 Jing Pei1†
Jing Pei1† Jing Li
Jing Li- 1Department of Otolaryngology, Head and Neck Surgery, The Affiliated Jiangning Hospital With Nanjing Medical University, Nanjing, Jiangsu, China
- 2Department of Otorhinolaryngology Head and Neck Surgery, Northern Jiangsu People’s Hospital Affiliated to Yangzhou University, Yangzhou, Jiangsu, China
- 3Department of Blood Purification Center, Northern Jiangsu People’s Hospital Affiliated to Yangzhou University, Yangzhou, Jiangsu, China
Basic research on chronic rhinosinusitis (CRS) has advanced significantly in the past two decades, yet a comprehensive understanding of its pathogenic mechanisms remains elusive. Concurrently, there is a growing interest among scientists in exploring the involvement of autophagy in various human diseases, including tumors and inflammatory conditions. While the role of autophagy in asthma has been extensively studied in airway inflammatory diseases, its significance in CRS with or without nasal polyps (NPs), a condition closely linked to asthma pathophysiology, has also garnered attention, albeit with conflicting findings across studies. This review delves into the role of autophagy in CRS, suggesting that modulating autophagy to regulate inflammatory responses could potentially serve as a novel therapeutic target.
Introduction
Chronic rhinosinusitis (CRS) is a highly heterogeneous disease characterized by long-term and recurrent mucosal inflammation of the nasal cavity and sinuses, presenting symptoms such as nasal congestion, rhinorrhea, headache, facial pressure, and hyposmia (Subspecialty Group of Rhinology, Editorial Board of Chinese Journal of Otorhinolaryngology Head and Neck Surgery, and Subspecialty Group of Rhinology, Society of Otorhinolaryngology Head and Neck Surgery, Chinese Medical Association, 2019; Fokkens et al., 2020) (Figure 1). Nasal polyps (NPs) are structures resembling lychee fruits that form due to swelling and hyperplasia of the nasal mucosa after prolonged inflammatory stimulation. Epidemiological studies indicate that approximately one-third of CRS patients also have NPs, leading to the diagnosis of chronic rhinosinusitis with nasal polyps (CRSwNP) (Chee et al., 2022; Sedaghat et al., 2022; Starry et al., 2022). Pathological subtyping of CRSwNP based on the infiltration of inflammatory cells in the polyp tissue lacks uniform international criteria. Eosinophilic chronic rhinosinusitis with nasal polyp (eCRSwNP) is suggested to have a diagnostic criterion of eosinophils equal to or greater than 10 per high magnification field of view (Fokkens et al., 2020). Other pathological types, such as lymphocyte-type, plasma cell-type, neutrophilic-type, and mixed CRSwNP, are collectively referred to as non-eosinophilic chronic rhinosinusitis with nasal polyp (non-eCRSwNP) (Subspecialty Group of Rhinology, Editorial Board of Chinese Journal of Otorhinolaryngology Head and Neck Surgery, and Subspecialty Group of Rhinology, Society of Otorhinolaryngology Head and Neck Surgery, Chinese Medical Association, 2019). Patients with CRSwNP, particularly eCRSwNP, generally exhibit a poorer prognosis and higher recurrence rates compared to those with chronic rhinosinusitis without polyps (CRSsNP) (Ho et al., 2018; Cai et al., 2023). eCRSwNP is typically associated with Th2 inflammation, while non-eCRSwNP and CRSsNP are commonly linked to Th1 inflammation and mixed (Th1 or Th17) type inflammation (Lin et al., 2023; Yang et al., 2023). The mechanisms underlying CRS are complex and involve interactions between the host immune system, microbiota, and environmental factors (Subspecialty Group of Rhinology, Editorial Board of Chinese Journal of Otorhinolaryngology Head and Neck Surgery, and Subspecialty Group of Rhinology, Society of Otorhinolaryngology Head and Neck Surgery, Chinese Medical Association, 2019; Fokkens et al., 2020; Goulioumis et al., 2023). Currently, targeted drugs focusing on Th2 type inflammatory factors like anti-IL-4 and anti-IL-5 have received approval in certain countries for treating CRSwNP. However, their limited use is attributed to high costs, specific indications, and uncertain long-term efficacy (Bachert et al., 2020). Thus, exploring new targets or pathways to regulate CRSwNP progression could lead to more efficient and cost-effective treatment options in clinical practice.

Figure 1. Mechanisms of autophagy in CRS.
Autophagy plays a crucial role in defending against external pathogenic microorganisms, removing aging and harmful substances from cells, and maintaining cellular homeostasis and function. It accomplishes this by delivering cytoplasmic components to lysosomes for degradation and amino acid recycling through macroautophagy, chaperone-mediated autophagy, and microautophagy pathways (Esclatine et al., 2009; Feng et al., 2014; Munz, 2014). Dysregulation of autophagy has been linked to various diseases such as cancer, neurodegenerative diseases, reproductive system diseases, and inflammatory diseases (Chandrasekaran et al., 2023; Debnath et al., 2023; Dong et al., 2023; Kirat et al., 2023). Recent research has highlighted the importance of autophagy in the immune system, where it aids in pathogen removal, immune cell differentiation, antigen presentation, and regulation of inflammatory responses (Germic et al., 2019; Munz, 2021; Shariq et al., 2023). The connection between airway inflammation and autophagy has been a focus of investigation, particularly in the context of asthma (Lee and Kim, 2019; Painter et al., 2020; Theofani and Xanthou, 2021; Wang et al., 2021). Autophagy has been shown to have both protective and detrimental effects in allergic asthma, with a balancing role to prevent excessive lung tissue damage while mounting a protective anti-pathogen response (Theofani and Xanthou, 2021). Moreover, autophagy has been identified as a key regulator of asthma airway remodeling, influencing airway wall thickening and rigidity, and can also contribute to mitigating fibrosis by degrading extracellular matrix (ECM) proteins (Herrera et al., 2018).
Due to the close relationship and similarities between upper and lower respiratory tract inflammation, otolaryngologists have recently started investigating the potential role of autophagy in CRS. As research progresses, there have been some controversies regarding the involvement of autophagy in CRS. This paper aims to summarize the existing studies on this topic in hopes of guiding future research.
Classical markers and pathways of autophagy
Autophagy, classified as a type II programmed cell death, is a crucial self-degradation process essential for energy maintenance during development and in response to stress. This process can be either selective or non-selective (Dikic and Elazar, 2018). The core of autophagy lies in intracellular membrane rearrangement, which occurs in four main stages: extension of a phagocytic bubble, formation of isolated membranes and autophagosomes, fusion of autophagosomes with lysosomes, and cleavage of autophagosomes (Li et al., 2020; McAlinden et al., 2019). Various autophagy-related proteins, synthesized by autophagy-related genes (ATG), play a role in regulating and controlling the different stages of autophagy formation (Li et al., 2020).
LC3, also known as MAP1LC3, serves as a well-established autophagy marker throughout the autophagic process. In mammals, LC3 exists in three types: LC3A, LC3B, and LC3C, with LC3B being the most commonly utilized. When the LC3 protein is synthesized, the C-terminal 5-peptide is sheared off by Atg4 to produce LC3-I. During autophagy, LC3-I is modified and processed by a ubiquitin-like system including Atg7 and Atg3, which couples with phosphatidylethanolamine (PE) to form LC3-II and localize to the autophagosome’s inner and outer membranes. Upon fusion of autophagosome and lysosome, LC3-II on the outer membrane is cleaved by Atg4 to produce LC3-I for recycling; LC3-II on the inner membrane is degraded by lysosomal enzymes, resulting in low LC3 content in autophagic lysosomes (Kiriyama and Nochi, 2015). The ratio of LC3 II/LC3I is a widely used indicator of autophagic activity, where a higher ratio corresponds to increased autophagic activity.
In addition to LC3, changes in the expression of other autophagic substrates can also be used to monitor autophagic flow. Among these, p62 is commonly utilized. During autophagosome formation, p62 acts as a bridge linking LC3 and polyubiquitinated proteins, being selectively enclosed within autophagosomes. Subsequently, it undergoes degradation by proteohydrolases in autophagic lysosomes, leading to a negative correlation between the expression of the p62 protein and autophagic activity (Ichimura et al., 2008). Beclin-1, a key regulatory protein of autophagy encoded by the BECN1 gene, participates in the formation of autophagosome membranes. Therefore, elevated levels of Beclin-1 indicate heightened autophagic activity (Xu and Qin, 2019).
Several autophagy-related pathways have been identified, with notable classical pathways including the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) and Mitogen-Activated Protein Kinases (MAPK)/Extracellular Signal-Regulated Kinase (ERK) pathways that activate mammalian target of rapamycin protein (mTOR), as well as the Adenosine Monophosphate Activated Protein Kinase (AMPK) and p53 pathways that inhibit mTOR (Yang and Klionsky, 2010; Yu et al., 2018; Ba et al., 2019). mTOR serves as a crucial negative regulator of cellular autophagy, being activated in nutrient-sufficient conditions to suppress autophagy and inhibited during starvation or stress to promote autophagy (Wang and Zhang, 2019). ULK, possessing serine/threonine kinase activity, is the core protein of the autophagy signaling pathway. The ULK1 complex acts as a link between upstream mTOR and AMPK and downstream autophagosome formation. Activation of AMPK or inhibition of mTOR phosphorylates ULK1, thereby enhancing autophagy (Kim et al., 2011).
Autophagy phenomenon in tissues of CRS patients
Autophagy phenomenon in tissues of CRS patients remains a topic of debate in the scientific community (Table 1). While some studies suggest autophagic insufficiency or excess in CRS, the majority lean towards a deficiency in autophagy. For example, Western blot (WB) analysis indicated a significant decrease in LC3 expression and an increase in p-Akt and p-mTOR in nasal tissue specimens of CRS patients compared to normal nasal mucosae. Immunohistochemical (IHC) analysis further supported these findings, showing weaker LC3 and stronger p-mTOR staining in nasal polyps (Chen et al., 2015). Analogously, Wang L F. et al. analyzed the nasal mucosa from six CRSsNP patients and the NPs obtained from five CRSwNP patients via WB, showing that low LC3 but high cyclooxygenase-2 (COX-2) expression in NPs. IHC results showed that most NPs (17/23) had negative to weak LC3 staining intensity, while more than half of them (12/23) had moderate to strong COX-2 staining intensity. Besides, an inverse relationship between LC3 and COX-2 expression was identified using Kappa statistic, suggesting that insufficient autophagy may have contributed to increased inflammation (Wang et al., 2015). Compared with normal inferior turbinate mucosa (n = 20), the expression of LC3B and Beclin-1 were deficient and p62 was stronger in specimens of CRSwNP (n = 50) both in IHC and reverse transcription-polymerase chain reaction (RT-PCR) results (Qi et al., 2016). IHC with 35 nasal tissue sections also revealed that LC3 immunoreactivity was decreased in the NPs compared to the mucosa from patients with compensatory turbinate hypertrophy. In their IHC results, mTOR also had stronger positive signal (Simsek et al., 2021). Overall, the evidence points towards a deficiency in autophagy in CRS patients, as supported by various molecular analyses and IHC results. The levels of p-Akt and PI3K proteins were significantly higher in NPs compared to nasal tissues from sinus cyst patients, indicating a potential downregulation of autophagy in CRSwNP patients. Additionally, increased myeloperoxidase (MPO) activity, mRNA levels of hypoxia-inducible factor-1α (HIF-1α), and IL-17A in NPs suggest a link between hypoxia-induced neutrophil inflammation and inadequate autophagy (Cheng et al., 2022). Wang et al. (2022)’s study comparing various protein markers in different CRSwNP groups revealed an activated Akt-mTOR pathway and decreased levels of beclin-1 and LC3II in CRSwNP patients, with no significant differences between eCRSwNP and non-eCRSwNP groups. Interestingly, p62 levels did not vary significantly between groups in IHC experiments but were notably reduced in control tissues compared to eCRSwNP tissues in WB experiments. In addition, Transmission Electron Microscopy (TEM) showed less frequent formation of autophagosomes in both CRSwNP endotypes compared to control tissues. The study’s findings suggest a deficiency in autophagy in CRSwNP patients, regardless of endotype. Furthermore, they also investigated mitophagy-related markers (PINK1, BNIP3, and FUNDC1) with WB and tissue remodeling conditions using Masson’s trichrome (MT) and Periodic acid–Schiff Alcian blue (PAS–AB) staining, and they found that the levels of PINK1 and BNIP3 were all negatively correlated with collagen deposition in non-eCRSwNP patients, suggesting that mitophagy may play an important role in tissue remodeling in patients with non-eCRSwNP (Wang et al., 2022).
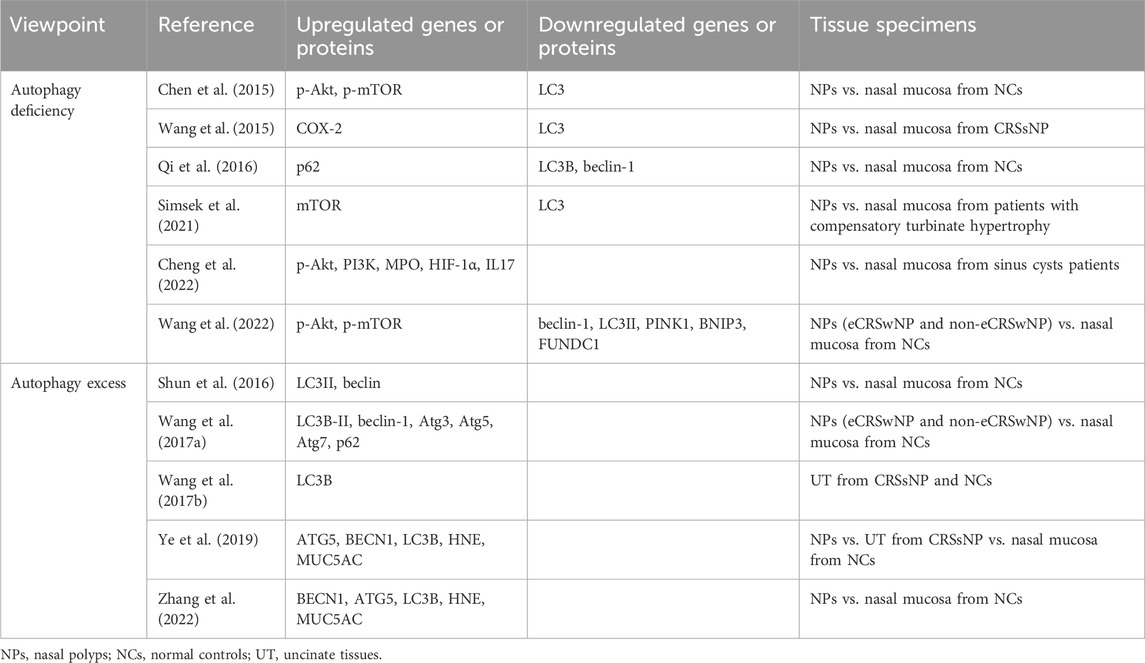
Table 1. Autophagy phenomenon in tissues of CRS patients.
Contrary to some beliefs, studies have shown that enhanced autophagy is present in patients with chronic rhinosinusitis (CRS). For example, WB results revealed increased expression of LC3II in 20 NPs specimens compared to 5 specimens of normal nasal mucosa, with IHC also indicating enhanced beclin levels in NPs, suggesting autophagic excess (Shun et al., 2016). Another study found elevated expression of autophagic proteins (LC3B-II, beclin-1, Atg3, Atg5, Atg7, and p62) and increased autophagic vacuole formation in both eCRSwNP and non-eCRSwNP tissues compared to normal controls (Wang et al., 2017a). Wang et al. (2017b) further investigated LC3B expression levels in uncinate tissues (UT) from CRSsNP patients and normal controls, revealing LC3B overexpression in CRSsNP patients. Ye et al. (2019) compared autophagic activity in polyp tissue from CRSwNP patients, UT from CRSsNP patients, and nasal mucosa from patients with simple septal deviation, showing increased mRNA levels of ATG5 and BECN1 in CRS, particularly in CRSwNP. Additionally, IHC analysis confirmed autophagic upregulation in CRS by comparing LC3B expression at the protein level in the three groups. Both human neutrophil elastase (HNE) and mucin 5AC (MUC5AC) were found to be upregulated simultaneously, suggesting that autophagy activation may promote HNE-induced mucus production (Ye et al., 2019). This was supported by another study showing increased mRNA levels of BECN1, ATG5, HNE, and MUC5AC by RT-PCR, as well as stronger positive signals of LC3B, HNE, and MUC5AC proteins in CRSwNP tissue samples compared to controls by IHC (Zhang et al., 2022).
Autophagy in cellular experiments and animal models
Autophagy marker levels in nasal mucosa or polyp samples from patients with CRS have been used to investigate the role of autophagy in the development of CRS and its underlying mechanisms (Table 2). Various cellular and animal experiments have been conducted by researchers to further understand this relationship.
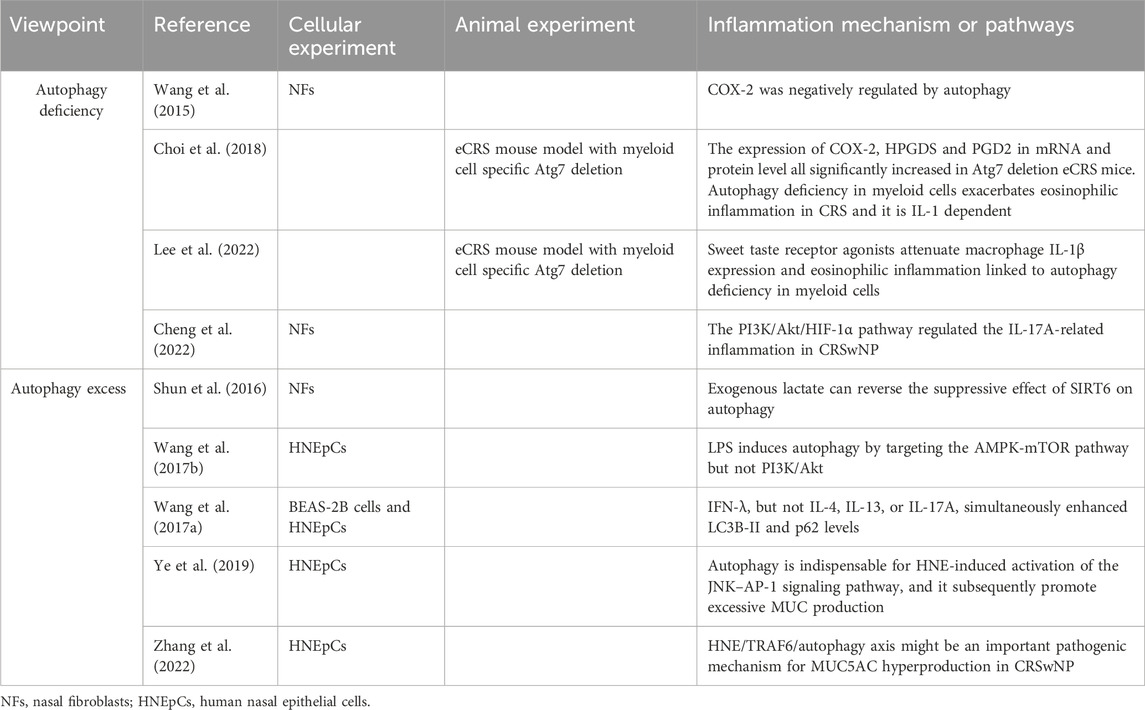
Table 2. Autophagy in cellular experiments and animal models.
Firstly, we summarize those studies in which CRS suffered from autophagy deficiency. Nasal fibroblasts (NFs) were isolated and cultured from NPs of CRSwNP patients (NPFs) and nasal mucosa of CRSsNP patients (NMFs). Autophagy induction through starvation and LC3 overexpression in both NFs led to a decrease in COX-2 expression, a common indicator of inflammation. Conversely, inhibiting autophagy with 3-methyladenine (3-MA) increased COX-2 expression. Interestingly, treatment with pro-inflammatory cytokines IL-1β and TNF-α did not significantly alter LC3 expression but did induce COX-2 expression in both NPFs and NMFs (Wang et al., 2015). These findings suggest that the chronic mucosal inflammation seen in CRSwNP may be due to a persistent autophagy deficiency, and manipulating autophagy could offer a promising therapeutic approach for CRSwNP. Choi et al. (2018) investigated the impact of impaired autophagy on eosinophilic inflammation in eosinophilic chronic rhinosinusitis (eCRS) mouse models through myeloid cell-specific deletion of Atg7. Their study revealed that autophagy deficiency in myeloid cells exacerbated eosinophilia, mucosal thickening, and epithelial hyperplasia in eCRS mice. While the counts of leukocytes, neutrophils, lymphocytes, basophils, and monocytes remained unchanged, there was a significant increase in eosinophil numbers in the blood of eCRS mice with impaired autophagy. Additionally, they explored the potential mechanisms and observed a substantial increase in the expression of COX-2, hematopoietic prostaglandin D synthase (HPGDS), and prostaglandin D2 (PGD2) at both mRNA and protein levels in Atg7 deletion eCRS mice. Moreover, the researchers demonstrated that blockade of the IL-1 receptor could alleviate eosinophilic inflammation, suggesting an IL-1 dependent pathway in eCRS due to autophagy deficiency. In a separate study, Lee et al. (2022) discovered that the anti-inflammatory effects of sweet taste receptor (STR) agonists, particularly trehalose, significantly mitigated eosinophilia, and disease pathogenesis in eCRS mice with autophagy deficiency in myeloid cells. Their mechanistic investigation revealed the involvement of T1R3 in reducing macrophage IL-1β production and eosinophilia in CRS, which was supported by genetic manipulation of T1R3 expression in macrophages and treatment with the T1R3 antagonist gurmarin. This research highlighted a previously overlooked anti-inflammatory target, offering a potential strategy for modulating autophagy in the treatment of eCRS (Lee et al., 2022). NFs isolated from CRSwNP patients were cultured to investigate the autophagy pathway. Upon stimulation with Tumor Necrosis Factor-alpha (TNF-α), levels of PI3K, p-AKT, HIF-1α, and IL-17A significantly increased in the fibroblasts. However, upon application of Wortmannin (a selective PI3K inhibitor), these indicators decreased significantly. This suggests that the PI3K/Akt/HIF-1α pathway may regulate IL-17A-related inflammation in CRSwNP (Cheng et al., 2022).
Several studies have indicated an increase in autophagy in CRS. Shun et al. (2016) discovered that Sirtuin 6 (SIRT6) expression was decreased while beclin was upregulated in NPFs. Additionally, under hypoxic conditions, the levels of LC3II in NPFs increased in a time-dependent manner, peaking at 12 h. Overexpression of SIRT6 was found to suppress hypoxia induced LC3II production in NPFs; however, the addition of exogenous lactate could reverse this suppressive effect on autophagy. In another study, human Nasal Epithelial Cells (HNEpCs) treated with lipopolysaccharide (LPS)—a common pathogenic element in CRS - showed dose- and time-dependent induction of autophagy. This was evidenced by increased levels of LC3B-II, decreased levels of p62 as observed through WB, and the presence of autophagosomes observed through TEM. Further exploration into the mechanism of autophagy involved Western blot analysis of proteins in the PI3K/Akt and AMPK pathways, revealing no significant changes in p-Akt, Akt, AMPK, and mTOR, but a notable increase in p-AMPK and decrease in p-mTOR. Treatment of HNEpC with Compound C—an AMPK inhibitor—after LPS exposure resulted in the inhibition of autophagy, indicating that LPS induces autophagy in HNEpC by targeting the AMPK-mTOR pathway (Wang et al., 2017b). Wang et al. (2017a) found that IFN-λ increased levels of autophagic proteins in both eCRSwNP and non-eCRSwNP tissues. Through cellular experiments, they discovered that IFN-λ enhanced LC3B-II and p62 levels in BEAS-2B cells and HNEpCs, while inhibitors like bafilomycin A1 led to increased levels of autophagic proteins. This suggests that IFN-λ induces activated but insufficient autophagy in CRSwNP. Autophagy activity was also found to be involved in regulating mucus secretion, with HNE inducing MUC5AC expression via the AP-1 pathway in primary HNEpCs. Inhibition of autophagy pathways and TRAF6 were shown to alleviate hypersecretion of MUC5AC, indicating the potential importance of the HNE/TRAF6/autophagy axis in CRSwNP pathogenesis (Zhang et al., 2022).
Transcriptome sequencing
Recent transcriptome sequencing has identified differentially expressed genes related to the autophagy pathway in CRS, suggesting a potential role of autophagy in the pathogenesis of CRS. For instance, Xiong et al. (2020) conducted transcriptome analysis and found that epithelial cell injury, autophagy, and the mTOR pathway (hsa04140 and hsa04150) may play a role in the development of non-eCRSwNP. Furthermore, bulk RNA sequencing and single cell RNA-seq datasets of epithelial cells (EpCs) from CRS patients revealed upregulation of mTOR complex 1 (mTORC1) activity and glycolytic activity in CRSwNP. Ex vivo inhibition experiments suggested that mTOR plays a critical role in EpC production of CXCL8, IL-33, and CXCL2. Enhanced mTORC1 signaling in CRSwNP EpCs was found to be associated with glycolysis, rather than mTORC2 (a typical PI3K/Akt/mTOR signature gene). Epithelial glycolysis can be induced by Th2 and Th17 cytokines in vitro and is correlated with Th2 cytokine response in vivo. These findings imply that CRSwNP may exhibit autophagy deficiency, with mTORC1 playing a significant role in its pathogenesis, potentially linked to glycolytic activity (Huang et al., 2023). Additionally, bulk and single cell RNA-seq data highlighted the importance of Migration Inhibitory Factor (MIF) and Transforming Growth Factor-Beta (TGF-β) pathways in mediating the effects of mitophagy in CRS, particularly emphasizing the role of MIF (Zhou et al., 2023).
Concluding remarks and future perspectives
From the overview above, autophagy displays both protective and detrimental effects on CRS. We speculate that autophagy is enhanced in the early stages of disease in CRS to remove cellular damage caused by pathogens or allergens to maintain organismal homeostasis, but as the damage persists, autophagy will be depleted and exhibit autophagic insufficiency. Current research leans towards autophagy deficiency in CRS, particularly in CRSwNP. However, the limited literature, small sample sizes in clinical studies, and scarcity of animal experiments highlight the need for further investigation. Studies have mainly focused on NFs from NPs tissues or control mucosa, as well as HNEpCs from healthy individuals, with conflicting findings on autophagy regulation. Most of the literature favors the presence of autophagy upregulation in HNEpCs and autophagy downregulation in NFs (Shun et al., 2016; Wang et al., 2017a; Wang et al., 2017b; Ye et al., 2019; Zhang et al., 2022). However, there is an exception which showed autophagy upregulation in NFs (Shun et al., 2016). Future in vitro experiments using both cell types could shed light on the differences in autophagy levels and their impact on CRS progression. The review focused on eCRS models, leaving room for further exploration of autophagy differences in non-eCRS models (Choi et al., 2018; Lee et al., 2022). Non-type 2 inflammation in CRS, represented by LPS, IFN-λ, and HNE-stimulated HNEpCs, often shows autophagy excess involving AMPK-mTOR, JNK–AP-1, and HNE-TRAF6 pathways (Wang et al., 2017a; Wang et al., 2017b; Ye et al., 2019). On the other hand, the PI3K/Akt/HIF-1α and COX-2/HPGDS/PGD2 pathway tends to regulate type-2 inflammation in CRS, leading to autophagy deficiency (Choi et al., 2018; Cheng et al., 2022). Interestingly, no significant differences in autophagy marker expression were observed between eCRSwNP and non-eCRSwNP tissue samples, although further confirmation with larger sample sizes is warranted (Wang et al., 2017a; Wang et al., 2022). The dual role of autophagy in CRS may vary among different cell types, as seen in lung inflammation. Myeloid lineage cells (especially neutrophils and eosinophils) and lymphoid lineage cells (e.g., CD4+, CD8+, ILC2, and B-cells) can enhance lung inflammation through autophagy modulation in distinct pathways, while epithelial cells and fibroblasts can influence lung remodeling via autophagy regulation (Painter et al., 2020). As current in vitro studies mainly focus on non-immune cells (HNEpCs and NFs), additional experiments are needed to understand the changes in autophagy levels in immune cells of CRS. The development of personalized autophagy modulators tailored to specific cell types and pathogenesis holds promise for future therapeutic interventions.
A recent study found that offspring of female mice exposed to OVA may reduce Th2 inflammation by enhancing autophagy in mast cells when exposed to OVA (Kim et al., 2021). Overexpression of Orosomucoid-like-3 (ORMDL3) increased the expression of SERCA2b, activating transcription factor 6 (ATF6), Beclin-1, and LC3BII. Conversely, inhibiting autophagy or knocking down ATF6 reversed the decrease in anaphylactic reaction caused by ORMDL3 overexpression (Li et al., 2021). Autophagy has also been linked to epithelial mesenchymal transition (EMT) and epigenetic changes in the fields of oncology, lung diseases, and diabetes (Ranieri et al., 2021; Zeng et al., 2021; Gundamaraju et al., 2022; Yu et al., 2024). Both EMT and epigenetic changes play roles in the pathogenesis of CRS (Li et al., 2022; Wang et al., 2023). These findings provide valuable insights for future research on CRS, highlighting the importance of further exploring the role of autophagy in this condition. While the current study is not without controversy, it sheds light on potential new therapeutic approaches for CRS by targeting autophagy.
Author contributions
JP: Investigation, Methodology, Software, Writing–original draft. ZD: Conceptualization, Formal Analysis, Methodology, Project administration, Writing–original draft. CJ: Formal Analysis, Methodology, Writing–original draft. YT: Formal Analysis, Investigation, Methodology, Software, Writing–original draft. HY: Data curation, Project administration, Supervision, Writing–review and editing. JL: Conceptualization, Data curation, Investigation, Methodology, Software, Writing–review and editing.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. The Project Supported by Science Foundation of Kangda College of Nanjing Medical University (No. KD2021KYJJZD027) and Nanjing Medical Science and Technology Development Fund (No. YKK22217).
Acknowledgments
We thank Kangda College of Nanjing Medical University and Nanjing Medical Science and Technology Development Fund for providing financial support to this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ba, L., Gao, J., Chen, Y., Qi, H., Dong, C., Pan, H., et al. (2019). Allicin attenuates pathological cardiac hypertrophy by inhibiting autophagy via activation of PI3K/Akt/mTOR and MAPK/ERK/mTOR signaling pathways. Phytomedicine 58, 152765. doi:10.1016/j.phymed.2018.11.025
Bachert, C., Zhang, N., Cavaliere, C., Weiping, W., Gevaert, E., and Krysko, O. (2020). Biologics for chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 145 (3), 725–739. doi:10.1016/j.jaci.2020.01.020
Cai, S., Lou, H., and Zhang, L. (2023). Prognostic factors for post-operative outcomes in chronic rhinosinusitis with nasal polyps: a systematic review. Expert Rev. Clin. Immunol. 19 (8), 867–881. doi:10.1080/1744666X.2023.2218089
Chandrasekaran, V., Hediyal, T. A., Anand, N., Kendaganna, P. H., Gorantla, V. R., Mahalakshmi, A. M., et al. (2023). Polyphenols, autophagy and neurodegenerative diseases: a review. Biomolecules 13 (8), 1196. doi:10.3390/biom13081196
Chee, J., Pang, K. W., Low, T., Wang, Y., and Subramaniam, S. (2022). Epidemiology and aetiology of chronic rhinosinusitis in Asia-A narrative review. Clin. otolaryngology official J. ENT-UK ; official J. Neth. Soc. Oto-Rhino-Laryngology Cervico-Facial Surg. 48 (2), 305–312. doi:10.1111/coa.13971
Chen, J. Y., Hour, T. C., Yang, S. F., Chien, C. Y., Chen, H. R., Tsai, K. L., et al. (2015). Autophagy is deficient in nasal polyps: implications for the pathogenesis of the disease. Int. Forum Allergy Rhinol. 5 (2), 119–123. doi:10.1002/alr.21456
Cheng, K. J., Zhou, M. L., Liu, Y. C., and Zhou, S. H. (2022). Roles played by the PI3K/Akt/HIF-1α pathway and IL-17a in the Chinese subtype of chronic sinusitis with nasal polyps. Mediat. Inflamm. 2022, 8609590. doi:10.1155/2022/8609590
Choi, G. E., Yoon, S. Y., Kim, J. Y., Kang, D. Y., Jang, Y. J., and Kim, H. S. (2018). Autophagy deficiency in myeloid cells exacerbates eosinophilic inflammation in chronic rhinosinusitis. J. Allergy Clin. Immunol. 141 (3), 938–950. doi:10.1016/j.jaci.2017.10.038
Debnath, J., Gammoh, N., and Ryan, K. M. (2023). Autophagy and autophagy-related pathways in cancer. Nat. Rev. Mol. Cell. Biol. 24 (8), 560–575. doi:10.1038/s41580-023-00585-z
Dikic, I., and Elazar, Z. (2018). Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell. Biol. 19 (6), 349–364. doi:10.1038/s41580-018-0003-4
Dong, H., Yang, W., Li, W., Zhu, S., Zhu, L., Gao, P., et al. (2023). New insights into autophagy in inflammatory subtypes of asthma. Front. Immunol. 14, 1156086. doi:10.3389/fimmu.2023.1156086
Esclatine, A., Chaumorcel, M., and Codogno, P. (2009). Macroautophagy signaling and regulation. Curr. Top. Microbiol. Immunol. 335, 33–70. doi:10.1007/978-3-642-00302-8_2
Feng, Y., He, D., Yao, Z., and Klionsky, D. J. (2014). The machinery of macroautophagy. Cell. Res. 24 (1), 24–41. doi:10.1038/cr.2013.168
Fokkens, W. J., Lund, V. J., Hopkins, C., Hellings, P. W., Kern, R., Reitsma, S., et al. (2020). European position paper on rhinosinusitis and nasal polyps 2020. Rhinology 58 (Suppl. S29), 1–464. doi:10.4193/Rhin20.600
Germic, N., Frangez, Z., Yousefi, S., and Simon, H. U. (2019). Regulation of the innate immune system by autophagy: monocytes, macrophages, dendritic cells and antigen presentation. Cell. Death Differ. 26 (4), 715–727. doi:10.1038/s41418-019-0297-6
Goulioumis, A. K., Kourelis, K., Gkorpa, M., and Danielides, V. (2023). Pathogenesis of nasal polyposis: current trends. Indian J. Otolaryngol. Head. Neck Surg. 75 (Suppl. 1), 733–741. doi:10.1007/s12070-022-03247-2
Gundamaraju, R., Lu, W., Paul, M. K., Jha, N. K., Gupta, P. K., Ojha, S., et al. (2022). Autophagy and EMT in cancer and metastasis: who controls whom? Biochim. Biophys. Acta Mol. Basis Dis. 1868 (9), 166431. doi:10.1016/j.bbadis.2022.166431
Herrera, J., Henke, C. A., and Bitterman, P. B. (2018). Extracellular matrix as a driver of progressive fibrosis. J. Clin. Investig. 128 (1), 45–53. doi:10.1172/JCI93557
Ho, J., Hamizan, A. W., Alvarado, R., Rimmer, J., Sewell, W. A., and Harvey, R. J. (2018). Systemic predictors of eosinophilic chronic rhinosinusitis. Am. J. Rhinol. Allergy 32 (4), 252–257. doi:10.1177/1945892418779451
Huang, G. X., Hallen, N. R., Lee, M., Zheng, K., Wang, X., Mandanas, M. V., et al. (2023). Increased epithelial mTORC1 activity in chronic rhinosinusitis with nasal polyps. bioRxiv. Prepr. Serv. Biol. 2023, 562288. doi:10.1101/2023.10.13.562288
Ichimura, Y., Kumanomidou, T., Sou, Y. S., Mizushima, T., Ezaki, J., Ueno, T., et al. (2008). Structural basis for sorting mechanism of p62 in selective autophagy. J. Biol. Chem. 283 (33), 22847–22857. doi:10.1074/jbc.M802182200
Kim, H. Y., Nam, S. Y., Kim, H. M., and Jeong, H. J. (2021). Exposure of ovalbumin during pregnancy prevents the development of allergic rhinitis in offspring through the induction of mast cell autophagy. Cell. Immunol. 365, 104362. doi:10.1016/j.cellimm.2021.104362
Kim, J., Kundu, M., Viollet, B., and Guan, K. L. (2011). AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell. Biol. 13 (2), 132–141. doi:10.1038/ncb2152
Kirat, D., Alahwany, A. M., Arisha, A. H., Abdelkhalek, A., and Miyasho, T. (2023). Role of macroautophagy in mammalian male reproductive physiology. Cells 12 (9), 1322. doi:10.3390/cells12091322
Kiriyama, Y., and Nochi, H. (2015). The function of autophagy in neurodegenerative diseases. Int. J. Mol. Sci. 16 (11), 26797–26812. doi:10.3390/ijms161125990
Lee, J., and Kim, H. S. (2019). The role of autophagy in eosinophilic airway inflammation. Immune Netw. 19 (1), e5. doi:10.4110/in.2019.19.e5
Lee, J., Kim, S. J., Choi, G. E., Park, H. J., Choi, W. S., et al. (2022). Sweet taste receptor agonists attenuate macrophage IL-1β expression and eosinophilic inflammation linked to autophagy deficiency in myeloid cells. Clin. Transl. Med. 12 (8), e1021. doi:10.1002/ctm2.1021
Li, J., Qiu, C. Y., Tao, Y. J., and Cheng, L. (2022). Epigenetic modifications in chronic rhinosinusitis with and without nasal polyps. Front. Genet. 13, 1089647. doi:10.3389/fgene.2022.1089647
Li, J., Ullah, M. A., Jin, H., Liang, Y., Lin, L., Wang, J., et al. (2021). ORMDL3 functions as a negative regulator of antigen-mediated mast cell activation via an ATF6-UPR-autophagy-dependent pathway. Front. Immunol. 12, 604974. doi:10.3389/fimmu.2021.604974
Li, X., He, S., and Ma, B. (2020). Autophagy and autophagy-related proteins in cancer. Mol. Cancer 19 (1), 12. doi:10.1186/s12943-020-1138-4
Lin, Y. T., Lin, C. F., Liao, C. K., and Yeh, T. H. (2023). Comprehensive evaluation of type 2 endotype and clinical features in patients with chronic rhinosinusitis with nasal polyps in Taiwan: a cross-sectional study. Eur. Arch. Otorhinolaryngol. 280 (12), 5379–5389. doi:10.1007/s00405-023-08118-2
McAlinden, K. D., Deshpande, D. A., Ghavami, S., Xenaki, D., Sohal, S. S., Oliver, B. G., et al. (2019). Autophagy activation in asthma airways remodeling. Am. J. Respir. Cell. Mol. Biol. 60 (5), 541–553. doi:10.1165/rcmb.2018-0169OC
Munz, C. (2014). Regulation of innate immunity by the molecular machinery of macroautophagy. Cell. Microbiol. 16 (11), 1627–1636. doi:10.1111/cmi.12358
Munz, C. (2021). The macroautophagy machinery in MHC restricted antigen presentation. Front. Immunol. 12, 628429. doi:10.3389/fimmu.2021.628429
Painter, J. D., Galle-Treger, L., and Akbari, O. (2020). Role of autophagy in lung inflammation. Front. Immunol. 11, 1337. doi:10.3389/fimmu.2020.01337
Qi, J. J., Han, X. F., Cai, X. L., Li, X. Z., Zhang, L. Q., Feng, X., et al. (2016). Expression and clinical significance of autophagy-related gene Beclin1 and P62 in nasal polyps. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 51 (6), 428–432. doi:10.3760/cma.j.issn.1673-0860.2016.06.006
Ranieri, D., Nanni, M., Guttieri, L., Torrisi, M. R., and Belleudi, F. (2021). The aberrant expression in epithelial cells of the mesenchymal isoform of FGFR2 controls the negative crosstalk between EMT and autophagy. J. Cell. Mol. Med. 25 (8), 4166–4172. doi:10.1111/jcmm.16309
Sedaghat, A. R., Kuan, E. C., and Scadding, G. K. (2022). Epidemiology of chronic rhinosinusitis: prevalence and risk factors. J. Allergy Clin. Immunol. Pract. 10, 1395–1403. doi:10.1016/j.jaip.2022.01.016
Shariq, M., Quadir, N., Alam, A., Zarin, S., Sheikh, J. A., Sharma, N., et al. (2023). The exploitation of host autophagy and ubiquitin machinery by Mycobacterium tuberculosis in shaping immune responses and host defense during infection. Autophagy 19 (1), 3–23. doi:10.1080/15548627.2021.2021495
Shun, C. T., Lin, S. K., Hong, C. Y., and Liu, C. M. (2016). Sirtuin 6 modulates hypoxia-induced autophagy in nasal polyp fibroblasts via inhibition of glycolysis. Am. J. Rhinol. Allergy 30 (3), 179–185. doi:10.2500/ajra.2016.30.4282
Simsek, F., Eren, E., Bahceci, S., and Aladag, I. (2021). High PI3K/mTOR and low MAPK/JNK activity results in decreased apoptosis and autophagy in nasal polyposis. Braz J. Otorhinolaryngol. 87 (5), 572–577. doi:10.1016/j.bjorl.2019.12.005
Starry, A., Hardtstock, F., Wilke, T., Weihing, J., Ultsch, B., Wernitz, M., et al. (2022). Epidemiology and treatment of patients with Chronic rhinosinusitis with nasal polyps in Germany-A claims data study. Allergy 77, 2725–2736. doi:10.1111/all.15301
Theofani, E., and Xanthou, G. (2021). Autophagy: a friend or foe in allergic asthma? Int. J. Mol. Sci. 22 (12), 6314. doi:10.3390/ijms22126314
Wang, B. F., Cao, P. P., Wang, Z. C., Li, Z. Y., Ma, J., et al. (2017a). Interferon-gamma-induced insufficient autophagy contributes to p62-dependent apoptosis of epithelial cells in chronic rhinosinusitis with nasal polyps. Allergy 72 (9), 1384–1397. doi:10.1111/all.13153
Wang, C., Zhou, M. L., Liu, Y. C., and Cheng, K. J. (2022). The roles of autophagy, mitophagy, and the Akt/mTOR pathway in the pathogenesis of chronic rhinosinusitis with nasal polyps. J. Immunol. Res. 2022, 2273121. doi:10.1155/2022/2273121
Wang, L. F., Chien, C. Y., Yang, Y. H., Hour, T. C., Yang, S. F., Chen, H. R., et al. (2015). Autophagy is deficient and inversely correlated with COX-2 expression in nasal polyps: a novel insight into the inflammation mechanism. Rhinology 53 (3), 270–276. doi:10.4193/Rhin14.232
Wang, S., Jiang, Z., Li, L., Zhang, J., Zhang, C., and Shao, C. (2021). Ameliorative effects of eosinophil deficiency on immune response, endoplasmic reticulum stress, apoptosis, and autophagy in fungus-induced allergic lung inflammation. Respir. Res. 22 (1), 173. doi:10.1186/s12931-021-01770-4
Wang, X. H., Zhang, Z. H., Cai, X. L., Ye, P., Feng, X., Liu, T. T., et al. (2017b). Lipopolysaccharide induces autophagy by targeting the AMPK-mTOR pathway in human nasal epithelial cells. Biomed. Pharmacother. 96, 899–904. doi:10.1016/j.biopha.2017.12.011
Wang, Y., Wang, X., Jin, M., and Lu, J. (2023). Activation of the hedgehog signaling pathway is associated with the promotion of cell proliferation and epithelial-mesenchymal transition in chronic rhinosinusitis with nasal polyps. Eur. Arch. Otorhinolaryngol. 280 (3), 1241–1251. doi:10.1007/s00405-022-07664-5
Wang, Y., and Zhang, H. (2019). Regulation of autophagy by mTOR signaling pathway. Adv. Exp. Med. Biol. 1206, 67–83. doi:10.1007/978-981-15-0602-4_3
Xiong, G., Xie, X., Wang, Q., Zhang, Y., Ge, Y., Lin, W., et al. (2020). Immune cell infiltration and related core genes expression characteristics in eosinophilic and non-eosinophilic chronic rhinosinusitis with nasal polyps. Exp. Ther. Med. 20 (6), 180. doi:10.3892/etm.2020.9310
Xu, H. D., and Qin, Z. H. (2019). Beclin 1, bcl-2 and autophagy. Adv. Exp. Med. Biol. 1206, 109–126. doi:10.1007/978-981-15-0602-4_5
Yang, S. K., Kim, J. W., Won, T. B., Rhee, C. S., Han, Y. B., and Cho, S. W. (2023). Differences in clinical and immunological characteristics according to the various criteria for tissue eosinophilia in chronic rhinosinusitis with nasal polyps. Clin. Exp. Otorhinolaryngol. 16 (4), 359–368. doi:10.21053/ceo.2023.00542
Yang, Z., and Klionsky, D. J. (2010). Mammalian autophagy: core molecular machinery and signaling regulation. Curr. Opin. Cell. Biol. 22 (2), 124–131. doi:10.1016/j.ceb.2009.11.014
Ye, Y., Zhao, J., Ye, J., Jiang, X., Liu, H., Xie, Y., et al. (2019). The role of autophagy in the overexpression of MUC5AC in patients with chronic rhinosinusitis. Int. Immunopharmacol. 71, 169–180. doi:10.1016/j.intimp.2019.03.028
Yu, L., Chen, Y., and Tooze, S. A. (2018). Autophagy pathway: cellular and molecular mechanisms. Autophagy 14 (2), 207–215. doi:10.1080/15548627.2017.1378838
Yu, S., Li, Y., Lu, X., Han, Z., Li, C., Yuan, X., et al. (2024). The regulatory role of miRNA and lncRNA on autophagy in diabetic nephropathy. Cell. Signal 118, 111144. doi:10.1016/j.cellsig.2024.111144
Zeng, X., Liu, F., Liu, K., Xin, J., and Chen, J. (2021). HMGB1 could restrict 1,3-β-glucan induced mice lung inflammation by affecting Beclin1 and Bcl2 interaction and promoting the autophagy of epithelial cells. Ecotoxicol. Environ. Saf. 222, 112460. doi:10.1016/j.ecoenv.2021.112460
Zhang, Y., Qi, J., Yan, D., Deng, Y., Zhang, J., and Luo, Q. (2022). HNE induces the hyperexpression of MUC5AC in chronic rhinosinusitis with nasal polyps by activating the TRAF6/autophagy regulatory axis. Am. J. Rhinol. Allergy 36 (6), 816–826. doi:10.1177/19458924221116939
Subspecialty Group of Rhinology, Editorial Board of Chinese Journal of Otorhinolaryngology Head and Neck Surgery, and Subspecialty Group of Rhinology, Society of Otorhinolaryngology Head and Neck Surgery, Chinese Medical Association (2019). Chinese guidelines for diagnosis and treatment of chronic rhinosinusitis (2018). Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 54 (2), 81–100. doi:10.3760/cma.j.issn.1673-0860.2019.02.001
Zhou, S., Fan, K., Lai, J., Tan, S., Zhang, Z., Li, J., et al. (2023). Comprehensive analysis of mitophagy-related genes in diagnosis and heterogeneous endothelial cells in chronic rhinosinusitis: based on bulk and single-cell RNA sequencing data. Front. Genet. 14, 1228028. doi:10.3389/fgene.2023.1228028
Glossary

Keywords: chronic rhinosinusitis, nasal polyps, autophagy, CRSsNP, CRSwNP
Citation: Pei J, Ding Z, Jiao C, Tao Y, Yang H and Li J (2024) Autophagy in chronic rhinosinusitis with or without nasal polyps. Front. Cell Dev. Biol. 12:1417735. doi: 10.3389/fcell.2024.1417735
Received: 15 April 2024; Accepted: 13 May 2024;
Published: 12 June 2024.
Edited by:
Xianwei Wang, Xinxiang Medical University, ChinaReviewed by:
Yunping Fan, The 7th Affiliated Hospital of Sun Yat-sen University, ChinaDaniel Peña-Oyarzun, Pontifical Catholic University of Chile, Chile
Copyright © 2024 Pei, Ding, Jiao, Tao, Yang and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huifen Yang, am55eXloZkAxNjMuY29t; Jing Li, bmptdWViaEAxNjMuY29t
†These authors have contributed equally to this work