
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol., 27 February 2023
Sec. Molecular and Cellular Pathology
Volume 11 - 2023 | https://doi.org/10.3389/fcell.2023.1125723
This article is part of the Research TopicTranscription Factors in Cardiovascular Development and RemodelingView all 5 articles
 Na Wang1†
Na Wang1† Jing Hua1†
Jing Hua1† Yuhua Fu2†Jun An3†Xiangyu Chen1Chuancui Wang4Yanghong Zheng1
Yuhua Fu2†Jun An3†Xiangyu Chen1Chuancui Wang4Yanghong Zheng1 Feilong Wang1Yingqun Ji1*Qiang Li1*
Feilong Wang1Yingqun Ji1*Qiang Li1*Pulmonary hypertension (PH) is a group of syndromes characterized by irreversible vascular remodeling and persistent elevation of pulmonary vascular resistance and pressure, leading to ultimately right heart failure and even death. Current therapeutic strategies mainly focus on symptoms alleviation by stimulating pulmonary vessel dilation. Unfortunately, the mechanism and interventional management of vascular remodeling are still yet unrevealed. Hypoxia plays a central role in the pathogenesis of PH and numerous studies have shown the relationship between PH and hypoxia-inducible factors family. EPAS1, known as hypoxia-inducible factor-2 alpha (HIF-2α), functions as a transcription factor participating in various cellular pathways. However, the detailed mechanism of EPAS1 has not been fully and systematically described. This article exhibited a comprehensive summary of EPAS1 including the molecular structure, biological function and regulatory network in PH and other relevant cardiovascular diseases, and furthermore, provided theoretical reference for the potential novel target for future PH intervention.
Pulmonary hypertension (PH) is a group of diseases caused by various etiologies, including genetic mutations and environmental and toxin exposures, that results in pulmonary vascular remodeling and the subsequent elevation of vascular resistance and pressure and right ventricular overload, ultimately leading to right heart decompensation and death (Zheng et al., 2020). Clinically, PH is categorized into five groups: pulmonary arterial hypertension (PAH), PH associated with left heart disease, PH associated with lung disease and/or hypoxia, PH associated with pulmonary artery obstructions, and PH with unclear or multifactorial mechanisms (Hoeper et al., 2016; Humbert et al., 2022). The diagnostic criteria for PH were updated as a mean pulmonary artery pressure (mPAP) > 20 mmHg at rest during right heart catheterization according to the 2022 European Society of Cardiology/European Respiratory Society PH guidelines (Humbert et al., 2022). The primary pathophysiology of pulmonary vascular remodeling involves the intimal endothelial cell (ECs) proliferation and apoptotic resistance, medial smooth muscle cell (SMCs) hypertrophy and proliferation, adventitial fibroblast proliferation and activation with excessive extracellular matrix (ECM) deposition, and interstitial or perivascular inflammatory infiltration (Tuder et al., 2013a; Tuder et al., 2013b; Galiè et al., 2019). Although multiple pharmacological therapies have been developed over the past few decades that mainly target vasomotor tones such as endothelin (ET), prostaglandin I2, and nitric oxide (NO)/cyclic guanosine monophosphate pathways (Zheng et al., 2020), PH patients have a relatively low survival rate and high mortality, about 57% (Benza et al., 2012; Tuder et al., 2013a; Tuder et al., 2013b; He et al., 2020), due to the complexity of etiologies and limited interventions for irreversible pulmonary vascular remodeling process.
Endothelial PAS domain-containing protein 1 (EPAS1), also known as hypoxia-inducible factor 2 alpha (HIF-2α), is a protein encoded by the EPAS1 gene on chromosome 2 of humans and almost exclusively expressed by ECs (Young et al., 2019). According to the bibliometric analysis (Figure 1), studies in the past 20 years revealed the transcriptional regulatory role of HIF-2α in different cellular pathways including hypoxic metabolism, inflammation, apoptosis, and angiogenesis (Luo et al., 2011), as well as various correlated diseases, including pheochromocytomas, Von-Hipple-Lindau disease, and other malignancies. The most specific pathological condition relevant to HIF-2α is high-altitude environmental adaptation, such as acute or chronic mountain sickness and subsequent secondary polycythemia with erythrocytosis.
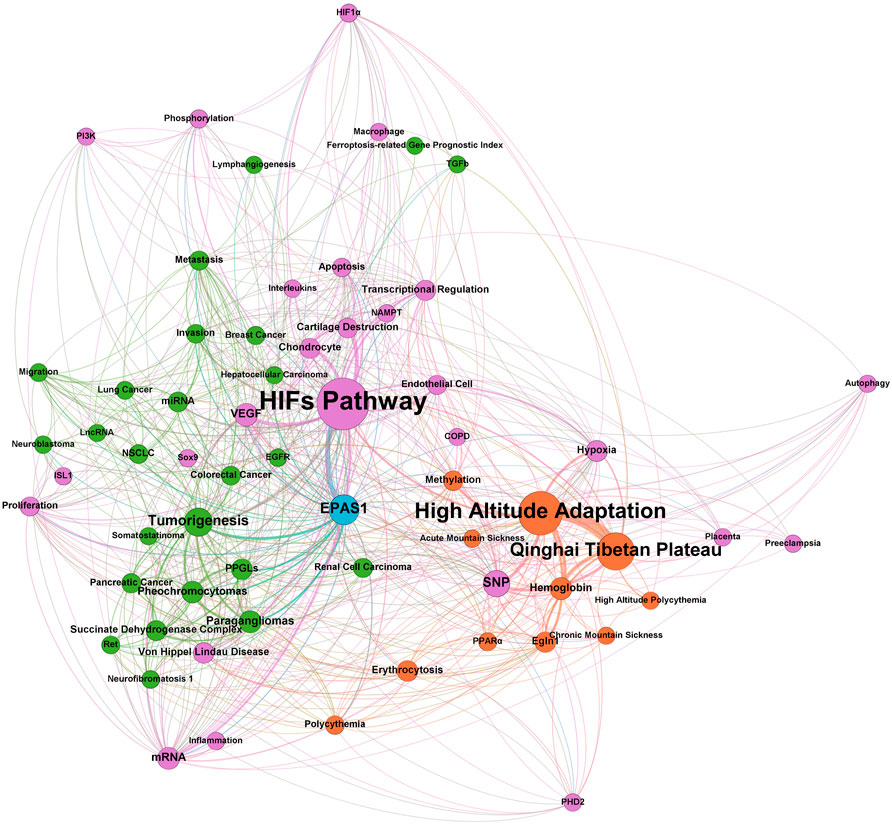
FIGURE 1. Bibliometrics analysis of EPAS1 in the past 20 years.
Furthermore, consecutive publications have illustrated the pathological role of EPAS1 in the development of PH in the past decade, but its mechanism of action has not been thoroughly explored. Therefore, this review aims to provide a comprehensive summary of EPAS1 in the pathogenesis and potential therapeutic interventions of PH.
As first identified by Semenza in Hep3 cells in 1992 (Semenza and Wang, 1992), the hypoxia-inducible factor (HIF) family is a group of transcription factors involved in the intracellular response sensing oxygen concentration, which is strongly associated with organism growth and development and disease pathogenesis.
Structurally, HIF is a heterodimer composed of an oxygen-sensitive alpha subunit and constitutively expressed beta subunit (Figure 2). Both alpha and beta subunits belong to the PER-ARNT-SIM (PAS) subfamily of basic helix-loop-helix (bHLH) transcription factors. EPAS1 is located on chromosome 2 (specifically p16–21 regions), shares 48% homology of the primary amino acid sequence with HIF-1α(Tian et al., 1997), and contains the following domains (Yang et al., 2005; Zhao et al., 2015): 1) a bHLH domain at the N-terminal for DNA binding to hypoxia-responsive elements (HREs) in the promoters or enhancers of target genes (Lim et al., 2013); 2) a PAS domain that facilitates heterodimer formation with beta subunit; 3) the oxygen-dependent degradation domain (ODDD) is responsible for the hydroxylation of proline residues (Pro-405 and Pro-531) and degradation under normoxic conditions; and 4) two transcriptional activation domains (TADs) recruit and interact with transcriptional coregulatory proteins such as CREB-binding protein/E1A binding protein p300 (CBP/p300) to maintain transcriptional activity. N-terminal TAD (N-TAD) lies within the sequence of ODDD, which confers specific target genes to HIF-2α, whereas C-terminal TAD (C-TAD) is responsible for the common target genes of HIF-1α (Daly et al., 2021).
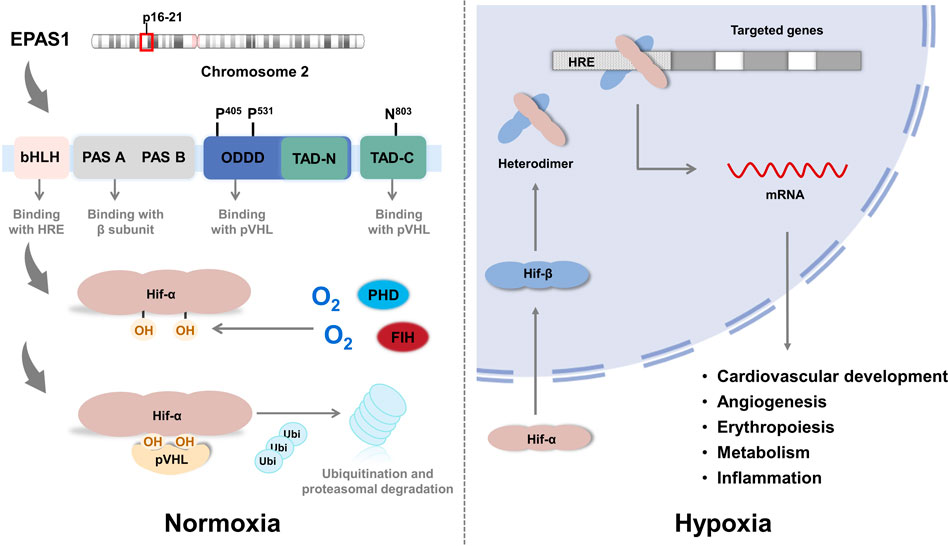
FIGURE 2. The structure of EPAS1.
Under sufficient oxygen availability, HIF alpha subunits are hydroxylated at conserved proline residues by HIF prolyl-hydroxylases (PHD), allowing for the recognition and ubiquitination by von Hippel-Lindau (VHL) E3 ubiquitin ligase, leading to degradation by the proteasome (Daly et al., 2021). HIF prolyl-hydroxylases are inhibited under hypoxic conditions because of the limited oxygen utilization as a cosubstrate, stabilizing the alpha subunit from ubiquitination and allowing it to translocate into the nucleus and form a transcriptionally active heterodimer with the beta subunit. This complex then binds to the HRE regions of the promoters of its downstream genes (Wang et al., 1995), which are involved in various physiological processes, including cardiovascular development, metabolism, inflammation, angiogenesis, and erythropoiesis, that are believed to be associated with PH development (Kim and Yang, 2015; Yoo et al., 2015). Notably, in the study of Petra Miikkulainen (Miikkulainen et al., 2019), a strong positive correlation between HIF-prolyl hydroxylase 3 (PHD3) and HIF-2α mRNA expression in renal clear cell carcinomas (RCCs) is observed, in contrast to the expected accumulation of HIF-2α after PHD3 knockdown in non-RCC cells, which also indicates the aggressiveness and poor prognosis of RCC. In addition to PHD-pVHL pathway, factor inhibiting HIF (FIH) participates the hydroxylation of asparagine-803 in the CTAD domain of HIF-2α and inhibits interactions with transcriptional coactivators such as CBP/p300 (Lando et al., 2002; Greer et al., 2012). Another pathway participating in the degradation of HIF-2α protein is PTEN/PI3K/AKT (Joshi et al., 2014), which phosphorylates E3 ligase of VHL and inactivates the ubiquitination in tumor associated macrophages (TAMs). In terms of transcription, studies revealed that the expression of HIF-2α is positively regulated by IGF induced PI3K-mTORC2 signaling (Mohlin et al., 2015) and peroxisome proliferator-activated receptor gamma coactivator (PGC)/Estrogen-related receptor (ERR) complex families (Hamidian et al., 2015) in neuroblastoma cells; IL-4 signal in macrophages (Takeda et al., 2010); deubiquitylase Cezanne (also known as OTUD7B) by stabilizing transcription factor E2F1 in various cell lines (Moniz et al., 2015); and suppressed by histone deacetylases (HDACs) in soft tissue sarcoma (Nakazawa et al., 2016). An anti-inflammatory cytokine 15-Deoxy-Delta-12,14-prostaglandin J2 (15d-PGJ2), identified by Michael Zimmer (Zimmer et al., 2010) and colleagues, inhibits HIF-2α translation by promoting the binding of iron regulatory protein-1 (IRP1) to iron responsive element (IRE) of HIF-2α message and exerts the anti-inflammatory and putative antineoplastic effects. Other pathways including post-translational modifications and miRNAs interference (Gradin et al., 2002; To et al., 2006; Dioum et al., 2009; Van Hagen et al., 2010; Chen et al., 2012; Poitz et al., 2013; Mathew et al., 2014; Zhang et al., 2014; Sohn et al., 2015; Pangou et al., 2016) are summarized in Table 1, these data disclose the complex regulatory networks of HIF-2α pending further completion.
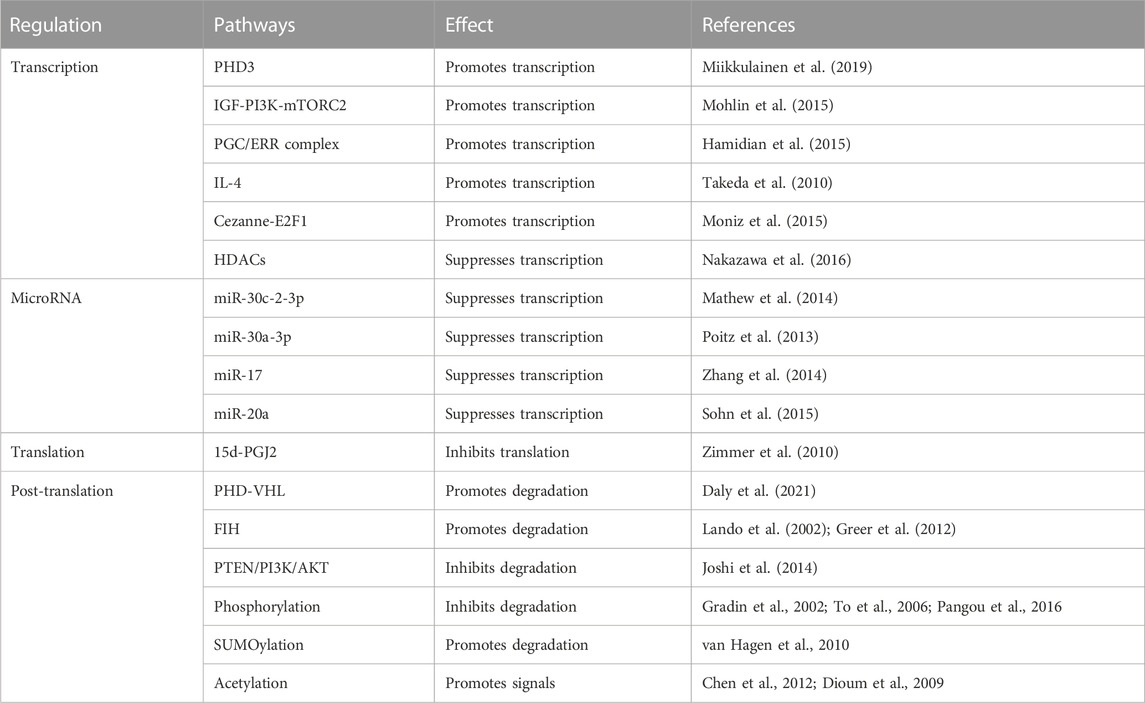
TABLE 1. Regulations of HIF-2α.
EPAS1 was first found by Tian (Tian et al., 1997) in 1997 as a transcription factor exclusively expressed in vascular ECs of the umbilical cord that regulates vascularization and response to hypoxia. The following year, Tian (Tian et al., 1998) further reported that EPAS1 is essential for the maintenance of cardiac output and circulating catecholamine levels from the endothelium under hypoxic conditions during embryonic development. During early embryonic development, as shown in Figure 3, HIF-2α is maintained at a high level in the chromaffin cells of the Organ of Zuckerkandl, the major source of fetal catecholamines. Homozygous EPAS1-deficient mice failed to survive in the mid-gestational stage owing to profound bradycardia and circulatory failure. Additionally, HIF-2α plays an indispensable role in angiogenesis compared to HIF-1α, which mainly activates the glucose metabolic pathways (Wang et al., 2005). Peng (Peng et al., 2000) found that EPAS1-deficient embryos failed to form large vessels or seal intact structures in the yolk sac, indicating improper vascular remodeling during vasculogenesis. Vadive (Vadivel et al., 2014) reported arrested growth of pulmonary vessels and alveoli after EPAS1 expression and HIF-1α was reduced by dominant-negative adenovirus-mediated gene transfer or chetomin. Mice with EC-specific EPAS1 deletion showed increased vascular permeability with ultrastructural abnormalities despite normal vascular anatomic development, resulting in PH(Skuli et al., 2009). Furthermore, EPAS1 regulates angiogenesis in various solid malignancies of the gastrointestinal and genitourinary tracts, such as colonic/hepatic/pancreatic cancer and breast/ovarian/prostate/renal carcinomas (Blancher et al., 2000; Palayoor et al., 2003; Zhang and Rigas, 2006; Osada et al., 2007; Bertout et al., 2008; Imamura et al., 2009; Menrad et al., 2010) by promoting ECs germination, migration, and adherens junctions (Majmundar et al., 2010; Kovacic et al., 2012; Park et al., 2013).
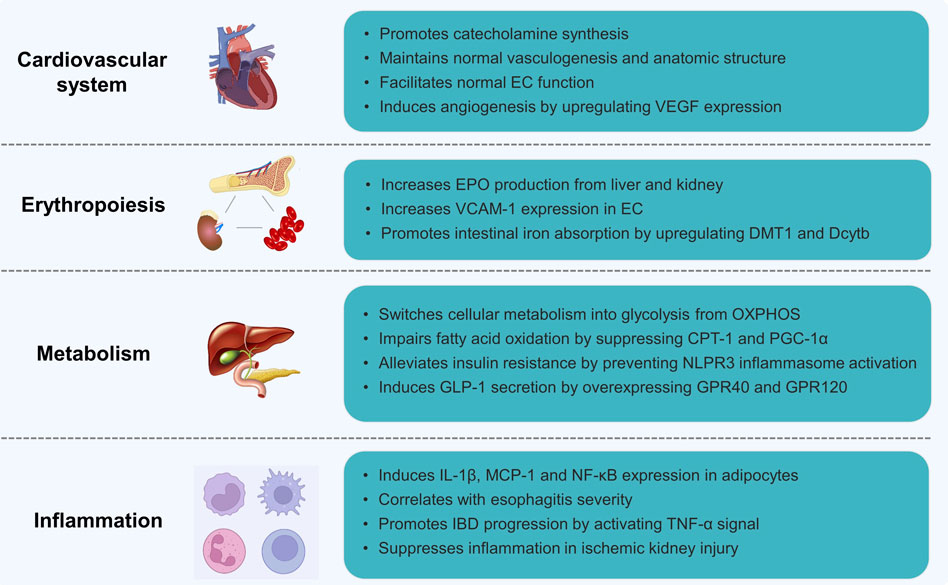
FIGURE 3. The biological functions of EPAS1.
Vascular endothelial growth factor (VEGF) is believed to be the primary target of HIF-2α during angiogenesis. Dumpa (Dumpa et al., 2019) found that caffeine therapy for premature apnea may reduce the incidence of bronchopulmonary dysplasia by enhancing VEGF and angiopoietin expression through the HIF-2α pathway and restoring pulmonary microvasculature and alveolarization in the adult lung. Weidemann (Weidemann et al., 2010) reported that retinal astrocyte-derived VEGF driven by EPAS1 was a key mediator of vascular proliferation in a model of hypoxia-induced retinopathy. Turner (Turner et al., 2002) also revealed consistency in cellular VEGF mRNA and HIF-2α protein levels, indicating the pro-angiogenic role of HIF-2α exertion on the HRE sequences of VEGF and VEGF receptors (VEGFR) and promotion of transcription under hypoxic conditions.
Etiologies triggering excessive erythropoiesis include acute hemorrhage, adaptation to high altitude, or pathological hypoxic conditions such as congenital cyanotic heart diseases and chronic lung diseases (Semenza, 2022). HIF-2α promotes erythropoiesis by regulating erythropoietin (EPO) transcription (Figure 3), a glycoprotein highly expressed in perisinusoidal cells in the liver during embryonic development and produced by interstitial fibroblasts in the kidney during adulthood (Wang et al., 1995; Haase, 2010). HIF-2α recognizes and binds to the core sequence 5′-TACGTGCT-3′ of the EPO HRE sequence, which activates transcription and mediates the cellular response to the hypoxic microenvironment (Hussain et al., 2008). In a study by Gruber M (Gruber et al., 2007), both newborn and adult rats with EPAS1 knockout presented with anemia or severe pancytopenia with suppressed EPO production, indicating the pro-erythrocytosis effect of HIF-2α by transcriptionally regulating EPO production (Scortegagna et al., 2003; Scortegagna et al., 2005). Toshiharu Y (Yamashita et al., 2008) illustrated, however, that HIF-2α governs erythropoiesis by specifically regulating vascular adhesion molecule-1(VCAM-1) expression in ECs to maintain the hematopoietic microenvironment compared to normocytic anemia in mice with EPAS1 knockdown.
In addition to EPO production, HIF-2α stimulates erythropoiesis by participating in intestinal iron absorption, which is in high demand during erythropoiesis and critical for oxygen-carrying red blood cell maturation under hypoxic conditions. Anderson (Anderson et al., 2011) demonstrated that mice with intestinal disruption of EPAS1 (EPAS1ΔIE) showed decreased serum iron levels and expression of iron absorption genes, such as divalent metal transporter 1 (DMT1) and Duodenal cytochrome b (Dcytb), in phenylhydrazine-induced erythropoiesis. In contrast, an iron responsive element (IRE) is found in the 5′untranslated region of the EPAS1 sequence, and the translation of the HIF-2α protein can be repressed by the binding of iron regulatory proteins to the 5′IRE of EPAS1 during iron deficiency (Percy et al., 2007). Additionally, elevated HIF-2α levels during hypoxia suppress hepcidin expression by inducing hepatic EPO production, leading to enhanced intestinal iron uptake and release from internal stores (Liu et al., 2012). Taken together, these data demonstrate the net regulation among EPAS1, erythropoiesis, and iron metabolism.
Metabolism is essential for the maintenance of biological activities, and metabolic reprogramming is a believed hallmark of cellular dysfunction of different etiologies (Faubert et al., 2020). Transformation from mitochondrial oxidative phosphorylation (OXPHOS) to cytoplasmic glycolysis is a major adaptive change in response to insufficient oxygen availability (Shaw, 2006). Unlike HIF-1α, which is considered the mediator of OXPHOS to glycolysis in hypoxic environments (Miska et al., 2019), limited evidence has shown the role of EPAS1 in glycolysis and OXPHOS balance. Farsijani (Farsijani et al., 2016) demonstrated that HIF-2α increased the expression of glycolytic enzymes including hexokinase 2(Hk2), glucose transporter 1 (GLUT1), aldolase C (Aldoc), phosphoglycerate mutase 1 (Pgam1), 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 (PFKFB3), pyruvate kinase M2 (Pkm2), and lactate dehydrogenase A (Ldha), and promoted the synthesis of EPO in renal tubular epithelial cells during hypoxia in a VHL-dependent manner (Figure 3). Early elevated and later downregulated EPAS1 are required for pluripotency induction of induced pluripotent stem cells (iPSC) by glycolysis reprogramming (Mathieu et al., 2014).
In addition to glycolysis, accumulated HIF-2α during chronic hypoxia impairs fatty acid β-oxidation in hepatocytes and ultimately leads to steatosis by suppressing the expression of carnitine palmitoyl transferase 1 (CPT-1) and proliferator-activated receptor-γ coactivator-1α (PGC-1α), rate-limiting enzymes for the β-oxidation pathway, which can be rescued after EPAS1 knockdown (Liu et al., 2014). Similarly, Li (Li et al., 2021) revealed that HIF-2α transcriptally suppresses CPT-1 expression and prevents NLPR3 inflammasome activation, ultimately alleviating insulin resistance in chronic metabolic diseases. Qu (Qu et al., 2011) demonstrated that mice with liver-specific disruption of Vhl achieved EPAS1 overexpression and presented steatosis accompanied by pro-inflammatory and fibrogenic cytokine overexpression. The latest finding by Mooli (Mooli et al., 2022) suggest that HIF-2α augments G-protein-coupled receptor 40 (GPR40) and 120 (GPR120) expressions in intestinal lipid-sensing L cells and induces GLP-1 secretion, regulating the lipid metabolism network. Moreover, constitutive activation of EPAS1 impairs both fatty acid β-oxidation and lipogenesis by downregulating associated genes, whereas it promotes gene expression within the lipid storage and gluconeogenesis pathways (Rankin et al., 2009).
Microenvironmental hypoxia is commonly observed in various systemic inflammatory diseases, such as atherosclerosis, diabetes mellitus, inflammatory bowel diseases (IBD), cancers, and PH. In adipocytes (Figure 3), EPAS1 overexpression upregulates pro-inflammatory cytokines, including interleukin (IL)-1β, monocyte chemotactic protein-1, and the nuclear factor κB (NF-κB) pathway (Lin et al., 2013). Mice with adipocyte-specific EPAS1 overexpression developed lethal cardiac hypertrophy, which is considered the molecular mechanism linking obesity and cardiomyopathy. Xue (Xue et al., 2013; Kerber et al., 2020) reported that highly activated EPAS1 in the intestinal epithelium of IBD patients and mice enhances tumor necrosis factor-α (TNF-α) expression and promotes experimental colitis by inducing TNF-α promoter activity through the myc-associated zinc-finger protein (MAZ) binding, sparing NF-κB pathway. Esophageal HIF-2α expression correlates with reflux esophagitis severity. Refluxed acidic bile salts stabilize epithelial HIF-2α promoting pro-inflammatory cytokine expression (Huo et al., 2017). In contrast, HIF-2α in renal endothelial cells suppresses inflammatory reactions and sustains recovery from ischemic kidney injury (Kapitsinou et al., 2014). The pro-inflammatory role of HIF-2α has also been reported in osteoarthritis (Inoue et al., 2015), hepatoma (Ahn et al., 2010), atopic dermatitis, and psoriasis (Tashiro et al., 2019).
Chronic hypoxia signaling plays a central role in the progression of pulmonary vascular remodeling, leading to irreversible PH and right heart failure. Investigators implied different cellular predominance of HIF-1 and HIF-2 in the development of PH. EPAS1 exclusively governs the ECs dysfunction compared with HIF-1α, whereas in SMCs and fibroblasts, both HIF-1α and EPAS1 orchestrate the pathogenesis of vascular remodeling (Waypa and Schumacker, 2019). Detailed aspects of EPAS1 in PH pathogenesis are listed as below (Figure 4).
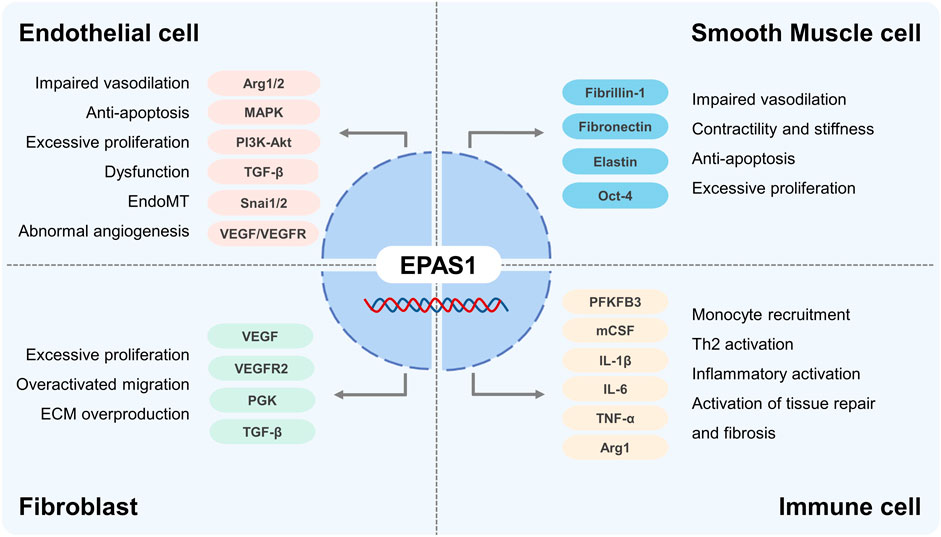
FIGURE 4. The role of EPAS1 in Pulmonary Hypertension.
As the primary initiation of pulmonary vasculopathy, different molecular mechanisms focusing on EC dysfunction, including a spatiotemporal imbalance between apoptosis and proliferation, impairment of migration, sprouting and angiogenesis, methanological abnormalities, and endothelial to mesenchymal transition (EndoMT), explain the development of PH. HIF-2α was significantly elevated in the lung tissues, especially in ECs from patients with idiopathic PH, activates the cellular growth pathways, including MAPK, PI3K-Akt, VEGF, and transforming growth factor-β, increases pulmonary artery EC proliferation as well as collagen synthesis, and stimulates fibrotic remodeling of the pulmonary vasculature (Rimon et al., 2008; Fijalkowska et al., 2010; Bryant et al., 2016; Dai et al., 2018). Upregulated HIF-2α in ECs increases arginase 1 transcription and competes with the endothelial NO complex enzyme for L-Arginine, decreasing NO synthesis, the predominant vasodilator, promoting vascular remodeling and PH progression (Xu et al., 2004; Girgis et al., 2005; Cowburn et al., 2016). Research (Macias et al., 2021) also shown that Arg2 enzyme activity is elevated and correlated with EPAS1 expression in vascular ECs derived from patients with PH. Using model of excessive accumulation of HIF-1 and HIF-2 proteins by inhibiting HIF prolyl hydroxylase domain-containing protein 2 (PHD2), investigators concluded that mice with EC knockout of PHD2 developed severe pulmonary hypertension and right ventricular failure in a HIF-2 dependent manner, despite excessive accumulations of both HIF-1 and HIF-2 proteins (Kapitsinou et al., 2016). Dai (Dai et al., 2016) reported that mice with EC and bone marrow hematopoietic cells knockout of PHD2 showed severe obliterative vascular remodeling and PH, by promoting SMCs proliferation through CXCL12 activation. HIF-2α (Tang et al., 2018) promotes ECs transition into mesenchymal myofibroblasts (EndoMT) by enhancing the expression of Snai1/Snai2, zinc-finger transcription factors family, and leads to remodeled pulmonary vasculopathy. EC-specific deletion of EPAS1 may reduce the expression of ECM proteins, fibronectin, integrin receptors, and ET-B, disrupt cellular overgrowth, block plexiform formation, and ultimately inhibit hypoxia-induced pulmonary vascular remodeling (Hu et al., 2019). Rodents exposed in chronic hypoxia plus inhibition of VEGFR signal by Sugen5416 present as severe pulmonary obliterative vasculopathy and ventricular failure, whereas selectively disrupting VEGFR2 Y949, the tyrosine at residual 949 of VEGFR2, prevents vascular permeability with subsequent myeloid cells infiltration and pulmonary arterioles muscularization (Zhou et al., 2022). HIF-2α regulates angiogenesis in ECs by promoting transcription of VEGF-A/C and VEGFR1/2 pathway, regardless VEGF-B and VEGFR3 regulated by HIF-1α(Downes et al., 2018). Although the correlation between HIF-2α and VEGF/VEGFR signal pathway in PH development is rarely reported, but the paradoxical influence in ECs and pulmonary vascular function is still required clarification.
The hyperplastic proliferation of pulmonary arterial SMCs (PASMCs) is attributed to the core pathogenesis of pulmonary vascular remodeling. Different studies showed that HIF-1α is overactivated in PASMCs in PH, whereas EPAS1 is rarely reported (Dai et al., 2018). In the study of Xin Yi Chan (Chan et al., 2021), gain-of-function mutation of EPAS1 in SMCs increases the expression of fibrillin-1, fibronectin, and elastic fiber components elastin (ELN) via activating endothelin 1 (EDN1) transcription, which contributes to the contractility and stiffness of SMCs and development of pulmonary hypertension in mice. Another data support from Raghavan (Raghavan et al., 2012) pointed out that HIF-2α stimulated PASMCs proliferation by upregulating the expression of transcription factor Oct-4 (Firth et al., 2010). Conclusively, the effects of EPAS1 on PASMCs in PH are still undetermined and correlations among different vascular cells governed by HIF families pend further investigations.
Adventitial fibroblast activation and transition to myofibroblasts, the major origin of collagen and ECM synthesis, are believed to promote vasculopathy in PH. With inhibition of HIF-1α and EPAS1 using RNA interference technology, Eul (Eul et al., 2006) reported that the proliferative response of adventitial fibroblasts relies only on HIF-2α by activation of VEGF, VEGFR2, Phosphoglycerate kinase (PGK) and TGFβ, while the migratory response is correlated with both HIF-1α and HIF-2α. Moreover, mutation of the VHL gene at codon 200 results in cellular HIF-2α accumulation, further induces pulmonary fibroblasts activation with ECM overproduction, and, thereby promoting pulmonary fibrosis (Hickey et al., 2010).
Myeloid cells, especially the monocyte-macrophage lineage, are increasingly reported to contribute to PH pathogenesis (Lim et al., 2013; He et al., 2020). During the early stage of PH, circulatory monocytes are recruited to the pulmonary interstitial space, infiltrating as a pro-inflammatory phenotype and mediating EC dysfunction and PASMC proliferation (Tian et al., 2013; Florentin et al., 2018). In addition to HIF-1α, Wang (Wang et al., 2021) demonstrated that myeloid activation of PFKFB3 impels inflammatory macrophage differentiation via the HIF-2α pathway. HIF-2α upregulates macrophage colony-stimulating factor (M-CSF) expression and promotes monocyte perivascular infiltration and differentiation into inflammatory macrophages (Wang et al., 2018), secreting cytokines including IL-1β, IL-6, IL-8, IL-13, IL-18, and TNF-α, which are believed to be related to PH severity and prognosis (Groth et al., 2014). Interestingly, phenotypic alterations in anti-inflammatory/profibrotic macrophages participate in the late stage of pulmonary vascular remodeling. Few studies have revealed the role of EPAS1 in anti-inflammatory macrophages in PH. Li and colleagues (Li et al., 2021) argued that NLRP3 inflammasome activation of M1 could be suppressed by HIF-2α, thereby preventing insulin resistance. Takeda (Takeda et al., 2010) explained that EPAS1 induced by T helper 2 cytokines (IL-4) specifically promoted arginase 1 expression during M2 macrophage polarization. Despite 48% structural similarity, several data revealed the opposing regulatory roles of HIF-1α and HIF-2α on macrophages phenotype dominance which destine outcomes of inflammatory diseases and malignant tumor (Takeda et al., 2010; Eubank et al., 2011).
Surveys by Scheuermann (Scheuermann et al., 2009; Scheuermann et al., 2013). have identified a hydrophobic cavity at PAS-B domain of EPAS1 which can be occupied by artificial ligands, disrupting the EPAS1-ARNT heterodimer formation, inhibiting the transcription of target genes, and potentially reverse the development of PH. Zimmer (Zimmer et al., 2008) reported a small molecule inhibitor of HIF-2α, C76, facilitates the binding of Iron Regulatory Protein 1 (IRP1) to the IRE of EPAS1 message and abolishes EPAS1 translation, showing as a potential strategy ameliorating the vascular remodeling and right ventricular hypertrophy in PH. Coincidentally, Hu demonstrated (Hu et al., 2019) mice with EPAS1 inducible deletion by antisense oligonucleotides (EPAS1-ASO) exhibited a decreased right ventricular hypertrophy index, reduced vascular remodeling and increased survival of PH. PT2567, an orally bioavailable compound of HIF-2α inhibitor, significantly diminished the early monocytes recruitment, pulmonary vascular cells proliferation, right ventricular remodeling, and plasma nitrite concentration in rats during hypoxia induced PH development. In vitro study, PT2567 reduces arginase1 activity induced by HIF-2α and attenuates inflammation and dysfunction of ECs(Macias et al., 2021). These data indicate the potential role of pharmacological agents targeting on HIF-2α in the interventional strategies of PH.
The current standard management of PH mainly focuses on regenerating normal vasomotor function instead of preventing vascular remodeling, which causes the high mortality and low quality of life. A new interventional strategy reversing pulmonary vascular remodeling is imperative for the treatment of PH. As a key regulator of PH, HIF-2α is involved in pulmonary vascular remodeling, erythropoiesis, the inflammatory response, and hypoxic metabolism during PH. Therapies targeting or blocking HIF-2α pathways have proven advantages in opposing vascular remodeling progression both in vivo and in vitro (Zheng et al., 2022). However, most exploration lines within the preclinical stage, and credible and valid data from clinical trials are required for further verification. In addition, more investigations on the biological function of the EPAS1 regulation network are required to develop a comprehensive picture of the pathophysiology of PH.
NW, writing—original draft, reviewing and editing. JH, literature review and data collection. YF, data collection and figures draft. JA, writing—original draft. XC, data collection. CW, data collection. YZ, writing—figures drafting. FW, writing—review and editing, formal analysis. YJ, conceptualization. QL, conceptualization.
This work was supported by the National Natural Science Foundation of China (NSFC) (82200060), the Key specialty Construction Project of Shanghai Pudong New Area Health Commission (Grant No. PWZzk2022-07), the Top-level Clinical Discipline Project of Shanghai Pudong New Area (Grant No. PWYgf 2021-05).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ahn, Y. T., Chua, M. S., Whitlock, J. P., Shin, Y. C., Song, W. H., Kim, Y., et al. (2010). Rodent-specific hypoxia response elements enhance PAI-1 expression through HIF-1 or HIF-2 in mouse hepatoma cells. Int. J. Oncol. 37, 1627–1638. doi:10.3892/ijo_00000817
Anderson, E. R., Xue, X., and Shah, Y. M. (2011). Intestinal hypoxia-inducible factor-2alpha (HIF-2alpha) is critical for efficient erythropoiesis. J. Biol. Chem. 286, 19533–19540. doi:10.1074/jbc.M111.238667
Benza, R. L., Miller, D. P., Barst, R. J., Badesch, D. B., Frost, A. E., and Mcgoon, M. D. (2012). An evaluation of long-term survival from time of diagnosis in pulmonary arterial hypertension from the REVEAL Registry. Chest 142, 448–456. doi:10.1378/chest.11-1460
Bertout, J. A., Patel, S. A., and Simon, M. C. (2008). The impact of O2 availability on human cancer. Nat. Rev. Cancer 8, 967–975. doi:10.1038/nrc2540
Blancher, C., Moore, J. W., Talks, K. L., Houlbrook, S., and Harris, A. L. (2000). Relationship of hypoxia-inducible factor (HIF)-1alpha and HIF-2alpha expression to vascular endothelial growth factor induction and hypoxia survival in human breast cancer cell lines. Cancer Res. 60, 7106–7113.
Bryant, A. J., Carrick, R. P., Mcconaha, M. E., Jones, B. R., Shay, S. D., Moore, C. S., et al. (2016). Endothelial HIF signaling regulates pulmonary fibrosis-associated pulmonary hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 310, L249–L262. doi:10.1152/ajplung.00258.2015
Chan, X. Y., Volkova, E., Eoh, J., Black, R., Fang, L., Gorashi, R., et al. (2021). HIF2A gain-of-function mutation modulates the stiffness of smooth muscle cells and compromises vascular mechanics. iScience 24, 102246. doi:10.1016/j.isci.2021.102246
Chen, R., Xu, M., Hogg, R. T., Li, J., Little, B., and Gerard et al., R. D. (2012). The acetylase/deacetylase couple CREB-binding protein/Sirtuin 1 controls hypoxia-inducible factor 2 signaling. J Biol Chem 287, 30800–11.
Cowburn, A. S., Crosby, A., Macias, D., Branco, C., Colaço, R. D., Southwood, M., et al. (2016). HIF2α-arginase axis is essential for the development of pulmonary hypertension. Proc. Natl. Acad. Sci. U. S. A. 113, 8801–8806. doi:10.1073/pnas.1602978113
Dai, Z., Li, M., Wharton, J., Zhu, M. M., and Zhao, Y. Y. (2016). Prolyl-4 hydroxylase 2 (PHD2) deficiency in endothelial cells and hematopoietic cells induces obliterative vascular remodeling and severe pulmonary arterial hypertension in mice and humans through hypoxia-inducible factor-2α. Circulation 133, 2447–2458. doi:10.1161/CIRCULATIONAHA.116.021494
Dai, Z., Zhu, M. M., Peng, Y., Machireddy, N., Evans, C. E., Machado, R., et al. (2018). Therapeutic targeting of vascular remodeling and right heart failure in pulmonary arterial hypertension with a HIF-2α inhibitor. Am. J. Respir. Crit. Care Med. 198, 1423–1434. doi:10.1164/rccm.201710-2079OC
Daly, L. A., Brownridge, P. J., Batie, M., Rocha, S., Sée, V., and Eyers, C. E. (2021). Oxygen-dependent changes in binding partners and post-translational modifications regulate the abundance and activity of HIF-1α/2α. Sci. Signal 14, eabf6685. doi:10.1126/scisignal.abf6685
Dioum, E. M., Chen, R., Alexander, M. S., Zhang, Q., Hogg, R. T., and Gerard et al., R. D. (2009). Regulation of hypoxia-inducible factor 2alpha signaling by the stress-responsive deacetylase sirtuin 1. Science 324, 1289–93.
Downes, N. L., Laham-Karam, N., Kaikkonen, M. U., and Ylä-Herttuala, S. (2018). Differential but complementary HIF1α and HIF2α transcriptional regulation. Mol. Ther. 26, 1735–1745. doi:10.1016/j.ymthe.2018.05.004
Dumpa, V., Nielsen, L., Wang, H., and Kumar, V. H. S. (2019). Caffeine is associated with improved alveolarization and angiogenesis in male mice following hyperoxia induced lung injury. BMC Pulm. Med. 19, 138. doi:10.1186/s12890-019-0903-x
Eubank, T. D., Roda, J. M., Liu, H., O'neil, T., and Marsh, C. B. (2011). Opposing roles for HIF-1α and HIF-2α in the regulation of angiogenesis by mononuclear phagocytes. Blood 117, 323–332. doi:10.1182/blood-2010-01-261792
Eul, B., Rose, F., Krick, S., Savai, R., Goyal, P., Klepetko, W., et al. (2006). Impact of HIF-1alpha and HIF-2alpha on proliferation and migration of human pulmonary artery fibroblasts in hypoxia. Faseb J. 20, 163–165. doi:10.1096/fj.05-4104fje
Farsijani, N. M., Liu, Q., Kobayashi, H., Davidoff, O., Sha, F., Fandrey, J., et al. (2016). Renal epithelium regulates erythropoiesis via HIF-dependent suppression of erythropoietin. J. Clin. Invest. 126, 1425–1437. doi:10.1172/JCI74997
Faubert, B., Solmonson, A., and Deberardinis, R. J. (2020). Metabolic reprogramming and cancer progression. Science 368, eaaw5473. doi:10.1126/science.aaw5473
Fijalkowska, I., Xu, W., Comhair, S. A., Janocha, A. J., Mavrakis, L. A., Krishnamachary, B., et al. (2010). Hypoxia inducible-factor1alpha regulates the metabolic shift of pulmonary hypertensive endothelial cells. Am. J. Pathol. 176, 1130–1138. doi:10.2353/ajpath.2010.090832
Firth, A. L., Yao, W., Remillard, C. V., Ogawa, A., and Yuan, J. X. (2010). Upregulation of Oct-4 isoforms in pulmonary artery smooth muscle cells from patients with pulmonary arterial hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 298, L548–L557. doi:10.1152/ajplung.00314.2009
Florentin, J., Coppin, E., Vasamsetti, S. B., Zhao, J., Tai, Y. Y., Tang, Y., et al. (2018). Inflammatory macrophage expansion in pulmonary hypertension depends upon mobilization of blood-borne monocytes. J. Immunol. 200, 3612–3625. doi:10.4049/jimmunol.1701287
Galiè, N., Mclaughlin, V. V., Rubin, L. J., and Simonneau, G. (2019). An overview of the 6th world symposium on pulmonary hypertension. Eur. Respir. J. 53, 1802148. doi:10.1183/13993003.02148-2018
Girgis, R. E., Champion, H. C., Diette, G. B., Johns, R. A., Permutt, S., and Sylvester, J. T. (2005). Decreased exhaled nitric oxide in pulmonary arterial hypertension: Response to bosentan therapy. Am. J. Respir. Crit. Care Med. 172, 352–357. doi:10.1164/rccm.200412-1684OC
Gradin, K., Takasaki, C., Fujii-Kuriyama, Y., and Sogawa, K. (2002). The transcriptional activation function of the HIF-like factor requires phosphorylation at a conserved threonine. J Biol Chem 277, 23508–14.
Greer, S. N., Metcalf, J. L., Wang, Y., and Ohh, M. (2012). The updated biology of hypoxia-inducible factor. Embo J. 31, 2448–2460. doi:10.1038/emboj.2012.125
Groth, A., Vrugt, B., Brock, M., Speich, R., Ulrich, S., and Huber, L. C. (2014). Inflammatory cytokines in pulmonary hypertension. Respir. Res. 15, 47. doi:10.1186/1465-9921-15-47
Gruber, M., Hu, C. J., Johnson, R. S., Brown, E. J., Keith, B., and Simon, M. C. (2007). Acute postnatal ablation of Hif-2alpha results in anemia. Proc. Natl. Acad. Sci. U. S. A. 104, 2301–2306. doi:10.1073/pnas.0608382104
Haase, V. H. (2010). Hypoxic regulation of erythropoiesis and iron metabolism. Am. J. Physiol. Ren. Physiol. 299, F1–F13. doi:10.1152/ajprenal.00174.2010
Hamidian, A., Von Stedingk, K., Munksgaard Thorén, M., Mohlin, S., and Påhlman, S. (2015). Differential regulation of HIF-1α and HIF-2α in neuroblastoma: Estrogen-related receptor alpha (ERRα) regulates HIF2A transcription and correlates to poor outcome. Biochem. Biophys. Res. Commun. 461, 560–567. doi:10.1016/j.bbrc.2015.04.083
He, W., Kapate, N., Shields, C. W. T., and Mitragotri, S. (2020). Drug delivery to macrophages: A review of targeting drugs and drug carriers to macrophages for inflammatory diseases. Adv. Drug Deliv. Rev. 165-166, 15–40. doi:10.1016/j.addr.2019.12.001
Hickey, M. M., Richardson, T., Wang, T., Mosqueira, M., Arguiri, E., Yu, H., et al. (2010). The von Hippel-Lindau Chuvash mutation promotes pulmonary hypertension and fibrosis in mice. J. Clin. Invest. 120, 827–839. doi:10.1172/JCI36362
Hoeper, M. M., Humbert, M., Souza, R., Idrees, M., Kawut, S. M., Sliwa-Hahnle, K., et al. (2016). A global view of pulmonary hypertension. Lancet Respir. Med. 4, 306–322. doi:10.1016/S2213-2600(15)00543-3
Hu, C. J., Poth, J. M., Zhang, H., Flockton, A., Laux, A., Kumar, S., et al. (2019). Suppression of HIF2 signalling attenuates the initiation of hypoxia-induced pulmonary hypertension. Eur. Respir. J. 54, 1900378. doi:10.1183/13993003.00378-2019
Humbert, M., Kovacs, G., Hoeper, M. M., Badagliacca, R., Berger, R. M. F., Brida, M., et al. (2022). 2022 ESC/ERS Guidelines for the diagnosis and treatment of pulmonary hypertension. Eur. Respir. J. 61, 2200879. doi:10.1183/13993003.00879-2022
Huo, X., Agoston, A. T., Dunbar, K. B., Cipher, D. J., Zhang, X., Yu, C., et al. (2017). Hypoxia-inducible factor-2α plays a role in mediating oesophagitis in GORD. Gut 66, 1542–1554. doi:10.1136/gutjnl-2016-312595
Hussain, S., Slevin, M., Matou, S., Ahmed, N., Choudhary, M. I., Ranjit, R., et al. (2008). Anti-angiogenic activity of sesterterpenes; natural product inhibitors of FGF-2-induced angiogenesis. Angiogenesis 11, 245–256. doi:10.1007/s10456-008-9108-2
Imamura, T., Kikuchi, H., Herraiz, M. T., Park, D. Y., Mizukami, Y., Mino-Kenduson, M., et al. (2009). HIF-1alpha and HIF-2alpha have divergent roles in colon cancer. Int. J. Cancer 124, 763–771. doi:10.1002/ijc.24032
Inoue, H., Arai, Y., Kishida, T., Terauchi, R., Honjo, K., Nakagawa, S., et al. (2015). Hydrostatic pressure influences HIF-2 alpha expression in chondrocytes. Int. J. Mol. Sci. 16, 1043–1050. doi:10.3390/ijms16011043
Joshi, S., Singh, A. R., Zulcic, M., and Durden, D. L. (2014). A macrophage-dominant PI3K isoform controls hypoxia-induced HIF1α and HIF2α stability and tumor growth, angiogenesis, and metastasis. Mol. Cancer Res. 12, 1520–1531. doi:10.1158/1541-7786.MCR-13-0682
Kapitsinou, P. P., Rajendran, G., Astleford, L., Michael, M., Schonfeld, M. P., Fields, T., et al. (2016). The endothelial prolyl-4-hydroxylase domain 2/hypoxia-inducible factor 2 Axis regulates pulmonary artery pressure in mice. Mol. Cell Biol. 36, 1584–1594. doi:10.1128/MCB.01055-15
Kapitsinou, P. P., Sano, H., Michael, M., Kobayashi, H., Davidoff, O., Bian, A., et al. (2014). Endothelial HIF-2 mediates protection and recovery from ischemic kidney injury. J. Clin. Invest. 124, 2396–2409. doi:10.1172/JCI69073
Kerber, E. L., Padberg, C., Koll, N., Schuetzhold, V., Fandrey, J., and Winning, S. (2020). The importance of hypoxia-inducible factors (HIF-1 and HIF-2) for the pathophysiology of inflammatory bowel disease. Int. J. Mol. Sci. 21, 8551. doi:10.3390/ijms21228551
Kim, S. Y., and Yang, E. G. (2015). Recent advances in developing inhibitors for hypoxia-inducible factor prolyl hydroxylases and their therapeutic implications. Molecules 20, 20551–20568. doi:10.3390/molecules201119717
Kovacic, J. C., Mercader, N., Torres, M., Boehm, M., and Fuster, V. (2012). Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition: From cardiovascular development to disease. Circulation 125, 1795–1808. doi:10.1161/CIRCULATIONAHA.111.040352
Lando, D., Peet, D. J., Whelan, D. A., Gorman, J. J., and Whitelaw, M. L. (2002). Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch. Science 295, 858–861. doi:10.1126/science.1068592
Li, X., Zhang, X., Xia, J., Zhang, L., Chen, B., Lian, G., et al. (2021). Macrophage HIF-2α suppresses NLRP3 inflammasome activation and alleviates insulin resistance. Cell Rep. 36, 109607. doi:10.1016/j.celrep.2021.109607
Lim, C. S., Kiriakidis, S., Sandison, A., Paleolog, E. M., and Davies, A. H. (2013). Hypoxia-inducible factor pathway and diseases of the vascular wall. J. Vasc. Surg. 58, 219–230. doi:10.1016/j.jvs.2013.02.240
Lin, Q., Huang, Y., Booth, C. J., Haase, V. H., Johnson, R. S., Celeste Simon, M., et al. (2013). Activation of hypoxia-inducible factor-2 in adipocytes results in pathological cardiac hypertrophy. J. Am. Heart Assoc. 2, e000548. doi:10.1161/JAHA.113.000548
Liu, Q., Davidoff, O., Niss, K., and Haase, V. H. (2012). Hypoxia-inducible factor regulates hepcidin via erythropoietin-induced erythropoiesis. J. Clin. Invest. 122, 4635–4644. doi:10.1172/JCI63924
Liu, Y., Ma, Z., Zhao, C., Wang, Y., Wu, G., Xiao, J., et al. (2014). HIF-1α and HIF-2α are critically involved in hypoxia-induced lipid accumulation in hepatocytes through reducing PGC-1α-mediated fatty acid β-oxidation. Toxicol. Lett. 226, 117–123. doi:10.1016/j.toxlet.2014.01.033
Luo, S. H., Weng, L. H., Xie, M. J., Li, X. N., Hua, J., Zhao, X., et al. (2011). Defensive sesterterpenoids with unusual antipodal cyclopentenones from the leaves of Leucosceptrum canum. Org. Lett. 13, 1864–1867. doi:10.1021/ol200380v
Macias, D., Moore, S., Crosby, A., Southwood, M., Du, X., Tan, H., et al. (2021). Targeting HIF2α-ARNT hetero-dimerisation as a novel therapeutic strategy for pulmonary arterial hypertension. Eur. Respir. J. 57, 1902061. doi:10.1183/13993003.02061-2019
Majmundar, A. J., Wong, W. J., and Simon, M. C. (2010). Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 40, 294–309. doi:10.1016/j.molcel.2010.09.022
Mathew, L. K., Lee, S. S., Skuli, N., Rao, S., Keith, B., Nathanson, K. L., et al. (2014). Restricted expression of miR-30c-2-3p and miR-30a-3p in clear cell renal cell carcinomas enhances HIF2α activity. Cancer Discov. 4, 53–60. doi:10.1158/2159-8290.CD-13-0291
Mathieu, J., Zhou, W., Xing, Y., Sperber, H., Ferreccio, A., Agoston, Z., et al. (2014). Hypoxia-inducible factors have distinct and stage-specific roles during reprogramming of human cells to pluripotency. Cell Stem Cell 14, 592–605. doi:10.1016/j.stem.2014.02.012
Menrad, H., Werno, C., Schmid, T., Copanaki, E., Deller, T., Dehne, N., et al. (2010). Roles of hypoxia-inducible factor-1alpha (HIF-1alpha) versus HIF-2alpha in the survival of hepatocellular tumor spheroids. Hepatology 51, 2183–2192. doi:10.1002/hep.23597
Miikkulainen, P., Högel, H., Seyednasrollah, F., Rantanen, K., Elo, L. L., and Jaakkola, P. M. (2019). Hypoxia-inducible factor (HIF)-prolyl hydroxylase 3 (PHD3) maintains high HIF2A mRNA levels in clear cell renal cell carcinoma. J. Biol. Chem. 294, 3760–3771. doi:10.1074/jbc.RA118.004902
Miska, J., Lee-Chang, C., Rashidi, A., Muroski, M. E., Chang, A. L., Lopez-Rosas, A., et al. (2019). HIF-1α is a metabolic switch between glycolytic-driven migration and oxidative phosphorylation-driven immunosuppression of tregs in glioblastoma. Cell Rep. 27, 226–237.e4. doi:10.1016/j.celrep.2019.03.029
Mohlin, S., Hamidian, A., Von Stedingk, K., Bridges, E., Wigerup, C., Bexell, D., et al. (2015). PI3K-mTORC2 but not PI3K-mTORC1 regulates transcription of HIF2A/EPAS1 and vascularization in neuroblastoma. Cancer Res. 75, 4617–4628. doi:10.1158/0008-5472.CAN-15-0708
Moniz, S., Bandarra, D., Biddlestone, J., Campbell, K. J., Komander, D., Bremm, A., et al. (2015). Cezanne regulates E2F1-dependent HIF2α expression. J. Cell Sci. 128, 3082–3093. doi:10.1242/jcs.168864
Mooli, R. G. R., Mukhi, D., Pasupulati, A. K., Evers, S. S., Sipula, I. J., Jurczak, M., et al. (2022). Intestinal HIF-2α regulates GLP-1 secretion via lipid sensing in L-cells. Cell Mol. Gastroenterol. Hepatol. 13, 1057–1072. doi:10.1016/j.jcmgh.2021.12.004
Nakazawa, M. S., Eisinger-Mathason, T. S., Sadri, N., Ochocki, J. D., Gade, T. P., Amin, R. K., et al. (2016). Epigenetic re-expression of HIF-2α suppresses soft tissue sarcoma growth. Nat. Commun. 7, 10539. doi:10.1038/ncomms10539
Osada, R., Horiuchi, A., Kikuchi, N., Yoshida, J., Hayashi, A., Ota, M., et al. (2007). Expression of hypoxia-inducible factor 1alpha, hypoxia-inducible factor 2alpha, and von Hippel-lindau protein in epithelial ovarian neoplasms and allelic loss of von Hippel-lindau gene: Nuclear expression of hypoxia-inducible factor 1alpha is an independent prognostic factor in ovarian carcinoma. Hum. Pathol. 38, 1310–1320. doi:10.1016/j.humpath.2007.02.010
Palayoor, S. T., Tofilon, P. J., and Coleman, C. N. (2003). Ibuprofen-mediated reduction of hypoxia-inducible factors HIF-1alpha and HIF-2alpha in prostate cancer cells. Clin. Cancer Res. 9, 3150–3157.
Pangou, E., Befani, C., Mylonis, I., Samiotaki, M., Panayotou, G., and Simos et al., G. (2016). HIF-2α phosphorylation by CK1δ promotes erythropoietin secretion in liver cancer cells under hypoxia. J Cell Sci 129, 4213–4226.
Park, C., Kim, T. M., and Malik, A. B. (2013). Transcriptional regulation of endothelial cell and vascular development. Circ. Res. 112, 1380–1400. doi:10.1161/CIRCRESAHA.113.301078
Peng, J., Zhang, L., Drysdale, L., and Fong, G. H. (2000). The transcription factor EPAS-1/hypoxia-inducible factor 2alpha plays an important role in vascular remodeling. Proc. Natl. Acad. Sci. U. S. A. 97, 8386–8391. doi:10.1073/pnas.140087397
Percy, M. J., Sanchez, M., Swierczek, S., Mcmullin, M. F., Mojica-Henshaw, M. P., Muckenthaler, M. U., et al. (2007). Is congenital secondary erythrocytosis/polycythemia caused by activating mutations within the HIF-2 alpha iron-responsive element? Blood 110, 2776–2777. doi:10.1182/blood-2007-03-082503
Poitz, D. M., Augstein, A., Gradehand, C., Ende, G., Schmeisser, A., and Strasser, R. H. (2013). Regulation of the Hif-system by micro-RNA 17 and 20a - role during monocyte-to-macrophage differentiation. Mol. Immunol. 56, 442–451. doi:10.1016/j.molimm.2013.06.014
Qu, A., Taylor, M., Xue, X., Matsubara, T., Metzger, D., Chambon, P., et al. (2011). Hypoxia-inducible transcription factor 2α promotes steatohepatitis through augmenting lipid accumulation, inflammation, and fibrosis. Hepatology 54, 472–483. doi:10.1002/hep.24400
Raghavan, A., Zhou, G., Zhou, Q., Ibe, J. C., Ramchandran, R., Yang, Q., et al. (2012). Hypoxia-induced pulmonary arterial smooth muscle cell proliferation is controlled by forkhead box M1. Am. J. Respir. Cell Mol. Biol. 46, 431–436. doi:10.1165/rcmb.2011-0128OC
Rankin, E. B., Rha, J., Selak, M. A., Unger, T. L., Keith, B., Liu, Q., et al. (2009). Hypoxia-inducible factor 2 regulates hepatic lipid metabolism. Mol. Cell Biol. 29, 4527–4538. doi:10.1128/MCB.00200-09
Rimon, E., Chen, B., Shanks, A. L., Nelson, D. M., and Sadovsky, Y. (2008). Hypoxia in human trophoblasts stimulates the expression and secretion of connective tissue growth factor. Endocrinology 149, 2952–2958. doi:10.1210/en.2007-1099
Scheuermann, T. H., Li, Q., Ma, H. W., Key, J., Zhang, L., Chen, R., et al. (2013). Allosteric inhibition of hypoxia inducible factor-2 with small molecules. Nat. Chem. Biol. 9, 271–276. doi:10.1038/nchembio.1185
Scheuermann, T. H., Tomchick, D. R., Machius, M., Guo, Y., Bruick, R. K., and Gardner, K. H. (2009). Artificial ligand binding within the HIF2alpha PAS-B domain of the HIF2 transcription factor. Proc. Natl. Acad. Sci. U. S. A. 106, 450–455. doi:10.1073/pnas.0808092106
Scortegagna, M., Ding, K., Zhang, Q., Oktay, Y., Bennett, M. J., Bennett, M., et al. (2005). HIF-2alpha regulates murine hematopoietic development in an erythropoietin-dependent manner. Blood 105, 3133–3140. doi:10.1182/blood-2004-05-1695
Scortegagna, M., Morris, M. A., Oktay, Y., Bennett, M., and Garcia, J. A. (2003). The HIF family member EPAS1/HIF-2alpha is required for normal hematopoiesis in mice. Blood 102, 1634–1640. doi:10.1182/blood-2003-02-0448
Semenza, G. L. (2022). Regulation of erythropoiesis by the hypoxia-inducible factor pathway: Effects of genetic and pharmacological perturbations. Annu. Rev. Med. 74, 307–319. doi:10.1146/annurev-med-042921-102602
Semenza, G. L., and Wang, G. L. (1992). A nuclear factor induced by hypoxia via de novo protein synthesis binds to the human erythropoietin gene enhancer at a site required for transcriptional activation. Mol. Cell Biol. 12, 5447–5454. doi:10.1128/mcb.12.12.5447-5454.1992
Shaw, R. J. (2006). Glucose metabolism and cancer. Curr. Opin. Cell Biol. 18, 598–608. doi:10.1016/j.ceb.2006.10.005
Skuli, N., Liu, L., Runge, A., Wang, T., Yuan, L., Patel, S., et al. (2009). Endothelial deletion of hypoxia-inducible factor-2alpha (HIF-2alpha) alters vascular function and tumor angiogenesis. Blood 114, 469–477. doi:10.1182/blood-2008-12-193581
Sohn, E. J., Won, G., Lee, J., Lee, S., and Kim, S. H. (2015). Upregulation of miRNA3195 and miRNA374b mediates the anti-angiogenic properties of melatonin in hypoxic PC-3 prostate cancer cells. J. Cancer 6, 19–28. doi:10.7150/jca.9591
Takeda, N., O'dea, E. L., Doedens, A., Kim, J. W., Weidemann, A., Stockmann, C., et al. (2010). Differential activation and antagonistic function of HIF-{alpha} isoforms in macrophages are essential for NO homeostasis. Genes Dev. 24, 491–501. doi:10.1101/gad.1881410
Tang, H., Babicheva, A., Mcdermott, K. M., Gu, Y., Ayon, R. J., Song, S., et al. (2018). Endothelial HIF-2α contributes to severe pulmonary hypertension due to endothelial-to-mesenchymal transition. Am. J. Physiol. Lung Cell Mol. Physiol. 314, L256–l275. doi:10.1152/ajplung.00096.2017
Tashiro, N., Segawa, R., Tobita, R., Asakawa, S., Mizuno, N., Hiratsuka, M., et al. (2019). Hypoxia inhibits TNF-α-induced TSLP expression in keratinocytes. PLoS One 14, e0224705. doi:10.1371/journal.pone.0224705
Tian, H., Hammer, R. E., Matsumoto, A. M., Russell, D. W., and Mcknight, S. L. (1998). The hypoxia-responsive transcription factor EPAS1 is essential for catecholamine homeostasis and protection against heart failure during embryonic development. Genes Dev. 12, 3320–3324. doi:10.1101/gad.12.21.3320
Tian, H., Mcknight, S. L., and Russell, D. W. (1997). Endothelial PAS domain protein 1 (EPAS1), a transcription factor selectively expressed in endothelial cells. Genes Dev. 11, 72–82. doi:10.1101/gad.11.1.72
Tian, W., Jiang, X., Tamosiuniene, R., Sung, Y. K., Qian, J., Dhillon, G., et al. (2013). Blocking macrophage leukotriene b4 prevents endothelial injury and reverses pulmonary hypertension. Sci. Transl. Med. 5, 200ra117. doi:10.1126/scitranslmed.3006674
To, K. K., Sedelnikova, O. A., Samons, M., Bonner, W. M., and Huang, L. E. (2006). The phosphorylation status of PAS-B distinguishes HIF-1alpha from HIF-2alpha in NBS1 repression. Embo j 25, 4784–94.
Tuder, R. M., Archer, S. L., Dorfmüller, P., Erzurum, S. C., Guignabert, C., Michelakis, E., et al. (2013a). Relevant issues in the pathology and pathobiology of pulmonary hypertension. J. Am. Coll. Cardiol. 62, D4–D12. doi:10.1016/j.jacc.2013.10.025
Tuder, R. M., Stacher, E., Robinson, J., Kumar, R., and Graham, B. B. (2013b). Pathology of pulmonary hypertension. Clin. Chest Med. 34, 639–650. doi:10.1016/j.ccm.2013.08.009
Turner, K. J., Moore, J. W., Jones, A., Taylor, C. F., Cuthbert-Heavens, D., Han, C., et al. (2002). Expression of hypoxia-inducible factors in human renal cancer: Relationship to angiogenesis and to the von Hippel-lindau gene mutation. Cancer Res. 62, 2957–2961.
Vadivel, A., Alphonse, R. S., Etches, N., Van Haaften, T., Collins, J. J., O'reilly, M., et al. (2014). Hypoxia-inducible factors promote alveolar development and regeneration. Am. J. Respir. Cell Mol. Biol. 50, 96–105. doi:10.1165/rcmb.2012-0250OC
Van Hagen, M., Overmeer, R. M., Abolvardi and , S. S., and Vertegaal, A. C. (2010). RNF4 and VHL regulate the proteasomal degradation of SUMO-conjugated Hypoxia-Inducible Factor-2alpha. Nucleic Acids Res 38, 1922–31.
Wang, G. L., Jiang, B. H., Rue, E. A., and Semenza, G. L. (1995). Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. U. S. A. 92, 5510–5514. doi:10.1073/pnas.92.12.5510
Wang, L., Zhang, X., Cao, Y., Ma, Q., Mao, X., Xu, J., et al. (2021). Mice with a specific deficiency of Pfkfb3 in myeloid cells are protected from hypoxia-induced pulmonary hypertension. Br. J. Pharmacol. 178, 1055–1072. doi:10.1111/bph.15339
Wang, Q., He, Z., Huang, M., Liu, T., Wang, Y., Xu, H., et al. (2018). Vascular niche IL-6 induces alternative macrophage activation in glioblastoma through HIF-2α. Nat. Commun. 9, 559. doi:10.1038/s41467-018-03050-0
Wang, V., Davis, D. A., Haque, M., Huang, L. E., and Yarchoan, R. (2005). Differential gene up-regulation by hypoxia-inducible factor-1alpha and hypoxia-inducible factor-2alpha in HEK293T cells. Cancer Res. 65, 3299–3306. doi:10.1158/0008-5472.CAN-04-4130
Waypa, G. B., and Schumacker, P. T. (2019). Roles of HIF1 and HIF2 in pulmonary hypertension: It all depends on the context. Eur. Respir. J. 54, 1901929. doi:10.1183/13993003.01929-2019
Weidemann, A., Krohne, T. U., Aguilar, E., Kurihara, T., Takeda, N., Dorrell, M. I., et al. (2010). Astrocyte hypoxic response is essential for pathological but not developmental angiogenesis of the retina. Glia 58, 1177–1185. doi:10.1002/glia.20997
Xu, W., Kaneko, F. T., Zheng, S., Comhair, S. A., Janocha, A. J., Goggans, T., et al. (2004). Increased arginase II and decreased NO synthesis in endothelial cells of patients with pulmonary arterial hypertension. Faseb J. 18, 1746–1748. doi:10.1096/fj.04-2317fje
Xue, X., Ramakrishnan, S., Anderson, E., Taylor, M., Zimmermann, E. M., Spence, J. R., et al. (2013). Endothelial PAS domain protein 1 activates the inflammatory response in the intestinal epithelium to promote colitis in mice. Gastroenterology 145, 831–841. doi:10.1053/j.gastro.2013.07.010
Yamashita, T., Ohneda, O., Sakiyama, A., Iwata, F., Ohneda, K., and Fujii-Kuriyama, Y. (2008). The microenvironment for erythropoiesis is regulated by HIF-2alpha through VCAM-1 in endothelial cells. Blood 112, 1482–1492. doi:10.1182/blood-2007-11-122648
Yang, J., Zhang, L., Erbel, P. J., Gardner, K. H., Ding, K., Garcia, J. A., et al. (2005). Functions of the Per/ARNT/Sim domains of the hypoxia-inducible factor. J. Biol. Chem. 280, 36047–36054. doi:10.1074/jbc.M501755200
Yoo, S., Takikawa, S., Geraghty, P., Argmann, C., Campbell, J., Lin, L., et al. (2015). Integrative analysis of DNA methylation and gene expression data identifies EPAS1 as a key regulator of COPD. PLoS Genet. 11, e1004898. doi:10.1371/journal.pgen.1004898
Young, J. M., Williams, D. R., and Thompson, A. a. R. (2019). Thin air, thick vessels: Historical and current perspectives on hypoxic pulmonary hypertension. Front. Med. (Lausanne) 6, 93. doi:10.3389/fmed.2019.00093
Zhang, H., Pu, J., Qi, T., Qi, M., Yang, C., Li, S., et al. (2014). MicroRNA-145 inhibits the growth, invasion, metastasis and angiogenesis of neuroblastoma cells through targeting hypoxia-inducible factor 2 alpha. Oncogene 33, 387–397. doi:10.1038/onc.2012.574
Zhang, Z., and Rigas, B. (2006). NF-kappaB, inflammation and pancreatic carcinogenesis: NF-kappaB as a chemoprevention target (review). Int. J. Oncol. 29, 185–192.
Zhao, J., Du, F., Shen, G., Zheng, F., and Xu, B. (2015). The role of hypoxia-inducible factor-2 in digestive system cancers. Cell Death Dis. 6, e1600. doi:10.1038/cddis.2014.565
Zheng, Q., Lu, W., Yan, H., Duan, X., Chen, Y., Zhang, C., et al. (2022). Established pulmonary hypertension in rats was reversed by a combination of a HIF-2α antagonist and a p53 agonist. Br. J. Pharmacol. 179, 1065–1081. doi:10.1111/bph.15696
Zheng, W., Wang, Z., Jiang, X., Zhao, Q., and Shen, J. (2020). Targeted drugs for treatment of pulmonary arterial hypertension: Past, present, and future perspectives. J. Med. Chem. 63, 15153–15186. doi:10.1021/acs.jmedchem.0c01093
Zhou, W., Liu, K., Zeng, L., He, J., Gao, X., Gu, X., et al. (2022). Targeting VEGF-A/VEGFR2 Y949 signaling-mediated vascular permeability alleviates hypoxic pulmonary hypertension. Circulation 146, 1855–1881. doi:10.1161/CIRCULATIONAHA.122.061900
Zimmer, M., Ebert, B. L., Neil, C., Brenner, K., Papaioannou, I., Melas, A., et al. (2008). Small-molecule inhibitors of HIF-2a translation link its 5'UTR iron-responsive element to oxygen sensing. Mol. Cell 32, 838–848. doi:10.1016/j.molcel.2008.12.004
Zimmer, M., Lamb, J., Ebert, B. L., Lynch, M., Neil, C., Schmidt, E., et al. (2010). The connectivity map links iron regulatory protein-1-mediated inhibition of hypoxia-inducible factor-2a translation to the anti-inflammatory 15-deoxy-delta12,14-prostaglandin J2. Cancer Res. 70, 3071–3079. doi:10.1158/0008-5472.CAN-09-2877
Keywords: EPAS1, pulmonary hypertension, hypoxia, vascular remodeling, transcription factor
Citation: Wang N, Hua J, Fu Y, An J, Chen X, Wang C, Zheng Y, Wang F, Ji Y and Li Q (2023) Updated perspective of EPAS1 and the role in pulmonary hypertension. Front. Cell Dev. Biol. 11:1125723. doi: 10.3389/fcell.2023.1125723
Received: 16 December 2022; Accepted: 14 February 2023;
Published: 27 February 2023.
Edited by:
Yuqin Chen, First Affiliated Hospital of Guangzhou Medical University, ChinaReviewed by:
Kai Yang, First Affiliated Hospital of Guangzhou Medical University, ChinaCopyright © 2023 Wang, Hua, Fu, An, Chen, Wang, Zheng, Wang, Ji and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yingqun Ji, aml5aW5ncXVuQDE2My5jb20=; Qiang Li, bGlxcmVzc2hAaG90bWFpbC5jb20=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.