 Jingliang Gu
Jingliang Gu Wu Rao1†
Wu Rao1† Deta Chen
Deta Chen
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol. , 13 December 2022
Sec. Molecular and Cellular Pathology
Volume 10 - 2022 | https://doi.org/10.3389/fcell.2022.1092776
This article is part of the Research Topic New Insight on Development and Degeneration of Bone and Cartilage: Molecular Mechanisms and Translational Medicine View all 7 articles
During the last decade, osteoarthritis (OA) has become one of the most prevalent musculoskeletal diseases worldwide. OA is characterized by progressive loss of articular cartilage, abnormal remodeling of subchondral bone, hyperplasia of synovial cells, and growth of osteophytes, which lead to chronic pain and disability. The pathological mechanisms underlying OA initiation and progression are still poorly understood. Non-coding RNAs (ncRNAs) constitute a large portion of the transcriptome that do not encode proteins but function in numerous biological processes. Cumulating evidence has revealed a strong association between the changes in expression levels of ncRNA and the disease progression of OA. Moreover, loss- and gain-of-function studies utilizing transgenic animal models have demonstrated that ncRNAs exert vital functions in regulating cartilage homeostasis, degeneration, and regeneration, and changes in ncRNA expression can promote or decelerate the progression of OA through distinct molecular mechanisms. Recent studies highlighted the potential of ncRNAs to serve as diagnostic biomarkers, prognostic indicators, and therapeutic targets for OA. MiRNAs and lncRNAs are two major classes of ncRNAs that have been the most widely studied in cartilage tissues. In this review, we focused on miRNAs and lncRNAs and provided a comprehensive understanding of their functional roles as well as molecular mechanisms in cartilage homeostasis and OA pathogenesis.
The maintenance of cartilage homeostasis is crucial for our joint function because the loss of cartilage homeostasis can cause extracellular matrix (ECM) degradation and chondrocyte death, leading to irreversible cartilage damage and the onset of osteoarthritis (OA) (Tong et al., 2022). During the last decade, OA has become the most prevalent degenerative joint condition affecting more than 25% of the population over 18 years of age (Chen et al., 2017). Typical pathological changes in an OA joint include gradual loss and degradation of articular cartilage, sclerosis of the subchondral bone, growth of osteophytes, synovial inflammation, ligament, and meniscal lesions, and enlargement of the joint capsule (Chen et al., 2017; Snoeker et al., 2020; Tong et al., 2022). The risk factors for developing OA include genetic susceptibility, aging, obesity, and joint injury (Palazzo et al., 2016). There are currently no effective therapies to repair damaged cartilage or slow the progression of the disease since the precise molecular mechanisms of OA pathogenesis are still poorly understood. Interestingly, cumulating evidence has recently revealed that non-coding RNAs (ncRNAs) play multiple central roles in maintaining cartilage homeostasis and are also deeply involved in OA pathogenesis (Razmara et al., 2019; Xie et al., 2020). The ncRNAs refer to functional RNAs that are transcribed from DNA but not translated into proteins (Mattick and Makunin, 2006). It is well-known that the ncRNAs can function as epigenetic regulators in regulating gene and protein expressions and thus participate in numerous fundamental biological processes (Wei et al., 2017). Epigenetic-related ncRNAs include microRNA (miRNA), small interfering RNA (siRNA), piwi-interacting RNA (piRNA), circular RNA (circRNA), and long non-coding RNA (lncRNA) (Wei et al., 2017). The ncRNAs can act as vital regulators in cartilage formation and homeostasis by mediating a series of physiological processes, such as chondrocyte proliferation, differentiation, and ECM biosynthesis (Gibson and Asahara, 2013; Huang et al., 2019; Razmara et al., 2019). Dysregulation in ncRNAs expression can lead to ECM degradation, chondrocyte hypertrophy, senescence, and apoptosis, ultimately resulting in OA initiation and progression (Weilner et al., 2015; Xie et al., 2020; Pekacova et al., 2022). Recent studies underscored the potential of ncRNAs as diagnostic biomarkers, prognostic indicators, and therapeutic targets for OA (Ghafouri-Fard et al., 2021).
MiRNAs and lncRNAs are two major classes of ncRNAs that have been the most widely studied in the regulatory mechanisms of skeletal homeostasis and diseases. MiRNAs are small non-coding RNAs (∼21–24 nucleotides in length) with essential biological activities, such as post-translational regulation of the gene expression (Saquib et al., 2021). The synthesis of miRNAs begins with the transcription of DNA sequences into primary miRNAs (pri-miRNAs). Then the pri-miRNAs are processed into precursor miRNAs (pre-miRNAs) and eventually mature miRNAs through several sophisticated molecular mechanisms (O'Brien et al., 2018; Adams, 2017; Joyce et al., 2018). LncRNAs, which are RNA transcripts longer than 200 nucleotides, represent a considerable portion of ncRNAs. The biogenesis of lncRNA is highly similar to that of messenger RNAs (mRNAs), a process that includes polymerase II-mediated transcription, polyadenylation, splicing, and 5′-capping (Kung et al., 2013; Aliperti et al., 2021). The processes controlling lncRNA synthesis are cell type- and stage-specific and governed by cell type- and stage-specific stimuli (Dahariya et al., 2019). Once lncRNAs finish their transcription, they fold into thermodynamically stable secondary structures and display distinct expression patterns and specific nuclear localization, which differs from mRNAs (Saxena and Carninci, 2011). In this review, we focused on miRNAs and lncRNAs and provided a comprehensive understanding of their functional roles in cartilage homeostasis and OA pathogenesis.
Recent studies have highlighted a series of crucial functions of miRNAs in maintaining cartilage homeostasis (Razmara et al., 2019; Fujii et al., 2022). In vitro chondrogenesis of mesenchymal stem cells is the most common experiment used to study the role of miRNAs in chondrocyte proliferation and chondrogenic differentiation. Meng and coworkers showed that miR-193b-3p expression was elevated in chondrogenic and hypertrophic human mesenchymal stem cells (hMSC) and was significantly decreased in degenerated human cartilage. MiR-193b-3p regulates hMSC chondrogenesis and the metabolism of primary human chondrocytes by directly targeting the 3′-untranslated region (3′-UTR) of histone deacetylase 3 (HDAC3) mRNA (Meng et al., 2018). MiR-200a was reported to have a role in controlling the proliferation and differentiation of mandibular condylar chondrocytes (MCCs) (Umeda et al., 2015). Transfection of miR-200a mimics inhibits MCC differentiation and promotes cell proliferation, while miR-200a inhibitors enhance MCC differentiation. MiR-9 regulates the survival of chondroblasts by targeting protogenin and thus maintains the cartilage homeostasis (Song et al., 2013). MiR-142-3p inhibits chondrocyte apoptosis and inflammation through targeting the expression of high mobility group box 1 (HMGB1), an essential pathological factor leading to the loss of cartilage homeostasis (Wang et al., 2016). MiR-140 is specifically expressed in healthy cartilage and has crucial functions in maintaining ECM homeostasis, probably by promoting the production of type II collagen and inhibiting the expressions of matrix metalloproteinase 13 (Mmp13) and a disintegrin and metalloproteinase with thrombospondin motifs 5 (Adamts5) (Si et al., 2017). Using genetically modified mouse models, Huang et al. showed that two homologous miRNAs, miR-204 and miR-211, synergistically maintain cartilage homeostasis and protect cartilage from OA-like lesions (Huang et al., 2019). The absence of miR-204/-211 in mesenchymal progenitor cells (MPCs) causes abnormal accumulation of Runx2, a vital transcriptional factor for promoting chondrocyte hypertrophy in multi-type joint cells, which leads to OA-like degeneration of the whole joint. Moreover, loss of miR-204/-211 expression strongly stimulates matrix-degrading proteases, such as Mmp13 and Adamts5 in articular chondrocytes and synoviocytes and thus promote the degradation of articular cartilage. Furthermore, overexpression of miR-204 in the articular joint essentially restores cartilage homeostasis and decelerates the progression of OA in mice. Both strands of miR-455, i.e., miR-455-5p and miR-455-3p, are highly expressed in chondrocytes of healthy articular cartilage (Ito et al., 2021). Genetic ablation of miR-455 disrupts cartilage homeostasis and results in considerable OA-like damages, whereas overexpression of both strands of miR-455 protects against surgery-induced cartilage degradation in mice (Ito et al., 2021). Zhang et al. recently reported that miR-17 is strongly expressed in chondrocytes of both superficial and middle zone cartilages under homeostatic conditions (Zhang et al., 2022). The expression of miR-17 is essential for maintaining the balance between ECM anabolism and catabolism.
In response to mechanical loading, chondrocyte-mediated mechanotransduction is necessary for cartilage health and homeostasis (Campbell et al., 2007). Recent studies have revealed the participation of miRNAs in mechanotransduction within articular cartilage. By utilizing the miRNA microarray technology, the expression pattern of miRNAs in the anterior weight-bearing zone and posterior non-weight-bearing area of bovine articular cartilage was analyzed (Dunn et al., 2009). The data showed that miR-221 and miR-222 were markedly higher in the anterior weight-bearing medical condylar cartilage compared to the posterior non-weight-bearing medial condylar cartilage, implicating potential roles of these miRNAs in chondrocyte mechanotransduction. Shang and coworkers further studied the extracellular vehicles (EVs)-mediated intercellular communication between chondrocytes and osteoblasts (Shang et al., 2021). They found that chondrocyte-derived EVs contain miR-221-3p, and the latter can be transferred to osteoblasts to regulate gene expressions (Shang et al., 2021). Collectively, these findings suggest that the expressions of specific miRNA clusters are necessary for the maintenance of cartilage homeostasis. Loss of these homeostatic miRNAs may lead to cartilage degeneration and OA.
Articular cartilage has a limited potential for self-repair. Cartilage regeneration technology has been developed to produce resilient cartilage-like tissue where cartilage has been worn away or destroyed (Xiang et al., 2022). Surgical intervention is often required for cartilage regeneration (Murphy et al., 2020). The most common arthroscopic treatment for cartilage injuries is chondroplasty or removing loose cartilage fragments (Martin et al., 2019). This gives temporary symptomatic alleviation; nevertheless, the remaining cartilage is more prone to wear and rapid degeneration. Microfracture is another frequently used method that pierces the subchondral bone to let bone marrow fill the cartilage defects. This method leads to the production of fibrocartilage tissue that is mechanically inferior. Recent research has underscored miRNAs as a potential therapeutic agent for promoting cartilage regeneration (Foo et al., 2021). For instance, Zhu and colleagues have developed a hydrogel-based miRNAs delivery system to regenerate damaged cartilage by providing a regenerative milieu to limit chondrocyte senescence that predominantly leads to cartilage lesions in OA (Zhu et al., 2022). MiR-23a-3p is the most highly expressed in human umbilical cord mesenchymal stem cells-derived small extracellular vesicles. The latter demonstrates an effect in promoting cartilage regeneration by transferring miR-23a-3p (Hu et al., 2020). In a rat OA model, exosomes origin from miR-140-5p-overexpressing human synovial mesenchymal stem cells significantly promote cartilage regeneration and decelerate knee OA progression (Tao et al., 2017). Furthermore, Wang et al. have shown a strong cartilage regenerative capacity of the miR-221-3p-containing EVs, which are derived from chondrogenic progenitor cells of MRL/MpJ superhealer mice, in the destabilization of the medial meniscus (DMM)-induced OA model (Wang et al., 2020).
The miRNA profiles during the initiation and progression of OA have been characterized in human OA cartilage and two surgically-induced mouse OA models, including the DMM model and the anterior cruciate ligament transection (ALCT) model. By serum miRNAs microarray analysis, Ntoumou et al. identified 279 differentially expressed miRNAs in serum samples of OA patients as compared with healthy controls, among which miR-140-3p, miR-33b-3p, and miR-571-3p could be used as potential biomarkers of OA (Ntoumou et al., 2017). Almeida and colleagues performed RNA sequencing in human-preserved and lesioned OA cartilage to profile the changes in miRNA and mRNA (Coutinho de Almeida et al., 2019). Their data showed a total of 142 miRNAs and 2387 mRNA that were differentially expressed between preserved and damaged OA cartilage. By bioinformatic approaches, a miRNA-mRNA interactome network was drawn consisting of 62 miRNAs targeting 238 mRNA to elucidate the molecular regulatory network during OA pathogenesis (Coutinho de Almeida et al., 2019). In the DMM model, a total of 139 miRNAs were determined to be differentially expressed in OA articular cartilage at one and/or 6 weeks after surgery (Kung et al., 2018). A paired miRNA/mRNA expression analysis confirmed an overlapping dysregulation of miRNAs between mouse OA cartilage and human end-staged OA cartilage. The overlapped miRNAs include miR-15/16-5p, miR-26p-5p, miR-30c-5p, miR-98-5p, miR-149-5p, miR-210-3p and miR-342-3p. Moreover, some unreported OA-associated miRNAs were discovered, including miR-574-5p, miR-31-5p, and let-7d-5p. In a rat ALCT model, Cheng and colleagues analyzed the miRNA profile after ALCT surgery with or without extracorporeal shockwave (SW) therapy (Cheng et al., 2016). By the next-generation sequencing technology, 118 differentially expressed miRNAs were identified in articular cartilage, and 214 differentially expressed miRNAs were identified in the subchondral bone. Global functional analysis revealed that the identified miRNAs were associated with cartilage development, inflammatory response, cell adhesion, transcription and translation, cell cycle, signal transduction, apoptotic process, collagen fibril organization, and chondrocyte differentiation (Cheng et al., 2016). Moreover, Zheng et al. demonstrated a significantly higher level of serum miR-98 expression in OA patients than in healthy individuals, suggesting miR-98 as a potential diagnostic biomarker for OA (Zheng et al., 2018). In a rat model of OA, upregulated expression of miR-98 was also observed, and the knockdown of miR-98 had an inhibitory effect on cartilage deterioration and chondrocyte death (Wang et al., 2016b).
Low-grade inflammation is a key player in OA pathogenesis (Robinson et al., 2016). IL-1β is the most important pro-inflammatory cytokine during early OA onset, and IL-1β treatment has been used to stimulate chondrocyte apoptosis and ECM catabolism to mimic OA phenotype in vitro (Wojdasiewicz et al., 2014). Cumulating evidence has revealed the participation of miRNAs in IL-1β-stimulated OA-like phenotypes in articular chondrocytes. For instance, miR-377-3p alleviates the chondrocyte apoptosis under IL-1β stimulation in vitro and the damage of synovial tissues in facet joint OA in vivo (Tu et al., 2020). An association between increased miR-448 and decreased expression of matrilin-3 has been found in human OA cartilage tissues compared to normal cartilage (Yang et al., 2018). The deletion of miR-448 significantly ameliorates the IL-1β-induced catabolic phenotype in primary cultured chondrocytes, whereas overexpression of miR-448 shows the opposite effects (Yang et al., 2018). Martilin-3 is the target gene of miR-448, and the genetic deletion of matrilin-3 reverses the regulatory effects of miR-448 on the chondrocyte catabolism (Yang et al., 2018). The expression level of miR-320 is reduced in the IL-1β-treated primary mouse chondrocytes (PMCs) (Meng et al., 2016). Forced expression of miR-320 inhibits the expression of matrix proteinase Mmp13 by targeting its 3′-UTR of mRNA, whereas anti-miR-320 treatment enhances the Mmp13 expression (Meng et al., 2016). Activating the NFκB/MAPK pathway downregulates the miR-320 expression in the IL-1β-treated PMCs (Meng et al., 2016). MiR-320c inhibits the expression and transcriptional activity of β-catenin, whereas loss of miR-320c expression leads to OA-like phenotypes in chondrocytes and late-stage chondrogenic differentiation in human adipose-derived stem cells (Zhang et al., 2018a; Hu et al., 2019a). Injection of miR-320-3p agonist attenuates OA progression in the OA mouse model (Hu et al., 2019a). However, controversial results have also been reported. Peng et al. reported an increased miR-320a expression in OA chondrocytes as compared with normal chondrocytes. They further showed that transfection of miR-320 antagonists inhibits the Mmp13 expression in human OA chondrocytes in vitro by regulating the expressions of BMI-1 and Runx2 mRNA (Peng et al., 2017). The possible reasons for these controversial results could be the small sample size (N = 5–6) in these studies and the heterogeneity of primary human chondrocytes isolated from OA patients. IL-1β treatment downregulates the level of miR-27a-3p and increases the expression of its target gene ADAMTS5 mRNA in the primary human chondrocytes (Li et al., 2018). Overexpression of miR-27a-3p abolishes the expression of Adamts5 induced by the IL-1β treatment (Li et al., 2018). Furthermore, Qiu and colleagues have demonstrated that miR-27a suppresses the inflammatory response and catabolic activity during IL-1β stimulation in chondrocytes by targeting toll-like receptor 4 (TLR4) (Qiu et al., 2019). Transfection of miR-27a mimics significantly reduces the production of reactive oxygen species (ROS) and a series of pro-inflammatory cytokines, including IL-6, IL-8, and TNF-α in chondrocytes (Qiu et al., 2019).
TNF-α is another crucial inflammatory cytokine in OA pathogenesis (Robinson et al., 2016). Hu et al. reported a role of miR-145 in TNF-α-driven cartilage matrix degradation during the OA progression (Hu et al., 2017). MiR-145 expression is dramatically decreased in TNF-α-treated chondrocytes and in OA cartilages (Hu et al., 2017). MiR-145 could directly target the 3′UTR of mitogen-activated protein kinase 4 (MKK4) mRNA to inhibit the production of several TNF-α-induced matrix-degrading proteinases, including Mmp3, Mmp13, and Adamts5 (Hu et al., 2017). Intraarticular injection of miR-145 alleviates cartilage degradation and matrix catabolism in a rat DMM model (Hu et al., 2017).
Lipopolysaccharide (LPS) is one of the most crucial pro-inflammatory factors, stimulating strong inflammatory responses in synovial tissues and cartilage in arthritic diseases. In LPS-stimulated primary chondrocytes, Ding et al. have shown that overexpression of miR-93 significantly enhances chondrocyte viability and inhibits chondrocyte apoptosis (Ding et al., 2019). Transfection of miR-93 mimics in primary chondrocytes depresses the expression of several LPS-induced inflammatory cytokines, including TNF-α, IL-1β, and IL-6 both in vitro and in vivo (Ding et al., 2019). Another report from Zhang et al. has shown that miR-9 is significantly reduced in articular cartilage tissues from OA patients (Zhang et al., 2018b). In a sodium iodoacetate-induced rat OA model, the authors demonstrate that intraarticular injection of miR-9 agomir significantly downregulates the expression level of matrix proteinase Mmp13 and upregulates the expression level of Col2a1. Mechanistically, miR-9 directly binds to the 3′-UTR of the Mmp13 mRNA (Zhang et al., 2018b). Furthermore, miR-26a has also been reduced in rat OA models (Zhao et al., 2019a). MiR-26a plays a vital role in inhibiting the phosphorylation levels of IκBα and p65, two important factors in the NFκB inflammatory signaling pathway, thus ameliorating synovial hyperplasia and cartilage injury during OA progression (Zhao et al., 2019a). A low expression level of miR-107 has been found in chondrocytes from OA patients compared with normal controls (Zhao et al., 2019b). Transfection of miR-107 mimics into chondrocytes inhibits apoptosis and promotes autophagy by activating AKT/mTOR and NF-κB pathway (Zhao et al., 2019b).
Recently, changes in miRNAs have been linked with epigenetic regulations. In primary cultured human OA chondrocytes, Wu and coworkers have demonstrated that miR-200b-3p targets the expression of DNA methyltransferase 3 alpha (DNMT3A) to regulate the secretion of matrix proteinases and synthesis of type II collagen (Wu et al., 2017). In IL-1β-stimulated chondrocytes, Ma et al. have shown that miR-33b-3p also targets DNMT3A to inhibit IL-1β-induced chondrocyte apoptosis as well as ECM degradation (Ma et al., 2019). Chen and colleagues have reported that miR-455-3p regulates the expression of histone deacetylase 2/8 (HDAC2/8) (Chen et al., 2016). Their results further showed that miR-445-3p downregulates the expression of HDAC2/8 and promotes histone H3 acetylation at the COL2A1 promoter in human chondrogenic cells (Chen et al., 2016). Results from a different research group demonstrated that miR-455-3p targets P21-activated kinases 2(PKA2) to promote the TGF-β signaling pathway and inhibit OA progression (Hu et al., 2019b). MiR-95-5p promotes chondrogenesis and thus inhibits the progression of OA by targeting HDAC2/8 (Mao et al., 2018a). The same research group further reported a role of miR-92a-3p overexpressing exosomes from human mesenchymal stem cells in promoting chondrogenesis and preventing cartilage degradation through regulating histone deacetylase 2 and Wnt5a (Mao et al., 2018b).
Exosomal miRNAs are identified to be pivotal mediators of intercellular communication and thus involved in OA pathogenesis (Fan et al., 2022). Interestingly, Liu and coworkers found a significant increase in circulating exosomal osteoclast-derived miRNAs during the early OA onset (Liu et al., 2021). Targeted deletion of osteoclast-derived miRNAs by genetic knockout of the essential miRNA-processing enzyme Dicer or blocking the release of osteoclast-derived exosomes by siRNA-mediated silencing of Rab27a markedly limited ECM degeneration, osteochondral angiogenesis, and sensory innervation in a surgery-induced mouse OA model (Liu et al., 2021). It has been further demonstrated that osteoclast-derived miRNAs caused OA-like lesions through suppressing the tissue inhibitor of metalloproteinase-2 (TIMP-2) and TIMP-3 (Liu et al., 2021). Furthermore, the authors utilized their previously established osteoclast-targeted delivery system to show that systemic inhibition of osteoclast exosome largely mitigates the surgery-induced OA lesions in mice (Liu et al., 2015; Liu et al., 2021). On the other hand, it has been reported that exosomes derived from mesenchymal stem cells contain certain clusters of miRNAs, such as miR-100-5p and miR-127-3p, which can protect against OA damages and ameliorate gait abnormalities (Tao et al., 2017; Wu et al., 2019; Qiu et al., 2020; Dong et al., 2021; Tao et al., 2021; Xu and Xu, 2021). The above studies have clearly shown that exosomal miRNAs play a central role in the pathogenesis of OA; nonetheless, the complex networks of these exosomal miRNAs still warrant further investigations.
Recent studies have highlighted miRNAs as the potential indicators of therapeutic effects for OA-treating drugs. In an ex-vivo porcine cartilage impact injury model, Genemaras et al. have tested whether interleukin-1 receptor antagonist protein (IRAP) inhibits impact injury-induced inflammation and catabolism in chondrocytes (Genemaras et al., 2015). The data showed that IRAP significantly decreases the expression levels of matrix-degrading enzymes (i.e., Adamts4/5 and Mmp13) as well as inflammatory cytokines (i.e., IL-1β and TNF-α) via downregulation of a series of miRNAs, including miR-140, miR-125b and miR-27b (Genemaras et al., 2015). In 2018, the same research group further reported the effects of anti-inflammatory agents, including IRAP, hyaluronan, dexamethasone, and mesenchymal stem cell treatment on genetic markers of miRNAs, cartilage matrix degradation, apoptosis, and inflammation in an ex-vivo porcine model of acute knee cartilage injury (Genemaras et al., 2018). IRAP significantly increases the expression of miR-140, miR-125b, miR-27b, miR-146a, and miR-22 in injured cartilage. Hyaluronan treatment increases the expression of miR-34a in addition to the above miRNAs. MiR-140 has been linked to chondrogenesis and cartilage formation (Miyaki et al., 2010). Loss of miR-140 in mice causes dysplasia due to impaired chondrocyte proliferation. Results from two groups have demonstrated that estrogen treatment inhibits cartilage degradation and the secretion of Mmp13 from chondrocytes by regulating the expression level of miR-140 (Liang et al., 2016; Xu et al., 2019). The expression level of miR-140 was positively correlated with the therapeutic effects of estrogen in an ovariectomized (OVX) rat model of postmenopausal OA. Estrogen treatment significantly increases the miR-140-5p level compared with the control group, whereas knockdown of miR-140 expression abolishes the inhibitory effect of estrogen on cartilage degradation. Moreover, miR-140 expression can be upregulated by melatonin treatment, leading to enhanced cell proliferation, promoted expression of cartilage ECM proteins (e.g., type II collagen and aggrecan), and inhibited levels of proteinases, including MMP9/13 and ADAMTS4/5, in IL-1β-treated human chondrocytes (Zhang et al., 2019). Taken together, the above findings suggest that miRNAs play central roles in the pathological mechanisms of OA and could serve as diagnostic and prognostic biomarkers and potential therapeutic targets for OA treatment. The functions of miRNAs in cartilage homeostasis and OA pathogenesis are summarized in Table 1.
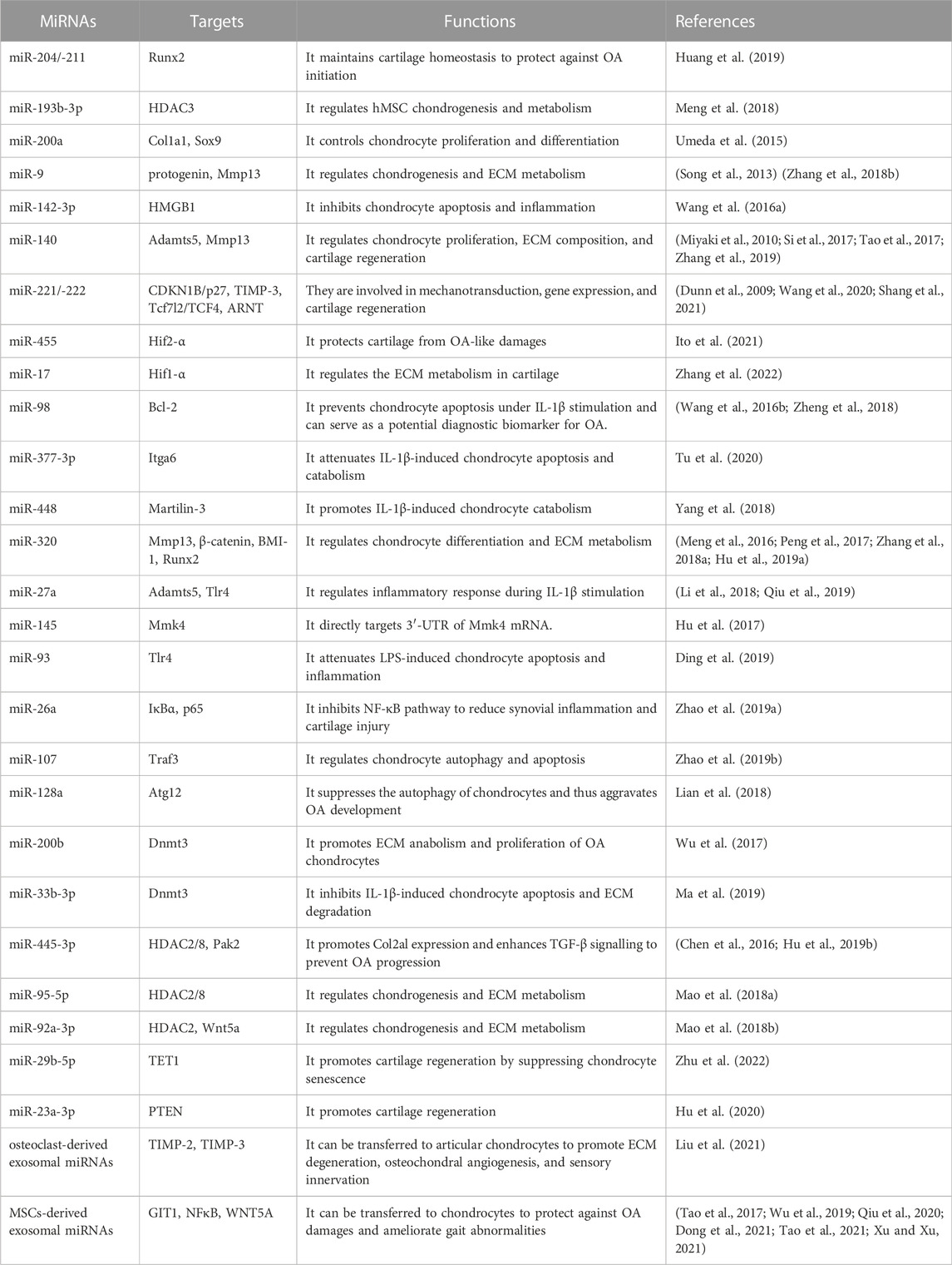
TABLE 1. Roles of miRNAs in cartilage homeostasis and OA pathogenesis.
Under physiological conditions, lncRNA expression is pivotal for regulating gene expression and maintaining cartilage homeostasis. LncRNAs can regulate chondrocyte differentiation by mediating the expression of Sox9, BMP7, and IGF-2 (Zhu et al., 2019). Interestingly, a recent study demonstrated that lncRNA MM2P, stimulated by IL-4 or IL-13, induces expression and exosomal transfer of Sox9 mRNA from monocyte-derived cells to primary chondrocytes to maintain a healthy phenotype of chondrocytes (Bai et al., 2020). The expressions of LncRNA KLF3-AS1 (KLF3 antisense RNA 1) and metastasis-associated lung adenocarcinoma transcript 1 (MALAT-1) are essential for the regulatory effects of hMSCs-derived exosomes on ECM metabolism, chondrocyte differentiation, inflammation, to maintain cartilage homeostasis (Liu et al., 2018; Pan et al., 2021). Some studies have reported that lncRNAs promote chondrocyte proliferation while inhibiting the chondrocyte apoptosis (Jiang et al., 2019; Wang et al., 2021; Xie et al., 2021). For instance, lncRNA small nucleolar RNA host gene 5 (SNHG5) can act as a sponger of miR-10a-5p to suppress chondrocytes apoptosis (Jiang et al., 2021). Loss of SNHG5 expression leads to enhanced IL-1β-induced chondrocyte apoptosis. SNHG15 maintains ECM homeostasis to protect against OA damages (Chen et al., 2020a). Zhang et al. also confirmed the protective role of SNHG15 during OA initiation and progression as SNHG15 can competitively bind with miR-141-3p to upregulate the expression of BCL2L13 (Zhang et al., 2020a). LncRNA maternally expressed 3 (MEG3) promotes chondrocyte proliferation and migration and inhibits apoptosis and inflammation (Huang et al., 2021). Collectively, current evidence has suggested that lncRNA expression is essential for the maintenance of cartilage homeostasis.
Several studies have recently shown that lncRNAs play an essential role in cartilage regeneration. Wang et al. reported that lncRNA colorectal neoplasia differentially expressed gene (CRNDE) regulates the levels of silent information regulator factor 2-related enzyme 1 (SIRT1) and Sox9 to promote the chondrogenic differentiation of bone marrow mesenchymal stem cells (BMSCs) in vitro and enhance cartilage regeneration in a rat model of OA (Shi et al., 2021). LncRNA KLF3-AS1 is markedly enriched in hMSCs-derived exosomes, and the latter is demonstrated to promote cartilage repair via enhancing chondrocyte proliferation in a rat model of OA (Pan et al., 2021). LncRNA H19 can transfer from umbilical cord mesenchymal stem cells (UMSCs) to chondrocytes via exosomes and act as a competing endogenous sponge of miR-29b-3p and miR-29b-3p to regulate FOXO3 expression (Yan et al., 2021). Intra-articular injection of H19-containing exosomes facilitates cartilage repair in an in vivo SD rat cartilage defect model (Yan et al., 2021). Moreover, exosomal H19 derived from UMSCs and fibroblast-like synoviocytes-derived promotes chondrocyte migration and ECM synthesis while suppressing chondrocyte senescence and apoptosis, both in vitro and in vivo (Tan et al., 2020; Yan et al., 2021).
Recent studies have suggested lncRNAs as promising candidates for the diagnosis and therapy of OA as their expression is significantly changed during OA initiation and progression (Okuyan and Begen, 2022). The lncRNA profile under physiological and OA conditions has been analyzed by Hoolwerff and colleagues (van Hoolwerff et al., 2020). LncRNA sequencing was conducted in preserved and lesioned OA articular cartilage tissues from patients taking total knee replacement. The data showed a difference of 191 lncRNAs between preserved and lesioned OA cartilage. The identified lncRNAs were classified into trans-acting lncRNA or cis-acting lncRNA based on their functional location relative to the transcription site. By comparing the distribution of dysregulation of lncRNAs with all transcriptional mRNAs and all sense genes with differentially expressed antisense lncRNAs, the data suggested a cis-regulation mechanism for both intergenic and antisense lncRNAs in OA cartilage. To validate the above cis-regulation mechanism of lncRNAs, the authors chose P3H2-AS1 as an example for proof of concept. A P3H2-AS1 targeting locked nucleic acid (LNA) GapmeRs was transfected into primary chondrocytes, and the expression level of P3H2 was determined. The results showed a significant decrease in P3H2 expression in cells transfected with targeting LNA GapmeRs compared to those transfected with non-targeting LNA GapmeRs. Chen and colleagues Field also analyzed the dysregulation of lncRNAs and related mRNA network (Chen and Chen, 2020). They detected a total of 49 lncRNAs and 1212 mRNAs that were differentially expressed in OA knee articular cartilage as compared with normal controls. In this lncRNA-mRNA network, 7 hub lncRNAs were identified, including MIR210HG, LINC00313, LINC00839, TBC1D3P1-DHX40P1, ISM1-AS1, LINC00654, and HCP5. Further Kyoto encyclopedia of genes and genomes (KEGG) analysis revealed that these OA-related hub lncRNAs were associated with osteoclast differentiation, the FoxO signaling pathway, the TNF signaling pathway, the P53 signaling pathway, and extracellular matrix organization.
LncRNA ADP-ribosylation factor-related protein 1 (ARFRP1) was found to be increased in OA cartilage and in the LPS-treated chondrocytes (Zhang et al., 2020b). Loss of ARFRP1 ameliorated LPS-induced chondrocyte injury via regulating miR-15a-5p/TLR axis. Growth arrest specificity 5 (GAS5) was found to be upregulated in serum and cartilage tissues from knee OA patients (Gao et al., 2020). Overexpression of GAS5 induced chondrocyte apoptosis and inhibited chondrocyte proliferation through downregulation of miR-137. The expression of MALAT1 was reported to be reduced in OA patients (Li et al., 2020). Loss of MALAT1 promotes the production of cyclooxygenase-2 (COX-2), IL-6, and Mmp13 and inhibits the level of type II collagen in the LPS-treated chondrocytes (Li et al., 2020). MALAT1 regulates miR-146a to control PI3K/Akt/mTOR pathway during LPS-induced chondrocyte catabolism, inflammation, and apoptosis (Li et al., 2020). However, another report from Liu et al. showed a controversial role of MALAT1 in the OA pathogenesis (Liu et al., 2019). Their data showed upregulation of MALAT1 in OA cartilages and IL-1β-stimulated chondrocytes. Overexpression of MALAT1 depressed chondrocyte viability and enhanced cartilage catabolism via upregulation of Adamts5 during IL-1β treatment. The nuclear enriched abundant transcript 1 (NEAT1), the prostate cancer-associated transcript 1 (PCAT1), and the SNHG5 can respectively regulate chondrocyte proliferation and apoptosis via controlling several downstream miRNAs (Jiang et al., 2021; Xiao et al., 2021; Zhou et al., 2021). An association between decreased expression of OIP5 antisense RNA 1 (OIP5-ASI) and increased expression of miR-29b-3p was found in IL-1β-treated CHON-001 and ATDC5 chondrocyte-like cells (Zhi et al., 2021). Overexpression of OIP5-ASI increases chondrocyte viability and proliferation and decreases the production of inflammatory cytokines. OIP5-ASI directly binds to miR-29-3p to control the expression of PGRN.
The plasmacytoma variant translocation 1 (PVT1) and the HOXA transcript at the distal tip (HOTTIP) can participate in the ECM degradation during OA progression (Mao et al., 2019; Lu et al., 2020). PVT1 expression is elevated in OA patients and IL-1β-stimulated C28/I2 chondrocytes (Lu et al., 2020). Deletion of PVT1 enhanced cell survival and autophagy and depressed IL-1β-induced apoptosis and inflammation. PVT1 loss upregulated the levels of miR-27b-3p and downregulated downstream target mRNAs of miR-27b-3p. Another study suggests that PVT1 is involved in the hyperglycemia-induced collagen degradation, probably through regulation of the miR-26b-TGF-β1-axis (Ding et al., 2020). LncRNA LINC00671 exacerbates OA lesions by enhancing ONECUT2-mediated Smurf2 expression and ECM degradation (Chen and Xu, 2021).
HOX antisense intergenic RNA (HOTAIR) is found to be elevated in OA chondrocytes (Yang et al., 2020). Loss- and gain-of-function studies demonstrated that HOTAIR directly targets Wnt inhibitory factor 1 (WIF-1) by promoting the H3K27 trimethylation (Yang et al., 2020). Overexpression of HOTAIR increased expression of BMP2, Mmp13, and Adamts5 and decreased expression of Sox9 in SW1353 chondrocyte-like cells, and silencing of HOTAIR exerted opposite effects (Yang et al., 2020). HOTAIR acts as a sponge to regulate the expression of miR-130a-3p and miR-20b to control chondrocyte autophagy and catabolism (Chen et al., 2020b; He and Jiang, 2020; Yang et al., 2020). LncRNA X inactive specific transcript (XIST) contributes to OA progression via miR-149-5p/DNMT3A axis (Liu et al., 2020). Lysyl oxidase-like 1 antisense RNA 1 (LOXL1-AS1) can sponge miR-423-5p and abolish miR-423-5p-dependent inhibition on lysine demethylase 5C (KDM5C) to promote OA progression (Chen et al., 2020c). Taken together, these findings suggest lncRNAs as potential therapeutic targets for OA treatment. The roles of lncRNAs in cartilage homeostasis and OA pathogenesis are summarized in Table 2.
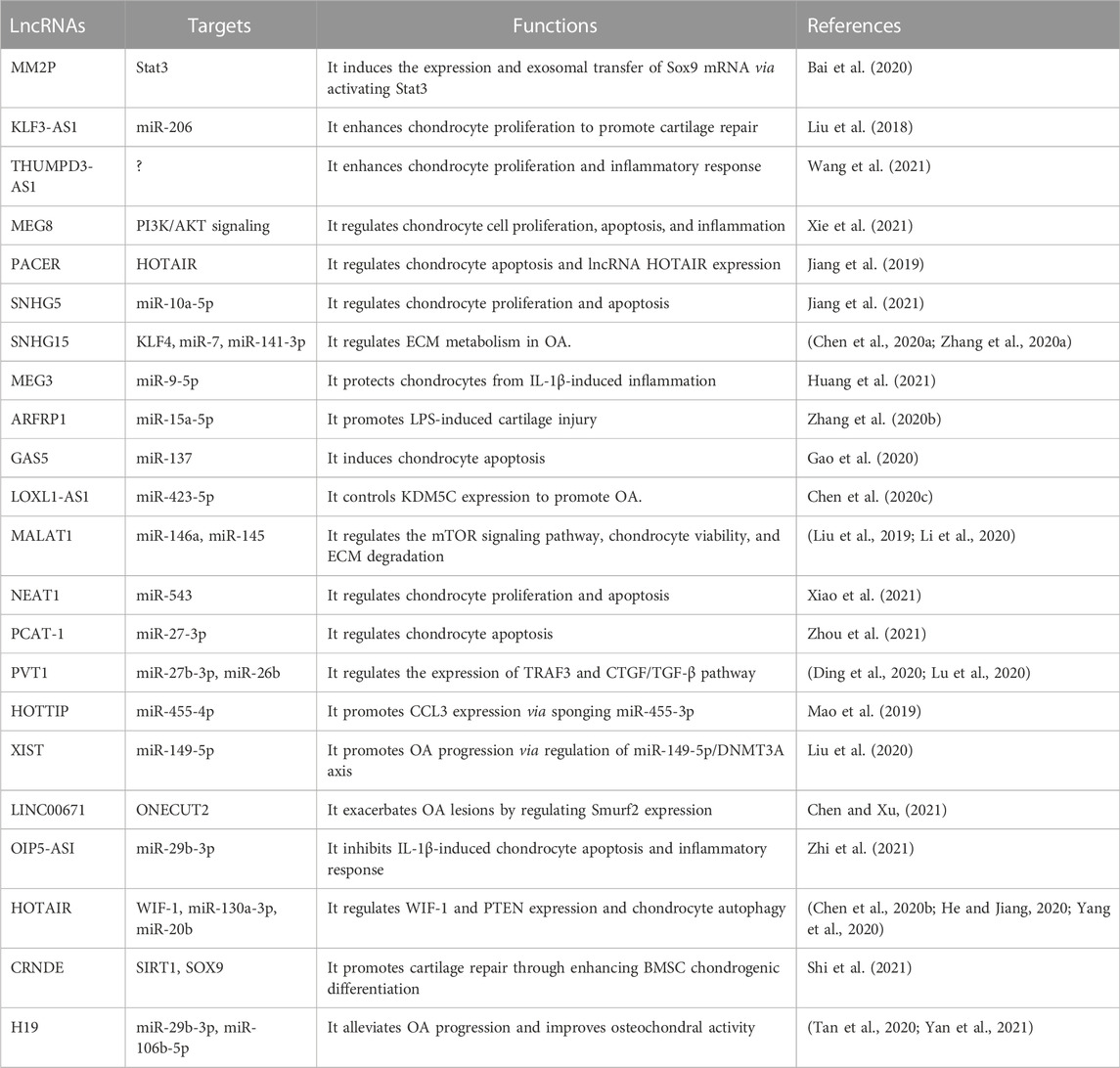
TABLE 2. Role of lncRNAs in cartilage homeostasis and OA pathogenesis.
In this review, we discussed the functional roles of ncRNAs, especially miRNAs and lncRNAs, in cartilage homeostasis and OA pathogenesis (Figure 1). RNA sequencing data reveal a bunch of ncRNAs associated with OA initiation and progression. Nonetheless, the mechanism underlying how ncRNAs regulate cartilage homeostasis and participate in OA pathogenesis remains elusive. Several loss- and gain-of-function studies utilizing transgenic animal models have suggested that ncRNAs exert vital functions in maintaining cartilage homeostasis, and manipulations on ncRNA expression can promote or decelerate the progression of OA through direct or indirect molecular mechanisms. Moreover, ncRNAs can serve as promising diagnostic biomarkers, prognostic indicators, and therapeutic targets for OA. However, we must point out that the quality of current evidence regarding ncRNAs and their functions in OA is relatively low due to inappropriate study design, controversial results, and the lack of direct in vivo evidence. Further high-quality investigations are still needed to confirm and characterize the functional role of ncRNAs in OA pathogenesis in the future.

FIGURE 1. Involvement of miRNAs and lncRNAs in cartilage homeostasis and OA pathogenesis.
JG, WR, and XS wrote the draft of the manuscript. TF, MQ, HZ, DC, and SH contributed to the literature research for the manuscript. JG, WR, SH, and XS revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by Shanghai three year clinical plan on TCM development, the clinical major project of Integrative Medicine (Grant No. SHDC2020CR1051B) and the National Natural Science Foundation of China (No. 82004395).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adams, L. (2017). Non-coding RNA: Pri-miRNA processing: Structure is key. Nat. Rev. Genet. 18, 145. doi:10.1038/nrg.2017.6
Aliperti, V., Skonieczna, J., and Cerase, A. (2021). Long non-coding RNA (lncRNA) roles in cell Biology, neurodevelopment and neurological disorders. Noncoding. RNA 7, 36. doi:10.3390/ncrna7020036
Bai, J., Zhang, Y., Zheng, X., Huang, M., Cheng, W., Shan, H., et al. (2020). LncRNA MM2P-induced, exosome-mediated transfer of Sox9 from monocyte-derived cells modulates primary chondrocytes. Cell Death Dis. 11, 763. doi:10.1038/s41419-020-02945-5
Campbell, J. J., Blain, E. J., Chowdhury, T. T., and Knight, M. M. (2007). Loading alters actin dynamics and up-regulates cofilin gene expression in chondrocytes. Biochem. Biophys. Res. Commun. 361, 329–334. doi:10.1016/j.bbrc.2007.06.185
Chen, C., and Xu, Y. (2021). Long noncoding RNA LINC00671 exacerbates osteoarthritis by promoting ONECUT2-mediated Smurf2 expression and extracellular matrix degradation. Int. Immunopharmacol. 90, 106846. doi:10.1016/j.intimp.2020.106846
Chen, D., Shen, J., Zhao, W., Wang, T., Han, L., Hamilton, J. L., et al. (2017). Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 5, 16044. doi:10.1038/boneres.2016.44
Chen, H., and Chen, L. (2020). An integrated analysis of the competing endogenous RNA network and co-expression network revealed seven hub long non-coding RNAs in osteoarthritis. Bone Jt. Res. 9, 90–98. doi:10.1302/2046-3758.93.BJR-2019-0140.R2
Chen, K., Fang, H., and Xu, N. (2020). LncRNA LOXL1-AS1 is transcriptionally activated by JUND and contributes to osteoarthritis progression via targeting the miR-423-5p/KDM5C axis. Life Sci. 258, 118095. doi:10.1016/j.lfs.2020.118095
Chen, W., Chen, L., Zhang, Z., Meng, F., Huang, G., Sheng, P., et al. (2016). MicroRNA-455-3p modulates cartilage development and degeneration through modification of histone H3 acetylation. Biochim. Biophys. Acta 1863, 2881–2891. doi:10.1016/j.bbamcr.2016.09.010
Chen, Y., Guo, H., Li, L., Bao, D., Gao, F., Li, Q., et al. (2020). Long non-coding RNA (lncRNA) small nucleolar RNA host gene 15 (SNHG15) alleviates osteoarthritis progression by regulation of extracellular matrix homeostasis. Med. Sci. Monit. 26, e923868. doi:10.12659/MSM.923868
Chen, Y., Zhang, L., Li, E., Zhang, G., Hou, Y., Yuan, W., et al. (2020). Long-chain non-coding RNA HOTAIR promotes the progression of osteoarthritis via sponging miR-20b/PTEN axis. Life Sci. 253, 117685. doi:10.1016/j.lfs.2020.117685
Cheng, J-H., Wang, C-J., Su, S-H., Huang, C-Y., and Hsu, S-L. (2016). Next-generation sequencing identifies articular cartilage and subchondral bone miRNAs after ESWT on early osteoarthritis knee. Oncotarget 20, 84398–84407. doi:10.18632/oncotarget.11331
Coutinho de Almeida, R., Ramos, Y. F. M., Mahfouz, A., den Hollander, W., Lakenberg, N., Houtman, E., et al. (2019). RNA sequencing data integration reveals an miRNA interactome of osteoarthritis cartilage. Ann. Rheum. Dis. 78, 270–277. doi:10.1136/annrheumdis-2018-213882
Dahariya, S., Paddibhatla, I., Kumar, S., Raghuwanshi, S., Pallepati, A., and Gutti, R. K. (2019). Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 112, 82–92. doi:10.1016/j.molimm.2019.04.011
Ding, L. B., Li, Y., Liu, G. Y., Li, T. H., Li, F., Guan, J., et al. (2020). Long non-coding RNA PVT1, a molecular sponge of miR-26b, is involved in the progression of hyperglycemia-induced collagen degradation in human chondrocytes by targeting CTGF/TGF-β signal ways. Innate Immun. 26, 204–214. doi:10.1177/1753425919881778
Ding, Y., Wang, L., Zhao, Q., Wu, Z., and Kong, L. (2019). MicroRNA‑93 inhibits chondrocyte apoptosis and inflammation in osteoarthritis by targeting the TLR4/NF‑κB signaling pathway. Int. J. Mol. Med. 43, 779–790. doi:10.3892/ijmm.2018.4033
Dong, J., Li, L., Fang, X., and Zang, M. (2021). Exosome-encapsulated microRNA-127-3p released from bone marrow-derived mesenchymal stem cells alleviates osteoarthritis through regulating CDH11-mediated wnt/β-catenin pathway. J. Pain Res. 14, 297–310. doi:10.2147/JPR.S291472
Dunn, W., DuRaine, G., and Reddi, A. H. (2009). Profiling microRNA expression in bovine articular cartilage and implications for mechanotransduction. Arthritis Rheum. 60, 2333–2339. doi:10.1002/art.24678
Fan, W. J., Liu, D., Pan, L. Y., Wang, W. Y., Ding, Y. L., Zhang, Y. Y., et al. (2022). Exosomes in osteoarthritis: Updated insights on pathogenesis, diagnosis, and treatment. Front. Cell Dev. Biol. 10, 949690. doi:10.3389/fcell.2022.949690
Foo, J. B., Looi, Q. H., How, C. W., Lee, S. H., Al-Masawa, M. E., Chong, P. P., et al. (2021)., 14. Pharmaceuticals (Basel), 1093. doi:10.3390/ph14111093Mesenchymal stem cell-derived exosomes and MicroRNAs in cartilage regeneration: Biogenesis, efficacy, miRNA enrichment and deliveryPharmaceuticals
Fujii, Y., Liu, L., Yagasaki, L., Inotsume, M., Chiba, T., and Asahara, H. (2022). Cartilage homeostasis and osteoarthritis. Int. J. Mol. Sci. 23, 6316. doi:10.3390/ijms23116316
Gao, S. T., Yu, Y. M., Wan, L. P., Liu, Z. M., and Lin, J. X. (2020). LncRNA GAS5 induces chondrocyte apoptosis by down-regulating miR-137. Eur. Rev. Med. Pharmacol. Sci. 24, 10984–10991. doi:10.26355/eurrev_202011_23582
Genemaras, A. A., Ennis, H., Bradshaw, B., Kaplan, L., and Huang, C-Y. C. (2018). Effects of anti-inflammatory agents on expression of early responsive inflammatory and catabolic genes in ex vivo porcine model of acute knee cartilage injury. Cartilage 9, 293–303. doi:10.1177/1947603516684589
Genemaras, A. A., Reiner, T., Huang, C. Y., and Kaplan, L. (2015). Early intervention with Interleukin-1 Receptor Antagonist Protein modulates catabolic microRNA and mRNA expression in cartilage after impact injury. Osteoarthr. Cartil. 23, 2036–2044. doi:10.1016/j.joca.2015.05.010
Ghafouri-Fard, S., Poulet, C., Malaise, M., Abak, A., Mahmud Hussen, B., Taheriazam, A., et al. (2021). Regulatory role of non-coding RNAs on immune responses during sepsis. Front. Immunol. 12, 798713–773171. doi:10.3389/fimmu.2021.798713
Gibson, G., and Asahara, H. (2013). microRNAs and cartilage. J. Orthop. Res. 31, 1333–1344. doi:10.1002/jor.22397
He, B., and Jiang, D. (2020). HOTAIR-induced apoptosis is mediated by sponging miR-130a-3p to repress chondrocyte autophagy in knee osteoarthritis. Cell Biol. Int. 44, 524–535. doi:10.1002/cbin.11253
Hu, G., Zhao, X., Wang, C., Geng, Y., Zhao, J., Xu, J., et al. (2017). MicroRNA-145 attenuates TNF-alpha-driven cartilage matrix degradation in osteoarthritis via direct suppression of MKK4. Cell Death Dis. 8, e3140. doi:10.1038/cddis.2017.522
Hu, H., Dong, L., Bu, Z., Shen, Y., Luo, J., Zhang, H., et al. (2020). miR-23a-3p-abundant small extracellular vesicles released from Gelma/nanoclay hydrogel for cartilage regeneration. J. Extracell. Vesicles 9, 1778883. doi:10.1080/20013078.2020.1778883
Hu, S., Mao, G., Zhang, Z., Wu, P., Wen, X., Liao, W., et al. (2019). MicroRNA-320c inhibits development of osteoarthritis through downregulation of canonical Wnt signaling pathway. Life Sci. 228, 242–250. doi:10.1016/j.lfs.2019.05.011
Hu, S., Zhao, X., Mao, G., Zhang, Z., Wen, X., Zhang, C., et al. (2019). MicroRNA-455-3p promotes TGF-beta signaling and inhibits osteoarthritis development by directly targeting PAK2. Exp. Mol. Med. 51, 1–13. doi:10.1038/s12276-019-0322-3
Huang, J., Zhao, L., Fan, Y., Liao, L., Ma, P. X., Xiao, G., et al. (2019). The microRNAs miR-204 and miR-211 maintain joint homeostasis and protect against osteoarthritis progression. Nat. Commun. 10, 2876. doi:10.1038/s41467-019-10753-5
Huang, Y., Chen, D., Yan, Z., Zhan, J., Xue, X., Pan, X., et al. (2021). LncRNA MEG3 protects chondrocytes from IL-1β-induced inflammation via regulating miR-9-5p/KLF4 Axis. Front. Physiol. 12–617654. doi:10.3389/fphys.2021.617654
Ito, Y., Matsuzaki, T., Ayabe, F., Mokuda, S., Kurimoto, R., Matsushima, T., et al. (2021). Both microRNA-455-5p and -3p repress hypoxia-inducible factor-2α expression and coordinately regulate cartilage homeostasis. Nat. Commun. 12, 4148. doi:10.1038/s41467-021-24460-7
Jiang, H., Pang, H., Wu, P., Cao, Z., Li, Z., and Yang, X. (2021). LncRNA SNHG5 promotes chondrocyte proliferation and inhibits apoptosis in osteoarthritis by regulating miR-10a-5p/H3F3B axis. Connect. Tissue Res. 62, 605–614. doi:10.1080/03008207.2020.1825701
Jiang, M., Liu, J., Luo, T., Chen, Q., Lu, M., and Meng, D. (2019). LncRNA PACER is down-regulated in osteoarthritis and regulates chondrocyte apoptosis and lncRNA HOTAIR expression. Biosci. Rep. 39, BSR20190404. doi:10.1042/BSR20190404
Joyce, B. T., Zheng, Y., Zhang, Z., Liu, L., Kocherginsky, M., Murphy, R., et al. (2018). miRNA-processing gene methylation and cancer risk. Cancer Epidemiol. Biomarkers Prev. 27, 550–557. doi:10.1158/1055-9965.EPI-17-0849
Kung, J. T., Colognori, D., and Lee, J. T. (2013). Long noncoding RNAs: Past, present, and future. Genetics 193, 651–669. doi:10.1534/genetics.112.146704
Kung, L. H. W., Ravi, V., Rowley, L., Angelucci, C., Fosang, A. J., Bell, K. M., et al. (2018). Cartilage MicroRNA dysregulation during the onset and progression of mouse osteoarthritis is independent of aggrecanolysis and overlaps with candidates from end-stage human disease. Arthritis Rheumatol. 70, 383–395. doi:10.1002/art.40378
Li, H., Xie, S., Li, H., Zhang, R., and Zhang, H. (2020). LncRNA MALAT1 mediates proliferation of LPS treated-articular chondrocytes by targeting the miR-146a-PI3K/Akt/mTOR axis. Life Sci. 254, 116801. doi:10.1016/j.lfs.2019.116801
Li, X., He, P., Li, Z., Wang, H., Liu, M., Xiao, Y., et al. (2018). Interleukin‑1β‑mediated suppression of microRNA‑27a‑3p activity in human cartilage via MAPK and NF‑κB pathways: A potential mechanism of osteoarthritis pathogenesis. Mol. Med. Rep. 18, 541–549. doi:10.3892/mmr.2018.8970
Lian, W. S., Ko, J. Y., Wu, R. W., Sun, Y. C., Chen, Y. S., Wu, S. L., et al. (2018). MicroRNA-128a represses chondrocyte autophagy and exacerbates knee osteoarthritis by disrupting Atg12. Cell Death Dis. 9, 919. doi:10.1038/s41419-018-0994-y
Liang, Y., Duan, L., Xiong, J., Zhu, W., Liu, Q., Wang, D., et al. (2016). E2 regulates MMP-13 via targeting miR-140 in IL-1β-induced extracellular matrix degradation in human chondrocytes. Arthritis Res. Ther. 18, 105. doi:10.1186/s13075-016-0997-y
Liu, C., Ren, S., Zhao, S., and Wang, Y. (2019). LncRNA MALAT1/MiR-145 adjusts IL-1β-induced chondrocytes viability and cartilage matrix degradation by regulating ADAMTS5 in human osteoarthritis. Yonsei Med. J. 60, 1081–1092. doi:10.3349/ymj.2019.60.11.1081
Liu, J., Dang, L., Li, D., Liang, C., He, X., Wu, H., et al. (2015). A delivery system specifically approaching bone resorption surfaces to facilitate therapeutic modulation of microRNAs in osteoclasts. Biomaterials 52, 148–160. doi:10.1016/j.biomaterials.2015.02.007
Liu, J., Wu, X., Lu, J., Huang, G., Dang, L., Zhang, H., et al. (2021). Exosomal transfer of osteoclast-derived miRNAs to chondrocytes contributes to osteoarthritis progression. Nat. Aging 1, 368–384. doi:10.1038/s43587-021-00050-6
Liu, Y., Lin, L., Zou, R., Wen, C., Wang, Z., and Lin, F. (2018). MSC-derived exosomes promote proliferation and inhibit apoptosis of chondrocytes via lncRNA-KLF3-AS1/miR-206/GIT1 axis in osteoarthritis. Cell Cycle 17, 2411–2422. doi:10.1080/15384101.2018.1526603
Liu, Y., Liu, K., Tang, C., Shi, Z., Jing, K., and Zheng, J. (2020). Long non-coding RNA XIST contributes to osteoarthritis progression via miR-149-5p/DNMT3A axis. Biomed. Pharmacother. 128, 110349. doi:10.1016/j.biopha.2020.110349
Lu, X., Yu, Y., Yin, F., Yang, C., Li, B., Lin, J., et al. (2020). Knockdown of PVT1 inhibits IL-1β-induced injury in chondrocytes by regulating miR-27b-3p/TRAF3 axis. Int. Immunopharmacol. 79, 106052. doi:10.1016/j.intimp.2019.106052
Ma, F., Li, G., Yu, Y., Xu, J., and Wu, X. (2019). MiR-33b-3p promotes chondrocyte proliferation and inhibits chondrocyte apoptosis and cartilage ECM degradation by targeting DNMT3A in osteoarthritis. Biochem. Biophys. Res. Commun. 519, 430–437. doi:10.1016/j.bbrc.2019.09.022
Mao, G., Hu, S., Zhang, Z., Wu, P., Zhao, X., Lin, R., et al. (2018). Exosomal miR-95-5p regulates chondrogenesis and cartilage degradation via histone deacetylase 2/8. J. Cell. Mol. Med. 22, 5354–5366. doi:10.1111/jcmm.13808
Mao, G., Kang, Y., Lin, R., Hu, S., Zhang, Z., Li, H., et al. (2019). Long non-coding RNA HOTTIP promotes CCL3 expression and induces cartilage degradation by sponging miR-455-3p. Front. Cell Dev. Biol. 7, 161. doi:10.3389/fcell.2019.00161
Mao, G., Zhang, Z., Hu, S., Zhang, Z., Chang, Z., Huang, Z., et al. (2018). Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res. Ther. 9, 247. doi:10.1186/s13287-018-1004-0
Martin, A. R., Patel, J. M., Zlotnick, H. M., Carey, J. L., and Mauck, R. L. (2019). Emerging therapies for cartilage regeneration in currently excluded 'red knee' populations. NPJ Regen. Med. 4, 12. doi:10.1038/s41536-019-0074-7
Mattick, J. S., and Makunin, I. V. (2006). Non-coding RNA. Hum. Mol. Genet. 1, R17–R29. doi:10.1093/hmg/ddl046
Meng, F., Li, Z., Zhang, Z., Yang, Z., Kang, Y., Zhao, X., et al. (2018). MicroRNA-193b-3p regulates chondrogenesis and chondrocyte metabolism by targeting HDAC3. Theranostics 8, 2862–2883. doi:10.7150/thno.23547
Meng, F., Zhang, Z., Chen, W., Huang, G., He, A., Hou, C., et al. (2016). MicroRNA-320 regulates matrix metalloproteinase-13 expression in chondrogenesis and interleukin-1β-induced chondrocyte responses. Osteoarthr. Cartil. 24, 932–941. doi:10.1016/j.joca.2015.12.012
Miyaki, S., Sato, T., Inoue, A., Otsuki, S., Ito, Y., Yokoyama, S., et al. (2010). MicroRNA-140 plays dual roles in both cartilage development and homeostasis. Genes Dev. 24, 1173–1185. doi:10.1101/gad.1915510
Murphy, M. P., Koepke, L. S., Lopez, M. T., Tong, X., Ambrosi, T. H., Gulati, G. S., et al. (2020). Articular cartilage regeneration by activated skeletal stem cells. Nat. Med. 26, 1583–1592. doi:10.1038/s41591-020-1013-2
Ntoumou, E., Tzetis, M., Braoudaki, M., Lambrou, G., Poulou, M., Malizos, K., et al. (2017). Serum microRNA array analysis identifies miR-140-3p, miR-33b-3p and miR-671-3p as potential osteoarthritis biomarkers involved in metabolic processes. Clin. Epigenetics 9, 127. doi:10.1186/s13148-017-0428-1
O'Brien, J., Hayder, H., Zayed, Y., and Peng, C. (2018). Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 9, 402. doi:10.3389/fendo.2018.00402
Okuyan, H. M., and Begen, M. A. (2022). LncRNAs in osteoarthritis. Clin. Chim. Acta. 532, 145–163. doi:10.1016/j.cca.2022.05.030
Palazzo, C., Nguyen, C., Lefevre-Colau, M. M., Rannou, F., and Poiraudeau, S. (2016). Risk factors and burden of osteoarthritis. Ann. Phys. Rehabil. Med. 59, 134–138. doi:10.1016/j.rehab.2016.01.006
Pan, C., Huang, W., Chen, Q., Xu, J., Yao, G., Li, B., et al. (2021). LncRNA malat-1 from MSCs-derived extracellular vesicles suppresses inflammation and cartilage degradation in osteoarthritis. Front. Bioeng. Biotechnol. 9, 772002. doi:10.3389/fbioe.2021.772002
Pekacova, A., Baloun, J., Svec, X., and Senolt, L. (2022). Non-coding RNAs in diseases with a focus on osteoarthritis. Wiley Interdiscip Rev RNA. 1, e1756.
Peng, H., Liang, D., Li, B., Liang, C., Huang, W., and Lin, H. (2017). MicroRNA-320a protects against osteoarthritis cartilage degeneration by regulating the expressions of BMI-1 and RUNX2 in chondrocytes. Pharmazie 72, 223–226. doi:10.1691/ph.2017.6890
Qiu, B., Xu, X., Yi, P., and Hao, Y. (2020). Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J. Cell. Mol. Med. 24, 10855–10865. doi:10.1111/jcmm.15714
Qiu, W. J., Xu, M. Z., Zhu, X. D., and Ji, Y. H. (2019). MicroRNA-27a alleviates IL-1β-induced inflammatory response and articular cartilage degradation via TLR4/NF-κB signaling pathway in articular chondrocytes. Int. Immunopharmacol. 76, 105839. doi:10.1016/j.intimp.2019.105839
Razmara, E., Bitaraf, A., Yousefi, H., Nguyen, T. H., Garshasbi, M., Cho, W. C., et al. (2019). Non-coding RNAs in cartilage development: An updated review. Int. J. Mol. Sci. 20, 4475. doi:10.3390/ijms20184475
Robinson, W. H., Lepus, C. M., Wang, Q., Raghu, H., Mao, R., Lindstrom, T. M., et al. (2016). Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 12, 580–592. doi:10.1038/nrrheum.2016.136
Saquib, M., Agnihotri, P., Monu, , and Biswas, S. (2021). Exogenous miRNA: A perspective role as therapeutic in rheumatoid arthritis. Curr. Rheumatol. Rep. 23, 43. doi:10.1007/s11926-021-01009-7
Saxena, A., and Carninci, P. (2011). Long non-coding RNA modifies chromatin: Epigenetic silencing by long non-coding RNAs. Bioessays 33, 830–839. doi:10.1002/bies.201100084
Shang, X., Böker, K. O., Taheri, S., Lehmann, W., and Schilling, A. F. (2021). Extracellular vesicles allow epigenetic mechanotransduction between chondrocytes and osteoblasts. Int. J. Mol. Sci. 22, 13282. doi:10.3390/ijms222413282
Shi, C., Zheng, W., and Wang, J. (2021). lncRNA-CRNDE regulates BMSC chondrogenic differentiation and promotes cartilage repair in osteoarthritis through SIRT1/SOX9. Mol. Cell. Biochem. 476, 1881–1890. doi:10.1007/s11010-020-04047-4
Si, H. B., Zeng, Y., Liu, S. Y., Zhou, Z. K., Chen, Y. N., Cheng, J. Q., et al. (2017). Intra-articular injection of microRNA-140 (miRNA-140) alleviates osteoarthritis (OA) progression by modulating extracellular matrix (ECM) homeostasis in rats. Osteoarthr. Cartil. 25, 1698–1707. doi:10.1016/j.joca.2017.06.002
Snoeker, B., Turkiewicz, A., Magnusson, K., Frobell, R., Yu, D., Peat, G., et al. (2020). Risk of knee osteoarthritis after different types of knee injuries in young adults: A population-based cohort study. Br. J. Sports Med. 54, 725–730. doi:10.1136/bjsports-2019-100959
Song, J., Kim, D., Chun, C-H., and Jin, E-J. (2013). MicroRNA-9 regulates survival of chondroblasts and cartilage integrity by targeting protogenin. Cell Commun. Signal. 5, 66. doi:10.1186/1478-811X-11-66
Tan, F., Wang, D., and Yuan, Z. (2020). The fibroblast-like synoviocyte derived exosomal long non-coding RNA H19 alleviates osteoarthritis progression through the miR-106b-5p/TIMP2 Axis. Inflammation 43, 1498–1509. doi:10.1007/s10753-020-01227-8
Tao, S. C., Yuan, T., Zhang, Y. L., Yin, W. J., Guo, S. C., and Zhang, C. Q. (2017). Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics 7, 180–195. doi:10.7150/thno.17133
Tao, Y., Zhou, J., Wang, Z., Tao, H., Bai, J., Ge, G., et al. (2021). Human bone mesenchymal stem cells-derived exosomal miRNA-361-5p alleviates osteoarthritis by downregulating DDX20 and inactivating the NF-κB signaling pathway. Bioorg. Chem. 113, 104978. doi:10.1016/j.bioorg.2021.104978
Tong, L., Yu, H., Huang, X., Shen, J., Xiao, G., Chen, L., et al. (2022). Current understanding of osteoarthritis pathogenesis and relevant new approaches. Bone Res. 10, 60. doi:10.1038/s41413-022-00226-9
Tu, Y., Ma, T., Wen, T., Yang, T., Xue, L., Cai, M., et al. (2020). MicroRNA-377-3p alleviates IL-1β-caused chondrocyte apoptosis and cartilage degradation in osteoarthritis in part by downregulating ITGA6. Biochem. Biophys. Res. Commun. 523, 46–53. doi:10.1016/j.bbrc.2019.11.186
Umeda, M., Terao, F., Miyazaki, K., Yoshizaki, K., and Takahashi, I. (2015). MicroRNA-200a regulates the development of mandibular condylar cartilage. J. Dent. Res. 94, 795–802. doi:10.1177/0022034515577411
van Hoolwerff, M., Metselaar, P. I., Tuerlings, M., Suchiman, H. E. D., Lakenberg, N., Ramos, Y. F. M., et al. (2020). Elucidating epigenetic regulation by identifying functional cis-acting long noncoding RNAs and their targets in osteoarthritic articular cartilage. Arthritis Rheumatol. 72, 1845–1854. doi:10.1002/art.41396
Wang, J., Chen, L., Jin, S., Lin, J., Zheng, H., Zhang, H., et al. (2016). MiR-98 promotes chondrocyte apoptosis by decreasing Bcl-2 expression in a rat model of osteoarthritis. Acta Biochim. Biophys. Sin. 48, 923–929. doi:10.1093/abbs/gmw084
Wang, R., Jiang, W., Zhang, L., Xie, S., Zhang, S., Yuan, S., et al. (2020). Intra-articular delivery of extracellular vesicles secreted by chondrogenic progenitor cells from MRL/MpJ superhealer mice enhances articular cartilage repair in a mouse injury model. Stem Cell Res. Ther. 11, 93. doi:10.1186/s13287-020-01594-x
Wang, X., Guo, Y., Wang, C., Yu, H., Yu, X., and Yu, H. (2016). MicroRNA-142-3p inhibits chondrocyte apoptosis and inflammation in osteoarthritis by targeting HMGB1. Inflammation 39, 1718–1728. doi:10.1007/s10753-016-0406-3
Wang, Y., Li, T., Yang, Q., Feng, B., Xiang, Y., Lv, Z., et al. (2021). LncRNA THUMPD3-AS1 enhances the proliferation and inflammatory response of chondrocytes in osteoarthritis. Int. Immunopharmacol. 100, 108138. doi:10.1016/j.intimp.2021.108138
Wei, J-W., Huang, K., Yang, C., and Kang, C-S. (2017). Non-coding RNAs as regulators in epigenetics (Review). Oncol. Rep. 37, 3–9. doi:10.3892/or.2016.5236
Weilner, S., Grillari-Voglauer, R., Redl, H., Grillari, J., and Nau, T. (2015). The role of microRNAs in cellular senescence and age-related conditions of cartilage and bone. Acta Orthop. 86, 92–99. doi:10.3109/17453674.2014.957079
Wojdasiewicz, P., Poniatowski, L. A., and Szukiewicz, D. (2014). The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 561459. doi:10.1155/2014/561459
Wu, J., Kuang, L., Chen, C., Yang, J., Zeng, W. N., Li, T., et al. (2019). miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials 206, 87–100. doi:10.1016/j.biomaterials.2019.03.022
Wu, J., Tao, Y., Shang, A., Wang, W., Zhang, Y., Hu, L., et al. (2017). Effect of the interaction between MiR-200b-3p and DNMT3A on cartilage cells of osteoarthritis patients. J. Cell. Mol. Med. 21, 2308–2316. doi:10.1111/jcmm.13152
Xiang, X. N., Zhu, S. Y., He, H. C., Yu, X., Xu, Y., and He, C. Q. (2022). Mesenchymal stromal cell-based therapy for cartilage regeneration in knee osteoarthritis. Stem Cell Res. Ther. 13, 14. doi:10.1186/s13287-021-02689-9
Xiao, P., Zhu, X., Sun, J., Zhang, Y., Qiu, W., Li, J., et al. (2021). LncRNA NEAT1 regulates chondrocyte proliferation and apoptosis via targeting miR-543/PLA2G4A axis. Hum. Cell 34, 60–75. doi:10.1007/s13577-020-00433-8
Xie, F., Liu, Y. L., Chen, X. Y., Li, Q., Zhong, J., Dai, B. Y., et al. (2020). Role of MicroRNA, LncRNA, and exosomes in the progression of osteoarthritis: A review of recent literature. Orthop. Surg. 12, 708–716. doi:10.1111/os.12690
Xie, W., Jiang, L., Huang, X., Shang, H., Gao, M., You, W., et al. (2021). lncRNA MEG8 is downregulated in osteoarthritis and regulates chondrocyte cell proliferation, apoptosis and inflammation. Exp. Ther. Med. 22, 1153. doi:10.3892/etm.2021.10587
Xu, H., and Xu, B. (2021). BMSC-derived exosomes ameliorate osteoarthritis by inhibiting pyroptosis of cartilage via delivering miR-326 targeting HDAC3 and STAT1//NF-κB p65 to chondrocytes. Mediat. Inflamm. 2021, 9972805. doi:10.1155/2021/9972805
Xu, X., Li, X., Liang, Y., Ou, Y., Huang, J., Xiong, J., et al. (2019). Estrogen modulates cartilage and subchondral bone remodeling in an ovariectomized rat model of postmenopausal osteoarthritis. Med. Sci. Monit. 25, 3146–3153. doi:10.12659/MSM.916254
Yan, L., Liu, G., and Wu, X. (2021). The umbilical cord mesenchymal stem cell-derived exosomal lncRNA H19 improves osteochondral activity through miR-29b-3p/FoxO3 axis. Clin. Transl. Med. 11, e255. doi:10.1002/ctm2.255
Yang, H., Wu, D., Li, H., Chen, N., and Shang, Y. (2018). Downregulation of microRNA-448 inhibits IL-1β-induced cartilage degradation in human chondrocytes via upregulation of matrilin-3. Cell. Mol. Biol. Lett. 23, 7. doi:10.1186/s11658-018-0072-6
Yang, Y., Xing, D., Wang, Y., Jia, H., Li, B., and Li, J. J. (2020). A long non-coding RNA, HOTAIR, promotes cartilage degradation in osteoarthritis by inhibiting WIF-1 expression and activating Wnt pathway. BMC Mol. Cell Biol. 21, 53. doi:10.1186/s12860-020-00299-6
Zhang, G., Zhang, Q., Zhu, J., Tang, J., and Nie, M. (2020). LncRNA ARFRP1 knockdown inhibits LPS-induced the injury of chondrocytes by regulation of NF-κB pathway through modulating miR-15a-5p/TLR4 axis. Life Sci. 261, 118429. doi:10.1016/j.lfs.2020.118429
Zhang, H., Song, B., and Pan, Z. (2018). Downregulation of microRNA-9 increases matrix metalloproteinase-13 expression levels and facilitates osteoarthritis onset. Mol. Med. Rep. 17, 3708–3714. doi:10.3892/mmr.2017.8340
Zhang, H-X., Sun, C., Yu, . H-C., Song, B., and Pan, Z-X. (2018). Targeted inhibition of β-catenin by miR-320 and decreased MMP-13 expression in suppressing chondrocyte collagen degradation. Eur. Rev. Med. Pharmacol. Sci. 22, 5828–5835. doi:10.26355/eurrev_201809_15909
Zhang, X., Huang, C. R., Pan, S., Pang, Y., Chen, Y. S., Zha, G. C., et al. (2020). Long non-coding RNA SNHG15 is a competing endogenous RNA of miR-141-3p that prevents osteoarthritis progression by upregulating BCL2L13 expression. Int. Immunopharmacol. 83, 106425. doi:10.1016/j.intimp.2020.106425
Zhang, Y., Li, S., Jin, P., Shang, T., Sun, R., Lu, L., et al. (2022). Dual functions of microRNA-17 in maintaining cartilage homeostasis and protection against osteoarthritis. Nat. Commun. 13, 2447. doi:10.1038/s41467-022-30119-8
Zhang, Y., Lin, J., Zhou, X., Chen, X., Chen, A. C., Pi, B., et al. (2019). Melatonin prevents osteoarthritis-induced cartilage degradation via targeting MicroRNA-140. Oxid. Med. Cell. Longev. 2019, 9705929. doi:10.1155/2019/9705929
Zhao, X., Li, H., and Wang, L. (2019). MicroRNA-107 regulates autophagy and apoptosis of osteoarthritis chondrocytes by targeting TRAF3. Int. Immunopharmacol. 71, 181–187. doi:10.1016/j.intimp.2019.03.005
Zhao, Z., Dai, X. S., Wang, Z. Y., Bao, Z. Q., and Guan, J. Z. (2019). MicroRNA-26a reduces synovial inflammation and cartilage injury in osteoarthritis of knee joints through impairing the NF-κB signaling pathway. Biosci. Rep. 39, BSR20182025. doi:10.1042/BSR20182025
Zheng, W-D., Zhou, F-L., Lin, N. J. L., and Liu, J. (2018). Investigation for the role of CTX-III and microRNA-98 in diagnosis and treatment of osteoarthritis. Eur. Rev. Med. Pharmacol. Sci. 22, 5424–5428. doi:10.26355/eurrev_201809_15801
Zhi, L., Zhao, J., Zhao, H., Qing, Z., Liu, H., and Ma, J. (2021). Downregulation of LncRNA OIP5-AS1 induced by IL-1β aggravates osteoarthritis via regulating miR-29b-3p/PGRN. Cartilage 13, 1345s–1355s. doi:10.1177/1947603519900801
Zhou, L., Gu, M., Ma, X., Wen, L., Zhang, B., Lin, Y., et al. (2021). Long non-coding RNA PCAT-1 regulates apoptosis of chondrocytes in osteoarthritis by sponging miR-27b-3p. J. Bone Min. Metab. 39, 139–147. doi:10.1007/s00774-020-01128-8
Zhu, J., Yang, S., Qi, Y., Gong, Z., Zhang, H., Liang, K., et al. (2022). Stem cell-homing hydrogel-based miR-29b-5p delivery promotes cartilage regeneration by suppressing senescence in an osteoarthritis rat model. Sci. Adv. 8–eabk0011. doi:10.1126/sciadv.abk0011
Keywords: osteoarthritis, cartilage homeostasis, microRNA, lncRNA, regeneration
Citation: Gu J, Rao W, Huo S, Fan T, Qiu M, Zhu H, Chen D and Sheng X (2022) MicroRNAs and long non-coding RNAs in cartilage homeostasis and osteoarthritis. Front. Cell Dev. Biol. 10:1092776. doi: 10.3389/fcell.2022.1092776
Received: 08 November 2022; Accepted: 29 November 2022;
Published: 13 December 2022.
Edited by:
Jin Liu, Hong Kong Baptist University, Hong Kong SAR, ChinaReviewed by:
Maolin Wang, First Affiliated Hospital of Shantou University Medical College, ChinaCopyright © 2022 Gu, Rao, Huo, Fan, Qiu, Zhu, Chen and Sheng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoping Sheng, c2hlbmd4aWFvcEAxMjYuY29t; Shaochuan Huo, aHVvc2hhb2NodWFuc3pAMTYzLmNvbSYjeDAyMDBhOw==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.