
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell Dev. Biol., 20 March 2023
Sec. Cell Adhesion and Migration
Volume 11 - 2023 | https://doi.org/10.3389/fcell.2023.959611
This article is part of the Research TopicMulticellularity: Views from Cellular Signaling and MechanicsView all 10 articles
 Ignacio Montenegro-Rojas1†
Ignacio Montenegro-Rojas1† Guillermo Yañez2,3†
Guillermo Yañez2,3† Emily Skog1†
Emily Skog1† Oscar Guerrero-Calvo1
Oscar Guerrero-Calvo1 Martin Andaur-Lobos1Luca Dolfi4,5Alessandro Cellerino6,7
Martin Andaur-Lobos1Luca Dolfi4,5Alessandro Cellerino6,7 Mauricio Cerda8,9,10
Mauricio Cerda8,9,10 Miguel L. Concha8,9,11
Miguel L. Concha8,9,11 Cristina Bertocchi12,13
Cristina Bertocchi12,13 Nicolás O. Rojas1
Nicolás O. Rojas1 Andrea Ravasio1*Timothy J. Rudge2,3*
Andrea Ravasio1*Timothy J. Rudge2,3*Introduction: Deciphering the biological and physical requirements for the outset of multicellularity is limited to few experimental models. The early embryonic development of annual killifish represents an almost unique opportunity to investigate de novo cellular aggregation in a vertebrate model. As an adaptation to seasonal drought, annual killifish employs a unique developmental pattern in which embryogenesis occurs only after undifferentiated embryonic cells have completed epiboly and dispersed in low density on the egg surface. Therefore, the first stage of embryogenesis requires the congregation of embryonic cells at one pole of the egg to form a single aggregate that later gives rise to the embryo proper. This unique process presents an opportunity to dissect the self-organizing principles involved in early organization of embryonic stem cells. Indeed, the physical and biological processes required to form the aggregate of embryonic cells are currently unknown.
Methods: Here, we developed an in silico, agent-based biophysical model that allows testing how cell-specific and environmental properties could determine the aggregation dynamics of early Killifish embryogenesis. In a forward engineering approach, we then proceeded to test two hypotheses for cell aggregation (cell-autonomous and a simple taxis model) as a proof of concept of modeling feasibility. In a first approach (cell autonomous system), we considered how intrinsic biophysical properties of the cells such as motility, polarity, density, and the interplay between cell adhesion and contact inhibition of locomotion drive cell aggregation into self-organized clusters. Second, we included guidance of cell migration through a simple taxis mechanism to resemble the activity of an organizing center found in several developmental models.
Results: Our numerical simulations showed that random migration combined with low cell-cell adhesion is sufficient to maintain cells in dispersion and that aggregation can indeed arise spontaneously under a limited set of conditions, but, without environmental guidance, the dynamics and resulting structures do not recapitulate in vivo observations.
Discussion: Thus, an environmental guidance cue seems to be required for correct execution of early aggregation in early killifish development. However, the nature of this cue (e.g., chemical or mechanical) can only be determined experimentally. Our model provides a predictive tool that could be used to better characterize the process and, importantly, to design informed experimental strategies.
Annual killifish have a unique early developmental pattern that differs from most teleost species. Unlike non-annual species, for whom most morphogenetic movements are concomitant, annual killifish have separated epiboly from embryo formation, resulting in an initial phase of cell dispersal that is followed by a process of cell aggregation, with embryonic cells occurring through active cell migration in the confined space between the enveloping layer (EVL) and the yolk syncytial layer (YSL) (Figure 1) (Dolfi et al., 2014; Concha and Reig, 2022). The embryonic cells remain undifferentiated, with stem-like properties during epiboly and until the end of the dispersion phase, when the cells start to aggregate at one pole of the embryo and initiate the genetic and morphogenetic processes leading to the formation of germ layers and the establishment of the embryonic axis (Wourms, 1972; Pereiro et al., 2017; Márquez et al., 2019) (Figure 1). These processes, which span between several hours and a few days depending on the specific killifish species and the environmental conditions (Dolfi et al., 2014; Márquez et al., 2019), occur in the context of a nearly spherical egg (about 1 mm diameter) and can be easily visualized in a living animal, as eggs are optically transparent and develop outside the mother (Wourms, 1972; Pereiro et al., 2017; Reig et al., 2017; Concha and Reig, 2022). The dispersion phase is characterized by a random walk of embryonic cells moving at a very low density (Márquez et al., 2019), while the cellular processes and morphogenetic mechanisms that form the aggregate are still unknown. It has been proposed that self-organizing processes may break the initial symmetry of the embryo and initiate the aggregation process (Pereiro et al., 2017; Abitua et al., 2021), since the molecular signals involved in embryo formation are apparently non-polarized during the stages prior to the aggregate formation. However, it cannot be ruled out that an organizing center, possibly located in extraembryonic structures, provides the signals that initiate the aggregate formation (Pereiro et al., 2017) as has been shown in other non-annual teleost species (Concha and Reig, 2022).
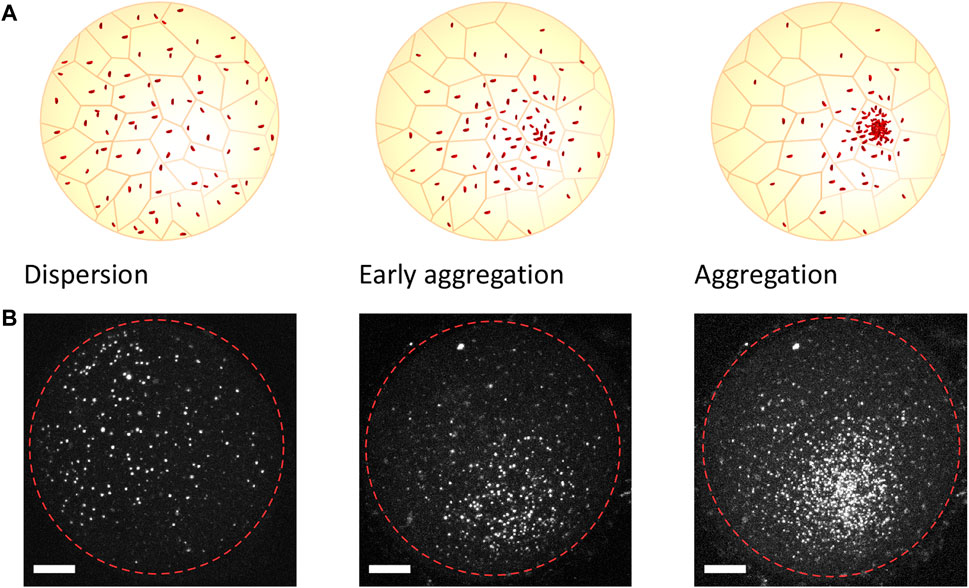
FIGURE 1. Stages of the early development of annual killifish as recorded in vivo. Cartoon representation (A) and confocal images of embryonic cell nuclei (B) stained using the FUCCI construct, illustrating the stages of the early development of killifish embryos. Following the dispersive state (left), the cells move directionally toward a pole of the embryo (center) and form an aggregate (right). The dashed red circles indicate the approximate outline of the embryo. The time between the stages varies between several hours to a few days, depending on the environmental condition and specific killifish species (Dolfi et al., 2014; Márquez et al., 2019). Scale bar, 200 μm.
In silico modeling proved to be a powerful tool to accurately capture the essential features of various biological systems, such as wound healing (Ravasio et al., 2015a), tissue expansion (Ravasio et al., 2015b), cancer invasion (Stichel et al., 2017), and embryonic development (CONTE et al., 2008; Cai et al., 2016; Pereiro et al., 2017; Stepien et al., 2019). Thus, it has been proposed that they could be used as predictive tools to design informed experimental strategies (Kabla, 2012; Phillips, 2015). Here, we used an in silico model to understand the mechanical requirements for killifish cells to 1) remain in a dispersed state and 2) aggregate at the embryo pole to initiate embryogenesis. In our model, motile cells are represented by three-dimensional self-propelled particle spheres, which is a 3D framework commonly used to model collective cell dynamics (Belmonte et al., 2008; Henkes et al., 2011; Sepúlveda et al., 2013; Méhes and Vicsek, 2014; Tarle et al., 2015). This approach has been extended to incorporate biologically relevant interactions such as cell–substrate friction, intercellular and cell–substrate adhesions (Kanchanawong et al., 2010; Bertocchi et al., 2017), and contact inhibition of locomotion (CIL) (Abercrombie and Heaysman, 1954; Roycroft and Mayor, 2016). When generalized, this model can exhibit diverse dynamic states, such as gas phases, polar liquids, and 3D aggregates, depending on the parameter explored (Bray, 1993; Mladek et al., 2006; Moreno and Likos, 2007; Redner et al., 2013). Although these states were experimentally observed at high cell densities, it is an open question as to whether such mechanisms could account for the aggregation behavior observed at low cell densities found in the early stages of killifish development. Furthermore, to date, models have not considered the specific geometry of this process, such as the cells moving in confinement and on curved surfaces with spherical topology. Typically, these studies use periodic boundary conditions on a plane, giving a toroidal topology (Bellomo et al., 2015). Our model incorporates realistic conditions in terms of cell density, geometrical and mechanical properties of the EVL and the YSL, and their effect on the dynamics of embryonic cells. Thus, the in silico investigation presented aims to provide a flexible framework to model the early teleost development, which can help predict the minimal mechanical requirements for cell aggregation under the specific conditions of annual killifish early development. As the in vivo system is poorly understood and presents various intrinsic experimental challenges (e.g., coriaceous chorion), our forward engineering approach can provide useful information, enabling an informed experimental investigation of the biological system.
During the early stages of killifish embryo development, undifferentiated stem cells move tangentially between the inner surface of the epithelial enveloping cell layer and the yolk syncytial layer (Figure 2A). The system is modeled here in two distinct ways: a cell-autonomous system that includes mechanisms that are intrinsic to the cells and the same cells that are under the influence of guidance from the environment toward an organizing center.
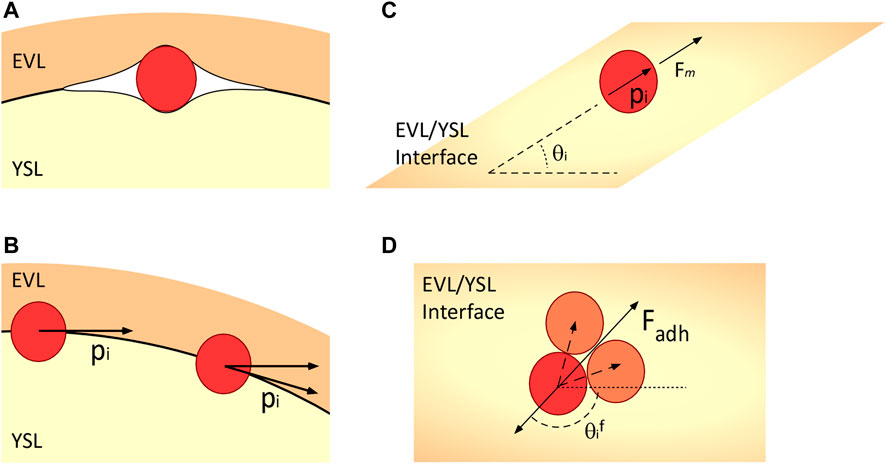
FIGURE 2. Schematic representation of the model’s major components. (A) Embryonic cells move between the EVL and the YSL, which are roughly spherical. They are subject to forces due to the deformation of the adjacent cell layers. (B) Polarity of the cells is constrained to the tangential plane as they move. (C) Contact inhibition of locomotion repolarizes cells away from the average position of their neighbors, and adhesion forces attract them to their neighbors. (D) Angle with respect to the average neighbor position. The dashed vectors indicate the directions to the neighbors of the red cell, the bottom solid arrow indicates the reference direction, and
We present, here, a variation of the model proposed by Smeets et al. (2016) (Basan et al., 2013) for autonomous motile cells, extended to three dimensions and including the physical and geometrical constraints imposed by the EVL and the YSL, where the cells migrate tangentially to the surface of the YSL (Figure 2B).
The cells have an intrinsic motile force,
with the left-hand term and the motile force with direction
with
The adhesion and repulsion between cells may play a significant role in the formation of aggregates. These forces are determined by the interaction between the cell–cell adhesion energy
for pairs of cells i and j in contact (
which is the force applied to cell i by the YSL when
which is the force applied to cell i by the EVL when
In our model, the cell polarity vector
where
where
Contact inhibition of locomotion (CIL), which works as a repulsion interaction causing cells to steer away from each other, is another important determinant of cell migration, which we wish to test in our model. For CIL, we model the repolarization direction,
for the neighboring cells j, the desired direction vector in the tangent plane with normal
Thus, the cells will turn away from contacting cells at a rate of
The system can again be normalized by the dimensionless parameter
This model can be solved numerically (see Methods) for a range of parameters, providing a powerful tool to simulate developmental processes occurring in a realistic geometrical setting. Using the model described previously, we first examined the effect of the cell density on the collective behavior of cells, with other parameters for cell–cell adhesion and contact inhibition being fixed at Wc = 1 and Ψ = 0.5, respectively. Spherical cells of radius 1 were initialized at random positions on the surface of a sphere with a radius 25, representing the YSL, and their migration simulated 500 time steps. At a cell packing density (
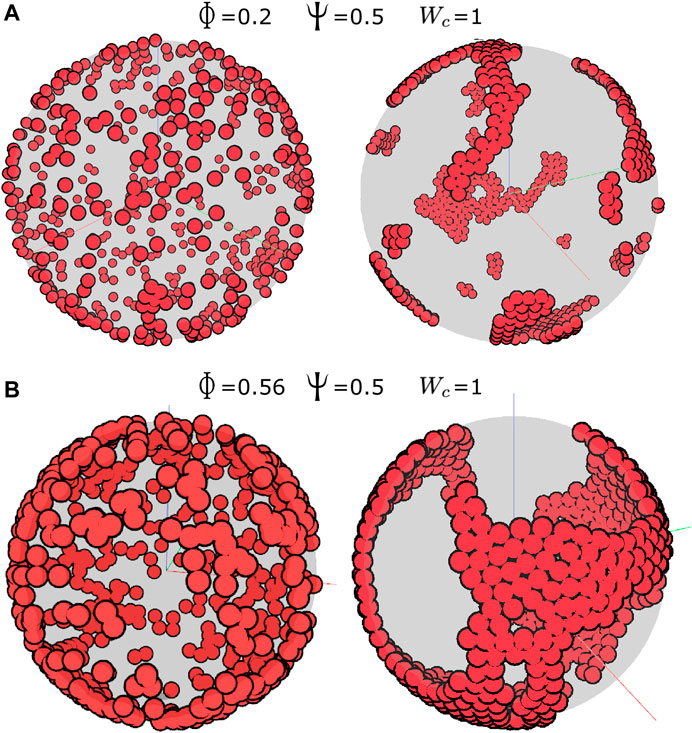
FIGURE 3. Aggregation in the cell-autonomous system. (A) Cell density
To understand the effect of cell–cell adhesion and CIL on these dynamics, we scanned the parameter space, simulating the system with a range of values of Wc and Ψ for each density (Φ = 0.2 or 0.56). Figure 4 shows the final configurations of cells after 500 time steps for each parameter combination. A clear pattern emerges, where at low values of Wc, the system maintains its dispersed condition, and at high Wc, aggregation occurs (Figures 4A, B). Similar to what was shown previously, large single aggregates can be obtained at high densities and strong cell–cell adhesions, whereas at a first approximation, Ψ appears to have only a marginal effect on the qualitative appearance of the cell aggregate. To quantify these effects, we computed the maximum cluster size at the end of simulations for each of the parameter combinations (Figure 5), and we also analyzed its time evolution (Figure 6). This analysis clearly confirms that larger aggregates are formed at a higher Wc, where both cell densities are considered, and that Wc has a greater influence on the formation of aggregates as compared to Ψ. Interestingly, it also showed a sort of weak biphasic effect of Ψ on the cluster size, with larger aggregates being formed at intermediate values (between 0.75 and 1.5). However, high variations in final cluster sizes can be seen at these values, which may be due to the more pronounced stochasticity of the process.
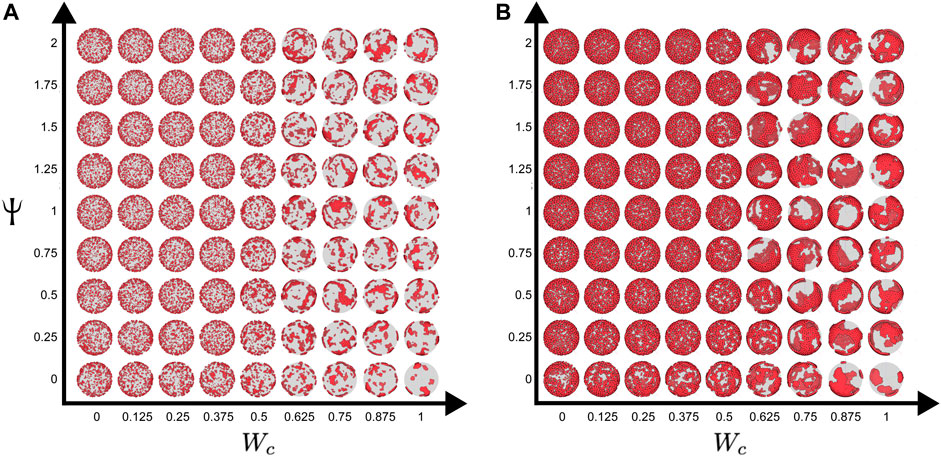
FIGURE 4. Parameter scan of the cell-autonomous system over different cell adhesion and CIL parameters. (A) Cell density of
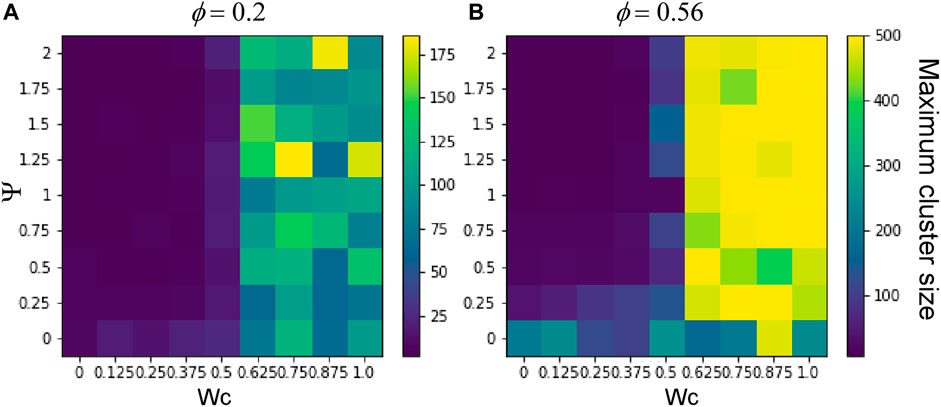
FIGURE 5. Maximum cluster size as a function of CIL and cell–cell adhesion. The maximum cluster sizes for the final states (after 500 time steps, each grid point represents the average of 25 simulations) of simulations with cell densities of (A)
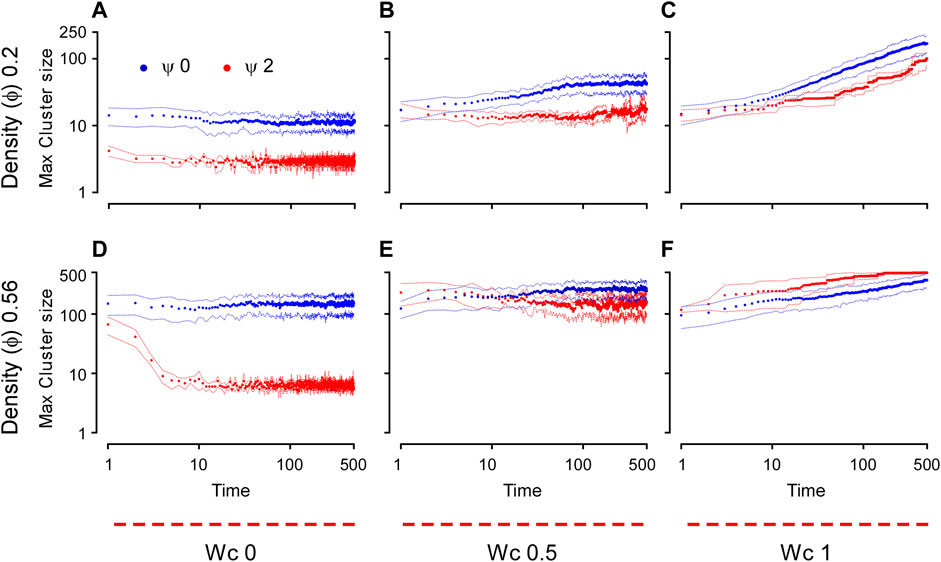
FIGURE 6. Time dynamics of the aggregation of the cell-autonomous system measured by the maximum cluster size. The cluster size is plotted here as a function of time for different cell densities, both conditions include 500 cells in each simulation (
Numerical simulations also allow us detailed insights into the dynamics of the aggregation process, which can be quantified by the time variation in the maximum cluster size (Figure 6). With no cell–cell adhesion (Wc = 0), the system was largely static, with no increase in the cluster size at either density tested after 10 dimensionless time units. This is a condition that closely resembles the dynamics observed during the dispersion state. However, as Wc was increased, the maximum size of the clusters tended toward the power law dynamics. On the other hand, CIL appeared to have a marginal effect on the dynamics (rate) of aggregation for intermediate and high values of Wc (Figures 6B, C, E, F), whereas a marked effect of CIL can be seen when Wc is absent (Figures 6A, D). In these conditions, a high CIL (Ψ = 2) caused an initial decline in the size of the cluster and a generally low aggregation as compared to Ψ = 0. This phenomenon, which was more pronounced at high cell densities, most likely reflects the scattering effect provided by CIL.
The results presented previously show that purely cell-autonomous behaviors could explain the dispersion state by keeping low cell–cell adhesion, but they were not sufficient to lead to the formation of a single aggregate at one of the embryonic poles, as seen in annual killifish early embryogenesis. It is, therefore, possible that the information provided by environmental cues is needed in the form of an organizing center that causes cells to orient toward a specific position of the embryo, where possibly a site-specific increase in Wc for cells reaching the location could initiate the aggregation process. The cells might preferentially move toward the organizing center by a variety of mechanisms including chemotaxis, durotaxis, and haptotaxis (CARTER, 1967; Bellomo et al., 2015; Espina et al., 2022). A powerful feature of our model and its software implementation is that such external environmental factors can easily be included. As a proof of concept, we show, here, a simple model of repolarization that shows a bias for the cells to repolarize toward the organizing center. However, experiments support a variety of possible guidance mechanisms (Sarris and Sixt, 2015a). As a feasibility study, in the Supplementary Material, we further expand two more models (i.e., “adjustment of directional speed along gradient” and “slowing down at the source”) to demonstrate that our modeling framework provides a robust and flexible tool to model a variety of taxis models (see Supplementary Material, Figures S1, S2).
If the organizing center is at position
Then, the angular equation of motion becomes
In this simple model, the rate of repolarization does not depend on the distance from the organizing center, nor are there any effects on the speed of cell motion. Various mechanisms and models of taxis have been proposed (Sarris and Sixt, 2015b), which while not considered here, are straightforward to implement in our modeling framework.
To test the effect of this simple external guidance (taxis), we then performed a series of simulations using the more realistic (i.e., closest to in vivo) conditions with cell number = 500 and sphere radius = 25, for density
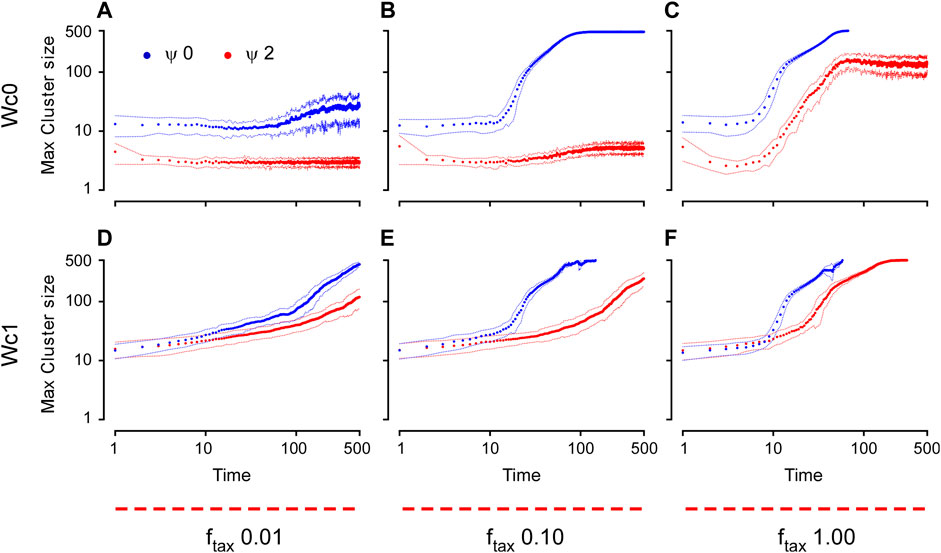
FIGURE 7. Time dynamics of aggregation for the environmental guidance model measured by the maximum cluster size. The cluster size is plotted here as a function of time for different
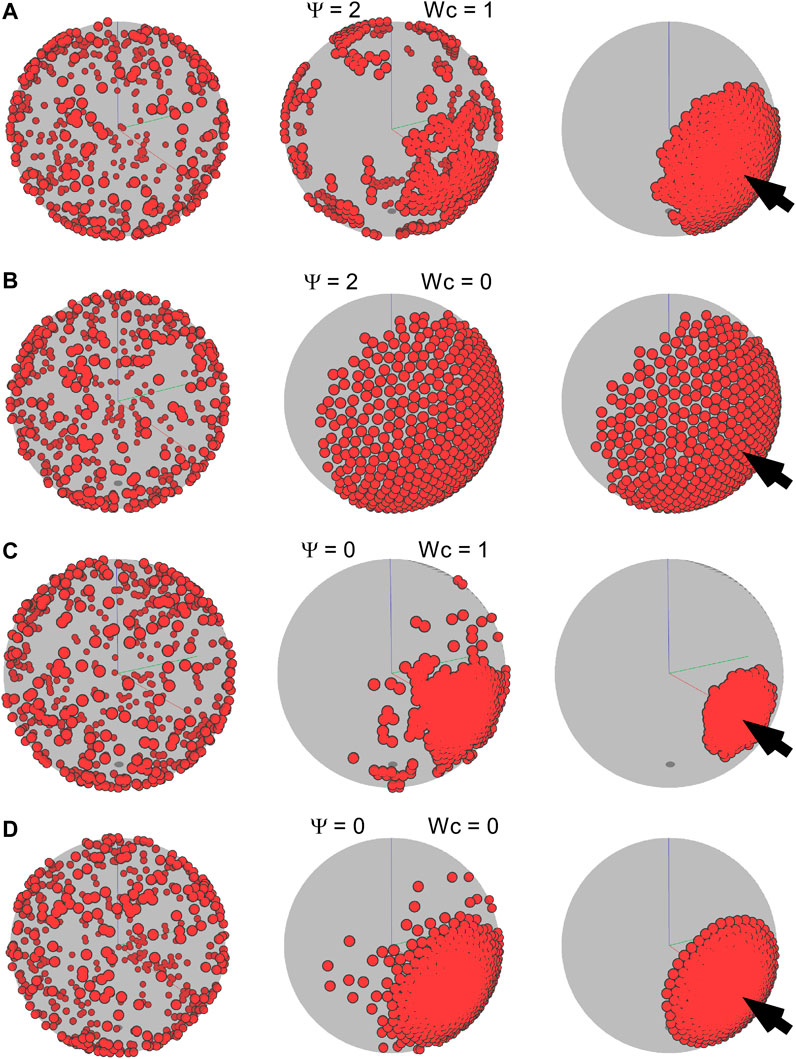
FIGURE 8. Frames of different time steps for an environmental guidance system (external taxis). For all simulations, ftax is 1 and
We have developed a computational model of cell migration and aggregation mechanisms for cells trapped between two concentric spheres, as is the case of the in vivo conditions of the annual killifish early embryo development. We demonstrate that numerical simulations of this model are an essential tool to test the role of physical mechanisms based on both the cell-autonomous and environmental guidance factors in driving the complex self-organizing processes during morphogenesis. Our computational model includes the full geometry of the embryo, including the spherical topology due to the constraint between the EVL and the YSL. This model was solved numerically to build phase space maps that reveal the effect of key mechanical parameters on the aggregation process.
For cell-autonomous mechanisms, the simulated aggregate phenotype was comparable to the experimental observation of the dispersed state, which could be achieved simply if the cells had low adhesion but failed to recapitulate the aggregation into a single cluster (Figures 3–6). At high adhesion, smaller clusters fused into a larger cluster, but the type of coalescence observed in vivo was never observed under these simple conditions. Thus, we explored the possibility that in addition to cell-autonomous factors, external cues possibly arising from a putative organizing center are also at play. The environmental taxis cues that guide cell migration could, in principle, be of many kinds, possibly chemical (i.e., diffusible or substrate-bound gradients of a chemoattractant) or physical (i.e., stiffness gradients). Using our model, we simulated a simple external cue guiding the direction of the migration of cells and showed that, under some conditions, it does indeed form single localized clusters (Figures 7, 8). In particular, our simulations determined that in order to achieve the dynamic and resulting phenotype of aggregation observed in early killifish development, an organizing center and low levels of CIL are essential. In addition, it appears that adhesive forces must remain low throughout the process and possibly increase only after the aggregate has formed.
Importantly, our strategy aims at building knowledge using a bottom-up approach by making predictions starting from the minimal condition for aggregation. Thus, at present, our model is kept to the most simplistic conditions possible in order to avoid overfitting of the system. However, more detailed models of cell motility or taxis, for example, may easily be constructed using our framework, given the relevant mechanistic details and more complete experimental evidence. Another aspect that might be considered is lateral inhibition, a mechanism that implicates the cell inhibition of adjacent cell activity. We are currently exploring all these mechanisms to simulate an aggregation process that is most like the experimental phenomena seen in annual killifish. Most importantly, despite its simplicity, our model is able to recapitulate the major features of cell aggregation in early annual killifish embryo development, a nearly unique model of cell aggregation in vertebrates.
The equations of motion (1) were solved using an implicit Euler method, with
so that we solve for
where
For each set of parameters, 50 replicates were made for cell-autonomous and 50 for external taxis models.
All codes and models were implemented using the open-source CellModeller multicellular modeling framework (Rudge et al., 2012).
The in vivo experiments depicted in Figure 1 were performed under the license for animal housing, breeding, and manipulation that was issued by Umwelt-und Verbraucherschutzamt der Stadt Köln, with the authorization no. 576.1.36.6.G28/13 Be. All the fish used were raised in 35-L tanks at 24°C–26°C and belonged to the N. furzeri FUCCI transgenic strain published by Dolfi et al. (2019). They were fed two to three times a day with frozen Chironomus larvae or living nauplii of Artemia salina, depending on their size. The breeders were kept in 8-L tanks with one or more boxes (9 cm × 9 cm × 4 cm) filled with 2 cm of river sand and were left to spawn eggs for 1 h. The embryos were collected by sieving the sand with a plastic net and were then embedded in 2% low melting agar. The fluorescence images shown in Figure 1 were acquired using a Leica TCS SP5-X confocal microscope and the red emission channel with 543-nm lasers. A total of 40 to 60 images per embryo were acquired at a depth distance of 6 μm. The maximum intensity projections shown have been generated using ImageJ. The contrast in the images has been adjusted, but not altered, to optimize visualization.
The raw data supporting the conclusion of this article will be made available by the authors upon request.
The animal study was reviewed and approved by Umwelt-und Verbraucherschutzamt der Stadt Köln. Authorization No. 576.1.36.6.G28/13 Be.
TR wrote the simulation software; IM-R, GY, MA-L, OG-C, and ES performed all the numerical simulations; GY, AR, and TR designed and performed the analyses; IM-R and NOR contributed with theoretical formulations; LD and AC performed and supervised the animal experiments; MC, MLC, and CB contributed with critical theoretical and analytical knowledge; AR and TR designed, directed, and supervised the project; GY, IM-R, MLC, AR, and TR wrote the manuscript. All authors contributed to commenting and writing the manuscript.
This work was funded by ANID SCIA/ACT192015, ANID FONDECYT Regular 1211598, 1210872, 1221696, 1230919, and 1190806; ANID FONDEQUIP EMQ210101, EQM210020, and EQM130051; FONDAP 15150012; ICN09_015; NCN19_170; REDES170212, and PUENTE-2022-13 from Pontificia Universidad Católica de Chile.
AR and TR are grateful to PUC/VRI, PUC IIBM, and the graduate program at IIBM for the seed funding and support. NOR is thankful for the support received from PUC IIBM.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2023.959611/full#supplementary-material
Abercrombie, M., and Heaysman, J. E. M. (1954). Observations on the social behaviour of cells in tissue culture. II. Monolayering of fibroblasts. Exp. Cell Res. 6, 293–306. doi:10.1016/0014-4827(54)90176-7
Abitua, P. B., Aksel, D. C., and Schier, A. F. (2021). Axis formation in annual killifish: Nodal coordinates morphogenesis in absence of Huluwa prepatterning. bioRxiv. doi:10.1101/2021.04.16.440199
Basan, M., Elgeti, J., Hannezo, E., Rappel, W. J., and Levine, H. (2013). Alignment of cellular motility forces with tissue flow as a mechanism for efficient wound healing. Proc. Natl. Acad. Sci. U. S. A. 110, 2452–2459. doi:10.1073/pnas.1219937110
Bellomo, N., Bellouquid, A., Tao, Y., and Winkler, M. (2015). Toward a mathematical theory of Keller–Segel models of pattern formation in biological tissues. Math. Models Methods Appl. Sci.25, 1663-1763. doi:10.1142/S021820251550044X
Belmonte, J. M., Thomas, G. L., Brunnet, L. G., de Almeida, R. M. C., and Chaté, H. (2008). Self-propelled particle model for cell-sorting phenomena. Phys. Rev. Lett. 100, 248702. doi:10.1103/PhysRevLett.100.248702
Bertocchi, C., Wang, Y., Ravasio, A., Hara, Y., Wu, Y., Sailov, T., et al. (2017). Nanoscale architecture of cadherin-based cell adhesions. Nat. Cell Biol. 19, 28–37. doi:10.1038/ncb3456
Bray, A. J. (1993). Theory of phase ordering kinetics. Phys. A Stat. Mech. its Appl. 194, 41–52. doi:10.1016/0378-4371(93)90338-5
Cai, D., Dai, W., Prasad, M., Luo, J., Gov, N. S., and Montell, D. J. (2016). Modeling and analysis of collective cell migration in an in vivo three-dimensional environment. Proc. Natl. Acad. Sci. 113. doi:10.1073/pnas.1522656113
Carter, S. B. (1967). Haptotaxis and the mechanism of cell motility. Nature 213, 256–260. doi:10.1038/213256a0
Concha, M. L., and Reig, G. (2022). Origin, form and function of extraembryonic structures in teleost fishes. Philos. Trans. R. Soc. Lond B Biol. Sci. 377, 20210264. doi:10.1098/rstb.2021.0264
Conte, V., Munoz, J., and Miodownik, M. (2008). A 3D finite element model of ventral furrow invagination in the Drosophila melanogaster embryo. J. Mech. Behav. Biomed. Mater. 1, 188–198. doi:10.1016/j.jmbbm.2007.10.002
Dolfi, L., Ripa, R., Antebi, A., Valenzano, D. R., and Cellerino, A. (2019). Cell cycle dynamics during diapause entry and exit in an annual killifish revealed by FUCCI technology. EvoDevo 10, 29. doi:10.1186/s13227-019-0142-5
Dolfi, L., Ripa, R., and Cellerino, A. (2014). Transition to annual life history coincides with reduction in cell cycle speed during early cleavage in three independent clades of annual killifish. EvoDevo 5, 32. doi:10.1186/2041-9139-5-32
Espina, J. A., Marchant, C. L., and Barriga, E. H. (2022). Durotaxis: The mechanical control of directed cell migration. FEBS J. 289, 2736–2754. doi:10.1111/febs.15862
Henkes, S., Fily, Y., and Marchetti, M. C. (2011). Active jamming: Self-propelled soft particles at high density. Phys. Rev. E 84, 040301. doi:10.1103/PhysRevE.84.040301
Kabla, A. J. (2012). Collective cell migration: Leadership, invasion and segregation. J. R. Soc. Interface 9, 3268–3278. doi:10.1098/rsif.2012.0448
Kanchanawong, P., Shtengel, G., Pasapera, A. M., Ramko, E. B., Davidson, M. W., Hess, H. F., et al. (2010). Nanoscale architecture of integrin-based cell adhesions. Nature 468, 580–584. doi:10.1038/nature09621
Márquez, S., Reig, G., Concha, M., and Soto, R. (2019). Cell migration driven by substrate deformation gradients. Phys. Biol. 16, 066001. doi:10.1088/1478-3975/ab39c7
Méhes, E., and Vicsek, T. (2014). Collective motion of cells: From experiments to models. Integr. Biol. 6, 831–854. doi:10.1039/c4ib00115j
Mladek, B. M., Gottwald, D., Kahl, G., Neumann, M., and Likos, C. N. (2006). formation of polymorphic cluster phases for a class of models of purely repulsive soft spheres. Phys. Rev. Lett. 96, 045701. doi:10.1103/PhysRevLett.96.045701
Moreno, A. J., and Likos, C. N. (2007). Diffusion and relaxation dynamics in cluster crystals. Phys. Rev. Lett. 99, 107801. doi:10.1103/PhysRevLett.99.107801
Pereiro, L., Loosli, F., Fernandez, J., Hartel, S., Wittbrodt, J., and Concha, M. L. (2017). Gastrulation in an annual killifish: Molecular and cellular events during germ layer formation in Austrolebias. Dev. Dyn. 246, 812–826. doi:10.1002/dvdy.24496
Phillips, R. (2015). Theory in biology: Figure 1 or figure 7? Trends Cell Biol. 25, 723–729. doi:10.1016/j.tcb.2015.10.007
Ravasio, A., Cheddadi, I., Chen, T., Pereira, T., Ong, H. T., Bertocchi, C., et al. (2015). Gap geometry dictates epithelial closure efficiency. Nat. Commun. 6, 7683–7713. doi:10.1038/ncomms8683
Ravasio, A., Le, A. P., Saw, T. B., Tarle, V., Ong, H. T., Bertocchi, C., et al. (2015). Regulation of epithelial cell organization by tuning cell-substrate adhesion. Integr. Biol. (United Kingdom) 7, 1228–1241. doi:10.1039/c5ib00196j
Redner, G. S., Baskaran, A., and Hagan, M. F. (2013). Reentrant phase behavior in active colloids with attraction. Phys. Rev. E 88, 012305. doi:10.1103/PhysRevE.88.012305
Reig, G., Cerda, M., Sepulveda, N., Flores, D., Castaneda, V., Tada, M., et al. (2017). Extra-embryonic tissue spreading directs early embryo morphogenesis in killifish. Nat. Commun. 8, 15431. doi:10.1038/ncomms15431
Roycroft, A., and Mayor, R. (2016). Molecular basis of contact inhibition of locomotion. Cell. Mol. Life Sci. 73, 1119–1130. doi:10.1007/s00018-015-2090-0
Rudge, T. J., Steiner, P. J., Phillips, A., and Haseloff, J. (2012). Computational modeling of synthetic microbial biofilms. ACS Synth. Biol. 1, 345–352. doi:10.1021/sb300031n
Sarris, M., and Sixt, M. (2015). Navigating in tissue mazes: Chemoattractant interpretation in complex environments. Curr. Opin. Cell Biol. 36, 93–102. doi:10.1016/j.ceb.2015.08.001
Sarris, M., and Sixt, M. (2015). Navigating in tissue mazes: Chemoattractant interpretation in complex environments. Curr. Opin. Cell Biol. 36, 93–102. doi:10.1016/j.ceb.2015.08.001
Sepúlveda, N., Petitjean, L., Cochet, O., Grasland-Mongrain, E., Silberzan, P., and Hakim, V. (2013). Collective cell motion in an epithelial sheet can Be quantitatively described by a stochastic interacting particle model. PLoS Comput. Biol. 9, e1002944. doi:10.1371/journal.pcbi.1002944
Smeets, B., Alert, R., Pešek, J., Pagonabarraga, I., Ramon, H., and Vincent, R. (2016). Emergent structures and dynamics of cell colonies by contact inhibition of locomotion. Proc. Natl. Acad. Sci. 113, 14621–14626. doi:10.1073/pnas.1521151113
Stepien, T. L., Lynch, H. E., Yancey, S. X., Dempsey, L., and Davidson, L. A. (2019). Using a continuum model to decipher the mechanics of embryonic tissue spreading from time-lapse image sequences: An approximate Bayesian computation approach. PLOS ONE 14, e0218021. doi:10.1371/journal.pone.0218021
Stichel, D., Middleton, A. M., Muller, B. F., Depner, S., Klingmuller, U., Breuhahn, K., et al. (2017). An individual-based model for collective cancer cell migration explains speed dynamics and phenotype variability in response to growth factors. npj Syst. Biol. Appl. 3, 5. doi:10.1038/s41540-017-0006-3
Tarle, V., Ravasio, A., Hakim, V., and Gov, N. S. (2015). Modeling the finger instability in an expanding cell monolayer. Integr. Biol. (Camb) 7, 1218–1227. doi:10.1039/c5ib00092k
Keywords: multicellularity, mechanics, biophysics, killifish, adhesion, modeling
Citation: Montenegro-Rojas I, Yañez G, Skog E, Guerrero-Calvo O, Andaur-Lobos M, Dolfi L, Cellerino A, Cerda M, Concha ML, Bertocchi C, Rojas NO, Ravasio A and Rudge TJ (2023) A computational framework for testing hypotheses of the minimal mechanical requirements for cell aggregation using early annual killifish embryogenesis as a model. Front. Cell Dev. Biol. 11:959611. doi: 10.3389/fcell.2023.959611
Received: 01 June 2022; Accepted: 08 February 2023;
Published: 20 March 2023.
Edited by:
Bojana Gligorijevic, Temple University, United StatesReviewed by:
Stuart A. Newman, New York Medical College, United StatesCopyright © 2023 Montenegro-Rojas, Yañez, Skog, Guerrero-Calvo, Andaur-Lobos, Dolfi, Cellerino, Cerda, Concha, Bertocchi, Rojas, Ravasio and Rudge. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Timothy J. Rudge, dGltLnJ1ZGdlQG5ld2Nhc3RsZS5hYy51aw==; Andrea Ravasio, YW5kcmVhLnJhdmFzaW9AdWMuY2w=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.