 Jianxiong Kang
Jianxiong Kang Peiyan Hua1
Peiyan Hua1 Xiaojing Wu
Xiaojing Wu Bin Wang
Bin Wang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol. , 09 April 2024
Sec. Cellular Biochemistry
Volume 12 - 2024 | https://doi.org/10.3389/fcell.2024.1271684
This article is part of the Research Topic Exosomal Biomarkers: Roles in Diagnostics and Therapeutics - Volume II View all 9 articles
Macrophages, the predominant immune cells in the lungs, play a pivotal role in maintaining the delicate balance of the pulmonary immune microenvironment. However, in chronic inflammatory lung diseases and lung cancer, macrophage phenotypes undergo distinct transitions, with M1-predominant macrophages promoting inflammatory damage and M2-predominant macrophages fostering cancer progression. Exosomes, as critical mediators of intercellular signaling and substance exchange, participate in pathological reshaping of macrophages during development of pulmonary inflammatory diseases and lung cancer. Specifically, in inflammatory lung diseases, exosomes promote the pro-inflammatory phenotype of macrophages, suppress the anti-inflammatory phenotype, and subsequently, exosomes released by reshaped macrophages further exacerbate inflammatory damage. In cancer, exosomes promote pro-tumor tumor-associated macrophages (TAMs); inhibit anti-tumor TAMs; and exosomes released by TAMs further enhance tumor proliferation, metastasis, and resistance to chemotherapy. Simultaneously, exosomes exhibit a dual role, holding the potential to transmit immune-modulating molecules and load therapeutic agents and offering prospects for restoring immune dysregulation in macrophages during chronic inflammatory lung diseases and lung cancer. In chronic inflammatory lung diseases, this is manifested by exosomes reshaping anti-inflammatory macrophages, inhibiting pro-inflammatory macrophages, and alleviating inflammatory damage post-reshaping. In lung cancer, exosomes reshape anti-tumor macrophages, inhibit pro-tumor macrophages, and reshaped macrophages secrete exosomes that suppress lung cancer development. Looking ahead, efficient and targeted exosome-based therapies may emerge as a promising direction for treatment of pulmonary diseases.
Pulmonary macrophages, the most abundant immune cells in the lungs, can be classified into distinct subtypes based on their functions, phenotypes, and secretory profiles. These subtypes include M1, M2, and other subgroups of macrophages (Aegerter et al., 2022). M1 macrophages, also known as “classically activated macrophages,” secrete proinflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-1β, IL-6, IL-12. In addition, M1 macrophages present foreign pathogens, cell debris, aging cells, and tumor cell antigens and engulf them (Yunna et al., 2020). However, excess of pro-inflammatory cytokines leads to an uncontrolled inflammatory response, resulting in damage to bronchial epithelial and alveolar cells, mucus obstruction of airways which restricts ventilation, and excessive activation of fibroblasts and collagen deposition (Dong et al., 2022). M2 macrophages, also known as “alternatively activated macrophages,” secrete anti-inflammatory cytokines including transforming growth factor beta (TGF-β), IL-10, CC motif chemokine ligand (CCL)18, and CCL22, exerting immunomodulatory functions (Cheng et al., 2021). They can further be divided into M2a, M2b, M2c, and M2d subtypes. M2a macrophages are associated with allergic reactions and secrete pro-fibrotic factors necessary for tissue repair, while M2b macrophages secrete anti-inflammatory cytokines such as IL-10 and have the ability to recruit regulatory T cells to combat inflammation (Yang et al., 2019; Gopalakrishnan et al., 2022). M2c macrophages suppress immune responses and promote tissue repair (Junior et al., 2021). M2d macrophages, also known as tumor-associated macrophages (TAMs), suppress inflammatory responses and promote angiogenesis and tumor growth (Zhang and Sioud, 2023).
Macrophages undergo substantial phenotypic and functional changes upon activation triggered by injury, stimulation, or alterations in the lung microenvironment. They play a significant role in the pathophysiology of diverse lung diseases, including inflammatory lung diseases such as chronic obstructive pulmonary disease (COPD), acute lung injury or acute respiratory distress syndrome (ALI/ARDS), pulmonary fibrosis, asthma, and lung cancer (TJonck and Bain, 2023). Prolonged injury and irritation induce macrophage population dysregulation, amplify inflammation or induce abnormal proliferation of tumor cells, and disrupt the pulmonary microenvironment, thereby contributing to development of various lung diseases (Melo et al., 2021).
Exosomes are membranous particles synthesized and secreted by cells. They can carry various genetic materials and signaling molecules, thereby regulating the functional state of recipient cells (Xu et al., 2021). Abnormalities in the quantity, cargo, or surface proteins of exosomes can reflect the status of donor cells and the extracellular microenvironment. Notably, exosomes isolated from blood, human bronchoalveolar lavage fluid (BALF), or sputum have shown promise as diagnostic markers for various lung diseases, thereby attracting growing interest as both mechanistic players and potential therapeutic targets in the context of pulmonary disorders (Zhao et al., 2022).
This review focuses on the dual regulatory role of exosomes in chronic pulmonary diseases. First, it provides an overview of exosomes, including their definition, biogenesis, structure, and composition. Subsequently, from the perspective of crosstalk between exosomes and macrophages, this review analyzes the mechanisms underlying the disruption of immune balance in chronic pulmonary inflammatory diseases and lung cancer. Finally, it analyzes the dual-edged nature of exosomes, highlighting their reparative effects on immune dysregulation associated with macrophages in chronic pulmonary inflammatory diseases and lung cancer.
Extracellular vesicles are membrane-bound structures released by cells into the extracellular space. Based on particle size and biogenesis processes, extracellular vesicles can be broadly categorized into three main groups: exosomes (30–150 nm in diameter), microvesicles (100–1,000 nm in diameter), and apoptotic bodies (100–5,000 nm in diameter) (Liu and Wang, 2023). Apoptotic bodies are vesicular structures formed by membrane wrinkling and invagination during cell apoptosis, directly budding off. Microvesicles are also vesicular structures formed through direct budding and shedding (Kalluri and LeBleu, 2020). Compared to microvesicles and apoptotic bodies, exosomes exhibit a more uniform particle size (Mathivanan et al., 2010). Exosomes have a diverse range of functions, including intercellular communication (immune suppression, antigen presentation, transfer of signaling components, inflammation, tumor growth, metastasis, angiogenesis, and intercellular exchange of functional genetic information), cell adhesion, and coagulation (van der Pol et al., 2012). Some functions of microvesicles overlap with those of exosomes, such as cell adhesion and coagulation (Wolf, 1967; Merten et al., 1999).
Exosome biogenesis involves a complex series of membrane fusion and sorting processes to encapsulate and sort contents. Initially, there is an invagination of the cell membrane, encapsulating extracellular components, to form early sorting endosomes (ESEs). These ESEs can then fuse with each other or engage in material exchange with internally synthesized substances, resulting in the formation of late sorting endosomes (LSEs). Further sorting and fusion lead to the formation of multivesicular bodies (MVBs) within the cell. MVB biogenesis involves various mechanisms, including the endosomal sorting complex required for transport (ESCRT) machinery for the transport-associated intraluminal vesicle (ILV) sorting, triggered by ESCRT-I/II, as well as non-ESCRT mechanisms (Colombo et al., 2014). After maturation and sorting of intraluminal vesicles (ILVs, also known as exosomes) within MVBs, two potential fates emerge: MVBs can either fuse with the cell membrane, budding internally and releasing exosomes into the extracellular space, or they may undergo fusion with lysosomes for cargo degradation (Broad et al., 2023). Upon transport and docking at the plasma membrane, secretory MVBs bind to the cell’s inner membrane receptors (such as soluble N-ethylmaleimide-sensitive factor attachment protein receptor, SNARE), initiating release (Bebelman et al., 2018). As depicted in Figure 1, exosome formation involves multiple rounds of sorting and assembly, resulting in significant heterogeneity among exosomes. This heterogeneity is not only evident in their varying particle sizes but, more importantly, in their unique structures and compositions.
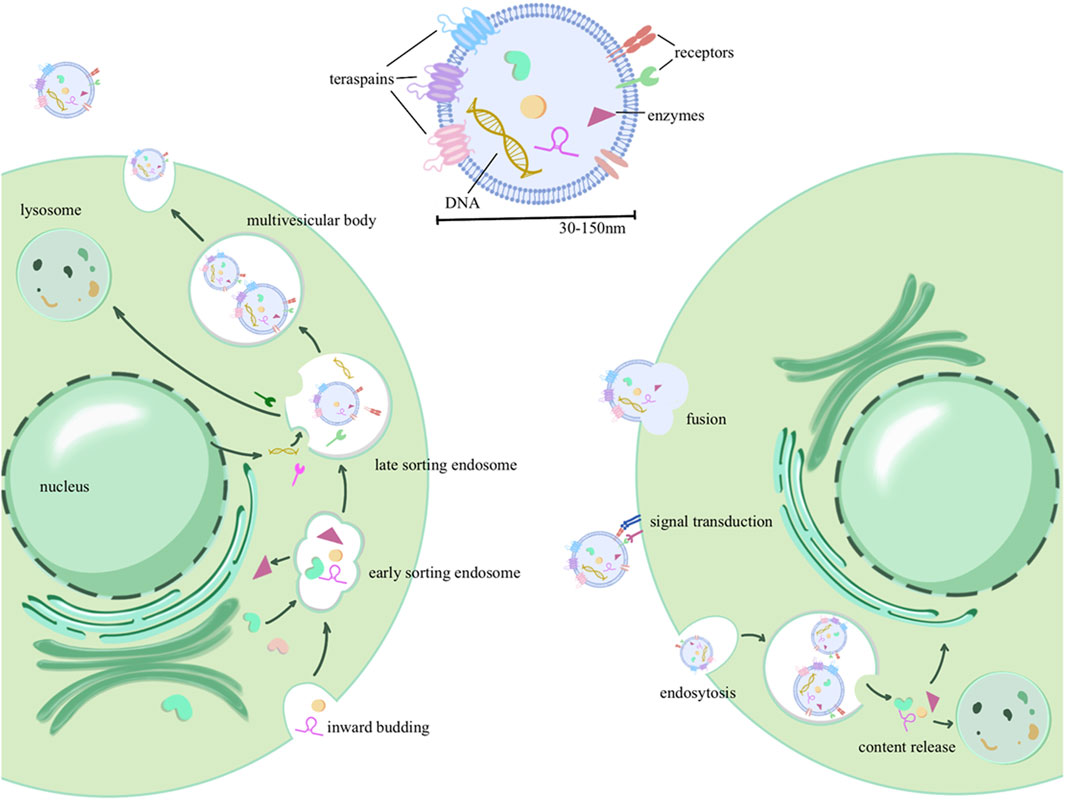
Figure 1. Exosome production process, contents, and delivery: the process initiates with invagination of the plasma membrane, forming early sorting endosomes (ESEs) that encapsulate extracellular components. These ESEs can fuse with intracellularly synthesized substances or exchange biomaterials, leading to the formation of late sorting endosomes (LSEs). Further maturation gives rise to intracellular multivesicular bodies (MVBs), from which intraluminal vesicles (ILVs) are selectively sorted and eventually released into the extracellular space as extracellular vesicles. In addition, three forms of extracellular vesicle uptake by recipient cells were described, including membrane fusion, receptor binding (signal transduction), and endocytosis.
Generally, exosomes are primarily composed of a phospholipid membrane forming the outer shell, along with various cargo molecules such as RNA, DNA, and proteins (Mondal et al., 2023). The phospholipid bilayer of exosomes is relatively stable, allowing for long-distance transportation. In addition to maintaining the stability of the membrane skeleton, membrane proteins also participate in information exchange, such as selective binding with receptor cells. These membrane-encapsulated proteins can be transported to recipient cells, serving diverse functions such as enzymatic modification of downstream pathways or assisting in viral transmission by transporting viral antigens (Gurunathan et al., 2021). Some specific proteins are used for exosome identification, including tetraspanins CD63, CD9, and CD81; heat shock proteins HSP70 and HSP90; as well as apoptosis-linked gene 2-interacting protein X(ALIX) and tumor susceptibility gene 101(TSG101) (Pegtel and Gould, 2019). In addition to proteins, exosomes also carry various types of RNAs and DNA. Non-coding RNAs, in particular, can epigenetically modify gene expression frequencies in recipient cells, leading to altered functionality. The DNA and RNA in exosomes may carry genetic information from donor cells and viruses (Zaiets et al., 2023) and can serve as diagnostic markers for inflammation, cancer, and viral infections (Zhu et al., 2020).
As mentioned earlier, macrophages are primarily divided into two subtypes, M1 and M2. M1 macrophages are typically associated with inflammation and immune response, while M2 macrophages are linked to anti-inflammatory and tissue repair processes (Aegerter et al., 2022). In inflammatory lung diseases such as asthma and COPD, as well as in lung cancer, the roles of M1 and M2 macrophages exhibit complex interrelationships.
In inflammatory lung diseases such as asthma and COPD, M1 macrophages are typically involved in the inflammatory process, producing inflammatory mediators and engulfing pathogens. The inflammatory response of M1 macrophages may be necessary in the early stages. However, excessive or prolonged inflammation may lead to lung tissue damage (Chen et al., 2020). M2 macrophages are generally associated with anti-inflammation and promotion of tissue repair processes. They may play a protective role in the later stages of inflammation, but excessive activated M2 macrophages can produce pro-fibrotic mediators, leading to sustained activation of fibroblasts, promoting myofibroblast proliferation, and facilitating collagen deposition (Zhang et al., 2018). In summary, maintaining a balance between M1 and M2 macrophages in inflammatory lung diseases can help resist the invasion of pulmonary pathogens, protect tissue cells from excessive inflammatory damage, and prevent fibrosis.
In lung cancer, tumor-associated macrophages (TAMs) are closely associated with tumor development, invasion, and metastasis, influencing the tumor microenvironment, immune suppression, and angiogenesis (Hu et al., 2022). TAMs are generally classified into two main subtypes: M1 macrophages and M2 macrophages. In the early stages of lung cancer, M1 macrophages may exhibit anti-tumor activity. They identify and destroy cancer cells, produce inflammatory mediators, activate immune cells, and participate in anti-tumor immune responses (Ma et al., 2010). M2 macrophages: as the tumor progresses, TAMs gradually transform into M2 macrophages, displaying immunosuppressive characteristics and promoting tumor growth. They release immunosuppressive molecules; inhibit anti-tumor immune responses; and simultaneously facilitate angiogenesis, invasion, and metastasis of tumor cells (Basak et al., 2023).
In summary, the roles of macrophages in inflammatory lung diseases and lung cancer may be complex and dynamic. The contradictory nature of these roles primarily depends on the specific functions of macrophages at different disease stages and their interactions with other immune cells and inflammatory mediators (Boutilier and Elsawa, 2021; Lee et al., 2021; Basak et al., 2023).
As crucial immune cells regulating the pulmonary microenvironment, macrophages play a pivotal role in promptly recognizing and engulfing invading pathogens, as well as senescent and aberrant cells entering the airways. They secrete cytokines to activate various immune cells, present antigens to initiate subsequent immune responses, and maintain a normal immune microenvironment in the lungs (Shotland et al., 2021). In this section, we elucidate the role of exosomes in macrophage-related immune dysregulation in inflammatory lung diseases and lung cancer, as shown in Table 1. In inflammatory lung diseases such as asthma, acute respiratory distress syndrome (ARDS), chronic obstructive pulmonary disease (COPD), and pulmonary fibrosis, exosomes reshape macrophages toward a pro-inflammatory phenotype, suppressing the anti-inflammatory phenotype. The reshaped macrophages release exosomes associated with inflammatory damage (Jiang et al., 2023), as shown in Figure 2. In lung cancer, exosomes reshape macrophages to promote a pro-tumorigenic phenotype while suppressing the anti-tumor phenotype. Tumor-associated macrophages release exosomes that further enhance tumor proliferation, metastasis, and resistance to chemotherapy (Liang et al., 2020), as shown in Figure 3.
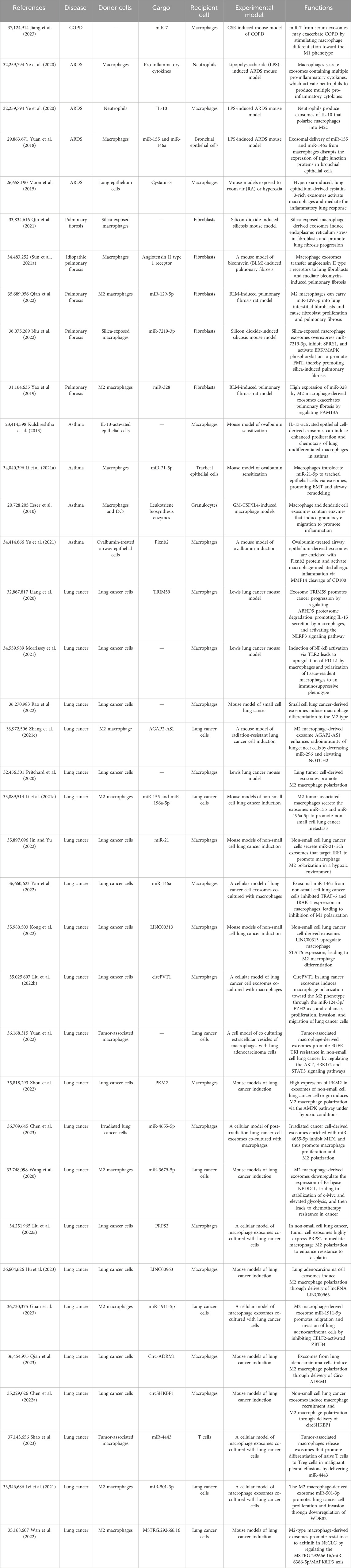
Table 1. Exosomes are involved in macrophage-mediated formation of the pulmonary pathological microenvironment.
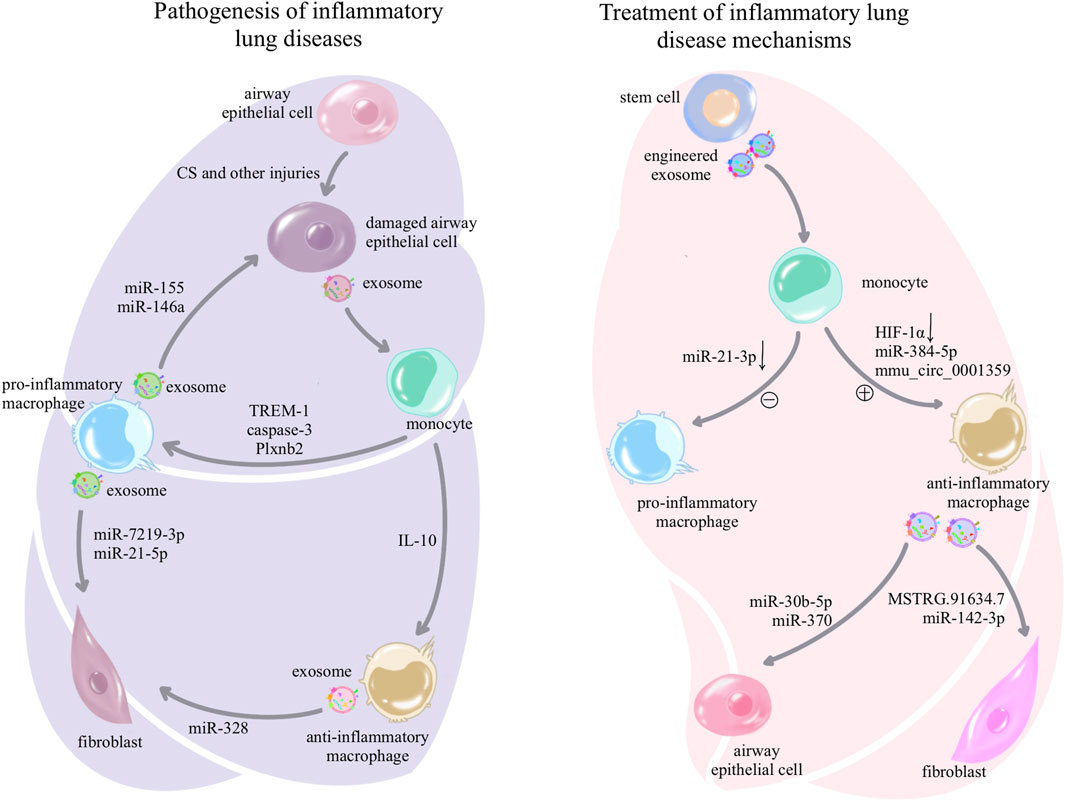
Figure 2. Pathogenesis and treatment of exosome-associated inflammatory lung disease: in inflammatory lung diseases, exosomes can be secreted by damaged airway epithelial cells, prompting macrophages to differentiate toward the pro-inflammatory phenotype and suppressing anti-inflammatory differentiation; while pro-inflammatory macrophages in turn use exosomes for further airway epithelial damage, fibroblast activation, and activation of other immune cells, forming a vicious circle to amplify inflammation and injury. However, the treatment of inflammatory lung diseases is delivered to macrophages through a variety of stem cell exosomes and synthetic drug-encapsulated exosomes, which induce anti-inflammatory differentiation and inhibit pro-inflammatory differentiation of macrophages, or the use of anti-inflammatory macrophage-secreted exosomes that act on other cells to mitigate the damage and destruction of lung tissue.
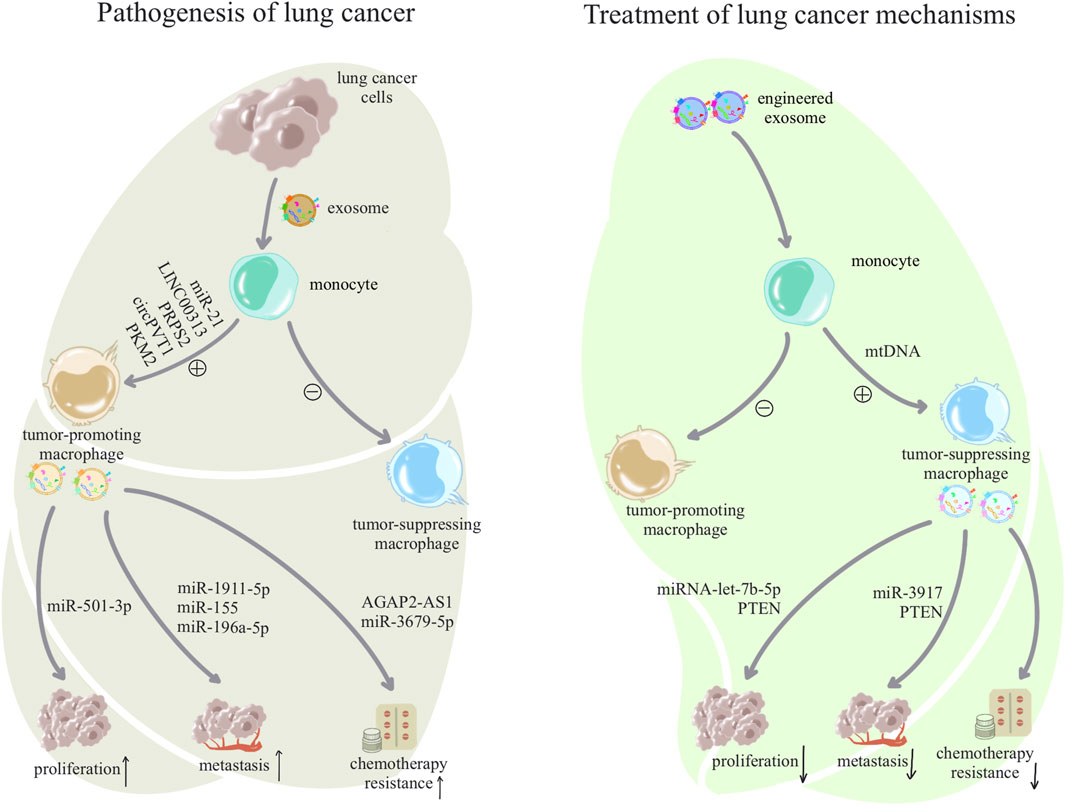
Figure 3. Pathogenesis and treatment of exosome-associated lung cancer: lung cancer cells use exosomes to alter macrophage subpopulations to form a tumor microenvironment, and dysregulated macrophages facilitate a variety of malignant behaviors such as growth, metastasis, and drug resistance of lung cancer cells through exosomes. In contrast, treatment of lung cancer can begin by awakening anti-tumor macrophages in macrophages and inhibiting tumor-assisting macrophages, and anti-tumor macrophages will also inhibit the proliferation and development of tumor cells through exosomes.
In COPD, exosomes activate macrophages and induce differentiation toward the pro-inflammatory M1 type, leading to persistent inflammatory damage and formation of a pathological microenvironment in the lungs. Cigarette smoke exposure is an important risk factor for COPD. Wang et al. (2021) demonstrated that mouse airway epithelial cells treated with cigarette smoke extract (CSE) released exosomes expressing high levels of triggering receptor expressed on myeloid cells-1 (TREM-1), which influenced macrophages and promoted M1 polarization. Subsequently, the excessive inflammatory response mediated by M1 macrophages leads to bronchial wall thickening and alveolar structural damage. Additionally, in COPD mice, highly expressed miR-7 in serum exosomes acted on macrophages, regulated macrophage activation through proto-oncogene Pim-1(PIM1), and promoted differentiation into the M1 phenotype, exacerbating the inflammatory response (Jiang et al., 2023).
ALI/ARDS is characterized by direct or indirect lung injury and an excessive and uncontrolled systemic inflammatory response (Chen et al., 2020). The pathogenesis of ALI/ARDS is complex, involving both infectious and non-infectious factors. High oxygen injury is a significant non-infectious etiology, where pulmonary epithelial cells, upon exposure to high oxygen injury, secrete exosomes rich in caspase-3. These exosomes, through the ROCK1 pathway, activate macrophages, leading to an increased secretion of inflammatory proteins, such as macrophage inflammatory protein 2 (MIP-2), thereby exacerbating the inflammatory lung response and worsening lung injury (Moon et al., 2015).
In asthma, Persistent airway inflammation and inflammation-induced airway remodeling are critical factors. Damaged airway epithelial cells can activate monocytes–macrophages and various inflammatory cells through exosomes. For instance, IL-13-induced epithelial cells exacerbate asthma inflammation by inducing monocyte proliferation and chemotaxis via exosomal signaling. Conversely, inhibition of exosome production by GW4869 (exosome inhibitor) reduces monocyte proliferation and chemotaxis (Kulshreshtha et al., 2013). Additionally, exosomes derived from ovalbumin-primed airway epithelial cells are enriched in Plexin B2 (Plxnb2) protein and cleave CD100 via matrix metalloproteinase 14 (MMP14), leading to increased levels of soluble CD100, which activate macrophage-mediated allergic inflammation and enhance the recruitment of lung neutrophils, eosinophils, and dendritic cells (Yu et al., 2021).
In inflammatory lung diseases, various etiological factors induce epithelial cell damage, promoting polarization of M1 macrophages, exacerbating inflammatory injury, and concurrently inhibiting the polarization of M2 macrophages. This imbalance between M1 and M2 cells further aggravates inflammation (Song et al., 2019). BEAS-2B cells (human bronchial epithelial cells) treated with CSE can reduce the polarization of M2 macrophages through modulated extracellular vesicles (He et al., 2019). Numerous studies have indicated that therapeutic exosomes can alleviate lung inflammation by promoting the polarization of M2 macrophages (Deng et al., 2020; Harrell et al., 2020). However, persistent inflammatory damage can also induce the polarization of M2 macrophages, leading to airway remodeling and fibrosis. For instance, in acute respiratory distress syndrome (ARDS), activated neutrophils release exosomes containing IL-10, polarizing macrophages into the M2c subtype, thereby inducing tissue remodeling and fibrosis after ALI (Ye et al., 2020).
In ARDS, hyperactivated macrophages secrete exosomes that contribute to the activation of other inflammatory cells and damage airway epithelial cells, exacerbating inflammation and injury. For instance, macrophage-derived exosomes contain pro-inflammatory cytokines that activate neutrophils and enhance their inflammatory response, leading to uncontrolled inflammation in ARDS (45). Additionally, exosomes released by macrophages disrupt the expression of tight junction proteins in bronchial epithelial cells, compromising the structural barrier (Yuan et al., 2018).
In pulmonary fibrosis, dysregulated macrophage subpopulations secrete exosomes that contribute to fibroblast activation and collagen deposition. For example, in silicosis, macrophage-derived exosomes induced endoplasmic reticulum stress in fibroblasts, upregulated type I collagen and alpha-smooth muscle actin (α-SMA) expression, and exacerbated fibrosis progression (Qin et al., 2021). Moreover, silica-exposed macrophage exosomes express high levels of miR-7219-3p, which promotes fibroblast-to-myofibroblast trans-differentiation (FMT) and enhances silicosis-induced pulmonary fibrosis through Spouty1 (SPRY1) inhibition and extracellular signal-regulated protein kinase (ERK)/mitogen-activated protein kinase (MAPK) pathway activation (Niu et al., 2022). In a mouse model of bleomycin-induced fibrosis, macrophages exhibited increased levels of angiotensin II (Ang II) and angiotensin II type 1 receptor (AT1R). Furthermore, exosomes facilitated the transportation of Ang II from macrophages to fibroblasts, contributing to the progression of fibrosis (Sun et al., 2021). M2 macrophages can release pro-fibrotic factors and activate fibroblasts through the exosomal pathway. For example, M2 macrophage-derived exosomes containing high levels of miR-328 exacerbate pulmonary fibrosis by regulating Family with sequence similarity 13, member A (FAM13A) (Yao et al., 2019).
In asthma, macrophages are activated by exosomes and, in turn, activate and recruit more inflammatory cells through exosome-mediated signaling. For example, macrophages and dendritic cells secrete exosomes containing enzymes involved in leukotriene biosynthesis and hence promote granulocyte migration (Esser et al., 2010). Alveolar macrophages transport miR-21-5p via exosomes to tracheal epithelial cells, promoting epithelial–mesenchymal transition (EMT) and airway remodeling through the TGF-β1/Smad signaling pathway targeting Smad7(52).
As mentioned Section 3, TAMs gradually shift toward an M2-dominant phenotype with the progression of tumors (Basak et al., 2023). M2-polarized macrophages, as an alternative activated form, have been implicated in various malignant processes, including promoting tumor cell proliferation and anti-apoptosis (Ye et al., 2018), metastasis (Zhang et al., 2021), increased vascular permeability and edema formation (Zhang et al., 2021), angiogenesis, and immune suppression (Mohapatra et al., 2021). Exosomes, as crucial mediators of communication between tumor cells and the immune microenvironment in the lungs, play a significant role in assisting tumor cells in reshaping M2-type TAMs and creating an immune microenvironment conducive to tumor growth (Qiu et al., 2022).
First, tumor cell-derived exosomes can deliver various non-coding RNAs, altering the epigenetics of macrophages and activating the polarization of M2-type macrophages. Examples include exosomes secreted by non-small cell lung cancer, which are rich in miR-21 (Jin and Yu, 2022), LINC00313 (Kong et al., 2022), and PRPS2 (Liu et al., 2022), promoting M2 polarization in macrophages. Liu et al. (2022) also found that exosomes from lung cancer contain circPVT1, which induces M2 polarization by inhibiting the expression of miR-124-3p in macrophages, leading to enhanced proliferation, invasion, and migration capabilities of lung cancer cells. Additionally, glycolysis is a crucial mechanism influencing macrophage polarization. For instance, pyruvate kinase M2 (PKM2) is a crucial molecule in macrophage metabolic adaptation, and tumor cells can activate PKM2 to regulate macrophage glycolysis, promoting the transition toward the M2 phenotype (Wu et al., 2022). Under hypoxic conditions, non-small cell lung cancer cells can directly secrete exosomes rich in PKM2 to induce M2 macrophage polarization through the AMPK pathway in macrophages (Zhou et al., 2022). Indirectly, through the delivery of exosomes, miR-1294 can upregulate PKM2 expression, promoting M2 macrophage polarization in an HIF-1α-dependent manner by regulating glycolysis (Chen et al., 2022). Furthermore, E-box-binding homeobox 1 (Zeb1) can induce the accumulation of M2-like tumor-associated macrophages (TAMs) through its involvement in glycolysis regulation (Jiang et al., 2022). Zeb1 can also induce the transcription of macrophage colony-stimulating factor (M-CSF) in cancer cells, leading to the secretion of M-CSF, driving M2-TAM polarization (Long et al., 2021). In lung adenocarcinoma, cancer cells can transfer LINC00963 through exosomes, stabilizing Zeb1 and stimulating M2 macrophage polarization (Hu et al., 2023). Additionally, matrix metalloproteinase 14 (MMP14), known as a target in various cancers (Liang et al., 2022), can not only promote tumor cell proliferation and migration but also facilitate the polarization of M2 macrophages. Exosomes from lung adenocarcinoma cells transmit Circ-ADRM1, recruiting USP12 to prevent ubiquitination of MMP14 protein and enhance MMP14 protein stability, thereby promoting M2-type macrophage polarization (Qian et al., 2023).
Additionally, TAMs exhibit a non-classical M2/M1 phenotype (Qian and Pollard, 2010). It is inappropriate to solely consider M2-polarized macrophages in TAMs as favorable for tumor growth and M1-polarized macrophages as inhibitory to tumor growth, as TAMs undergo reprogramming by tumor-derived exosomes. For instance, tumor-derived exosomes transfer tripartite motif-containing 59 (TRIM59) to macrophages, inducing the ubiquitination of abhydrolase domain-containing 5 (ABHD5), activating pro-tumor functions in macrophages. This activation occurs through the secretion of IL-1β, which activates the NLRP3 inflammasome signaling pathway, promoting the inflammatory microenvironment and cancer progression (Liang et al., 2020).
Tumor-derived exosomes have been shown to exert immunosuppressive effects on macrophages, creating a microenvironment that facilitates tumor growth by suppressing “thermal immunity.” One mechanism involves the upregulation of programmed death ligand 1 (PD-L1), which promotes immunosuppression of macrophages. Tumor-derived exosomes activate NF-κB through Toll-like receptor-2 (TLR2), leading to the upregulation of PD-L1 expression. Moreover, increased glucose uptake and lactate conversion induced by these exosomes further enhance NF-κB activity, resulting in elevated PD-L1 levels. Consequently, tissue-resident macrophages are polarized toward an immunosuppressive phenotype (Morrissey et al., 2021). Furthermore, exosomes released by non-small cell lung cancer cells inhibit the expression of TNF receptor-associated factor 6 (TRAF6) and interleukin-1 receptor-associated kinase (IRAK1) in M0 macrophages, impairing M1 polarization and diminishing their ability to eliminate tumor cells (Yan et al., 2022). These findings underscore the role of tumor-derived exosomes in modulating macrophage function and shaping an immunosuppressive microenvironment that promotes tumor survival.
As mentioned above, tumor-derived exosomes can reshape TAMs, particularly promoting the formation of immunosuppressive TAMs that dominate over anti-tumor TAMs (Qiu et al., 2022). TAMs, reshaped by tumor cells and the tumor microenvironment, secrete exosomes that promote tumor proliferation, invasion, metastasis, and resistance to chemotherapy or radiotherapy (Xu et al., 2022). This section primarily analyzes the impact of reshaped TAM-derived exosomes on tumor development.
First, TAMs promote the proliferation of lung cancer cells through exosomes. Specifically, exosomes derived from M2 macrophages have the ability to transport miR-501-3p to tumor cells. It has been discovered that WD repeat-containing 82 (WDR82) serves as the target gene for miR-501-3p, and its tumor-suppressive function has been demonstrated in rectal cancer (Liu et al., 2018; Li et al., 2021) Similarly, M2 macrophage-derived exosomes carrying miR-501-3p enhance lung cancer cell proliferation and invasion by downregulating WDR82 (76).
Second, TAMs also contribute to the invasion and metastasis of lung cancer cells through exosomes. For example, exosomes derived from M2 macrophages containing miR-1911-5p facilitate the migration and invasion of lung adenocarcinoma cells by silencing zinc finger and BTB domain-containing 4 (ZBTB4) (Guan et al., 2023), and ZBTB4 has been shown to regulate glycolipid metabolism and inhibit proliferation and invasion in malignancies such as pancreatic adenocarcinoma (Yang et al., 2023) and glioma (Dong et al., 2021). In lung cancer, ZBTB4 is also a target of CS-induced EMT (Cheng et al., 2021). Therefore, we speculated that M2 macrophage-derived exosomes reduce the expression of ZBTB4, thereby promoting the formation of a tumor-friendly microenvironment and facilitating tumor metastasis in advance. Additionally, another potential therapeutic target for inhibiting cancer cell invasion and EMT is RAS association domain family 4 (RASSF4) (Zhang et al., 2017). Exosomes carrying miR-155 and miR-196a-5p, secreted by M2 tumor-associated macrophages, negatively regulate the expression of RASSF4, thus promoting metastasis and EMT in non-small cell lung cancer (Li et al., 2021c).
Third, macrophages play a significant role in promoting chemotherapy resistance or radiation resistance in lung cancer cells through release of exosomes. For instance, the resistance of non-small cell lung cancer cells to epidermal growth factor receptor (EGFR)-targeted drugs could be attributed to the reactivation of AKT, ERK1/2, and signal transducer and activator of transcription 3 (STAT3) signaling pathways facilitated by exosomes released from macrophages (Yuan et al., 2022). Moreover, M2 macrophages can activate the MAPK signaling pathway via exosomes, leading to resistance against the EGFR-targeted drug osimertinib (Wan et al., 2022). Another factor contributing to drug resistance is heightened glycolysis. Increased expression of miR-3679-5p carried by M2 macrophage-derived exosomes downregulates the expression of E3 ligase NEDD4L (neural precursor cell-expressed developmentally downregulated gene 4-like), resulting in the stabilization of MYC proto-oncogene (c-Myc) and enhanced glycolysis. Enhanced glycolysis, in turn, leads to chemotherapy resistance in cancer cells (Wang et al., 2020). Apart from conferring chemotherapy resistance, exosomes can also induce radiation resistance. For instance, the M2 macrophage-derived exosome AGAP2 antisense RNA 1 (AGAP2-AS1) promotes the malignant phenotype of radiation-resistant cancer cells by reducing the levels of miR-296 and increasing the expression of notch homolog protein 2 (NOTCH2) (Rao et al., 2022).
Lastly, TAMs can modulate immune cells through exosomes, reshaping the immunosuppressive microenvironment and promoting tumor progression (Xu et al., 2022). For instance, TAMs with predominant M2 polarization secrete exosomes rich in miR-4443, facilitating the differentiation of naïve T cells into regulatory T (Treg) cells in malignant pleural effusion (Shao et al., 2023).
As mentioned above, regulation of macrophages by exosomes plays a pathogenic role in promoting inflammation and tumor growth in inflammatory lung diseases and lung cancer. However, exosomes, as a double-edged sword, derived from stem cells or artificially manipulated exosomes, can also exert anti-inflammatory and anti-tumor effects (Gunassekaran et al., 2021; Wang et al., 2023). As shown in Table 2. This section will discuss the role of exosomes in reshaping macrophages toward an anti-inflammatory phenotype, inhibiting pro-inflammatory phenotypes, and reshaped macrophages release exosomes to alleviate inflammatory damage in inflammatory lung disease, as shown in Figure 2. Additionally, it will elaborate on how exosomes reshape anti-tumor macrophages, inhibit tumor-promoting macrophages, and how reshaped macrophages secrete exosomes to suppress tumor proliferation in lung cancer, as shown in Figure 3.
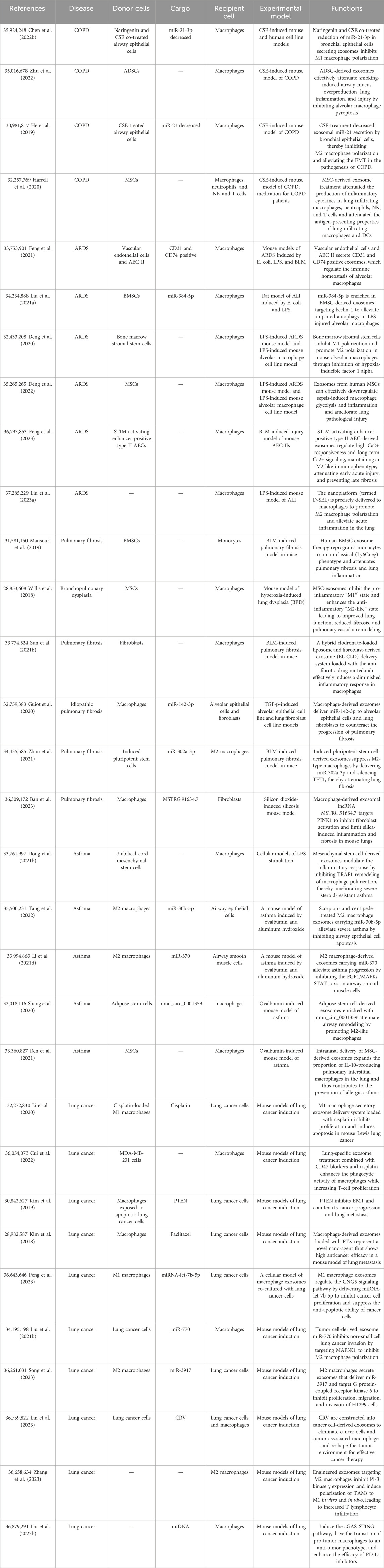
Table 2. Exosomes target in macrophage-mediated repair of the pulmonary pathological microenvironment.
In COPD, exosomes derived from stem cells can repair macrophage immune dysregulation and alleviate inflammatory damage. For instance, exosomes derived from mesenchymal stem cells (MSCs) have been found to interact with alveolar macrophages, inhibiting apoptosis and effectively mitigating persistent airway inflammation induced by cigarette smoke exposure (Zhu et al., 2022). Moreover, exosomes derived from MSCs facilitate the expansion of M2 macrophages and enhance the secretion of IL-10. This, in turn, induces the expansion of regulatory dendritic cells (DCs) and regulatory T cells, resulting in alterations to the immune microenvironment within the airways and consequently reducing chronic airway inflammation (Harrell et al., 2020).
In ARDS, M2 macrophages play a critical role in tissue repair and reducing inflammatory damage. Therefore, various engineered exosomes or stem cell-derived exosomes can alleviate inflammation in the acute phase of ARDS by promoting M2 macrophage polarization. For instance, an inhalable biomimetic sustained drug release nanoplatform, known as D-SEL, consisting of a combination of serum exosomes and liposomes encapsulating methylprednisolone succinate (MPS), can be precisely delivered to macrophages to promote M2 macrophage polarization and alleviate acute lung inflammation (Liu et al., 2023). Exosomes released by bone marrow mesenchymal stem cells (BMSCs) downregulate glycolysis by inhibiting HIF-1α, thereby promoting M2 polarization and attenuating sepsis-induced lung injury. These exosomes possess potent immunomodulatory and immunosuppressive properties (Deng et al., 2020). Exosomes derived from STIM-activating enhancer-positive type II alveolar epithelial cells (AEC II) regulate high Ca2+ responsiveness and long-term Ca2+ signaling, thereby maintaining an M2-like immune phenotype and metabolic selection. This modulation helps attenuate early acute injury and helps prevent late-stage fibrosis (Feng et al., 2023). Additionally, macrophages can be classified based on Ly6C into Ly6Clow, Ly6Chi, and Ly6C- macrophages. Exosomes derived from endothelial cells (EnCs) and AEC II suppress the expression of regulator of G protein signaling-1 (RGS1) in macrophages. RGS1 regulates macrophage Ca2+-dependent immune responses and modulates the recruitment of macrophages with different immune phenotypes during lung infection. Adjusting the pro-inflammatory or anti-fibrotic phenotype of Ly6C- macrophages promotes an increase in anti-inflammatory cytokines and tissue repair or fibrotic factors (Feng et al., 2021). Moreover, reducing macrophage autophagy also alleviates the progression of ARDS. BMSC-derived exosomes enriched in miR-384-5p target beclin-1 to alleviate autophagic stress in lipopolysaccharide (LPS)-injured alveolar macrophages and thereby alleviate inflammation (Liu et al., 2021).
In asthma, various stem cell-derived exosomes can promote M2 macrophage polarization to attenuate the inflammatory response and airway remodeling. For instance, exosomes derived from MSCs inhibit tumor necrosis factor receptor-associated factor 1 (TRAF1)-mediated macrophage polarization and promote M2 polarization, thereby modulating the inflammatory response and ameliorating severe steroid-resistant asthma (Dong et al., 2021). Exosomes derived from ADSCs modified with mmu_circ_0001359 absorb miR-183-5p to enhance forkhead box transcription factor 1 (FOXO1) signaling-mediated activation of M2 macrophages for treatment of asthma (Shang et al., 2020). Furthermore, intranasal delivery of mesenchymal stem cell-derived exosomes increases the proportion of IL-10-producing interstitial macrophages in the lungs, exerting anti-inflammatory effects and effectively treating allergic asthma (Ren et al., 2021).
In COPD, the inhibition of M1 macrophage polarization has been shown to effectively reduce inflammatory infiltration. For instance, the co-treatment of bronchial epithelial cells with naringenin and CSE resulted in the secretion of exosomes with reduced levels of miR-21-3p, which targets phosphatase and tensin homolog (PTEN)/AKT, leading to inhibition of M1 macrophage polarization. This treatment also resulted in decreased secretion of TNF-α, IL-6, IL-1β, inducible nitric oxide synthase (iNOS), and IL-12 (98). Furthermore, the administration of exosomes derived from MSCs attenuated the production of inflammatory cytokines in lung-infiltrating macrophages, neutrophils, natural killer cells, and natural killer T cells, reducing the antigen-presenting capacity of lung-infiltrating macrophages and dendritic cells (Harrell et al., 2020). M2 macrophages can secrete anti-inflammatory cytokines but exacerbate EMT. Treatment with CSE resulted in a decrease in secretion of exosomal miR-21 by bronchial epithelial cells, thereby inhibiting M2 macrophage polarization and alleviating the pathogenesis of EMT in COPD (99).
In ARDS, exosomes derived from various stem cells exhibit the ability to inhibit pro-inflammatory macrophages or suppress the expression of pro-inflammatory factors, thereby mitigating early acute inflammation and reducing mortality. For instance, exosomes derived from human MSCs effectively downregulate macrophage glycolysis and the expression of pro-inflammatory factors induced by sepsis, leading to the amelioration of pulmonary pathological injury (Deng et al., 2022). As mentioned earlier, BMSCs promote M2 polarization by releasing exosomes that downregulate glycolysis and also exhibit inhibitory effects on M1 polarization. This modulation suppresses pro-inflammatory cytokines and prevents the escalation of the inflammatory response (Deng et al., 2020). Moreover, the previously mentioned D-SEL also promotes M2 polarization. Additionally, localized and sustained release of DNase I degrades dysregulated neutrophil extracellular traps (NETs), inhibiting neutrophil activation and the formation of a mucus-clogged microenvironment. This process further suppresses the recruitment of pro-inflammatory macrophages and reinforces M2 polarization (Liu et al., 2023).
In pulmonary fibrosis, exosomes derived from MSCs have the ability to suppress pro-inflammatory macrophages, resulting in the amelioration of chronic inflammation and pulmonary fibrosis. For instance, exosomes derived from human BMSCs suppress the pro-inflammatory monocyte phenotype and shift the distribution of pulmonary classical and non-classical monocytes toward that observed in control mice, including alveolar macrophages (Mansouri et al., 2019). MSC exosomes regulate and inhibit pro-inflammatory M1 alveolar macrophages both in vitro and in vivo, leading to improved lung function, reduced fibrosis, pulmonary vascular remodeling, and amelioration of pulmonary hypertension (Willis et al., 2018). A delivery system combining clodronate-loaded liposomes with fibroblast-derived exosomes (EL-CLD), loaded with the anti-fibrotic drug nintedanib, effectively induces a diminished inflammatory response in macrophages, providing a potential therapeutic approach for pulmonary fibrosis (Sun et al., 2021). Additionally, exosomes derived from induced pluripotent stem cells (iPSCs) suppress M2-type macrophages by delivering miR-302a-3p and silencing ten-eleven translocation 1 (TET1), thereby attenuating pulmonary fibrosis (Zhou et al., 2021).
In asthma, inhibiting the inflammatory response of macrophages can significantly alleviate disease progression and impaired ventilation. For example, exosomes derived from mesenchymal stem cells inhibit TRAF1 to remodel macrophage polarization and suppress M1 polarization, thereby modulating the inflammatory response and ameliorating severe steroid-resistant asthma (Dong et al., 2021).
In pulmonary fibrosis, exosomes derived from macrophages play a crucial role in targeting and inhibiting fibroblast activation and collagen deposition, thereby attenuating the exacerbation of fibrosis and impaired ventilation. For instance, macrophage-derived exosomes deliver miR-142-3p to alveolar epithelial cells and lung fibroblasts, effectively countering the progression of pulmonary fibrosis (Guiot et al., 2020). Moreover, macrophage-derived exosomes containing the lncRNA MSTRG.91634.7 target PTEN-induced putative kinase 1 (PINK1) to inhibit fibroblast activation, thereby limiting silica-induced lung inflammation and fibrosis in mice (Ban et al., 2023).
In asthma, exosomes derived from M2 macrophages have the ability to selectively target and act on airway epithelial cells and airway smooth muscle cells, thereby alleviating inflammation and fibrosis progression in asthma. For instance, M2 macrophage exosomes treated with the scorpion and centipede contain miR-30b-5p, which mitigates severe asthma by inhibiting apoptosis in airway epithelial cells (Tang et al., 2022). Additionally, M2 macrophage-derived exosomes carrying miR-370 alleviate asthma progression by suppressing the fibroblast growth factor-1 (FGF1) 1/MAPK/STAT1 axis in airway smooth muscle cells, resulting in the inhibition of abnormal proliferation, invasion, and the production of fibrosis-related proteins (Li et al., 2021).
Synthetic exosomes can activate tumor-killing macrophages. For instance, the macrophage phosphoinositide 3-kinase gamma (PI3Kγ) is a crucial target for stimulating macrophage immunity and inhibiting tumor growth (Kaneda et al., 2016). This combination of gene editing technology with specific exosomes significantly improves targeting specificity and compatibility while reducing off-target effects and has been applied to various diseases. These exosomes specifically target and inhibit the expression of PI3Kγ in TAMs, leading to M1 polarization and reshaping of the tumor microenvironment, ultimately impeding tumor growth (Zhang et al., 2023). Drug delivery with engineered exosomes also enhances targeted drug delivery and relieves tumor inhibition of macrophages. For example, lung-specific exosomes combined with CD47 blockers and cisplatin treatment enhance macrophage phagocytic activity and promote T cell proliferation (Cui et al., 2022). In addition to the aforementioned engineered exosomes, plant-derived exosomes also exhibit the ability to activate tumor-killing macrophages. Nanovesicles derived from Artemisia species carrying plant-derived mitochondrial DNA (mtDNA) induce the cGAS (cyclic GMP–AMP synthase)–STING (stimulator of interferon genes) pathway, thereby promoting the transition of pro-tumor macrophages to an anti-tumor phenotype and enhancing the efficacy of PD-L1 inhibitors (Liu et al., 2023). Although traditional Chinese medicine offers numerous herbal treatments for tumors, oral absorption of these remedies is often inefficient and slow. However, research on plant-derived exosomes remains limited, holding the potential for unforeseen therapeutic benefits in tumor treatment.
Interestingly, some tumor-derived exosomes have the ability to hinder their own proliferation and invasion by inhibiting macrophages. For example, cancer cell-derived exosomes engineered with tumor-associated antigens can inhibit M2 tumor-associated macrophages and myeloid-derived suppressor cells (MDSCs), while increasing CD8- and CD4-positive T cells. This leads to a remodeling of the tumor environment, enhancing the efficacy of cancer therapy (Lin et al., 2023). Additionally, tumor cell-derived exosomes containing miR-770 can impede M2 macrophage polarization by targeting MAP3K1, thereby suppressing invasion in non-small cell lung cancer (Liu et al., 2021). These oncogenic exosomes limit their own development by suppressing pro-tumor macrophage polarization. We speculate that this may involve a regulatory mechanism within the tumor itself, employing a negative feedback regulation to restrict growth for improved vascularization or to extend the latency period, thus favoring the long-term survival of cancer cells. Similarly, the hepatitis B virus can also exert negative regulation on its own replication through exosomes, enabling a more insidious replication to evade complete elimination by immune cells. Hepatitis B-infected hepatocytes release exosomes containing HBV-miR-3, which stimulate macrophages to secrete IL-6, consequently limiting HBV replication (Gan et al., 2022). Moreover, the target molecules involved in promoting macrophage activation and polarization, as mentioned above, can be targeted to inhibit the formation of an unfavorable tumor microenvironment and suppress tumor progression.
First, exosomes derived from anti-tumor tumor-associated macrophages (TAMs) can inhibit cancer cell proliferation and metastasis. For instance, M1 macrophages release exosomes containing miRNA-let-7b-5p, which regulate the G protein subunit gamma 5 (GNG5) signaling pathway, suppressing cancer cell proliferation and inhibiting their anti-apoptotic ability (Peng et al., 2023). Cancer apoptosis cells irradiated with ultraviolet light, when co-cultured with macrophages, stimulate the production of exosomes rich in PTEN. These exosomes inhibit epithelial–mesenchymal transition (EMT), thereby limiting cancer progression and lung metastasis (Kim et al., 2019).
Additionally, exosomes from anti-tumor TAMs themselves exhibit anti-tumor effects. After artificial intervention to load drugs into them, their anti-cancer effects become more significant and highly specific. For example, exosomes derived from macrophages engineered to carry the anti-cancer drug paclitaxel (PTX) can be modified by incorporating aminoethyl anisamide (AA), a ligand specific to cancer sigma receptors, to enhance the targeting efficiency. Furthermore, adding polyethylene glycol (PEG) to these exosomes helps reduce immunogenicity and prolong circulation time. The resulting AA-PEG-modified exosomes loaded with PTX (AA-PEG-exoPTX) can selectively accumulate in cancer cells, significantly improving prognosis (Kim et al., 2018). A delivery system utilizing exosomes from M1 macrophages loaded with cisplatin effectively inhibits the proliferation of Lewis lung cancer cells in mice and induces apoptosis (Li et al., 2020).
Interestingly, M2 macrophages and their secreted exosomes are typically associated with promoting tumor behavior, but in some cancer cell lines, they can also exhibit anti-proliferative effects. For example, M2 macrophages release exosomes carrying miR-3917, which targets G protein-coupled receptor kinase 6 and inhibits proliferation, migration, and invasion of H1299 cells, while showing the opposite effect in A549 cells (cancer cell line) (Song et al., 2023). This difference may be attributed to variations in the expression frequency of miR-3917 downstream genes in different cell lines, providing new clues for the role of M2 macrophages in tumors. As shown in Table 2, extracellular vesicles target macrophages, mediating the repair of the pathological microenvironment in the lungs.
Macrophages represent the predominant immune cell population, participating in nearly all physiological and pathological processes in the lungs. Exosomes, as carriers for cellular material and information exchange, play a crucial role in regulating lung macrophages and reshaping the immune balance of the pulmonary microenvironment. In inflammatory lung diseases and lung cancer, exosomes derived from damaged airway epithelial cells, activated neutrophils, and tumor cells reshape the phenotype of macrophages. The reshaped macrophages exert pro-inflammatory and pro-tumor effects through exosome-mediated mechanisms, contributing to severe damage and tumor growth. Conversely, exosomes derived from stem cells or engineered exosomes reshape macrophage phenotypes and exhibit anti-inflammatory and anti-tumor effects through exosome-mediated mechanisms.
For future exosome-based therapies targeting inflammatory lung diseases and lung cancer, the pivotal directions of development may revolve around enhancing the specificity and efficiency. Specificity can be enhanced by constructing various target cell receptors and ligands on the exosome membrane surface, facilitating specific binding to target cells. Efficiency can be improved by exosomes derived from the same cell type for drug delivery, promoting enhanced “homing” compared to other drug carriers and exosomes derived from the deferent cell type, thus increasing uptake efficiency. Additionally, loading exosomes with tumor antigens can specifically enhance the antigen presentation ability of immune cells against tumor cells, efficiently activating endogenous “hot immune responses” to eliminate tumor cells. However, due to the complex composition and high heterogeneity of exosomes, challenges persist in achieving both specificity and efficiency.
JK: writing–original draft. PH: writing–review and editing. XW: writing–review and editing, supervision, and conceptualization. BW: writing–review and editing, validation, and funding acquisition.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article from the Medical Foundation of China (project number: zgyxjjh-wcwk-2023062001).
We thank the Medical Foundation of China and all the authors for their support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aegerter, H., Lambrecht, B. N., and Jakubzick, C. V. (2022). Biology of lung macrophages in health and disease. Immunity 55 (9), 1564–1580. doi:10.1016/j.immuni.2022.08.010
Ban, J., Liu, F., Zhang, Q., Chang, S., Zeng, X., and Chen, J. (2023). Macrophage-derived exosomal lncrna Mstrg.91634.7 inhibits fibroblasts activation by targeting Pink1 in silica-induced lung fibrosis. Toxicol. Lett. 372, 36–44. doi:10.1016/j.toxlet.2022.10.004
Basak, U., Sarkar, T., Mukherjee, S., Chakraborty, S., Dutta, A., Dutta, S., et al. (2023). Tumor-associated macrophages: an effective player of the tumor microenvironment. Front. Immunol. 14, 1295257. doi:10.3389/fimmu.2023.1295257
Bebelman, M. P., Smit, M. J., Pegtel, D. M., and Baglio, S. R. (2018). Biogenesis and function of extracellular vesicles in cancer. Pharmacol. Ther. 188, 1–11. doi:10.1016/j.pharmthera.2018.02.013
Boutilier, A. J., and Elsawa, S. F. (2021). Macrophage polarization states in the tumor microenvironment. Int. J. Mol. Sci. 22 (13), 6995. doi:10.3390/ijms22136995
Broad, K., Walker, S. A., Davidovich, I., Witwer, K., Talmon, Y., and Wolfram, J. (2023). Unraveling multilayered extracellular vesicles: speculation on cause. J. Extracell. Vesicles 12 (2), e12309. doi:10.1002/jev2.12309
Chen, W., Tang, D., Lin, J., Huang, X., Lin, S., Shen, G., et al. (2022a). Exosomal Circshkbp1 participates in non-small cell lung cancer progression through pkm2-mediated glycolysis. Mol. Ther. Oncolytics 24, 470–485. doi:10.1016/j.omto.2022.01.012
Chen, X., Tang, J., Shuai, W., Meng, J., Feng, J., and Han, Z. (2020). Macrophage polarization and its role in the pathogenesis of acute lung injury/acute respiratory distress syndrome. Inflamm. Res. 69 (9), 883–895. doi:10.1007/s00011-020-01378-2
Chen, X., Wang, L., Yu, H., Shen, Q., Hou, Y., Xia, Y.-X., et al. (2023). Irradiated lung cancer cell-derived exosomes modulate macrophage polarization by inhibiting Mid1 via mir-4655-5p. Mol. Immunol. 155, 58–68. doi:10.1016/j.molimm.2023.01.009
Chen, Z., Wu, H., Fan, W., Zhang, J., Yao, Y., Su, W., et al. (2022b). Naringenin suppresses beas-2b-derived extracellular vesicular cargoes disorder caused by cigarette smoke extract thereby inhibiting M1 macrophage polarization. Front. Immunol. 13, 930476. doi:10.3389/fimmu.2022.930476
Cheng, C., Wu, Y., Xiao, T., Xue, J., Sun, J., Xia, H., et al. (2021b). Mettl3-Mediated M6a modification of Zbtb4 mrna is involved in the smoking-induced emt in cancer of the lung. Mol. Ther. Nucleic Acids 23, 487–500. doi:10.1016/j.omtn.2020.12.001
Cheng, P., Li, S., and Chen, H. (2021a). Macrophages in lung injury, repair, and fibrosis. Cells 10 (2), 436. Epub 20210218. doi:10.3390/cells10020436
Colombo, M., Raposo, G., and Théry, C. (2014). Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 30, 255–289. doi:10.1146/annurev-cellbio-101512-122326
Cui, Z., Ruan, Z., Zeng, J., Sun, J., Ye, W., Xu, W., et al. (2022). Lung-specific exosomes for Co-delivery of Cd47 blockade and cisplatin for the treatment of non-small cell lung cancer. Thorac. Cancer 13 (19), 2723–2731. doi:10.1111/1759-7714.14606
Deng, H., Wu, L., Liu, M., Zhu, L., Chen, Y., Zhou, H., et al. (2020). Bone marrow mesenchymal stem cell-derived exosomes attenuate lps-induced ards by modulating macrophage polarization through inhibiting glycolysis in macrophages. Shock 54 (6), 828–843. doi:10.1097/SHK.0000000000001549
Deng, H., Zhu, L., Zhang, Y., Zheng, L., Hu, S., Zhou, W., et al. (2022). Differential lung protective capacity of exosomes derived from human adipose tissue, bone marrow, and umbilical cord mesenchymal stem cells in sepsis-induced acute lung injury. Oxid. Med. Cell Longev. 2022, 7837837. doi:10.1155/2022/7837837
Dong, B., Wang, C., Zhang, J., Zhang, J., Gu, Y., Guo, X., et al. (2021b). Exosomes from human umbilical cord mesenchymal stem cells attenuate the inflammation of severe steroid-resistant asthma by reshaping macrophage polarization. Stem Cell Res. Ther. 12 (1), 204. doi:10.1186/s13287-021-02244-6
Dong, T., Chen, X., Xu, H., Song, Y., Wang, H., Gao, Y., et al. (2022). Mitochondrial metabolism mediated macrophage polarization in chronic lung diseases. Pharmacol. Ther. 239, 108208. doi:10.1016/j.pharmthera.2022.108208
Dong, W., Liu, X., Yang, C., Wang, D., Xue, Y., Ruan, X., et al. (2021a). Glioma glycolipid metabolism: msi2-snord12b-fip1l1-zbtb4 feedback loop as a potential treatment target. Clin. Transl. Med. 11 (5), e411. doi:10.1002/ctm2.411
Esser, J., Gehrmann, U., D'Alexandri, F. L., Hidalgo-Estévez, A. M., Wheelock, C. E., Scheynius, A., et al. (2010). Exosomes from human macrophages and dendritic cells contain enzymes for leukotriene biosynthesis and promote granulocyte migration. J. Allergy Clin. Immunol. 126 (5), 1032–1040. doi:10.1016/j.jaci.2010.06.039
Feng, Z., Jing, Z., Li, Q., Chu, L., Jiang, Y., Zhang, X., et al. (2023). Exosomal stimate derived from type ii alveolar epithelial cells controls metabolic reprogramming of tissue-resident alveolar macrophages. Theranostics 13 (3), 991–1009. doi:10.7150/thno.82552
Feng, Z., Zhou, J., Liu, Y., Xia, R., Li, Q., Yan, L., et al. (2021). Epithelium- and endothelium-derived exosomes regulate the alveolar macrophages by targeting Rgs1 mediated calcium signaling-dependent immune response. Cell Death Differ. 28 (7), 2238–2256. doi:10.1038/s41418-021-00750-x
Gan, W., Chen, X., Wu, Z., Zhu, X., Liu, J., Wang, T., et al. (2022). The relationship between serum exosome hbv-mir-3 and current virological markers and its dynamics in chronic hepatitis B patients on antiviral treatment. Ann. Transl. Med. 10 (10), 536. doi:10.21037/atm-22-2119
Gopalakrishnan, A., Joseph, J., Shirey, K. A., Keegan, A. D., Boukhvalova, M. S., Vogel, S. N., et al. (2022). Protection against influenza-induced acute lung injury (ali) by enhanced induction of M2a macrophages: possible role of pparγ/Rxr ligands in il-4-induced M2a macrophage differentiation. Front. Immunol. 13, 968336. Epub 20220816. doi:10.3389/fimmu.2022.968336
Guan, B., Dai, X., Zhu, Y., and Geng, Q. (2023). M2 macrophage-derived exosomal mir-1911-5p promotes cell migration and invasion in lung adenocarcinoma by down-regulating Celf2 -activated Zbtb4 expression. Anticancer Drugs 34 (2), 238–247. doi:10.1097/CAD.0000000000001414
Guiot, J., Cambier, M., Boeckx, A., Henket, M., Nivelles, O., Gester, F., et al. (2020). Macrophage-derived exosomes attenuate fibrosis in airway epithelial cells through delivery of antifibrotic mir-142-3p. Thorax 75 (10), 870–881. doi:10.1136/thoraxjnl-2019-214077
Gunassekaran, G. R., Poongkavithai Vadevoo, S. M., Baek, M. C., and Lee, B. (2021). M1 macrophage exosomes engineered to foster M1 polarization and target the il-4 receptor inhibit tumor growth by reprogramming tumor-associated macrophages into M1-like macrophages. Biomaterials 278, 121137. Epub 20210917. doi:10.1016/j.biomaterials.2021.121137
Gurunathan, S., Kang, M.-H., and Kim, J.-H. (2021). A comprehensive review on factors influences biogenesis, functions, therapeutic and clinical implications of exosomes. Int. J. Nanomedicine 16, 1281–1312. doi:10.2147/IJN.S291956
Harrell, C. R., Miloradovic, D., Sadikot, R., Fellabaum, C., Markovic, B. S., Miloradovic, D., et al. (2020). Molecular and cellular mechanisms responsible for beneficial effects of mesenchymal stem cell-derived product "Exo-D-Mapps" in attenuation of chronic airway inflammation. Anal. Cell Pathol. (Amst) 2020, 3153891. doi:10.1155/2020/3153891
He, S., Chen, D., Hu, M., Zhang, L., Liu, C., Traini, D., et al. (2019). Bronchial epithelial cell extracellular vesicles ameliorate epithelial-mesenchymal transition in copd pathogenesis by alleviating M2 macrophage polarization. Nanomedicine 18, 259–271. Epub 20190411. doi:10.1016/j.nano.2019.03.010
Hu, Q., Wu, G., Wang, R., Ma, H., Zhang, Z., and Xue, Q. (2022). Cutting edges and therapeutic opportunities on tumor-associated macrophages in lung cancer. Front. Immunol. 13, 1007812. doi:10.3389/fimmu.2022.1007812
Hu, R., Xu, B., Ma, J., Li, L., Zhang, L., Wang, L., et al. (2023). Linc00963 promotes the malignancy and metastasis of lung adenocarcinoma by stabilizing Zeb1 and exosomes-induced M2 macrophage polarization. Mol. Med. 29 (1), 1. doi:10.1186/s10020-022-00598-y
Jiang, H., Wei, H., Wang, H., Wang, Z., Li, J., Ou, Y., et al. (2022). Zeb1-Induced metabolic reprogramming of glycolysis is essential for macrophage polarization in breast cancer. Cell Death Dis. 13 (3), 206. doi:10.1038/s41419-022-04632-z
Jiang, Y., Wang, J., Zhang, H., Min, Y., and Gu, T. (2023). Serum exosome-derived mir-7 exacerbates chronic obstructive pulmonary disease by regulating macrophage differentiation. Iran. J. Public Health 52 (3), 563–574. doi:10.18502/ijph.v52i3.12139
Jin, J., and Yu, G. (2022). Hypoxic lung cancer cell-derived exosomal mir-21 mediates macrophage M2 polarization and promotes cancer cell proliferation through targeting Irf1. World J. Surg. Oncol. 20 (1), 241. doi:10.1186/s12957-022-02706-y
Junior, L. Y.-S., Nguyen, H. T., Salmanida, F. P., and Chang, K.-T. (2021). Mertk+/Hi M2c macrophages induced by baicalin alleviate non-alcoholic fatty liver disease. Int. J. Mol. Sci. 22 (19), 10604. doi:10.3390/ijms221910604
Kalluri, R., and LeBleu, V. S. (2020). The Biology, function, and biomedical applications of exosomes. Science 367 (6478), eaau6977. doi:10.1126/science.aau6977
Kaneda, M. M., Messer, K. S., Ralainirina, N., Li, H., Leem, C. J., Gorjestani, S., et al. (2016). Pi3kγ is a molecular switch that controls immune suppression. Nature 539 (7629), 437–442. doi:10.1038/nature19834
Kim, M. S., Haney, M. J., Zhao, Y., Yuan, D., Deygen, I., Klyachko, N. L., et al. (2018). Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations. Nanomedicine 14 (1), 195–204. doi:10.1016/j.nano.2017.09.011
Kim, Y.-B., Ahn, Y.-H., Jung, J.-H., Lee, Y.-J., Lee, J.-H., and Kang, J. L. (2019). Programming of macrophages by uv-irradiated apoptotic cancer cells inhibits cancer progression and lung metastasis. Cell. Mol. Immunol. 16 (11), 851–867. doi:10.1038/s41423-019-0209-1
Kong, W., Zhang, L., Chen, Y., Yu, Z., and Zhao, Z. (2022). Cancer cell-derived exosomal Linc00313 induces M2 macrophage differentiation in non-small cell lung cancer. Clin. Transl. Oncol. 24 (12), 2395–2408. doi:10.1007/s12094-022-02907-7
Kulshreshtha, A., Ahmad, T., Agrawal, A., and Ghosh, B. (2013). Proinflammatory role of epithelial cell-derived exosomes in allergic airway inflammation. J. Allergy Clin. Immunol. 131 (4), 1194–1203. doi:10.1016/j.jaci.2012.12.1565
Lee, J.-W., Chun, W., Lee, H. J., Min, J.-H., Kim, S.-M., Seo, J.-Y., et al. (2021). The role of macrophages in the development of acute and chronic inflammatory lung diseases. Cells 10 (4), 897. doi:10.3390/cells10040897
Lei, J., Chen, P., Zhang, F., Zhang, N., Zhu, J., Wang, X., et al. (2021). M2 macrophages-derived exosomal microrna-501-3p promotes the progression of lung cancer via targeting wd repeat domain 82. Cancer Cell Int. 21 (1), 91. doi:10.1186/s12935-021-01783-5
Li, C., Deng, C., Zhou, T., Hu, J., Dai, B., Yi, F., et al. (2021d). Microrna-370 carried by M2 macrophage-derived exosomes alleviates asthma progression through inhibiting the fgf1/mapk/stat1 Axis. Int. J. Biol. Sci. 17 (7), 1795–1807. doi:10.7150/ijbs.59715
Li, J., Li, N., and Wang, J. (2020). M1 macrophage-derived exosome-encapsulated cisplatin can enhance its anti-lung cancer effect. Minerva Med. 114, 634–641. doi:10.23736/S0026-4806.20.06564-7
Li, P., Weng, Z., Li, P., Hu, F., Zhang, Y., Guo, Z., et al. (2021b). Batf3 promotes malignant phenotype of colorectal cancer through the S1pr1/P-Stat3/Mir-155-3p/Wdr82 Axis. Cancer Gene Ther. 28 (5), 400–412. Epub 20201014. doi:10.1038/s41417-020-00223-2
Li, X., Chen, Z., Ni, Y., Bian, C., Huang, J., Chen, L., et al. (2021c). Tumor-associated macrophages secret exosomal mir-155 and mir-196a-5p to promote metastasis of non-small-cell lung cancer. Transl. Lung Cancer Res. 10 (3), 1338–1354. doi:10.21037/tlcr-20-1255
Li, X., Yang, N., Cheng, Q., Zhang, H., Liu, F., and Shang, Y. (2021a). Mir-21-5p in macrophage-derived exosomes targets Smad7 to promote epithelial mesenchymal transition of airway epithelial cells. J. Asthma Allergy 14, 513–524. doi:10.2147/JAA.S307165
Liang, M., Chen, X., Wang, L., Qin, L., Wang, H., Sun, Z., et al. (2020). Cancer-derived exosomal Trim59 regulates macrophage Nlrp3 inflammasome activation to promote lung cancer progression. J. Exp. Clin. Cancer Res. 39 (1), 176. doi:10.1186/s13046-020-01688-7
Liang, Z. W., Yu, J., Gu, D. M., Liu, X. M., Liu, J., Wu, M. Y., et al. (2022). M2-Phenotype tumour-associated macrophages upregulate the expression of prognostic predictors Mmp14 and inhba in pancreatic cancer. J. Cell Mol. Med. 26 (5), 1540–1555. Epub 20220212. doi:10.1111/jcmm.17191
Lin, Y., Yi, M., Guan, X., Chen, E., Yang, L., Li, S., et al. (2023). Two birds with One stone" strategy for the lung cancer therapy with bioinspired aie aggregates. J. Nanobiotechnology 21 (1), 49. doi:10.1186/s12951-023-01799-1
Liu, C., Xi, L., Liu, Y., Mak, J. C. W., Mao, S., Wang, Z., et al. (2023a). An inhalable hybrid biomimetic nanoplatform for sequential drug release and remodeling lung immune homeostasis in acute lung injury treatment. ACS Nano 17, 11626–11644. doi:10.1021/acsnano.3c02075
Liu, G., Luo, Y., and Hou, P. (2022a). Prps2 enhances resistance to cisplatin via facilitating exosomes-mediated macrophage M2 polarization in non-small cell lung cancer. Immunol. Invest. 51 (5), 1423–1436. doi:10.1080/08820139.2021.1952217
Liu, H., Li, Y., Li, J., Liu, Y., and Cui, B. (2018). H3k4me3 and Wdr82 are associated with tumor progression and a favorable prognosis in human colorectal cancer. Oncol. Lett. 16 (2), 2125–2134. Epub 20180605. doi:10.3892/ol.2018.8902
Liu, J., Luo, R., Wang, J., Luan, X., Wu, D., Chen, H., et al. (2021b). Tumor cell-derived exosomal mir-770 inhibits M2 macrophage polarization via targeting Map3k1 to inhibit the invasion of non-small cell lung cancer cells. Front. Cell Dev. Biol. 9, 679658. doi:10.3389/fcell.2021.679658
Liu, J., Xiang, J., Jin, C., Ye, L., Wang, L., Gao, Y., et al. (2023b). Medicinal plant-derived mtdna via nanovesicles induces the cgas-sting pathway to remold tumor-associated macrophages for tumor regression. J. Nanobiotechnology 21 (1), 78. doi:10.1186/s12951-023-01835-0
Liu, X., Gao, C., Wang, Y., Niu, L., Jiang, S., and Pan, S. (2021a). Bmsc-derived exosomes ameliorate lps-induced acute lung injury by mir-384-5p-controlled alveolar macrophage autophagy. Oxid. Med. Cell Longev. 2021, 9973457. doi:10.1155/2021/9973457
Liu, Y., Li, L., and Song, X. (2022b). Exosomal Circpvt1 derived from lung cancer promotes the progression of lung cancer by targeting mir-124-3p/ezh2 Axis and regulating macrophage polarization. Cell Cycle 21 (5), 514–530. doi:10.1080/15384101.2021.2024997
Liu, Y.-J., and Wang, C. (2023). A review of the regulatory mechanisms of extracellular vesicles-mediated intercellular communication. Cell Commun. Signal 21 (1), 77. doi:10.1186/s12964-023-01103-6
Long, L., Hu, Y., Long, T., Lu, X., Tuo, Y., Li, Y., et al. (2021). Tumor-associated macrophages induced spheroid formation by ccl18-zeb1-M-csf feedback loop to promote transcoelomic metastasis of ovarian cancer. J. Immunother. Cancer 9 (12), e003973. doi:10.1136/jitc-2021-003973
Ma, J., Liu, L., Che, G., Yu, N., Dai, F., and You, Z. (2010). The M1 form of tumor-associated macrophages in non-small cell lung cancer is positively associated with survival time. BMC Cancer 10, 112. doi:10.1186/1471-2407-10-112
Mansouri, N., Willis, G. R., Fernandez-Gonzalez, A., Reis, M., Nassiri, S., Mitsialis, S. A., et al. (2019). Mesenchymal stromal cell exosomes prevent and revert experimental pulmonary fibrosis through modulation of monocyte phenotypes. JCI Insight 4 (21), e128060. doi:10.1172/jci.insight.128060
Mathivanan, S., Ji, H., and Simpson, R. J. (2010). Exosomes: extracellular organelles important in intercellular communication. J. Proteomics 73 (10), 1907–1920. doi:10.1016/j.jprot.2010.06.006
Melo, E. M., Oliveira, V. L. S., Boff, D., and Galvão, I. (2021). Pulmonary macrophages and their different roles in health and disease. Int. J. Biochem. Cell Biol. 141, 106095. doi:10.1016/j.biocel.2021.106095
Merten, M., Pakala, R., Thiagarajan, P., and Benedict, C. R. (1999). Platelet microparticles promote platelet interaction with subendothelial Matrix in a glycoprotein iib/iiia-dependent mechanism. Circulation 99 (19), 2577–2582. doi:10.1161/01.cir.99.19.2577
Mohapatra, S., Pioppini, C., Ozpolat, B., and Calin, G. A. (2021). Non-coding rnas regulation of macrophage polarization in cancer. Mol. Cancer 20 (1), 24. doi:10.1186/s12943-021-01313-x
Mondal, J., Pillarisetti, S., Junnuthula, V., Saha, M., Hwang, S. R., Park, I.-K., et al. (2023). Hybrid exosomes, exosome-like nanovesicles and engineered exosomes for therapeutic applications. J. Control Release 353, 1127–1149. doi:10.1016/j.jconrel.2022.12.027
Moon, H. G., Cao, Y., Yang, J., Lee, J. H., Choi, H. S., and Jin, Y. (2015). Lung epithelial cell-derived extracellular vesicles activate macrophage-mediated inflammatory responses via Rock1 pathway. Cell Death Dis. 6 (12), e2016. doi:10.1038/cddis.2015.282
Morrissey, S. M., Zhang, F., Ding, C., Montoya-Durango, D. E., Hu, X., Yang, C., et al. (2021). Tumor-derived exosomes drive immunosuppressive macrophages in a pre-metastatic niche through glycolytic dominant metabolic reprogramming. Cell Metab. 33 (10), 2040–2058.e10. doi:10.1016/j.cmet.2021.09.002
Niu, Z., Wang, L., Qin, X., Ye, Z., Xie, B., and Hu, Y. (2022). Macrophage derived mir-7219-3p-containing exosomes mediate fibroblast trans-differentiation by targeting Spry1 in silicosis. Toxicology 479, 153310. doi:10.1016/j.tox.2022.153310
Pegtel, D. M., and Gould, S. J. (2019). Exosomes. Annu. Rev. Biochem. 88, 487–514. doi:10.1146/annurev-biochem-013118-111902
Peng, J., Li, S., Li, B., Hu, W., and Ding, C. (2023). Exosomes derived from M1 macrophages inhibit the proliferation of the A549 and H1299 lung cancer cell lines via the mirna-let-7b-5p-gng5 Axis. PeerJ 11, e14608. doi:10.7717/peerj.14608
Pritchard, A., Tousif, S., Wang, Y., Hough, K., Khan, S., Strenkowski, J., et al. (2020). Lung tumor cell-derived exosomes promote M2 macrophage polarization. Cells 9 (5), 1303. doi:10.3390/cells9051303
Qian, B.-Z., and Pollard, J. W. (2010). Macrophage diversity enhances tumor progression and metastasis. Cell 141 (1), 39–51. doi:10.1016/j.cell.2010.03.014
Qian, J., Li, J., Ma, H., and Ji, W. (2023). Exosomal circ-adrm1 promotes lung adenocarcinoma progression and induces macrophage M2 polarization through regulating Mmp14 mrna and protein. Anticancer Drugs 34 (3), 333–343. doi:10.1097/CAD.0000000000001430
Qian, Q., Ma, Q., Wang, B., Qian, Q., Zhao, C., Feng, F., et al. (2022). Downregulated mir-129-5p expression inhibits rat pulmonary fibrosis by upregulating Stat1 gene expression in macrophages. Int. Immunopharmacol. 109, 108880. doi:10.1016/j.intimp.2022.108880
Qin, X., Lin, X., Liu, L., Li, Y., Li, X., Deng, Z., et al. (2021). Macrophage-derived exosomes mediate silica-induced pulmonary fibrosis by activating fibroblast in an endoplasmic reticulum stress-dependent manner. J. Cell Mol. Med. 25 (9), 4466–4477. doi:10.1111/jcmm.16524
Qiu, S., Xie, L., Lu, C., Gu, C., Xia, Y., Lv, J., et al. (2022). Gastric cancer-derived exosomal mir-519a-3p promotes liver metastasis by inducing intrahepatic M2-like macrophage-mediated angiogenesis. J. Exp. Clin. Cancer Res. 41 (1), 296. Epub 20221010. doi:10.1186/s13046-022-02499-8
Rao, X., Zhou, X., Wang, G., Jie, X., Xing, B., Xu, Y., et al. (2022). Nlrp6 is required for cancer-derived exosome-modified macrophage M2 polarization and promotes metastasis in small cell lung cancer. Cell Death Dis. 13 (10), 891. doi:10.1038/s41419-022-05336-0
Ren, J., Liu, Y., Yao, Y., Feng, L., Zhao, X., Li, Z., et al. (2021). Intranasal delivery of msc-derived exosomes attenuates allergic asthma via expanding il-10 producing lung interstitial macrophages in mice. Int. Immunopharmacol. 91, 107288. doi:10.1016/j.intimp.2020.107288
Shang, Y., Sun, Y., Xu, J., Ge, X., Hu, Z., Xiao, J., et al. (2020). Exosomes from Mmu_Circ_0001359-modified adscs attenuate airway remodeling by enhancing Foxo1 signaling-mediated M2-like macrophage activation. Mol. Ther. Nucleic Acids 19, 951–960. doi:10.1016/j.omtn.2019.10.049
Shao, M.-M., Pei, X.-B., Chen, Q.-Y., Wang, F., Wang, Z., and Zhai, K. (2023). Macrophage-derived exosome promotes regulatory T cell differentiation in malignant pleural effusion. Front. Immunol. 14, 1161375. doi:10.3389/fimmu.2023.1161375
Shotland, A. M., Fontenot, A. P., and McKee, A. S. (2021). Pulmonary macrophage cell death in lung health and disease. Am. J. Respir. Cell Mol. Biol. 64 (5), 547–556. doi:10.1165/rcmb.2020-0420TR
Song, C., Li, H., Li, Y., Dai, M., Zhang, L., Liu, S., et al. (2019). Nets promote ali/ards inflammation by regulating alveolar macrophage polarization. Exp. Cell Res. 382 (2), 111486. Epub 20190628. doi:10.1016/j.yexcr.2019.06.031
Song, S., Zhao, Y., Wang, X., Tong, X., Chen, X., and Xiong, Q. (2023). M2 macrophages-derived exosomal mir-3917 promotes the progression of lung cancer via targeting Grk6. Biol. Chem. 404 (1), 41–57. doi:10.1515/hsz-2022-0162
Sun, L., Fan, M., Huang, D., Li, B., Xu, R., Gao, F., et al. (2021b). Clodronate-loaded liposomal and fibroblast-derived exosomal hybrid system for enhanced drug delivery to pulmonary fibrosis. Biomaterials 271, 120761. doi:10.1016/j.biomaterials.2021.120761
Sun, N.-N., Zhang, Y., Huang, W.-H., Zheng, B.-J., Jin, S.-Y., Li, X., et al. (2021a). Macrophage exosomes transfer angiotensin ii type 1 receptor to lung fibroblasts mediating bleomycin-induced pulmonary fibrosis. Chin. Med. J. Engl. 134 (18), 2175–2185. doi:10.1097/CM9.0000000000001605
Tang, B., Wu, Y., Zhang, Y., Cheng, Y., Wu, Y., and Fang, H. (2022). Scorpion and centipede alleviates severe asthma through M2 macrophage-derived exosomal mir-30b-5p. Aging (Albany NY) 14 (9), 3921–3940. doi:10.18632/aging.204053
TJonck, W., and Bain, C. C. (2023). The role of monocyte-derived macrophages in the lung: it's all about context. Int. J. Biochem. Cell Biol. 159, 106421. doi:10.1016/j.biocel.2023.106421
van der Pol, E., Böing, A. N., Harrison, P., Sturk, A., and Nieuwland, R. (2012). Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 64 (3), 676–705. doi:10.1124/pr.112.005983
Wan, X., Xie, B., Sun, H., Gu, W., Wang, C., Deng, Q., et al. (2022). Exosomes derived from M2 type tumor-associated macrophages promote osimertinib resistance in non-small cell lung cancer through mstrg.292666.16-mir-6836-5p-mapk8ip3 Axis. Cancer Cell Int. 22 (1), 83. doi:10.1186/s12935-022-02509-x
Wang, H., Wang, L., Pan, H., Wang, Y., Shi, M., Yu, H., et al. (2020). Exosomes derived from macrophages enhance aerobic glycolysis and chemoresistance in lung cancer by stabilizing C-myc via the inhibition of Nedd4l. Front. Cell Dev. Biol. 8, 620603. doi:10.3389/fcell.2020.620603
Wang, L., Chen, Q., Yu, Q., Xiao, J., and Zhao, H. (2021). Cigarette smoke extract-treated airway epithelial cells-derived exosomes promote M1 macrophage polarization in chronic obstructive pulmonary disease. Int. Immunopharmacol. 96, 107700. doi:10.1016/j.intimp.2021.107700
Wang, Z., Zhang, C., Meng, J., Jiao, Z., Bao, W., Tian, H., et al. (2023). A targeted exosome therapeutic confers both cfdna scavenging and macrophage polarization for ameliorating rheumatoid arthritis. Adv. Mater 35 (48), e2302503. Epub 20231027. doi:10.1002/adma.202302503
Willis, G. R., Fernandez-Gonzalez, A., Anastas, J., Vitali, S. H., Liu, X., Ericsson, M., et al. (2018). Mesenchymal stromal cell exosomes ameliorate experimental bronchopulmonary dysplasia and restore lung function through macrophage immunomodulation. Am. J. Respir. Crit. Care Med. 197 (1), 104–116. doi:10.1164/rccm.201705-0925OC
Wolf, P. (1967). The nature and significance of platelet products in human plasma. Br. J. Haematol. 13 (3), 269–288. doi:10.1111/j.1365-2141.1967.tb08741.x
Wu, J., Yuan, M., Shen, J., Chen, Y., Zhang, R., Chen, X., et al. (2022). Effect of modified jianpi yangzheng on regulating content of Pkm2 in gastric cancer cells-derived exosomes. Phytomedicine 103, 154229. doi:10.1016/j.phymed.2022.154229
Xu, K., Zhang, C., Du, T., Gabriel, A. N. A., Wang, X., Li, X., et al. (2021). Progress of exosomes in the diagnosis and treatment of lung cancer. Biomed. Pharmacother. = Biomedecine Pharmacother. 134, 111111. doi:10.1016/j.biopha.2020.111111
Xu, Z., Chen, Y., Ma, L., Chen, Y., Liu, J., Guo, Y., et al. (2022). Role of exosomal non-coding rnas from tumor cells and tumor-associated macrophages in the tumor microenvironment. Mol. Ther. 30 (10), 3133–3154. Epub 20220409. doi:10.1016/j.ymthe.2022.01.046
Yan, Z., Wen, J.-X., Cao, X.-S., Zhao, W., Han, Y.-L., Wen, X.-H., et al. (2022). Tumor cell-derived exosomal microrna-146a promotes non-small cell lung cancer cell invasion and proliferation by inhibiting M1 macrophage polarization. Ann. Transl. Med. 10 (24), 1307. doi:10.21037/atm-22-5565
Yang, R., Liao, Y., Wang, L., He, P., Hu, Y., Yuan, D., et al. (2019). Exosomes derived from M2b macrophages attenuate dss-induced colitis. Front. Immunol. 10, 2346. doi:10.3389/fimmu.2019.02346
Yang, Z., Chen, F., Wang, F., Chen, X., Zheng, B., Liao, X., et al. (2023). Identification of Zbtb4 as an immunological biomarker that can inhibit the proliferation and invasion of pancreatic cancer. BMC Cancer 23 (1), 263. doi:10.1186/s12885-023-10749-x
Yao, M.-Y., Zhang, W.-H., Ma, W.-T., Liu, Q.-H., Xing, L.-H., and Zhao, G.-F. (2019). Microrna-328 in exosomes derived from M2 macrophages exerts a promotive effect on the progression of pulmonary fibrosis via Fam13a in a rat model. Exp. Mol. Med. 51 (6), 1–16. doi:10.1038/s12276-019-0255-x
Ye, C., Li, H., Bao, M., Zhuo, R., Jiang, G., and Wang, W. (2020). Alveolar macrophage - derived exosomes modulate severity and outcome of acute lung injury. Aging (Albany NY) 12 (7), 6120–6128. doi:10.18632/aging.103010
Ye, Y., Xu, Y., Lai, Y., He, W., Li, Y., Wang, R., et al. (2018). Long non-coding rna cox-2 prevents immune evasion and metastasis of hepatocellular carcinoma by altering M1/M2 macrophage polarization. J. Cell Biochem. 119 (3), 2951–2963. doi:10.1002/jcb.26509
Yu, Y., Zhou, Y., Di, C., Zhao, C., Chen, J., Su, W., et al. (2021). Increased airway epithelial cell-derived exosomes activate macrophage-mediated allergic inflammation via Cd100 shedding. J. Cell Mol. Med. 25 (18), 8850–8862. doi:10.1111/jcmm.16843
Yuan, S., Chen, W., Yang, J., Zheng, Y., Ye, W., Xie, H., et al. (2022). Tumor-associated macrophage-derived exosomes promote egfr-tki resistance in non-small cell lung cancer by regulating the akt, erk1/2 and Stat3 signaling pathways. Oncol. Lett. 24 (4), 356. doi:10.3892/ol.2022.13476
Yuan, Z., Bedi, B., and Sadikot, R. T. (2018). Bronchoalveolar lavage exosomes in lipopolysaccharide-induced septic lung injury. J. Vis. Exp., 57737. doi:10.3791/57737
Yunna, C., Mengru, H., Lei, W., and Weidong, C. (2020). Macrophage M1/M2 polarization. Eur. J. Pharmacol. 877, 173090. doi:10.1016/j.ejphar.2020.173090
Zaiets, I., Gunewardena, S., Menne, S., Weinman, S. A., and Gudima, S. O. (2023). Sera of individuals chronically infected with hepatitis B virus (hbv) contain diverse rna types produced by hbv replication or derived from integrated hbv DNA. J. Virol. 97 (3), e0195022. doi:10.1128/jvi.01950-22
Zhang, F., Sang, Y., Chen, D., Wu, X., Wang, X., Yang, W., et al. (2021c). M2 macrophage-derived exosomal long non-coding rna agap2-as1 enhances radiotherapy immunity in lung cancer by reducing microrna-296 and elevating Notch2. Cell Death Dis. 12 (5), 467. doi:10.1038/s41419-021-03700-0
Zhang, L., Lin, Y., Li, S., Guan, X., and Jiang, X. (2023). In situ reprogramming of tumor-associated macrophages with internally and externally engineered exosomes. Angew. Chem. Int. Ed. Engl. 62 (11), e202217089. doi:10.1002/anie.202217089
Zhang, L., Wang, Y., Wu, G., Xiong, W., Gu, W., and Wang, C.-Y. (2018). Macrophages: friend or foe in idiopathic pulmonary fibrosis? Respir. Res. 19 (1), 170. doi:10.1186/s12931-018-0864-2
Zhang, M., Wang, D., Zhu, T., and Yin, R. (2017). Rassf4 overexpression inhibits the proliferation, invasion, emt, and wnt signaling pathway in osteosarcoma cells. Oncol. Res. 25 (1), 83–91. doi:10.3727/096504016X14719078133447
Zhang, Q., and Sioud, M. (2023). Tumor-associated macrophage subsets: shaping polarization and targeting. Int. J. Mol. Sci. 24 (8), 7493. doi:10.3390/ijms24087493
Zhang, S., Xie, B., Wang, L., Yang, H., Zhang, H., Chen, Y., et al. (2021b). Macrophage-mediated vascular permeability via vla4/vcam1 pathway dictates ascites development in ovarian cancer. J. Clin. Invest. 131 (3), e140315. doi:10.1172/JCI140315
Zhang, Y., Fan, Y., Jing, X., Zhao, L., Liu, T., Wang, L., et al. (2021a). Otud5-Mediated deubiquitination of yap in macrophage promotes M2 phenotype polarization and favors triple-negative breast cancer progression. Cancer Lett. 504, 104–115. doi:10.1016/j.canlet.2021.02.003
Zhao, L., Wang, H., Fu, J., Wu, X., Liang, X.-Y., Liu, X.-Y., et al. (2022). Microfluidic-based exosome isolation and highly sensitive aptamer exosome membrane protein detection for lung cancer diagnosis. Biosens. Bioelectron. 214, 114487. doi:10.1016/j.bios.2022.114487
Zhou, S., Lan, Y., Li, Y., Li, Z., Pu, J., and Wei, L. (2022). Hypoxic tumor-derived exosomes induce M2 macrophage polarization via pkm2/ampk to promote lung cancer progression. Cell Transpl. 31, 9636897221106998. doi:10.1177/09636897221106998
Zhou, Y., Gao, Y., Zhang, W., Chen, Y., Jin, M., and Yang, Z. (2021). Exosomes derived from induced pluripotent stem cells suppresses M2-type macrophages during pulmonary fibrosis via mir-302a-3p/tet1 Axis. Int. Immunopharmacol. 99, 108075. doi:10.1016/j.intimp.2021.108075
Zhu, L., Sun, H.-T., Wang, S., Huang, S.-L., Zheng, Y., Wang, C.-Q., et al. (2020). Isolation and characterization of exosomes for cancer research. J. Hematol. Oncol. 13 (1), 152. doi:10.1186/s13045-020-00987-y
Keywords: exosomes, macrophage, immunomodulator, chronic lung disease, immune balance
Citation: Kang J, Hua P, Wu X and Wang B (2024) Exosomes: efficient macrophage-related immunomodulators in chronic lung diseases. Front. Cell Dev. Biol. 12:1271684. doi: 10.3389/fcell.2024.1271684
Received: 02 August 2023; Accepted: 15 March 2024;
Published: 09 April 2024.
Edited by:
Dwijendra K. Gupta, Allahabad University, IndiaReviewed by:
Patrick Fordjour Asare, University of Adelaide, AustraliaCopyright © 2024 Kang, Hua, Wu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaojing Wu, d3V4aWFvamluZzkxNEAxNjMuY29t; Bin Wang, YndhbmdAamx1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.