
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell Dev. Biol. , 30 April 2021
Sec. Signaling
Volume 9 - 2021 | https://doi.org/10.3389/fcell.2021.601929
 Sanjay Mishra1†
Sanjay Mishra1† Manish Charan1†
Manish Charan1† Ajeet Kumar Verma1Bhuvaneswari Ramaswamy2Dinesh Kumar Ahirwar1
Ajeet Kumar Verma1Bhuvaneswari Ramaswamy2Dinesh Kumar Ahirwar1 Ramesh K. Ganju1,2*
Ramesh K. Ganju1,2*Recent studies revealed that ethnic differences in mechanistic target of rapamycin (mTOR) and extracellular signal-regulated kinase (ERK-1/2) signaling pathways might be associated with the development and progression of different human malignancies. The African American (AA) population has an increased rate of cancer incidence and mortality compared to the Caucasian American (CA) population. Although the socioeconomic differences across different ethnic groups contribute to the disparity in developing different cancers, recent scientific evidence indicates the association of molecular and genetic variations in racial disparities of different human malignancies. The mTOR and ERK-1/2 signaling pathways are one of the well-known oncogenic signaling mechanisms that regulate diverse molecular and phenotypic aspects of normal as well as cancer cells in response to different external or internal stimuli. To date, very few studies have been carried out to explore the significance of racial disparity with abnormal mTOR and ERK-1/2 kinase signaling pathways, which may contribute to the development of aggressive human cancers. In this review, we discuss the differential regulation of mTOR and ERK-1/2 kinase signaling pathways across different ethnic groups, especially between AA and CA populations. Notably, we observed that key signaling proteins associated with mTOR and ERK-1/2 pathway including transforming growth factor-beta (TGF-β), Akt, and VEGFR showed racially disparate expression in cancer patients. Overall, this review article encompasses the significance of racially disparate signaling molecules related to mTOR/ERK1/2 and their potential in developing tailor-made anti-cancer therapies.
Recent reports on cancer research highlighted the significant impact of racial disparities on the clinical outcome of human malignancies (Esnaola and Ford, 2012; Heath et al., 2018; Freedman et al., 2021; Zavala et al., 2021). Cancer is one of the leading causes of death worldwide and witnessed major advancements in terms of cancer prevention, management, and newer treatments over the past few decades. Although these advancements have indeed improved the quality of life of cancer patients, statistical data on cancer-related mortalities, pointed out an inconsistency among certain ethnic groups regarding the disease incidence, aggressiveness, and clinical outcome (Gross et al., 2008; Esnaola and Ford, 2012). Cancer-related racial disparities have been reported among all ethnicities but the racial disparity has been extensively analyzed between the Caucasian American (CA) and African American (AA) population.
Many clinical factors such as aberrant genomic mutations, immune cell response, epigenetic modifications, and deregulated cell signaling pathways show racially disparate expression in several human cancers (Charan et al., 2020b). In recent years, numerous research studies around the world, have established a crucial role of the mechanistic target of rapamycin (mTOR) pathway in cellular growth and survival of mammalian cells (Soliman, 2013). mTOR is a Ser/Thr protein kinase that plays a key role in regulating cellular metabolism and immunity (Soliman, 2013). The dysregulation of mTOR pathway often leads to the initiation and progression of cancer. Several novel functions of the mTOR pathway in cancers have been discovered lately. mTOR is another essential kinase whose targeting has shown improved clinical efficacy against renal carcinoma (Battelli and Cho, 2011). Racial differences in African American renal cell carcinoma patients are well-recognized (Krishnan et al., 2016). Temsirolimus and everolimus are two FDA-approved drugs that exert anti-tumor effects through mTOR pathway inhibition (Voss et al., 2011). In a phase I study, everolimus showed higher renal clearance in AA population and, therefore, higher doses were required to achieve equivalent effects as observed in non-black patients (Kovarik et al., 2001).
The mitogen-activated protein kinase (MAPK) signaling is one of the most extensively investigated pathways that constitute Ras, Raf, mitogen-activated protein/extracellular signal-regulated kinase (MEK), and extracellular signal-regulated kinase (ERK-1/2; Morrison, 2012). MAPK signaling cascade is highly evolutionarily conserved signaling and partakes in a myriad of cellular functions including cell survival, proliferation, growth, and apoptosis (Braicu et al., 2019). MAPK pathway acts as a connecting link between extracellular signals and intracellular responses. Atypical genetic and epigenetic changes that lead to dysregulation of this pathway often result in the development of cancer (Braicu et al., 2019). Altered expression of MAPK family kinases has been reported in various cancers and many targeted therapies against these kinases have been tested in multiple human malignancies (Lee et al., 2020). Recently, a research study indicated MAPK/MEK1 mutations in AA colorectal cancer patients (Heath et al., 2018). Although there is no direct evidence of the racially disparate expression of the ERK-1/2 pathway in cancers, ERK-1/2 might be activated indirectly via different signaling cascades including MAPK/MEK pathway in AA patients. However, the development of resistance and redundancy in mTOR and ERK-1/2 pathways still present a major hurdle for precision medicine.
The interplay between mTOR and ERK-1/2 signaling pathways is a key determinant of cancer progression and metastasis (Mendoza et al., 2011). However, the exact underlying molecular mechanism connecting mTOR and ERK-1/2 signaling is not yet established. In this review article, we have summarized the current understanding of the mTOR pathway and its varied signaling networks. Until date, not much is known about the racially disparate expression of mTOR and ERK-1/2 pathways in cancers. We have comprehensively reviewed cancer-related racial disparities and the expression or activation status of the upstream regulator and downstream effector proteins that regulate the mTOR and ERK-1/2 pathway.
Here, we focus on current understanding of mTOR and MAPK pathways in cancer, with a particular emphasis on racially disparate regulation of these signaling pathways and their crosstalk with other key signaling molecules in different solid tumors. We highlight the mTOR pathway in response to intra- and extracellular signals that regulate energy homeostasis, cell growth, proliferation, and survival. Although mTOR targeting is clinically approved for treatment of renal cancers, overall survival for AA renal cancer patients compared to CA counterpart has not been improved. We will review the current approaches of targeting mTOR and ERK-1/2 signaling using pharmacological inhibitors and their therapeutic relevance in AA cancer patients.
Rapamycin was first extracted from the soil of Easter Island during an expedition. Later, the Ayerst Pharmaceuticals identified an antifungal macrolide from this soil, which was produced from Streptomyces hygroscopicus bacterium (Seto, 2012). This compound was named Rapamycin attributed to its place of origin, Rapa Nui (Easter Island). Further characterization of Rapamycin revealed its potential as an immunosuppressive and anti-tumor agent (Xie et al., 2016). However, its mechanism of action remained obscure for almost two decades until 1994 when biochemical analysis characterized the mTOR as the direct target of the rapamycin-FKBP12 complex in mammals and established a critical role of mTOR in cell signaling nexus (Kennedy and Lamming, 2016). mTOR plays a central role in regulating a variety of fundamental cell processes, including cell survival, proliferation, protein synthesis, and autophagy (Kim and Guan, 2019).
Genetic and metabolic changes influence hallmark characteristics of cancer that are indispensable for tumor initiation, growth, and metastasis. mTOR is a key determinant of cellular metabolism and signaling pathways (Figure 1). Cellular defects in energy homeostasis cause the activation of phosphatidylinositol 3-kinase (PI3K)/AKT/mTOR pathway (Fruman et al., 2017). mTOR is a Ser/Thr protein kinase and belongs to the PI3K-related kinase family and forms two distinct multi-protein complexes, mTOR complex 1 (mTORC1) and 2 (mTORC2; Zhou and Huang, 2011). mTORC1 consists of mTOR, mLST8, and Raptor is the most extensively characterized mTOR component (Saxton and Sabatini, 2017). In presence of specific growth factors, mTORC1 is activated and regulates cellular growth and proliferation through eukaryotic translation initiation factor 4E (eIF4E), ribosomal protein S6 kinases, and eIF4E-binding proteins (eIF4E-BP1, 2, and 3; Josse et al., 2016).
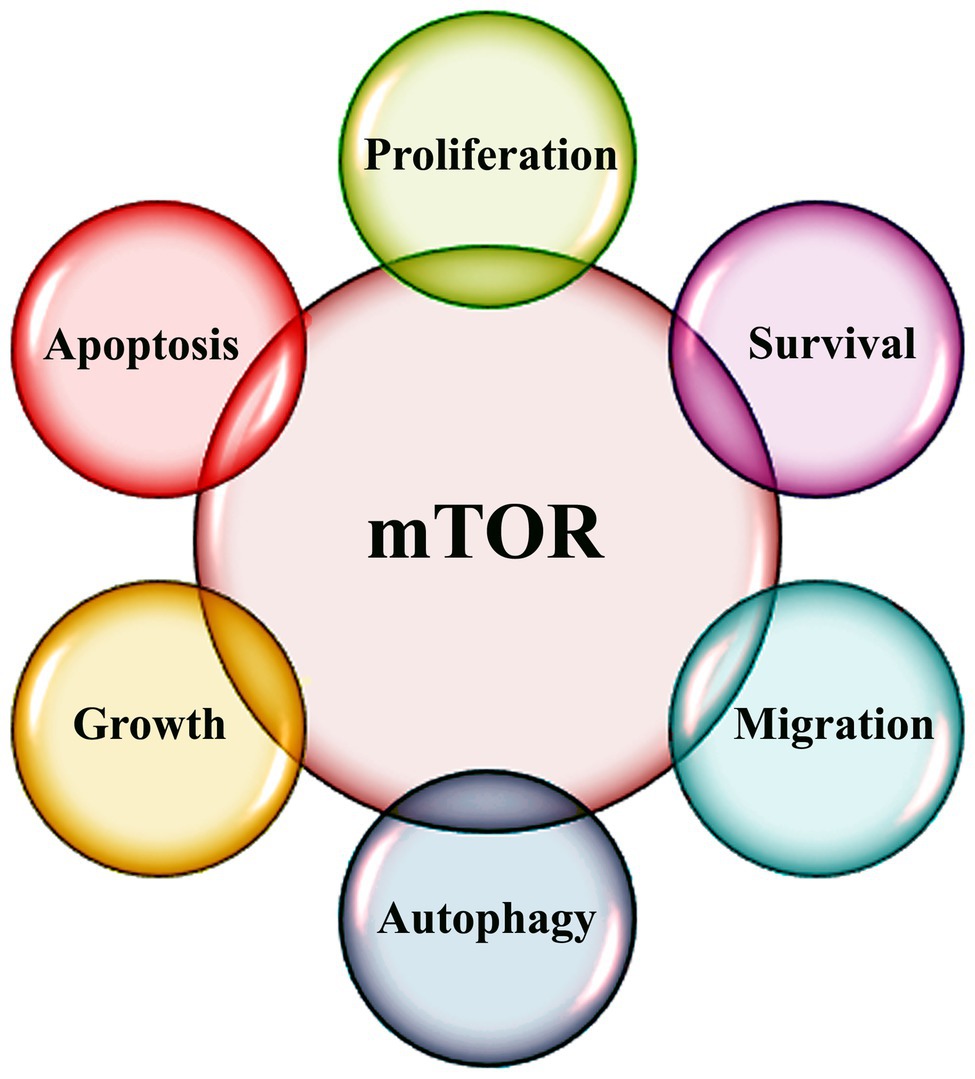
Figure 1. Schematic diagram highlighting various functions of mTOR signaling.
Besides, p53 family members have been reported to regulate mTOR signaling (Rosenbluth et al., 2008; Berkers et al., 2013). p53 and its family members; p63 and p73 are well-known tumor suppressor proteins that play a crucial role in preventing the oncogenic transformation of cells (Charan et al., 2020a). Also, p53, p63, and p73 ensure normal cellular fate by mediating a critical role in sensing different cellular cues and coordinating their downstream responses including cell senescence, cell-cycle arrest, differentiation, and cell death (Collavin et al., 2010; Cam et al., 2020). Moreover, an integrated network of p53 family members and mTOR pathway ensures homeostasis in a normal cell. However, the exact molecular mechanism of p53 family members mediated mTOR regulation is still not clear. mTOR pathway is often deregulated in different cancers (Kim et al., 2017). Dysregulation of the mTOR pathway disturbs the homeostasis of cells that may help in the metastasis of cancers.
Mitogen-activated protein kinases regulate key cellular processes and signaling pathways, including cell growth, survival, differentiation, and programmed cell death. To date, six different types of MAPKs have been identified in mammalian systems, which include extracellular signal-regulated kinase (ERK)1/2, ERK3/4, ERK5, ERK7/8, Jun N-terminal kinase (JNK)1/2/3, and the p38 isoforms α/β/Ɣ(ERK6)/ẟ (Schaeffer and Weber, 1999; Krens et al., 2006; Dhillon et al., 2007; Kyriakis and Avruch, 2012). ERK-1/2 was the first discovered MAPK that plays the main role in survival and proliferation signaling pathways (Wortzel and Seger, 2011). MAPK signaling pathways play a key role in regulating the responses to various extra and intracellular stress stimuli (Figure 2; Munshi and Ramesh, 2013).
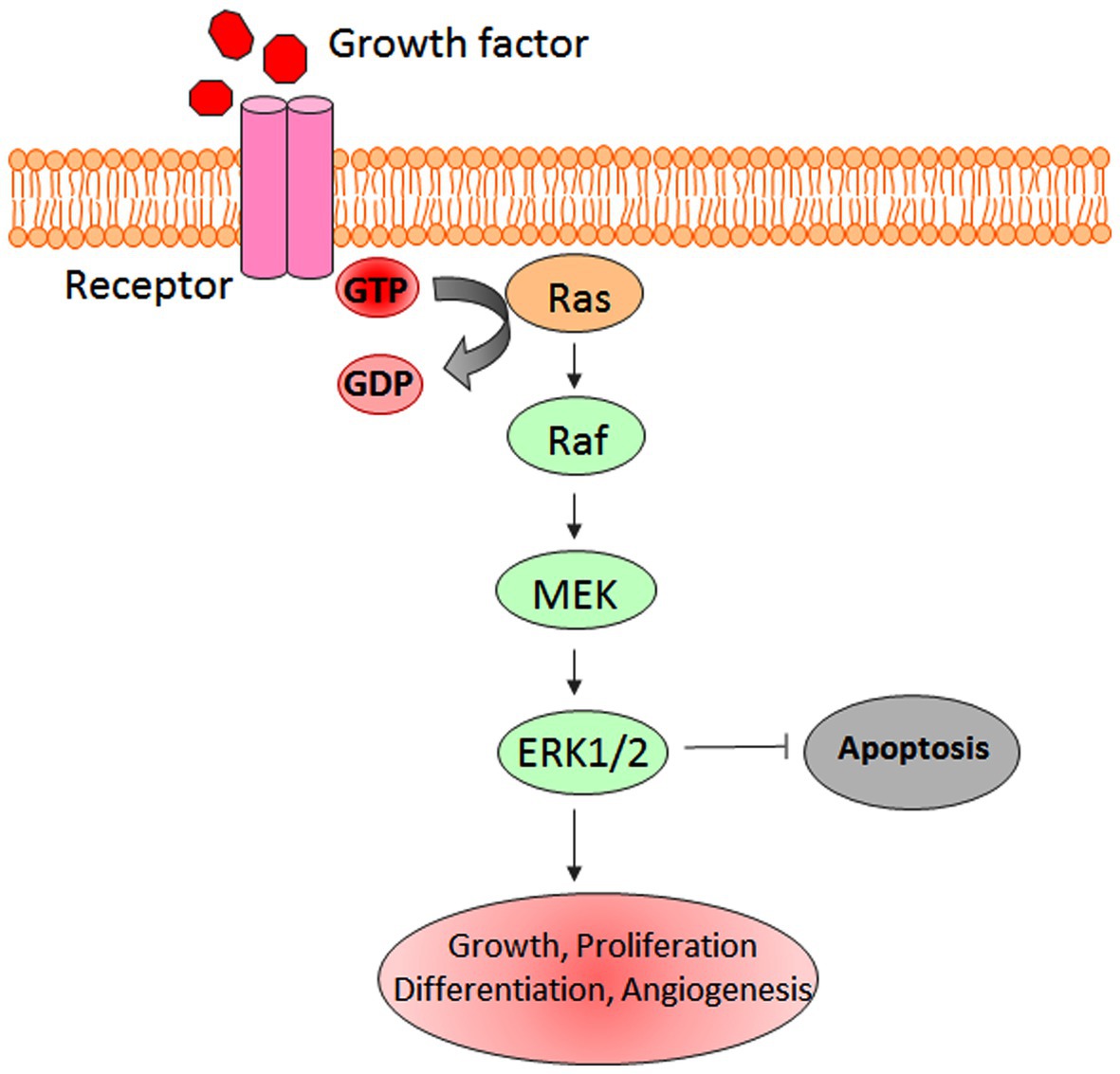
Figure 2. Schematic representation of extracellular signal-regulated kinase (ERK-1/2) pathway.
Anomalous activation of the ERK-1/2 pathway has been associated with tumorigenesis (Deschenes-Simard et al., 2014). Hyperactivation of ERK-1/2 signaling has been reported in several malignancies including neuroblastoma, renal, ovarian, breast, lung, and colorectal cancers (Steinmetz et al., 2004; Vicent et al., 2004; Bartholomeusz et al., 2012; Roseweir et al., 2018; Xie et al., 2019; Zhu et al., 2020). ERK-1/2 signaling cascade also promotes the invasive and metastatic abilities of cancer cells (Ward et al., 2001). Besides, ERK-1/2 plays a key role in enabling cancer cells with development of chemo- and radio-resistance and regulates the tumorigenic potential through the maintenance of cancer stem cells (Ciccarelli et al., 2016; Salaroglio et al., 2019).
Although mTOR and ERK-1/2 signaling pathways have been well-studied in several human cancers, their association with racial disparity has never been explored. AA population tends to have a higher number of gene mutations compared to the CA cohort (Chang et al., 2019). Genomic analyses revealed that AA breast cancer patients show 28,847 single-nucleotide polymorphisms (SNPs) in intronic regions of 61 genes that were associated with the mTOR pathway (Cheng et al., 2016). However, not much has been investigated on mTOR/ERK-1/2 genomic alterations in different human malignancies. Although a large number of signaling molecules are associated with mTOR/ERK-1/2 signaling pathways, very few studies have reported their differential expression in different races of cancer patients.
Recent reports have highlighted the disparate expression of transforming growth factor-beta (TGFβ) in different pathophysiological conditions among the AA population. TGFβ1 serves as a central molecule for various cellular signaling events including the pathways associated with mTOR and ERK-1/2. Racially disparate expression of TGFβ1 has already been reported and it is highly plausible that TGFβ1 might mediate its effects by modulating mTOR/ERK-1/2 pathways. TGFβ1 promotes epithelial to mesenchymal transition (EMT) through the activation of PI3K/Akt and mTOR signaling cascades, which further enhances distant metastasis (Lamouille and Derynck, 2007; Xu et al., 2009). The activation of the Akt and mTOR-signaling pathways leads to enhanced cancer cell growth (increased protein synthesis), and proliferation (increase in cell number; Lamouille and Derynck, 2007, 2011).
Interestingly, TGFβ is considered a double-edged sword in various human cancers including renal cancer (Akhurst and Derynck, 2001). Initially, TGFβ was reported to be a tumor suppressor, but later its novel oncogenic functions were discovered (Akhurst and Derynck, 2001). So far, there are no direct reports on the racially disparate expression or regulation of mTOR or ERK1/2 pathway. This manuscript is the first report connecting unappreciated essential molecules that are associated with mTOR and ERK-1/2 pathways that are involved in the regulation of various cellular processes including cell survival, differentiation, metabolism, and motility. TGFβ influences major cellular processes in cancer cells including proliferation, survival, epithelial-mesenchymal transition (EMT), and distant metastasis. TGFβ affects the regulation of the ERK-1/2 pathway. Treatment of TGFβ increases the phosphorylation of ERK-1/2, which is reported to further induce EMT in normal murine mammary gland epithelial cells and mouse cortical tubule epithelial cells (Xie et al., 2004).
Several studies have demonstrated TGFβ1 overexpression in AA patients with different disease conditions including renal disease, diabetes, hypertension, and breast cancer compared to CA counterparts (August et al., 2000; Suthanthiran et al., 2000; August et al., 2009; Huan et al., 2010; Quan et al., 2014). Importantly, AA population entails a SNP at codon 10 that is linked to higher levels of both mRNA and protein expression of TGF-β (Suthanthiran et al., 2000; Kim et al., 2004; Stuelten et al., 2005). Furthermore, an increased level of TGFβ1 expression was reported in AA prostate cancer patients compared to the CA counterparts (Elliott et al., 2018). Additionally, increased expression of TGFβ3 was detected in AA-derived prostate cancer cell lines compared to CA-derived cell lines (Elliott et al., 2018). AA prostate cancer patients also exhibited a higher level of TGFβ3 in blood sera compared to the CA cohort (Elliott et al., 2018). TGF-β-1, 2, and 3 are isoforms that interacts with each other and regulate a variety of cellular processes including cell differentiation, migration, invasion, and immune cell functions. However, there is no report on their disparate expression and redundant functions in cancer (De Caestecker et al., 2000; Jung et al., 2017).
Cancer cells often evade immune surveillance and immune clearance through a number of ways, one such approach is the expression of TGF-β. Considering its role in immunosuppression, TGF-β has been developed as a potential therapeutic target in onco-immunotherapy (Ungefroren, 2019). Moreover, TGF-β has been reported to increase the expression of an immune checkpoint molecule, programmed death ligand (PD-L1) in different cancers including lung cancer and pancreatic ductal adenocarcinoma (Hussain et al., 2021; Pan et al., 2021). TGF-β signaling in the tumor microenvironment is associated with immunologically cold tumors and contributes in the development of immune checkpoint resistance (Larson et al., 2020). Preclinical data using mouse models revealed that combination of immune checkpoint blockade with TGFβ inhibition improved the overall anti-tumor response (Ganesh and Massague, 2018). Further, TGF-β could induce immune suppression by promoting T cell-exclusion and Th1 effector functions in various cancers including colon and urothelial cancer (Tauriello et al., 2018; Bai et al., 2019). Taken together, the disparate high expression of TGFβ protein may be used as a personalized biomarker for AA prostate cancer patients for better diagnosis and prognosis of the disease.
Tumor suppressors like PTEN are associated with favorable prognosis and lower risks for cancer development (Maxwell et al., 2000; Charan et al., 2020b). PTEN negatively regulates PI3K and its downstream signaling, mediated through Akt and mTOR. Mutation in PTEN gene abolishes its protective functions and is often reported in various human cancers. The frequency of PTEN mutations in endometrial cancer patients was significantly higher in CA population than AA cohort (Maxwell et al., 2000). In addition, the rate of PTEN loss in prostate cancer patients was significantly higher in European American (EA) men than AA counterpart (Tosoian et al., 2017). KRAS is a family member of Ras genes. RAS proteins are GTPases that regulate survival, cell cycle, migration, cytoskeletal reorganization, and vesicular transport. KRAS is activated by epidermal growth factor receptor (EGFR) that further activates MEK/ERK1/2 and mTOR pathway (Tomasini et al., 2016). KRAS mutations in lung cancers are more common in AA patients than CA patients (Staudacher et al., 2017) that make mTOR a potential therapeutic target for KRAS-mutant lung cancer.
Recently, the differential expression of matrix metalloproteinases (MMP) family proteins, especially MMP2 and 9 has been studied in the different ethnic groups of cancer patients. Interestingly, the role of mTOR and ERK-1/2 signaling in regulating the expression of MMPs has been reported in different solid tumors (Chen et al., 2009; Akter et al., 2015; Xia et al., 2015; Park et al., 2016; Tsao et al., 2018). Besides, TGFβ that is a key mediator of mTOR/ERK-1/2 associated signaling cascades, has been shown to regulate MMP2 and MMP9 expression (Moore-Smith et al., 2017). The function of MMP2 and MMP9 are very well-characterized in cancer cell metastasis. Racially disparate expression of MMP2 and 9 has been reported in AA prostate cancer patients (Elliott et al., 2018; Ganaie et al., 2018). In addition, AA-derived prostate cancer cell lines showed higher expression of MMP2 and MMP9 both at transcriptome and proteome levels compared to CA counterparts (Elliott et al., 2018).
Several chemical inhibitors against different MMPs including MMP2 and 9 have been developed as potential antitumor agents (Tsao et al., 2018). mTOR and ERK-1/2 signaling pathways regulate metastasis of cancer cells by modulating MMPs (Park et al., 2016; Zou et al., 2020). Salmonella treatment inhibited AKT/mTOR signaling pathways, which further reduced the expression of MMP9 in the mouse tumor models (Tsao et al., 2018). Interestingly, the treatment of Chrysin (ERK-1/2 and JNK inhibitor) also reduced the expression of MMP9 in gastric cancer cells (Xia et al., 2015). Altogether, TGFβ and MMPs may act as key players in influencing mTOR and ERK-1/2 functions in cancer cells and their disparate expressions may predict different clinical outcomes.
AA-prostate cancer patients have also been shown to express higher levels of EGFR compared to CA prostate cancer patients (Shuch et al., 2004). EGFR is an upstream regulator of mTOR and ERK-1/2 signaling cascade and its overexpression and aberrant mutational status have been reported in several human cancers including renal cancer (Cossu-Rocca et al., 2016; Sooro et al., 2018). Furthermore, mTOR inhibitors reduced the proliferating potential of EGFR mutant drug-resistant lung cancer cells (Ishikawa et al., 2013). Moreover, ERK-1/2 signaling chemo-sensitizes drug-resistant lung cancer cells to WZ4002 (EGFR inhibitor). In addition, ERK-1/2 signaling attenuates the emergence of drug resistance in lung cancer cells. Another important regulator of the mTOR and ERK-1/2 signaling pathway is AKT, which is commonly altered among the CA cohort of breast cancer patients compared to AA counterparts (Khan et al., 2013). AKT promotes Ras/Raf/ERK-1/2 and mTOR signaling cascades, which in turn enhances the proliferation and survival of cancer cells (Chang et al., 2019).
Cross-talk between MAPK signaling pathways with other signaling pathways significantly impacts the clinical outcome of targeted anti-cancer therapies. ERK1/2 shares a number of proteins that interact with MST kinase, which is a key protein molecule of the Hippo signaling pathway. The Hippo signaling pathway primarily plays an important role in development. Dysregulation of this pathway can leads to the formation of tumor (Han, 2019). ERK1/2 and MST commonly mediate their effects through MYC, FOXO3, and TP53 proteins for regulating metabolism, growth, and cell death (Harvey et al., 2013). Higher genetic variations in the Hippo signaling pathway were reported in AA women than CA counterpart (Zhang et al., 2016). Hippo pathway alterations may also contributes in elevating breast cancer risks in AA women. In addition, Son of Sevenless 1 (SOS1) is an activator of Ras/MAPK, which is overexpressed in AA prostate cancer cells. SOS1 increased levels caused higher cancer cell proliferation and migration through increased activation of ERK signaling pathways (Timofeeva et al., 2009).
PI3K-AKT is one of the most important signaling pathways, which is differentially activated among different races and may modulate the mTOR pathway in several human cancers. AA renal cancer patients, especially, clear cell type-B (ccB), demonstrate higher activation of von Hippel-Lindau (VHL) and reduced expression of hypoxia-inducible factor (HIF) gene signature compared to CA patients (Krishnan et al., 2016). Such AA clear cell renal cancer patients show minimal response to VEGF-directed therapy as compared to CA counterparts. This could be one of the important reasons that contribute to the worst survival rate of AA patients receiving anti-VEGF therapy for clear cell renal carcinoma. In addition to VHL, mTOR and PI3KCA genes were altered in tumor samples of clear cell renal cell carcinoma (ccRCC) patients. The upregulation of the HIF pathway also leads to activation of tyrosine receptor kinases (RTKs), which initiates the signaling cascade of the Raf/MEK/ERK-1/2 pathway in renal carcinoma cells (Oka et al., 1995). In conclusion, the cross talk between HIF and mTOR/ERK-1/2 signaling pathways may regulate the aggressive properties of renal carcinoma in the AA population. In addition, higher activation of ERK in colorectal cancer and melanoma indicates poor responses to both neoadjuvant and adjuvant chemotherapy (Mirmohammadsadegh et al., 2007; Holck et al., 2019). Hence, ERK1/2 inhibition in combination with chemotherapy may turn out more effective anti-tumor therapeutic strategy. Overall, disparate expression of mTOR and ERK-1/2 associated signaling proteins can be selectively targeted along with mTOR or ERK-1/2 inhibitors to efficiently inhibit the tumor growth, metastasis, and to increase the overall survival depending upon the specific race.
In this review article, we attempted to shed light on disparate expression of unrecognized essential signaling molecules related to mTOR and ERK-1/2 pathway that play an important role in regulation of various cellular processes. AA groups of patients are at higher risk of developing highly invasive and aggressive cancers than CA cancer patients. mTOR and ERK-1/2 pathways have been the most extensively studied pathways in different cancers and their targeting has given promising results. mTOR and ERK-1/2 regulate cancer cells aggressiveness through modulating the multiple signaling pathways in human body. Furthermore, the differential expression of mTOR and ERK-1/2 associated signaling molecules may significantly affect the clinical outcome of certain AA cancer patients and could be used in precision medicine to improve the therapeutic response and overall survival. Moreover, this information will help in designing innovative treatment strategies against various aggressive cancers to improve the clinical outcome for different racial groups. Overall, this comprehensive study will unleash unappreciated regulators or pathways involved in various pathophysiological processes and could lead to the development of effective targeted therapy aimed at improving the quality of life of cancer patients from different ethnicity.
All authors contributed to writing and drafting the manuscript. SM, MC, BR, AV, DA, and RG conceived and reviewed the final version of the manuscript. All authors contributed to the article and approved the submitted version.
This manuscript was supported in part by NIH R01 grants CA109527 (RG) and CA231857 (BR and RG).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
AA, African American; CA, Caucasian American; SNP, Single-nucleotide polymorphism; mTOR, Mechanistic target of rapamycin; ERK-1/2, Extracellular-signal-regulated kinase; TGF-β, Transforming growth factor-β; EMT, Epithelial to mesenchymal transition; EGFR, Epidermal growth factor receptor; MMP, Matrix metallo proteinase; MAPK, Mitogen activated protein kinase.
Akhurst, R. J., and Derynck, R. (2001). TGF-beta signaling in cancer--a double-edged sword. Trends Cell Biol. 11, S44–S51. doi: 10.1016/s0962-8924(01)02130-4
Akter, H., Park, M., Kwon, O. S., Song, E. J., Park, W. S., and Kang, M. J. (2015). Activation of matrix metalloproteinase-9 (MMP-9) by neurotensin promotes cell invasion and migration through ERK pathway in gastric cancer. Tumour Biol. 36, 6053–6062. doi: 10.1007/s13277-015-3282-9
August, P., Leventhal, B., and Suthanthiran, M. (2000). Hypertension-induced organ damage in African Americans: transforming growth factor-beta(1) excess as a mechanism for increased prevalence. Curr. Hypertens. Rep. 2, 184–191. doi: 10.1007/s11906-000-0080-5
August, P., Sharma, V., Ding, R., Schwartz, J. E., and Suthanthiran, M. (2009). Transforming growth factor beta and excess burden of renal disease. Trans. Am. Clin. Climatol. Assoc. 120, 61–72.
Bai, X., Yi, M., Jiao, Y., Chu, Q., and Wu, K. (2019). Blocking TGF-beta signaling to enhance the efficacy of immune checkpoint inhibitor. Onco. Targets. Ther. 12, 9527–9538. doi: 10.2147/OTT.S224013
Bartholomeusz, C., Gonzalez-Angulo, A. M., Liu, P., Hayashi, N., Lluch, A., Ferrer-Lozano, J., et al. (2012). High ERK protein expression levels correlate with shorter survival in triple-negative breast cancer patients. Oncologist 17, 766–774. doi: 10.1634/theoncologist.2011-0377
Battelli, C., and Cho, D. C. (2011). mTOR inhibitors in renal cell carcinoma. Therapy 8, 359–367. doi: 10.2217/thy.11.32
Berkers, C. R., Maddocks, O. D., Cheung, E. C., Mor, I., and Vousden, K. H. (2013). Metabolic regulation by p53 family members. Cell Metab. 18, 617–633. doi: 10.1016/j.cmet.2013.06.019
Braicu, C., Buse, M., Busuioc, C., Drula, R., Gulei, D., Raduly, L., et al. (2019). A comprehensive review on MAPK: a promising therapeutic target in cancer. Cancers 11:1618. doi: 10.3390/cancers11101618
Cam, M., Charan, M., Welker, A. M., Dravid, P., Studebaker, A. W., Leonard, J. R., et al. (2020). DeltaNp73/ETS2 complex drives glioblastoma pathogenesis-targeting downstream mediators by rebastinib prolongs survival in preclinical models of glioblastoma. Neuro-Oncology 22, 345–356. doi: 10.1093/neuonc/noz190
Chang, C. S., Kitamura, E., Johnson, J., Bollag, R., and Hawthorn, L. (2019). Genomic analysis of racial differences in triple negative breast cancer. Genomics 111, 1529–1542. doi: 10.1016/j.ygeno.2018.10.010
Charan, M., Dravid, P., Cam, M., Audino, A., Gross, A. C., Arnold, M. A., et al. (2020a). GD2-directed CAR-T cells in combination with HGF-targeted neutralizing antibody (AMG102) prevent primary tumor growth and metastasis in Ewing sarcoma. Int. J. Cancer 146, 3184–3195. doi: 10.1002/ijc.32743
Charan, M., Verma, A. K., Hussain, S., Misri, S., Mishra, S., Majumder, S., et al. (2020b). Molecular and cellular factors associated with racial disparity in breast cancer. Int. J. Mol. Sci. 21:5936. doi: 10.3390/ijms21165936
Chen, J. S., Wang, Q., Fu, X. H., Huang, X. H., Chen, X. L., Cao, L. Q., et al. (2009). Involvement of PI3K/PTEN/AKT/mTOR pathway in invasion and metastasis in hepatocellular carcinoma: association with MMP-9. Hepatol. Res. 39, 177–186. doi: 10.1111/j.1872-034X.2008.00449.x
Cheng, T. D., Ambrosone, C. B., Hong, C. C., Lunetta, K. L., Liu, S., Hu, Q., et al. (2016). Genetic variants in the mTOR pathway and breast cancer risk in African American women. Carcinogenesis 37, 49–55. doi: 10.1093/carcin/bgv160
Ciccarelli, C., Vulcano, F., Milazzo, L., Gravina, G. L., Marampon, F., Macioce, G., et al. (2016). Key role of MEK/ERK pathway in sustaining tumorigenicity and in vitro radioresistance of embryonal rhabdomyosarcoma stem-like cell population. Mol. Cancer 15:16. doi: 10.1186/s12943-016-0501-y
Collavin, L., Lunardi, A., and Del Sal, G. (2010). p53-family proteins and their regulators: hubs and spokes in tumor suppression. Cell Death Differ. 17, 901–911. doi: 10.1038/cdd.2010.35
Cossu-Rocca, P., Muroni, M. R., Sanges, F., Sotgiu, G., Asunis, A., Tanca, L., et al. (2016). EGFR kinase-dependent and kinase-independent roles in clear cell renal cell carcinoma. Am. J. Cancer Res. 6, 71–83.
De Caestecker, M. P., Piek, E., and Roberts, A. B. (2000). Role of transforming growth factor-beta signaling in cancer. J. Natl. Cancer Inst. 92, 1388–1402. doi: 10.1093/jnci/92.17.1388
Deschenes-Simard, X., Kottakis, F., Meloche, S., and Ferbeyre, G. (2014). ERKs in cancer: friends or foes? Cancer Res. 74, 412–419. doi: 10.1158/0008-5472.CAN-13-2381
Dhillon, A. S., Hagan, S., Rath, O., and Kolch, W. (2007). MAP kinase signalling pathways in cancer. Oncogene 26, 3279–3290. doi: 10.1038/sj.onc.1210421
Elliott, B., Zackery, D. L., Eaton, V. A., Jones, R. T., Abebe, F., Ragin, C. C., et al. (2018). Ethnic differences in TGFbeta-signaling pathway may contribute to prostate cancer health disparity. Carcinogenesis 39, 546–555. doi: 10.1093/carcin/bgy020
Esnaola, N. F., and Ford, M. E. (2012). Racial differences and disparities in cancer care and outcomes: where’s the rub? Surg. Oncol. Clin. N. Am. 21, 417–437. doi: 10.1016/j.soc.2012.03.012
Freedman, J. A., Al Abo, M., Allen, T. A., Piwarski, S. A., Wegermann, K., and Patierno, S. R. (2021). Biological aspects of cancer health disparities. Annu. Rev. Med. 72, 229–241. doi: 10.1146/annurev-med-070119-120305
Fruman, D. A., Chiu, H., Hopkins, B. D., Bagrodia, S., Cantley, L. C., and Abraham, R. T. (2017). The PI3K pathway in human disease. Cell 170, 605–635. doi: 10.1016/j.cell.2017.07.029
Ganaie, A. A., Beigh, F. H., Astone, M., Ferrari, M. G., Maqbool, R., Umbreen, S., et al. (2018). BMI1 drives metastasis of prostate cancer in Caucasian and African-American men and is a potential therapeutic target: hypothesis tested in race-specific models. Clin. Cancer Res. 24, 6421–6432. doi: 10.1158/1078-0432.CCR-18-1394
Ganesh, K., and Massague, J. (2018). TGF-beta inhibition and immunotherapy: checkmate. Immunity 48, 626–628. doi: 10.1016/j.immuni.2018.03.037
Gross, C. P., Smith, B. D., Wolf, E., and Andersen, M. (2008). Racial disparities in cancer therapy: did the gap narrow between 1992 and 2002? Cancer 112, 900–908. doi: 10.1002/cncr.23228
Han, Y. (2019). Analysis of the role of the hippo pathway in cancer. J. Transl. Med. 17:116. doi: 10.1186/s12967-019-1869-4
Harvey, K. F., Zhang, X., and Thomas, D. M. (2013). The hippo pathway and human cancer. Nat. Rev. Cancer 13, 246–257. doi: 10.1038/nrc3458
Heath, E. I., Lynce, F., Xiu, J., Ellerbrock, A., Reddy, S. K., Obeid, E., et al. (2018). Racial disparities in the molecular landscape of cancer. Anticancer Res. 38, 2235–2240. doi: 10.21873/anticanres.12466
Holck, S., Klarskov, L. L., and Larsson, L. I. (2019). Phospho-ERK levels as predictors for chemotherapy of rectal carcinoma. Oncotarget 10, 1745–1755. doi: 10.18632/oncotarget.26741
Huan, Y., Deloach, S., Daskalakis, C., Dunn, S. R., Sharma, K., and Falkner, B. (2010). Regulation of transforming growth factor-beta1 by insulin in prediabetic African Americans. Kidney Int. 78, 318–324. doi: 10.1038/ki.2010.109
Hussain, S. M., Kansal, R. G., Alvarez, M. A., Hollingsworth, T. J., Elahi, A., Miranda-Carboni, G., et al. (2021). Role of TGF-beta in pancreatic ductal adenocarcinoma progression and PD-L1 expression. Cell. Oncol. doi: 10.1007/s13402-021-00594-0 [Epub ahead of print]
Ishikawa, D., Takeuchi, S., Nakagawa, T., Sano, T., Nakade, J., Nanjo, S., et al. (2013). mTOR inhibitors control the growth of EGFR mutant lung cancer even after acquiring resistance by HGF. PLoS One 8:e62104. doi: 10.1371/journal.pone.0062104
Josse, L., Xie, J., Proud, C. G., and Smales, C. M. (2016). mTORC1 signalling and eIF4E/4E-BP1 translation initiation factor stoichiometry influence recombinant protein productivity from GS-CHOK1 cells. Biochem. J. 473, 4651–4664. doi: 10.1042/BCJ20160845
Jung, B., Staudacher, J. J., and Beauchamp, D. (2017). Transforming growth factor beta superfamily signaling in development of colorectal cancer. Gastroenterology 152, 36–52. doi: 10.1053/j.gastro.2016.10.015
Kennedy, B. K., and Lamming, D. W. (2016). The mechanistic target of Rapamycin: the grand ConducTOR of metabolism and aging. Cell Metab. 23, 990–1003. doi: 10.1016/j.cmet.2016.05.009
Khan, K. H., Yap, T. A., Yan, L., and Cunningham, D. (2013). Targeting the PI3K-AKT-mTOR signaling network in cancer. Chin. J. Cancer 32, 253–265. doi: 10.5732/cjc.013.10057
Kim, L. C., Cook, R. S., and Chen, J. (2017). mTORC1 and mTORC2 in cancer and the tumor microenvironment. Oncogene 36, 2191–2201. doi: 10.1038/onc.2016.363
Kim, J., and Guan, K. L. (2019). mTOR as a central hub of nutrient signalling and cell growth. Nat. Cell Biol. 21, 63–71. doi: 10.1038/s41556-018-0205-1
Kim, E. S., Kim, M. S., and Moon, A. (2004). TGF-beta-induced upregulation of MMP-2 and MMP-9 depends on p38 MAPK, but not ERK signaling in MCF10A human breast epithelial cells. Int. J. Oncol. 25, 1375–1382.
Kovarik, J. M., Hsu, C. H., Mcmahon, L., Berthier, S., and Rordorf, C. (2001). Population pharmacokinetics of everolimus in de novo renal transplant patients: impact of ethnicity and comedications. Clin. Pharmacol. Ther. 70, 247–254. doi: 10.1067/mcp.2001.118022
Krens, S. F., Spaink, H. P., and Snaar-Jagalska, B. E. (2006). Functions of the MAPK family in vertebrate-development. FEBS Lett. 580, 4984–4990. doi: 10.1016/j.febslet.2006.08.025
Krishnan, B., Rose, T. L., Kardos, J., Milowsky, M. I., and Kim, W. Y. (2016). Intrinsic genomic differences between African American and white patients with clear cell renal cell carcinoma. JAMA Oncol. 2, 664–667. doi: 10.1001/jamaoncol.2016.0005
Kyriakis, J. M., and Avruch, J. (2012). Mammalian MAPK signal transduction pathways activated by stress and inflammation: a 10-year update. Physiol. Rev. 92, 689–737. doi: 10.1152/physrev.00028.2011
Lamouille, S., and Derynck, R. (2007). Cell size and invasion in TGF-beta-induced epithelial to mesenchymal transition is regulated by activation of the mTOR pathway. J. Cell Biol. 178, 437–451. doi: 10.1083/jcb.200611146
Lamouille, S., and Derynck, R. (2011). Emergence of the phosphoinositide 3-kinase-Akt-mammalian target of rapamycin axis in transforming growth factor-beta-induced epithelial-mesenchymal transition. Cells Tissues Organs 193, 8–22. doi: 10.1159/000320172
Larson, C., Oronsky, B., Carter, C. A., Oronsky, A., Knox, S. J., Sher, D., et al. (2020). TGF-beta: a master immune regulator. Expert Opin. Ther. Targets 24, 427–438. doi: 10.1080/14728222.2020.1744568
Lee, S., Rauch, J., and Kolch, W. (2020). Targeting MAPK Signaling in cancer: mechanisms of drug resistance and sensitivity. Int. J. Mol. Sci. 21:1102. doi: 10.3390/ijms21031102
Maxwell, G. L., Risinger, J. I., Hayes, K. A., Alvarez, A. A., Dodge, R. K., Barrett, J. C., et al. (2000). Racial disparity in the frequency of PTEN mutations, but not microsatellite instability, in advanced endometrial cancers. Clin. Cancer Res. 6, 2999–3005.
Mendoza, M. C., Er, E. E., and Blenis, J. (2011). The Ras-ERK and PI3K-mTOR pathways: cross-talk and compensation. Trends Biochem. Sci. 36, 320–328. doi: 10.1016/j.tibs.2011.03.006
Mirmohammadsadegh, A., Mota, R., Gustrau, A., Hassan, M., Nambiar, S., Marini, A., et al. (2007). ERK1/2 is highly phosphorylated in melanoma metastases and protects melanoma cells from cisplatin-mediated apoptosis. J. Invest. Dermatol. 127, 2207–2215. doi: 10.1038/sj.jid.5700870
Moore-Smith, L. D., Isayeva, T., Lee, J. H., Frost, A., and Ponnazhagan, S. (2017). Silencing of TGF-beta1 in tumor cells impacts MMP-9 in tumor microenvironment. Sci. Rep. 7:8678. doi: 10.1038/s41598-017-09062-y
Morrison, D. K. (2012). MAP kinase pathways. Cold Spring Harb. Perspect. Biol. 4:a011254. doi: 10.1101/cshperspect.a011254
Munshi, A., and Ramesh, R. (2013). Mitogen-activated protein kinases and their role in radiation response. Genes Cancer 4, 401–408. doi: 10.1177/1947601913485414
Oka, H., Chatani, Y., Hoshino, R., Ogawa, O., Kakehi, Y., Terachi, T., et al. (1995). Constitutive activation of mitogen-activated protein (MAP) kinases in human renal cell carcinoma. Cancer Res. 55, 4182–4187.
Pan, L. N., Ma, Y. F., Li, Z., Hu, J. A., and Xu, Z. H. (2021). KRAS G12V mutation upregulates PD-L1 expression via TGF-beta/EMT signaling pathway in human non-small-cell lung cancer. Cell Biol. Int. 45, 795–803. doi: 10.1002/cbin.11524
Park, J. H., Cho, Y. Y., Yoon, S. W., and Park, B. (2016). Suppression of MMP-9 and FAK expression by pomolic acid via blocking of NF-kappaB/ERK/mTOR signaling pathways in growth factor-stimulated human breast cancer cells. Int. J. Oncol. 49, 1230–1240. doi: 10.3892/ijo.2016.3585
Quan, L., Gong, Z., Yao, S., Bandera, E. V., Zirpoli, G., Hwang, H., et al. (2014). Cytokine and cytokine receptor genes of the adaptive immune response are differentially associated with breast cancer risk in American women of African and European ancestry. Int. J. Cancer 134, 1408–1421. doi: 10.1002/ijc.28458
Rosenbluth, J. M., Mays, D. J., Pino, M. F., Tang, L. J., and Pietenpol, J. A. (2008). A gene signature-based approach identifies mTOR as a regulator of p73. Mol. Cell. Biol. 28, 5951–5964. doi: 10.1128/MCB.00305-08
Roseweir, A. K., Halcrow, E. S., Chichilo, S., Powell, A. G., Mcmillan, D. C., Horgan, P. G., et al. (2018). ERK and p38MAPK combine to improve survival in patients with BRAF mutant colorectal cancer. Br. J. Cancer 119, 323–329. doi: 10.1038/s41416-018-0174-y
Salaroglio, I. C., Mungo, E., Gazzano, E., Kopecka, J., and Riganti, C. (2019). ERK is a pivotal player of chemo-immune-resistance in cancer. Int. J. Mol. Sci. 20:2505. doi: 10.3390/ijms20102505
Saxton, R. A., and Sabatini, D. M. (2017). mTOR signaling in growth, metabolism, and disease. Cell 168, 960–976. doi: 10.1016/j.cell.2017.02.004
Schaeffer, H. J., and Weber, M. J. (1999). Mitogen-activated protein kinases: specific messages from ubiquitous messengers. Mol. Cell. Biol. 19, 2435–2444. doi: 10.1128/MCB.19.4.2435
Seto, B. (2012). Rapamycin and mTOR: a serendipitous discovery and implications for breast cancer. Clin. Transl. Med. 1:29. doi: 10.1186/2001-1326-1-29
Shuch, B., Mikhail, M., Satagopan, J., Lee, P., Yee, H., Chang, C., et al. (2004). Racial disparity of epidermal growth factor receptor expression in prostate cancer. J. Clin. Oncol. 22, 4725–4729. doi: 10.1200/JCO.2004.06.134
Soliman, G. A. (2013). The role of mechanistic target of rapamycin (mTOR) complexes signaling in the immune responses. Nutrients 5, 2231–2257. doi: 10.3390/nu5062231
Sooro, M. A., Zhang, N., and Zhang, P. (2018). Targeting EGFR-mediated autophagy as a potential strategy for cancer therapy. Int. J. Cancer 143, 2116–2125. doi: 10.1002/ijc.31398
Staudacher, J. J., Yazici, C., Bul, V., Zeidan, J., Khalid, A., Xia, Y., et al. (2017). Increased frequency of KRAS mutations in African Americans compared with Caucasians in sporadic colorectal cancer. Clin. Transl. Gastroenterol. 8:e124. doi: 10.1038/ctg.2017.48
Steinmetz, R., Wagoner, H. A., Zeng, P., Hammond, J. R., Hannon, T. S., Meyers, J. L., et al. (2004). Mechanisms regulating the constitutive activation of the extracellular signal-regulated kinase (ERK) signaling pathway in ovarian cancer and the effect of ribonucleic acid interference for ERK1/2 on cancer cell proliferation. Mol. Endocrinol. 18, 2570–2582. doi: 10.1210/me.2004-0082
Stuelten, C. H., Dacosta Byfield, S., Arany, P. R., Karpova, T. S., Stetler-Stevenson, W. G., and Roberts, A. B. (2005). Breast cancer cells induce stromal fibroblasts to express MMP-9 via secretion of TNF-alpha and TGF-beta. J. Cell Sci. 118, 2143–2153. doi: 10.1242/jcs.02334
Suthanthiran, M., Li, B., Song, J. O., Ding, R., Sharma, V. K., Schwartz, J. E., et al. (2000). Transforming growth factor-beta 1 hyperexpression in African-American hypertensives: a novel mediator of hypertension and/or target organ damage. Proc. Natl. Acad. Sci. U. S. A. 97, 3479–3484. doi: 10.1073/pnas.050420897
Tauriello, D. V. F., Palomo-Ponce, S., Stork, D., Berenguer-Llergo, A., Badia-Ramentol, J., Iglesias, M., et al. (2018). TGFbeta drives immune evasion in genetically reconstituted colon cancer metastasis. Nature 554, 538–543. doi: 10.1038/nature25492
Timofeeva, O. A., Zhang, X., Ressom, H. W., Varghese, R. S., Kallakury, B. V., Wang, K., et al. (2009). Enhanced expression of SOS1 is detected in prostate cancer epithelial cells from African-American men. Int. J. Oncol. 35, 751–760. doi: 10.3892/ijo_00000388
Tomasini, P., Walia, P., Labbe, C., Jao, K., and Leighl, N. B. (2016). Targeting the KRAS pathway in non-small cell lung cancer. Oncologist 21, 1450–1460. doi: 10.1634/theoncologist.2015-0084
Tosoian, J. J., Almutairi, F., Morais, C. L., Glavaris, S., Hicks, J., Sundi, D., et al. (2017). Prevalence and prognostic significance of PTEN loss in African-American and European-American men undergoing radical prostatectomy. Eur. Urol. 71, 697–700. doi: 10.1016/j.eururo.2016.07.026
Tsao, Y. T., Kuo, C. Y., Cheng, S. P., and Lee, C. H. (2018). Downregulations of AKT/mTOR signaling pathway for Salmonella-mediated suppression of matrix Metalloproteinases-9 expression in mouse tumor models. Int. J. Mol. Sci. 19:1630. doi: 10.3390/ijms19061630
Ungefroren, H. (2019). Blockade of TGF-beta signaling: a potential target for cancer immunotherapy? Expert Opin. Ther. Targets 23, 679–693. doi: 10.1080/14728222.2019.1636034
Vicent, S., Lopez-Picazo, J. M., Toledo, G., Lozano, M. D., Torre, W., Garcia-Corchon, C., et al. (2004). ERK1/2 is activated in non-small-cell lung cancer and associated with advanced tumours. Br. J. Cancer 90, 1047–1052. doi: 10.1038/sj.bjc.6601644
Voss, M. H., Molina, A. M., and Motzer, R. J. (2011). mTOR inhibitors in advanced renal cell carcinoma. Hematol. Oncol. Clin. North Am. 25, 835–852. doi: 10.1016/j.hoc.2011.04.008
Ward, Y., Wang, W., Woodhouse, E., Linnoila, I., Liotta, L., and Kelly, K. (2001). Signal pathways which promote invasion and metastasis: critical and distinct contributions of extracellular signal-regulated kinase and Ral-specific guanine exchange factor pathways. Mol. Cell. Biol. 21, 5958–5969. doi: 10.1128/MCB.21.17.5958-5969.2001
Wortzel, I., and Seger, R. (2011). The ERK cascade: distinct functions within various subcellular organelles. Genes Cancer 2, 195–209. doi: 10.1177/1947601911407328
Xia, Y., Lian, S., Khoi, P. N., Yoon, H. J., Joo, Y. E., Chay, K. O., et al. (2015). Chrysin inhibits tumor promoter-induced MMP-9 expression by blocking AP-1 via suppression of ERK and JNK pathways in gastric cancer cells. PLoS One 10:e0124007. doi: 10.1371/journal.pone.0124007
Xie, L., Law, B. K., Chytil, A. M., Brown, K. A., Aakre, M. E., and Moses, H. L. (2004). Activation of the Erk pathway is required for TGF-beta1-induced EMT in vitro. Neoplasia 6, 603–610. doi: 10.1593/neo.04241
Xie, J., Wang, X., and Proud, C. G. (2016). mTOR inhibitors in cancer therapy. F1000Res 5:F1000. doi: 10.12688/f1000research.9207.1
Xie, G., Wu, H., Cai, W., Chen, M., Huang, W., Yan, W., et al. (2019). RDM1 promotes neuroblastoma growth through the RAS-Raf-MEK-ERK pathway. FEBS Open Bio 9, 490–497. doi: 10.1002/2211-5463.12586
Xu, J., Lamouille, S., and Derynck, R. (2009). TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 19, 156–172. doi: 10.1038/cr.2009.5
Zavala, V. A., Bracci, P. M., Carethers, J. M., Carvajal-Carmona, L., Coggins, N. B., Cruz-Correa, M. R., et al. (2021). Cancer health disparities in racial/ethnic minorities in the United States. Br. J. Cancer 124, 315–332. doi: 10.1038/s41416-020-01038-6
Zhang, J., Yao, S., Hu, Q., Zhu, Q., Liu, S., Lunetta, K. L., et al. (2016). Genetic variations in the hippo signaling pathway and breast cancer risk in African American women in the AMBER consortium. Carcinogenesis 37, 951–956. doi: 10.1093/carcin/bgw077
Zhou, H., and Huang, S. (2011). Role of mTOR signaling in tumor cell motility, invasion and metastasis. Curr. Protein Pept. Sci. 12, 30–42. doi: 10.2174/138920311795659407
Zhu, L., Ding, R., Yan, H., Zhang, J., and Lin, Z. (2020). ZHX2 drives cell growth and migration via activating MEK/ERK signal and induces sunitinib resistance by regulating the autophagy in clear cell renal cell carcinoma. Cell Death Dis. 11:337. doi: 10.1038/s41419-020-2541-x
Keywords: racial disparity, cancer, mTOR, ERK, MAPK
Citation: Mishra S, Charan M, Verma AK, Ramaswamy B, Ahirwar DK and Ganju RK (2021) Racially Disparate Expression of mTOR/ERK-1/2 Allied Proteins in Cancer. Front. Cell Dev. Biol. 9:601929. doi: 10.3389/fcell.2021.601929
Edited by:
Gianluca Aguiari, University of Ferrara, ItalyReviewed by:
Christian Gomez, University of Mississippi Medical Center, United StatesCopyright © 2021 Mishra, Charan, Verma, Ramaswamy, Ahirwar and Ganju. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ramesh K. Ganju, cmFtZXNoLmdhbmp1QG9zdW1jLmVkdQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.