 Johannes Jun Wei Low
Johannes Jun Wei Low Siti Aishah Sulaiman
Siti Aishah Sulaiman Nor Adzimah Johdi
Nor Adzimah Johdi Nadiah Abu
Nadiah Abu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Cell Dev. Biol. , 16 November 2022
Sec. Molecular and Cellular Pathology
Volume 10 - 2022 | https://doi.org/10.3389/fcell.2022.996805
This article is part of the Research Topic Extracellular Vesicles as Potent Modulators of Immunity View all 5 articles
Glioblastoma (GB) is a type of brain cancer that can be considered aggressive. Glioblastoma treatment has significant challenges due to the immune privilege site of the brain and the presentation of an immunosuppressive tumor microenvironment. Extracellular vesicles (EVs) are cell-secreted nanosized vesicles that engage in intercellular communication via delivery of cargo that may cause downstream effects such as tumor progression and recipient cell modulation. Although the roles of extracellular vesicles in cancer progression are well documented, their immunomodulatory effects are less defined. Herein, we focus on glioblastoma and explain the immunomodulatory effects of extracellular vesicles secreted by both tumor and immune cells in detail. The tumor to immune cells, immune cells to the tumor, and intra-immune cells extracellular vesicles crosstalks are involved in various immunomodulatory effects. This includes the promotion of immunosuppressive phenotypes, apoptosis, and inactivation of immune cell subtypes, which affects the central nervous system and peripheral immune system response, aiding in its survival and progression in the brain.
Central nervous system (CNS) cancers comprise both primary and secondary CNS tumors in which the former are derived from CNS cells while the latter emerges from the spread of cancerous cells from the peripheral body to the brain (Lapointe et al., 2018; Achrol et al., 2019). The incidence rate for secondary CNS tumors is expected to be larger than primary CNS tumors (Davis et al., 2012), with the survival rate determined by the primary cancer location (Cagney et al., 2017). Nevertheless, the low incidence rate of primary CNS cancer is confounded by its high mortality rate (Miller et al., 2021). Gliomas, representing only 24.5% of all primary brain and CNS tumors, hold 80.9% of recorded malignant tumors, signifying the importance of this cancer as a modern-day killer (Ostrom et al., 2021). Currently, both glioblastoma, isocitrate dehydrogenase-wildtype (IDH-wildtype) and astrocytoma, IDH-mutant (previously glioblastoma, IDH-mutant) are classified as WHO Grade 4. Astrocytoma, IDH-mutant is a highly malignant astrocytic glioma with a low survival rate (6.8% survival rate post-diagnosis, median survival of 8 months) and portrayed mutations to either IDH1/2 (Louis et al., 2021; Ostrom et al., 2021). It was previously mentioned that glioblastoma emerges in two different types: 1) primary, in which it arises de novo, or 2) secondary, where it progresses from a lower grade astrocytoma (Ohgaki and Kleihues, 2013) but following the 2021 classification, the term “secondary glioblastoma” which commonly involves R132H mutation in IDH1 (92.7% of various brain tumors) was annulled (Balss et al., 2008; Louis et al., 2021). Glioblastoma (GB) arises de novo with a very fast progression rate (mean 6.3 months from the first symptom to definitive diagnosis) (Ohgaki and Kleihues, 2005; Ohgaki and Kleihues, 2007). Growth dynamics analysis conducted by Stensjoen et al. showed that untreated GB has a median specific growth rate of 1.4% every day, leading to an equivalent volume doubling time of 49.6 days, further showcasing the expedited growth of GB tumors (Stensjøen et al., 2015). GB tumors need to mediate an immunosuppressive microenvironment (Brown et al., 2018) through bidirectional communication with surrounding resident cells via several approaches including soluble factors, direct cell-cell contact, and extracellular vesicles (Broekman et al., 2018), where the latter is increasingly recognized as an important mediator of cell-cell communication (Gao et al., 2020).
The CNS is traditionally known as an “immune privileged” site due to several factors, including 1) the presence of a blood-brain barrier (BBB) that limits access to peripheral immune cells, 2) the lack of a lymphatic vessel serving the CNS which limits antigen trafficking and presentation in lymph nodes, 3) paucity of antigen-presenting cells (APC) in the CNS, 4) downregulation of major histocompatibility complex (MHC) expression in normal brain parenchyma leading to diluted T cell immune response, and 5) presence of anti-inflammatory modulators (Fabry et al., 2008; Brown et al., 2018). The CNS facilitates the entry and continued presence of immune cells to survey and respond against foreign entities such as tumors (Papadopoulos et al., 2020). Still, GB has long been considered a “cold” tumor with high intrinsic and adaptive resistance to immunotherapy (Jackson et al., 2019) due to intratumoral heterogeneity and lack of high-quality neoantigens, as well as severe dysregulation of immune cells favoring the immunosuppressive phenotype (Hao et al., 2002). Nevertheless, efforts have been taken to convert the “cold” GB phenotype to a more immunotherapy-susceptible “hot” phenotype (Tomaszewski et al., 2019). Glioma cells often engage with multiple glial cell types, including immune cells, to create an immunosuppressive tumor microenvironment (TME) via the secretion of pro-tumorigenic mediators (Quail and Joyce, 2017; Brown et al., 2018). Mechanisms relating to immune cells’ response in the GB microenvironment remain scarce and incompletely defined (Lim et al., 2018) although extracellular vesicles are found to be implicated in several instances stated further along in this review.
EVs are small lipid-enclosed membrane vesicles secreted from virtually all kinds of cells into the extracellular spaces to engage in various cellular processes (Urabe et al., 2020). EVs can be classified based on their size: small EVs consist of particles <200 nm in diameter, medium EVs consist of particles between 200–400 nm in diameter, and large EVs consist of particles larger than 400 nm in diameter (Théry et al., 2018). Tumor cells-derived EVs play important roles in modulating the tumor microenvironment (TME) and promoting tumor progression via the transfer of tumor-specific molecules to recipient cells (Ricklefs et al., 2016). Effects of this intercellular communication include the establishment of a premetastatic niche, promoting angiogenesis, disruption to the peritoneum or BBB, chemotherapeutic drug resistance, and formation of heterogenous cancer-associated fibroblast (Urabe et al., 2020). Several studies have also described the variety of cargo carried within the vesicle including proteins and nucleic acids in both small and large EVs (Thakur et al., 2014; Hurwitz et al., 2016; García-Romero et al., 2017; Vagner et al., 2018). In essence, EVs allow biomolecules to be transported in a stable and protected format, allowing liquid biopsy utilizing patients’ blood. García-Romero et al. reported on the presence of glioma tumor-derived genomic DNA (gDNA) despite the presence of the BBB, denoting EV’s capability to bypass the anatomical restriction (García-Romero et al., 2017).
Biomolecules present in and on the extracellular vesicles are representative of parental cells and are often functional (Yekula et al., 2020). A study conducted by Kucharzewska et al. demonstrated that exosomes reflect the hypoxic status of glioma cells evidently through the cargo makeup of the exosome (Kucharzewska et al., 2013). Exosomal enzymes’ mRNA levels correspond to levels in parental cells, further denoting the presence of an elaborate cargo selection machinery in cells (Shao et al., 2015). Several mechanisms have been determined to influence cargo selection in EVs, notably the ADP-ribosylation factor 6 (ARF-6)-Exportin-5 axis, where ARF6-GTP interacts with Exportin-5 to deliver miRNA into tumor microvesicles (MVs) in an ARF6-GTP dependent manner (Clancy et al., 2019). Other than that, target proteins can also be sorted into EVs via ubiquitination, which is mainly mediated by endosomal sorting complexes required for transport (ESCRT) complex and partly by other proteins displaying Ubiquitin (Ub)-binding domains (Piper et al., 2014). Lipid sorting is not well understood, but Bissig and Gruenberg (Bissig and Gruenberg, 2013) summarized several important factors such as lipid-lipid/lipid-protein interaction, differing membrane biophysical traits, and metabolic enzyme turnover rate/distribution to be deeply involved with differing lipid populations in endosomes (Bissig and Gruenberg, 2013). Various other cargoes can also get sorted into exosomes via post-translational modifications (Moreno-Gonzalo et al., 2018). The schematic representation of GB-derived EVs immunosuppression is described in Figure 1.
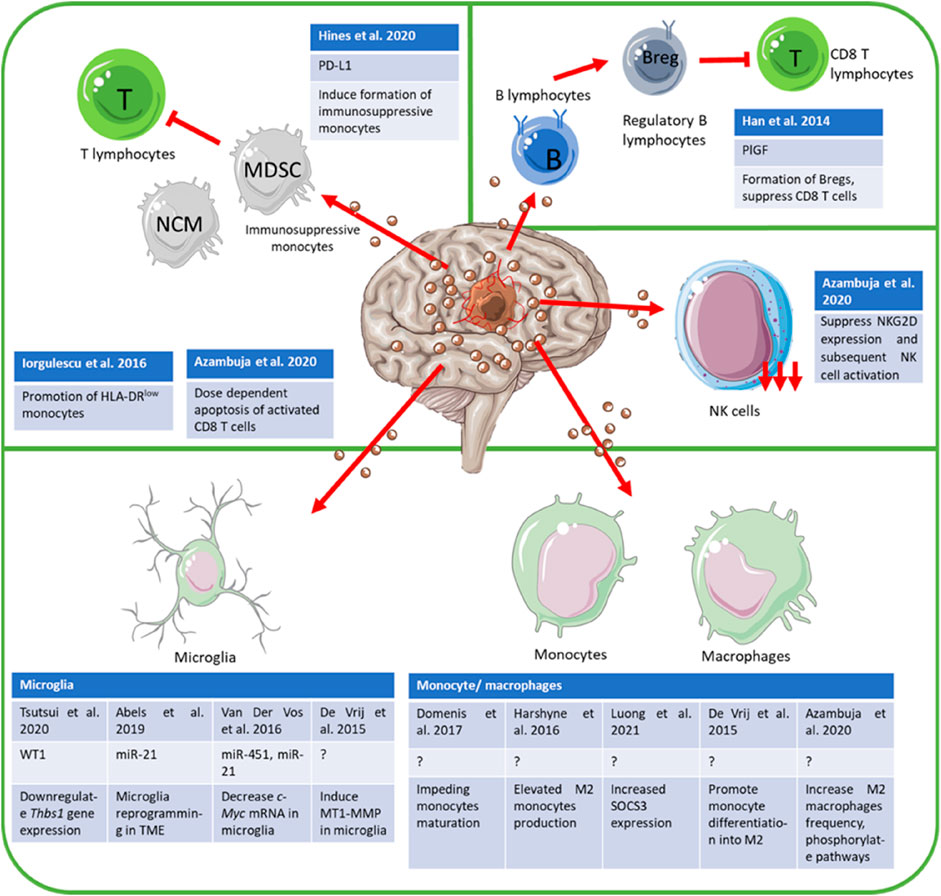
FIGURE 1. Schematic description of GB-derived EVs immunomodulation. GB performs immunosuppression by interfering with the expression of regulatory proteins via either expression of miRNAs or other proteins. Other than that, GM also impedes the maturation of immune cells such as monocytes, in addition to promoting the expression of immunosuppressive phenotypes in immune cells while downregulating the presence of immune-promoting phenotypes. Other immunosuppressive mechanisms employed included dose-dependent apoptosis of activated immune cells.
EV is also instrumental in the metabolism modulation within the GB TME. Tumor-activated stromal cells (TASC), also known as cancer-associated fibroblasts have been determined to transfer mitochondria to primary GB cells via various mechanisms including EVs (Salaud et al., 2020). This led to increased glycolysis which translates to better GB proliferation (Salaud et al., 2020). Under favorable metabolic conditions, GB cells also utilize EVs as a vehicle to reduce intracellular miRNA content. This is evident when GB cells secrete exosomal miR-451, which is >40-fold more abundant in EVs compared to in cells (Van Der Vos et al., 2016). Although said EVs were absorbed by microglia, the increased release of exosomal miR-451 from GB cells coincides with a GB cell self-preservation mechanism when glucose is limited. Godlewski et al. (Godlewski et al., 2010) demonstrated when glucose is limited, miR-451 levels in the cell decline, leading to heightened CAB39 expression, activation of AMPK and consequently cell survival by reducing cell proliferation. GB-derived EVs are also capable of modulating other cells in the CNS to facilitate the TME. A study by Oushy et al. (Oushy et al., 2018) found that GB EV-treated normal human astrocytes (NHA) displayed enhanced migration ability and cytokine production, which are tumor-promoting phenotypes favored by GBM cells. The EV-mediated NHA drive to tumor-supporting phenotype was further elucidated by Hallal et al. (Hallal et al., 2019) when it was postulated that EV-treated NHA displayed a senescence-associated secretory profile (SASP) along with enhanced migration capabilities through enhanced podosome and gelatin matrix degradation. EV-mediated cell modulation in the GBM TME also occurs in immune cells, where tumor-associated macrophages (TAMs) are found to secrete CHD7-targeting microRNAs to glioma stem cells (GSCs) to trigger a proneural-to-mesenchymal transition (PMT) (Zhang et al., 2020). As PMT confers resistance to therapy, this contributes to worsening diagnosis in recurrent GBM (Fedele et al., 2019). Other than that, GSCs-derived EVs also induce the growth of brain endothelial cells (BEC), further enhancing their survival (Spinelli et al., 2018) which may be beneficial to the GBM TME. A study comparing glioma-derived human ECs (GhECs) and normal human ECs (NhECs) determined that GhEC-EV significantly induced LN229 GB cell line migration in vitro via MYO1C transfer to recipient cells (Tian et al., 2020). A study by Lucero et al. (Lucero et al., 2020) also discovered that GSC-derived EVs carry vasculature-associated miRNAs that reprogram brain EC to perform angiogenesis. EC is also vital for the formation of the blood-brain barrier (Kadry et al., 2020) yet a study by Treps et al. (Treps et al., 2016) discovered that Semaphorin3A expressed on GBM EVs’ surface causes increased vascular permeability in vivo, which can jeopardize the integrity of the BBB. GSC also secrete VEGF-A, a known angiogenic and permeability factor in the exosomal form to human BECs (Treps et al., 2017).
Immunotherapy-based strategies for GB have shown some preclinical successes that were not translated into Phase 3 clinical efficacies. (Yu and Quail, 2021). Therefore, looking into the TME might provide clues to targeting this issue. A paper by Ali et al. (Ali et al., 2021) showed that TME-targeting treatments including combination strategies led to a variety of post-treatment GB TME. GB cells have been shown to utilize extracellular vesicles (EVs) to circumvent immunotherapies and radiation therapy. For instance, U87 glioma cell line treated with bevacizumab has been shown to secrete out EVs containing bevacizumab on the surface (Simon et al., 2018) albeit with a slight influence on cell viability and proliferation in clinical dosages. Hypoxic glioma cells also secrete exosomal miR-301a that promotes resistance to radiotherapy (Yue et al., 2019). In terms of metabolism, the post-irradiation brain allows a tumor-tolerant microenvironment via metabolic shift with high production of energy carriers and low production of antioxidants (Gupta et al., 2020), which might also be translated into extracellular vesicles as well. This has been documented when GB cell lines of multiple subtypes exposed to either acute or chronic irradiation also exhibit metabolic changes that translate into the alteration of microvesicle cargo, ultimately modulating said vesicles’ paracrine signaling towards untreated glioma cells (Baulch et al., 2016).
Radiation therapy and chemotherapy might also cause glioma cells to be more malignant. U87MG cells exposed to radiation also secrete more exosomes with enhanced cell migration capability due to an increased abundance of cell motility-related mRNA and proteins (Arscott et al., 2013). Pavlyukov et al. (Pavlyukov et al., 2018) also demonstrated apoptotic cell-derived EVs (apoEVs) from irradiated or chemo-treated glioma cells promote malignancy by phenotypic changes induced by splicing factor transfer to recipient glioma cells. A study by Ramakrishnan et al. (Ramakrishnan et al., 2020) also demonstrated that irradiation causes glioma cells to adopt a stem cell state by EV-mediated release of miR-603, causing resistance to ionizing radiation and DNA alkylating drugs. In terms of chemotherapy, temozolomide administered to glioma stem cells also induce secretion of GSC-derived EVs that are enriched with cell adhesion proteins which might aid in tumor progression (Andre-Gregoire et al., 2018). TMZ-induced EV cargo change is also reported by Garnier et al. (Garnier et al., 2018) when they determined that EVs secreted by GB cell lines contained TMZ resistance transcripts. A comprehensive study by Cuperlovic-Culf et al. (Cuperlovic-Culf et al., 2020) on GB cell line-derived EVs also showed that metabolome cargoes are implicated in immune response and metabolism amid being varied depending on the cell line.
T cells’ secretion of EVs is documented to be upregulated post T cell receptor (TCR) triggering, which possibly mediates surface TCR/CD3-mediated cell homing (Blanchard et al., 2002). Other than that, overactive T cells are also capable of secreting Fas ligand (FasL) and Apo2 ligand (APO2L) via EV release, suggesting EV-mediated autocrine or paracrine immune regulation (Monleón et al., 2001). Delivery of specific cargo to recipient cells is also evident with mechanisms including sequence motifs-dependent microRNA localization into exosomes (Villarroya-Beltri et al., 2013) and monophosphorylation-dependent FasL sorting into secreted lysosomes (Zuccato et al., 2007). Nevertheless, death ligands need to be bound on the membrane to crosslink efficiently with their corresponding death receptors (Anel et al., 2019). In general, T cells EVs deliver cargoes such as enzymes, transmembrane proteins, members of the immunoglobulins (Ig) superfamily, and MHC molecules to target cells such as APCs (Choudhuri et al., 2014; Mittelbrunn et al., 2011; Nolte-'t Hoen et al., 2004) and B cells (Yang et al., 2019). Multiple responses in target cells were recorded, including activation-induced cell death (AICD) where microvesicles containing FasL and APO2L were released shortly before cell apoptosis (Martínez-Lorenzo et al., 1999). Positive and negative regulation of T cell responses were also recorded. T cell-derived EVs carry surface receptors and molecules to APCs, causing transcellular signaling and modulation of APCs (Choudhuri et al., 2014; Nolte-'t Hoen et al., 2004). T cell-derived exosomes are also capable of transporting miRNA to APCs, causing changes to recipient cells’ gene expressions (Mittelbrunn et al., 2011).
Tregs, owning to their immunosuppressive traits, also secrete exosomes that perform similar functions. Immunosuppression mediated by Tregs involves several different mechanisms, including adenosine (Ado) mediated immunomodulation (Smyth et al., 2013; Schuler et al., 2014), cyclooxygenase-2 (Cox-2) mediated regulation of interferon-gamma (IFNγ) secretion in T effector cells (Okoye et al., 2014), target cell cycle arrest and apoptosis, conversion of T cells into Tregs (Aiello et al., 2017), induction of tolerogenic dendritic cells (DCs) (Tung et al., 2018), and increased resistance to apoptosis by Tregs cells (Czystowska et al., 2010). GB cell line-derived exosomes contain inhibitory proteins such as CD39, FasL, CTLA-4, TRAIL, and CD73 that attenuate the normal function of all immune cells (Azambuja et al., 2020). GB has been documented to release PD-L1+ EVs which inhibit T cell proliferation via myeloid-derived suppressor cells (MDSC) and nonclassical monocytes (NCM) instead of direct T cell inhibition (Himes et al., 2020). The mechanism of action is in contrast to findings by Ricklefs et al. who argue GB-derived EVs block TCR-mediated T cell activation (Ricklefs et al., 2018), although Hines et al. do point out the possible discrepancy with T cell stimulation employed by both researchers causing contrasting results regarding GB EVs role (Himes et al., 2020). GB-derived exosomes have limited ability to activate CD8+ T lymphocytes (Iorgulescu et al., 2016) yet it has been reported that the expression of CD86 on glioma cell lines can be bound competitively by cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), leading to possible T cell immunosuppression (Walker and Sansom, 2011). Therefore, more research is needed to elucidate the GB-derived EV effect on T cells. The crosstalk between EVs and T cells, and other immune cells are tabulated in Table 1.
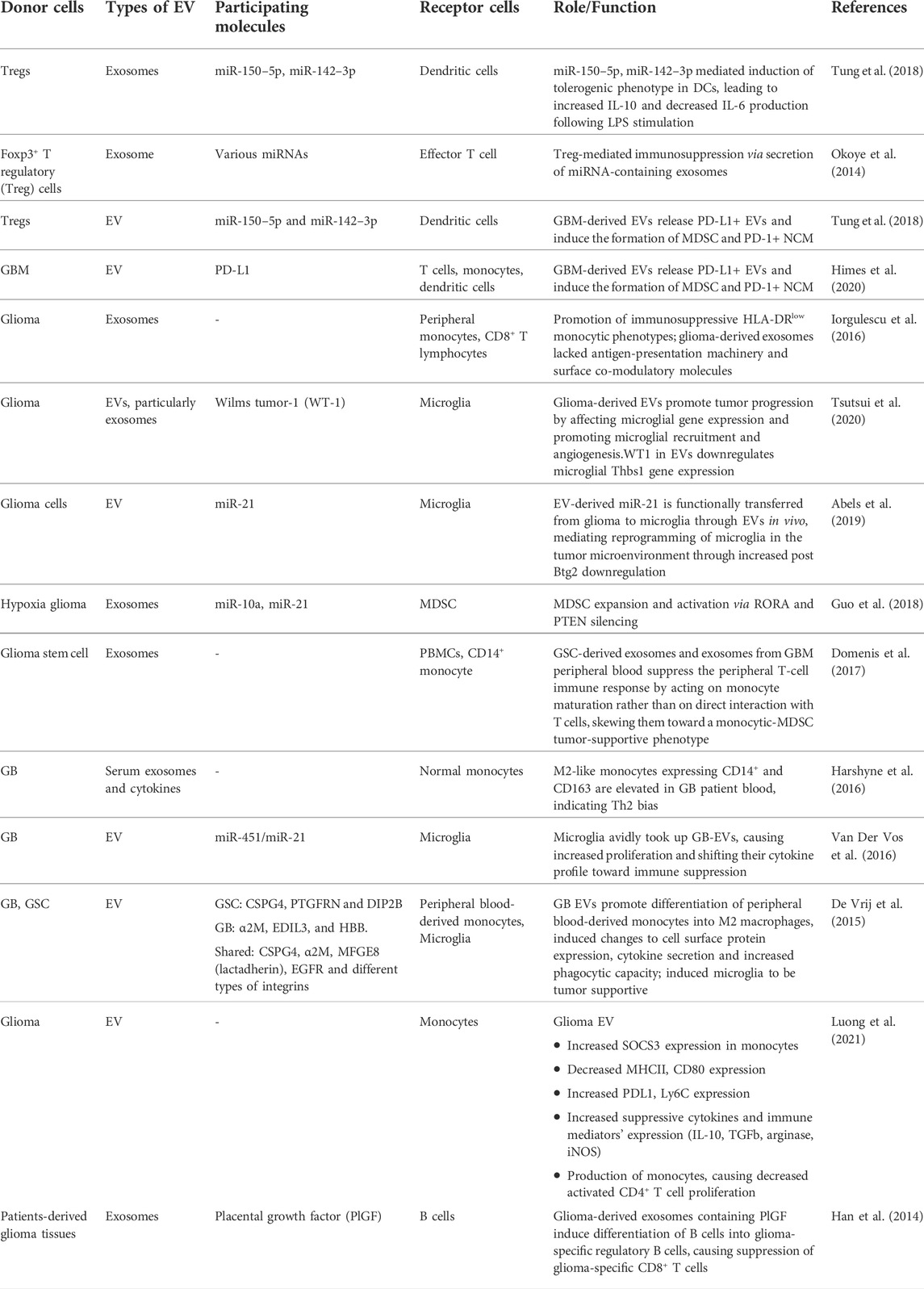
TABLE 1. Examples of immune cells -glioblastoma EV crosstalk.
Given myeloid cells’ extensive presence in the brain, gliomas’ interaction with them is considerably more frequent, where up to 30% of total glioma/glioblastoma mass is comprised of myeloid cells (Arcuri et al., 2017). Gliomas utilize this concentration of myeloid cells to their advantage by secreting EVs capable of modulating microglia. The WT-1 protein is found secreted in glioma-derived EVs and downregulates thrombospondin-1 (Thbs1) in microglia, subsequently promoting angiogenesis which is vital in glioma progression (Wagner et al., 2014; Tsutsui et al., 2020). Other than that, microglia proliferation is also promoted via downregulation of BTG anti-proliferation factor 2 (Btg2) expression in microglia post-delivery of miR-21 through EVs (Abels et al., 2019).
Glioma cells also exhibit EV-mediated malignancy under hypoxic conditions. Under hypoxic conditions, glioma cells secrete exosomes containing TERF2 interacting protein (TERF2IP) targeting-miR-1246, causing activation and inhibition of STAT3 and Nuclear factor-κB (NF-κB) signaling pathways respectively. This leads to the polarization of macrophages from M1 to immunosuppressive M2 phenotype (Qian et al., 2020). Glioma cells under hypoxic conditions have also been found to secrete exosomes containing miR-10a and miR-21 to MDSCs, causing its expansion and activation (Guo et al., 2018), which further exacerbates cancer progression via inhibition of immune cell functions, Tregs expansion and promotion of immunosuppressive regulatory B cells (Bregs) (Mi et al., 2020). Other than that, GB affects T-cell immune response through the modulation of monocytes. Systemic T cell suppression via glioma stem cells (GSCs) derived exosomes occurs with internalization by CD14+ monocyte which causes stunted maturation and formation of monocytic MDSCs, subsequently disrupting CD3+ and CD4+ T cell activation (Domenis et al., 2017). GB EVs also induces the proliferation of NCMs and MDSCs, eventually inhibiting T cell proliferation (Himes et al., 2020).
GB is also able to influence the peripheral immune environment via exosomes as M2-like monocytes expressing CD163 and CD14 are highly expressed in the GB patients’ peripheral blood, representing a Th2 bias (Harshyne et al., 2016). GB tumors also secrete miR-451 and miR-21 in EVs, which when internalized by microglia, cause an increase in microglia proliferation and a cytokine shift favoring immune suppression (Van Der Vos et al., 2016). GB EVs also contain leukocyte migration and focal adhesion-specific proteins that skewed peripheral monocytes’ differentiation towards M2 macrophages in addition to modification to macrophages’ cell surface protein expression, cytokine secretion, and phagocytic effect (De Vrij et al., 2015). De Vrij et al. also mentioned human microglia exhibit high expression of membrane-type 1-matrix metalloproteinase (MT1-MMP) post-incubation with GB EVs, which supports tumor growth, denoting the tumor proliferation aspect of cancer-associated immune cells in addition to their immunoregulatory role (De Vrij et al., 2015). Luong et al. (Luong et al., 2021) also determined that glioma-derived EVs perform several pro-tumorigenic functions in monocytes such as upregulated expression of suppressive cytokines, proteins, PD-L1 and lymphocyte antigen six complex (Ly6C), downregulation of proinflammatory cytokines, MHC II and costimulatory CD80 expression as well as the conversion of monocytes into suppressive cells involved in inhibition of activated CD4+ T cells (Luong et al., 2021). IFN-γ stimulation of GB cells also causes superinduction of GB-derived, immunosuppressive IDO-1 and PD-L1 expressing EVs, which led to immunosuppression of monocytes on top of differentiation of monocytes into immunosuppressive MDSCs and NCMs (Jung et al., 2022).
NK cells are also known to secrete extracellular vesicles containing a variety of biomolecules, including membrane and extracellular matrix (ECM) proteins (tetraspanins, integrins), death receptor ligands, cytolytic enzymes, and miRNA that translate into cytotoxic effect in cancer cells such as apoptosis while avoiding damage to normal PBMCs (Farcas and Inngjerdingen, 2020). Other than that, NK extracellular vesicles (NKEVs) cause immunomodulation in PBMCs including higher expression of HLA-DR and costimulatory molecules on monocytes, CD25 on T cells, and CD56 on NK cells, which translates into both pro-inflammation and anti-inflammation spectrum of the immune response (Federici et al., 2020). NK exosomes also contain FasL, Natural Killer Group 2D (NKG2D), and perforin molecules, which perform an individualistic function: perforin mediates tumor and activated immune cell death in a time and dose-dependent manner, while Fas is suggested to be involved in lymphocyte homeostasis regulation (Lugini et al., 2012). NKG2D, typically known as an activating receptor by cytolytic lymphoid cells (Pende et al., 2001), typically can induce a cytolytic effect without more specific natural cytotoxicity NK receptors, yet their functional role in exosomes is not known. Activated NKEVs induced dose-dependent caspase-mediated apoptosis in neuroblastoma via functional perforin, granulysin, and granzymes A and B mediation of the caspase pathway (Jong et al., 2017) which possibly translates into a similar result in GB. As per other immune cells, tumor-derived MVs secreted under hypoxic conditions are more potent in impairing NK cytotoxicity and cell function as compared to normoxic equivalent (Berchem et al., 2016) due to the delivery of CD107a targeting-miR23a and TGF-β (Alter et al., 2004; Viel et al., 2016). NK EVs are also multifaceted in immunomodulation. Azambuja et al. mentioned that GB-derived exosomes suppress NK cell activation by suppressing NKG2D expression levels (Azambuja et al., 2020). In contrast to this, NK cells also secrete cytotoxic EVs under the influence of pro-inflammatory cytokines (Enomoto et al., 2021) with IL-15 and IL-21 both playing significant roles in NK cell activation (Carson et al., 1994; Skak et al., 2008). All in all, studies have shown that NK cell activity can be modulated by tumor-derived EVs, including those of GB origin, with the effect dependent on external stimuli such as the balance between immune promoting- and inhibiting-signals, further confirming the notion that NK cell function is niche dependent.
B cell’s secretion of EVs has been documented with the release of MHC II-containing exosomes (Raposo et al., 1996), which it was suggested to involve in antigen presentation (Lindenbergh and Stoorvogel, 2018). Muntasell et al. reported on upregulated B cell-derived exosome carrying antigenic-peptide MHC II (pMHC II) by the antigen-specific CD4+ T cell, which can poise as a positive modulator for ongoing immune response and maintenance of antigenic memory in T cells (Muntasell et al., 2007). Given so, B cells exosomes are also implicated in anti-inflammatory responses such as the possible transfer of membrane-tethered CD73 to Tregs in the peripheral causing an increase in anti-inflammatory adenosine secretion (Schuler et al., 2014). A more recent paper by Zhang et al. also demonstrated that CD19+ EVs secreted from B cells induce hydrolysis of adenosine triphosphate (ATP) to adenosine via incorporated CD39 and CD73 action, thus inhibiting CD8+ T cell proliferation and subsequent reduction in chemotherapy efficacy (Zhang et al., 2019). Although both studies do not employ GB as their model, the GB tumor microenvironment is known to be hypoxic (Brown et al., 2018; Tomaszewski et al., 2019), and CD73 is found to be upregulated in both CNS (Kulesskaya et al., 2013) and hypoxic conditions (Li et al., 2006). B cells in the CNS might also participate in the upregulation of adenosine, especially when Zhang et al. also mentioned that hypoxia-inducible factor-1α (HIF-1α) mediated Rab27a expression causes heightened EV secretion by B cells (Zhang et al., 2019). Glioma cells also participated in B cell modulation. Glioma cells secrete placental growth factor (PlGF) in exosomal form, which in contact with naïve B cells can induce their differentiation to Bregs (Han et al., 2014). Induced Bregs suppress granzyme B and perforin secretion by the glioma-specific CD8+ T cell, denoting specificity in B cell action against glioma. However, relevant publications regarding B cells exosome against glioma are still lacking.
In conclusion, EVs play vital roles in both faces of the immunopathological aspects of GB. EVs typically reflect the physiological state of the donor cell and perform specific effector functions in recipient cells, thus modulating the recipient cells to have an abnormal phenotype. This plays well into the induction of the TME, which in the GB context involves the relationship between GB tumors and neighboring cells, promoting immunosuppression and proliferation within the brain. How the common cell adjusts its metabolic requirement to fit into the objective of the tumor microenvironment is an interesting avenue to research, especially with the usage of EVs, which can be a vehicle for both paracrine and endocrine signaling in the first place. More thorough EV research across all facets of cell biochemistry is needed to synthesize, elucidate, and magnify possible influential pathways in the cell, where such results can be translated into more optimized and effective screening and treatment strategies in the future.
NA wrote the initial draft, JW wrote the manuscript, NA, SS and NJ edited the manuscript.
JW was funded by the HiCoE grant JJ-2021–003. Parts of the figure were produced using pictures “Monocytes” from Servier Medical Art, provided by Servier, licensed under a Creative Commons Attribution 3.0 unported license.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abels, E. R., Maas, S. L. N., Nieland, L., Wei, Z., Cheah, P. S., Tai, E., et al. (2019). Glioblastoma-associated microglia reprogramming is mediated by functional transfer of extracellular miR-21. Cell Rep. 28, 3105–3119. doi:10.1016/j.celrep.2019.08.036
Achrol, A. S., Rennert, R. C., Anders, C., Soffietti, R., Ahluwalia, M. S., Nayak, L., et al. (2019). Brain metastases. Nat. Rev. Dis. Prim. 5, 5. doi:10.1038/s41572-018-0055-y
Aiello, S., Rocchetta, F., Longaretti, L., Faravelli, S., Todeschini, M., Cassis, L., et al. (2017). Extracellular vesicles derived from T regulatory cells suppress T cell proliferation and prolong allograft survival. Sci. Rep. 7, 11518–11519. doi:10.1038/s41598-017-08617-3
Ali, S., Borin, T. F., Piranlioglu, R., Ara, R., Lebedyeva, I., Angara, K., et al. (2021). Changes in the tumor microenvironment and outcome for TME-targeting therapy in glioblastoma: A pilot study. PLoS One 16, e0246646. doi:10.1371/journal.pone.0246646
Alter, G., Malenfant, J. M., and Altfeld, M. (2004). CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 294, 15–22. doi:10.1016/j.jim.2004.08.008
Andre-Gregoire, G., Bidere, N., and Gavard, J. (2018). Temozolomide affects extracellular vesicles released by glioblastoma cells. Biochimie 155, 11–15. doi:10.1016/j.biochi.2018.02.007
Anel, A., Gallego-Lleyda, A., de Miguel, D., Naval, J., and Martínez-Lostao, L. (2019). Role of exosomes in the regulation of T-cell mediated immune responses and in autoimmune disease. Cells 8, 154. doi:10.3390/cells8020154
Arcuri, C., Fioretti, B., Bianchi, R., Mecca, C., Tubaro, C., Beccari, T., et al. (2017). Microglia-glioma cross-talk: A two way approach to new strategies against glioma. Front. Biosci. 22, 268–309. doi:10.2741/4486
Arscott, W. T., Tandle, A. T., Zhao, S., Shabason, J. E., Gordon, I. K., Schlaff, C. D., et al. (2013). Ionizing radiation and glioblastoma exosomes: implications in tumor biology and cell migration. Transl. Oncol. 6, 638–648. doi:10.1593/tlo.13640
Azambuja, J. H., Ludwig, N., Yerneni, S., Rao, A., Braganhol, E., and Whiteside, T. L. (2020). Molecular profiles and immunomodulatory activities of glioblastoma-derived exosomes. Neurooncol. Adv. 2, vdaa056–11. doi:10.1093/noajnl/vdaa056
Balss, J., Meyer, J., Mueller, W., Korshunov, A., Hartmann, C., and von Deimling, A. (2008). Analysis of the IDH1 codon 132 mutation in brain tumors. Acta Neuropathol. 116, 597–602. doi:10.1007/s00401-008-0455-2
Baulch, J. E., Geidzinski, E., Tran, K. K., Yu, L., Zhou, Y. H., and Limoli, C. L. (2016). Irradiation of primary human gliomas triggers dynamic and aggressive survival responses involving microvesicle signaling. Environ. Mol. Mutagen. 57, 405–415. doi:10.1002/em.21988
Berchem, G., Noman, M. Z., Bosseler, M., Paggetti, J., Baconnais, S., Le cam, E., et al. (2016). Hypoxic tumor-derived microvesicles negatively regulate NK cell function by a mechanism involving TGF-β and miR23a transfer. OncoImmunology 5, e1062968. doi:10.1080/2162402X.2015.1062968
Bissig, C., and Gruenberg, J. (2013). Lipid sorting and multivesicular endosome biogenesis. Cold Spring Harb. Perspect. Biol. 5, a016816. doi:10.1101/cshperspect.a016816
Blanchard, N., Lankar, D., Faure, F., Regnault, A., Dumont, C., Raposo, G., et al. (2002). TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/zeta complex. J. Immunol. 168, 3235–3241. doi:10.4049/jimmunol.168.7.3235
Broekman, M. L., Maas, S. L. N. N., Abels, E. R., Mempel, T. R., Krichevsky, A. M., and Breakefield, X. O. (2018). Multidimensional communication in the microenvirons of glioblastoma. Nat. Rev. Neurol. 14, 482–495. doi:10.1038/s41582-018-0025-8
Brown, N. F., Carter, T. J., Ottaviani, D., and Mulholland, P. (2018). Harnessing the immune system in glioblastoma. Br. J. Cancer 119, 1171–1181. doi:10.1038/s41416-018-0258-8
Cagney, D. N., Martin, A. M., Catalano, P. J., Redig, A. J., Lin, N. U., Lee, E. Q., et al. (2017). Incidence and prognosis of patients with brain metastases at diagnosis of systemic malignancy: A population-based study. Neuro. Oncol. 19, 1511–1521. doi:10.1093/neuonc/nox077
Carson, W. E., Giri, J. G., Lindemann, M. J., Linett, M. L., Ahdieh, M., Paxton, R., et al. (1994). Interleukin (IL) 15 is a novel cytokine that activates human natural killer cells via components of the IL-2 receptor. J. Exp. Med. 180, 1395–1403. doi:10.1084/jem.180.4.1395
Choudhuri, K., Llodrá, J., Roth, E. W., Tsai, J., Gordo, S., Wucherpfennig, K. W., et al. (2014). Polarized release of T-cell-receptor-enriched microvesicles at the immunological synapse. Nature 507, 118–123. doi:10.1038/nature12951
Clancy, J. W., Zhang, Y., Sheehan, C., and D’Souza-Schorey, C. (2019). An ARF6–Exportin-5 axis delivers pre-miRNA cargo to tumour microvesicles. Nat. Cell Biol. 21, 856–866. doi:10.1038/s41556-019-0345-y
Cuperlovic-Culf, M., Khieu, N. H., Surendra, A., Hewitt, M., Charlebois, C., and Sandhu, J. K. (2020). Analysis and simulation of glioblastoma cell lines-derived extracellular vesicles metabolome. Metabolites 10, E88. doi:10.3390/metabo10030088
Czystowska, M., Strauss, L., Bergmann, C., Szajnik, M., Rabinowich, H., and Whiteside, T. L. (2010). Reciprocal granzyme/perforin-mediated death of human regulatory and responder T cells is regulated by interleukin-2 (IL-2). J. Mol. Med. 88, 577–588. doi:10.1007/s00109-010-0602-9
Davis, F. G., Dolecek, T. A., McCarthy, B. J., and Villano, J. L. (2012). Toward determining the lifetime occurrence of metastatic brain tumors estimated from 2007 United States cancer incidence data. Neuro. Oncol. 14, 1171–1177. doi:10.1093/neuonc/nos152
De Vrij, J., Niek Maas, S. L., Kwappenberg, K. M. C., Schnoor, R., Kleijn, A., Dekker, L., et al. (2015). Glioblastoma-derived extracellular vesicles modify the phenotype of monocytic cells. Int. J. Cancer 137, 1630–1642. doi:10.1002/ijc.29521
Domenis, R., Cesselli, D., Toffoletto, B., Bourkoula, E., Caponnetto, F., Manini, I., et al. (2017). Systemic T cells immunosuppression of glioma stem cell-derived exosomes is mediated by monocytic myeloid-derived suppressor cells. PLoS ONE 12, e0169932. doi:10.1371/journal.pone.0169932
Enomoto, Y., Li, P., Jenkins, L. M., Anastasakis, D., Lyons, G. C., Hafner, M., et al. (2021). Cytokine-enhanced cytolytic activity of exosomes from NK Cells. Cancer Gene Ther. 3, 734–749. doi:10.1038/s41417-021-00352-2
Fabry, Z., Schreiber, H. A., Harris, M. G., and Sandor, M. (2008). Sensing the microenvironment of the central nervous system: immune cells in the central nervous system and their pharmacological manipulation. Curr. Opin. Pharmacol. 8, 496–507. doi:10.1016/j.coph.2008.07.009
Farcas, M., and Inngjerdingen, M. (2020). Natural killer cell–derived extracellular vesicles in cancer therapy. Scand. J. Immunol. 92, 129388–7. doi:10.1111/sji.12938
Fedele, M., Cerchia, L., Pegoraro, S., Sgarra, R., and Manfioletti, G. (2019). Proneural-mesenchymal transition: Phenotypic plasticity to acquire multitherapy resistance in glioblastoma. Int. J. Mol. Sci. 20, E2746. doi:10.3390/ijms20112746
Federici, C., Shahaj, E., Cecchetti, S., Camerini, S., Casella, M., Iessi, E., et al. (2020). Natural-killer-derived extracellular vesicles: Immune sensors and interactors. Front. Immunol. 11, 262–324. doi:10.3389/fimmu.2020.00262
Gao, X., Zhang, Z., Mashimo, T., Shen, B., Nyagilo, J., Wang, H., et al. (2020). Gliomas interact with non-glioma brain cells via extracellular vesicles. Cell Rep. 30, 2489–2500. doi:10.1016/j.celrep.2020.01.089
García-Romero, N., Carrión-Navarro, J., Esteban-Rubio, S., Lázaro-Ibáñez, E., Peris-Celda, M., Alonso, M. M., et al. (2017). DNA sequences within glioma-derived extracellular vesicles can cross the intact blood-brain barrier and be detected in peripheral blood of patients. Oncotarget 8, 1416–1428. doi:10.18632/oncotarget.13635
Garnier, D., Meehan, B., Kislinger, T., Daniel, P., Sinha, A., Abdulkarim, B., et al. (2018). Divergent evolution of temozolomide resistance in glioblastoma stem cells is reflected in extracellular vesicles and coupled with radiosensitization. Neuro. Oncol. 20, 236–248. doi:10.1093/neuonc/nox142
Godlewski, J., Nowicki, M. O., Bronisz, A., Nuovo, G., Palatini, J., De Lay, M., et al. (2010). MicroRNA-451 regulates LKB1/AMPK signaling and allows adaptation to metabolic stress in glioma cells. Mol. Cell 37, 620–632. doi:10.1016/j.molcel.2010.02.018
Guo, X., Qiu, W., Liu, Q., Qian, M., Wang, S., Zhang, Z., et al. (2018). Immunosuppressive effects of hypoxia-induced glioma exosomes through myeloid-derived suppressor cells via the miR-10a/Rora and miR-21/Pten Pathways. Oncogene 37, 4239–4259. doi:10.1038/s41388-018-0261-9
Gupta, K., Vuckovic, I., Zhang, S., Xiong, Y., Carlson, B. L., Jacobs, J., et al. (2020). Radiation induced metabolic alterations associate with tumor aggressiveness and poor outcome in glioblastoma. Front. Oncol. 10, 535. doi:10.3389/fonc.2020.00535
Hallal, S., Mallawaaratchy, D. M., Wei, H., Ebrahimkhani, S., Stringer, B. W., Day, B. W., et al. (2019). Extracellular vesicles released by glioblastoma cells stimulate normal astrocytes to acquire a tumor-supportive phenotype via p53 and MYC signaling pathways. Mol. Neurobiol. 56, 4566–4581. doi:10.1007/s12035-018-1385-1
Han, S., Feng, S., Ren, M., Ma, E., Wang, X., Xu, L., et al. (2014). Glioma cell-derived placental growth factor induces regulatory B cells. Int. J. Biochem. Cell Biol. 57, 63–68. doi:10.1016/j.biocel.2014.10.005
Hao, C., Parney, I. F., Roa, W. H., Turner, J., Petruk, K. C., and Ramsay, D. A. (2002). Cytokine and cytokine receptor mRNA expression in human glioblastomas: Evidence of Th1, Th2 and Th3 cytokine dysregulation. Acta Neuropathol. 103, 171–178. doi:10.1007/s004010100448
Harshyne, L. A., Nasca, B. J., Kenyon, L. C., Andrews, D. W., and Hooper, D. C. (2016). Serum exosomes and cytokines promote a T-helper cell type 2 environment in the peripheral blood of glioblastoma patients. Neuro. Oncol. 18, 206–215. doi:10.1093/neuonc/nov107
Himes, B. T., Peterson, T. E., de Mooij, T., Garcia, L. M. C., Jung, M. Y., Uhm, S., et al. (2020). The role of extracellular vesicles and PD-L1 in glioblastoma-mediated immunosuppressive monocyte induction. Neuro. Oncol. 22, 967–978. doi:10.1093/neuonc/noaa029
Hurwitz, S. N., Rider, M. A., Bundy, J. L., Liu, X., Singh, R. K., and Meckes, D. G. (2016). Proteomic profiling of NCI-60 extracellular vesicles uncovers common protein cargo and cancer type-specific biomarkers. Oncotarget 7, 86999–87015. doi:10.18632/oncotarget.13569
Iorgulescu, J. B., Ivan, M. E., Safaee, M., and Parsa, A. T. (2016). The limited capacity of malignant glioma-derived exosomes to suppress peripheral immune effectors. J. Neuroimmunol. 290, 103–108. doi:10.1016/j.jneuroim.2015.11.025
Jackson, C. M., Choi, J., and Lim, M. (2019). Mechanisms of immunotherapy resistance: Lessons from glioblastoma. Nat. Immunol. 20, 1100–1109. doi:10.1038/s41590-019-0433-y
Jong, A. Y., Wu, C. H., Li, J., Sun, J., Fabbri, M., Wayne, A. S., et al. (2017). Large-scale isolation and cytotoxicity of extracellular vesicles derived from activated human natural killer cells. J. Extracell. Vesicles 6, 1294368. doi:10.1080/20013078.2017.1294368
Jung, M. Y., Aibaidula, A., Brown, D. A., Himes, B. T., Cumba Garcia, L. M., and Parney, I. F. (2022). Superinduction of immunosuppressive glioblastoma extracellular vesicles by IFN-gamma through PD-L1 and IDO1. Neurooncol. Adv. 4, vdac017. doi:10.1093/noajnl/vdac017
Kadry, H., Noorani, B., and Cucullo, L. (2020). A blood-brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS 17, 69. doi:10.1186/s12987-020-00230-3
Kucharzewska, P., Christianson, H. C., Welch, J. E., Svensson, K. J., Fredlund, E., Ringnér, M., et al. (2013). Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. Proc. Natl. Acad. Sci. U. S. A. 110, 7312–7317. doi:10.1073/pnas.1220998110
Kulesskaya, N., Võikar, V., Peltola, M., Yegutkin, G. G., Salmi, M., Jalkanen, S., et al. (2013). CD73 is a major regulator of adenosinergic signalling in mouse brain. PLoS ONE 8, e66896. doi:10.1371/journal.pone.0066896
Lapointe, S., Perry, A., and Butowski, N. A. (2018). Primary brain tumours in adults. Lancet 392, 432–446. doi:10.1016/S0140-6736(18)30990-5
Li, X., Zhou, T., Zhi, X., Zhao, F., Yin, L., and Zhou, P. (2006). Effect of hypoxia/reoxygenation on CD73 (ecto-5′-nucleotidase) in mouse microvessel endothelial cell lines. Microvasc. Res. 72, 48–53. doi:10.1016/j.mvr.2006.04.005
Lim, M., Xia, Y., Bettegowda, C., and Weller, M. (2018). Current state of immunotherapy for glioblastoma. Nat. Rev. Clin. Oncol. 15, 422–442. doi:10.1038/s41571-018-0003-5
Lindenbergh, M. F. S., and Stoorvogel, W. (2018). Antigen presentation by extracellular vesicles from professional antigen-presenting cells. Annu. Rev. Immunol. 36, 435–459. doi:10.1146/annurev-immunol-041015-055700
Louis, D. N., Perry, A., Wesseling, P., Brat, D. J., Cree, I. A., Figarella-Branger, D., et al. (2021). The 2021 WHO classification of tumors of the central nervous system: a summary. Neuro. Oncol. 23, 1231–1251. doi:10.1093/neuonc/noab106
Lucero, R., Zappulli, V., Sammarco, A., Murillo, O. D., Cheah, P. S., Srinivasan, S., et al. (2020). Glioma-derived miRNA-containing extracellular vesicles induce angiogenesis by reprogramming brain endothelial cells. Cell Rep. 30, 2065–2074. doi:10.1016/j.celrep.2020.01.073
Lugini, L., Cecchetti, S., Huber, V., Luciani, F., Macchia, G., Spadaro, F., et al. (2012). Immune surveillance properties of human NK cell-derived exosomes. J. Immunol. 189, 2833–2842. doi:10.4049/jimmunol.1101988
Luong, N., Lenz, J. A., Modiano, J. F., and Olson, J. K. (2021). Extracellular vesicles secreted by tumor cells promote the generation of suppressive monocytes. ImmunoHorizons 5, 647–658. doi:10.4049/immunohorizons.2000017
Martínez-Lorenzo, M. J., Anel, A., Gamen, S., Monle n, I., Lasierra, P., Larrad, L., et al. (1999). Activated human T cells release bioactive Fas ligand and APO2 ligand in microvesicles. J. Immunol. Baltim. Md, 1950) 163, 1274–1281.
Mi, Y., Guo, N., Luan, J., Cheng, J., Hu, Z., Jiang, P., et al. (2020). The emerging role of myeloid-derived suppressor cells in the glioma immune suppressive microenvironment. Front. Immunol. 11, 737–811. doi:10.3389/fimmu.2020.00737
Miller, K. D., Ostrom, Q. T., Kruchko, C., Patil, N., Tihan, T., Cioffi, G., et al. (2021). Brain and other central nervous system tumor statistics, 2021. CA A Cancer J. Clin. 71, 381–406. doi:10.3322/caac.21693
Mittelbrunn, M., Gutiérrez-Vázquez, C., Villarroya-Beltri, C., González, S., Sánchez-Cabo, F., González, M. Á., et al. (2011). Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2, 282. doi:10.1038/ncomms1285
Monleón, I., Martínez-Lorenzo, M. J., Monteagudo, L., Lasierra, P., Taulés, M., Iturralde, M., et al. (2001). Differential secretion of Fas ligand- or APO2 ligand/TNF-related apoptosis-inducing ligand-carrying microvesicles during activation-induced death of human T cells. J. Immunol. 167, 6736–6744. doi:10.4049/jimmunol.167.12.6736
Moreno-Gonzalo, O., Fernandez-Delgado, I., and Sanchez-Madrid, F. (2018). Post-translational add-ons mark the path in exosomal protein sorting. Cell. Mol. Life Sci. 75, 1–19. doi:10.1007/s00018-017-2690-y
Muntasell, A., Berger, A. C., and Roche, P. A. (2007). T cell-induced secretion of MHC class II–peptide complexes on B cell exosomes. EMBO J. 26, 4263–4272. doi:10.1038/sj.emboj.7601842
Nolte-'t Hoen, E. N., Wagenaar-Hilbers, J. P., Peters, P. J., Gadella, B. M., van Eden, W., and Wauben, M. H. (2004). Uptake of membrane molecules from T cells endows antigen-presenting cells with novel functional properties. Eur. J. Immunol. 34, 3115–3125. doi:10.1002/eji.200324711
Ohgaki, H., and Kleihues, P. (2005). Population-based studies on incidence, survival rates, and genetic alterations in astrocytic and oligodendroglial gliomas. J. Neuropathol. Exp. Neurol. 64, 479–489. doi:10.1093/jnen/64.6.479
Ohgaki, H., and Kleihues, P. (2007). Genetic pathways to primary and secondary glioblastoma. Am. J. Pathol. 170, 1445–1453. doi:10.2353/ajpath.2007.070011
Ohgaki, H., and Kleihues, P. (2013). The definition of primary and secondary glioblastoma. Clin. Cancer Res. 19, 764–772. doi:10.1158/1078-0432.CCR-12-3002
Okoye, I. S., Coomes, S. M., Pelly, V. S., Czieso, S., Papayannopoulos, V., Tolmachova, T., et al. (2014). MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity 41, 503–103. doi:10.1016/j.immuni.2014.08.008
Ostrom, Q. T., Cioffi, G., Waite, K., Kruchko, C., and Barnholtz-Sloan, J. S. (2021). CBTRUS statistical report: Primary brain and other central nervous system tumors diagnosed in the United States in 2014-2018. Neuro. Oncol. 23, III1–III105. doi:10.1093/neuonc/noab200
Oushy, S., Hellwinkel, J. E., Wang, M., Nguyen, G. J., Gunaydin, D., Harland, T. A., et al. (2018). Glioblastoma multiforme-derived extracellular vesicles drive normal astrocytes towards a tumour-enhancing phenotype. Philos. Trans. R. Soc. Lond. B Biol. Sci. 373, 20160477. doi:10.1098/rstb.2016.0477
Papadopoulos, Z., Herz, J., and Kipnis, J. (2020). Meningeal lymphatics: From anatomy to central nervous system immune surveillance. J. Immunol. 204, 286–293. doi:10.4049/jimmunol.1900838
Pavlyukov, M. S., Yu, H., Bastola, S., Minata, M., Shender, V. O., Lee, Y., et al. (2018). Apoptotic cell-derived extracellular vesicles promote malignancy of glioblastoma via intercellular transfer of splicing factors. Cancer Cell 34, 119–135. doi:10.1016/j.ccell.2018.05.012
Pende, D., Cantoni, C., Rivera, P., Vitale, M., Castriconi, R., Marcenaro, S., et al. (2001). Role of NKG2D in tumor cell lysis mediated by human NK cells: Cooperation with natural cytotoxicity receptors and capability of recognizing tumors of nonepithelial origin. Eur. J. Immunol. 31, 1076–1086. doi:10.1002/1521-4141(200104)31:4<1076:aid-immu1076>3.0.co;2-y
Piper, R. C., Dikic, I., and Lukacs, G. L. (2014). Ubiquitin-dependent sorting in endocytosis. Cold Spring Harb. Perspect. Biol. 6, a016808. doi:10.1101/cshperspect.a016808
Qian, M., Wang, S., Guo, X., Wang, J., Zhang, Z., Qiu, W., et al. (2020). Hypoxic glioma-derived exosomes deliver microRNA-1246 to induce M2 macrophage polarization by targeting TERF2IP via the STAT3 and NF-κB pathways. Oncogene 39, 428–442. doi:10.1038/s41388-019-0996-y
Quail, D. F., and Joyce, J. A. (2017). The microenvironmental landscape of brain tumors. Cancer Cell 31, 326–341. doi:10.1016/j.ccell.2017.02.009
Ramakrishnan, V., Xu, B., Akers, J., Nguyen, T., Ma, J., Dhawan, S., et al. (2020). Radiation-induced extracellular vesicle (EV) release of miR-603 promotes IGF1-mediated stem cell state in glioblastomas. EBioMedicine 55, 102736. doi:10.1016/j.ebiom.2020.102736
Raposo, G., Nijman, H. W., Stoorvogel, W., Liejendekker, R., Harding, C. V., Melief, C. J., et al. (1996). B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 183, 1161–1172. doi:10.1084/jem.183.3.1161
Ricklefs, F., Mineo, M., Rooj, A. K., Nakano, I., Charest, A., Weissleder, R., et al. (2016). Extracellular vesicles from high-grade glioma exchange diverse pro-oncogenic signals that maintain intratumoral heterogeneity. Cancer Res. 76, 2876–2881. doi:10.1158/0008-5472.CAN-15-3432
Ricklefs, F. L., Alayo, Q., Krenzlin, H., Mahmoud, A. B., Speranza, M. C., Nakashima, H., et al. (2018). Immune evasion mediated by PD-L1 on glioblastoma-derived extracellular vesicles. Sci. Adv. 4, eaar2766. doi:10.1126/sciadv.aar2766
Salaud, C., Alvarez-Arenas, A., Geraldo, F., Belmonte-Beitia, J., Calvo, G. F., Gratas, C., et al. (2020). Mitochondria transfer from tumor-activated stromal cells (TASC) to primary Glioblastoma cells. Biochem. Biophys. Res. Commun. 533, 139–147. doi:10.1016/j.bbrc.2020.08.101
Schuler, P. J., Saze, Z., Hong, C. S., Muller, L., Gillespie, D. G., Cheng, D., et al. (2014). Human CD4+CD39+ regulatory T cells produce adenosine upon co-expression of surface CD73 or contact with CD73+ exosomes or CD73+ cells. Clin. Exp. Immunol. 177, 531–543. doi:10.1111/cei.12354
Shao, H., Chung, J., Lee, K., Balaj, L., Min, C., Carter, B. S., et al. (2015). Chip-based analysis of exosomal mRNA mediating drug resistance in glioblastoma. Nat. Commun. 6, 6999–9. doi:10.1038/ncomms7999
Simon, T., Pinioti, S., Schellenberger, P., Rajeeve, V., Wendler, F., Cutillas, P. R., et al. (2018). Shedding of bevacizumab in tumour cells-derived extracellular vesicles as a new therapeutic escape mechanism in glioblastoma. Mol. Cancer 17, 132. doi:10.1186/s12943-018-0878-x
Skak, K., Frederiksen, K. S., and Lundsgaard, D. (2008). Interleukin-21 activates human natural killer cells and modulates their surface receptor expression. Immunology 123, 575–583. doi:10.1111/j.1365-2567.2007.02730.x
Smyth, L. A., Ratnasothy, K., Tsang, J. Y., Boardman, D., Warley, A., Lechler, R., et al. (2013). CD73 expression on extracellular vesicles derived from CD4+ CD25+ Foxp3+ T cells contributes to their regulatory function. Eur. J. Immunol. 43, 2430–2440. doi:10.1002/eji.201242909
Spinelli, C., Montermini, L., Meehan, B., Brisson, A. R., Tan, S., Choi, D., et al. (2018). Molecular subtypes and differentiation programmes of glioma stem cells as determinants of extracellular vesicle profiles and endothelial cell-stimulating activities. J. Extracell. Vesicles 7, 1490144. doi:10.1080/20013078.2018.1490144
Stensjøen, A. L., Solheim, O., Kvistad, K. A., Håberg, A. K., Salvesen, Ø., and Berntsen, E. M. (2015). Growth dynamics of untreated glioblastomas in vivo. Neuro. Oncol. 17, 1402–1411. doi:10.1093/neuonc/nov029
Thakur, B. K., Zhang, H., Becker, A., Matei, I., Huang, Y., Costa-Silva, B., et al. (2014). Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 24, 766–769. doi:10.1038/cr.2014.44
Théry, C., Witwer, K. W., Aikawa, E., Alcaraz, M. J., Anderson, J. D., Andriantsitohaina, R., et al. (2018). Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 7, 1535750. doi:10.1080/20013078.2018.1535750
Tian, Y., Wang, Z., Wang, Y., Yin, B., Yuan, J., Qiang, B., et al. (2020). Glioma-derived endothelial cells promote glioma cells migration via extracellular vesicles-mediated transfer of MYO1C. Biochem. Biophys. Res. Commun. 525, 155–161. doi:10.1016/j.bbrc.2020.02.017
Tomaszewski, W., Sanchez-Perez, L., Gajewski, T. F., and Sampson, J. H. (2019). Brain tumor microenvironment and host state: Implications for immunotherapy. Clin. Cancer Res. 25, 4202–4210. doi:10.1158/1078-0432.CCR-18-1627
Treps, L., Edmond, S., Harford-Wright, E., Galan-Moya, E. M., Schmitt, A., Azzi, S., et al. (2016). Extracellular vesicle-transported Semaphorin3A promotes vascular permeability in glioblastoma. Oncogene 35, 2615–2623. doi:10.1038/onc.2015.317
Treps, L., Perret, R., Edmond, S., Ricard, D., and Gavard, J. (2017). Glioblastoma stem-like cells secrete the pro-angiogenic VEGF-A factor in extracellular vesicles. J. Extracell. Vesicles 6, 1359479. doi:10.1080/20013078.2017.1359479
Tsutsui, T., Kawahara, H., Kimura, R., Dong, Y., Jiapaer, S., Sabit, H., et al. (2020). Glioma-derived extracellular vesicles promote tumor progression by conveying WT1. Carcinogenesis 41, 1238–1245. doi:10.1093/carcin/bgaa052
Tung, S. L., Boardman, D. A., Sen, M., Letizia, M., Peng, Q., Cianci, N., et al. (2018). Regulatory T cell-derived extracellular vesicles modify dendritic cell function. Sci. Rep. 8, 6065–6112. doi:10.1038/s41598-018-24531-8
Urabe, F., Kosaka, N., Ito, K., Kimura, T., Egawa, S., and Ochiya, T. (2020). Extracellular vesicles as biomarkers and therapeutic targets for cancer. Am. J. Physiol. Cell Physiol. 318, C29–C39. doi:10.1152/ajpcell.00280.2019
Vagner, T., Spinelli, C., Minciacchi, V. R., Balaj, L., Zandian, M., Conley, A., et al. (2018). Large extracellular vesicles carry most of the tumour DNA circulating in prostate cancer patient plasma. J. Extracell. Vesicles 7, 1505403. doi:10.1080/20013078.2018.1505403
Van Der Vos, K. E., Abels, E. R., Zhang, X., Lai, C., Carrizosa, E., Oakley, D., et al. (2016). Directly visualized glioblastoma-derived extracellular vesicles transfer RNA to microglia/macrophages in the brain. Neuro. Oncol. 18, 58–69. doi:10.1093/neuonc/nov244
Viel, S., Marçais, A., Guimaraes, F. S. F., Loftus, R., Rabilloud, J., Grau, M., et al. (2016). TGF-β inhibits the activation and functions of NK cells by repressing the mTOR pathway. Sci. Signal. 9, ra19–14. doi:10.1126/scisignal.aad1884
Villarroya-Beltri, C., Gutiérrez-Vázquez, C., Sánchez-Cabo, F., Pérez-Hernández, D., Vázquez, J., Martin-Cofreces, N., et al. (2013). Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 4, 2980–3010. doi:10.1038/ncomms3980
Wagner, K. D., Cherfils-Vicini, J., Hosen, N., Hohenstein, P., Gilson, E., Hastie, N. D., et al. (2014). The Wilms tumour suppressor Wt1 is a major regulator of tumour angiogenesis and progression. Nat. Commun. 5, 5852. doi:10.1038/ncomms6852
Walker, L. S. K., and Sansom, D. M. (2011). The emerging role of CTLA4 as a cell-extrinsic regulator of T cell responses. Nat. Rev. Immunol. 11, 852–863. doi:10.1038/nri3108
Yang, J., Dang, G., Lü, S., Liu, H., Ma, X., Han, L., et al. (2019). T‐cell–derived extracellular vesicles regulate B‐cell IgG production via pyruvate kinase muscle isozyme 2. FASEB J. 33, 12780–12799. doi:10.1096/fj.201900863R
Yekula, A., Minciacchi, V. R., Morello, M., Shao, H., Park, Y., Zhang, X., et al. (2020). Large and small extracellular vesicles released by glioma cells in vitro and in vivo. J. Extracell. Vesicles 9, 1689784. doi:10.1080/20013078.2019.1689784
Yu, M. W., and Quail, D. F. (2021). Immunotherapy for glioblastoma: Current progress and challenges. Front. Immunol. 12, 676301. doi:10.3389/fimmu.2021.676301
Yue, X., Lan, F., and Xia, T. (2019). Hypoxic glioma cell-secreted exosomal miR-301a activates wnt/β-catenin signaling and promotes radiation resistance by targeting TCEAL7. Mol. Ther. 27, 1939–1949. doi:10.1016/j.ymthe.2019.07.011
Zhang, F., Li, R., Yang, Y., Shi, C., Shen, Y., Lu, C., et al. (2019). Specific decrease in B-Cell-Derived extracellular vesicles enhances post-chemotherapeutic CD8 + T cell responses. Immunity 50, 738–750. doi:10.1016/j.immuni.2019.01.010
Zhang, Z., Xu, J., Chen, Z., Wang, H., Xue, H., Yang, C., et al. (2020). Transfer of MicroRNA via macrophage-derived extracellular vesicles promotes proneural-to-mesenchymal transition in glioma stem cells. Cancer Immunol. Res. 8, 966–981. doi:10.1158/2326-6066.CIR-19-0759
Keywords: glioma, exosome, immune cells, Tumor microenvironment, microvesicle (MV)
Citation: Low JJW, Sulaiman SA, Johdi NA and Abu N (2022) Immunomodulatory effects of extracellular vesicles in glioblastoma. Front. Cell Dev. Biol. 10:996805. doi: 10.3389/fcell.2022.996805
Received: 18 July 2022; Accepted: 26 September 2022;
Published: 16 November 2022.
Edited by:
Katarzyna Nazimek, Jagiellonian University, PolandReviewed by:
Takashi Minami, Kumamoto University, JapanCopyright © 2022 Low, Sulaiman, Johdi and Abu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nadiah Abu, bmFkaWFoLmFidUBwcHVrbS51a20uZWR1Lm15
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.