significantly higher than those of monocultures, indicating that mixed cultures have a strong potential to produce protein rich biomass from TV, aiding in the transition of both the tequila and the livestock industries to a sustainable circular bioeconomy model by the reintegration of organic material flows into productive processes, reducing raw resource intake and waste generation. The present circular bioeconomy approach could represent a potential to produce 45,664 tons of protein feed yearly, based on the current tequila vinasses generated in the state of Jalisco.
Agro-industrial waste management is a challenging task for highly productive regions across the globe, as it can potentially cause severe environmental and health problems (Bedoić et al., 2019). In Mexico, the state of Jalisco is an intensive agro-industrial region where most tequila (a fermented alcoholic beverage made from Agave tequilana Weber var. azul) is produced. In 2019, 374 million liters of tequila were produced in Mexico, of which 70% was exported internationally (CRT, 2019). Tequila vinasses (TV) are the main by-product produced from tequila distillation; between 7 and 15 L of TV are generated for each liter of tequila produced (España-Gamboa et al., 2011). In 2019, this resulted in a TV production of 2,645,655.70 m3 year−1 in Mexico (Díaz-Vázquez et al., 2021). TV display high acidity as well as high concentrations of total solids (TS), chemical oxygen demand (COD), and nutrients (nitrogen and phosphorus). Furthermore, their characteristic dark brown color is due to a high concentration of phenolic compounds that are produced during the distillation process (López-López et al., 2010). TV are rarely treated by small and mid-sized tequila producers due to technical and economic limitations, and, as a result, the direct discharge of untreated or partially treated TV into soils and waterbodies is a widespread practice among producers (Díaz-Vázquez et al., 2021).
A circular bioeconomy model aims to integrate the biological recovery of organic resources and nutrients from waste into a circular economy scheme (Sherwood, 2020). Accordingly, the resources contained in TV can be recovered into a variety of subproducts, such as fertilizers, biogas, molecular hydrogen, and certain aromatic compounds (Buitrón et al., 2014; Díaz-Vázquez et al., 2021). The production of yeast-derived single-cell protein (SCP) has gained special interest for livestock applications due to its high protein content, unique amino acid profile, low nucleic acid content, and lipid, mineral and vitamin compositions suitable for livestock diet supplementation (Ugalde & Castrillo, 2002). Furthermore, mixed yeast cultures have shown great capability to remove higher loads of contaminants in wastewater when compared to yeast monocultures (Wang et al., 2018). Therefore, mixed yeast cultures present an excellent alternative for the treatment of TV, while simultaneously producing cattle feed supplements. Additionally, high-throughput sequencing allows the evolution of microbial cultures to be observed during particular processes and to correlate the abundance of microbial communities with specific conditions of their environment (Ercolini, 2013; Gill, 2017). A deeper comprehension of microbial dynamics is required to treat TV using mixed yeast cultures on a large scale. This would allow for simultaneous pollutant removal and SCP production, increasing the economic feasibility for the integrated management of TV though a circular bioeconomy approach. Therefore, the goals of the present study were to assess the treatment and revalorization potential of TV using mono and mixed yeast cultures to produce SCP and to analyze yeast community composition using high-throughput sequencing during the mixed-culture fermentation of tequila vinasses.
R. mucilaginosa (ATCC 9450) (R), C. utilis (ATCC 9950) (C), and K. marxianus (ATCC 2512) (K) were selected due to their ability to use TV as a substrate, as well as for their potential to produce biomass that can be used as a livestock feed supplement. R, C and K were obtained from the National Microbiological Strain and Cell Culture Collection (CDBB, 2021) and were evaluated as mono- and mixed-culture treatments to assess possible interactions between yeast communities and their specific pollutant degrading mechanisms. The two mixed culture combinations used in the current study (C + K and C + K + R) were selected based on previous results reported by Díaz-Vázquez et al. (2022) for the treatment and revalorization of TV. The inoculum used for the benchtop-scale cultures was initially prepared at a concentration of 10−6 cells ml−1 at 30°C for 16 h and was mixed using a shaking incubator (Ecotron, Infors HT). All yeast species were cultured in YM broth (Merck).
The TV used in these experiments were provided by a large-scale tequila production facility (Tierra de Agaves; 20.870292 N, −103.792093 W) located in the municipality of Tequila, Jalisco. The TV were generated from the fermentation of 100% agave (no additional sugars) and distilled in a continuous distillation column. The TV were characterized for chemical oxygen demand (COD), total nitrogen (TN), and total phosphorus (TP), based on the current applicable legislation set by Mexico’s Ministry of the Environment (DOF, 1997, 2021). After characterization, all TV were stored at −80°C prior to the yest culture treatment.
Benchtop cultures were performed on a 5 L ez-Control bioreactor (Applikon) using a working volume of 3 L of autoclave sterilized undiluted TV. 5% v/v of each respective yeast inoculum was added at a concentration of 106 cells ml−1, using equal microorganism proportions for all mixed cultures. The temperature, agitation, and aeration rates remained constant throughout each experiment at 30°C, 200 rpm, and 1 vvm, respectively. Experimental runs were 48 h long in duration with culture samples taken in triplicate every 24 h after the initial inoculation. The samples used for the assessment of pollutant removal were stored in 50 ml sterile conical tubes at -20°C prior to their analysis. The samples used for DNA extraction were collected in 2 ml microtubes and stored at −80°C prior to processing. Experimental duplicates were performed for all benchtop-scale treatments to assess between-run reproducibility.
Viable biomass (VB) was monitored using a Futura 6531-52 (ABER) capacitance annular probe, with readings performed using the microbial reading preset (580 and 15,650 kHz). The sensor was calibrated individually for each of the three yeast cultures (C, K and R) using YM broth alone to correlate capacitance readings with dry weight biomass under the same culture conditions as used in the experimental setup. A single linear regression (R = 0.9638) was obtained that integrated the individual monoculture (C, K and R) capacitance-biomass regressions. A unified regression was used for all experimental runs in the present study (mono and mixed) since the cell morphology of all three microorganisms was similar and therefore presented a similar capacitance-biomass response, as previously reported (Díaz-Vázquez et al., 2022).
Uncentrifuged broth samples were homogenized for 1 min at 16,000 rpm (IKA T25 Ultra Turrax homogenizer) prior to protein (P) determination by the Bradford assay, as reported by Nouroozi et al. (2015), using 96-well microplate. A microplate spectrophotometer (Varioskan Lux, Thermo Scientific) was used to perform the readings at 595 nm. Bovine serum albumin (BSA) (Merck) was employed as the standard for the construction of the protein content calibration curve (R = 9952). A fraction of the culture broth samples was centrifuged for 5 min at 4,000 rpm (1580R centrifuge, Gyrozen) to separate the media from the generated biomass. The resulting centrifuged media was used to evaluate reducing sugars (RS), COD, TN, and TP. Reducing sugar concentrations were determined by the dinitrosalicylic acid method, as reported by Wood et al. (2012), on 96-well microplates with readings performed at 540 nm using a microplate spectrophotometer (Varioskan Lux, Thermo Scientific). Glucose was employed as the standard for the construction of the reducing sugar calibration curve (R = 0.9981). Commercial colorimetric kits were used for the determination of chemical oxygen demand (HACH 2125915), total nitrogen (HACH 2714100), and total phosphorus (HACH 2767245). All colorimetric kits were incubated on a heating block (HACH DRB200) following the manufacturer’s instructions. Readings were performed on a HACH DR3900 spectrophotometer using the chemical oxygen demand, total nitrogen, and total phosphorus presets, respectively.
Samples were taken from the mixed culture treatments (C + K and C + K + R) for DNA extraction at initial (0 h), middle (24 h) and final (48 h) times to evaluate the temporal changes in the relative abundance (in terms of ribosomal DNA sequence reads) of individual yeast communities within the mixed cultures. Samples were taken from each of the experimental culture treatment duplicates resulting in a total of 12 samples (6 for each of the mixed culture treatments). The DNA extraction procedure was performed using the FastDNA Spin Kit for Soil (MP Biomedicals, Solon, OH, United States). For this, 500 µl of the sample were placed on a matrix tube for DNA extraction following the manufacturer’s specifications using three washings with SEWS-M solution. DNA extractions were concentrated with Microcon Centrifugal Filters (information of the kit), where 80 µl of the sample were filtered following manufacturer´s instructions. DNA extractions were stored at −80°C. High-throughput sequencing of the ITS2 rDNA region was performed by Novogene Corporation Inc. using NovaSeq 6000 PE250 employing the ITS3-2024F (GCATCGATGAAGAACGCAGC) and ITS4-2409R (TCCTCCGCTTATTGATATGC) primers.
Raw sequences were analyzed using QIIME 2 software (Quantitative Insights into Microbial Ecology) (Bolyen et al., 2019) following a bioinformatic pipeline for the fungal nuclear ribosomal internal transcribed spacer (ITS) region (Bokulich, 2018). The raw sequences were imported to QIIME 2 in Cassava format, then demultiplexed sequences were denoised into amplicon sequence variants (ASV) using the DADA 2 algorithm with the following parameters: p-trunc-len-f 0, p-trunc-len-r 0, p-trim-left-f 0, p-trim-left-r 0. Afterwards, the UNITE database (version 8.3) was used as reference database (Abarenkov et al., 2021) to construct sequence and taxonomy tables. The classifier was trained using the sequences and length of the primers through the Naives Bayes classifier method (Bolyen et al., 2019). Subsequently, the sequences and the trained classifiers were aligned using the classify-sklearn algorithm (Bolyen et al., 2019). Finally, the classified ASV were downloaded in CVS format (comma-separated value file) from the QIIME 2 visualizer (view.qiime2.org) for further analysis.
Minitab 19.2020.1 was used to perform the one-way ANOVA and Tukey tests presented in this study, which were employed to evaluate statistical differences in pollutant removal efficiencies (COD, TN, TP) and revalorization potential parameters for livestock feed applications (VB, RS, P) between treatments. Moreover, one-way ANOVA and Tukey tests were utilized to evaluate significant differences in amino acid composition and total protein content between the culture treatments.
Additionally, a redundancy analysis (RDA) was performed to analyze the effects of various physicochemical parameters on the composition of the yeast communities. RDA is an analysis that models the response variables as a function of the explanatory variables (Zuur et al., 2007). In this case, the response variables are the yeast communities at the genus level and the physicochemical parameters were the explanatory variables, respectively. A correlation triplot was used to extract further information from the RDA results; this plot consists of two superimposed biplots, which include quantitative explanatory and response variables (vectors) and observation points (Zuur et al., 2007; Borcard et al., 2011). R studio version 4.1.0 was used to perform the RDA and to develop the correlation triplot using the scales (Wickham, 2016), vegan (Dixon, 2003) and ggplot2 packages (Wickham, 2016).
The six monitored parameters (COD removal, TN removal, TP removal, P, RS, VB) for the mono (C, K and R) and mixed (C + K and C + K + R) yeast treatments are presented in Table 1 for the 0, 24, and 48 h timepoints from inoculation. This data was used to compare pollutant removal capacities between the various mono- and mixed-cultures, as well as their revalorization potential for use in the livestock industry as a protein-rich feed supplement.
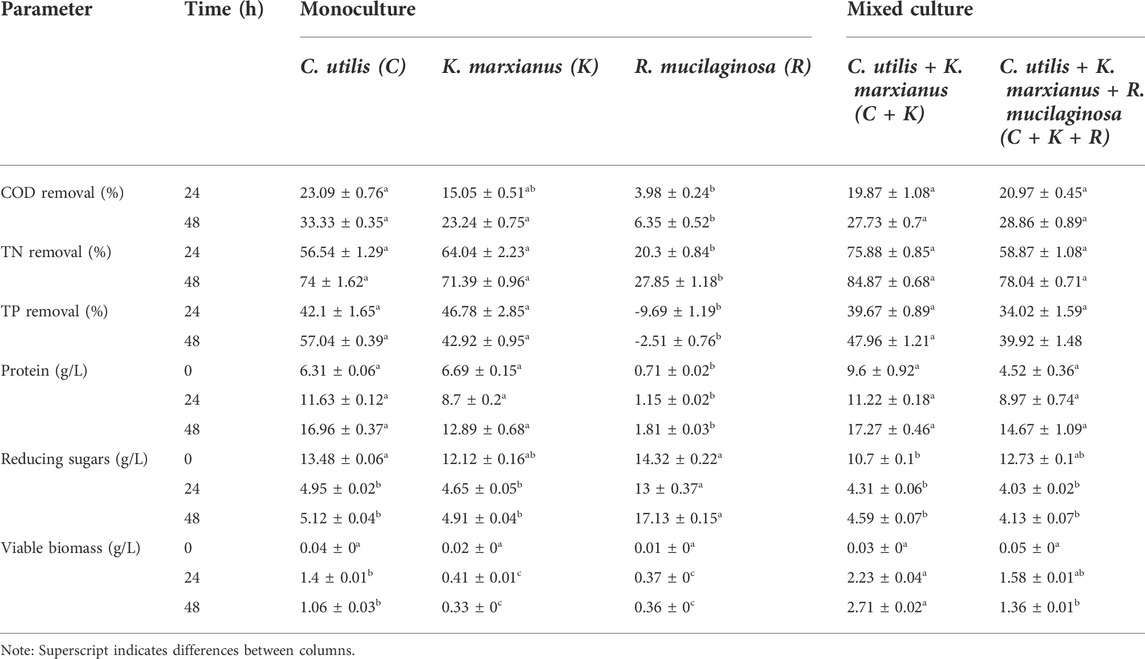
TABLE 1. Pollutant removal and revalorization potential of TV through five different yeast culture treatments.
Among the monocultures, C. utilis displayed the highest COD removal rate, reaching 33% after 48 h, followed by K. marxianus at 23%. R. mucilaginosa reached a COD removal rate of only 6% after 48 h, which correlates with the overall poor biomass and protein accumulation displayed by this yeast species. Although it has previously been reported that R. mucilaginosa is able to withstand and degrade phenolic compounds in wastewaters (Jarboui et al., 2012), the increased acidity and phenolic composition of the TV could have impaired its growth. Future research should, therefore, address phenolic composition during TV fermentation to understand its influence on yeast growth. The C + K and C + K + R mixed cultures reached similar COD removal efficiencies of 28% and 29%, respectively. This indicates that the influence of R. mucilaginosa in the mixed culture was minimal due to the low levels of this species detected after 24 h. Additionally, the C + K and C + K + R mixed cultures displayed lower COD removal capabilities compared to the C. utilis monoculture, potentially as a result of competition between the species for the easy-access nutrients and carbon sources (Viljoen, 2006). Likewise, the presence of specific metabolites within the media can trigger predatory behavior in yeast, such as in Rhodotorula sp. and Candida sp. (Lachance & Pang, 1997).
The highest TN removal rate within the monocultures was displayed by C. utilis (74% after 48 h), followed by K. marxianus (71% after 48 h) and R. mucilaginosa (28% after 48 h). However, the C + K and C + K + R mixed cultures reached TN removal efficiencies of 85% and 78%, respectively, after 48 h. These increased TN removal rates of the mixed cultures compared to the monocultures indicates that the interaction between C. utilis and K. marxianus potentially promoted a high utilization of nitrogen. This elevated nitrogen uptake can be correlated with an increase in biomass and protein accumulation in the C + K treatment. This may be attributed to extracellular substrate degradation mechanisms, which increase assimilable nitrogen concentrations within the media, thereby increasing nitrogen uptake for all species (Boze et al., 1992; Viljoen, 2006). However, the introduction of R. mucilaginosa resulted in a decreased TN removal rate, which was lower in the C + K + R culture compared to the C + K culture. This could potentially be due to competition for carbon and nutrients, along with predatory behavior among the cultured yeast species (Lachance & Pang, 1997; Viljoen, 2006). Following the same trend as observed for COD and TN, the C. utilis monoculture showed the highest TP removal rate (57% after 48 h) followed by K. marxianus (43% after 48 h). Conversely, R. mucilaginosa displayed an increase in the TP content, resulting in negative removal rates (Table 1), which could be due to the release of intracellular phosphorus after cellular death. Furthermore, the C + K and C + K + R mixed cultures displayed TP removal efficiencies of 48% and 40% after 48 h, respectively, which were lower than the C. utilis monoculture. Similar to COD, the interaction between strains potentially inhibited TP removal. The protein content in the C. utilis monoculture was the highest (17 mg/L) after 48 h, followed by K. marxianus (13 mg/L), and R. mucilaginosa (1.8 mg/L). The C + K mixed culture displayed a similar protein content (17.3 mg/L) to that of the C. utilis monoculture, while the C + K + R culture displayed a lower protein content (of 14.7 mg/L). Lower protein levels reduce the nutritional value of the recovered biomass that can be used as an animal feed supplement and, thus, the economic viability for the proposed circular bioeconomy scheme.
When evaluating reducing sugar utilization, the C. utilis and K. marxianus monocultures displayed similar behaviors. The initial reducing sugar concentration (13.48 and 12.12 mg/L, respectively) was reduced to more than half (4.95 and 4.65 mg/L) after 24 h and then remained at a similar level (5.12 and 4.91 mg/L) after 48 h. The R. mucilaginosa monoculture, however, displayed a fluctuating reducing sugar trend, as reducing sugars decreased from 14.3 mg/L (initial concentration) to 13 mg/L after 24 h, and then raised to 17.1 mg/L after 48 h. This indicates sugar liberation, likely due to cellular death and the subsequent release of intracellular components (Alexandre & Guilloux-Benatier, 2006). However, a total sugar assay must be performed to further understand the changes in the sugar composition of the media. The mixed cultures (C + K and C + K + R) displayed very similar trends to one another, with initial concentrations of 10.7 and 12.73 mg/L, respectively, which were reduced to 4.3 and 4.0 mg/L after 24 h, and then slightly increased to 4.6 mg/L and 4.1 mg/L, respectively, after 48 h. The sugar utilization by both mixed cultures was likewise very similar to that of the C. utilis and K. marxianus monocultures. All culture treatments, except for the R. mucilaginosa monoculture, displayed stable reducing sugar concentrations between the 24–48 h period. During this same time frame, however, protein concentrations increased significatively, which can be attributed to the degradation and uptake of complex carbon sources by the cultured yeast once the easy-access carbon sources are depleted. Although reducing sugar concentrations remained stable, the COD was consumed during the 24–48 h period. The viable biomass in the C. utilis monoculture was the highest among the three monocultures, reaching a concentration of 1.06 mg/L after 48 h. The viable biomass concentration in the K. marxianus and R. mucilaginosa monoculture were very similar to one another with values of 0.33 and 0.36 mg/L after 48 h, respectively. The mixed cultures displayed a greater viable biomass compared to monocultures. The C + K and C + K + R treatments displayed biomass contents of 2.71 and 1.36 mg/L after 48 h, which was 155% and 28% higher than that of the C. utilis monoculture, respectively.
Overall, C. utilis and K. marxianus demonstrated the best potential to grow using TV as substrate while simultaneously removing pollutants (COD, TN and TP) in order to synthesize protein-rich biomass as previously reported by (Díaz-Vázquez et al., 2022). C. utilis has been reported to have robust fermentation characteristics and a high tolerance to environmental changes and can produce biomass rich in protein, vitamins, and amino acid diversity, which are ideal characteristics for an animal feed protein supplement (Buerth et al., 2016; Sousa-Silva et al., 2021). Furthermore, C. utilis has the capacity to assimilate carbon (organic acids, alcohol, propionaldehyde, acetaldehyde) and nitrogen (nitrate, nitrite, ammonium hydroxide, amino acids) sources (Buerth et al., 2016) producing high protein yields (dos Santos et al., 2019). Similarly, K. marxianus has been shown to possess important features for industrial exploitation, such as an ability to utilize a broad range of sugars, thermotolerance, secretion of lytic enzymes, and ethanol production by fermentation (Karim et al., 2020). The Kluyveromyces genus of ascomycetous yeasts has been classified as a potential candidate for animal feed supplementation since species of this genus are known to be able to produce protein-rich biomass with useful amino acid compositions (Fonseca et al., 2008). Conversely, in this study, R. mucilaginosa displayed a significatively lower growth and substrate consumption compared to the other two monoculture treatments. Species of the Rhodotorula genus, such as R. mucilaginosa, are known to have the capacity to degrade phenolic compounds (Jarboui et al., 2012) and reduce 50%–90% of the COD and BOD from phenolic-rich wastewaters (Shailubhai et al., 1985; Jarboui et al., 2012). However, in the present research, R. mucilaginosa displayed the lowest performance in terms of pollutant removal and biomass production rates within the monocultures. When comparing monocultures to mixed cultures, overall, the C. utilis monoculture demonstrated better performance (COD, TN and TP removal) than any other culture tested. However, the C + K culture displayed other important advantages, such as higher nitrogen removal and biomass generation rates, which are essential attributes for a potential livestock feed supplement. Therefore, when all of these results are taken into consideration, the C + K mixed culture seems very promising, especially considering that the differences between the C + K mixed culture and C. utilis monoculture were low in terms of COD and TP removal rates.
The fungal nuclear ribosomal internal transcribed spacer (ITS) region was characterized to further analyze the behavior of the yeast communitiess within the mixed cultures and to understand their correlation with physicochemical parameters during the fermentation of TV. 2,023,911 raw read sequences were obtained through high-throughput sequencing of the ITS2 region with a mean length of 329 bp. A total of 1,481,358 (73%) were classified as fungi through the Unite database, while the remainder were described as unclassified. Figure 1 shows the genus-level yeast communities’ proportions for the C + K and C + K + R cultures throughout the experiment and supports that there was no fungal contamination during fermentation, as no abundance greater than 1% was observed for any other microorganism. K. marxianus was dominant throughout the experiment with relative abundances of 67.9%, 70.3% and 60.7% within the C + K culture and 59.9%, 62.8% and 57.5% within the C + K + R at 0, 24 and 48 h, respectively. This was followed by Candida with abundances of 31.7%, 29.6%, 39.2% within the C + K culture and 26.5%, 37.06% and 42.4% within the C + K + R culture at 0, 24 and 48 h, respectively (Figure 1). Fittingly, K. marxianus has been reported to possess one of the fastest growth rates of all known eukaryotic microorganisms (Karim et al., 2020).
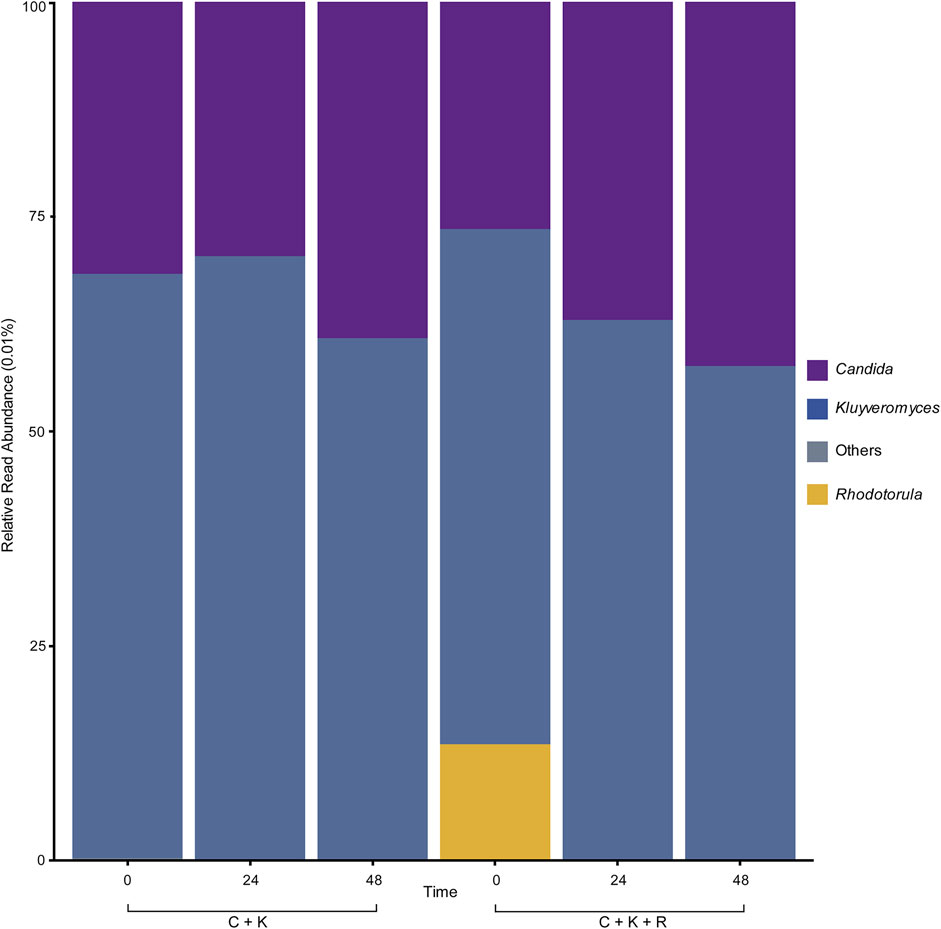
FIGURE 1. Genus-level relative read abundances of the mixed yeast cultures throughout the experiment. ‘Others’ refers to yeast genera with a relative read abundance of <1%.
These results indicate that both K. marxianus and C. utilis were able to adapt to TV as a growth medium. The relative abundance of K. marxianus and C. utilis within the C + K and C + K + R cultures remained consistent from 0 to 24 h. However, the relative abundance of C. utilis increased from 29.6% to 39.2% in the C + K culture and from 37.1% to 42.4% in the C + K + R culture after 24 h, while the relative abundance of K. marxianus decreased from 70.3% to 60.7% in the C + K culture and from 62.8% to 57.5% in the C + K + R culture. Even though K. marxianus was the most abundant species at all times evaluated, the metabolism of C. utilis appeared to be favored since it showed an increasing growth trend. Furthermore, while the pollutant removal and biomass production rates (Table 1) in both mixed cultures were higher than those of the monocultures alone, both mixed cultures displayed similar overall behaviors to the Candida monoculture. Although R. mucilaginosa possessed the ability to grow on TV when in monoculture, the results from the C + K + R mixed culture (where R. mucilaginosa was not detectable at either 24 h or 48 h) indicate that it´s adaptation to the TV growth media was weaker compared to K. marxianus and C. utilis. Additionally, the removal efficiencies and biomass generation rates for the C + K + R culture (Table 1) were lower than C + K mixed culture. These results suggest that R. mucilaginosa did not adapt well to the TV medium, and its interactions with other better adapted yeast species blocked its growth. The growth of R. mucilaginosa could have also been affected by the pH (3.0–5.0) and the presence of phenolic compounds in TV, as has previously been reported for the Rhodotorula genus of yeast (Rodríguez-Félix et al., 2018). Indeed, high concentrations of phenolic compounds can lead to precipitation of lignocellulose-hydrolyzing enzymes affecting fungi’s growth (Aksu & Eren, 2005; Rodríguez-Félix et al., 2018; Arreola et al., 2020). Additionally, the growth of R. mucilaginosa is known to be inhibited by catechol and gallic acid compounds (Jarboui et al., 2012).
A redundancy analysis (RDA) is presented in Figure 2. The angles between the blue vectors of the three yeast represent correlations between the yeast, and angles between the yellow vectors represent correlations between the physicochemical parameters. Likewise, angles between the blue and yellow vectors represent correlations between specific yeast and physicochemical parameters. Each point represents an observation or a sequenced sample and indicates the corresponding vector value. The origin represents the mean value, projections in the same direction as the vector indicate values above average, and projections in the opposite direction of the vector represent values below average. This analysis was performed to analyze correlations between the parameters (COD, TN, TP, RS, and P) and the C + K and C + K + P yeast cultures. The two main redundancy components explained 58% of the total variability. The RDA (Figure 2) showed a correlation between the reducing sugars and the TN concentrations. Moreover, an inverse correlation was observed between the COD and the total biomass concentrations. The consumption of reducing sugars (Table 1) in all cultures was observed after 24 h and then decreased after 48 h, except for the R. mucilaginosa monoculture, where the sugars increased[p Additionally, the relative abundances of K. marxianus and C. utilis in the C + K culture (Figure 1) were almost equal at 0 and 24 h and significantly increased after 48 h. These results indicate that in the first growth phase (before 24 h), the preferred substrates were simple sugars and nitrogen, and biomass generation was inhibited in the mixed cultures. Then, in a second growth phase (between 24 and 48 h), the preferred carbon source shifted to more complex organic compounds (expressed as COD), nitrogen was consumed at lower rates, and higher biomass generation was promoted. Additionally, as shown through the RDA (Figure 2), C. utilis and K. marxianus displayed an inverse correlation to TP concentration, indicating that both strains participated in phosphorus metabolization. However, no significant increase was observed in TP removal in mixed cultures compared to the monocultures.
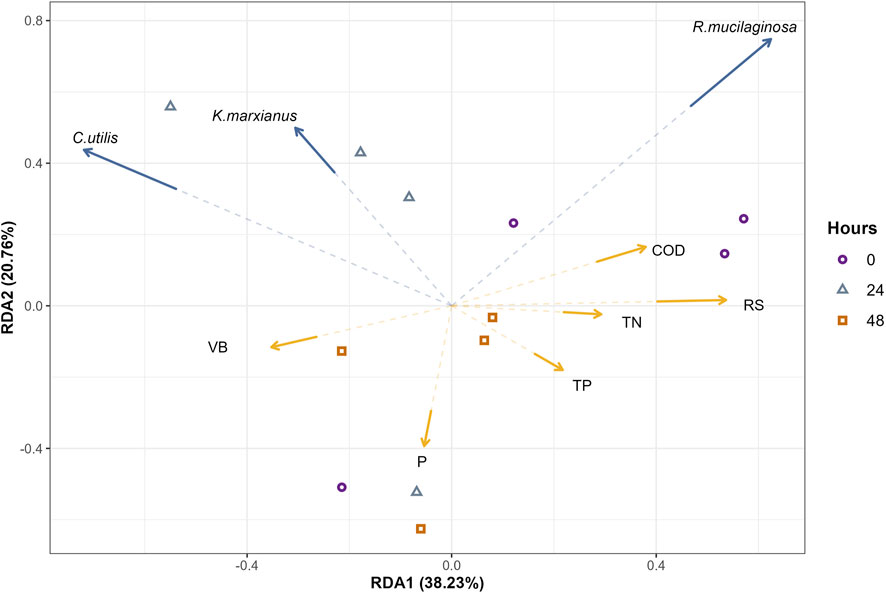
FIGURE 2. Redundancy analysis (RDA) showing correlations between yeast and physicochemical parameters. Yellow vectors represent the physicochemical parameters and blue vectors represent yeast communities.
Figure 3 presents a comparison of the amino acid compositions of the spray dried biomass recovered from the mono (C, K and R) and mixed (C + K and C + K + R) yeast cultures for potential application as a livestock feed protein supplement. Both mixed culture treatments (C + K and C + K + R) displayed the highest amino acid concentrations among the yeast culture treatments with 16.46 and 16.34 g 100 g DW−1 (dry weight) for C + K and C + K + R, respectively. In comparison, the monoculture treatments displayed significantly lower (p <0.05) amino acid concentrations of 12.56, 9.97 and 8.10 g 100 g DW−1 for the respective K, R, and C treatments. Several specific amino acids were also significatively higher in the mixed cultures compared to the monocultures. For example, the threonine content in the mixed cultures (1,057.7 and 918.1 mg 100 g DW−1 for C + K and C + K + R, respectively) was significatively higher (p <0.05) than those of the monoculture treatments (715.5, 502.5 and 362.8 for K, R, and C treatments, respectively). Similarly, the tyrosine and alanine concentrations in the mixed cultures were significatively higher than those of the monocultures (639.49 and 635.03 mg 100 g DW−1 of tyrosine and 735.30 and 635.70 mg 100gDW−1 of alanine within the C + K and C + K + R, respectively). This increased content of total and specific amino acids in the mixed cultures can significatively improve the nutritional potential and economic value of the recovered biomass compared to the monocultures. Threonine, arginine, cysteine, glutamine, lysine, valine, methionine, isoleucine and tryptophan supplementation in ruminants, poultry and swine has been associated with an increase in macrophage activation, T and natural killer lymphocyte proliferation, and antibody production, which are all key indicators of a healthy immune response (Montout et al., 2021). Increases in milk protein and fat content in dairy cows, muscle mass and fat accumulation in swine, and egg mass in poultry have likewise been linked to protein and specific amino acid supplementation (Harms & Russell, 1993; Doepel et al., 2004; Cheng et al., 2017). Therefore, specific amino acid supplementation offers multiple benefits to livestock producers, such as increasing the quality of the meat and reducing production costs.
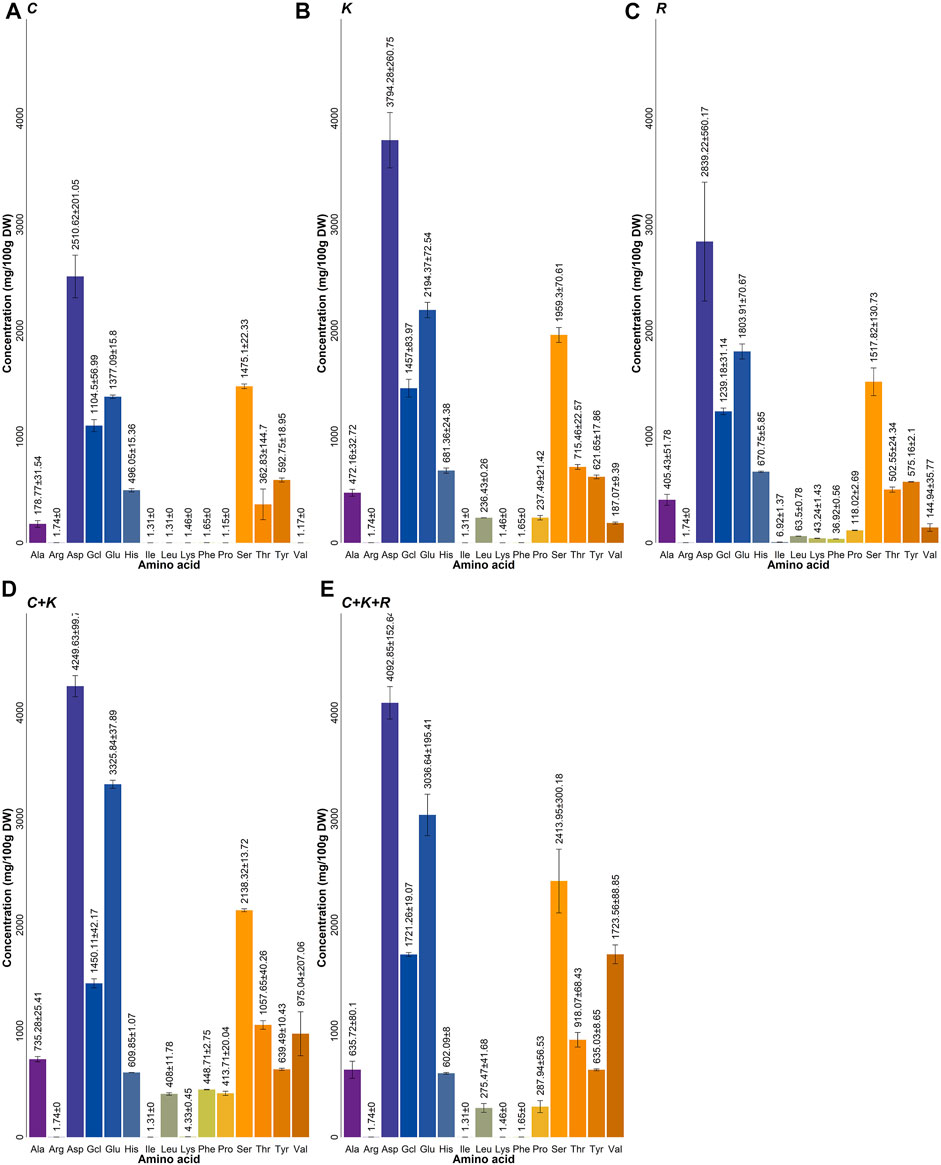
FIGURE 3. Amino acid composition of the yeast culture treatments. (A), C. utilis; (B), K. marxianus; (C), R. mucilaginosa; (D), Candida utilis + K. marxianus; (E), C. utilis + K. marxianus + R. mucilaginosa.
The amino acid profiles of the two mixed cultures (Figures 3D,E) show that the proline, leucine, and glutamic acid contents in the recovered biomass were significantly higher in the C + K culture compared to the C + K + R treatment. Conversely, the valine, glycine and serine contents were significantly higher in the C + K + R culture compared to the C + K culture. No significant differences (p <0.05) were observed in the arginine content among these treatments. Although the R. mucilaginosa population in the C + K + R mixed culture rapidly decreased after the initial inoculation, significant differences in amino acid composition were still identified between the C + K and C + K + R treatments. These differences can be explained by metabolic changes that result from the release of intracellular components during the cellular death of R. mucilaginosa (Viljoen, 2006). Easy-access TN and TP could have increased in availability during the death of the remaining R. mucilaginosa population, reducing the degradation and intake of complex TN and TP forms and thereby decreasing TN and TP removal rates in the C + K + R culture during the 24 and 48 h time points (Table 1), as compared to the C + K culture where R. mucilaginosa was absent. Ammonia is one of the main forms of nitrogen that can be easily assimilated by yeast (Boze et al., 1992). However, the TN present in TV is mainly in complex organic forms, which require degradation prior to assimilation (España-Gamboa et al., 2011). Therefore, the presence of easy-access nitrogen sources derived from disrupted cells could have negatively affected TN and TP removal rates in the C + K + R treatment.
Yeasts offer a promising way to maximize the production of desired amino acids in TV, and the specific amino acid content can be heavily influenced by the unique yeast communities present in the media. Therefore, carefully considering communities composition is a key factor for the optimization of this TV treatment method. Specific livestock species have particular amino acid feed needs in order to increase both the immune response of the living animals and the final product quality (Harms & Russell, 1993; Cheng et al., 2017; Park et al., 2020; Montout et al., 2021). Therefore, mixed yeast cultures may offer new alternatives to meet specific amino acid demands in the livestock industry, while simultaneously allowing for the reintegration of bio-based waste and nutrients into the value chain, following the principals of a circular bioeconomy (Carus & Dammer, 2018). The resulting biomass can be exploited for cattle feed supplementation with an estimated potential to produce 45,664 tons of protein feed yearly, based on the current tequila vinasses generated in the state of Jalisco (Díaz-Vázquez et al., 2021) and the protein composition determined for the C + K treatment in the present study.
The results presented in this experiment show that C. utilis and K. marxianus can successfully use tequila vinasse as a growth medium. Of all the treatments tested (mixed- and monocultures), C. utilis displayed the greatest ability to remove contaminants. However, the C + K mixed culture demonstrated the highest biomass and protein generation rates, characteristics which are highly advantageous for cattle feed supplement production. Two distinct growth phases were distinguished within the C + K mixed culture. During the first phase (before 24 h), simple sugars and nitrogen were the main substrates consumed and biomass generation was prominent compared to the second phase (between 24 and 48 h), where more complex organic compounds (COD) were consumed while lower nitrogen consumption rates were observed, and higher biomass generation was promoted. Additionally, even though R. mucilaginosa did not influence the C + K + R mixed culture in terms biomass, protein generation or pollutant removal, the amino acid composition was nonetheless significantly altered compared to the C + K culture without R. mucilaginosa. These findings offer an opportunity for the maximization of desired amino acid content based on the presence of specific yeast species in the media.
Yeast cultures offer a promising alternative way to treat and revalorize TV, and the resulting biomass can be exploited for cattle feed supplementation with an estimated potential to produce 45,664 tons of protein feed yearly, based on the current generation of TV in the state of Jalisco. Further work is required regarding the influence of native microbial populations and their influence in pollutant degradation in unsterilized vinasses to assess the technical and economic feasibility of the presented approach. This work presents an innovative and cost-effective solution for the integrated management of TV, however, additional research is needed to assess the technological and environmental feasibility, commercial quality of the produced feed as well as the financial sustainability of this approach.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA866992.
Conceptualization; DD-V, and MSG-H; data curation; DD-V, and MG, formal analysis; DD-V, and MSG-H; funding acquisition; MSG-H investigation; DD-V, and MSG-H; methodology; DD-V, MG, and AF, project administration; CS-G, resources; MSG-H; software; MG supervision; CS-G, and MSG-H; validation; CS-G, and MSG-H, visualization; DD-V, and MG roles/writing—original draft; DD-V, MG and AF writing—review and editing. DO-N, and MSG-H.
This research was funded by the Innovation, Science and Technology Ministry of the State of Jalisco (SICYT by its acronym in Spanish) and the Science and Technology State Council (COECyTJAL by its acronym in Spanish) through Jalisco’s Scientific Development Found (FODECIJAL by its acronym in Spanish), assigned as the Project 7947: “Vinazas tequileras como sustratos para la produccion ´de aditivos alimenticios de alto valor proteico para animales monogastricos.´”
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abarenkov, K., Zirk, A., Piirmann, T., Pöhönen, R., Ivanov, F., Nilsson, R. H., et al. (2021). UNITE QIIME release for Fungi Application/gzip. Tartu, Estonia: UNITE Community. doi:10.15156/BIO/1264708
Aksu, Z., and Eren, A. T. (2005). Carotenoids production by the yeast Rhodotorula mucilaginosa: Use of agricultural wastes as a carbon source. Process Biochem. 40 (9), 2985–2991. doi:10.1016/j.procbio.2005.01.011
Alexandre, H., and Guilloux-Benatier, M. (2006). Yeast autolysis in sparkling wine – A review. Aust. J. Grape Wine Res. 12 (2), 119–127. doi:10.1111/j.1755-0238.2006.tb00051.x
Arreola, A. R., Tizapa, M. S., Zurita, F., Morán-Lázaro, J. P., Valderrama, R. C., Rodríguez-López, J. L., et al. (2020). Treatment of tequila vinasse and elimination of phenol by coagulation–flocculation process coupled with heterogeneous photocatalysis using titanium dioxide nanoparticles. Environ. Technol. 41 (8), 1023–1033. doi:10.1080/09593330.2018.1518994
Bedoić, R., Ćosić, B., and Duić, N. (2019). Technical potential and geographic distribution of agricultural residues, co-products and by-products in the European Union. Sci. Total Environ. 686, 568–579. doi:10.1016/j.scitotenv.2019.05.219
Bokulich, N. A. (2018). Fungal ITS analysis tutorial, QIIME 2 forum. Available at: https://forum.qiime2.org/t/fungal-its-analysis-tutorial/7351 (Accessed June, 2022).
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37 (8), 852–857. doi:10.1038/s41587-019-0209-9
Boze, H., Moulin, G., and Galzy, P. (1992). Production of food and fodder yeasts. Crit. Rev. Biotechnol. 12 (1–2), 65–86. doi:10.3109/07388559209069188
Buerth, C., Tielker, D., and Ernst, J. F. (2016). Candida utilis and Cyberlindnera (Pichia) jadinii: Yeast relatives with expanding applications. Appl. Microbiol. Biotechnol. 100 (16), 6981–6990. doi:10.1007/s00253-016-7700-8
Buitrón, G., Kumar, G., Martinez-Arce, A., and Moreno, G. (2014). Hydrogen and methane production via a two-stage processes (H2-SBR + CH4-UASB) using tequila vinasses. Int. J. Hydrogen Energy 39 (33), 19249–19255. doi:10.1016/j.ijhydene.2014.04.139
Carus, M., and Dammer, L. (2018). The circular bioeconomy—concepts, opportunities, and limitations. Ind. Biotechnol. 14 (2), 83–91. doi:10.1089/ind.2018.29121.mca
CDBB. (2021). Colección de Cultivos microbianos. Available at: http://cdbb.cinvestav.mx/cdbb/files/1683/index.html (Accessed July, 2022).
Cheng, C., Liu, Z., Zhou, Y., Wei, H., Zhang, X., Xia, M., et al. (2017). Effect of oregano essential oil supplementation to a reduced-protein, amino acid-supplemented diet on meat quality, fatty acid composition, and oxidative stability of Longissimus thoracis muscle in growing-finishing pigs. Meat Sci. 133, 103–109. doi:10.1016/j.meatsci.2017.06.011
CRT. (2019). Estadisticas CRT. Available at: https://www.crt.org.mx/files/949/EstadisticasCRTweb.html (Accessed January, 2022).
Díaz-Vázquez, D., Carrillo-Nieves, D., Orozco-Nunnelly, D. A., Senés-Guerrero, C., and Gradilla-Hernández, M. S. (2021). An integrated approach for the assessment of environmental sustainability in agro-industrial waste management practices: The case of the tequila industry. Front. Environ. Sci. 9. 682093. doi:10.3389/fenvs.2021.682093
Díaz-Vázquez, D., Orozco-Nunnelly, D. A., Yebra-Montes, C., Senés-Guerrero, C., and Gradilla-Hernández, M. S. (2022). Using yeast cultures to valorize tequila vinasse waste: An example of a circular bioeconomy approach in the agro-industrial sector. Biomass Bioenergy 161, 106471. doi:10.1016/j.biombioe.2022.106471
Dixon, P. (2003). VEGAN, a package of R functions for community ecology. J. Veg. Sci. 14 (6), 927–930. doi:10.1111/j.1654-1103.2003.tb02228.x
Doepel, L., Pacheco, D., Kennelly, J. J., Hanigan, M. D., López, I. F., and Lapierre, H. (2004). Milk protein synthesis as a function of amino acid supply. J. Dairy Sci. 87 (5), 1279–1297. doi:10.3168/jds.S0022-0302(04)73278-6
DOF. (1997). NOM-001-SEMARNAT-1996, NOM-002-SEMARNAT-1996 y NOM-003-SEMARNAT-1997. Available at: http://www.conagua.gob.mx/CONAGUA07/Publicaciones/Publicaciones/SGAA-15-13/files/1366/SGAA-15-13.pdf (Accessed June, 2022).
DOF. (2021). NOM-001-SEMARNAT-2021. Available at: https://www.dof.gob.mx/nota_detalle.php?codigo=5645374&fecha=11/03/2022#gsc.tab=0 (Accessed June, 2022).
dos Santos, J. F., Canettieri, E. V., Souza, S. M. A., Rodrigues, R. C. L. B., and Martínez, E. A. (2019). Treatment of sugarcane vinasse from cachaça production for the obtainment of Candida utilis CCT 3469 biomass. Biochem. Eng. J. 148, 131–137. doi:10.1016/j.bej.2019.04.009
Ercolini, D. (2013). High-throughput sequencing and metagenomics: Moving forward in the culture-independent analysis of food microbial ecology. Appl. Environ. Microbiol. 79 (10), 3148–3155. doi:10.1128/AEM.00256-13
España-Gamboa, E., Mijangos-Cortes, J., Barahona-Perez, L., Dominguez-Maldonado, J., Hernández-Zarate, G., and Alzate-Gaviria, L. (2011). Vinasses: Characterization and treatments. Waste Manag. Res. 29 (12), 1235–1250. doi:10.1177/0734242X10387313
Fonseca, G. G., Heinzle, E., Wittmann, C., and Gombert, A. K. (2008). The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 79 (3), 339–354. doi:10.1007/s00253-008-1458-6
Gill, A. (2017). The importance of bacterial culture to food microbiology in the age of genomics. Front. Microbiol. 8, 777. doi:10.3389/fmicb.2017.00777
Harms, R. H., and Russell, G. B. (1993). Optimizing egg mass with amino acid supplementation of a low-protein diet. Poult. Sci. 72 (10), 1892–1896. doi:10.3382/ps.0721892
Jarboui, R., Baati, H., Fetoui, F., Gargouri, A., Gharsallah, N., and Ammar, E. (2012). Yeast performance in wastewater treatment: Case study of Rhodotorula mucilaginosa. Environ. Technol. 33 (8), 951–960. doi:10.1080/09593330.2011.603753
Karim, A., Gerliani, N., and Aïder, M. (2020). Kluyveromyces marxianus: An emerging yeast cell factory for applications in food and biotechnology. Int. J. Food Microbiol. 333, 108818. doi:10.1016/j.ijfoodmicro.2020.108818
Lachance, M.-A., and Pang, W.-M. (1997). Predacious yeasts. Yeast 13 (3), 225–232. doi:10.1002/(SICI)1097-0061(19970315)13:3<225::AID-YEA87>3.0.CO;2-I
López-López, A., Davila-Vazquez, G., León-Becerril, E., Villegas-García, E., and Gallardo-Valdez, J. (2010). Tequila vinasses: Generation and full scale treatment processes. Rev. Environ. Sci. Biotechnol. 9 (2), 109–116. doi:10.1007/s11157-010-9204-9
Montout, L., Poullet, N., and Bambou, J.-C. (2021). Systematic review of the interaction between nutrition and immunity in livestock: Effect of dietary supplementation with synthetic amino acids. Animals 11 (10), 2813. doi:10.3390/ani11102813
Nouroozi, R. V., Noroozi, M. V., and Ahmadizadeh, M. (2015). Determination of protein concentration using Bradford microplate protein quantification assay. Dis. Diagnosis 4 (1), 11–17. doi:10.31661/iejm158
Park, J. K., Yeo, J.-M., Bae, G.-S., Kim, E. J., and Kim, C.-H. (2020). Effects of supplementing limiting amino acids on milk production in dairy cows consuming a corn grain and soybean meal-based diet. J. Anim. Sci. Technol. 62 (4), 485–494. doi:10.5187/jast.2020.62.4.485
Rodríguez-Félix, E., Contreras-Ramos, S. M., Davila-Vazquez, G., Rodríguez-Campos, J., and Marino-Marmolejo, E. N. (2018). Identification and quantification of volatile compounds found in vinasses from two different processes of tequila production. Energies 11 (3), 490. doi:10.3390/en11030490
Shailubhai, K., Rao, N. N., and Modi, V. V. (1985). Degradation of petroleum industry oil sludge by Rhodotorula rubra and Pseudomonas aeruginosa. Oil Petrochem. Pollut. 2 (2), 133–136. doi:10.1016/S0143-7127(85)90478-8
Sherwood, J. (2020). The significance of biomass in a circular economy. Bioresour. Technol. 300, 122755. doi:10.1016/j.biortech.2020.122755
Sousa-Silva, M., Vieira, D., Soares, P., Casal, M., and Soares-Silva, I. (2021). Expanding the knowledge on the skillful yeast cyberlindnera jadinii. J. Fungi (Basel). 7 (1), 36. doi:10.3390/jof7010036
Ugalde, U. O., and Castrillo, J. I. (2002). “Single cell proteins from fungi and yeasts,” in Applied mycology and biotechnology. Editors G. G. Khachatourians, and D. K. Arora (Elsevier), 2, 123–149. doi:10.1016/S1874-5334(02)80008-9
Viljoen, B. C. (2006). “Yeast ecological interactions. Yeast’Yeast, Yeast’Bacteria, Yeast’Fungi interactions and yeasts as biocontrol agents,” in Yeasts in food and beverages. Editors A. Querol, and G. Fleet (Springer), 83–110. doi:10.1007/978-3-540-28398-0_4
Wang, Y., Qiu, L., and Hu, M. (2018). Application of yeast in the wastewater treatment. E3S Web Conf. 53, 04025. doi:10.1051/e3sconf/20185304025
Wickham, H. (2016). Ggplot2: Elegant graphics for data analysis. 2nd ed. Heidelberg Germany: Springer International PublishingSpringer. doi:10.1007/978-3-319-24277-4
Wood, I. P., Elliston, A., Ryden, P., Bancroft, I., Roberts, I. N., and Waldron, K. W. (2012). Rapid quantification of reducing sugars in biomass hydrolysates: Improving the speed and precision of the dinitrosalicylic acid assay. Biomass and Bioenergy, 44, 117–121. doi:10.1016/j.biombioe.2012.05.003
Keywords: tequila vinasses, yeast community composition, Candida utilis, contaminant removal effectiveness, Kluyveromyces marxianus, Rhodotorula mucilaginosa
Citation: Díaz-Vázquez D, Garibay MV, Fernández del Castillo A, Orozco-Nunnelly DA, Senés-Guerrero C and Gradilla-Hernández MS (2022) Yeast community composition impacts on tequila industry waste treatment for pollution control and waste-to-product synthesis. Front. Chem. Eng. 4:1013873. doi: 10.3389/fceng.2022.1013873
Received: 07 August 2022; Accepted: 08 September 2022;
Published: 27 September 2022.
Edited by:
Jose A Perez-Pimienta, Autonomous University of Nayarit, MexicoReviewed by:
Jorge Arreola-Vargas, Texas A&M University, United StatesCopyright © 2022 Díaz-Vázquez, Garibay, Fernández del Castillo, Orozco-Nunnelly, Senés-Guerrero and Gradilla-Hernández. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Misael Sebastián Gradilla-Hernández, bXNncmFkaWxsYUB0ZWMubXg=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.