 Chunyue Hao1
Chunyue Hao1 Bin Zhan
Bin Zhan Zixia Wang
Zixia Wang Xinping Zhu
Xinping Zhu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell Dev. Biol., 15 July 2021
Sec. Molecular and Cellular Pathology
Volume 9 - 2021 | https://doi.org/10.3389/fcell.2021.695015
This article is part of the Research TopicCellular and Molecular Basis in Parasitic Diseases Control: Research TrendsView all 42 articles
Helminth infection modulates host regulatory immune responses to maintain immune homeostasis. Our previous study identified Trichinella spiralis paramyosin (TsPmy) as a major immunomodulatory protein with the ability to induce regulatory T cells (Tregs). However, whether TsPmy regulates gut Tregs and contributes to intestinal immune homeostasis remains unclear. Here we investigated the therapeutic effect of recombinant TsPmy protein (rTsPmy) on experimental colitis in mice, and elucidated the roles and mechanisms of colonic Tregs induced by rTsPmy in ameliorating colitis. Acute colitis was induced by dextran sodium sulfate (DSS) in C57BL/6J mice, and chronic colitis was induced by naïve T cells in Rag1 KO mice. Mice with colitis were pre-treated with rTsPmy intraperitoneally, and clinical manifestations and colonic inflammation were evaluated. Colonic lamina propria (cLP) Tregs phenotypes and functions in DSS-induced colitis were analyzed by flow cytometry. Adoptive transfer of cLP Tregs treated by rTsPmy into Rag1 KO chronic colitis was utilized to verify Tregs suppressive function. rTsPmy ameliorated the disease progress of DSS-induced colitis, reduced pro-inflammatory responses but enhanced regulatory cytokines production in DSS-induced colitis. Moreover, rTsPmy specifically stimulated the expansion of thymic-derived Tregs (tTregs) rather than the peripherally derived Tregs (pTregs) in the inflamed colon, enhanced the differentiation of effector Tregs (eTregs) with higher suppressive function and stability in colitis. This study describes the mechanisms of colonic Tregs induced by the Trichinella-derived protein rTsPmy in maintaining gut immune homeostasis during inflammation. These findings provide further insight into the immunological mechanisms involved in the therapeutic effect of helminth-derived proteins in inflammatory bowel diseases.
Inflammatory bowel disease (IBD) is widely described as an autoimmune disease characterized by chronic recurrent inflammation in the gastrointestinal tract as a result of excessive immune responses to gut lumen antigens in genetically susceptible individuals. The two most common IBDs are Crohn’s disease (CD) and ulcerative colitis (UC) (Xavier and Podolsky, 2007; Maloy and Powrie, 2011). Abdominal pain, diarrhea, and weight loss are the most common symptoms in patients with IBD. The incidence and prevalence of IBD are rising worldwide, particularly in newly industrialized regions (Kaplan, 2015; Zuo et al., 2018). Despite an increasing repertoire of therapeutic targets focusing on lymphocyte trafficking and activation, gut barrier function and matrix remodeling, and even manipulation of gut microbiota (Neurath, 2017), a large number of patients still have chronic intestinal inflammation.
Foxp3+ regulatory T cells (Tregs) represent a unique CD4+ T-cell subset essential for maintaining immune balance and homeostasis (Ohkura et al., 2013). The gut, especially the colon, has highly abundant Tregs in the lamina propria (LP), accounting for about 20–30% of total CD4+ T cells, and is responsible for adapting to resident antigens in the intestine such as commensal bacteria and food antigens (Tanoue et al., 2016). Tregs restrict inflammatory responses and maintain immune tolerance via various molecular mechanisms at a multitude of cellular levels, including (i) inhibition of T-cell proliferation and effector functions through the anti-inflammatory cytokines IL-10, TGF-β, and IL-35, (ii) apoptosis of effector T cells (Teffs) induced by starvation of IL-2, (iii) metabolic disruption of Teffs by converting ATP into adenosine, (iv) direct cytolysis against Teffs by galectin-1, granzyme B and perforin, and (v) contact-dependent neutralization of dendritic cell (DC) function by the interaction of Treg-derived CTLA4/TIGIT and CD80/CD155 on DCs (Shevach, 2009, 2018).
There is a shred of definitive evidence that patients with IBD have defects in quantity and quality of Tregs (Abdel-Motal et al., 2019), coupled with the fact that the colon is a primary site of inflammation; therefore, enhancing colonic Treg activity has become a compelling strategy for the immunotherapy of IBD. Several studies demonstrated that it was feasible to expand polyclonal Tregs from patients in vitro under good manufacturing practice (GMP) condition as a source of therapy, in which these Tregs sustained their stability and suppression capabilities (Canavan et al., 2016; Mathew et al., 2018). Antigen-specific Tregs are believed to be more effective than polyclonal Tregs (Clough et al., 2020); however, no antigen has yet been identified as a definite cause in IBD. Nonetheless, a French study reported a clinical trial with ovalbumin (OVA)-specific Tregs showing dose-related therapeutic efficacy in refractory CD (Desreumaux et al., 2012). More clinical trials using autologous Tregs expanded in vitro are ongoing for immunotherapy of CD (ClinicalTrails.gov Identifier: NCT03185000).
The increase in the prevalence of IBD has been attributed to improved hygiene and the disappearance of intestinal helminth and other infections. Based on the “hygiene hypothesis,” a lack of early childhood exposure to multiple antigens increases susceptibility to allergies and autoimmune diseases including IBD. It is possible that intestinal immune homeostasis requires the presence of helminth infections (Gause and Maizels, 2016; Bach, 2018). The differentiation, maintenance, and functional maturation of gut Tregs are orchestrated via various specific signals from the lumen environment (Tanoue et al., 2016). The activation of Tregs has emerged as a central explanation for the beneficial role of helminth infections in moderating inflammatory diseases such as allergies and autoimmune disorders including IBD (Weinstock, 2015; Gazzinelli-Guimaraes and Nutman, 2018). There is growing evidence demonstrating that helminths resident in the gut continuously release immunoregulatory small molecules to modulate intestinal Tregs to benefit immune homeostasis locally or systemically (Reynolds et al., 2015; Gause and Maizels, 2016; Lopes et al., 2016). Natural infection with intestinal helminths reduced pathology in patients with UC (Buning et al., 2008; Broadhurst et al., 2010). Clinical trials with eggs of Trichuris suum and hookworm Necator americanus decreased symptoms in patients with chronic CD or UC (Cheifetz et al., 2017). Although the benefits of helminth infection in colitis have been extensively investigated, the mechanism related to effect of helminth-expanded colonic Tregs on the mitigation of IBD has not been studied extensively (Lopes et al., 2016), even though several studies have showed that infection with intestinal helminths stimulated Tregs in the colon (Taylor et al., 2012) and were associated with therapeutic effects in colitis (Hang et al., 2013). Because of safety concerns in helminth therapy, it is more feasible and practical to identify the specific helminth-derived molecules that stimulate Tregs’ response as an immunotherapeutic target for chronic IBD and other inflammatory diseases.
Trichinella spiralis is a tissue-dwelling nematode with adult worms residing in small intestinal mucosa and larvae migrating to striated muscle to form encapsulated larvae. In order to survive in a hostile environment, T. spiralis adult worms and larvae secrete various proteins or other small molecules to modulate the host’s immune system to counter the host’s immunological attack (Qi et al., 2018; Song et al., 2018). Our previous study found that excretory/secretory (ES) products of T. spiralis adult worms are able to induce Treg proliferation through activating dendritic cells (Sun et al., 2019). Experimental treatment with T. spiralis ES products significantly reduced colitis induced by dextran sodium sulfate (DSS) in a mouse model through stimulating Tregs (Yang et al., 2014; Wang et al., 2020) or macrophage M2 polarization (Wang et al., 2020) to reduce inflammation in the colon. The adult worm extracts of T. spiralis also stimulated regulatory cytokines and reduced OVA-induced airway inflammation (Yuan et al., 2019). In an effort to identify the effective components in the nematode-secreted proteins that play immunomodulatory functions in host immune systems, it was found that paramyosin as a structural protein expressed on the surface and secreted products of T. spiralis larval and adult worms (TsPmy) is one of the major proteins involved in the immunomodulation of the host immune response through binding to human complement C1q (Sun et al., 2015; Wang et al., 2018) and C8/C9 (Zhang et al., 2011; Zhao et al., 2014) to inhibit complement response. The C9 binding domain of rTsPmy coupled with a membrane-bound signal reduced complement-related arthritis in a mouse model (Chen et al., 2020). Also, rTsPmy was able to activate systemic Tregs and induce the differentiation of Tregs through tolerogenic DCs (Guo et al., 2016). However, there is little knowledge about whether rTsPmy modulates intestinal Tregs and how these Tregs induced by rTsPmy maintains intestinal immune homeostasis in the inflammatory environment. In the present study, we demonstrated for the first time that rTsPmy promotes the expansion of thymic-derived Treg cells (tTregs), and maintained the stability and suppressive function of Tregs in the colon as the major mechanism involved in the therapeutic effect of rTsPmy in experimental colitis in mice.
Female C57BL/6J mice (6–8 weeks) were provided by Beijing Vital River Laboratory Animal Technology Co., Ltd (Beijing, China). Female Foxp3eGFP reporter mice (B6.Cg-Foxp3TM 2Tch/J) and Rag1 KO mice (B6.129S7-Rag1TM 1Mom/J) were purchased from Jackson Laboratory (Bar Harbor, ME, United States). All mice were housed under specific pathogen-free conditions in the Laboratory Animal Services Center of Capital Medical University (Beijing, China) according to the NIH Guidelines for the Care and Use of Laboratory Animals. All experimental procedures were approved by the Capital Medical University Animal Care and Use Committee under projects AEEI-2017-140, AEEI-2016-008 and AEEI-2017-133.
Trichinella spiralis paramyosin (GenBank# ABO09862.1) was expressed as recombinant protein (rTsPmy) in the baculovirus insect expression system (Invitrogen, Carlsbad, CA, United States). The rTsPmy with hexahistidine-tag at the C-terminus was expressed as partially soluble protein. After being denatured with guanidine hydrochloride, rTsPmy was purified by Ni-affinity chromatography (GE Healthcare, Boston, MA, United States) followed by a refolding process using a Protein Refolding Kit (Novagen/Merck KGaA, Darmstadt, Germany). The refolded soluble rTsPmy was characterized by SDS-PAGE and could be recognized by anti-rTsPmy sera by western blot.
Acute colitis was induced in female C57BL/6J mice with DSS as previously described (Wirtz et al., 2017). In brief, mice received 3% DSS (36,000–50,000 MW, MP Biomedicals, Solon, OH, United States) in drinking water for 6 days followed by regular drinking water for 2 days. The pathological changes of DSS-induced colitis were evaluated by the following criteria:
The clinical scoring of colitis was measured by the weight loss, stool consistency (diarrhea), and rectal bleeding as described (Yang et al., 2014; Table 1) from day 0 to day 8 using following criteria. The disease activity index (DAI) was calculated based on the sum of these scoring criteria.

Table 1. The criteria for scoring DSS-induced colitis.
On day 8, all mice were euthanized, and the distal colons were fixed in 4% paraformaldehyde. Paraffin-embedded sections (5 μm) were stained with hematoxylin and eosin (H&E). Histological activity score was assessed as the sum of two parameters as follows (Okayasu et al., 1990): Crypt damage, 0 = none; 1 = basal 1/3 lost; 2 = basal 2/3 lost; 3 = crypts lost but surface epithelium present; 4 = entire crypt and epithelium lost. Inflammatory cell infiltration, 0 = none; 1 = infiltration around crypt bases; 2 = infiltration in the muscularis mucosa; 3 = extensive infiltration in the muscularis mucosa with edema; 4 = infiltration of the submucosa.
The colon tissue sections were reacted with rat anti-MPO mAb (Servicebio, Wuhan, China) at 1:500 dilution at 4°C overnight and then incubated with HRP-conjugated rabbit anti-rat IgG at room temperature for 50 min, followed by the diaminobenzidin (DAB) chromogenic reaction for MPO detection. The nuclei on all slides were counterstained with hematoxylin.
Female C57BL/6J mice were intraperitoneally treated with 20 μg of rTsPmy for three times at 2-week intervals. Right after the last treatment, all mice were challenged with DSS daily as described above. Each mouse was boosted with another treatment of rTsPmy (20 μg) on the day 5 of DSS induction. The pathological changes of DSS-induced colitis were measured as above.
To identify CD4+ T cells, colon sections were obtained from Foxp3eGFP reporter mice and reacted with rabbit anti-CD4 mAb (Servicebio, Wuhan, China) at 1:400 dilution at 4°C overnight and then incubated with Cy3-conjugated goat anti-rabbit IgG secondary antibody for CD4 detection. All slides were counterstained for nuclei with DAPI (1:5000 dilution, Sigma, St. Louis, MO, United States). Slides were observed using the Panoramic 250 Flash system (3DHISTECH, Budapest, Hungary) with Panoramic Viewer software (3DHISTECH, Budapest, Hungary).
The colons were removed and placed in Ca2+, Mg2+-free Hank’s balanced salt solution (HBSS; Gibco, Carlsbad, CA, United States). Colonic lamina propria (cLP) cells were isolated using the Lamina Propria Dissociation Kit (Miltenyi Biotec, GmBH, Cologne, Germany) in a gentleMACS Octo Dissociator at 37°C. In brief, the colons were incubated with Ca2+/Mg2+-free HBSS containing 1 mM dithiothreitol (DTT, Absin, Shanghai, China), 5 mM EDTA (Calbiochem, Merck, Darmstadt, Germany), 5% FBS (Gibco, Carlsbad, CA, United States), 100 U/mL penicillin, 100 U/mL streptomycin and 10 mM HEPES (Absin, Shanghai, China) for 50 min for two times at 37°C on a MACSmix Tube Rotator to remove epithelial layers. The remaining colon pieces were digested with an enzyme mix from a Lamina Propria Dissociation Kit for 30 min. The suspension was filtered through a MACS SmartStrainer (40 μm) and the single-cell suspension was washed once with PBS containing 5% FBS.
Epithelial layer cells were cultured with 25 ng/ml PMA (phorbol 12-myristate-13-acetate) and 1 μg/ml ionomycin (Sigma) for 48 h. The cLP cells were stimulated with 2 μg/mL anti-CD3 and 1 μg/mL anti-CD28 mAbs (Biolegend, San Diego, CA, United States) for 48 h. Different cytokines in cell culture supernatants were measured by the multiplex Luminex immunoassay platform or ELISA.
All detected antibodies are listed in Table 2. The suspended cells were pre-incubated with Zombie Violet Fixable Viability Kit (Biolegend, San Diego, CA, United States) to detect dead cells. FcγR was blocked with anti-CD16/CD32 antibody (clone 93, eBioscience, San Diego, CA, United States). For intracellular cytokine staining, LP cells (2 × 106/mL) were stimulated with a Cell Activation Cocktail (with Brefeldin A) (Biolegend) in complete medium (RPMI 1640 containing 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin) in 24-well plates for 5 h, then stained for intracellular cytokines IL-17A and IFN-γ using a Cytofix/Cytoperm kit (BD Bioscience, Franklin Lakes, NJ, United States) after staining surface antigens CD3ε and CD4. For intranuclear transcription factors staining, LP cells were stained for surface antigens CD3ε, CD4, and CD25, followed by staining intranuclear transcription factors for Foxp3, GATA3, and Helios using the Foxp3/Transcription Factor Fixation/Permeabilization staining buffer kit (eBioscience).
Table 2. The key resources in this study.
The stained samples were detected on BD LSRFortessa with DIVA software (BD Biosciences) or Cytek Aurora Spectral Flow Cytometry with SpectroFlo2.2.0 (Cytek Biosciences, San Jose, CA, United States). For Treg subsets, the lymphocytes were gated by FSC vs. SSC, and the singlets were gated by FSC-A and FSC-H, and CD3+CD4+ T cells were gated by CD3 and CD4 fluorescencent antibodies, Foxp3+ Treg cells were gated by Foxp3 and CD4 fluorescencent antibodies. For Th17, Th1, and Treg cells, the lymphocytes were gated by FSC vs. SSC, and the singlets were gated by FSC-A and FSC-H, and CD3+CD4+ T cells were gated by CD3 and CD4 fluorescencent antibodies. In each experiment, the collected cells were pooled and stained with one specific directly labeled antibody to set up the compensation parameters. Background fluorescence was assessed by staining with isotype-matched control mAbs. Data were analyzed using FlowJo software (Tomy Digital Biology, Tokyo, Japan).
To establish T-cell-induced colitis in Rag1 KO mice lacking mature T and B lymphocytes, CD4+ T cells were isolated from spleens of Foxp3eGFP reporter mice using the MACS CD4+ T-cell Isolation Kit (Miltenyi Biotec). CD45RBhiCD4+Foxp3– naïve T cells were purified via FACS sorting on FACSAria IIIu (BD Biosciences) to reach over 98%. The naïve T cells were adoptively transferred into Rag1 KO mice (1 × 106/0.1 mL per mouse) through tail vein injection. To observe the role of rTsPmy-induced Tregs in reducing naïve T cell-induced colitis in Rag1 KO mice, 1 week after being transferred with naïve T cells, the Rag1 KO mice were passively transferred with 1 × 105 Treg cells isolated from cLP of Foxp3eGFP reporter mice treated with 20 μg rTsPmy or PBS for four times. The body weight was recorded weekly. At 4 weeks after Treg cell transfer, the severity of colitis was evaluated by H&E staining and ELISA.
Data are expressed as the mean ± SEM, and the differences between groups were analyzed using either unpaired two-tailed Student’s t-test or one-way ANOVA with the Bonferroni correction. A value of p < 0.05 is considered statistically significant.
To evaluate whether rTsPmy has prophylactic and therapeutic effects on DSS-induced acute colitis, mice were each intraperitoneally administered 20 μg rTsPmy for three times before DSS induction and boosted with one more treatment 5 days after DSS induction. The DSS-induced mice without treatment had typical clinical colitis, including progressive weight loss and diarrhea with bloody feces from day 3 to day 8 after DSS induction. In contrast, treatment with rTsPmy significantly reduced these clinical manifestations and the disease activity index as well compared to DSS-induced mice without treatment (receiving PBS only) (Figures 1A,B). In terms of pathological changes, the colons in mice with DSS-induced colitis were significantly shortened due to severe inflammatory edema, whereas, treatment with rTsPmy inhibited the colon shortening (Figure 1C). Histopathological sections displayed severe loss of crypts and infiltration of inflammatory cells (Figures 2A,B), especially neutrophils within the cLP extending to the muscularis mucosae and submucosa (Figures 2C,D), whereas administration of rTsPmy significantly alleviated the pathologic damage and reduced infiltration of inflammatory cells, including neutrophils, in the colon (Figures 2A–D). The colitis-characterized pro-inflammatory cytokines such as TNF-α secreted by epithelial layer cells and IL-1β of colon homogenate were significantly decreased in rTsPmy-treated mice in comparison to PBS-treated mice (Figures 2E,F). These observations indicate that rTsPmy suppresses the development of acute DSS-induced colitis and reduces colon inflammation.
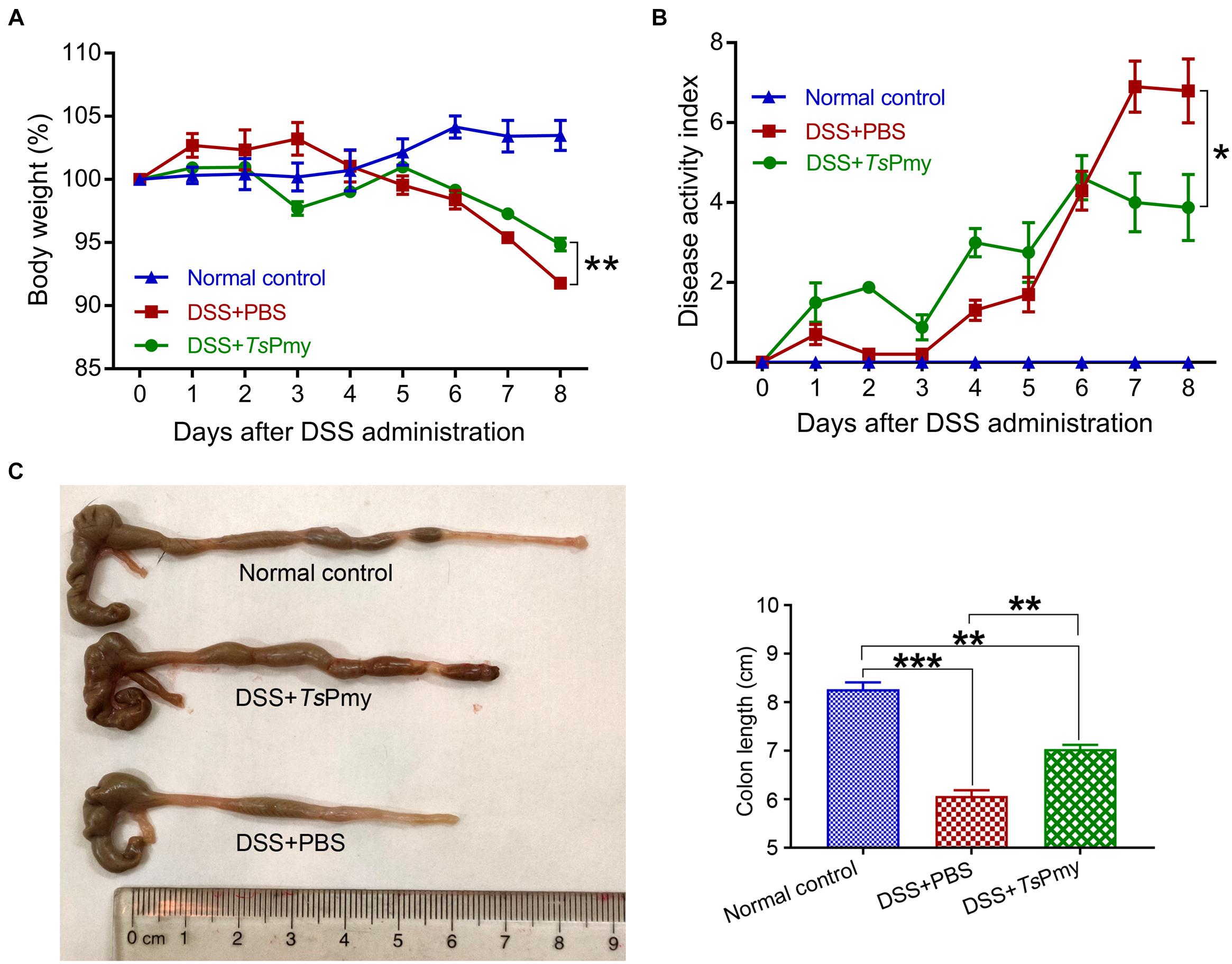
Figure 1. Treatment with rTsPmy alleviates clinical manifestations and colon shortening in DSS-induced acute colitis. Each mouse was treated with 20 μg of rTsPmy three times before being challenged with 3% DSS for 6 days. (A) Body weight changes (relative to initial weight) (n = 5). (B) The whole diseases activity index changes (n = 5). (C) Representative colons from each group are shown on the left; the average colon length from each group is shown on the right (n = 5). The data represent the mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
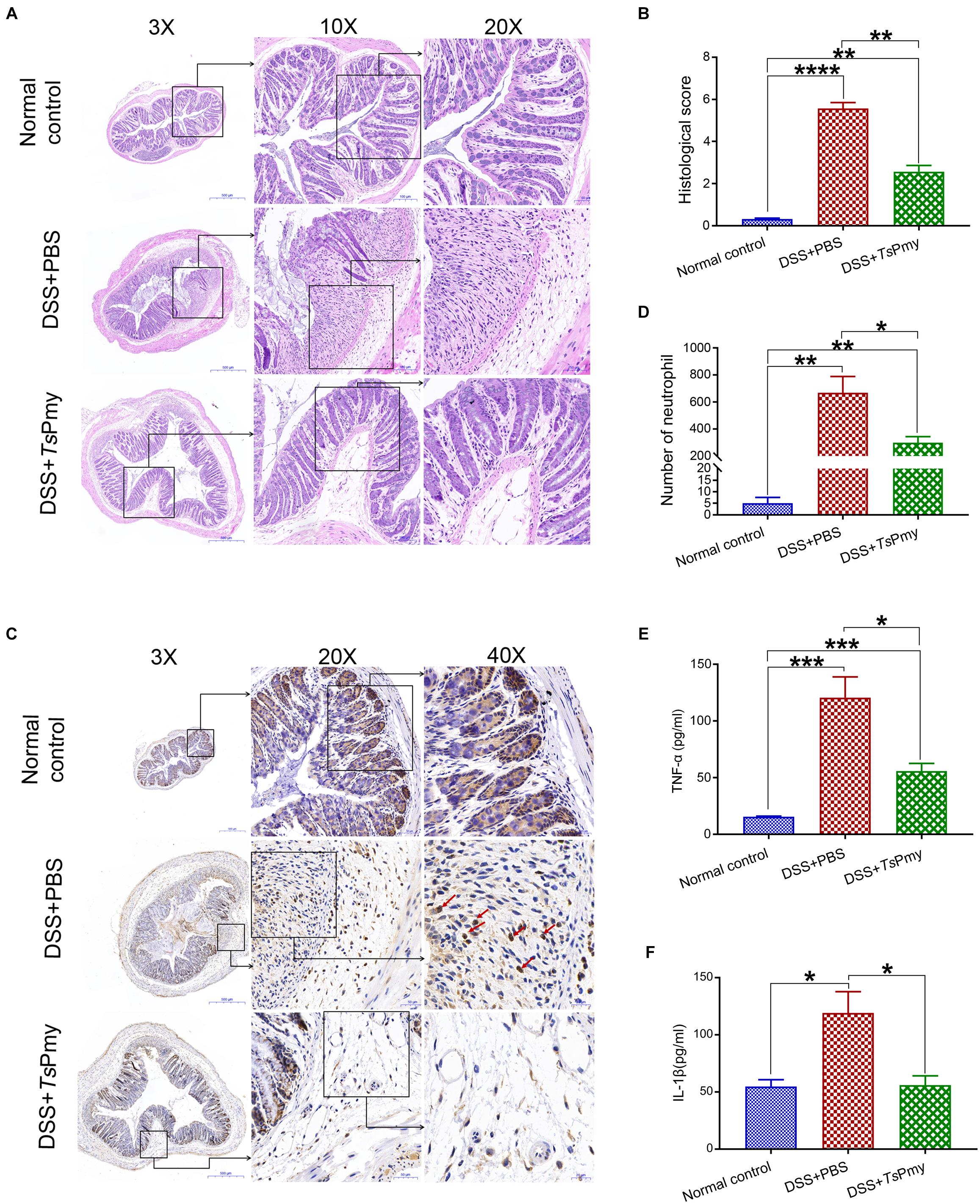
Figure 2. Treatment with rTsPmy improves the histopathological changes of DSS-induced acute colitis. (A) The representative histopathological sections of distal colon at objective 3x, 10x, and 20x. (B) The histological scores of colon in each group (n = 5). (C) Immunohistochemical (IHC) staining of myeloperoxidase (MPO) reflecting neutrophil infiltration (red arrows) in the distal colon. (D) The number of neutrophils of whole cross-section of distal colon in each group (n = 5). (E) The level of cytokines TNF-α derived by colonic epithelial layers cells (n = 5). (F) The level of cytokines IL-1β of colon homogenate (n = 5). The data represent the mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Excessive and prolonged activation of CD4+ effector T cells is the direct cause of intestinal inflammation and tissue damage in patients with IBD (Caruso et al., 2020). In this study, we observed that CD4+ T cells were significantly recruited to the isolated lymphoid follicles (ILFs) in the colon of mice with DSS-induced colitis (Figure 3A), indicating CD4+ effector T cells are involved in the inflammatory process of colitis. Treatment with rTsPmy significantly reduced the infiltration of CD4+ effector T cells in the colon of DSS-treated mice (Figure 3B). After being treated with rTsPmy, the CD3+CD4+ T cells expressing IL-17A and IFN-γ were significantly reduced in the colonic lamina propria mononuclear cells (LPMCs) of mice with DSS-induced colitis compared to group treated with PBS only (Figures 4A,B), indicating the colitis-related inflammatory Th1 and Th17 responses were inhibited by TsPmy treatment.
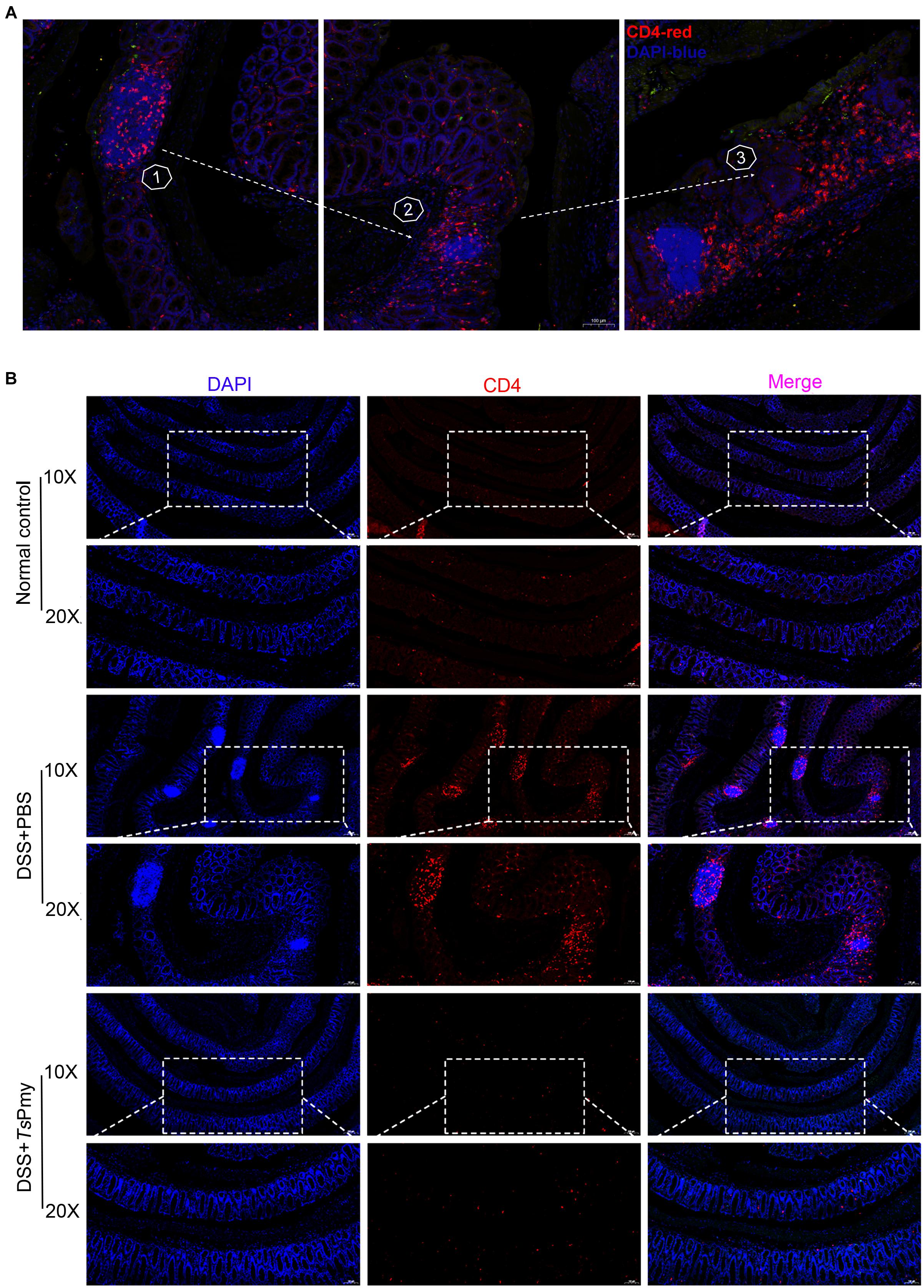
Figure 3. Treatment with rTsPmy reduces the recruitment of CD4+ effector cells in the isolated lymphoid follicles of mice with DSS-induced colitis. Immunofluorescent staining of the distal colon sections with antibodies for CD4 (red), counterstained with DAPI (blue). (A) Pathological process of DSS-induced colitis with increased recruitment of CD4+ effector cells in ILFs. Foxp3eGFP mice were used. (B) Distal colon sections of mice from different groups on day 5. Illustrations are representative of 3 mice per group. ILFs, isolated lymphoid follicles.
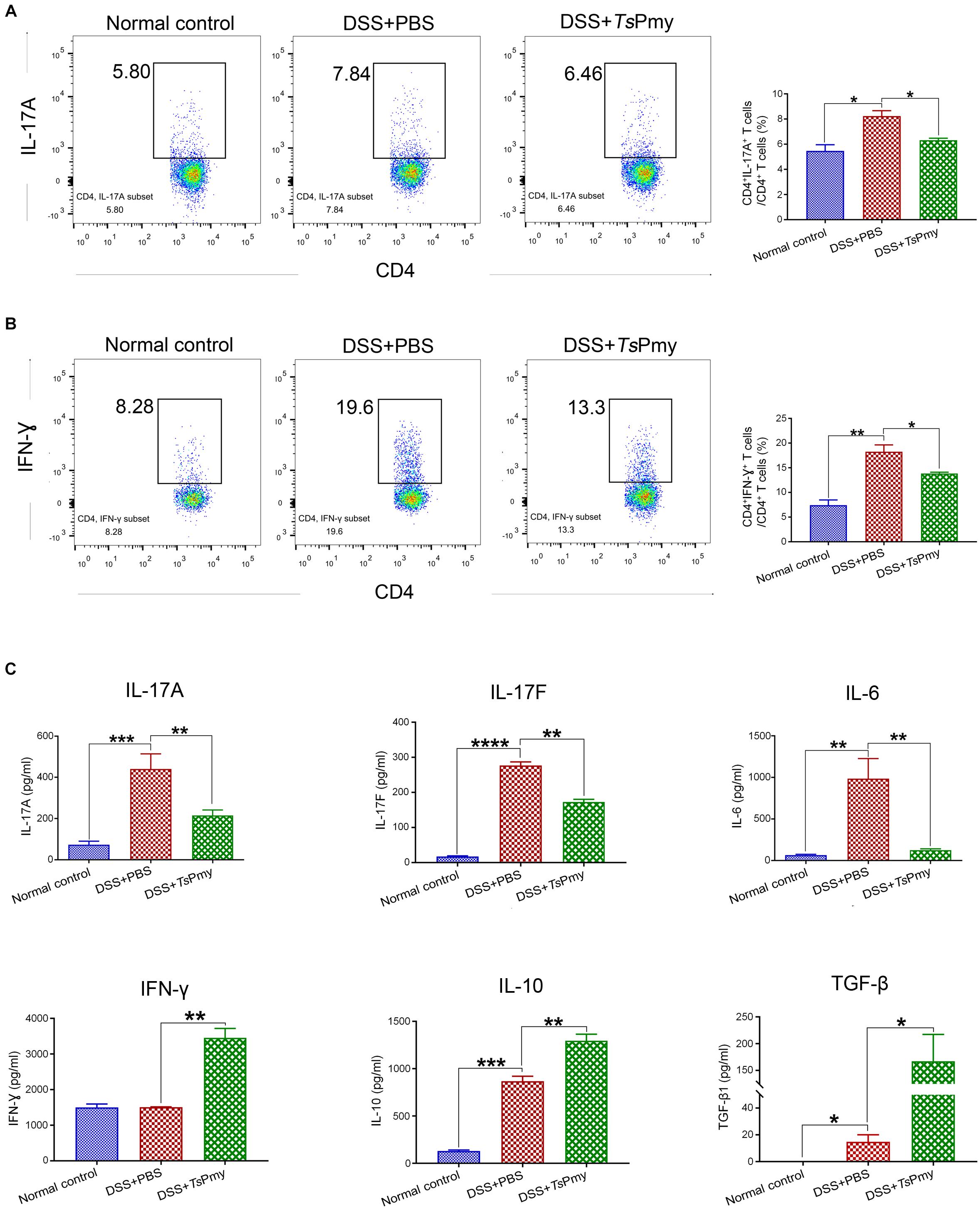
Figure 4. Treatment with rTsPmy reduces pro-inflammatory responses and enhances regulatory cytokines in cLP of mice with DSS-induced colitis. Lamina propria mononuclear cells (LPMCs) were isolated from distal colons of mice treated with rTsPmy or PBS on day 8. Flow cytometry shows CD3+CD4+ expressed with IL-17A (A) or IFN-γ (B). The gating strategy is shown in Supplementary Figure 1. The statistical analyses of the percentage of cells are shown on the right (n = 5). (C) After being stimulated with anti-CD3/CD28 mAbs for 48 h, LPMCs secretions of IL-17A, IL-17F, IL-6, IFN-γ, IL-10, and TGF-β were measured in the culture supernatants by Luminex or ELISA. The data represent the mean ± SEM (n = 3–5). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. cLP, colonic lamina propria.
The pro-inflammatory cytokine IL-17A, IL-17F, and IL-6 derived by LPMCs were significantly reduced in rTsPmy-treated mice; however, the level of IFN-γ was unexpectedly increased (Figure 4C). Meanwhile, the regulatory cytokines IL-10 and TGF-β were significantly increased in LPMCs of mice treated with rTsPmy compared to group that received PBS only (Figure 4C).
CD4+Foxp3+ Tregs are most abundant in the colonic mucosa and regulate the immune system in the intestine (Tanoue et al., 2016). Given that rTsPmy can suppress the development of acute DSS-induced colitis, it is suggested that rTsPmy may target the colonic Foxp3+ Tregs compartment involved with potential benefits for intestinal homeostasis. Four treatments with rTsPmy led to the increase of Foxp3+ Tregs in the colon, especially in the proximal colon of Foxp3eGFP reporter mice (data not shown). However, in the inflammatory condition of colitis, the number of CD4+Foxp3+ Tregs was reduced in the colon of mice with colitis treated with rTsPmy compared to those mice with colitis that received PBS only (Figure 5A). Further investigation identified that rTsPmy actually stimulated thymus-derived Tregs (tTregs) expressed with the transcription factor Helios (Helios+) rather than the peripherally derived Tregs (pTregs) in the cLP of mice with DSS-induced colitis (Figure 5B); the former plays a more suppressive and regulatory role in the extreme inflammatory environment (Clough et al., 2020). These results demonstrate that rTsPmy promotes tTregs expansion but not total Tregs pool in the colon.
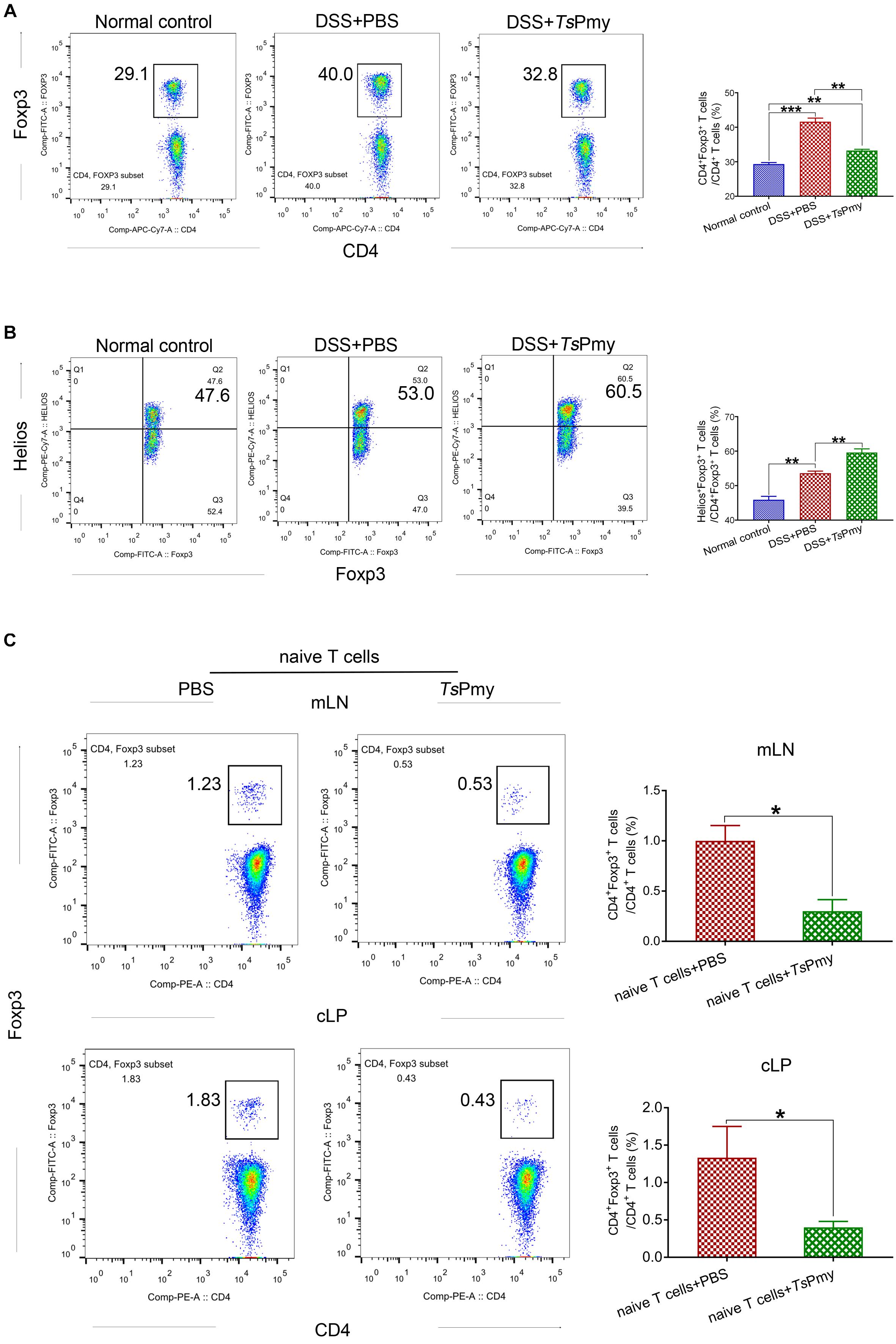
Figure 5. Treatment with rTsPmy specifically increases Helios+ tTreg expansion but could not induce pTreg differentiation in mLN and cLP in colitis. (A) The percentage of CD4+Foxp3+ Tregs in cLP from rTsPmy- or PBS-treated mice. The gating strategy is shown in Supplementary Figure 2. Statistical analysis is shown on the right (n = 5). (B) The frequency of Helios+ cells in Foxp3+ T cells of cLP from rTsPmy- or PBS-treated mice. The gate strategy is shown in Supplementary Figure 3. Statistical analysis is shown on the right (n = 5). (C) The percentage of Foxp3+ (eGFP) cells in CD3+CD4+ T cells of mLN (top) and cLP (bottom) at 5 weeks after CD4+Foxp3–CD45RBhi naïve T-cell adoptive transfer into Rag1 KO mice. The gate strategy is shown in Supplementary Figure 4. Frequencies are shown on the right (n = 3). Naïve T cells were from Foxp3eGFP mice. Results are shown as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001. mLN, mesenteric lymph nodes. cLP, colonic lamina propria. See also in Supplementary Figure 5.
To further confirm that rTsPmy could not induce the de novo generation of pTregs under inflammatory conditions, we adoptively transferred CD45RBhiFoxp3–CD4+ naïve T cells from Foxp3eGFP reporter mice into T/B lymphocyte-deficient Rag1 KO mice to induce T-cell-mediated inflammation in the colon (Chen et al., 2017), followed by treatment with rTsPmy to assess the differentiation of pTregs in colon in lymphopenia/inflammation. Analysis of pTregs in recipient mice 5 weeks post transfer revealed the sparse population of Foxp3+ (eGFP) pTregs converted from transferred naïve T cells in the cLP, mLN (Figure 5C), and spleen (Supplementary Figure 5) in rTsPmy-treated mice. These results suggest rTsPmy promotes tTregs expansion but is unable to induce the de novo generation of pTregs in the colon.
The characteristic Th2 cell transcription factor GATA binding protein 3 (GATA3) forms a complex with Foxp3 to enhance the stability of Tregs and facilitates their accumulation in inflamed intestines (Wohlfert et al., 2011; Rudra et al., 2012). GATA3 expression among Foxp3+ Tregs was assessed in this study and results showed that the proportion of Foxp3+ Tregs expressing GATA3 was significantly elevated in cLP from rTsPmy-treated mice with colitis compared with mice with colitis that received only PBS (Figure 6A). Further investigation showed that rTsPmy increased the GATA3+Helios+ tTregs subset in cLP of mice with colitis (Figure 6B).
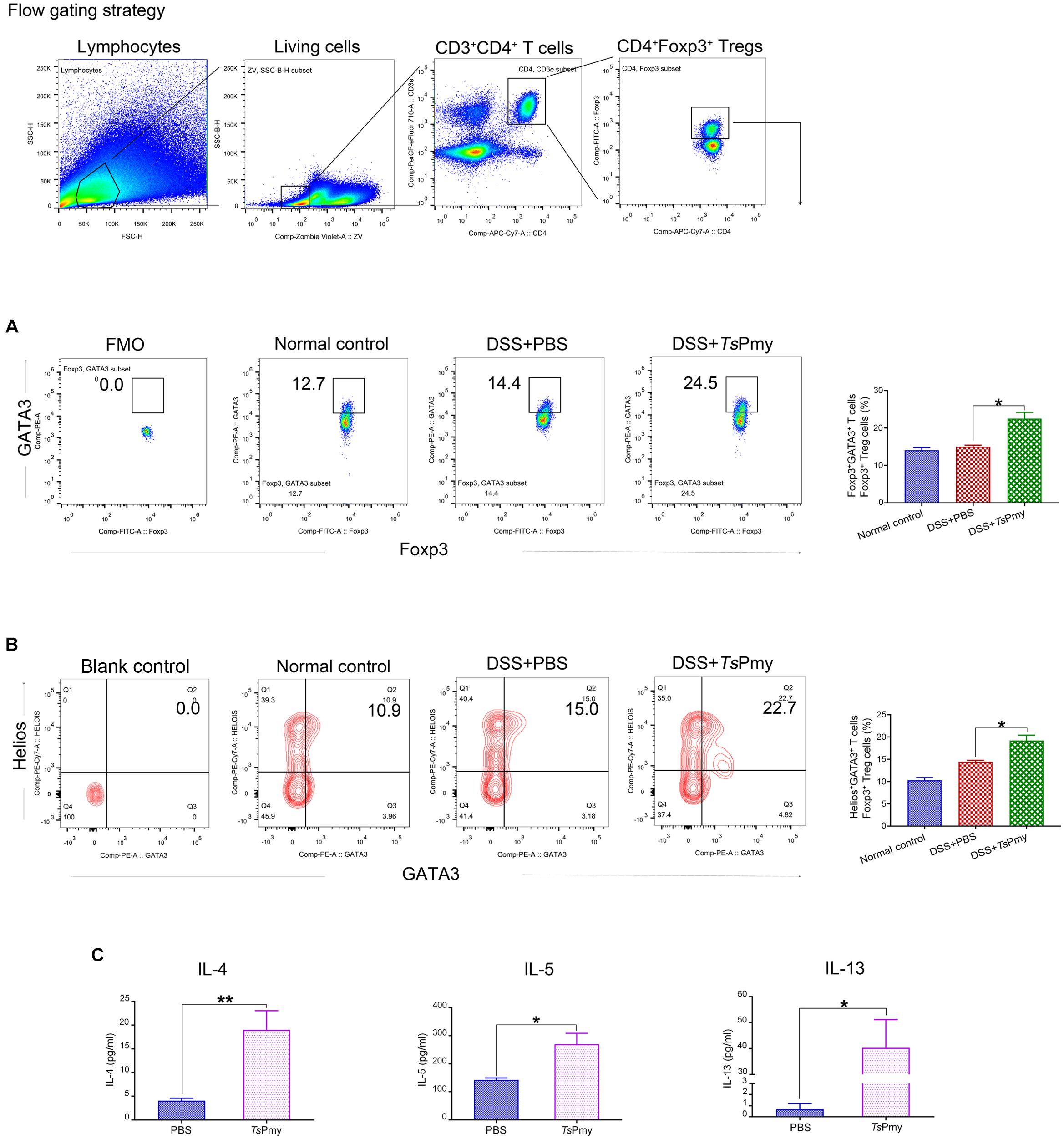
Figure 6. rTsPmy increases the GATA3+Helios+ Tregs subset in the Th2 environment. The percentage of GATA3+ cells (A) and Helios+GATA3+ cells (B) in Foxp3+ T cells of cLP from rTsPmy- or PBS-treated mice. The antibody of GATA3 (A) was cut in FMO control samples. Blank control of antibodies of GATA3 and Helios (B) was shown. Statistical analyses are shown on the right (n = 5). (C) The LPMCs were isolated from rTsPmy- or PBS-treated mice and stimulated with anti-CD3/CD28 mAb for 48 h. IL-4, IL-5, and IL-13 in the culture supernatants were measured by ELISA (n = 3). Error bars represent the mean ± SEM. *p < 0.05; **p < 0.01. LPMCs, Lamina propria mononuclear cells. See also in Supplementary Figure 6.
IL-4Rα signaling is crucial for the accumulation of GATA3+ Tregs in the inflamed intestine during helminth infections (Abdel et al., 2018); therefore we analyzed the Th2 cytokines IL-4, IL-5, and IL-13 secreted by LPMCs in the colon. LPMCs from rTsPmy-treated mice secreted higher levels of IL-4, IL-5, and IL-13 (Figure 6C). We also analyzed and found that the proportion of GATA3+Foxp3– Th2 subset in the colon was increased in rTsPmy-treated mice compared with their counterparts (Supplementary Figure 6). Taken together, these results suggest that treatment with rTsPmy up-regulates GATA3 expression in Foxp3+ Tregs in the Th2 environment, possibly enhancing the stability of Tregs in cLP of mice with colitis.
The Tregs (CD4+Foxp3+) expressing CD62LloCD44hi are considered effector regulatory T cells (eTregs) with suppressive functions in inflammatory sites (Tanoue et al., 2016). To determine if rTsPmy plays a role in maintaining Tregs’ suppressive status in the inflamed colon, the Tregs expressing CD62LloCD44hi in cLP were measured with flow cytometry. The results showed that the eTregs were decreased in cLP when mice had inflammatory colitis. Treatment with rTsPmy significantly increased the percentage of eTregs expressing CD62LloCD44hi in cLP in mice with colitis compared to mice with colitis that received PBS only (Figure 7A), suggesting that rTsPmy maintains the Tregs in an activated state in the colon. Tregs exert robust suppressive functions via signature phenotype molecules, such as CD25, TIGIT (T-cell immunoreceptor with Ig and ITIM domains), CTLA4 (cytotoxic T-lymphocyte antigen 4), and GITR (TNFR-related protein) (Clough et al., 2020). In this study, we found that treatment with rTsPmy significantly increased the expression of TIGIT (Figure 7B) within CD4+Foxp3+ T cells and the proportion of CTLA4+Foxp3+ Treg cells in CD4+ T cells (Figure 7C) in mice with colitis compared with mice with colitis that received PBS only.
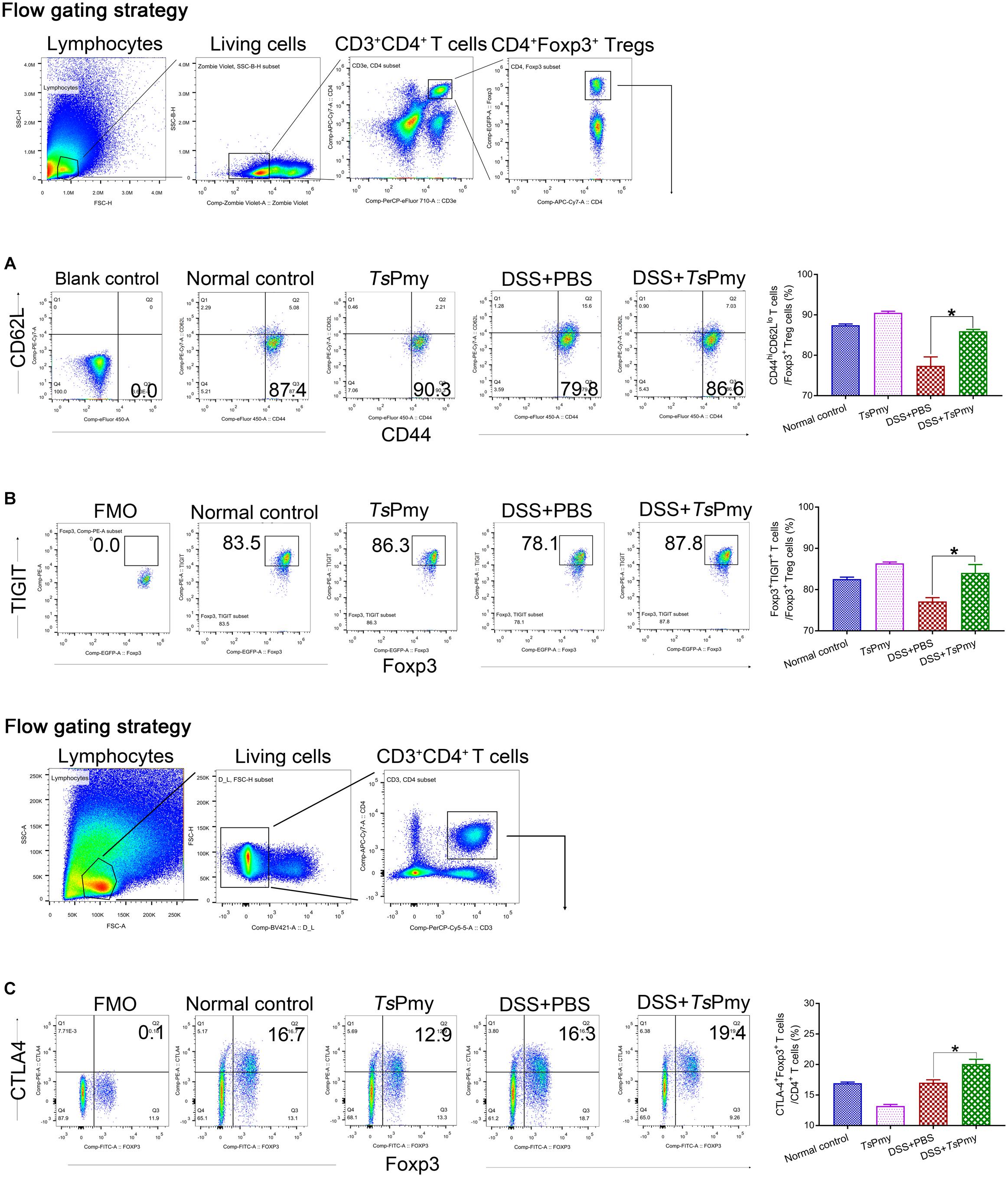
Figure 7. rTsPmy maintains majority of Tregs in an effective suppressor status (CD62LloCD44hi eTregs) with greater expression of functional phenotypes of TIGIT and CTLA4 in cLP with DSS-induced colitis. Flow cytometry results showed the frequencies of CD44hiCD62Llo cells (A) and TIGIT+ cells (B) in CD4+Foxp3+ T cells of cLP from rTsPmy or PBS-treated mice with or without DSS colitis. The blank control of antibodies of CD62L and CD44 (A) was shown, and the antibody of TIGIT (B) were cut in FMO control samples. The corresponding percentages are shown on the right (n = 5). (C) The frequency of Foxp3+CTLA4+ cells in CD3+CD4+ T cells. The antibody of CTLA4 was cut in FMO control samples. The corresponding percentage is shown on the right (n = 3–5). The bars represent the mean ± SEM. *p < 0.05. cLP, colonic lamina propria.
To determine if the inhibitory ability of rTsPmy-induced Tregs in colitis can be transferred to mice not receiving rTsPmy, we used a well-defined adoptive transfer model of Rag1 KO mice passively receiving Tregs isolated from cLP of Foxp3eGFP reporter mice treated with rTsPmy or PBS. Chronic colitis was induced in Rag1 KO mice by passive transfer with CD45RBhiCD4+CD25– naïve T cells from C57BL/6J mice exhibiting severe body weight loss (Figure 8A), loss of crypts in the colonic mucosa, infiltration of inflammatory cells (Figure 8B), and higher levels of IL-17A and IFN-γ production by LPMCs (Figure 8C). However, after being adoptively transferred with Tregs from cLP of Foxp3eGFP reporter donor mice treated with rTsPmy or PBS, these pathological conditions were significantly reduced in Rag1 KO mice with T-cell-induced colitis. More strikingly, the reduced pathology was more significant in Rag1 KO mice receiving rTsPmy-treated Tregs than in those receiving PBS-treated Tregs (Figures 8A–C). The results support that the inhibitory ability of rTsPmy-induced Tregs is transferable to mice to mitigate the inflammation in colon tissue.
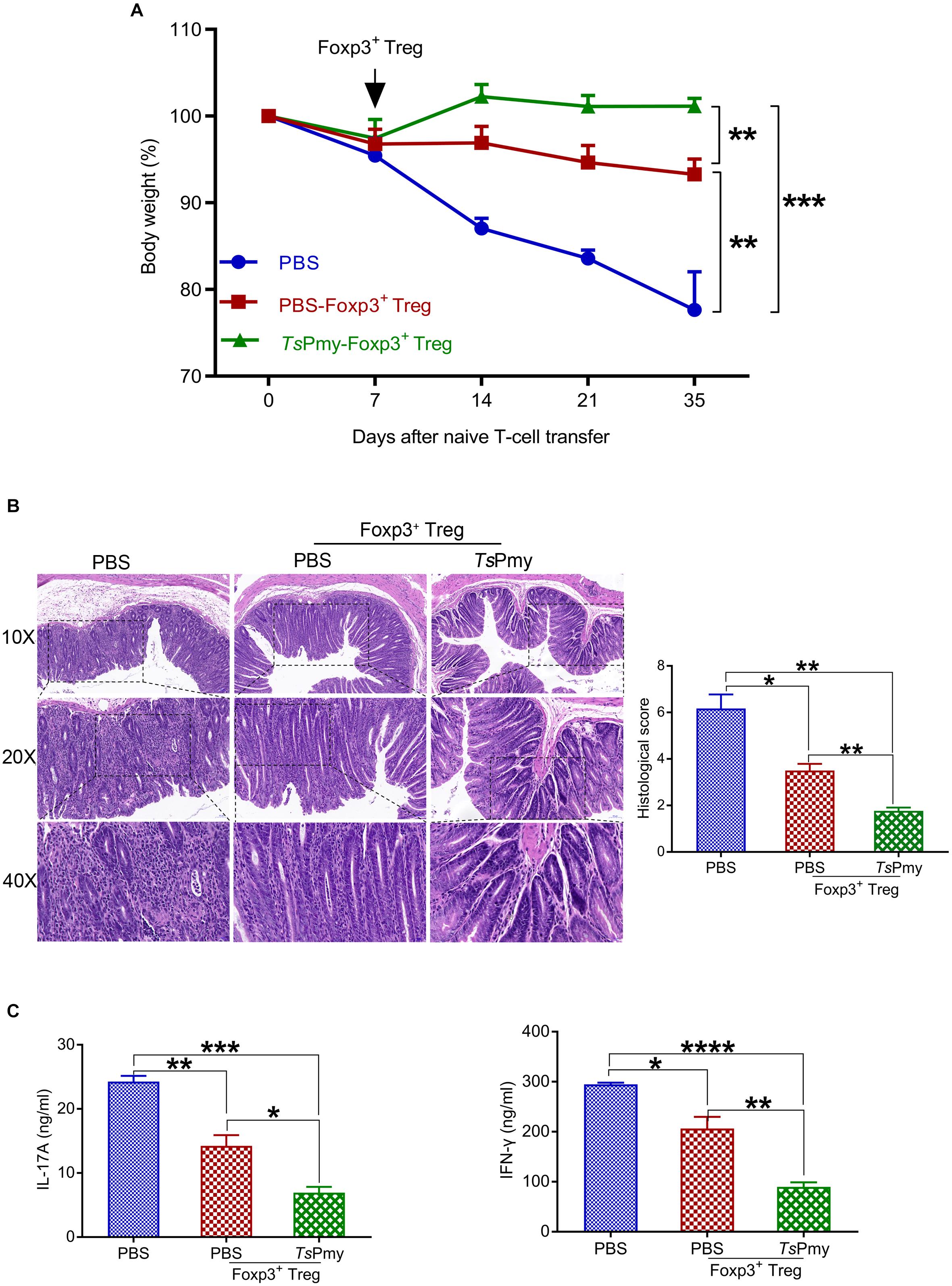
Figure 8. Adoptive transfer of rTsPmy-induced Tregs reduces the pathology and inflammation of naïve T-cell-induced colitis in Rag1 KO mice. (A) Changes in body weight (n = 3). (B) Hematoxylin and eosin (H&E) images of the distal colon at objective 10x, 20x, and 40x at 4 weeks after adoptive transfer of rTsPmy- or PBS-treated cLP Tregs compared to Rag1 KO mice receiving PBS. Histological scores of each group are displayed on the right (n = 3). (C) IL-17A and IFN-γ production of LPMCs in recipient Rag1 KO mice stimulated with anti-CD3/CD28 mAb for 48 h was determined by ELISA (n = 3). Data are presented as the mean ± SEM. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. LPMCs, Lamina propria mononuclear cells.
Inflammatory bowel disease is intestinal immune disorder caused by environmental factors and genetic susceptibility. Excessive immune response dominated by Th17 and Th1 is the major driver of intestinal tissue inflammation and damage in IBD patients (Xavier and Podolsky, 2007). Tregs play a critical role in suppressing these immune responses and maintaining immune homeostasis in the intestine (Tanoue et al., 2016). Decrease in Treg number and impaired Treg function have been related to the occurrence of IBD and other inflammatory diseases (Raffin et al., 2020). Immunomodulatory effects of helminth infection and helminth-derived molecules have been widely applied in the experimental immunotherapy of allergic or autoimmune inflammatory diseases; some of them have even been used in the clinical trials of allergic diseases (Feary et al., 2010; Daveson et al., 2011) or IBD (Summers et al., 2005; Mortimer et al., 2006). “Helminth therapy” in IBD includes the use of viable ova/larvae, helminth-derived crude extracts, purified molecules and cellular immunotherapy with helminth extract/antigen-pulsed immune cells (Lopes et al., 2016).
In this study, we demonstrated that treatment with rTsPmy, the major immunomodulatory protein expressed by T. spiralis, significantly reduced the development of DSS-induced colitis in a mouse model, including improvements in clinical manifestations and mitigation of pathology in the inflamed colon (Figures 1, 2). We observed the dynamic changes in CD4+ effector T cells involved in the pathological process of DSS-induced colitis in the mouse colon. Upon exposure to DSS, the effector CD4+ T cells are recruited to the ILFs in the colon. As disease progresses, the influx of CD4+ effector T cells from ILFs to the colonic mucosa leads to mucosal erosion (Figure 3A). After being treated with rTsPmy, the recruitment of the CD4+ effector T cells was significantly reduced and the pathological process of DSS-induced colitis was mitigated (Figure 3B). The activated excessive CD4+ effector T cells producing IL-17A and IFN-γ (Th17 and Th1) are believed to play pivotal roles in the pathogenesis of IBD (Xavier and Podolsky, 2007). The reduced pathology is correlated with reductions in Th1 and Th17 cells, neutrophil infiltration, and the corresponding cytokines such as IL-17A, IL-17F, and IL-6 in colonic LPMCs (Figures 2, 4). Even though we found that rTsPmy reduced the proportion of Th1 (CD4+IFN-γ+) cells in LPMCs, the IFN-γ level secreted by LPMCs was increased regardless of reductions in other pro-inflammatory cytokines IL-17A, IL-17F, and IL-6. The increased level of IFN-γ in colitis is possibly related to the increased level of T-bet that we observed in rTsPmy-treated colonic Tregs during colitis (data not shown). Tregs have been shown to up-regulate the Th1-related transcription factor T-bet among Tregs that may stimulate the secretion of the pro-inflammatory cytokine IFN-γ (Di Giovangiulio et al., 2019). Interestingly, we observed significantly increased IL-10 and TGF-β levels secreted by LPMCs of mice with colitis treated with rTsPmy, indicating the regulatory pathway may be involved in the reduced Th1/Th17 inflammatory response induced by the treatment of rTsPmy (Figure 4), which is consistent with the previous study that recombinant Schistosoma japonicum secreted protein Sj16 diminishes pro-inflammatory cytokine production but up-regulates immunoregulatory cytokine production IL-10 and TGF-β (Wang et al., 2017) in colonic mucosa. Therefore, we would like to explore if the regulatory T cells (Tregs) are involved in the alleviation of colitis during treatment with rTsPmy.
Tregs are critical for maintaining intestinal immune homeostasis, especially in the colon (Tanoue et al., 2016). It has been observed that helminth infections elevate the number of colonic Tregs and Foxp3 expression (Taylor et al., 2012) that are important in maintaining the immune balance in the colon (Gause and Maizels, 2016). In our study, we observed significantly increased abundance of Tregs in the colon in Foxp3eGFP reporter mice after treatment with rTsPmy (data not shown), and these Tregs were able to significantly inhibit chronic colitis induced by naïve T cells in Rag1 KO mice (Figure 8). However, the number of total Tregs was not increased in the colon of mice with colitis treated with rTsPmy (Figure 5A) even though the levels of IL-10 and TGF-β were significantly higher in the inflamed colon (Figure 4C). Further investigation of the phenotype of Tregs found that the thymus-derived Tregs (tTregs) expressed with Helios were significantly increased in cLP of mice with DSS-induced colitis treated with rTsPmy (Figure 5B). Passive transfer with naïve T cells from Foxp3eGFP reporter mice into T/B lymphocyte cell-deficient Rag1 KO mice followed by the treatment of rTsPmy showed that treatment with rTsPmy did not induce, and even inhibited, the de novo generation of pTregs in the colon of recipient mice (Figure 5C). The results indicate that rTsPmy only promotes tTregs expansion but not the differentiation of pTregs in the colon in an inflammatory environment. Colonic Foxp3+ Tregs comprise thymus-derived Tregs (tTregs) and peripherally derived Tregs (pTregs) (Tanoue et al., 2016). The tTregs complete their development in the thymus where they escape negative selection and then migrate to the colon, while pTregs are generated from naïve T cells in the colon microenvironment that is rich in TGF-β, microbial antigens and metabolites (Tanoue et al., 2016). The tTregs, rather than pTregs maintain stable suppressive activity in the extreme inflammatory environment (Clough et al., 2020). The specific stimulation of tTregs by rTsPmy indicates the specific regulation of tTregs in the inflamed colon by helminth-derived protein. A recent study reported that Nippostrongylus brasiliensis and Heligmosomoides polygyrus bakeri drove the ST2+GATA3+ tTreg responses in intestinal mucosa through DC-derived IL-33 that suppressed helminth immunity (Schiering et al., 2014; Hung et al., 2020). During helminth infection, tTregs are believed to produce the initial regulation of Th2 protective immune responses, while pTregs are developed more slowly. Mice depleted of natural Tregs prior to filarial or S. mansoni infections showed an increased Th2-mediated worm expulsion, suggesting that tTregs play an important role in the early stage of infection (Hesse et al., 2004; Baumgart et al., 2006). More studies showed that helminth infection rapidly expanded natural CD4+Foxp3+ regulatory T cells (tTregs) to inhibit host protective immunity against helminth infection while adaptive pTregs were differentiated slowly with limited protection against helminth infection (Taylor et al., 2012). The adaptive pTregs responded slowly to helminth infection through a TGF-β mimic derived from a helminth antigen and may suppress protective immunity during a later infection stage (Grainger et al., 2010). Blockade of TGF-βR signaling after 4 weeks of H. polygyrus infection inhibited the generation of pTregs thereby increasing parasite killing (Grainger et al., 2010). The tTregs and pTregs may have distinct yet overlapping functions as regulators during helminth infection. In this study, we demonstrated that rTsPmy induced and expanded tTregs in the colon; however, it is still unknown if the induced tTregs are rTsPmy antigen-specific. It will be important to determine the antigenic specificity of rTsPmy-activated tTregs and its specific role in the inhibition of inflammatory diseases.
After we showed that rTsPmy was able to stimulate and expand the tTregs in colitis, the question remains if rTsPmy can maintain the stability of Tregs in the colon so they can exert their regulatory functions. The tTregs expressing GATA3 exhibit stable suppressive activity and do not undergo reprogramming in the extreme inflammatory environment (Tanoue et al., 2016). Intrinsic GATA3 expression by Tregs is essential to sustain high levels of Foxp3 expression in an inflammatory setting and is required for efficient accumulation of Tregs at inflamed sites (Rudra et al., 2012). In this study, we showed that treatment with rTsPmy significantly increased GATA3 expression in Foxp3+ Tregs in cLP of mice with colitis (Figure 6A). Nearly all GATA3+ Tregs also expressed Helios in this study (Figure 6B), indicating their origin in tTregs. These rTsPmy-induced GATA3+Helios+ Tregs should be a functional tTregs subset with more stability (Tanoue et al., 2016). It is believed that IL-4Rα signaling is crucial to up-regulate GATA3 in Tregs and promote the accumulation of GATA3+ Tregs in the inflamed intestine during helminth infection (Abdel et al., 2018). We investigated Th2 responses in LPMCs and found that rTsPmy elicited robust Th2 immune responses in the colon including strong secretion of IL-4, IL-5, and IL-13 (Figure 6C). The results indicate that the rTsPmy-induced Th2 environment facilitates GATA3 expression in Tregs, and therefore increases the stability of Tregs in the colon to exert a suppressive function on DSS-induced colitis.
To determine if rTsPmy plays a role in stimulating Tregs to maintain their suppressive status in inflammatory colon, we measured the effector regulatory T cells (eTregs) which express CD62LloCD44hi (Tanoue et al., 2016) in the colon in colitis. Our results identified that rTsPmy significantly increased the percentage of eTregs expressing CD62LloCD44hi in cLP of mice with colitis compared to mice treated with PBS only. The increased eTregs also expressed suppressive signature phenotype molecules of TIGIT and CTLA4 (Joller et al., 2014; Clough et al., 2020; Figure 7), further suggesting rTsPmy up-regulates eTregs to maintain effective suppressive status in the inflammatory colon. However, the increased expression of CTLA4 was observed only in CD4+Foxp3+ T cells in colitis. In normal mice, treatment with rTsPmy increased only CTLA4 in CD4+Foxp3– T cells (data not shown), not in CD4+Foxp3+ T cells, which is consistent with the results of infections with H. polygyrus, Strongyloides ratti, Brugia malayi, and Litomosoides sigmodontis (Finney et al., 2007; Taylor et al., 2007; Walsh et al., 2007; McSorley et al., 2008). The up-regulation of CTLA4 in Tregs in inflammatory conditions indicates that the helminth-secreted immunomodulatory molecules regulate Treg function more effectively when there is a greater degree of inflammation (D’Elia et al., 2009).
The suppressive effect of rTsPmy-induced Tregs was also confirmed by passive transfer to Rag1 KO mice with chronic colitis induced by CD45RBhiCD4+CD25– naïve T cells; they had fewer clinical manifestations and pathological damage in colon that was associated with reduced levels of IL-17A and IFN-γ production in colonic LPMCs. The findings further confirm that the therapeutic efficacy of rTsPmy in colitis results from induction of Tregs. The results are consistent with the investigation of H. polygyrus infection-induced Tregs, in which adoptive transfer of Tregs from the cLP of H. polygyrus-infected mice as opposed to uninfected-mice significantly inhibited chronic colitis induced by CD4+CD25– T cells in Rag1 KO mice (Hang et al., 2013).
We demonstrated the prophylactic and therapeutic effects of rTsPmy in the experimental colitis associated with colonic Tregs activation in this study. The therapeutic effect of rTsPmy-induced Tregs was transferrable to mice with chronic colitis. The more significant finding in this study is that rTsPmy specifically stimulated the differentiation and expansion of thymic-derived Tregs but not peripherally derived Tregs; the former play more suppressive and regulatory roles in the extreme inflammatory environment. Treatment with rTsPmy also enhanced the differentiation of effector Tregs with greater suppressive function in the colon with colitis. In addition, rTsPmy enhanced the stability of effective Tregs in the inflamed colon by stimulating the expression of GATA3 on Foxp3+ Tregs in Th2 environment. The findings in this study describe the systemic and dynamic activation of effective Tregs induced by a major immunomodulatory protein rTsPmy and the roles of rTsPmy-induced Tregs in maintaining gut immune homeostasis during inflammation, providing further insight into the immunological mechanisms involved in the therapeutic effect of helminth-derived proteins in inflammatory bowel diseases.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
The animal study was reviewed and approved by the Capital Medical University Animal Care and Use Committee under projects AEEI-2017-140, AEEI-2016-008, and AEEI-2017-133.
CH: investigation, methodology, formal analysis, and writing – original draft. WW: validation and methodology. BZ: writing – review and editing. ZW: methodology. JH: resources. XS and XZ: conceptualization, supervision, funding acquisition, and writing – review and editing. All authors discussed the results, commented on the manuscript text, and approved the final version submitted.
This work was supported by the National Natural Science Foundation of China (81772213 and 81672042) and the Natural Science Foundation of Beijing (7144192).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Yuli Cheng, Yuan Gu, Sha Liu, Qing Sun, Zhihui Jia, Shuai Shao, SuQin Du, and Yan Yu for their technical assistance and helpful suggestions. We also thank the Fluorescence Microscopy Center and Flow Cytometry Center of Capital Medical University, Beijing, China for technical support and assistance.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2021.695015/full#supplementary-material
Abdel, A. N., Nono, J. K., Mpotje, T., and Brombacher, F. (2018). The Foxp3+ regulatory T-cell population requires IL-4Ralpha signaling to control inflammation during helminth infections. PLoS Biol. 16:e2005850. doi: 10.1371/journal.pbio.2005850
Abdel-Motal, U. M., Al-Shaibi, A., Elawad, M., and Lo, B. (2019). Zero tolerance! A perspective on monogenic disorders with defective regulatory T cells and IBD-like disease. Immunol. Rev. 287, 236–240. doi: 10.1111/imr.12717
Bach, J. F. (2018). The hygiene hypothesis in autoimmunity: the role of pathogens and commensals. Nat. Rev. Immunol. 18, 105–120. doi: 10.1038/nri.2017.111
Baumgart, M., Tompkins, F., Leng, J., and Hesse, M. (2006). Naturally occurring CD4+Foxp3+ regulatory T cells are an essential, IL-10-independent part of the immunoregulatory network in Schistosoma mansoni egg-induced inflammation. J. Immunol. 176, 5374–5387. doi: 10.4049/jimmunol.176.9.5374
Broadhurst, M. J., Leung, J. M., Kashyap, V., McCune, J. M., Mahadevan, U., McKerrow, J. H., et al. (2010). IL-22+ CD4+ T cells are associated with therapeutic trichuris trichiura infection in an ulcerative colitis patient. Sci. Transl. Med. 2:60ra88. doi: 10.1126/scitranslmed.3001500
Buning, J., Homann, N., von Smolinski, D., Borcherding, F., Noack, F., Stolte, M., et al. (2008). Helminths as governors of inflammatory bowel disease. Gut 57, 1182–1183. doi: 10.1136/gut.2008.152355
Canavan, J. B., Scotta, C., Vossenkamper, A., Goldberg, R., Elder, M. J., Shoval, I., et al. (2016). Developing in vitro expanded CD45RA+ regulatory T cells as an adoptive cell therapy for Crohn’s disease. Gut 65, 584–594. doi: 10.1136/gutjnl-2014-306919
Caruso, R., Lo, B. C., and Nunez, G. (2020). Host-microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 20, 411–426. doi: 10.1038/s41577-019-0268-7
Cheifetz, A. S., Gianotti, R., Luber, R., and Gibson, P. R. (2017). Complementary and alternative medicines used by patients with inflammatory bowel diseases. Gastroenterology 152, 415–429. doi: 10.1053/j.gastro.2016.10.004
Chen, W., Xu, Z., Zheng, Y., Wang, J., Qian, W., Olsen, N., et al. (2017). A protocol to develop T helper and Treg cells in vivo. Cell. Mol. Immunol. 14, 1013–1016. doi: 10.1038/cmi.2017.116
Chen, Y., Shao, S., Huang, J., Gu, Y., Cheng, Y., and Zhu, X. (2020). Therapeutic efficacy of a trichinella spiralis Paramyosin-Derived peptide modified with a Membrane-Targeting signal in mice with Antigen-Induced arthritis. Front. Microbiol. 11:608380. doi: 10.3389/fmicb.2020.608380
Clough, J. N., Omer, O. S., Tasker, S., Lord, G. M., and Irving, P. M. (2020). Regulatory T-cell therapy in Crohn’s disease: challenges and advances. Gut 69, 942–952. doi: 10.1136/gutjnl-2019-319850
Daveson, A. J., Jones, D. M., Gaze, S., McSorley, H., Clouston, A., Pascoe, A., et al. (2011). Effect of hookworm infection on wheat challenge in celiac disease–a randomised double-blinded placebo controlled trial. PLoS One 6:e17366. doi: 10.1371/journal.pone.0017366
D’Elia, R., Behnke, J. M., Bradley, J. E., and Else, K. J. (2009). Regulatory T cells: a role in the control of helminth-driven intestinal pathology and worm survival. J. Immunol. 182, 2340–2348. doi: 10.4049/jimmunol.0802767
Desreumaux, P., Foussat, A., Allez, M., Beaugerie, L., Hebuterne, X., Bouhnik, Y., et al. (2012). Safety and efficacy of antigen-specific regulatory T-cell therapy for patients with refractory Crohn’s disease. Gastroenterology 143, 1207–1217. doi: 10.1053/j.gastro.2012.07.116
Di Giovangiulio, M., Rizzo, A., Franze, E., Caprioli, F., Facciotti, F., Onali, S., et al. (2019). Tbet expression in regulatory t cells is required to initiate Th1-Mediated colitis. Front. Immunol. 10:2158. doi: 10.3389/fimmu.2019.02158
Feary, J. R., Venn, A. J., Mortimer, K., Brown, A. P., Hooi, D., Falcone, F. H., et al. (2010). Experimental hookworm infection: a randomized placebo-controlled trial in asthma. Clin. Exp. Allergy 40, 299–306. doi: 10.1111/j.1365-2222.2009.03433.x
Finney, C. A., Taylor, M. D., Wilson, M. S., and Maizels, R. M. (2007). Expansion and activation of CD4(+)CD25(+) regulatory T cells in Heligmosomoides polygyrus infection. Eur. J. Immunol. 37, 1874–1886. doi: 10.1002/eji.200636751
Gause, W. C., and Maizels, R. M. (2016). Macrobiota - helminths as active participants and partners of the microbiota in host intestinal homeostasis. Curr. Opin. Microbiol. 32, 14–18. doi: 10.1016/j.mib.2016.04.004
Gazzinelli-Guimaraes, P. H., and Nutman, T. B. (2018). Helminth parasites and immune regulation. F1000Res. 7:F1000FacultyRev–1685. doi: 10.12688/f1000research.15596.1
Grainger, J. R., Smith, K. A., Hewitson, J. P., McSorley, H. J., Harcus, Y., Filbey, K. J., et al. (2010). Helminth secretions induce de novo T cell Foxp3 expression and regulatory function through the TGF-beta pathway. J. Exp. Med. 207, 2331–2341. doi: 10.1084/jem.20101074
Guo, K., Sun, X., Gu, Y., Wang, Z., Huang, J., and Zhu, X. (2016). Trichinella spiralis paramyosin activates mouse bone marrow-derived dendritic cells and induces regulatory T cells. Parasit. Vectors 9, 569. doi: 10.1186/s13071-016-1857-y
Hang, L., Blum, A. M., Setiawan, T., Urban, J. J., Stoyanoff, K. M., and Weinstock, J. V. (2013). Heligmosomoides polygyrus bakeri infection activates colonic Foxp3+ T cells enhancing their capacity to prevent colitis. J. Immunol. 191, 1927–1934. doi: 10.4049/jimmunol.1201457
Hesse, M., Piccirillo, C. A., Belkaid, Y., Prufer, J., Mentink-Kane, M., Leusink, M., et al. (2004). The pathogenesis of schistosomiasis is controlled by cooperating IL-10-producing innate effector and regulatory T cells. J. Immunol. 172, 3157–3166. doi: 10.4049/jimmunol.172.5.3157
Hung, L. Y., Tanaka, Y., Herbine, K., Pastore, C., Singh, B., Ferguson, A., et al. (2020). Cellular context of IL-33 expression dictates impact on anti-helminth immunity. Sci. Immunol. 5:eabc6259. doi: 10.1126/sciimmunol.abc6259
Joller, N., Lozano, E., Burkett, P. R., Patel, B., Xiao, S., Zhu, C., et al. (2014). Treg cells expressing the coinhibitory molecule TIGIT selectively inhibit proinflammatory Th1 and Th17 cell responses. Immunity 40, 569–581. doi: 10.1016/j.immuni.2014.02.012
Kaplan, G. G. (2015). The global burden of IBD: from 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 12, 720–727. doi: 10.1038/nrgastro.2015.150
Lopes, F., Matisz, C., Reyes, J. L., Jijon, H., Al-Darmaki, A., Kaplan, G. G., et al. (2016). Helminth regulation of immunity: a three-pronged approach to treat colitis. Inflamm. Bowel Dis. 22, 2499–2512. doi: 10.1097/MIB.0000000000000889
Maloy, K. J., and Powrie, F. (2011). Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 474, 298–306. doi: 10.1038/nature10208
Mathew, J. M., H-Voss, J., LeFever, A., Konieczna, I., Stratton, C., He, J., et al. (2018). A phase i clinical trial with ex vivo expanded recipient regulatory t cells in living donor kidney transplants. Sci. Rep. 8:7428. doi: 10.1038/s41598-018-25574-7
McSorley, H. J., Harcus, Y. M., Murray, J., Taylor, M. D., and Maizels, R. M. (2008). Expansion of Foxp3+ regulatory T cells in mice infected with the filarial parasite Brugia malayi. J. Immunol. 181, 6456–6466. doi: 10.4049/jimmunol.181.9.6456
Mortimer, K., Brown, A., Feary, J., Jagger, C., Lewis, S., Antoniak, M., et al. (2006). Dose-ranging study for trials of therapeutic infection with Necator americanus in humans. Am. J. Trop. Med. Hyg. 75, 914–920. doi: 10.1016/j.actatropica.2006.09.009
Neurath, M. F. (2017). Current and emerging therapeutic targets for IBD. Nat. Rev. Gastroenterol. Hepatol. 14, 269–278. doi: 10.1038/nrgastro.2016.208
Ohkura, N., Kitagawa, Y., and Sakaguchi, S. (2013). Development and maintenance of regulatory T cells. Immunity 38, 414–423. doi: 10.1016/j.immuni.2013.03.002
Okayasu, I., Hatakeyama, S., Yamada, M., Ohkusa, T., Inagaki, Y., and Nakaya, R. (1990). A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 98, 694–702. doi: 10.1016/0016-5085(90)90290-h
Qi, X., Yue, X., Han, Y., Jiang, P., Yang, F., Lei, J. J., et al. (2018). Characterization of two trichinella spiralis Adult-Specific DNase II and their capacity to induce protective immunity. Front. Microbiol. 9:2504. doi: 10.3389/fmicb.2018.02504
Raffin, C., Vo, L. T., and Bluestone, J. A. (2020). Treg cell-based therapies: challenges and perspectives. Nat. Rev. Immunol. 20, 158–172. doi: 10.1038/s41577-019-0232-6
Reynolds, L. A., Finlay, B. B., and Maizels, R. M. (2015). Cohabitation in the intestine: interactions among helminth parasites, bacterial microbiota, and host immunity. J. Immunol. 195, 4059–4066. doi: 10.4049/jimmunol.1501432
Rudra, D., DeRoos, P., Chaudhry, A., Niec, R. E., Arvey, A., Samstein, R. M., et al. (2012). Transcription factor Foxp3 and its protein partners form a complex regulatory network. Nat. Immunol. 13, 1010–1019. doi: 10.1038/ni.2402
Schiering, C., Krausgruber, T., Chomka, A., Frohlich, A., Adelmann, K., Wohlfert, E. A., et al. (2014). The alarmin IL-33 promotes regulatory T-cell function in the intestine. Nature 513, 564–568. doi: 10.1038/nature13577
Shevach, E. M. (2009). Mechanisms of foxp3+ T regulatory cell-mediated suppression. Immunity 30, 636–645. doi: 10.1016/j.immuni.2009.04.010
Shevach, E. M. (2018). Foxp3(+) T Regulatory Cells: still Many Unanswered Questions-A Perspective After 20 Years of Study. Front. Immunol. 9:1048. doi: 10.3389/fimmu.2018.01048
Song, Y. Y., Zhang, Y., Yang, D., Ren, H. N., Sun, G. G., Jiang, P., et al. (2018). The immune protection induced by a serine protease inhibitor from the foodborne parasite trichinella spiralis. Front. Microbiol. 9:1544. doi: 10.3389/fmicb.2018.01544
Summers, R. W., Elliott, D. E., Urban, J. J., Thompson, R. A., and Weinstock, J. V. (2005). Trichuris suis therapy for active ulcerative colitis: a randomized controlled trial. Gastroenterology 128, 825–832. doi: 10.1053/j.gastro.2005.01.005
Sun, R., Zhao, X., Wang, Z., Yang, J., Zhao, L., Zhan, B., et al. (2015). Trichinella spiralis paramyosin binds human complement c1q and inhibits classical complement activation. PLoS Negl. Trop. Dis. 9:e4310. doi: 10.1371/journal.pntd.0004310
Sun, X. M., Guo, K., Hao, C. Y., Zhan, B., Huang, J. J., and Zhu, X. (2019). Trichinella spiralis Excretory-Secretory products stimulate host regulatory t cell differentiation through activating dendritic cells. Cells 8:1404. doi: 10.3390/cells8111404
Tanoue, T., Atarashi, K., and Honda, K. (2016). Development and maintenance of intestinal regulatory T cells. Nat. Rev. Immunol. 16, 295–309. doi: 10.1038/nri.2016.36
Taylor, M. D., Harris, A., Babayan, S. A., Bain, O., Culshaw, A., Allen, J. E., et al. (2007). CTLA-4 and CD4+ CD25+ regulatory T cells inhibit protective immunity to filarial parasites in vivo. J. Immunol. 179, 4626–4634. doi: 10.4049/jimmunol.179.7.4626
Taylor, M. D., van der Werf, N., and Maizels, R. M. (2012). T cells in helminth infection: the regulators and the regulated. Trends Immunol. 33, 181–189. doi: 10.1016/j.it.2012.01.001
Walsh, C. M., Smith, P., and Fallon, P. G. (2007). Role for CTLA-4 but not CD25+ T cells during Schistosoma mansoni infection of mice. Parasite Immunol. 29, 293–308. doi: 10.1111/j.1365-3024.2007.00947.x
Wang, L., Xie, H., Xu, L., Liao, Q., Wan, S., Yu, Z., et al. (2017). RSj16 protects against DSS-Induced colitis by inhibiting the PPAR-alpha signaling pathway. Theranostics 7, 3446–3460. doi: 10.7150/thno.20359
Wang, Z., Hao, C., Huang, J., Zhuang, Q., Zhan, B., and Zhu, X. (2018). Mapping of the complement C1q binding site on Trichinella spiralis paramyosin. Parasit. Vectors 11:666. doi: 10.1186/s13071-018-3258-x
Wang, Z., Hao, C., Zhuang, Q., Zhan, B., Sun, X., Huang, J., et al. (2020). Excretory/Secretory products from trichinella spiralis adult worms attenuated DSS-Induced colitis in mice by driving PD-1-Mediated m2 macrophage polarization. Front. Immunol. 11:563784. doi: 10.3389/fimmu.2020.563784
Weinstock, J. V. (2015). Do we need worms to promote immune health? Clin. Rev. Allergy Immunol. 49, 227–231. doi: 10.1007/s12016-014-8458-3
Wirtz, S., Popp, V., Kindermann, M., Gerlach, K., Weigmann, B., Fichtner-Feigl, S., et al. (2017). Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 12, 1295–1309. doi: 10.1038/nprot.2017.044
Wohlfert, E. A., Grainger, J. R., Bouladoux, N., Konkel, J. E., Oldenhove, G., Ribeiro, C. H., et al. (2011). GATA3 controls Foxp3(+) regulatory T cell fate during inflammation in mice. J. Clin. Invest. 121, 4503–4515. doi: 10.1172/JCI57456
Xavier, R. J., and Podolsky, D. K. (2007). Unravelling the pathogenesis of inflammatory bowel disease. Nature 448, 427–434. doi: 10.1038/nature06005
Yang, X., Yang, Y., Wang, Y., Zhan, B., Gu, Y., Cheng, Y., et al. (2014). Excretory/secretory products from Trichinella spiralis adult worms ameliorate DSS-induced colitis in mice. PLoS One 9:e96454. doi: 10.1371/journal.pone.0096454
Yuan, Y., Wang, L. Y., Mei, J., Cheng, Y., Wang, W., Chu, L., et al. (2019). [Protective effect of excretory-secretory protein from adult Trichinella spiralis on ovalbumin-induced allergic rhinitis in mice]. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 31, 504–509. doi: 10.16250/j.32.1374.2019069
Zhang, Z., Yang, J., Wei, J., Yang, Y., Chen, X., Zhao, X., et al. (2011). Trichinella spiralis paramyosin binds to C8 and C9 and protects the tissue-dwelling nematode from being attacked by host complement. PLoS Negl. Trop. Dis. 5:e1225. doi: 10.1371/journal.pntd.0001225
Zhao, X., Hao, Y., Yang, J., Gu, Y., and Zhu, X. (2014). Mapping of the complement C9 binding domain on Trichinella spiralis paramyosin. Parasit. Vectors 7:80. doi: 10.1186/1756-3305-7-80
Keywords: rTsPmy, Trichinella spiralis, regulatory T cells, inflammatory bowel disease, immunomodulation
Citation: Hao C, Wang W, Zhan B, Wang Z, Huang J, Sun X and Zhu X (2021) Trichinella spiralis Paramyosin Induces Colonic Regulatory T Cells to Mitigate Inflammatory Bowel Disease. Front. Cell Dev. Biol. 9:695015. doi: 10.3389/fcell.2021.695015
Received: 14 April 2021; Accepted: 07 June 2021;
Published: 15 July 2021.
Edited by:
Qingfeng Zhang, Tongji University, ChinaReviewed by:
Jun Zou, Georgia State University, United StatesCopyright © 2021 Hao, Wang, Zhan, Wang, Huang, Sun and Zhu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xinping Zhu, emh1eHBpbmdAY2NtdS5lZHUuY24=; Ximeng Sun, c3VueGltZW5nQGNjbXUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.