 Lin Hu
Lin Hu Xin Wu
Xin Wu Dongjie Chen
Dongjie Chen Zhenyu Cao4
Zhenyu Cao4 Qiangqiang Zhao
Qiangqiang Zhao
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell Dev. Biol. , 20 September 2022
Sec. Molecular and Cellular Oncology
Volume 10 - 2022 | https://doi.org/10.3389/fcell.2022.814722
Osteosarcoma (OS) is one of the most common types of solid sarcoma with a poor prognosis. Solid tumors are often exposed to hypoxic conditions, while hypoxia is regarded as a driving force in tumor recurrence, metastasis, progression, low chemosensitivity and poor prognosis. Pytoptosis is a gasdermin-mediated inflammatory cell death that plays an essential role in host defense against tumorigenesis. However, few studies have reported relationships among hypoxia, pyroptosis, tumor immune microenvironment, chemosensitivity, and prognosis in OS. In this study, gene and clinical data from Therapeutically Applicable Research to Generate Effective Treatments (TARGET) and Gene Expression Omnibus (GEO) databases were merged to develop a hypoxia risk model comprising four genes (PDK1, LOX, DCN, and HMOX1). The high hypoxia risk group had a poor prognosis and immunosuppressive status. Meanwhile, the infiltration of CD8+ T cells, activated memory CD4+ T cells, and related chemokines and genes were associated with clinical survival outcomes or chemosensitivity, the possible crucial driving forces of the OS hypoxia immune microenvironment that affect the development of pyroptosis. We established a pyroptosis risk model based on 14 pyroptosis-related genes to independently predict not only the prognosis but also the chemotherapy sensitivities. By exploring the various connections between the hypoxic immune microenvironment and pyroptosis, this study indicates that hypoxia could influence tumor immune microenvironment (TIM) remodeling and promote pyroptosis leading to poor prognosis and low chemosensitivity.
Osteosarcoma (OS), currently the most common primary solid bone malignancy, is derived from primitive mesenchymal cells and usually occurs in children and young adults (Li et al., 2011; Smeland et al., 2019). The incidence of OS is over 8 million/year at 15–19 years of age in Europe (Ritter and Bielack, 2010). OS is severely malignant; a considerable number of patients with the seemingly localized disease will develop metastases mostly in the lungs (Goguet-Surmenian et al., 2013). Thus, a multidisciplinary treatment approach is very important. To date, the 5-year survival of OS has improved and is achieved by almost 70% of the patients via the advancement of neoadjuvant and adjuvant chemotherapy combined with surgical resection (Rosen, 1985; Lilienthal and Herold, 2020). However, OS is a chemotherapy-resistant solid tumor with many patients being insensitive to chemotherapy due to the highly dynamic immune landscape in patients with OS ((Wu et al., 2021a), (Wu et al., 2021b)). Thus, a deep understanding of the tumor immune status in OS is crucial for improving prognosis and selecting effective treatments.
Hypoxia plays an important role in solid tumor development including OS. Specifically, hypoxia is a significant factor that affects tumor cell proliferation and invasion (Guan et al., 2015; Chen et al., 2020). The imbalance between oxygen consumption and supply results in hypoxia, an inevitable phenomenon in the majority of tumors (Zeng et al., 2011). Hypoxia is a relevant factor in chemosensitivity, metastatic potential, malignant phenotype, and poor survival (Song et al., 2006; Guan et al., 2015; Prudowsky and Yustein, 2020). Several studies have also found that the complicated relationship between hypoxia and the tumor microenvironment has been shown to occur in various tumors such as hepatocellular carcinoma, glioma, gastric cancer, acute myeloid leukemia, and breast cancer (Gibadulinova et al., 2020; Lin et al., 2020; Liu et al., 2020; Du et al., 2021; Zeng et al., 2021; Jiang et al., 2022). Hypoxia can decrease immune cell (e.g., CD8+ T cells) proliferation and promote disadvantageous expression of immune factors (e.g., HMGB1 and TLR4) (Yang et al., 2014; Liu et al., 2015; Wu et al., 2018; Scharping et al., 2021).
Pyroptosis, a type of proinflammatory cell death, is executed by the gasdermin family of pore-forming proteins (Zhou et al., 2020a). Pyroptosis can lead to the continuous expansion of cells until the cell membrane ruptures and is associated with the release of cell contents and inflammatory response (Shi et al., 2015; Xia et al., 2019). Increasing evidence indicates a close relationship between pyroptosis and tumors. The occurrence of pyroptosis is often accompanied by multiple signaling pathways and inflammatory responses that may be associated with the regulation of tumor immune and is involved in tumorigenesis, antitumor immune function, and chemosensitivity (Man, 2018; Xia et al., 2019; Hou et al., 2020; Tan et al., 2020; Wu et al., 2021c). Interestingly, several studies suggest that hypoxia causes pyroptosis through multiple potential pathways, which may due to the production of reactive oxygen species (ROS) or the mediation of circ-Calm4/mir-124-3p/Pdcd6 axis (Yu et al., 2019; Jiang et al., 2021). Hypoxia-inducible factor 1-α (HIF-1A) is a major transcriptional regulator of cell and developmental responses to hypoxia (Iyer et al., 1998). The activity of HIF-1A is directly regulated by cellular oxygen levels (Semenza, 1999). Jorgensen et al. (2016) found mitochondria with collapsed cristae in pyroptotic cells. Mitochondria are the main sites of ROS production. They have a four-layer structure, and in the process of oxidative phosphorylation (OXPHOS), ROS production mainly occurs on the electron transport chain (ETC) inside the mitochondrial membrane, which folds inward to form ridges, known as cristae (Li et al., 2013). Therefore, collapsed cristae may lead to reduced production of reactive oxygen species (Madamanchi and Runge, 2007), leading to hypoxia of cells. In fact, hypoxia can lead to pyroptosis (Jiao et al., 2018). TNF induced apoptosis has been observed in cell lines of multiple cancer types under normoxic conditions, while the same treatment induces necrotic cell death under hypoxia. It was found that necrotic cell death depended on Caspase-8 and GSDMC, suggesting that increased GSDMC expression under hypoxia could transform caspase-8-induced apoptosis into pyroptosis (Tsuchiya, 2021). A recent study showed that nuclear PD-L1 was found to enhance the expression of GSDMC under hypoxia conditions, while breast cancer patients with increased GSDMC expression had a poor prognosis, suggesting that GSDMC in the center of hypoxia regions may promote tumor progression by inducing the chronic tumor necrosis (Hou et al., 2020). Although pyroptosis has attracted considerable attention because of its potential effect on the tumor immune response, the relationship between hypoxia and pyroptosis and the role of pyroptosis in OS is unclear.
In his study, we established and validated a hypoxia risk model and a pyroptosis scoring model to predict prognosis, changes in the tumor immune microenvironment, and treatment sensitivity. Additionally, we categorized patients with OS into two cluster types based on pyroptosis-related genes that could predict prognosis and immune infiltration. These studies may help assist clinicians in their decision-making and provide insight into the relationships among hypoxia, tumor immune microenvironment, pyroptosis, and OS progression.
The RNA sequence and osteosarcoma patient characteristics were collected from the TARGET database. RNA expression data of normal human tissues were obtained from the GTEx. The “sva” R package (version 3.8) was used to correct biases after TARGET and GTEx data sets had been pooled. The merged data were used as the training dataset. The external validation GSE cohorts were selected from GEO, named GSE21257 (hypoxia), with detailed survival data. In the establishment of the pyroptosis risk model, TARGET data was separated into two parts: 70% used were for the training dataset, and 30% for the testing dataset. Perl scripts were used to transform the names of microarray probes into gene names.
Differentially expressed genes (DEGs) related to hypoxia and pyroptosis in normal tissues and osteosarcoma were identified via the “limma” package (version 3.48.3) with logFC >1 and p < 0.05 as the criteria. The protein-protein interaction (PPI) network was constructed with the online-supported tool STRING (version 11.5) (Szklarczyk et al., 2015). The ClusterProfiler” package in R software was used to execute the gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses. The heatmap was generated using the “Pheatmap” package (version 1.0.12).
To evaluate the expression of hypoxia, hypoxia-related genes (HRG) were obtained from hallmark-hypoxia gene sets obtained from the gene set enrichment analyses (GSEA). The statistically significant DEGs related to hypoxia were tested using univariable and multivariable Cox regression. The hypoxia risk score formula is as follows:
Preliminary screening of the survival-related genes was performed by univariate Cox regression analysis, and an optimum pyroptosis-related signature with robust distinguishing ability was established according to the optimum λ value via LASSO Cox regression analysis. The following formula was used to calculate the hypoxia risk score for each patient:
GSEA was utilized to examine the significant difference between the high hypoxia risk and low hypoxia risk groups of the expressed set of genes via GSEA software (version 4.1.0). Enrichment of MSigDB Collection (c7. all.v7.4. symbols) provided by the Molecular Signatures Database was regarded as the reference gene set. Kaplan-Meier survival curves were used for the survival analysis through the “survival” and “survminer” packages in R. Identification of potential prognostic factors was performed by Univariate Cox analysis. The multivariate Cox analysis was used to further determine independent prognostic factors. Time-dependent receiver operating characteristic (ROC) curves were used to determine the predictive efficiency of the model via the survivalROC R package.
To quantify the relative infiltration of various immune cell types and immune-related pathways in the high hypoxia risk and low hypoxia risk groups, ssGSEA analysis was performed.
The enrichment score determined by ssGSEA represents the level of immune-related cell infiltration in each sample. The CIBERSORT tool distinguished 22 types of immune cells via a gene signature matrix including 547 genes, named LM22 ((Newman et al., 2015)). To identify the characteristics of immune status related to pyroptosis, the infiltration of various immune cells in different groups was evaluated by CIBERSORT (version 1.0649).
The NCI-60 database was obtained from the CellMiner web site (https://discover.nci.nih.gov/cellminer), which covered 9 cancer categories and 60 different cancer cell lines. Besides, Pearson correlation analysis was used for exploring the relevance between the independent prognostic PRGs and drug sensitivity. Drugs used in this sensitivity analysis are those approved by the FDA or those in clinical tests.
Two pairs of Small interfering RNA (siRNA) targeting human CCL28/HMGB1/KLF2/TLR4/TNFSF18 were synthesized by Invitrogen, named Gene-SiRNA1and Gene-SiRNA2. The primer sequences are provided in Supplementary Table S1. Osteosarcoma cells were seeded with a density of (Rosen, 1985; Ritter and Bielack, 2010; Li et al., 2011; Goguet-Surmenian et al., 2013; Smeland et al., 2019) × 105 per well in 12-well plates. According to the manufacturer’s instruction, transfection of siRNAs was carried out using Lipofectamine™ 3,000 transfection with the when cells reached 80% confluence. Total RNA was extracted with TRIzol reagent and reverse transcribed to cDNA. Based on mRNA levels determined by RT-qPCR analysis. The most effective siRNA was determined. This experiment was carried out with 2 replicates.
Transfection was conducted as describe above and the supernatant was collected after 48 h of incubation. Subsequently, cells were digested by trypsin without EDTA and harvested the single cell suspension. The cells were resuspended with cold PBS and cell precipitates were collected by centrifugation. After washing with binding buffer, cells were stained with Annexin V−FITC and propidium iodide. Then, apoptotic analysis was immediately performed by flow cytometer.
First, paraffin tumor sections were dewaxed in water. Then, a microwave thermal process was used to repair the antigen. The slides were washed thrice in phosphate buffer solution (PBS) for 5 min each time, blocked with 10% goat serum, and incubated in primary antibody at 4°C overnight, then added secondary antibodies for incubated 50 min at room temperature. The nuclei were stained by DAPI solution after three times wash and incubated 10 min. Sections were then observed and images were collected using a fluorescence microscope (NIKON ECLIPSE C1). The paraffin tumor sections were obtained from Wuhan Sevier Biotechnology Co., Ltd.
R software (version x64 4.1.0) was used for the statistical analysis. Correlations between variables were analyzed using Pearson or Spearman coefficients. Comparisons between the two groups were performed using the Wilcoxon test. The Kruskal Wallis test was conducted to compare more than two groups. The Kaplan-Meier analysis was carried out via “survival” and “survminer” packages in R to compare the overall survival between groups. All statistical p values were two-sided, and the level of significance was set at p < 0.05.
The flow chart illustrates the entire study design (Figure 1). Two hundred hypoxia-related genes (HRGs) were downloaded from GSEA software. We evaluated the number of adjacent nodes and identified the top 50 genes with most interactions that might play a significant role in the response to hypoxia (Figure 2A). A more detailed PPI analysis was conducted to visualize the internal links among these HRGs (Figure 2B).
FIGURE 1. Flow chart of experimental design.
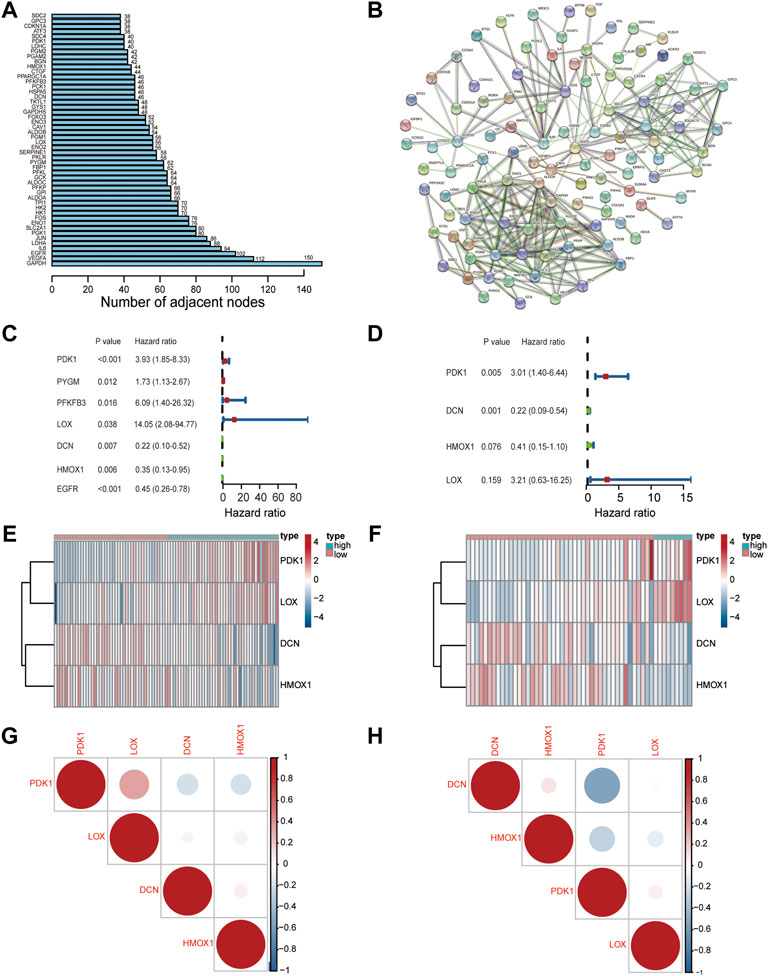
FIGURE 2. Characterization of a hypoxia risk model to predict prognosis of OS. (A) The number of adjacent nodes per hypoxia-related gene; (B) PPI network indicating the correlation of HRGs; (C,D) A hypoxia-risk model was evaluated by univariate and multivariate Cox regression; (E,F) Heatmap displaying expressions of hypoxia-related DEGs and comparison between high and low hypoxia risk groups in TARGET (E) and GEO databases (F); (G,H) Correlation analysis of hub gene in TARGET (G) and GEO databases (H).
Univariate Cox regression analysis was first performed to screen out prognostic genes in the training set that was merged with the TARGET and GTEx datasets (Figure 2C). Then, four genes (PDK1, LOX, DCN, and HMOX1) were selected to construct an optimal predictive model by utilizing the multivariate Cox regression analysis (Figure 2D). The formula for the hypoxia risk score calculation is as follows:
A heatmap of the four genes expression was used to visualize the different expression patterns. PDK1 and LOX were upregulated in the high hypoxia risk score group in both TARGET (training) (Figure 2E) and GEO (testing) datasets (Figure 2F). Correlations among all four genes were evaluated and were found not to be significant in both the dataset and the testing GEO dataset (Figures 2G,H). To determine the prognostic value of the four genes, overall survival curves for patients with different levels of individual gene expression in the training dataset were generated. The results indicated that higher PDK1 expression was associated with low survival probability (p = 0.002, Supplementary Figure S1), while there was no difference between the high- and low-expression groups for the other genes. The above results suggested that these four HRGs could be used to build the hypoxia risk model.
To further explore the signaling pathways for the HRGs, KEGG pathway enrichment (Figures 3A,B) and GO enrichment (Figures 3C,D) were conducted using TARGET dataset. The results indicated that the HRGs were primarily associated with mesenchyme development, extracellular matrix or structure organization, external encapsulating structure organization.
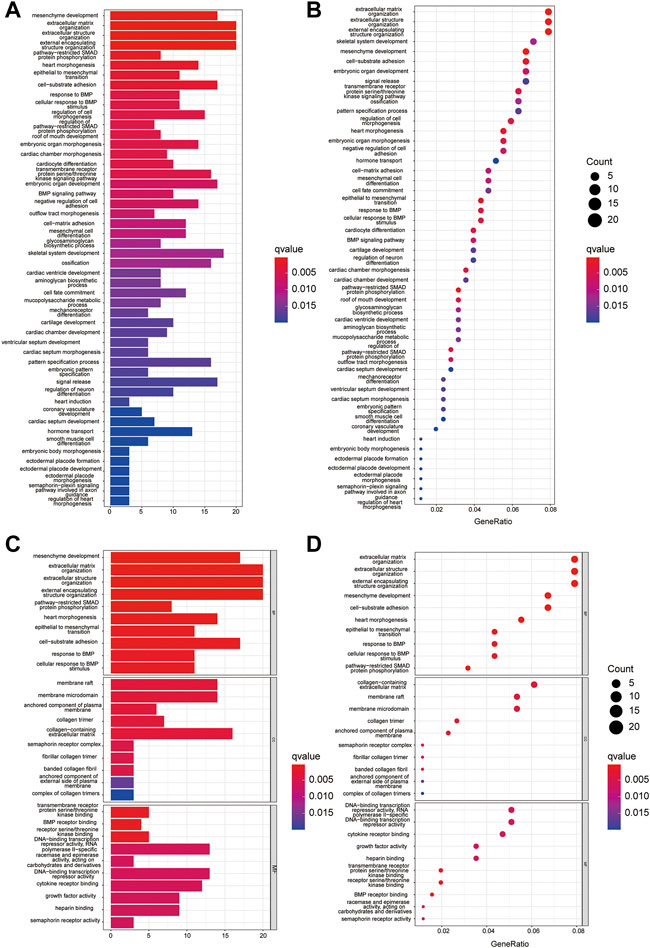
FIGURE 3. GO and KEGG enrichment analysis in TARGET dataset. (A,C) The KEGG (A) and GO (C) analysis by bar graph; (B,D) Bubble chart shows significantly enriched KEGG (B) pathway and GO (D).
Hypoxia often correlates with cancer progression and poor survival outcomes; thus, we further examined the predictive ability of the hypoxia risk model. Based on the median value of the hypoxia risk score, patients were assigned to the high or low hypoxia risk group in the TARGET and GEO datasets.
Patients in the training (Figure 4A) and testing datasets (Figure 4B) were stratified into high and low groups according to the median value of the HRS. The results showed that the mortality of the high hypoxia risk group was significantly higher than that of the low hypoxia risk group in training dataset (Figures 4C,E) and testing dataset (Figures 4D,F). Then, Kaplan-Meier analysis was performed to further assess the predictive ability of the hypoxia risk model in patients with OS. The results showed that the overall survival was worse in the high hypoxia risk group than in the low hypoxia risk group in the TARGET dataset (Figure 4G, p = 0.009); this was further confirmed in the GEO cohort (p = 0.035, Figure 4H). Then, the ROC curves were obtained for the TARGET and GEO cohorts to assess the predictive accuracy of the hypoxia risk model for the 1-, 3- and 5- year survival rates. In the TARGET dataset, the area under the ROC curve (AUC) was 0.679 at 1-year, 0.765 at 3-year, 0.740 at 5-year, indicating the robustness and accuracy of the hypoxia risk model (Figure 3I). This was confirmed in the GEO dataset (Figure 4J). Therefore, the hypoxia risk model is closely related to OS prognosis.
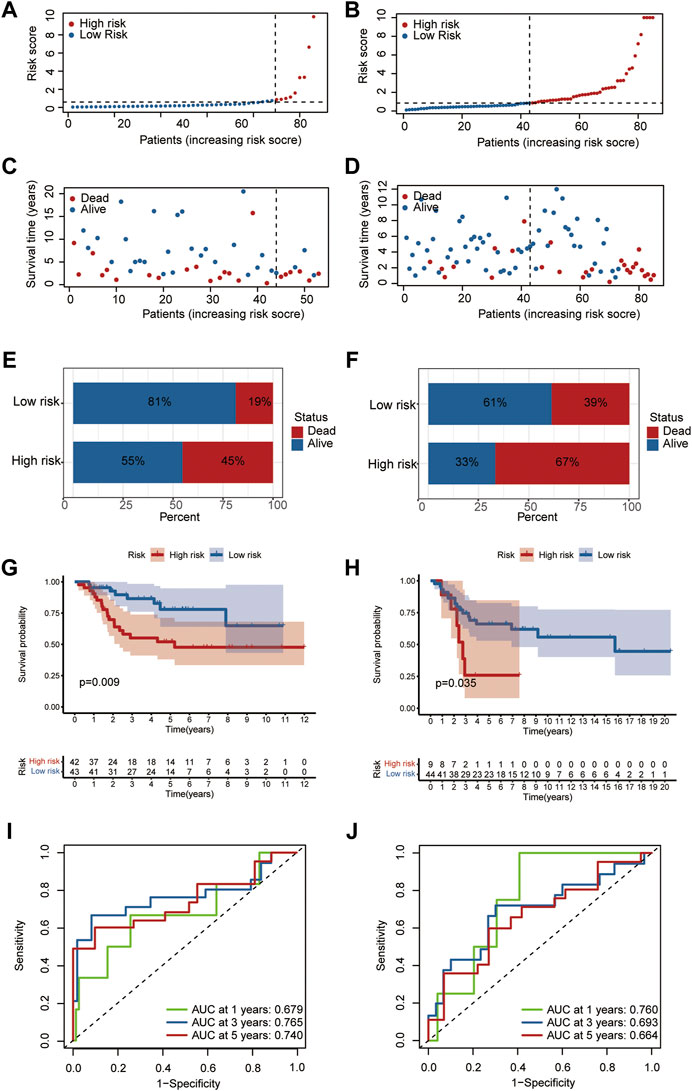
FIGURE 4. Predictive ability of the hypoxia risk model in OS. (A,C) Distribution of survival status and risk score in the high- and low-risk groups in TARGET dataset; (B,D) Distribution of survival status and risk score in the high- and low-risk group in the GEO database; (E,F) Mortality rates of the high- and low-risk group in TARGET (E) and GEO (F) databases; (G,H) The curves for predicting overall survival for patients in TARGET (G) and GEO (H) databases; (I,J) ROC curves of TARGET (I) and GEO (J) cohorts.
Previous studies indicated that the microenvironment in hypoxic conditions may help tumors resist antitumor immunity by reshaping TIM and enhancing immune evasion. We performed a CIBERSORT analysis to identify immune composition differences between the high- and low-risk groups. The heatmap of immune composition in patients from the training and testing datasets is shown in Figures 5A,B. The results suggest a lower immune cell infiltration in high hypoxia risk patients from the TARGET dataset such as CD8+ T cells, activated memory CD4+ T cells, and macrophage M2 cells, while M0 macrophages showed higher infiltration in the high hypoxia risk score group in the TARGET dataset (Figures 5C–F). Furthermore, the expression of Granzyme B (GZMB) and Interferon Gamma (IFNG) negatively correlated with hypoxia risk score in TARGET dataset (Supplementary Figures S2A, B) and GEO dataset (Supplementary Figures S2C, D). These results show that hypoxia may participate in TIM rebuilding and could be a target for immune therapy.
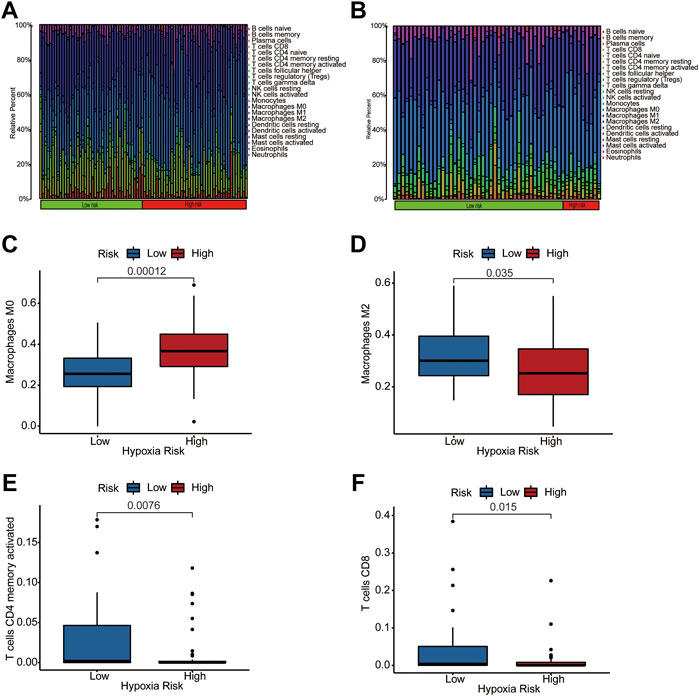
FIGURE 5. The immune landscape of two hypoxia score subtypes. (A,B) Distribution of immune infiltration in patients of high- and low-hypoxia risk groups from TARGET (A) and GEO (B) databases; (C–F) Comparison of various immune cells between high- and low-hypoxia risk groups from training dataset, visualized by Box plots.
The cancer-immunity cycle has guided studies of immune therapy and describes a series of stepwise events for an antitumor immune response such as antigen release and activation, immune cell migration, identification, and death of cancer cells (Chen and Mellman, 2013). To investigate the relationship between hypoxia and the genes that positively regulate these processes, we downloaded gene signatures from Tracking Tumor Immunophenotype (http://biocc.hrbmu.edu.cn/TIP/) (Xu et al., 2018). As shown in Figure 6, most positively regulating genes of the cancer-immunity cycle were downregulated in the patients with a high hypoxia risk score, suggesting that hypoxia is associated with low activities of these antitumor immune processes.
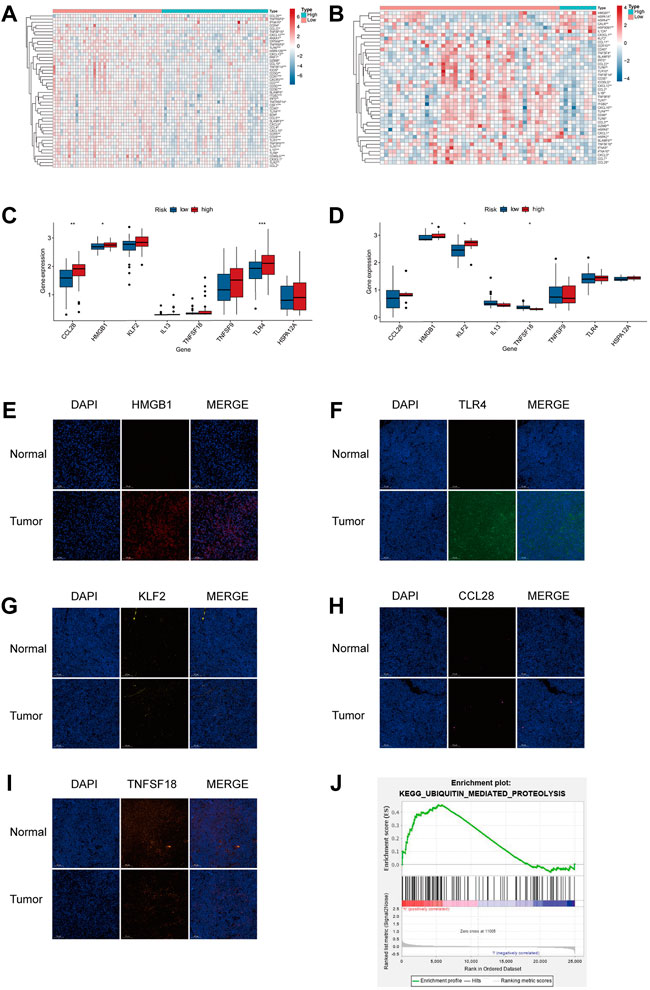
FIGURE 6. TIM of the high hypoxia risk group exhibited immunosuppressive characteristics. (A,B) Heatmap showing the positively regulated genes of the Cancer-Immunity Cycle in two subtypes in the training (A) and testing (B) databases. (C,D) Expression of immune-related signature in the training and testing dataset. (E–I) Expression of immune-related regulators were verified by immunofluorescence in tumor and normal tissues. (J) GSEA in TARGET cohort. ***p < 0.001.
Based on the results of the hypoxic microenvironment, we further explored the links between our model and immune-related factors. As shown in Figure 6A, the high hypoxia risk group had high expression of HMGB1 and TLR4 in the TARGET dataset. In the GEO dataset, the HMGB1 and KLF2 were highly expressed in the high hypoxia risk group, the expression of TNFSF18 was lower in the high hypoxia risk group than in the low hypoxia risk group (Figure 6B). Immunofluorescence was determined in tumor and normal tissues to further verify the immune regulator expression. As shown in Figures 6E–I, the expression of HMGB1 and TLR4 was significantly high in tumor tissue compared with that in normal tissue, the expression of CCL28, KLF2 and TNFSF18 in tumor were similar to normal tissues. In an attempt to investigate the potential pathway related to hypoxia, we conducted GSEA and found that the ubiquitin-mediated proteolysis pathway was enriched in high hypoxia risk patients in the TARGET dataset (Figure 6J).
The above results demonstrated that hypoxia might participate in establishing a TIM with immunosuppression by affecting immune regulators.
To identify the effects of HMGB1, TLR4, CCL28, KLF2 and TNFSF18 knockdown on apoptosis of osteosarcoma cells, the flow cytometry analysis was conducted. The result indicated that CCL28, KLF2 and TNFSF18 did not significantly affect cell apoptosis (Figures 7A,C,E,F), while the knockdown of HMGB1 and TLR4 significantly increased the proportion of apoptotic of osteosarcoma cells (Figures 7B,D,F).
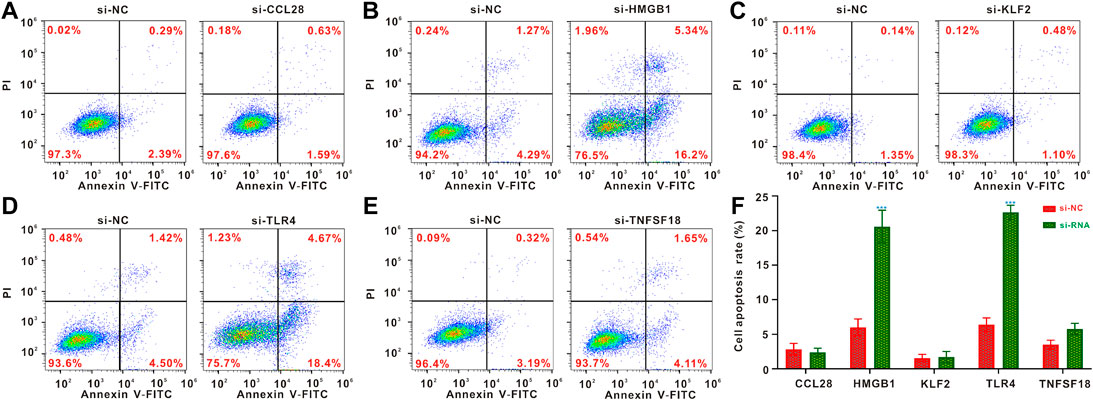
FIGURE 7. Flow cytometric analysis of cell apoptosis. (A–E) Flow cytometric analysis for osteosarcoma cells following CCL28, HMGB1, KLF2, TLR4 and TNFSF18 knockout (F) Quantified results of the flow cytometric analysis. *p < 0.05; **p < 0.01; ***p < 0.001.
According to previous evidence of the close relationship between immune regulators and pyroptosis, for example, HMGB1 and TLR4 may promote the development of pyroptosis (Wang et al., 2019a; Wang et al., 2020a). To determine whether there was any association between the hypoxic microenvironment and pyroptosis, we identified 46 DEGs by comparing the levels of 52 pyroptosis-related genes (PRGs) in the training dataset. The differential levels of gene expression in tumor and normal tissues were analyzed using a heatmap (Figure 8A). As shown in the heatmap, IL1A, GSDMD, IL1B, PYCARD, GZMA, IRF2, CASP8, CASP5, CHMP4C, TP53, NLRP7, TIRAP, GSDMA, AIM2, CHMP4B, BAK1, BAX, CASP3, and CASP6 showed upregulated expression patterns, while other genes were expressed at low levels in patients with OS. The correlation network of the DEGs is presented in Figure 8B. Moreover, we analyzed the PPI network using the STRING online tool (Figure 8C).
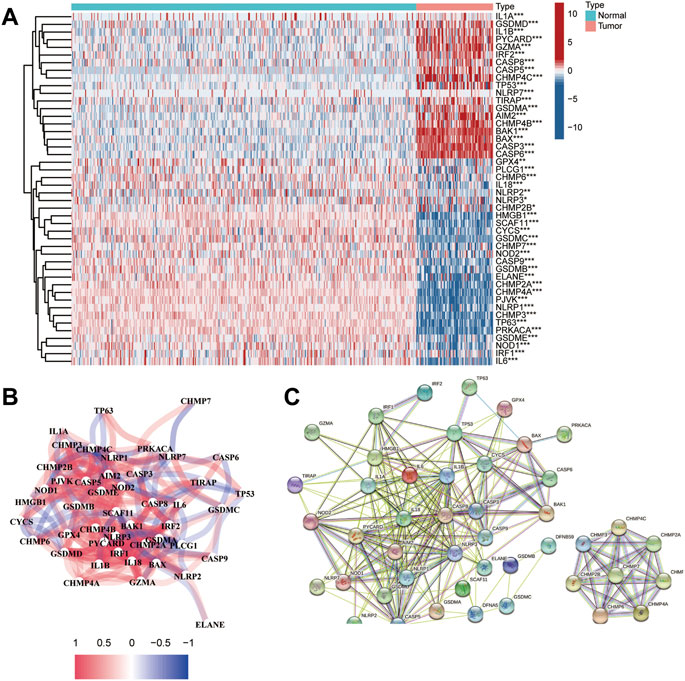
FIGURE 8. Identification of DEGs between healthy and tumor tissues. (A) Heatmap of the PRGs expressed in normal and tumor tissues; (B) The network of the PRGs showing interactions among the genes; (C) PPI network of the PRGs.
A consensus clustering analysis in the TARGET dataset was conducted to further understand the link between PRG and OS. According to the level of pyroptosis-related DEGs expression, we determined that the intergroup correlations were the lowest and the intragroup correlations were the highest, while the clustering variable (k) value was 2, suggesting that patients could be classified into two separate clusters (Figure 9A). We then compared the overall survival times of the two clusters. As shown in Figure 9B, cluster 1 was associated with a higher survival advantage than cluster 2. The analysis results for different k values are shown in Supplementary Figure S3.
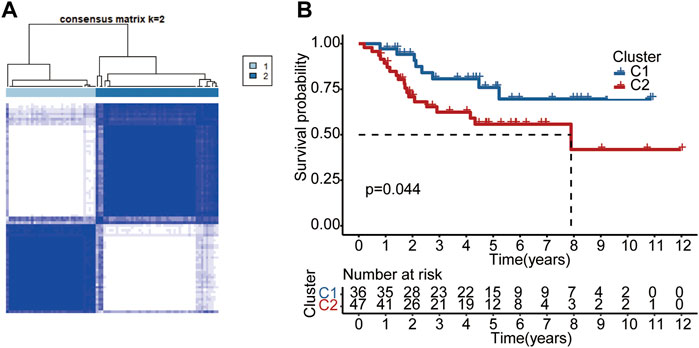
FIGURE 9. Subtypes of OS according to DEGs. (A) Based on the consensus clustering matrix, OS patients were divided into two clusters (k = 2). (B) Kaplan-Meier analysis of the two clusters.
For preliminary screening of the PRGs, univariate Cox regression analysis was used in the TARGET dataset (Supplementary Figure S4). A group of 14 genes (HSD11B2, KIF25, BST1, SNORA75, GBP1, CLDN11, ZNF692, ARMC4, FPR1, PCDHB6, IHH, TPD52, RAMP1, and TAC4) was classified using the optimal λ value (Lossos et al., 2004) by performing the LASSO Cox regression analysis (Figures 10A,B). The coefficient values for each gene are presented in Supplementary Table S2.
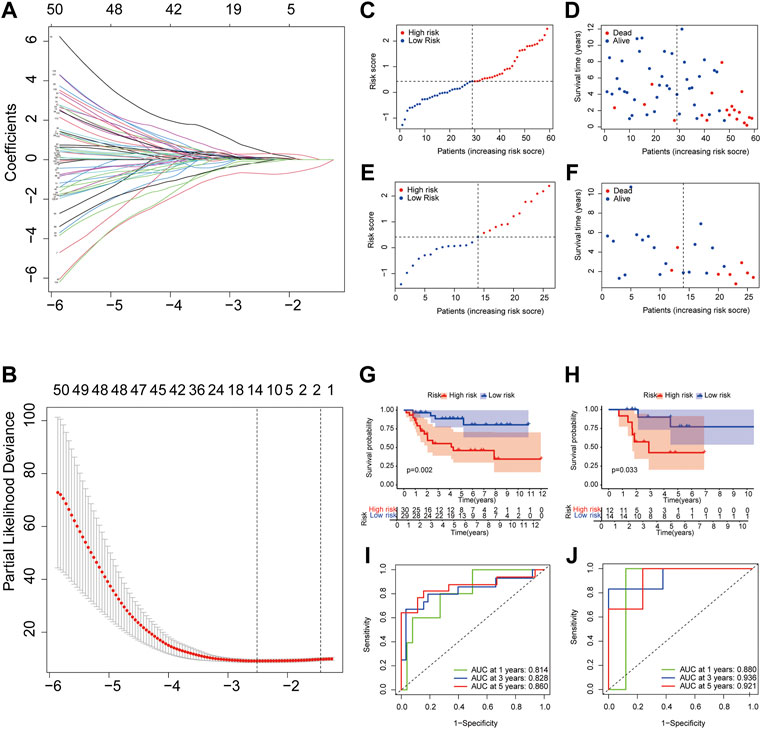
FIGURE 10. Construction and validation of pyroptosis risk signature. (A,B) LASSO regression and cross-validation of genes; (C,E) OS patients assigned in subtypes by the median value of the pyroptosis risk score in training (C) and testing cohorts (E); (D,F) Distribution of survival status in the high and low pyroptosis risk group from training cohort; (G,H) Overall survival curves for patients with high and low pyroptosis risk scores in training (G) and testing cohorts (H); (I,J) ROC curves of the pyroptosis risk signature indicating the predictive ability in training (I) and testing cohorts (J).
To evaluate further the prognostic value of the pyroptosis risk model, we assigned patients into low and high pyroptosis risk score groups in the training and testing cohorts, applying the cut-off median value from training (Figure 10C) and testing (Figure 10E) datasets. The results of the principal component analysis showed that patients with different pyroptosis risk scores were divided into two groups in both the training and testing datasets (Supplementary Figure S5).
To compare survival outcomes between the two pyroptosis risk groups, we performed overall survival and survival status analyses in the training cohort. The low pyroptosis risk score group was associated with better survival status (Figure 10D) and higher overall survival time (Figure 10G) than the high pyroptosis risk group; this was validated further by the testing dataset (Figures 10F,H). The ROC curve shows that AUCs were 0.814 at 1-year, 0.828 at 3-year, 0.860 at 5-year points, suggesting that the risk model has a high predictive value in the training dataset (Figure 10I). This was further confirmed by analyzing the testing cohort (Figure 10J).
To determine the functional enrichment and pathways in groups classified by the pyroptosis risk score, we used R software to identify 172 DEGs conforming to the criteria of FDR <0.05 and |log2FC | ≥ 1.
Using these DEGs, we performed the GO (Figures 11A,B) and KEGG (Figures 11C,D) pathway analysis on the training dataset and determined that the DEGs were mainly associated with the extracellular matrix or structure organization, ossification, BMP, TGF-beta signaling pathway and protein digestion and absorption.
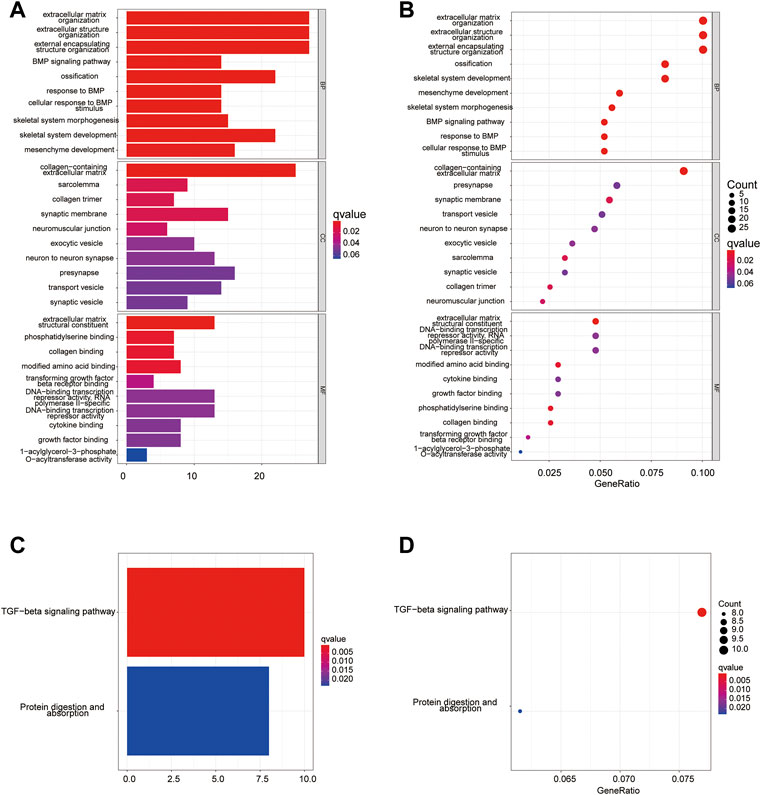
FIGURE 11. Functional analysis of training cohort (from TARGET dataset). (A,C) Barplot graph for GO (A) and KEGG (C) pathways; (B,D) Bubble graph for GO (B) and KEGG (D) enrichment.
To further investigate the potential correlation between the different pyroptosis-related risk and immune activities, we evaluated the level of 13 immune-related pathway activities and the enrichment of various immune cells between the two risk-score groups in the training dataset by ssGSEA (Figure 12A). The results suggested a lower infiltration of immune cells (e.g., IDCs cells, Th1 cells, Th2 cells) in the high pyroptosis risk score group. Moreover, we find lower infiltration of CD8+ T cells, Th1 cells and Th2 cells in the high pyroptosis risk score group in the testing dataset (Figure 12B). The high pyroptosis risk score group showed lower activities in some immune pathways such as antigen-presenting cell (APC) co-inhibition and T cell co-stimulation (Figure 12C). These results indicate that a higher pyroptosis score may be linked to a low antitumor immune response; this was also validated by performing the above examination with the testing dataset (Figure 12D).
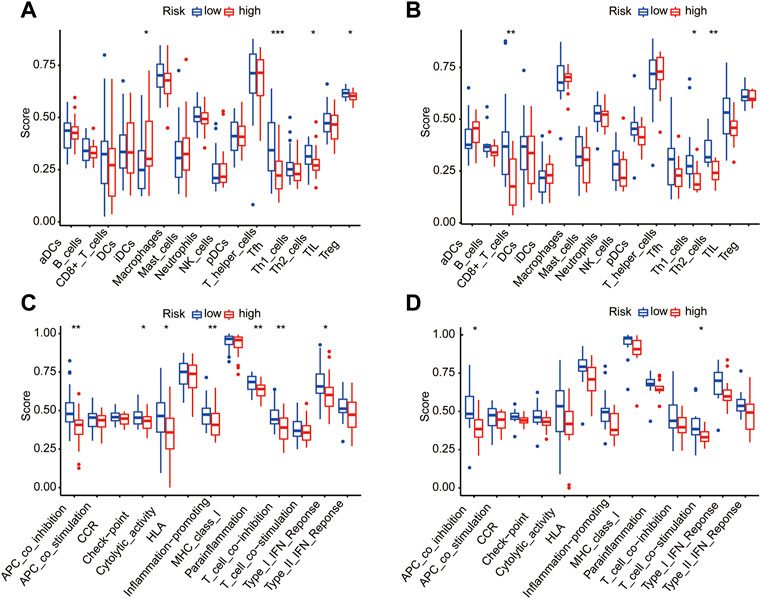
FIGURE 12. Correlation between immune activities and risk score in training and testing datasets. (A/B) Difference of infiltrating immune cell subpopulations and levels between the two pyroptosis risk groups in the training dataset (A) and testing cohorts (B). (C/D) The relationship between pyroptosis risk score and 13 immune-related pathway activities in the training dataset (C) and testing cohorts (D).
Nowadays, with the increase in survival rate due to various chemotherapy combinations, multimodal treatment is considered the most effective strategy. Therefore, we explored whether the expression of the 14 genes pyroptosis risk signature was related to the sensitivity of common chemotherapies in the training dataset. As shown in Figure 13, the expression of FPR1, BTS1 and GBP1 exhibited a positive correlation with chemotherapy strategies, and the expression of HSD11B2, TPD52 and RAMP1 exhibited a negative correlation with chemotherapy strategies. The results indicated that pyroptosis might be a potential target for chemotherapy in patients with OS.
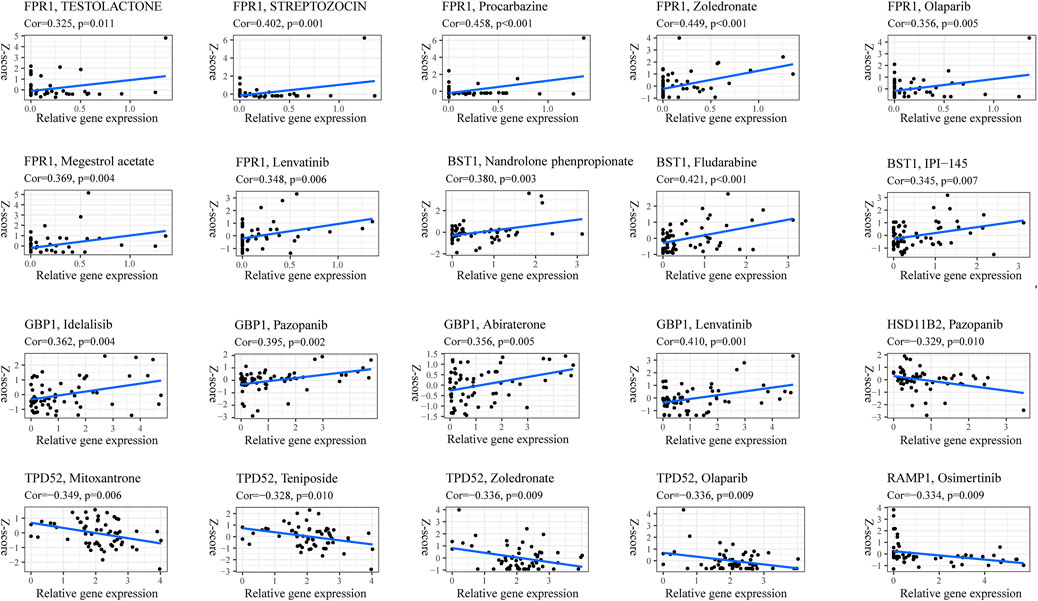
FIGURE 13. Correlation between pyroptosis score and chemotherapy sensitivities.
Osteosarcoma is a highly aggressive and malignant bone tumor with a poor prognosis that occurs primarily in children and adolescents (Jo and Fletcher, 2014). Over the past half-century, new profound insight and effective strategies have been urgently sought to resolve the clinical limitations because the treatment efficacy of OS has plateaued (Ballatori and Hinds, 2016). Despite the growth of multimodal treatment regimens and diagnosis of OS in previous decades, the survival rate remains low; this could be attributed mainly to the limitations of diagnosis, early metastasis, and low chemosensitivity (Niu et al., 2019). Previous evidence indicates that the alteration of TIM and the development of pyroptosis might play a major role in tumor pathogenesis. However, most studies have focused only on individual correlations between hypoxia, TIM, pyroptosis, and OS, and the characteristics of the internal connections among all these factors have not been systematically evaluated; understanding of these interactions might shed new light on treatment methods.
Hypoxia frequently exists in TIM and has been considered a critical mediator of tumor aggressiveness, metastasis, drug sensitivity, and poor prognosis. In addition, hypoxia has been reported to be directly linked to effects on immunosuppressive cells and molecules (Kumar and Gabrilovich, 2014). Several studies also suggested a close linkage between hypoxia and pyroptosis. Meanwhile, as the awareness of pyroptosis increases, the role of pyroptosis in TIM, antitumor response, and tumor progression has received increasing attention. Based on this experience, we established a hypoxia-risk model linked with a pyroptosis-related risk model to predict prognosis, TIM, and chemoresistance, and to reveal the relationship among hypoxia, TIM, and OS.
We initially created a hypoxia-risk model consisting of four HRGs and established a pyroptosis-risk model with 14 PRGs. It has been demonstrated that pyruvate dehydrogenase kinase 1 (PDK1) and lysyl oxidase (LOX) were associated with chemoresistance, tumor metastasis, and poor survival (Umezaki et al., 2019; Siu et al., 2020). Our results show high hypoxia risk with high levels of expression of PDK1 and LOX, validating the suggestion that high hypoxia is linked with high PDK1 and LOX, as previously reported (Majmundar et al., 2010; Ji et al., 2013). In contrast, decorin (DCN) plays a role in impeding tumorigenesis (Sainio and Järveläinen, 2019). Heme oxygenase-1(HOMX1) is considered to be the main protein in diseases that may be driven by oxidative and inflammatory attacks (Waza et al., 2018). After several survival analyses, this hypoxia risk model showed robust accuracy in predicting tumor prognosis.
In addition to contributing to OS aggression and malignancy, hypoxia promotes tumor escape and immune resistance by reducing the infiltration of immune cells and establishing an immunosuppressive microenvironment that may protect tumors from antitumor immune attacks. Previous studies indicate that T cells, and especially CD8+ T cells perform an essential role in the antitumor immune response, for example, CD8+ T cells induce ferroptosis and pyroptosis resulting in the inhabitation of tumor growth and enhancement of antitumor immunity (Lang et al., 2019; Tang et al., 2020; Zhang et al., 2020); enrichment of CD4+ T cells is associated with good prognosis (Liu et al., 2021). However, CD8+ T cells can become rapidly dysfunctional due to hypoxia via mitochondrial stress induced by continuous stimulation (Scharping et al., 2021). We also founded that GZMB and IFNG are negatively correlated with hypoxia risk score. As a T cell activation marker, the expression of GZMB is important for tumor immunity (Russell and Ley, 2002). Moreover, patients with osteosarcoma and a high expression of GZMB have a long overall survival time (Yoshida et al., 2019). The expression of Neuronal pentraxin (NPTX2) is negatively correlated with the expression of GZMB and IFNG, Knockdown of NPTX2 in osteosarcoma cells inhibited tumor growth and increased tumor cell apoptosis (Yang et al., 2021). Besides, some of the immune-related factors associated with tumor invasion and metastasis such as HMGB1 and Toll-like receptor 4 (TLR4) play a considerable role in rebuilding TIM and promoting the development of pyroptosis that could also be upregulated by hypoxia (Liu et al., 2015; Wu et al., 2018; Tan et al., 2020; Zha et al., 2020; Yuan et al., 2021). For example, HMGB1 promotes lung metastasis in triple-negative breast cancer by inducing CD62Ldim neutrophil polarization (Wang et al., 2020b). The prognostic role of TLR4 in Osteosarcoma remain controversial. As we know, TLR4 recognizes cathepsin K to activate the M2 polarization of tumor-associated macrophages (TAMs) through an mTOR-dependent pathway, resulting in tumor metastasis (Li et al., 2019). It has been reported that the activation of TLR4 prevents progression of osteosarcoma, while some other evidence demonstrated the TLR4 related pathway could promote the invasion and migration of Osteosarcoma (Wang et al., 2019b; Zhou et al., 2020b; Yahiro et al., 2020). Thus, the role of TLR4 in Osteosarcoma need further more investigations. Consistent with the above evidence, our study suggests that CD8+ T cells and CD4+ T memory-activated cells were decreased in patients with high hypoxia risk, indicating an immunosuppressive status. In our study, the expression of HMGB1 is higher in normal tissues than that in osteosarcoma from TARGET dataset. However, the immunofluorescence study showed high expression of HMGB1 in osteosarcoma. It has been reported that the expression of HMGB1 mRNA is higher in moderately and poorly differentiated gastrointestinal adenocarcinoma tissues compared to adjacent pre-cancerous and well-differentiated tissues (Xiang et al., 1997). It reminds that the expression of HMGB1 may be associated with the level of differentiation. Besides, there was also high degree of heterogeneity within individual tumors or between tumors (Hasegawa et al., 1991; Klangjorhor et al., 2020). These discordances may have resulted in the difference. These results show that our hypoxia risk model distinguishes immune status and immune cell subtypes in different patients and may be helpful in the selection of appropriate treatments. On the other hand, the above evidence indicates that hypoxia is a predominant driver of TIM remodeling, not only downregulating subset of infiltrating immune cells (e.g., CD8+ T cells and CD4+ T memory activated cells) but further regulating the immune response-related chemokines and their receptors and thus upregulating pyroptosis. Above all, hypoxia may initiate a series of chain reactions that lead to the suppressive immune status.
As a novel form of programmed cell death, pyroptosis has received increasing attention because of its dual regulation of the tumor immune response. In this study, we developed a pyroptosis risk score model based on DEGs related to pyroptosis that predicts prognosis, chemoresistance, and subtype of immune cells. ssGSEA revealed that the low pyroptosis risk score group had a low proportion of CD8+ T cells, Th1 cells, Th2 cells, and low activity in some immune pathways, indicating that pyroptosis also contributes to a suppressive microenvironment. Moreover, T helper cells and CD8+ T cells are regarded as crucial immune cells in antitumor immune response, and their large infiltration into tumor tissues is often linked with better prognosis (Fridman et al., 2012). The application of chemotherapy has improved the survival outcomes of patients with OS. Therefore, we evaluated the ability of the pyroptosis risk score to predict the sensitivity of conventional chemotherapies (Strauss et al., 2021). As previously reported, high pyroptosis scores were associated with lower chemosensitivity. After a series of analyses, we found that the low pyroptosis risk score was often associated with better clinical outcomes such as survival, chemosensitivity, and immune status, while the pyroptosis signature showed good predictive ability in these aspects. The above results demonstrate that hypoxia can also contribute to poor prognosis and low chemosensitivity by promoting pyroptosis.
In summary, this study has revealed the relationship between hypoxia, pyroptosis, and OS. Hypoxia is closely connected to the development of OS because the HRGs are differentially expressed in healthy and OS tissues, and could promote an immunosuppressive microenvironment by modulating the infiltration of immune cells and immune-related factors. In addition, our HRG and PRG signatures can predict the prognosis and immune landscape status in OS patients, thereby providing novel insights into OS tumorigenesis and a foundation for novel OS detection and treatment strategies.
In conclusion, we have established and validated a prognostic model based on HRGs in patients with OS that can also evaluate the characteristics of the tumor immune landscape. Moreover, another pyroptosis risk-score model that includes 14 PRGs provides independent prognostic and chemosensitivity capabilities. Through the complicated connections between hypoxia and pyroptosis, this study has demonstrated that hypoxia could alter poor prognosis and chemoresistance by affecting TIM remodeling and the occurrence of pyroptosis.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The patient data in this work were acquired from the publicly available datasets of patients whose informed consent were provided to the data holders. Ethical approval was granted by the Human Research Ethical Board of Third Xiangya Hospital, Central South University.
QZ and YL conceived the idea, designed and oversaw the experimental study. LH and XW undertook the analysis and wrote the draft. DC, ZC, and ZL made contributions to perform the experiments. YL provided guidance for the design of the experiment. QZ provided a critical revision of the article. All authors read and approved the final manuscript.
The Fundamental Research Funds for the Central Universities of Central South University under Grant (No. 2020zzts896); Qinghai Key Laboratory of Laboratory Medicine, Qingke Fazheng (2019) No. 105; the guiding project of Qinghai Provincial Health and Family Planning Commission (2018-wjzdx17). This work was supported by grants from the Project of Kunlun Elite, High-End Innovation and Entrepreneurship Talents of Qinghai Province (2021 No. 13). National Natural Science Foundation of China (82260365).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2022.814722/full#supplementary-material
DEGs, Differential expressed genes; DCN, Decorin; GEO, Gene Expression Omnibus; GO, gene ontology; GTEx, genotype-tissue expression database; GSEA, gene set enrichment analyses; GZMB, Granzyme B; HRGs, Hypoxia-related genes; HRG, hypoxia-related genes; HOMX1, Heme oxygenase-1; IFNG, Interferon Gamma; KEGG, kyoto encyclopedia of genes and genomes; LOX, lysyl oxidase; OS, Osteosarcoma; PRGs, pyroptosis related genes; PPI, The protein-protein interaction; PDK1, pyruvate dehydrogenase kinase 1; TARGET, Therapeutically Applicable Research to Generate Effective Treatments; TIM, tumor immune microenvironment; ssGSEA, Single sample GSEA; PBS, phosphate buffer solution; TLR4, Toll-like receptor 4; TAMs, tumor-associated macrophages; NPTX2, Neuronal pentraxin.
Ballatori, S. E., and Hinds, P. W. (2016). Osteosarcoma: Prognosis plateau warrants retinoblastoma pathway targeted therapy. Signal Transduct. Target. Ther. 1, 16001. doi:10.1038/sigtrans.2016.1
Chen, D. S., and Mellman, I. (2013). Oncology meets immunology: The cancer-immunity cycle. Immunity 39, 1–10. doi:10.1016/j.immuni.2013.07.012
Chen, G., Gu, H., Fang, T., Zhou, K., Xu, J., and Yin, X. (2020). Hypoxia-induced let-7f-5p/TARBP2 feedback loop regulates osteosarcoma cell proliferation and invasion by inhibiting the Wnt signaling pathway. Aging (Albany NY) 12, 6891–6903. doi:10.18632/aging.103049
Du, A., Wu, X., Gao, Y., Jiang, B., Wang, J., Zhang, P., et al. (2021). m6A regulator-mediated methylation modification patterns and tumor microenvironment infiltration characterization in acute myeloid leukemia. Front. Immunol. 12, 789914. doi:10.3389/fimmu.2021.789914
Fridman, W. H., Pagès, F., Sautès-Fridman, C., and Galon, J. (2012). The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 12, 298–306. doi:10.1038/nrc3245
Gibadulinova, A., Bullova, P., Strnad, H., Pohlodek, K., Jurkovicova, D., Takacova, M., et al. (2020). CAIX-mediated control of LIN28/let-7 Axis contributes to metabolic adaptation of breast cancer cells to hypoxia. Int. J. Mol. Sci. 21, 4299. doi:10.3390/ijms21124299
Goguet-Surmenian, E., Richard-Fiardo, P., Guillemot, E., Benchetrit, M., Gomez-Brouchet, A., Buzzo, P., et al. (2013). CXCR7-mediated progression of osteosarcoma in the lungs. Br. J. Cancer 109, 1579–1585. doi:10.1038/bjc.2013.482
Guan, G., Zhang, Y., Lu, Y., Liu, L., Shi, D., Wen, Y., et al. (2015). The HIF-1α/CXCR4 pathway supports hypoxia-induced metastasis of human osteosarcoma cells. Cancer Lett. 357, 254–264. doi:10.1016/j.canlet.2014.11.034
Hasegawa, T., Hirose, T., Kudo, E., Hizawa, K., Usui, M., and Ishii, S. (1991). Immunophenotypic heterogeneity in osteosarcomas. Hum. Pathol. 22, 583–590. doi:10.1016/0046-8177(91)90236-i
Hou, J., Zhao, R., Xia, W., Chang, C.-W., You, Y., Hsu, J.-M., et al. (2020). PD-L1-mediated gasdermin C expression switches apoptosis to pyroptosis in cancer cells and facilitates tumour necrosis. Nat. Cell Biol. 22, 1264–1275. doi:10.1038/s41556-020-0575-z
Iyer, N. V., Kotch, L. E., Agani, F., Leung, S. W., Laughner, E., Wenger, R. H., et al. (1998). Cellular and developmental control of O2 homeostasis by hypoxia-inducible factor 1 alpha. Genes Dev. 12, 149–162. doi:10.1101/gad.12.2.149
Ji, F., Wang, Y., Qiu, L., Li, S., Zhu, J., Liang, Z., et al. (2013). Hypoxia inducible factor 1α-mediated LOX expression correlates with migration and invasion in epithelial ovarian cancer. Int. J. Oncol. 42, 1578–1588. doi:10.3892/ijo.2013.1878
Jiang, D., Wu, X., Sun, X., Tan, W., Dai, X., Xie, Y., et al. (2022). Bone mesenchymal stem cell-derived exosomal microRNA-7-5p inhibits progression of acute myeloid leukemia by targeting OSBPL11. J. Nanobiotechnology 20, 29. doi:10.1186/s12951-021-01206-7
Jiang, Y., Liu, H., Yu, H., Zhou, Y., Zhang, J., Xin, W., et al. (2021). Circular RNA Calm4 regulates hypoxia-induced pulmonary arterial smooth muscle cells pyroptosis via the circ-calm4/miR-124-3p/PDCD6 Axis. Arterioscler. Thromb. Vasc. Biol. 41, 1675–1693. doi:10.1161/ATVBAHA.120.315525
Jiao, Y., Zhao, H., Chen, G., Sang, X., Yang, L., Hou, Z., et al. (2018). Pyroptosis of MCF7 cells induced by the secreted factors of hUCMSCs. Stem Cells Int. 2018, 5912194. doi:10.1155/2018/5912194
Jo, V. Y., and Fletcher, C. D. M. (2014). WHO classification of soft tissue tumours: An update based on the 2013 (4th) edition. Pathology 46, 95–104. doi:10.1097/PAT.0000000000000050
Jorgensen, I., Zhang, Y., Krantz, B. A., and Miao, E. A. (2016). Pyroptosis triggers pore-induced intracellular traps (PITs) that capture bacteria and lead to their clearance by efferocytosis. J. Exp. Med. 213, 2113–2128. doi:10.1084/jem.20151613
Klangjorhor, J., Chaiyawat, P., Teeyakasem, P., Sirikaew, N., Phanphaisarn, A., Settakorn, J., et al. (2020). Mycophenolic acid is a drug with the potential to be repurposed for suppressing tumor growth and metastasis in osteosarcoma treatment. Int. J. Cancer 146, 3397–3409. doi:10.1002/ijc.32735
Kumar, V., and Gabrilovich, D. I. (2014). Hypoxia-inducible factors in regulation of immune responses in tumour microenvironment. Immunology 143, 512–519. doi:10.1111/imm.12380
Lang, X., Green, M. D., Wang, W., Yu, J., Choi, J. E., Jiang, L., et al. (2019). Radiotherapy and immunotherapy promote tumoral lipid oxidation and ferroptosis via synergistic repression of SLC7A11. Cancer Discov. 9, 1673–1685. doi:10.1158/2159-8290.CD-19-0338
Li, R., Zhou, R., Wang, H., Li, W., Pan, M., Yao, X., et al. (2019). Gut microbiota-stimulated cathepsin K secretion mediates TLR4-dependent M2 macrophage polarization and promotes tumor metastasis in colorectal cancer. Cell Death Differ. 26, 2447–2463. doi:10.1038/s41418-019-0312-y
Li, X., Fang, P., Mai, J., Choi, E. T., Wang, H., and Yang, X. F. (2013). Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers. J. Hematol. Oncol. 6, 19. doi:10.1186/1756-8722-6-19
Li, Y., Flores, R., Yu, A., Okcu, M. F., Murray, J., Chintagumpala, M., et al. (2011). Elevated expression of CXC chemokines in pediatric osteosarcoma patients. Cancer 117, 207–217. doi:10.1002/cncr.25563
Lilienthal, I., and Herold, N. (2020). Targeting molecular mechanisms underlying treatment efficacy and resistance in osteosarcoma: A review of current and future strategies. Int. J. Mol. Sci., 21, 6885. doi:10.3390/ijms21186885
Lin, W., Wu, S., Chen, X., Ye, Y., Weng, Y., Pan, Y., et al. (2020). Characterization of hypoxia signature to evaluate the tumor immune microenvironment and predict prognosis in glioma groups. Front. Oncol. 10, 796. doi:10.3389/fonc.2020.00796
Liu, D., Vadgama, J., and Wu, Y. (2021). Basal-like breast cancer with low TGFβ and high TNFα pathway activity is rich in activated memory CD4 T cells and has a good prognosis. Int. J. Biol. Sci. 17, 670–682. doi:10.7150/ijbs.56128
Liu, Y., Wu, J., Huang, W., Weng, S., Wang, B., Chen, Y., et al. (2020). Development and validation of a hypoxia-immune-based microenvironment gene signature for risk stratification in gastric cancer. J. Transl. Med. 18, 201. doi:10.1186/s12967-020-02366-0
Liu, Y., Yan, W., Tohme, S., Chen, M., Fu, Y., Tian, D., et al. (2015). Hypoxia induced HMGB1 and mitochondrial DNA interactions mediate tumor growth in hepatocellular carcinoma through Toll-like receptor 9. J. Hepatol. 63, 114–121. doi:10.1016/j.jhep.2015.02.009
Lossos, I. S., Czerwinski, D. K., Alizadeh, A. A., Wechser, M. A., Tibshirani, R., Botstein, D., et al. (2004). Prediction of survival in diffuse large-B-cell lymphoma based on the expression of six genes. N. Engl. J. Med. 350, 1828–1837. doi:10.1056/NEJMoa032520
Madamanchi, N. R., and Runge, M. S. (2007). Mitochondrial dysfunction in atherosclerosis. Circ. Res. 100, 460–473. doi:10.1161/01.RES.0000258450.44413.96
Majmundar, A. J., Wong, W. J., and Simon, M. C. (2010). Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 40, 294–309. doi:10.1016/j.molcel.2010.09.022
Man, S. M. (2018). Inflammasomes in the gastrointestinal tract: Infection, cancer and gut microbiota homeostasis. Nat. Rev. Gastroenterol. Hepatol. 15, 721–737. doi:10.1038/s41575-018-0054-1
Newman, A. M., Liu, C. L., Green, M. R., Gentles, A. J., Feng, W., Xu, Y., et al. (2015). Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 12, 453–457. doi:10.1038/nmeth.3337
Niu, J., Yan, T., Guo, W., Wang, W., and Zhao, Z. (2019). Insight into the role of autophagy in osteosarcoma and its therapeutic implication. Front. Oncol. 9, 1232. doi:10.3389/fonc.2019.01232
Prudowsky, Z. D., and Yustein, J. T. (2020). Recent insights into therapy resistance in osteosarcoma. Cancers (Basel), 13, 83. doi:10.3390/cancers13010083
Ritter, J., and Bielack, S. S. (2010). Osteosarcoma. Ann. Oncol. 21 (7), vii320–5. doi:10.1093/annonc/mdq276
Rosen, G. (1985). Preoperative (neoadjuvant) chemotherapy for osteogenic sarcoma: A ten year experience. Orthopedics 8, 659–664. doi:10.3928/0147-7447-19850501-19
Russell, J. H., and Ley, T. J. (2002). Lymphocyte-mediated cytotoxicity. Annu. Rev. Immunol. 20, 323–370. doi:10.1146/annurev.immunol.20.100201.131730
Sainio, A. O., and Järveläinen, H. T. (2019). Decorin-mediated oncosuppression - a potential future adjuvant therapy for human epithelial cancers. Br. J. Pharmacol. 176, 5–15. doi:10.1111/bph.14180
Scharping, N. E., Rivadeneira, D. B., Menk, A. V., Vignali, P. D. A., Ford, B. R., Rittenhouse, N. L., et al. (2021). Mitochondrial stress induced by continuous stimulation under hypoxia rapidly drives T cell exhaustion. Nat. Immunol. 22, 205–215. doi:10.1038/s41590-020-00834-9
Semenza, G. L. (1999). Regulation of mammalian O2 homeostasis by hypoxia-inducible factor 1. Annu. Rev. Cell Dev. Biol. 15, 551–578. doi:10.1146/annurev.cellbio.15.1.551
Shi, J., Zhao, Y., Wang, K., Shi, X., Wang, Y., Huang, H., et al. (2015). Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 526, 660–665. doi:10.1038/nature15514
Siu, M. K. Y., Jiang, Y.-X., Wang, J.-J., Leung, T. H. Y., Ngu, S. F., Cheung, A. N. Y., et al. (2020). PDK1 promotes ovarian cancer metastasis by modulating tumor-mesothelial adhesion, invasion, and angiogenesis via α5β1 integrin and JNK/IL-8 signaling. Oncogenesis 9, 24. doi:10.1038/s41389-020-0209-0
Smeland, S., Bielack, S. S., Whelan, J., Bernstein, M., Hogendoorn, P., Krailo, M. D., et al. (2019). Survival and prognosis with osteosarcoma: Outcomes in more than 2000 patients in the EURAMOS-1 (European and American osteosarcoma study) cohort. Eur. J. Cancer 109, 36–50. doi:10.1016/j.ejca.2018.11.027
Song, X., Liu, X., Chi, W., Liu, Y., Wei, L., Wang, X., et al. (2006). Hypoxia-induced resistance to cisplatin and doxorubicin in non-small cell lung cancer is inhibited by silencing of HIF-1alpha gene. Cancer Chemother. Pharmacol. 58, 776–784. doi:10.1007/s00280-006-0224-7
Strauss, S. J., Frezza, A. M., Abecassis, N., Bajpai, J., Bauer, S., Biagini, R., et al. (2021). Bone sarcomas: ESMO-EURACAN-GENTURIS-ERN PaedCan clinical practice guideline for diagnosis, treatment and follow-up. Ann. Oncol. 32, 1520–1536. doi:10.1016/j.annonc.2021.08.1995
Szklarczyk, D., Franceschini, A., Wyder, S., Forslund, K., Heller, D., Huerta-Cepas, J., et al. (2015). STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 43, D447–D452. doi:10.1093/nar/gku1003
Tan, G., Huang, C., Chen, J., and Zhi, F. (2020). HMGB1 released from GSDME-mediated pyroptotic epithelial cells participates in the tumorigenesis of colitis-associated colorectal cancer through the ERK1/2 pathway. J. Hematol. Oncol. 13, 149. doi:10.1186/s13045-020-00985-0
Tang, R., Xu, J., Zhang, B., Liu, J., Liang, C., Hua, J., et al. (2020). Ferroptosis, necroptosis, and pyroptosis in anticancer immunity. J. Hematol. Oncol. 13, 110. doi:10.1186/s13045-020-00946-7
Tsuchiya, K. (2021). Switching from apoptosis to pyroptosis: Gasdermin-elicited inflammation and antitumor immunity. Int. J. Mol. Sci. 22, 426. doi:10.3390/ijms22010426
Umezaki, N., Nakagawa, S., Yamashita, Y.-I., Kitano, Y., Arima, K., Miyata, T., et al. (2019). Lysyl oxidase induces epithelial-mesenchymal transition and predicts intrahepatic metastasis of hepatocellular carcinoma. Cancer Sci. 110, 2033–2043. doi:10.1111/cas.14010
Wang, N., Meng, X., Liu, Y., Chen, Y., and Liang, Q. (2019). LPS promote Osteosarcoma invasion and migration through TLR4/HOTAIR. Gene 680, 1–8. doi:10.1016/j.gene.2018.09.031
Wang, Y., Zhang, H., Chen, Q., Jiao, F., Shi, C., Pei, M., et al. (2020). TNF-α/HMGB1 inflammation signalling pathway regulates pyroptosis during liver failure and acute kidney injury. Cell Prolif. 53, e12829. doi:10.1111/cpr.12829
Wang, Y., Zhu, X., Yuan, S., Wen, S., Liu, X., Wang, C., et al. (2019). TLR4/NF-κB signaling induces GSDMD-related pyroptosis in tubular cells in diabetic kidney disease. Front. Endocrinol. 10, 603. doi:10.3389/fendo.2019.00603
Wang, Z., Yang, C., Li, L., Jin, X., Zhang, Z., Zheng, H., et al. (2020). Tumor-derived HMGB1 induces CD62Ldim neutrophil polarization and promotes lung metastasis in triple-negative breast cancer. Oncogenesis 9, 82. doi:10.1038/s41389-020-00267-x
Waza, A. A., Hamid, Z., Ali, S., Bhat, S. A., and Bhat, M. A. (2018). A review on heme oxygenase-1 induction: Is it a necessary evil. Inflamm. Res. 67, 579–588. doi:10.1007/s00011-018-1151-x
Wu, G., Xu, G., Chen, D.-W., Gao, W.-X., Xiong, J.-Q., Shen, H.-Y., et al. (2018). Hypoxia exacerbates inflammatory acute lung injury via the toll-like receptor 4 signaling pathway. Front. Immunol. 9, 1667. doi:10.3389/fimmu.2018.01667
Wu, X., Zhang, X., Feng, W., Feng, H., Ding, Z., Zhao, Q., et al. (2021). A targeted erythrocyte membrane-encapsulated drug-delivery system with anti-osteosarcoma and anti-osteolytic effects. ACS Appl. Mat. Interfaces 13, 27920–27933. doi:10.1021/acsami.1c06059
Wu, Z. H., Li, Z. W., Yang, D. L., and Liu, J. (2021). Development and validation of a pyroptosis-related long non-coding RNA signature for hepatocellular carcinoma. Front. Cell Dev. Biol. 9, 713925. doi:10.3389/fcell.2021.713925
Wu, Z., Zhang, X., Chen, D., Li, Z., Wu, X., Wang, J., et al. (2021). N6-Methyladenosine-Related LncRNAs are potential remodeling indicators in the tumor microenvironment and prognostic markers in osteosarcoma. Front. Immunol. 12, 806189. doi:10.3389/fimmu.2021.806189
Xia, X., Wang, X., Cheng, Z., Qin, W., Lei, L., Jiang, J., et al. (2019). The role of pyroptosis in cancer: Pro-cancer or pro-"host. Cell Death Dis. 10, 650. doi:10.1038/s41419-019-1883-8
Xiang, Y. Y., Wang, D. Y., Tanaka, M., Suzuki, M., Kiyokawa, E., Igarashi, H., et al. (1997). Expression of high-mobility group-1 mRNA in human gastrointestinal adenocarcinoma and corresponding non-cancerous mucosa. Int. J. Cancer 74, 1–6. doi:10.1002/(sici)1097-0215(19970220)74:1<1::aid-ijc1>3.0.co;2-6
Xu, L., Deng, C., Pang, B., Zhang, X., Liu, W., Liao, G., et al. (2018). Tip: A web server for resolving tumor Immunophenotype profiling. Cancer Res. 78, 6575–6580. doi:10.1158/0008-5472.CAN-18-0689
Yahiro, K., Matsumoto, Y., Yamada, H., Endo, M., Setsu, N., Fujiwara, T., et al. (2020). Activation of TLR4 signaling inhibits progression of osteosarcoma by stimulating CD8-positive cytotoxic lymphocytes. Cancer Immunol. Immunother. 69, 745–758. doi:10.1007/s00262-020-02508-9
Yang, H., Zhao, L., Zhang, Y., and Li, F. F. (2021). A comprehensive analysis of immune infiltration in the tumor microenvironment of osteosarcoma. Cancer Med. 10, 5696–5711. doi:10.1002/cam4.4117
Yang, L., Xie, M., Yang, M., Yu, Y., Zhu, S., Hou, W., et al. (2014). PKM2 regulates the Warburg effect and promotes HMGB1 release in sepsis. Nat. Commun. 5, 4436. doi:10.1038/ncomms5436
Yoshida, K., Okamoto, M., Sasaki, J., Kuroda, C., Ishida, H., Ueda, K., et al. (2019). Clinical outcome of osteosarcoma and its correlation with programmed death-ligand 1 and T cell activation markers. Onco. Targets. Ther. 12, 2513–2518. doi:10.2147/OTT.S198421
Yu, L. M., Zhang, W. H., Han, X. X., Li, Y. Y., Lu, Y., Pan, J., et al. (2019). Hypoxia-induced ROS contribute to myoblast pyroptosis during obstructive sleep apnea via the NF-κB/HIF-1α signaling pathway. Oxid. Med. Cell. Longev. 2019, 4596368. doi:10.1155/2019/4596368
Yuan, R., Zhao, W., Wang, Q.-Q., He, J., Han, S., Gao, H., et al. (2021). Cucurbitacin B inhibits non-small cell lung cancer in vivo and in vitro by triggering TLR4/NLRP3/GSDMD-dependent pyroptosis. Pharmacol. Res. 170, 105748. doi:10.1016/j.phrs.2021.105748
Zeng, F., Zhang, Y., Han, X., Zeng, M., Gao, Y., and Weng, J. (2021). Employing hypoxia characterization to predict tumour immune microenvironment, treatment sensitivity and prognosis in hepatocellular carcinoma. Comput. Struct. Biotechnol. J. 19, 2775–2789. doi:10.1016/j.csbj.2021.03.033
Zeng, W., Wan, R., Zheng, Y., Singh, S. R., and Wei, Y. (2011). Hypoxia, stem cells and bone tumor. Cancer Lett. 313, 129–136. doi:10.1016/j.canlet.2011.09.023
Zha, C., Meng, X., Li, L., Mi, S., Qian, D., Li, Z., et al. (2020). Neutrophil extracellular traps mediate the crosstalk between glioma progression and the tumor microenvironment via the HMGB1/RAGE/IL-8 axis. Cancer Biol. Med. 17, 154–168. doi:10.20892/j.issn.2095-3941.2019.0353
Zhang, Z., Zhang, Y., Xia, S., Kong, Q., Li, S., Liu, X., et al. (2020). Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature 579, 415–420. doi:10.1038/s41586-020-2071-9
Zhou, J., Liu, Q., Qian, R., Liu, S., Hu, W., and Liu, Z. (2020). Paeonol antagonizes oncogenesis of osteosarcoma by inhibiting the function of TLR4/MAPK/NF-κB pathway. Acta Histochem. 122, 151455. doi:10.1016/j.acthis.2019.151455
Keywords: hypoxia, osteosarcoma, pyroptosis, drug resistance, prognosis
Citation: Hu L, Wu X, Chen D, Cao Z, Li Z, Liu Y and Zhao Q (2022) The hypoxia-related signature predicts prognosis, pyroptosis and drug sensitivity of osteosarcoma. Front. Cell Dev. Biol. 10:814722. doi: 10.3389/fcell.2022.814722
Received: 14 November 2021; Accepted: 29 August 2022;
Published: 20 September 2022.
Edited by:
Frédéric Lézot, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Weiting Qin, Shanghai Jiao Tong University, ChinaCopyright © 2022 Hu, Wu, Chen, Cao, Li, Liu and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yanmin Liu, MTM5OTcyNDU2OTlAMTYzLmNvbQ==; Qiangqiang Zhao, emd4eXdzQDE2My5jb20=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.