 Tao Su
Tao Su Jiandong Liu3†
Jiandong Liu3† Nasha Zhang
Nasha Zhang Ming Yang
Ming Yang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol., 19 June 2023
Sec. Cancer Cell Biology
Volume 11 - 2023 | https://doi.org/10.3389/fcell.2023.1157797
N6-Methyladenosine (m6A) methylation is one of the most extremely examined RNA modifications. M6A modification evidently impacts cancer development by effecting RNA metabolism. Long non-coding RNAs (lncRNAs) and microRNAs (miRNAs) are involved in multiple essential biological processes by regulating gene expression at the transcriptional and post-transcriptional levels. Accumulated evidences indicated that m6A is involved in regulating the cleavage, stability, structure, transcription, and transport of lncRNAs or miRNAs. Additionally, ncRNAs also play significant roles in modulating m6A levels of malignant cells by participating in the regulation of m6A methyltransferases, the m6A demethylases and the m6A binding proteins. In this review, we systematically summarize the new insight on the interactions between m6A and lncRNAs or miRNAs, as well as their impacts on gastrointestinal cancer progression. Although there are still extensive studies on genome-wide screening of crucial lncRNAs or miRNAs involved in regulating m6A levels of mRNAs and disclosing differences on mechanisms of regulating m6A modification of lncRNAs, miRNAs or mRNAs in cancer cells, we believe that targeting m6A-related lncRNAs and miRNAs may provide novel options for gastrointestinal cancer treatments.
RNA modifications play pivotal roles in regulating stability and functions of various RNAs. RNA methylation is one of the most extensively and diligently studied RNA modifications. Previous reports have shown that N6-methyladenosine (m6A) modification affects tumor development and progression in a variety of ways, such as influencing cell proliferation, promoting of cancer stem cell regeneration, and leading to resistance of malignant cells to radiotherapy or chemotherapy (Lan et al., 2019a). Another abundant mRNA modification is N6,2-O-dimethyladenosine (m6A.m.), which occurs near the mRNA cap (Wei et al., 1975). M6A.m. negatively impacts cap-dependent translation of methylated mRNAs (Sendinc et al., 2019). In addition, m6A.m. has been identified as an internal modification and a crucial regulatory factor in snoRNA (Chen et al., 2020; Goh et al., 2020; Oerum et al., 2021). However, m6A is the main RNA modification in long non-coding RNAs (lncRNAs) and microRNAs (miRNAs) (Oerum et al., 2021). In this review, we mainly summarize the correlation between m6A modification and lncRNA/miRNA in gastrointestinal cancers.
M6A modification is a reversible process, which requires the participation of the m6A methyltransferases (writers), the m6A demethylases (erasers) and the m6A binding proteins (readers) (Fu et al., 2014). The main functions of the m6A methyltransferases, such as WTAP, METTL3, and METTL14, are to initiate methylation of adenosines in RNAs. The m6A demethylases, such as FTO and ALKBH5, are to recognize and delete m6A modification. The m6A binding proteins including YTHDC1/2, YTHDF1/2/3 and IGF2BP1/2/3, can recognize methylated base pairs and impact cellular functions including mRNA translation, RNA metabolism, as well as interactions between microRNA (miRNAs) and their target RNAs (Nettersheim et al., 2019). Divergent cell-type-specific and tissue-specific expression and localization of these proteins results in significant differences in the level of m6A modification in different cell types (Shi et al., 2019). In addition, environmental factors also play important regulatory roles in this process (Shi et al., 2019).
M6A modification is considered to be a critical posttranscriptional regulator of gene expression. M6A modifications of messenger RNA (mRNA) regulate various aspects of mRNA metabolism (Frye et al., 2018). Non-coding RNAs (ncRNAs), such as miRNAs and lncRNAs, have crucial biological functions in the tumorigenesis and progression of various types of cancers (Jacob et al., 2017). It has been found that m6A modifications on miRNAs and lncRNAs can also regulate their production, metabolism, and functions (Han et al., 2019). Interestingly, miRNAs and lncRNAs were involved in regulating m6A modifications as well (Cai et al., 2018). In several pre-clinical studies, targeting m6A and miRNAs or lncRNAs in cancer cells simultaneously showed synergistic effects (Wahlestedt, 2013; Zhang et al., 2016). In addition, m6A modification can regulate chromatin state via lncRNAs (Wei and He, 2021). For example, in mouse embryonic stem cells (mESCs), FTO mediates m6A demethylation of LINE1 regulating LINE1 RNA abundance and the local chromatin accessibility (Wei et al., 2022). In this review, we systematically summarized the interplays between m6A modification and miRNAs/lncRNAs, and described the significance how they impact tumorigenesis in gastrointestinal cancers.
LncRNAs are a class of ncRNA molecules with more than 200 nucleotides (nt). Compared with other types of ncRNAs, lncRNAs have more family numbers and more complicated functions (Schmitt and Chang, 2016). In different gastrointestinal cancers, lncRNAs regulate expression of protein-coding genes via distinct mechanisms (Figure 1). There are plenty of m6A modifications in lncRNAs, of which show temporal and spatial specificities (Xiao et al., 2019). M6A modification can regulate expression and functions of lncRNAs. Proteins involved in m6A modifications can impact m6A levels of lncRNAs and stability of lncRNAs, which eventually regulates various biological functions of malignant cells. Moreover, m6A modification can initiate RNA-protein binding via providing binding sites for m6A reader protein or adjusting RNA local structures (Quinn and Chang, 2016), which might also impact lncRNA functions (Table 1).
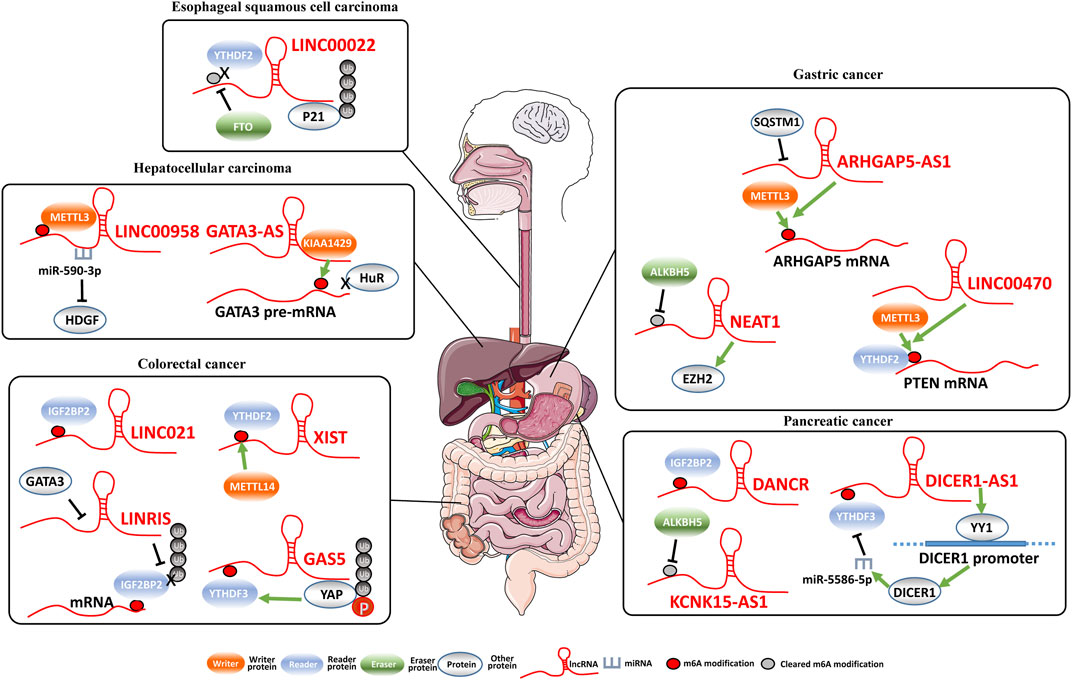
FIGURE 1. Regulation of lncRNAs by m6A modification and lncRNA-regulated m6A modification..
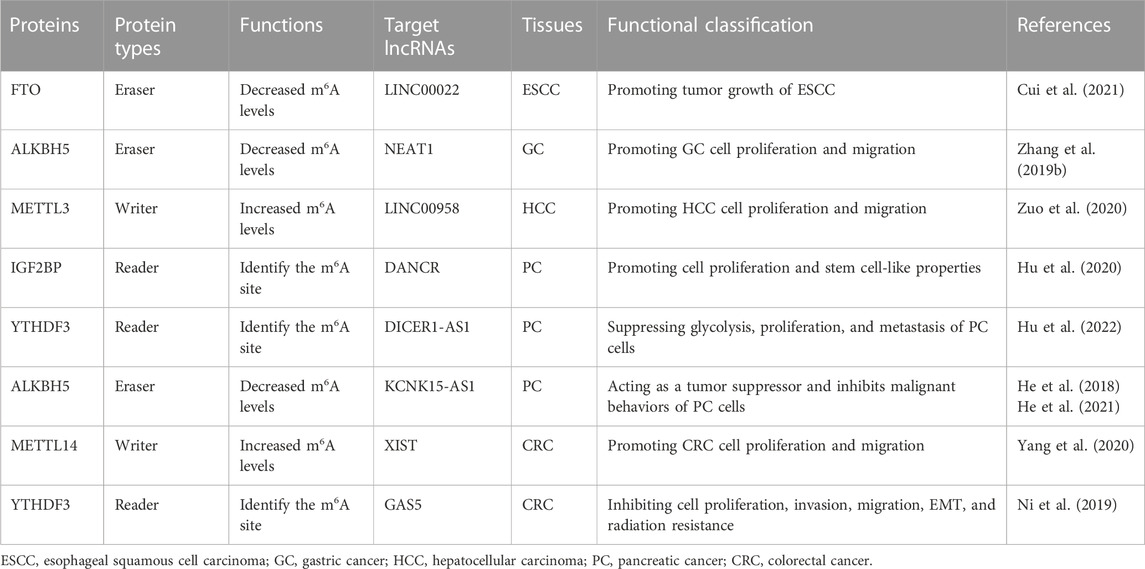
TABLE 1. M6A modification regulates lncRNA stability.
Esophageal squamous cell carcinoma (ESCC) is one major pathological subtype of esophageal cancer and one of the most lethal cancers worldwide. In East Asia including China, ESCC accounts for more than 90% of all esophageal cancer patients. However, how m6A impacts lncRNA functions in ESCC remains largely unexplored. Cui et al. (2021) found that LINC00022, which is demethylated by the m6A eraser FTO, promotes tumor growth of ESCC. LINC00022 is upregulated in primary ESCC samples and associated with poor outcomes for ESCC patients. In ESCC cells, LINC00022 interacts with P21 protein and enhances ubiquitination-mediated degradation of P21. The m6A eraser FTO reduces m6A methylation levels of LINC00022 and results in inhibited LINC00022 decay via the m6A reader YTHDF2. Therefore, over-expressed FTO drives LINC00022-dependent ESCC cell proliferation in vitro and in vivo.
As one of the most common digestive system cancers, gastric cancer (GC) is the leading cause of cancer-related death in the world. LncRNA NEAT1 is overexpressed in GC tissues compared to normal stomach tissues. Consistently, NEAT1 could significantly promotes invasion and metastasis capabilities of GC cells. Interestingly, lncRNA NEAT1 is a target RNA of the m6A demethylase ALKBH5. ALKBH5 eliminated the m6A modification of NEAT1 and, thus, influences the expression of EZH2 as well as GC progression (Zhang et al., 2019b).
In hepatocellular carcinoma (HCC), METTL3-mediated m6A modification stabilizes lncRNA LINC00958 to promote HCC progression. METTL3 increases m6A modification levels of LINC00958 and inhibits lncRNA degradation. High levels of LINC00958 promotes lipogenesis in HCC cells. Interestingly, LINC00958 upregulates hepatoma-derived growth factor (HDGF) expression through sponging miR-3619-5p (Zuo et al., 2020). As a result, the interaction between METTL3 and lipogenesis-related lncRNA LINC00958 enhances proliferation and metastasis capability of HCC cells.
Pancreatic cancer (PC) is the seventh leading cause of cancer death worldwide, with a 5-year survival rate of approximately 10% of patients (Hu et al., 2020). IGF2BPs family, as m6A readers, recognized the consensus GG (m6A)C motif of thousands of transcripts with m6A modification including lncRNAs (Huang et al., 2018). Among them, the RNA-binding protein IGF2BP2 play important roles in RNA localization, stability, and metabolism as a posttranscriptional regulatory factor. To investigate the clinical importance of IGF2BP2 in PC, Hu et al. (2020) examined IGF2BP2 expression through immunohistochemistry staining in the PC tissue microarrays containing 82 cancerous samples and 54 normal specimens. It has been found that evaluated expression of IGF2BP2 is significantly associated with poor outcomes of PC patients and suppression of IGF2BP2 inhibits cell proliferation. Indeed, IGF2BP2 can interact with lncRNA DANCR and enhances DANCR expression in PC cells. LncRNA DANCR has been reported to markedly increase stemness features of malignant cells to promote tumorigenesis in HCC, osteosarcoma and acute myeloid leukemia (Yuan et al., 2016; Jiang et al., 2017; Bill et al., 2019; Nance et al., 2020). Consistently, the high expression of lncRNA DANCR also accelerates PC cell proliferation and differentiation of cancer stem cells (Hu et al., 2020). LncRNA DANCR is modified with m6A in PC. IGF2BP2 serves as the reader for the m6A-modified DANCR and stabilizes DANCR RNA.
In PC, DICER1-AS1 is another m6A modified lncRNA which plays a pivotal role in glycolysis of PC cells (Hu et al., 2022). Overexpressed lncRNA DICER1-AS1 suppresses glycolysis, proliferation, and metastasis of PC cells via transcriptionally enhancing DICER1 expression. DICER1 enhances miR-5586-5p maturation and subsequently represses expression of multiple glycolytic genes. The m6A reader YTHDF3 recognizes the m6A-modified DICER1-AS1 and results in lncRNA DICER1-AS1 degradation in response to glucose depletion. Additionally, it has been found that the m6A reader YTHDF3 is a target gene of miR-5586-5p. Together, these results suggest that there is negative feedback with YTHDF3/DICER1-AS1/DICER1/miR-5586-5p to regulate PC glycolysis and pathogenesis.
In PC cells, KCNK15-AS1 is a target lncRNA of ALKBH5 (He et al., 2018; He et al., 2021). KCNK15-AS1 is significantly downregulated in PC tissues. Consistently, lncRNA KCNK15-AS1 acts as a tumor suppressor and inhibits malignant behaviors of PC cells. The m6A eraser ALKBH5, which was downregulated in PC cells, demethylates KCNK15-AS1 and regulates KCNK15-AS1-mediated cell motility (He et al., 2018). Moreover, He et al. (2021) found that lncRNA KCNK15-AS1 could bind with KCNK15 mRNA 5′-UTR to inhibit KCNK15 translation. Moreover, lncRNA KCNK15-AS1 recruits the E3 ligase MDM2 to REST protein, promotes REST ubiquitination and, thus, transcriptionally upregulates tumor suppressor PTEN expression to inactivate the AKT signaling (He et al., 2021).
Colorectal cancer (CRC) is one of the leading cancer types, ranking third for incidence, but second for mortality, and the incidence rate is rising worldwide. In CRC, the reduced levels of METTL14 expression were also markedly correlated with unfavorable prognosis of patients (Yang et al., 2020). Consistently, METTL14 could repress proliferative and invasive capabilities of CRC cells in vitro and in vivo. LncRNA XIST is a target of METTL14 in CRC. Knockdown of METTL14 substantially decreases oncogenic lncRNA XIST m6A modification levels and upregulates XIST expression. YTHDF2 acts as the m6A reader protein of XIST and mediates degradation of m6A-modifed XIST. In line with this, there was significantly negative expression correlations between lncRNA XIST and METTL14 or YTHDF2 in CRC tissues. These results highlighted the function and prognostic values of m6A modification and lncRNA XIST in CRC.
YAP activation is crucial for CRC tumorigenesis and progression (Johnson and Halder, 2014). Ni et al. identified GAS5 as a YAP-interacting lncRNA via analyzing the RIP-seq profiles (Ni et al., 2019). In CRC tissues, the expression levels of lncRNA GAS5 are negatively correlated with YAP. GAS5 directly interacts with the WW domain of YAP, thereby promoting endogenous YAP transport from the nucleus to the cytoplasm. In line with this, lncRNA GAS5 promotes YAP phosphorylation and subsequently ubiquitin-mediated YAP degradation, and thus inhibiting the CRC progression in vitro and in vivo. As one of the m6A “readers”, YTHDF3 collaborates with YTHDF1 to promote translation of protein synthesis and influence decay of methylated mRNA mediated through YTHDF2 (Shi et al., 2017; Ni et al., 2019). Notably, YTHDF3 recognizes the m6A modification site of lncRNA GAS5 and promotes its degradation, while GAS5 can repress YAP-mediated YTHDF3 transcriptional expression. In all, the study proposed a negative feedback regulation of lncRNA GAS5-YAP-YTHDF3, and established a new mechanism for m6A-induced degradation of lncRNA (Ni et al., 2019).
As described previously, the m6A modification of lncRNAs could impact expression and metabolism of lncRNAs. Interestingly, lncRNAs are also involved in regulation of expression of m6A-related proteins as well as functions of m6A-modified mRNAs at the post-transcriptional level in various cancers (Table 2; Figure 1).

TABLE 2. LncRNA-regulated m6A modification.
LncRNAs could impact the stability of their target RNAs by recruiting m6A writer proteins. For instance, lncRNA ARHGAP5-AS1 is highly expressed in chemoresistant GC cells and silencing ARHGAP5-AS1 reversed chemoresistance (Zhu et al., 2019). LncRNA ARHGAP5-AS1 can stimulate transcription of ARHGAP5 in the nucleus of GC cells, whereas stimulate m6A modification of ARHGAP5 mRNA to stabilize it through recruiting the m6A writer METTL3 in the cytoplasm. Interestingly, SQSTM1 can translocate lncRNA ARHGAP5-AS1 to autophagosomes to accelerate its degradation in cells. Therefore, the ARHGAP5-AS1/ARHGAP5 axis might be a possible target to reverse chemoresistance of GC cells.
LINC00470 is an oncogenic lncRNA in GC, with evidently upregulated expression in GC tissues and cell lines (Yan et al., 2020). It has been found that LINC00470 levels are significantly associated with metastasis, advanced TNM stages, and cancer progression of GC patients. LINC00470 could enhance proliferation, migration, and invasion of GC cells. Mechanistically, LINC0047 recruits the m6A writer METTL3 to increases m6A modification on PTEN mRNA, thereby decreasing its stability. As a result, LINC00470 inhibits its target gene PTEN expression and promotes tumorigenesis of GC cells.
LncRNA GATA3-AS, the antisense RNA of GATA3 gene, acts as an oncogene and accelerates metastasis in HCC (Lan et al., 2019b). Indeed, lncRNA GATA3-AS enhances the interaction of KIAA1429, a component of the m6A methyltransferase complex, with GATA3 pre-mRNA as the molecule scaffold. As a result, KIAA1429 induces GATA3 pre-mRNA methylation on the 3′-UTR, leading the isolation of RNA-binding protein HuR and degradation of GATA3 pre-mRNA in HCC cells.
LINRIS is upregulated in human CRC samples compared with their matched adjacent normal tissues. LncRNA LINRIS promotes disease progression by accelerating glycolysis of CRC cells (Wang et al., 2019). LINRIS inhibits K139 ubiquitination of IGF2BP2 protein and prevents the degradation of IGF2BP2 via the autophagy-lysosome pathway. IGF2BP2 can recognize m6A-modified mRNAs, such as ELAVL1, HUR, MATR3, and PABPC1, and maintain their stability. Interestingly, knockdown of LINRIS lessens the MYC-mediated glycolysis, which is regulated by IGF2BP2 in CRC cells. In summary, LINCRIS regulates the ubiquitination of the crucial m6A reader protein IGF2BP2 and the LINRIS-IGF2BP2-MYC axis might be a promising therapeutic target of CRC.
In CRC, LINC021 is remarkably upregulated in CRC tissues compared to normal tissues (Wu et al., 2022). Levels of LINC021 might be an independent prognostic factor of CRC patients. LINC021 functions as an oncogene to promote CRC malignant proliferation, colony formation, migration capabilities, and reduced cell apoptosis. Specifically, oncogenic LINC021 specifically interacts with IGF2BP2 to enhance the mRNA stability of MSX1 and JARID2. These findings demonstrate the important roles of LINC021/IGF2BP2/MSX1 axis in CRC tumorigenesis and shed light for CRC treatment.
MiRNAs are crucial regulatory small ncRNAs involved in tumorigenesis. M6A modification may impact synthesis and functions of miRNAs at various levels (Table 3; Figure 2). On one hand, m6A-related proteins regulate the expression of key microprocessor proteins in miRNA biosynthesis, such as Drosha and DGCR8, to affect miRNA expression. On the other hand, m6A-related proteins are also target genes of miRNAs in cancers (Burke et al., 2014; Alarcon et al., 2015; Catalanotto et al., 2016).
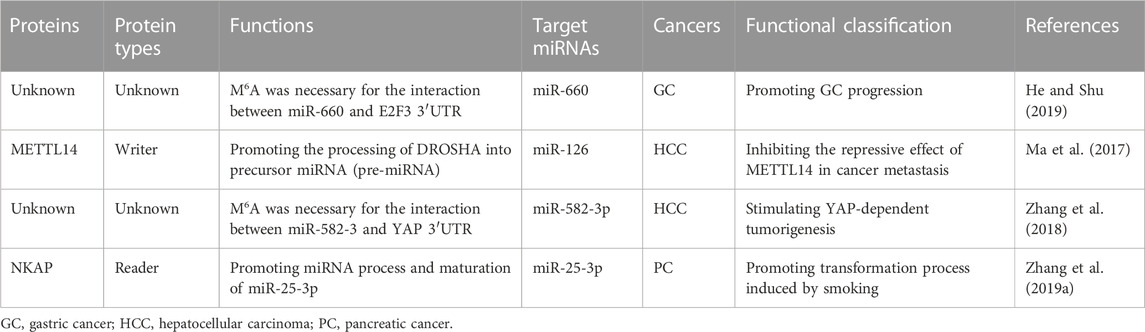
TABLE 3. M6A modification regulates miRNA.
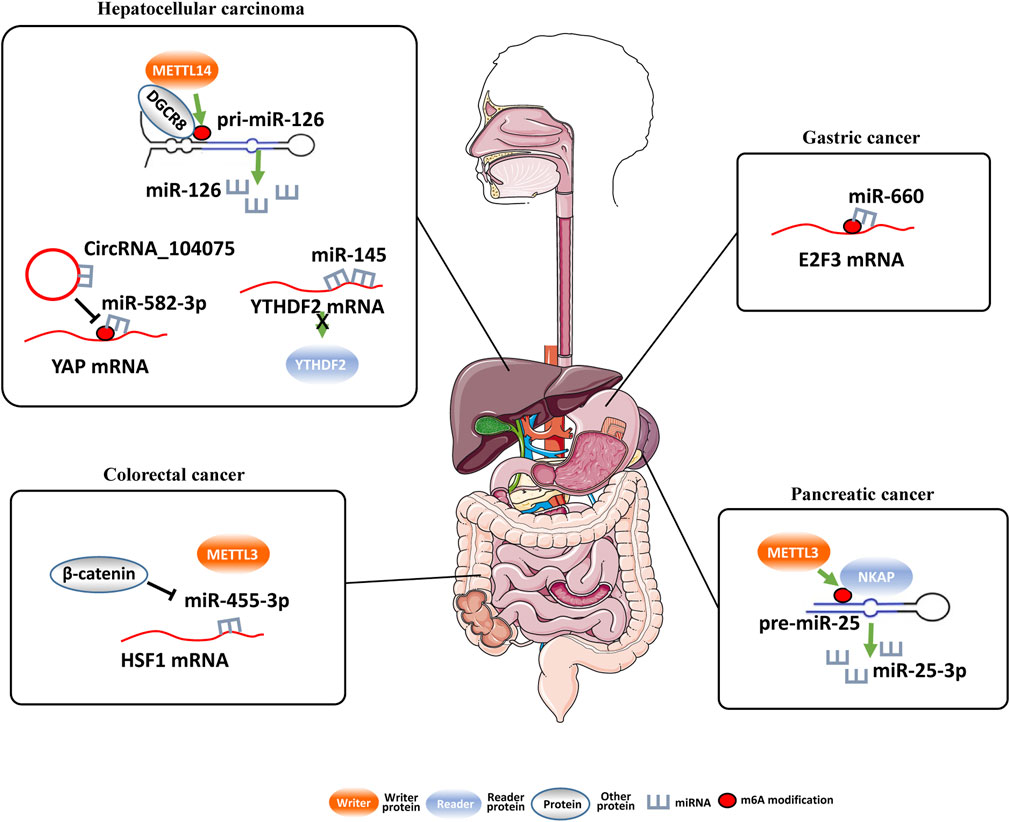
FIGURE 2. Regulation of miRNAs by m6A modification and miRNAs regulate m6A modification..
MiR-660 is significantly downregulated in GC tissues and associated with advanced TNM stage, larger tumor size, lymph node metastasis, and poor prognosis of GC patients (He and Shu, 2019). In line with these, miR-660 significantly repressed proliferation of GC cells and induced apoptosis of GC cells. In GC cells, miR-660 reduces expression of E2F3 via directly binding to E2F3 3′- UTR. It is worth noting that the m6A motif was found in E2F3 3′-UTR and the m6A site is necessary for the interaction between miR-660 and E2F3 mRNA. These findings further elucidate the crucial role of m6A in GC and highlights the regulatory functions of the miR-660/E2F3 pathway during GC progression.
The m6A modification is reduced in HCC and METTL14 is the main factor involved in abnormal m6A modification (Scholler et al., 2018). As miRNAs play important roles in tumorigenesis and metastasis, Ma et al. (2017) hypothesized that METTL14 might impact HCC progression by regulating miRNAs in an m6A-dependent, pri-miRNA-processing manner. Subsequent immunoprecipitation assays demonstrated that METTL14 directly interacted with the microprocessor protein DGCR8. Moreover, the interaction between METTL14 and DGCR8 was suppressed after ribonuclease treatment, elucidating that there were RNAs participating in their interaction. Interestingly, mature miR126 and METTL14 had the similarity expression trend in HCC cells. Consistently, unprocessed pri-miR126 accumulated upon METTL14-depleted cells and accelerated in METTL14-overexpressed cells. The overexpression of METTL14 increased the binding between pri-miR126 and DGCR8. These findings suggest that METTL14 regulated the recognition and binding of DGCR8 to pri-miRNAs to impact pri-miRNA processing in HCC.
Oncogenic circ_104,075 was evidently upregulated in HCC tissues, serum, and cell lines (Zhang et al., 2018). In HCC cells, circ_104,075 absorbed miR-582-3p as a molecular sponge to increase YAP expression. There is an m6A motif in the 353-357 nt region of YAP 3′-UTR, which is essential for the interaction between miR-582-3p and YAP 3′-UTR. Interestingly, circRNA_104,075 could be used for prediction of the occurrence of HCC, with the AUC-ROC of 0.973.
Cigarette smoke condensate enhanced m6A modification of oncogenic miR-25 to promote its maturation in pancreatic duct epithelial cells (Zhang et al., 2019a). MiR-25-3p are upregulated in smokers and in PC tissues, which are significantly assciated with poor prognosis of PC patients. NKAP (NF-κB related protein) is a novel m6A reader and recognizes the m6A-modified miR-25-3p precursor RNA in pancreatic cancer, leading to accelerated miRNA process and maturation of miR-25-3p and increased expression of miR-25-3p. miR-25-3p suppresses the expression of PHLPP2 and regulates the AKT-p70S6K signaling pathway. These results indicate that important roles of METTL3/miR-25-3p/PHLPP2/AKT axis during the transformation process induced by smoking in PC patients.
The m6A modification-related proteins play important roles in various cancers. Interestingly, dysregulated miRNAs can also regulate expression of these m6A modification-related genes and the m6A levels in cells, which may lead to tumorigenesis (Cai et al., 2018) (Table 4; Figure 2).

TABLE 4. MiRNA regulates m6A modification.
Yang et al. investigated the regulatory mechanism of YTHDF2, which was poorly understood in HCC (Yang et al., 2017). It has been found that miR-145 modifies m6A modification through suppressing YTHDF2 expression in HCC. On the contrary, inhibition of miR-145 upregulates YTHDF2 expression levels in HCC cells. In line with these data, the expression levels of miR-145 in HCC tissues was negatively correlated with the level of YTHDF2 mRNA. Additionally, inhibition of miR-145 evidently decreased m6A levels, which were rescued by silencing of YTHDF2 expression in cells. In summary, the miR-145/YTHDF2 axis plays a vital role in modulating m6A levels of HCC cells.
In colorectal cancer cells, miR-455-3p can interact with 3′-UTR of HSF1 mRNA to repress its translation (Song et al., 2020). The inhibition of WNT/β-catenin signaling by pyrvinium or β-catenin knockdown impaired HSF1 mRNA translation and its m6A modification. Importantly, inhibition of miR-455-3p can rescue the reduction of HSF1 m6A modification and METTL3 interaction caused by β-catenin depletion. β-catenin suppressed the biogenesis of miR-455-3p, which interrupt the interaction between METTL3 and HSF1 mRNA, thus promoting HSF1 m6A modification.
It has been found that m6A modification levels of lncRNAs or miRNAs might impact RNA stability. Conversely, lncRNAs and miRNAs are also involved in regulating m6A levels of mRNAs in cells. These m6A-related lncRNAs and miRNAs functions as novel oncogenes or rumor suppressors in malignant cells (Table 5). However, there are still no studies on genome-wide screening of crucial lncRNAs or miRNAs involved in regulating m6A levels of mRNAs in cancers. Moreover, it is still largely unclear if there are differences on mechanisms of regulating m6A modification of lncRNAs, miRNAs or mRNAs in different gastrointestinal cancer cells. In summary, there are still considerable studies on unveiling the regulatory relationships between lncRNAs/miRNAs and m6A modification. Though, we are certain of targeting m6A-related lncRNAs and miRNAs may provide novel options for gastrointestinal cancer treatments.
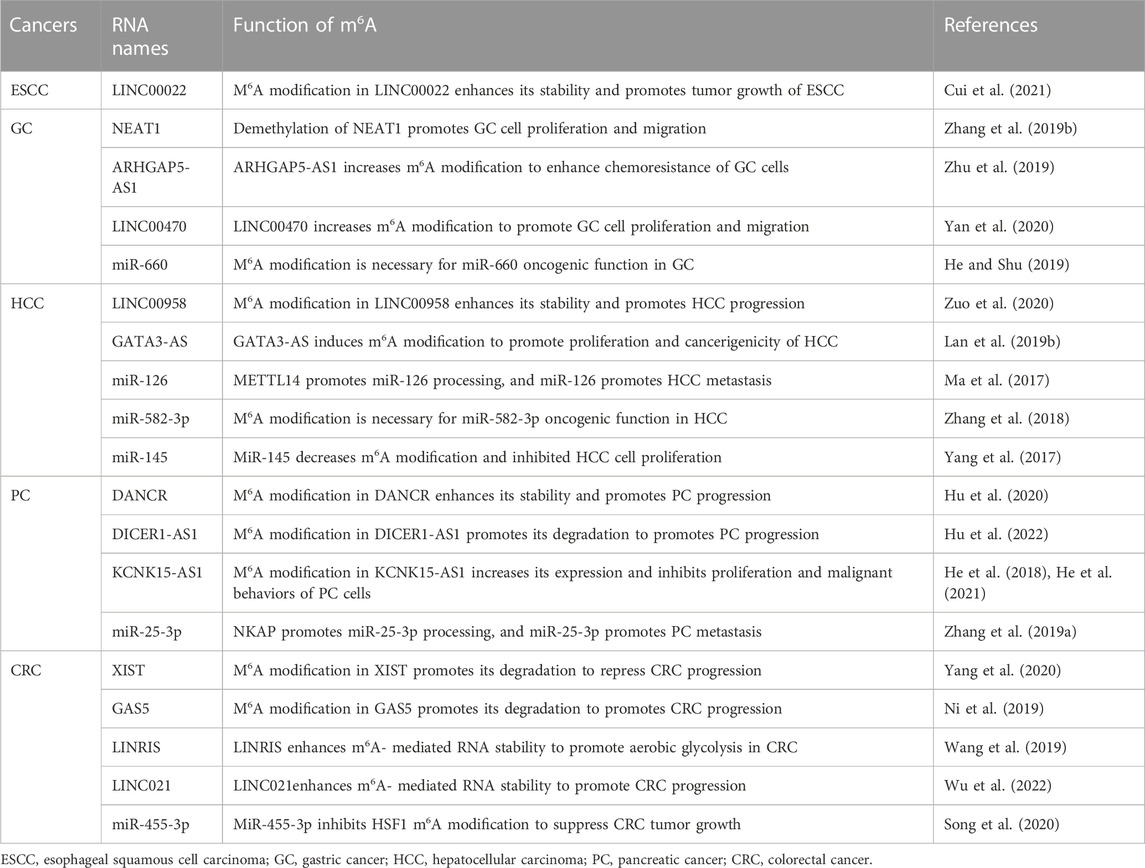
TABLE 5. Function of lncRNA and miRNA m6A modification in gastrointestinal cancers.
TS, JL, SW, and MY wrote sections of the original draft. All other authors provided input during early discussions and drafts. All authors contributed to the article and approved the submitted version.
This study was financially supported by the National Natural Science Foundation of China (82173070 and 82103291); Natural Science Foundation of Shandong Province (ZR2021LZL004 and ZR202102250889); Major Scientific and Technological Innovation Project of Shandong Province (2021ZDSYS04); Taishan Scholars Program of Shandong Province (tsqn202211340 and tstp20221141); Program of Science and Technology for the youth innovation team in universities of Shandong Province (2020KJL001 and 2022KJ316).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2023.1157797/full#supplementary-material
m6A, N6-Methyladenosine; lncRNAs, long non-coding RNAs; miRNAs, microRNAs; ncRNAs, non-coding RNAs; ceRNA, competing endogenous RNA; ESCC, esophageal squamous cell carcinoma: GC, gastric cancer; HCC, hepatocellular carcinoma; HDGF, hepatoma-derived growth factor; PC, pancreatic cancer; CRC, colorectal cancer.
Alarcon, C. R., Lee, H., Goodarzi, H., Halberg, N., and Tavazoie, S. F. (2015). N6-methyladenosine marks primary microRNAs for processing. Nature 519, 482–485. doi:10.1038/nature14281
Bill, M., Papaioannou, D., Karunasiri, M., Kohlschmidt, J., Pepe, F., Walker, C. J., et al. (2019). Expression and functional relevance of long non-coding RNAs in acute myeloid leukemia stem cells. Leukemia 33, 2169–2182. doi:10.1038/s41375-019-0429-5
Burke, J. M., Kelenis, D. P., Kincaid, R. P., and Sullivan, C. S. (2014). A central role for the primary microRNA stem in guiding the position and efficiency of Drosha processing of a viral pri-miRNA. RNA 20, 1068–1077. doi:10.1261/rna.044537.114
Cai, X., Wang, X., Cao, C., Gao, Y., Zhang, S., Yang, Z., et al. (2018). HBXIP-elevated methyltransferase METTL3 promotes the progression of breast cancer via inhibiting tumor suppressor let-7g. Cancer Lett. 415, 11–19. doi:10.1016/j.canlet.2017.11.018
Catalanotto, C., Cogoni, C., and Zardo, G. (2016). MicroRNA in control of gene expression: An overview of nuclear functions. Int. J. Mol. Sci. 17, 1712. doi:10.3390/ijms17101712
Chen, H., Gu, L., Orellana, E. A., Wang, Y., Guo, J., Liu, Q., et al. (2020). METTL4 is an snRNA m(6)Am methyltransferase that regulates RNA splicing. Cell Res. 30, 544–547. doi:10.1038/s41422-019-0270-4
Cui, Y., Zhang, C., Ma, S., Li, Z., Wang, W., Li, Y., et al. (2021). RNA m6A demethylase FTO-mediated epigenetic up-regulation of LINC00022 promotes tumorigenesis in esophageal squamous cell carcinoma. J. Exp. Clin. Cancer Res. 40, 294. doi:10.1186/s13046-021-02096-1
Frye, M., Harada, B. T., Behm, M., and He, C. (2018). RNA modifications modulate gene expression during development. Science 361, 1346–1349. doi:10.1126/science.aau1646
Fu, Y., Dominissini, D., Rechavi, G., and He, C. (2014). Gene expression regulation mediated through reversible m⁶A RNA methylation. Nat. Rev. Genet. 15, 293–306. doi:10.1038/nrg3724
Goh, Y. T., Koh, C. W. Q., Sim, D. Y., Roca, X., and Goh, W. S. S. (2020). METTL4 catalyzes m6Am methylation in U2 snRNA to regulate pre-mRNA splicing. Nucleic Acids Res. 48, 9250–9261. doi:10.1093/nar/gkaa684
Han, Y., Feng, J., Xia, L., Dong, X., Zhang, X., Zhang, S., et al. (2019)., 8. Cells, 168. CVm6A: A visualization and exploration database for m⁶As in cell linesCells doi:10.3390/cells8020168
He, X., and Shu, Y. (2019). RNA N6-methyladenosine modification participates in miR-660/E2F3 axis-mediated inhibition of cell proliferation in gastric cancer. Pathol. Res. Pract. 215, 152393. doi:10.1016/j.prp.2019.03.021
He, Y., Hu, H., Wang, Y., Yuan, H., Lu, Z., Wu, P., et al. (2018). ALKBH5 inhibits pancreatic cancer motility by decreasing long non-coding RNA KCNK15-AS1 methylation. Cell Physiol. Biochem. 48, 838–846. doi:10.1159/000491915
He, Y., Yue, H., Cheng, Y., Ding, Z., Xu, Z., Lv, C., et al. (2021). ALKBH5-mediated m(6)A demethylation of KCNK15-AS1 inhibits pancreatic cancer progression via regulating KCNK15 and PTEN/AKT signaling. Cell Death Dis. 12, 1121. doi:10.1038/s41419-021-04401-4
Hu, X., Peng, W. X., Zhou, H., Jiang, J., Zhou, X., Huang, D., et al. (2020). IGF2BP2 regulates DANCR by serving as an N6-methyladenosine reader. Cell Death Differ. 27, 1782–1794. doi:10.1038/s41418-019-0461-z
Hu, Y., Tang, J., Xu, F., Chen, J., Zeng, Z., Han, S., et al. (2022). A reciprocal feedback between N6-methyladenosine reader YTHDF3 and lncRNA DICER1-AS1 promotes glycolysis of pancreatic cancer through inhibiting maturation of miR-5586-5p. J. Exp. Clin. Cancer Res. 41, 69. doi:10.1186/s13046-022-02285-6
Huang, H., Weng, H., Sun, W., Qin, X., Shi, H., Wu, H., et al. (2018). Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 20, 285–295. doi:10.1038/s41556-018-0045-z
Jacob, R., Zander, S., and Gutschner, T. (2017). The dark side of the epitranscriptome: Chemical modifications in long non-coding RNAs. Int. J. Mol. Sci. 18, 2387. doi:10.3390/ijms18112387
Jiang, N., Wang, X., Xie, X., Liao, Y., Liu, N., Liu, J., et al. (2017). lncRNA DANCR promotes tumor progression and cancer stemness features in osteosarcoma by upregulating AXL via miR-33a-5p inhibition. Cancer Lett. 405, 46–55. doi:10.1016/j.canlet.2017.06.009
Johnson, R., and Halder, G. (2014). The two faces of hippo: Targeting the hippo pathway for regenerative medicine and cancer treatment. Nat. Rev. Drug Discov. 13, 63–79. doi:10.1038/nrd4161
Lan, Q., Liu, P. Y., Haase, J., Bell, J. L., Huttelmaier, S., and Liu, T. (2019a). The critical role of RNA m(6)A methylation in cancer. Cancer Res. 79, 1285–1292. doi:10.1158/0008-5472.CAN-18-2965
Lan, T., Li, H., Zhang, D., Xu, L., Liu, H., Hao, X., et al. (2019b). KIAA1429 contributes to liver cancer progression through N6-methyladenosine-dependent post-transcriptional modification of GATA3. Mol. Cancer 18, 186. doi:10.1186/s12943-019-1106-z
Ma, J. Z., Yang, F., Zhou, C. C., Liu, F., Yuan, J. H., Wang, F., et al. (2017). METTL14 suppresses the metastatic potential of hepatocellular carcinoma by modulating N(6) -methyladenosine-dependent primary MicroRNA processing. Hepatology 65, 529–543. doi:10.1002/hep.28885
Nance, D. J., Satterwhite, E. R., Bhaskar, B., Misra, S., Carraway, K. R., and Mansfield, K. D. (2020). Characterization of METTL16 as a cytoplasmic RNA binding protein. PLoS One 15, e0227647. doi:10.1371/journal.pone.0227647
Nettersheim, D., Berger, D., Jostes, S., Kristiansen, G., Lochnit, G., and Schorle, H. (2019). N6-Methyladenosine detected in RNA of testicular germ cell tumors is controlled by METTL3, ALKBH5, YTHDC1/F1/F2, and HNRNPC as writers, erasers, and readers. Andrology 7, 498–506. doi:10.1111/andr.12612
Ni, W., Yao, S., Zhou, Y., Liu, Y., Huang, P., Zhou, A., et al. (2019). Long noncoding RNA GAS5 inhibits progression of colorectal cancer by interacting with and triggering YAP phosphorylation and degradation and is negatively regulated by the m(6)A reader YTHDF3. Mol. Cancer 18, 143. doi:10.1186/s12943-019-1079-y
Oerum, S., Meynier, V., Catala, M., and Tisne, C. (2021). A comprehensive review of m6A/m6Am RNA methyltransferase structures. Nucleic Acids Res. 49, 7239–7255. doi:10.1093/nar/gkab378
Quinn, J. J., and Chang, H. Y. (2016). Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 17, 47–62. doi:10.1038/nrg.2015.10
Schmitt, A. M., and Chang, H. Y. (2016). Long noncoding RNAs in cancer pathways. Cancer Cell 29, 452–463. doi:10.1016/j.ccell.2016.03.010
Scholler, E., Weichmann, F., Treiber, T., Ringle, S., Treiber, N., Flatley, A., et al. (2018). Interactions, localization, and phosphorylation of the m(6)A generating METTL3-METTL14-WTAP complex. RNA 24, 499–512. doi:10.1261/rna.064063.117
Sendinc, E., Valle-Garcia, D., Dhall, A., Chen, H., Henriques, T., Navarrete-Perea, J., et al. (2019). PCIF1 catalyzes m6Am mRNA methylation to regulate gene expression. Mol. Cell 75, 620–630. doi:10.1016/j.molcel.2019.05.030
Shi, H., Wang, X., Lu, Z., Zhao, B. S., Ma, H., Hsu, P. J., et al. (2017). YTHDF3 facilitates translation and decay of N(6)-methyladenosine-modified RNA. Cell Res. 27, 315–328. doi:10.1038/cr.2017.15
Shi, H., Wei, J., and He, C. (2019). Where, when, and how: Context-dependent functions of RNA methylation writers, readers, and erasers. Mol. Cell 74, 640–650. doi:10.1016/j.molcel.2019.04.025
Song, P., Feng, L., Li, J., Dai, D., Zhu, L., Wang, C., et al. (2020). β-catenin represses miR455-3p to stimulate m6A modification of HSF1 mRNA and promote its translation in colorectal cancer. Mol. Cancer 19, 129. doi:10.1186/s12943-020-01244-z
Wahlestedt, C. (2013). Targeting long non-coding RNA to therapeutically upregulate gene expression. Nat. Rev. Drug Discov. 12, 433–446. doi:10.1038/nrd4018
Wang, Y., Lu, J. H., Wu, Q. N., Jin, Y., Wang, D. S., Chen, Y. X., et al. (2019). LncRNA LINRIS stabilizes IGF2BP2 and promotes the aerobic glycolysis in colorectal cancer. Mol. Cancer 18, 174. doi:10.1186/s12943-019-1105-0
Wei, C., Gershowitz, A., and Moss, B. (1975). N6, O2'-dimethyladenosine a novel methylated ribonucleoside next to the 5' terminal of animal cell and virus mRNAs. Nature 257, 251–253. doi:10.1038/257251a0
Wei, J., and He, C. (2021). Chromatin and transcriptional regulation by reversible RNA methylation. Curr. Opin. Cell Biol. 70, 109–115. doi:10.1016/j.ceb.2020.11.005
Wei, J., Yu, X., Yang, L., Liu, X., Gao, B., Huang, B., et al. (2022). FTO mediates LINE1 m(6)A demethylation and chromatin regulation in mESCs and mouse development. Science 376, 968–973. doi:10.1126/science.abe9582
Wu, H., Ding, X., Hu, X., Zhao, Q., Chen, Q., Sun, T., et al. (2022). LINC01021 maintains tumorigenicity by enhancing N6-methyladenosine reader IMP2 dependent stabilization of MSX1 and JARID2: Implication in colorectal cancer. Oncogene 41, 1959–1973. doi:10.1038/s41388-022-02189-x
Xiao, S., Cao, S., Huang, Q., Xia, L., Deng, M., Yang, M., et al. (2019). The RNA N(6)-methyladenosine modification landscape of human fetal tissues. Nat. Cell Biol. 21, 651–661. doi:10.1038/s41556-019-0315-4
Yan, J., Huang, X., Zhang, X., Chen, Z., Ye, C., Xiang, W., et al. (2020). LncRNA LINC00470 promotes the degradation of PTEN mRNA to facilitate malignant behavior in gastric cancer cells. Biochem. Biophys. Res. Commun. 521, 887–893. doi:10.1016/j.bbrc.2019.11.016
Yang, X., Zhang, S., He, C., Xue, P., Zhang, L., He, Z., et al. (2020). METTL14 suppresses proliferation and metastasis of colorectal cancer by down-regulating oncogenic long non-coding RNA XIST. Mol. Cancer 19, 46. doi:10.1186/s12943-020-1146-4
Yang, Z., Li, J., Feng, G., Gao, S., Wang, Y., Zhang, S., et al. (2017). MicroRNA-145 modulates N(6)-methyladenosine levels by targeting the 3'-untranslated mRNA region of the N(6)-methyladenosine binding YTH domain family 2 protein. J. Biol. Chem. 292, 3614–3623. doi:10.1074/jbc.M116.749689
Yuan, S. X., Wang, J., Yang, F., Tao, Q. F., Zhang, J., Wang, L. L., et al. (2016). Long noncoding RNA DANCR increases stemness features of hepatocellular carcinoma by derepression of CTNNB1. Hepatology 63, 499–511. doi:10.1002/hep.27893
Zhang, J., Bai, R., Li, M., Ye, H., Wu, C., Wang, C., et al. (2019a). Excessive miR-25-3p maturation via N(6)-methyladenosine stimulated by cigarette smoke promotes pancreatic cancer progression. Nat. Commun. 10, 1858. doi:10.1038/s41467-019-09712-x
Zhang, J., Guo, S., Piao, H. Y., Wang, Y., Wu, Y., Meng, X. Y., et al. (2019b). ALKBH5 promotes invasion and metastasis of gastric cancer by decreasing methylation of the lncRNA NEAT1. J. Physiol. Biochem. 75, 379–389. doi:10.1007/s13105-019-00690-8
Zhang, J., Tsoi, H., Li, X., Wang, H., Gao, J., Wang, K., et al. (2016). Carbonic anhydrase IV inhibits colon cancer development by inhibiting the Wnt signalling pathway through targeting the WTAP-WT1-TBL1 axis. Gut 65, 1482–1493. doi:10.1136/gutjnl-2014-308614
Zhang, X., Xu, Y., Qian, Z., Zheng, W., Wu, Q., Chen, Y., et al. (2018). circRNA_104075 stimulates YAP-dependent tumorigenesis through the regulation of HNF4a and may serve as a diagnostic marker in hepatocellular carcinoma. Cell Death Dis. 9, 1091. doi:10.1038/s41419-018-1132-6
Zhu, L., Zhu, Y., Han, S., Chen, M., Song, P., Dai, D., et al. (2019). Impaired autophagic degradation of lncRNA ARHGAP5-AS1 promotes chemoresistance in gastric cancer. Cell Death Dis. 10, 383. doi:10.1038/s41419-019-1585-2
Keywords: m6A modification, RNA modification, ncRNA, microRNA, lncRNA
Citation: Su T, Liu J, Zhang N, Wang T, Han L, Wang S and Yang M (2023) New insights on the interplays between m6A modifications and microRNA or lncRNA in gastrointestinal cancers. Front. Cell Dev. Biol. 11:1157797. doi: 10.3389/fcell.2023.1157797
Received: 03 February 2023; Accepted: 09 June 2023;
Published: 19 June 2023.
Edited by:
Simon Samuel McDade, Queen’s University Belfast, United KingdomReviewed by:
Jiangbo Wei, The University of Chicago, United StatesCopyright © 2023 Su, Liu, Zhang, Wang, Han, Wang and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Suzhen Wang, d2FuZ3N1emhlbjE5ODIwOTAzQDE2My5jb20=; Ming Yang, YWFyeW91bmdAeWVhaC5uZXQ=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.