 Maria M. Haykal
Maria M. Haykal Clara Nahmias
Clara Nahmias Christine Varon
Christine Varon Océane C. B. Martin
Océane C. B. Martin- 1Université Paris-Saclay, Institut Gustave Roussy, Inserm U981, Biomarqueurs Prédictifs et Nouvelles Stratégies Thérapeutiques en Oncologie, Villejuif, France
- 2Univ. Bordeaux, INSERM, BaRITOn, U1053, Bordeaux, France
Cancer is a complex disease and it is now clear that not only epithelial tumor cells play a role in carcinogenesis. The tumor microenvironment is composed of non-stromal cells, including endothelial cells, adipocytes, immune and nerve cells, and a stromal compartment composed of extracellular matrix, cancer-associated fibroblasts and mesenchymal cells. Tumorigenesis is a dynamic process with constant interactions occurring between the tumor cells and their surroundings. Even though all connections have not yet been discovered, it is now known that crosstalk between actors of the microenvironment drives cancer progression. Taking into account this complexity, it is important to develop relevant models to study carcinogenesis. Conventional 2D culture models fail to represent the entire tumor microenvironment properly and the use of animal models should be decreased with respect to the 3Rs rule. To this aim, in vitro organotypic models have been significantly developed these past few years. These models have different levels of complexity and allow the study of tumor cells alone or in interaction with the microenvironment actors during the multiple stages of carcinogenesis. This review depicts recent insights into organotypic modeling of the tumor and its microenvironment all throughout cancer progression. It offers an overview of the crosstalk between epithelial cancer cells and their microenvironment during the different phases of carcinogenesis, from the early cell autonomous events to the late metastatic stages. The advantages of 3D over classical 2D or in vivo models are presented as well as the most promising organotypic models. A particular focus is made on organotypic models used for studying cancer progression, from the less complex spheroids to the more sophisticated body-on-a-chip. Last but not least, we address the potential benefits of these models in personalized medicine which is undoubtedly a domain paving the path to new hopes in terms of cancer care and cure.
Introduction
Carcinogenesis is a complex multistep process, often described as somatic evolution. Typically, cancer progression involves the accumulation of genetic and/or epigenetic somatic modifications and exposition to environmental factors. Indeed, the development of many tumors is tightly linked with genotoxicity, chronic infections, dietary habits, or autoimmunity; which are all underlined by inflammation. Early on, Fearon and Vogelstein (1990) described a sequence of defined genetic events driving the formation of colorectal cancers. Afterward, the seminal works of Hanahan and Weinberg (2000, 2011) contributed to shift cancer research from a reductionist point of view with a sole focus on the cancer cell itself to a more comprehensive view involving cues from the neighboring niche. Therefore, carcinogenesis is the fruit of the interplay between multiple cell autonomous and non-autonomous processes, defined as “Hallmarks of cancer,” that include genomic instability, proliferative abnormality, stromal reprogramming, angiogenesis, immune suppression and tumor sustaining inflammation. In the following sections, we first define the tumor microenvironment (TME) and briefly depict its different components. We also summarize the recently described interactions between the TME actors and the tumor cells in the cancer progression cascade. In depth understanding of such interactions renders necessary the study of tumor cells within their microenvironment, as this is crucial for cancer progression. In this line of thought, we describe the most promising organotypic models used for modeling cancer progression stages from the initial tumor and its microenvironment to dissemination and metastasis.
Part I—Role of the Microenvironment in Tumoral Progression
The importance of the tumor microenvironment is embodied in the concept that cancer cells do not cause the disease alone, but rather corrupt recruited and neighboring normal cell types to serve as accessories to the crime (Hanahan and Coussens, 2012). In particular, interactions between cancer cells and their microenvironment represent a powerful relationship that influences disease initiation and progression and patient prognosis. For decades, the focus of cancer research has been almost exclusively on epithelial tumor cells. However, in the past few years, there has been a major shift toward the study of the TME, elucidating that tumor progression is dependent on an intricate network of interactions among cancer cells and their surroundings (McAllister and Weinberg, 2014; Taniguchi and Karin, 2018; Hinshaw and Shevde, 2019).
Tumors are unquestionably heterogenous entities, composed of phenotypically distinct cellular populations with different functions. This is illustrated by the clonal evolution theory (Nowell, 1976), TME heterogeneity (Junttila and de Sauvage, 2013) and hierarchal organization of cancer cell subpopulations that includes cancer stem cells (CSCs) and their progenies. Some studies have shown that CSCs are the driving force of tumor formation as they exhibit self-renewal and tumor−initiating capacities and phenotypic plasticity. Plasticity offers cancer cells the ability to switch from a differentiated state to an undifferentiated CSC-like state, responsible for long term tumor growth and drug resistance. Recently, observations of anatomically distinct niches of CSCs within tumors have emerged (reviewed in Plaks et al., 2015; Batlle and Clevers, 2017). These niches could have a role in preserving the plastic phenotype of CSCs and their protection from the immune system. Nonetheless, the heterogeneous tumor is a part of a larger society comprising many other actors that define the tumor microenvironment.
Defining the Tumor Microenvironment
The tumor microenvironment, a diversified compartment of differentiated and progenitor cells, comprises all the non-malignant host cellular and non-cellular components of the tumor niche including, but not restricted to, endothelial cells, adipocytes, cells of the immune and nervous systems, and the stroma.
Non-stromal Components
Endothelial Cells
The most well-known extrinsic modulator of cancer cell growth is neovascularization (Folkman, 1985). Early studies using mouse models show that the angiogenic switch increases the proliferation rate of cancer cells (Folkman et al., 1989). Angiogenesis is crucial to the ability of tumors to thrive and the vascular endothelium is an active participant in the formation of a growth-permissive tumor microenvironment. Vascularization is driven by the hypoxic center of the tumor where hyperproliferation results in increased oxygen demand. Consequently, low oxygen induces the expression of angiogenic proteins like vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF) (Papetti and Herman, 2002) that activate endothelial cells and attract them toward the tumor to form new vessels, allowing the delivery of nutrients and oxygen. Without angiogenesis, tumors are condemned to quiescence and cell death. Tumor vascularization requires the cooperation of different TME cells, mainly vascular endothelial cells that provide structural integrity to the newly formed vessels and pericytes that ensure their coverage and maturity (Weis and Cheresh, 2011). Endothelial cells also constitute routes to metastatic dissemination via angiogenesis and contribute to resistance to chemotherapies through an overexpression of drug efflux pumps thereby decreasing the tumor’s access to the drug (Hida et al., 2013).
Adipocytes
Cancer-associated adipocytes (CAAs) support cancer growth mainly through secretion of adipokines like adipsin (Goto et al., 2019), chemerin (Lu et al., 2019) as well as proinflammatory cytokines (Dirat et al., 2011) and growth factors. CAAs also supply lipids for cancer cell membranes and organelles, induce metabolic reprogramming in cancer cells and provide proteases for cancer cell invasion (reviewed in Deng et al., 2016). Moreover, through the production of tumor-promoting cytokines and factors, they have been shown to confer resistance to hormone therapies, chemotherapies, radiotherapies and targeted therapies in breast cancer (Choi et al., 2018), and to contribute to tumor progression across a variety of obesity-associated cancers (Park et al., 2014) such as esophagus, gastric, liver, kidney, colorectal, pancreatic, breast, ovarian, prostate, and thyroid cancers. Adipocytes from white adipose tissue are recruited to tumors, can differentiate into pericytes and incorporate into vessel walls contributing to angiogenesis and to tumor proliferation (Zhang et al., 2012).
Infiltrating Immune Cells
Variations in immune profiles are linked to prognosis and therapeutic responses (Gentles et al., 2015). All adult solid tumors contain infiltrates of diverse immune cell subsets that influence pro-tumorigenic and antitumor phenotypes. Of all infiltrating myeloid immune subsets, tumor-associated macrophages (TAMs) best represent this paradigm. TAMs are abundant in all stages of tumor progression and can be polarized into inflammatory M1 or immuno-suppressive M2 macrophages, depending on microenvironment stimuli (Ruffell and Coussens Lisa, 2015). While a subset of TAMs has antitumoral effects, others stimulate cancer cell proliferation by secreting growth factors, produce proteolytic enzymes that digest the ECM to facilitate tumor cell dissemination, and provide a supportive niche for metastatic tumor cells (Mantovani and Allavena, 2015). Eosinophils, primitive actors of innate immunity, have been shown to infiltrate tumors and influence tumor progression. Activated eosinophils secrete IL-10 and IL-12, to inhibit cancer cells growth, or can mediate cell death by direct cytotoxicity (Gatault et al., 2015; Lucarini et al., 2017). However, they can also promote tumor growth by secreting growth factors such as epidermal growth factor (EGF) and transforming growth factor-β1 (TGF-β1) (Grisaru-Tal et al., 2020). As tumors grow, myeloid-derived suppressor cells (MDSCs) (Kumar et al., 2016), immunosuppressive precursors of macrophages and dendritic cells (DCs), promote tumor vascularization and disrupt major mechanisms of immunosurveillance, including tumoral antigen presentation, T cell activation and cytotoxicity (Lindau et al., 2013).
The other major subset of tumor infiltrating immune cells is of lymphoid origin and includes T lymphocytes and natural killer (NK) cells. T lymphocytes can be grouped into 3 major subtypes: (i) TH lymphocytes divided mainly in two lineages: pro-inflammatory TH1 and anti-inflammatory TH2; (ii) Regulatory T cells (Treg), primarily pro-tumorigenic via their immunosuppressive activity; and (iii) cytotoxic T cells (TC) that destroy tumor cells through granzyme and perforin mediated apoptosis (Fridman et al., 2012; Lindau et al., 2013). A third lineage of effector TH cells, characterized by IL-17 secretion, called TH17 cells, acts as double-edged sword in anti-tumor immunity and tumorigenesis (Alizadeh et al., 2013).
Nerve Cells
Peripheral nerves are a common feature of the TME and emerging regulators of cancer progression. Innervated tumors are aggressive, have high proliferative indices and an increased risk of recurrence and metastasis (Magnon et al., 2015). Cancer cells can grow around nerves and invade them in a process called perineural invasion, which represents yet another route for dissemination (reviewed in Jobling et al., 2015). Recently, Zahalka et al. (2017) have shown that adrenergic nerves promote angiogenesis by activating the angiogenic switch in endothelial cells. Moreover, many studies described the formation of new nerve endings within tumors, showing that they stimulate their own innervation, a process termed axonogenesis, by expressing neurotrophic factors (Wang et al., 2014; Huang et al., 2015) or releasing exosomes containing axonal guidance molecules (Madeo et al., 2018). In return, nerves provide the tumor with neurotransmitters that enhance cancer cell growth.
Stromal Components
In healthy tissues, the stroma constitutes the main barrier against tumorigenesis. However, transformed cancer cells can direct stromal reprogramming to support tumor growth and progression.
The stroma is composed of the extracellular matrix (ECM) and specialized connective tissue cells, including fibroblasts, and mesenchymal stem cells.
The Extracellular Matrix
The ECM constitutes the scaffold of tissues and organs, providing the essential signals to maintain tissue architecture and to regulate cell growth and apoptosis. It is a complex network of glycoproteins, proteoglycans, glycosaminoglycans and other macromolecules. About 300 different proteins have been classified as ECM proteins, in what is called the matrisome (Hynes and Naba, 2012). The ECM undergoes constant remodeling by different actors, mainly enzymes such as collagenases and matrix metalloproteases (MMPs) and by fibroblasts. ECM stiffening, induced by increased collagen deposition and crosslinking, disrupts tissue morphogenesis contributing to malignant progression, but also facilitates metastasis and infiltration of immune cells in tumor sites (Bonnans et al., 2014).
Cancer-Associated Fibroblasts
Fibroblasts are widely distributed in all tissues. They constitute a multifunctional cell type residing in the ECM, shaping it by secreting collagens and fibrous macromolecules but also degrading it by releasing proteolytic enzymes, like MMPs.
Fibroblasts are known to modulate immune response by recruiting leucocyte infiltration and regulating inflammation via the secretion of growth factors, cytokines and chemokines and to play an important role in maintaining tissue homeostasis (Buckley et al., 2001). During wound healing or fibrosis, another type of specialized fibroblasts called myofibroblasts is present in the tissue (Tomasek et al., 2002). Tumors, for long considered as wounds that do not heal, are associated with a stroma similar to that observed in wound healing called the activated stroma, where fibroblasts resemble myofibroblasts and are called cancer-associated fibroblasts (CAFs). The activated stroma supports cancer progression (Hanahan and Coussens, 2012). Importantly, as for cancer cells, it has been described that CAF population is highly heterogeneous with tumor-promoting or tumor-suppressing CAFs and personalized anticancer therapies targeting CAFs could be of great interest (reviewed in Liu et al., 2019; Mhaidly and Mechta-Grigoriou, 2020).
Mesenchymal Stem Cells
The definition and characteristics of mesenchymal stem cells (MSCs) have been a matter of debate for a long time, and their characterization is an active field of research (Nombela-Arrieta et al., 2011). It is now established that MSCs are multipotent progenitor cells originating from the bone marrow that can migrate systemically through blood vessels and differentiate into osteoblasts, chondrocytes, or adipocytes. To date, the primary functions of MSCs within the TME are to regulate the immune response by the release of immunomodulatory cytokines and to promote tissue regeneration. Owing to their multipotent and cell fusion properties, they can also be at the origin of vascular cells, contributing to angiogenesis, of myofibroblasts and more rarely of cancer cells themselves.
Crosstalk Between Tumor Cells and Components of the TME in Cancer Progression
The tumor microenvironment plays a critical role in determining tumor fate, and stromal reprogramming has been recognized to be critical for carcinogenesis (Mantovani et al., 2008). Rudolf Virchow first proposed the possibility of a link between chronic inflammation and tumorigenesis in the nineteenth century after the observation of infiltrating leukocytes within tumors. This is now considered a hallmark of cancer. Cancer progression is associated with an ever-evolving tissue interface of direct epithelial–stromal interactions that regulate cancer cell metastasis and disease progression. This section describes the complex crosstalk between the actors of the TME and the cancer cells that take place during the different stages of cancer progression from the early cell autonomous events to the late metastatic stages (Figure 1).
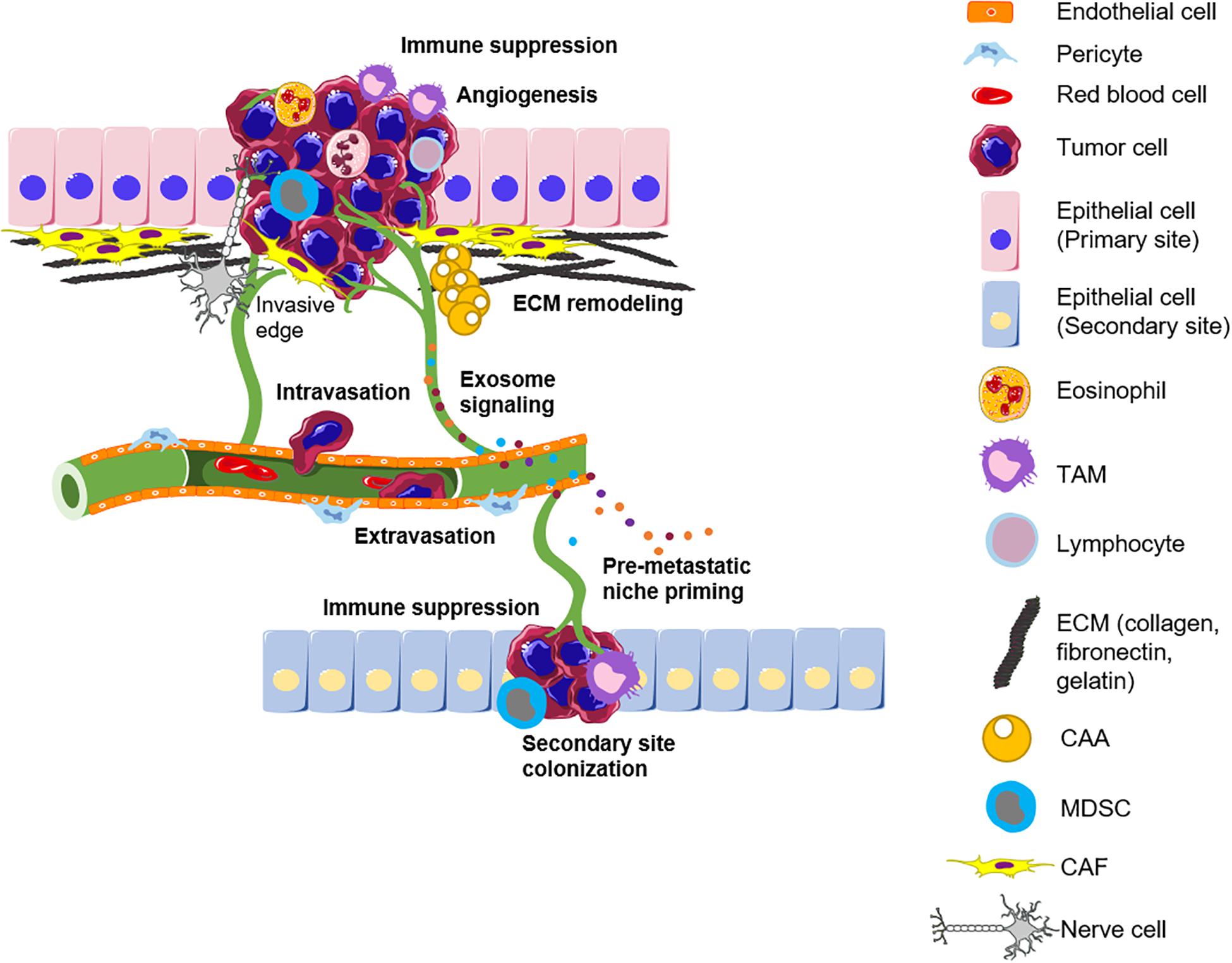
Figure 1. The tumor microenvironment influences the different stages of cancer progression. The primary tumor is infiltrated by different immune subsets and surrounded by a remodeled matrix. Angiogenesis ensures tumor growth by supplying nutrients and also provides a route for metastasis. Intravasation of tumor cells into blood vessels allows their shuttling to a novel site. The secondary site is primed by exosomes secreted by tumor cells and the different actors of the TME to allow the successful seeding of incoming tumor cells. TAM, Tumor-associated macrophage; ECM, Extracellular matrix; CAA, Cancer-associated adipocytes; MDSC, Myeloid-derived suppressor cells; CAF, Cancer-associated fibroblast; TME, Tumor microenvironment.
Primary Tumor Progression
Cancer cells reprogram the tumor-infiltrating stromal and immune cells, which facilitates primary tumor growth and progression. Therefore, it is important to decipher the reciprocal crosstalk between cancer cells and their heterotypic microenvironment.
Epithelial cancer progression is influenced by the cells’ contact with immune cells and a carcinogen-exposed stroma (Barcellos-Hoff and Ravani, 2000), by an overexpression of metalloproteases (Fukuda et al., 2011) which create a suitable environment for invasion, or by the stimulation with altered stromal cells like CAFs. In the skin, epigenetic modifications of fibroblasts are induced by ultraviolet exposure, leading to the production of inflammatory cytokines and matrix-remodeling enzymes that together influence the formation of epithelial tumors (Hu et al., 2012). CAFs accumulate in the TME along with tumor growth (Kalluri, 2016) and are activated by cytokines and growth factors of the TME, such as TGF-β (Taniguchi et al., 2020) and Fibroblast Growth Factor (FGF). In their turn, CAFs provide growth factors like VEGF to enhance angiogenesis and vascular permeability (Fukumura et al., 1998). Furthermore, TAMs can support many aspects of tumoral progression. They can secrete mediators that enhance tumor cell survival and proliferation such as growth factors and cytokines [epidermal growth factor (EGF), interleukin 6 (IL-6), and tumor necrosis factor (TNF) (Noy and Pollard Jeffrey, 2014)].
Another crucial step for cancer progression is immune evasion. This is supported mainly by the action of MDSCs. These cells infiltrate the developing tumors and inhibit the mechanisms of immune editing of cytotoxic immune cells, all the while promoting tumor vascularization (Talmadge and Gabrilovich, 2013). TAMs can also promote cancer immune escape by displaying immunosuppressive functions (Noy and Pollard Jeffrey, 2014). Other myeloid cells including neutrophils, monocytes, and eosinophils infiltrate the tumor and promote tumor growth by inhibiting antitumor immunity. Neutrophils can even induce genotoxic damages (Wilson et al., 2015) or recruit tumor-promoting TH17 lymphocytes (Ortiz et al., 2015). Additionally, invasion of the basement membrane underlying the epithelium by the tumor cells is a basic step for the upcoming dissemination. For this, CAFs have a physical impact on tumors that results in increased ECM stiffness around tumor cells and consequent mechanical stress. TAMs are also capable of driving invasive phenotypes (Condeelis and Pollard, 2006). In breast cancer, they facilitate invasion of tumor cells by sustaining a signaling paracrine loop involving CSF-1 and EGF (Goswami et al., 2005), and by the secretion of proteases (Gocheva et al., 2010). Thus, once the tumor cells evade the host immune system and gain the ability to invade the surrounding tissue, metastatic dissemination of cancer cells can take place.
Metastatic Dissemination
Metastasis is the leading cause of mortality among cancer patients (Mehlen and Puisieux, 2006). In epithelial tumors, metastasis begins with the cellular invasion of the basement membrane and the subsequent migration of cancer cells into the blood stream. One of the initial steps for primary tumor invasion is epithelial-mesenchymal transition (EMT). Under the influence of various signals, mainly TGF-β, cells gradually lose their epithelial traits while gaining mesenchymal ones that confer migratory capacities (Mani et al., 2008). CAFs participate in a TGF-β and platelet-derived growth factor (PDGF) signaling crosstalk with tumor cells to support EMT and the acquisition of an invasive phenotype (van Zijl et al., 2009). EMT can also enable the acquisition of CSC traits (Mani et al., 2008), suggesting that not only it causes cancer cells to disseminate from the primary tumor but also can provide these cells with the self-renewal properties needed for their subsequent implantation at secondary sites. Although CSCs are not be the only cells responsible for metastasis, the CSC-generated hierarchy of stem-like and differentiated tumor cells is able to initiate metastatic growth (Merlos-Suárez et al., 2011). However, EMT is not the only mechanism used by epithelial cells for migration. Epithelial cancer cells can migrate as single cells, as loosely attached cords or as highly organized collective entities (reviewed in Friedl et al., 2012). During early stages of cancer migration, CAFs increase the production of collagen in the underlying stroma and the fibers become aligned, giving rise to a stiffer ECM hence allowing the migration of cancer cells away from the primary tumor (Conklin et al., 2011). This is largely mediated by CAFs secreted factors that stiffen the ECM, namely enzymes of the Lysyl Oxidase (LOX) family (Kalluri and Zeisberg, 2006).
During metastasis, cancer cells cross the endothelial barrier during a step called intravasation to enter the blood stream, and by extravasation to exit from circulation into distant tissues, processes that involve different receptors, a plethora of signaling pathways, and interactions with the actors of the surrounding microenvironment (Reymond et al., 2013). Intravasation seems to require the cooperative work of a triad consisting of macrophages that localize to blood vessels where they help tumor cells intravasate into the blood stream (Harney et al., 2015). However, despite the help of macrophages, only 0.01% of cells that intravasate form detectable metastases (Chambers et al., 2002). Cancer cells in the blood stream can be shielded by platelets from NK-mediated cytotoxicity (Palumbo et al., 2005), and platelet binding enhances cancer cell adhesion to vessel wall and subsequent extravasation (Zhang et al., 2011; Schumacher et al., 2013). Inflammation also modulates endothelial crossings through TNF-induced vascular permeabilization, cyclooxygenase 2 (COX2)-dependent prostaglandin production and MMP-mediated tissue remodeling.
Secondary Organ Colonization
Docking of cells in organs to form secondary tumors is not a random process. Organ tropism has been first described by Stephen Paget in 1886 as the “seed-and-soil” theory, in which he suggests that metastasis is not the fruit of hazard but tumors have clear organ preferences for secondary colonization. Paget’s theory gave the basis for the description of the premetastatic niche: the primary tumor executes preparative events, preceding detectable metastasis, that render the secondary milieu less hostile for colonization by cancer cells. Studies of the premetastatic niche are still in their infancy but some traits and events are now clearer. Settlement of tumor cells at distant sites is dependent on tumor-secreted cytokines and extracellular vesicles, like exosomes, that enable the premetastatic microenvironment to support their colonization (Liu and Cao, 2016). These tumor-secreted factors communicate to both hematopoietic and mesenchymal stem cell compartments. It has been shown that bone marrow-derived VEGFR1+ cells are already present in premetastatic sites before tumor cell arrival, suggesting the communication between primary and secondary sites (Kaplan et al., 2005). Seeding is also facilitated by the LOX-mediated fibronectin upregulation in resident fibroblasts and recruitment of myeloid cells (Erler et al., 2009). Neutrophils may also be involved in the priming of metastatic sites. Neutrophils accumulate in premetastatic livers of mice bearing colorectal tumors (Wang et al., 2017) and their accumulation has been shown to be required for pancreatic cancer metastasis (Steele et al., 2016). Recently, it was also shown that omentum resident macrophages are required for ovarian cancer metastasis (Etzerodt et al., 2020). Neutrophils also serve as an energy source to fuel metastatic tumor cells. In a breast cancer model, infiltrating neutrophils are induced to store lipids upon interaction with resident mesenchymal cells in the lung so that when disseminated tumor cells (DTCs) arrive, neutrophils transfer their stored lipids to DTCs for their survival and proliferation (Li et al., 2020).
Colonization of secondary tissues requires the same elements as growth of the primary tumor namely, sufficient nutrients and oxygenation. One important step for metastatic tumor cell survival is the reversal to an epithelial phenotype via mesenchymal–epithelial transition (MET) to regain the ability of proliferation and differentiation. Once tumor cells colonize the secondary site, genetic instability inherent in neoplastic cells continues to operate at each cell division, and these cells continue the remodeling of the site, just as described above.
Accordingly, the crosstalk between cancer cells and their microenvironment provides valuable insights into cancer formation, progression and spread. Hence, it is necessary to study cancer as a whole process by modeling the interactions between tumor cells and their microenvironment to improve development of new therapies against cancer progression and metastasis.
Part II—Organotypic in vitro Models
Advantages of 3D Models Over 2D Models and Animal Experiments
Cancer research has long been based on two-dimensional (2D) cell culture, mainly in order to earn the right of passage to in vivo experiments. Conventional 2D cell cultures allowed the study of many mechanisms that drive tumor growth and the evaluation of optimal drug doses and toxicities. However, currently available cell lines fail to represent the genetic background across the range of human cancers (Huang A. et al., 2020) and may adapt to growth in culture, rather than mimic the behavior of the tumor in a complex microenvironment. Because they also lack all elements of the tumor stroma and surrounding tissue, they fail to mimic the complexity of the tumor microenvironment (Gillet et al., 2011). Owing to this, a large gap exists between the knowledge obtained in these models compared to in vivo cancer models because results of 2D experiments rarely predict therapeutic response in animals. This can be explained by the fact that cells cultured in 2D do not have the same architecture as cells in vivo that are arranged in three-dimensional (3D) structures unattached to planar surfaces. Furthermore, cultured monolayers lack the capacity to mimic in vivo tumoral hypoxia and exhibit a very different metabolism. Consequently, cells in monolayer cultures proliferate at unnaturally rapid rates (Langhans, 2018), differ in gene/protein expression compared to in vivo models, and alter their dynamic processes such as cell division and migration (Duval et al., 2017).
Even though in vivo experiments have the advantage of being physiologically relevant in contrast to cells cultured out of their bodily context, they have many flaws (Day et al., 2015). Aside from being long, expensive and ethically questionable, the use of human cancer cells in mouse models mostly requires the use of immunocompromised mice that lack, to varying extents, the immune components, thus limiting the advantages of these approaches in modeling tumoral progression and response to drugs. Indeed, the inflammatory immune cell component is lacking in immunocompromised mice. Although the engrafted tumors may exhibit a stromal response with the growth of endothelial cells and fibroblasts, these stromal cells originate from mice and therefore the implication of human TME could not be extrapolated. Moreover, it has recently been shown that patient-derived xenografts (PDXs) present genomic instability with continuously changing copy number alterations landscapes, and so their passaging causes a drift from the original tumor (Ben-David et al., 2017). As such, mouse co-clinical trials using PDXs have shown very little progress beyond proof of concept due to logistical issues (Clohessy and Pandolfi, 2015).
Even with strong supporting preclinical evidence, many targeted therapies produce modest clinical results, a fact now highlighted by the tremendous National Lung Matrix Trial that assessed personalized medicine in non–small cell lung cancer (NSCLC) (Middleton et al., 2020). The results have been fairly disappointing with a response rate of only 10% with some abandons due to lack of treatment efficacy. Genetically engineered mouse models of NSCLC, used for preclinical studies, have mutational burdens more than 100-fold lower than that of human disease (McFadden et al., 2016) arguing for the use of more appropriate preclinical models that integrate the immune and stromal landscapes beyond the genetic aberrations.
Another issue resides in the ability to translate results of immunotherapy from bench to clinic because of the high failure rate observed in human clinical trials after promising results obtained in mouse models. Even the durable clinical benefits observed with immune checkpoint blockers (ICBs) in some tumor types have been seen in a minority of patients (Cardin et al., 2014; Herbst et al., 2014; Hammel et al., 2016). Given the complexity of the tumor microenvironment, it is imperative to create models that include different immune cell types the administered compound may interact with.
Efforts have been made these last few years to “humanize” the mouse’s immune system by grafting human hematopoietic stem cells in mice or by transgenic expression of Human Leucocyte Antigen (HLA) (reviewed in Shultz et al., 2012; De La Rochere et al., 2018). However, the high cost of recipient mice, scarcity of human bone marrow acquisitions, engraftment variability, and laborious technical demands represent high inconveniences in a preclinical setting. Hence, optimal mouse studies are very cumbersome for simultaneous evaluation of numerous drugs and may be inefficient due to the different metabolic processing of drugs between humans and mice. Thus, high-throughput in vitro screening systems are essential precursors to in vivo evaluations. Developing 3D organotypic models that recapitulate physiological functions would allow further replacement and reduction of animal models as recommended by the 3Rs rule1.
In vitro 3D cultures recapitulate much better the architecture of tissues and capture the complexity of solid tumors than 2D counterparts, all the while allowing the modeling of different stages of the carcinogenic process (Yamada and Cukierman, 2007; Tanner and Gottesman, 2015). The concentric arrangement of cells in 3D cultures resembles initial avascular stages of solid tumors in vivo and non-vascularized micro-metastatic foci. More sophisticated 3D cultures also include different elements of the TME; allowing their use to study cellular interactions within tumors and to model stages of cancer progression. Additionally, genome-wide screens performed on 3D cultures showed improved detection of cancer genes and pathways compared with those performed in 2D (Han et al., 2020). Thus, increased biologically relevant behavior and characteristics could be acquired from genetic editing in organoids, cocultures, and 3D growth models. Moreover, the coalition between biologists, bioengineers and physicians inspired many strategies to reproduce ex vivo the complexity of biological systems. These approaches mimic organ topography, mechanical forces of tumor cells, matrix stiffness, functionality, and complexity much better than 2D or even 3D culture systems (van Duinen et al., 2015).
In the following section, we will describe the existing in vitro organotypic models for cell culture (Figure 2).
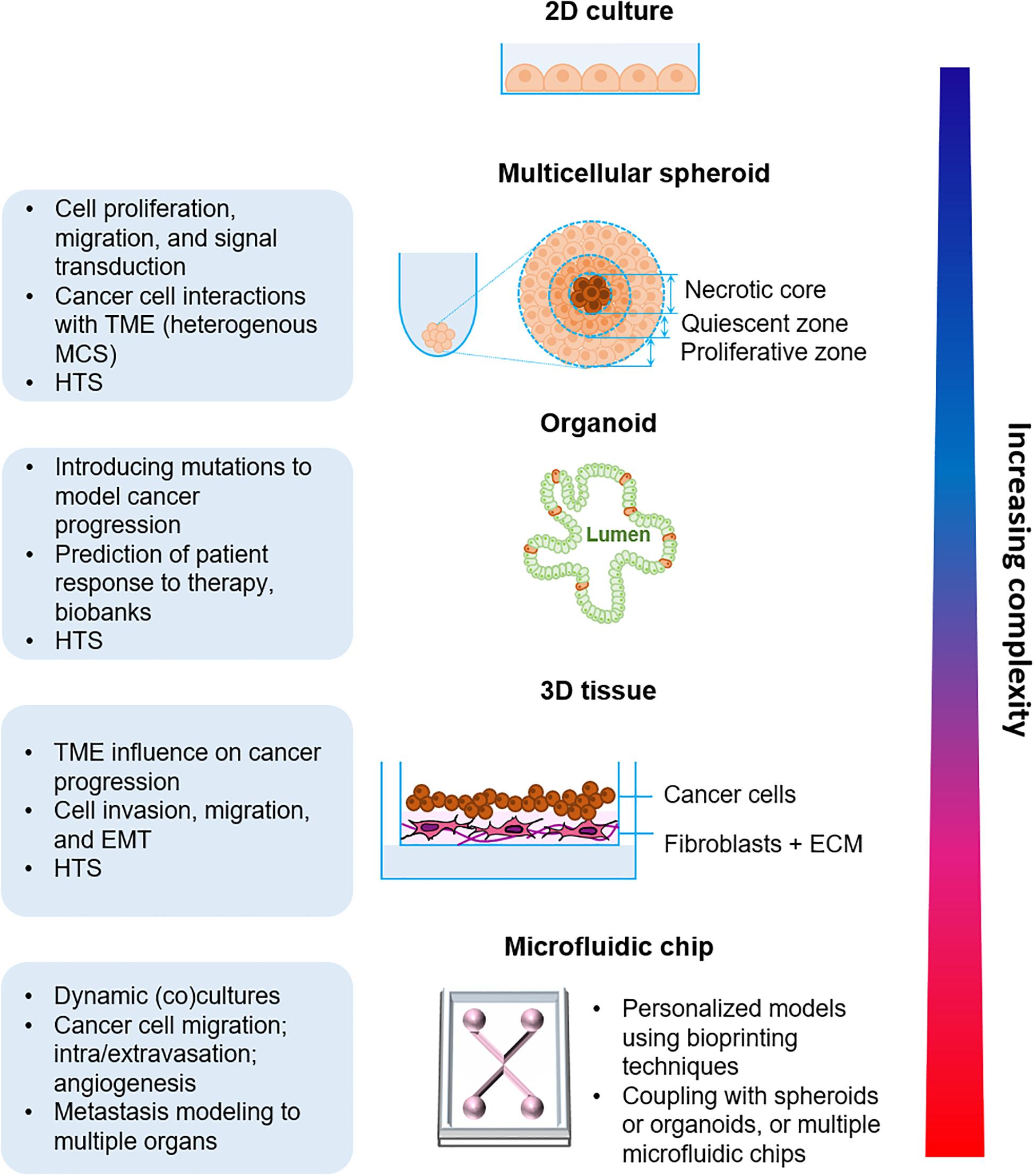
Figure 2. Organotypic in vitro models of cancer progression with increasing biological complexity from the simple multicellular spheroid model to the more complex microfluidic approaches. TME, Tumor microenvironment; MCS, Multicellular spheroid; HTS, High-throughput screening; ECM, Extracellular matrix.
Overview of in vitro Organotypic Cellular Models
Multicellular Spheroids
Multicellular spheroids (MCSs) or 3D cellular aggregates represent the bridge that fills the gap between 2D cultures and more elaborate 3D techniques. They are fairly representative of the in vivo situations because of their heterogeneity as they are composed of proliferating, non-proliferating, well-oxygenated, hypoxic and necrotic cells. Other features of MCSs like cell-cell signaling and interactions, the presence of different cellular layers, the genetic expression profiles, and drug resistance patterns are similar to characteristics of the natural cellular conditions. Currently, there exists many techniques for MCS production such as the forced floating methods in non-adherent plates, hanging drop method, the use of scaffolds and matrices, or even more sophisticated methods using microfluidic systems (reviewed in Ferreira et al., 2018).
MCS can be used for tumoral modeling by either forming homogenous cultures using solely cancer cells, or by more sophisticated cultures using cancer cells with components of the TME like fibroblasts, endothelial cells (Andrique et al., 2019) or immune cells, hence forming heterotypic spheroids. Encapsulating MCS in biomimetic hydrogel scaffolds offers biophysical and biochemical cues that simulate the behavior of extracellular matrix, essential for regulating cancer cell behavior (Li and Kumacheva, 2018).
Organoids
The term organoid, meaning resembling an organ, was first used in 1946 by Smith and Cochrane to describe a case of cystic teratoma. Ever since, it has been inaccurately used to describe some cell structures and aggregates, but the actual definition is now clearer: an organoid is a collection of organ-specific cell types that develops from stem cells, that possesses a minima of specific organ functions, and self-organizes to mimic the architecture of the organ itself (reviewed in Lancaster and Knoblich, 2014). Early pioneering works of Mina Bissell showed that primary epithelial cells derived from mouse mammary glands could self-organize into glandular structures and secrete milk proteins (Lee et al., 1984). These advances were followed by the works of Clevers’ lab, that described the generation of intestinal crypt organoids from Lgr5+ stem cells (Sato et al., 2009).
Now, it is recognized that organoids can be generated using two types of stem cells: pluripotent stem cells (PSCs) which can be embryonic or induced pluripotent stem cells or adult stem cells (ASCs) that reside in adult tissues and are tissue-specific, cultured under specific growth factor cocktails that allow their long-term expansion by mimicking the organ stem cell niche. To date, organoids have been developed for many organs including intestine (Spence et al., 2011), kidney (Takasato et al., 2015), brain (Lancaster et al., 2017), liver (Camp et al., 2017), stomach (McCracken et al., 2014), pancreas (Hohwieler et al., 2017), ovary (Kessler et al., 2015), and lung (Dye et al., 2015) among others. These organoids have been used for multiple approaches such as high-throughput drug screening efficacy and toxicity, host-microbe interactions and infectious diseases (Bartfeld et al., 2015; Leslie et al., 2015; Garcez et al., 2016), and disease modeling (reviewed in Dutta et al., 2017) in particular tumor development, which will be later discussed in detail.
Epithelial organoids recapitulate many aspects of organ development and disease and represent many opportunities for cancer modeling and anticancer drug testing. However, it is important to note the existence of some drawbacks and limitations. Organoids lack the native organ microenvironment: the stromal compartment, immune cells and vascularization, and they are mostly cultured in poorly defined animal matrices. Although, novel synthetic analogous ECM may constitute a better alternative as they are controllable and permit fine tuning of matrix constituents (Gjorevski et al., 2016).
3D-Tissues
The recreation of simple tissues has been described in a cell sheet engineering method using cells grown to confluence on culture dishes grafted with a temperature-responsive polymer, poly-(N-isopropylacrylamide). This technique allows cell growth at 37°C and cell harvest at room temperature as intact cell sheets and subsequently the stacking of different sheets to generate heterotypic thin 3D tissue analogs. Using this technique vascularized tissues (Asakawa et al., 2010) and liver tissue-like structures (Kim et al., 2012) were obtained.
Organotypic epithelial raft cultures, originally developed to study keratinocytes (Fuchs, 1990), represent an interesting approach to study epithelial cancer cell behavior, notably cancer cell invasion. These cultures are mechanically supported by semipermeable inserts and are either submerged in medium or maintained at an air–liquid interface. Epithelial tissues can be constructed in stages by first embedding stromal cells, mainly fibroblasts, for several days followed by seeding of epithelial cells on top (Kalabis et al., 2012), or also embed immune cells within the layers to obtain an integral tissue (Huang et al., 2017). These cultures generate a stratified tissue resembling the epithelium seen in vivo with a proliferating basal layer and differentiating supra-basal layers. The use of 3D-tissue revealed some advantages compared to organoids when the access to the epithelial cells’ apical surfaces is needed, for example to study host-pathogen interactions. To illustrate this using a colonic 3D-tissue, Martin and colleagues have shown that infection with genotoxin-producing Salmonella enterica synergises with the loss of APC to promote genomic instability and carcinogenesis (Martin et al., 2019). Although it should be noted that an elegant recent study has described the possibility to revert organoid polarity allowing access to the apical surfaces of the cells (Co et al., 2019). Miniaturized 3D tissues can be used to facilitate high-throughput drug screening (Dutta et al., 2017).
Microfluidic Approaches
The static nature of nutrients and metabolites in 3D cultures isn’t representative of the physiological conditions due to the lack of fluid shear stress and hydrostatic pressure that can greatly influence cell behavior (Polacheck et al., 2011). Microfluidic systems, based on the progress in synthetic biology, have enabled the development of in vitro assays that facilitate the study of cellular behavior under a spatiotemporally controlled microenvironment in which molecular, biophysical and cellular components can be tuned according to physiologically relevant parameters. These microfluidic cell culture systems, known under the term organ-on-a-chip, are usually made of continuously perfused hollow microchannels populated by living cells (reviewed in Bhatia and Ingber, 2014). To date, many organs have been successfully modeled in microfluidic devices. One of the first models was lung alveoli that responded to bacterial infection and inflammation (Huh et al., 2010), but also that reflected drug toxicity (Huh et al., 2012). Many studies followed that assessed nephrotoxicity in human kidney tubules-on-a-chip (Jang et al., 2013), liver function (Beckwitt et al., 2018), and more recently, a simulation of a body-on-a-chip multi-organ system (McAleer et al., 2019) to assess drug efficiency and toxicity.
Part III—Organotypic Models of Cancer Progression and Drug Response
Understanding the key aspects of tumoral progression is of utmost importance for the development of novel successful anticancer strategies. Organotypic modeling of these aspects alongside the interactions between the different actors of the TME would allow a better comprehension of the mechanisms that mediate tumoral progression and a first solid step toward preclinical drug screening in physiologically relevant situations. In this section, we describe how the previously mentioned organotypic models have been applied to study the different steps of tumor growth and metastasis (Table 1).
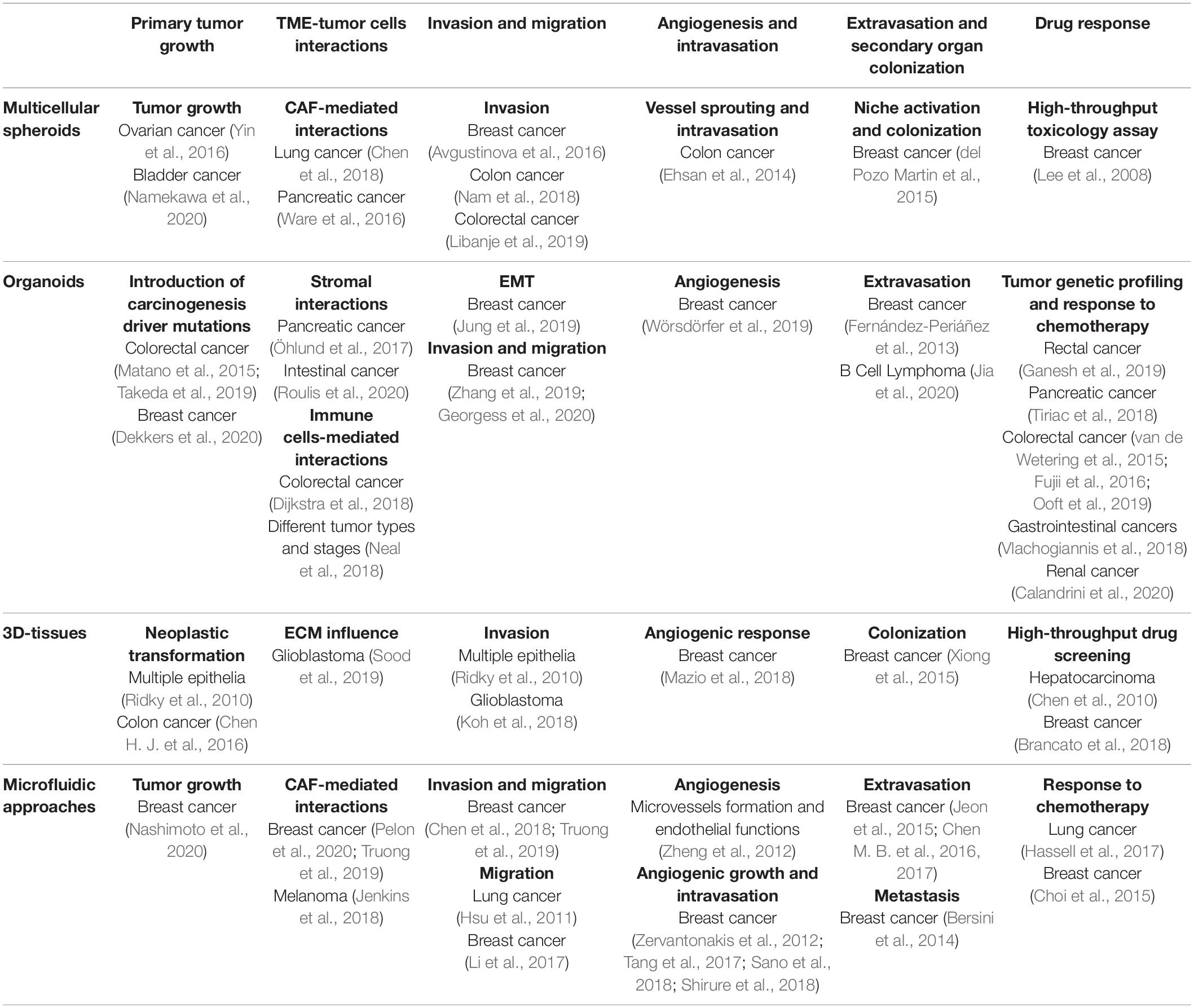
Table 1. Organotypic models used to study cancer progression stages and drug response.
Cancer Modeling Using Organotypic Models
Tumor Growth in situ—Interactions of Cancer Cells With the TME Elements
Many in vitro organotypic models have been used to study tumor initiation and growth and to identify how parenchymal cells (endothelial, epithelial, immune, nerve and stromal cells) and components (ECM, secreted factors) of the TME influence the growth in situ of different cancer types.
Modeling cancer initiation using organoid is highly attractive owing to the relative ease of genetic manipulation of cells. Using CRISPR-Cas9 genome editing, tumor suppressors have been identified (Michels et al., 2020), as well as the consequences of mutations in the DNA repair deficiency genes (Drost et al., 2017) or mutations that drive cancer progression (Fumagalli et al., 2017) have been elucidated. Such approaches allow the introduction of defined mutations to transform normal organoids and induce tumor growth, upon xenotransplantation. Matano and colleagues model human colon adenocarcinoma by introducing canonical colorectal cancer (CRC) driver mutations into primary human colon organoid cultures (Matano et al., 2015), revealing that mutations in APC, SMAD4, TP53, and KRAS simultaneously are sufficient to model colonic adenomas but not tumorigenesis, perhaps due to the lack of TME components within the organoids. Similarly engineered CRC organoids with APC and KRAS mutations formed dysplasia and could invade submucosa (Takeda et al., 2019), and transformed mammary organoids formed tumors upon xenotransplantation (Dekkers et al., 2020). Thus, deconstructing carcinogenesis into single genetic elements by engineering cancer genes in untransformed human organoids is a powerful tool for investigating how individual genetic aberrations contribute to the acquisition of cancer phenotypes.
Nevertheless, the genetic alterations driving cancer initiation are supplemented by the interactions of cancer cells with their microenvironment to ensure successful cancer progression. A refined cancer 3D-tissue model using cancer-associated genetic modifications and a stromal department showed the neoplastic transformation of normal epithelia which became invasive (Ridky et al., 2010). Indeed, many tumors are characterized by a prominent stromal compartment that modulates tissue architecture, due to extensive ECM remodeling mainly mediated by CAFs. Adding stromal fibroblasts to prostate organoids facilitated their branching (Richards et al., 2019), while the addition of CAFs to lung squamous carcinoma spheroids recapitulated the pathological changes of tumorigenesis, from invasion and hyperplasia to dysplasia (Chen et al., 2018). Additionally, CAFs were shown to enhance invasion and migration of breast cancer cells in a 3D microfluidic device (Nguyen et al., 2018; Truong et al., 2019).
Furthermore, the coculture of pancreatic stellate cells, a resident mesenchymal cell population that differentiates into CAFs, with pancreatic cancer patient-derived organoids (PDOs) (Öhlund et al., 2017) or with spheroids (Ware et al., 2016) produced a highly desmoplastic stroma, typical of pancreatic carcinomas. Equally investigating the role of the TME in CRC initiation using organoids, Roulis and colleagues performed single-cell RNA sequencing of the murine intestinal mesenchymal niche and found a population of fibroblasts in intestinal crypts that orchestrate intestinal tumorigenesis by exerting paracrine control over tumor initiating stem cells (Roulis et al., 2020).
Other key elements of the TME which significantly affect cancer cell behavior are immune cells. Tumor-immune system interactions have been widely studied by culturing immune cells recovered from patients together with established cancer cell lines in conventional monolayer cultures. However, these approaches fail to account for critical aspects of the TME. Indeed, microfluidic devices customized with human tumor spheroids containing immune cells recapitulate some features of response or resistance to immune checkpoint blockade in melanoma (Jenkins et al., 2018), but without features of the stromal compartment. The recent promise of therapies manipulating tumor-infiltrating immune cells created a particular exigency for human cancer models that recapitulate this TME diversity. In an effort to integrate an immune competent microenvironment to organoid cultures, a platform to induce and analyze tumor-specific T-cell responses to epithelial cancers was established (Dijkstra et al., 2018). Enrichment of functional tumor-reactive T lymphocytes from CRC or non-small cell lung cancer (NSCLC) patients was successfully established by cocultures of peripheral blood lymphocytes with autologous tumor organoids. These tumor-reactive T cells efficiently recognize and kill autologous tumor organoids, while leaving healthy organoids unharmed. Moreover, a recent study presents organoid modeling that preserves primary tumor epithelium with its endogenous immune and non-immune stromal elements (Neal et al., 2018).
Cancer Progression: EMT, Cancer Cell Migration and Invasion
The metastatic cascade initiates with invasion and migration of tumor cells away from the primary tumor. Invasion through the basement membrane is considered a differentiating step between neoplasia and malignant tumors. Because cancer cell contractility and matrix stiffness are critical parameters for invasion, accurate invasion models should include tunable matrix parameters (Wisdom et al., 2018). This is possible using organotypic 3D tissues, where virtually any component can be readily modulated. The stromal compartment can be enriched not only with fibroblasts but with myofibroblasts, endothelial cells or inflammatory cells (reviewed in Coleman, 2014).
To study the basis of cancer invasion, significant efforts have been made to recapitulate tumor–stroma interactions. Multicellular spheroids combined with ECM containing fibroblasts showed enhanced invasion (Avgustinova et al., 2016). However, the tumor and its environment being highly dynamic, microfluidic approaches are more fitted to study tumor cell migration. Indeed, the use of such approaches unveiled the contributions of different cell types to tumor cell migration and invasiveness. A 3D microfluidic coculture system containing side-by-side tumor and stroma regions showed that CAFs enhanced the migration and invasiveness of cancer cells (Truong et al., 2019; Pelon et al., 2020). TGFβ secreted by cancer cells was shown to stimulate fibroblasts to transform into myofibroblasts, which then produced soluble factors that fed back to increase the migration speed of the cancer cells (Hsu et al., 2011). Likewise, the cytokines secreted by macrophages cocultured with cancer cells in a microfluidic device, increased cancer cell migration speed and persistence in a MMP-dependent fashion (Li et al., 2017).
Angiogenesis and Cancer Cell Intravasation
Over the last decade, biomimetic 3D vascular models have been developed, contributing to the understanding of angiogenic processes. Rings of tissue from human umbilical arteries embedded into a 3D matrix were able to sprout in response to tumor-derived proangiogenic factors (Seano et al., 2013). However, vascular organotypic models should not be static as shear forces and blood flow are important for the vascularization process. So, microfluidic approaches have been developed in which endothelial cells are seeded into a channel within ECM to form a primitive vasculature that can be stimulated by angiogenic factors (Zheng et al., 2012; Nguyen et al., 2018), or with an incorporated layer of human bone marrow stromal cells around the channels to recapitulate perivascular barrier function (Alimperti et al., 2017). These microfluidic chips can also be used to trigger vasculogenesis; in that case, instead of seeding endothelial cells beside the matrix, endothelial cells, fibroblasts (Jeon et al., 2014) and tumor cells (Chen M. B. et al., 2017) are loaded within the matrix. Moreover, the ability of organoid-on-a-chip to mimic perfusable blood vessels may address an important issue of organoid use: the lack of nutrient supply. To surmount this, a tumoroid-on-a-chip was developed. It was created in a microfluidic device consisting of three interconnected chambers that enable the self-assembly of endothelial cells into a 3D network of blood vessels and their angiogenic growth toward the organoid-like structures from breast cancer patients (Shirure et al., 2018). However, in such approaches, endothelial cells may not always be free to interact with tumor cells because of the artificial membranes used in the organ-on-a-chip devices. To address this issue, endothelial cells were modified to produce ‘reset’ vascular endothelial cells (R-VECs) that grew into 3D branching vessels capable of transporting human blood in microfluidic chambers and when transplanted into mice (Palikuqi et al., 2020). These R-VECs adapted their growth upon their coculture with either normal colon organoids or patient-derived colorectal organoids. They arborized normal colon organoids and helped sustain their proliferation while they erratically infiltrated tumor-derived organoids, thus providing a novel physiological platform to study vasculogenesis and angiogenesis.
Entry of tumor cells into the blood stream is a critical step in cancer metastasis. Using microfluidic devices, interactions between invasive cancer cells and endothelial cells have been studied. It was shown that treatment of the endothelium with TNF or coculture with macrophages resulted in rapid and increased numbers of tumor cell–endothelial cell attachment events (Zervantonakis et al., 2012). The secretion of cytokines and chemokines by cancer cells increases the permeability of the endothelial barrier, allowing tumor cells to intravasate and extravasate (Reymond et al., 2013). This feature was modeled using a perfused microfluidic platform containing a vascular compartment with breast cancer cells and their associated endothelial cells separated via a micropillar array interface that allows direct communication of tumor and endothelial cells. The permeability of the vessels was greatly increased in response to the presence of tumor cells or tumor cell-conditioned medium (Tang et al., 2017). Moreover, a tissue-engineered model containing a realistic microvessel in coculture with mammary tumor organoids allowed real-time monitoring of tumor cell-vessel interactions. Using this model, it was shown that tumor cells can reshape, destroy, or intravasate into blood vessels (Silvestri et al., 2020).
Extravasation and Secondary Site Colonization
Cancer cells within vessels must extravasate to colonize new sites. This process is different from intravasation, because the vasculature to be breached is healthier and cancer cells experience fluid shear stresses due to blood flow. After extravasation, cancer cells have one final task to complete: colonization of secondary sites. Extravasation of tumor cells has been shown to occur via endothelial apoptosis in vitro (Heyder et al., 2002) but via necroptosis in vivo (Strilic et al., 2016). Thus, accurate modeling of the extravasation and colonization steps requires tissue-specific cell types, microenvironmental cues, and vascularization. Breast cancer cells extravasated through a vascular network into a bone-mimicking microenvironment generated by culturing osteo-differentiated MSCs within a hydrogel, or within a microfluidic device (Jeon et al., 2015; Sano et al., 2018). It was shown that extravasation rates were much higher to the bone microenvironment than to stromal matrices alone. Another similar model showed that β1 integrin expression is required for cancer cells to be able to invade through the endothelial basement membrane (Chen M. B. et al., 2016). Increased complexity and clinical relevance can be incorporated into organ-on-a-chip models, as devices have been developed to mimic interactions between circulating tumor cells (CTCs), endothelium and bone microenvironments as a model of metastasis to bone (Bersini et al., 2014).
Therapeutic Applications of Organotypic Models
Although the demand for anticancer drugs is constantly increasing, their development is slow and fastidious. Monolayer cultured cells are the most widely used in vitro models despite their inability to accurately reflect drug’s metabolism and pharmacokinetics in the human body. For years, cell-based drug discovery was based on monolayer cultures of authenticated cell lines (Smith et al., 2010; Barretina et al., 2012), but in this blooming era of precision medicine (Prasad et al., 2016), organotypic models represent great promise for anticancer drug discovery.
In line with this, using an organ-on-a-chip approach, a human lung cancer chip has been developed to study tumor growth patterns and drug response (Hassell et al., 2017). When lung cancer cells were cultured within a physiological-like microenvironment composed of lung endothelial cells, normal lung alveolar epithelium and ECM, they presented rampant growth and resistance to tyrosine kinase inhibitors (TKI) similar to NSCLC patient’s response, while they failed to do so in static conventional culture. Likewise, McAleer and colleagues designed a modulable five-chamber multi-organ system to monitor drug effects and simultaneously examine anticancer drug efficacy and off-target toxicity (McAleer et al., 2019). In two models incorporating an array of cancer and healthy human cell types, the system provided insight into the efficacy and toxicity of diclofenac, imatinib, and tamoxifen.
Beyond engineered organoids, organoids derived from patient biopsies or resected tumors, called patient-derived organoids (PDOs) have been successfully cultured with a high success rate and indefinite expansion. These contain tumor cells and stromal cells, thus providing a more realistic microenvironment and they seem to retain the tissue identity of the patient (Tiriac et al., 2018; Ganesh et al., 2019), indicating their great potential for personalized medicine approaches. Recent studies suggest that PDOs mirror clinical responses of individual patients to therapy within a clinically meaningful timeframe and even predict patient response to chemotherapy (Ooft et al., 2019; Pasch et al., 2019). Indeed, PDOs derived from glioblastoma samples were used to test responses to standard of care therapy as well as targeted treatments, like chimeric antigen receptor T (CAR-T) cell immunotherapy in a clinically relevant timescale (Jacob et al., 2020). These PDO properties laid the foundation of what is now known as organoid biobanks (van de Wetering et al., 2015; Calandrini et al., 2020) used for applications such as drug testing, cytological analyses, and xenografting.
With the significant need for biomarker identification of drug response, PDOs could also be considered as a tool for biomarker discovery by analyzing secreted factors such as extracellular vesicles (Huang L. et al., 2020) in contrast with PDX models, due to the presence of contaminating host factors. Although molecular diagnostic testing is now routinely used to determine the choice of targeted therapies for the treatment of cancer patients, patients in advanced stages who have exhausted standard clinical care approaches lack personalized therapeutics and will endure the arduous regimen of chemotherapy and see little or no benefit. Even if the use of functional testing in guiding personalized medicine in still in its infancy, the use of metastatic cancer site derived PDOs to evaluate drug response has proven its efficacy by recapitulating patient response (Weeber et al., 2015; Fujii et al., 2016; Pauli et al., 2017; Vlachogiannis et al., 2018). These evaluation platforms could be of great interest in orienting the treatment of advanced cancer patients.
Shortcomings and Future Directives of Organotypic Models in Translational and Preclinical Settings
The use of organotypic models for cancer modeling is a blooming area of research, however, there are still limitations to their use (Puca et al., 2018; Fujii and Sato, 2020). As an example, studying angiogenesis is rudimentary when it comes to organotypic models. Indeed, the use of vasculature is very basic and organotypic models with other surrounding tissue types are necessary to model more physiological situations. It would be of great interest to model angiogenesis and neovascularization within a transformed organoid. Additionally, complexifying organotypic models by engineering organoids surrounded by muscle, an immune system, and containing a neuronal network along with functional vasculature is something to look forward to in the near future.
When it comes to preclinical studies, organotypic models face many caveats. Spheroid-based 3D models must be used with caution when it comes to clinical relevance. Because they are generated from non-primary tumor cell lines (Friedrich et al., 2009), their use should be restricted to signaling pathways, mechanistic studies and first-line HTS drug screens. More sophisticated models like organoids could be used to validate drug candidates. Stem cell-derived organoids are important for modeling epithelial tumors. However, the lack of standardization and quality control of stem cell culture are an obstacle for their use in clinical studies. The use of pluripotent stem cells for organoid generation can be hampered by the presence of contaminating progenitors that can yield undesired cell types and a small population of undifferentiated PSCs can give rise to tumors that out-compete organ reconstitution in vivo (Fowler et al., 2020). Furthermore, due to different culture methods, organoids may present undesired phenotypic variabilities. Interestingly, the recent development of microwell arrays in a matrix-free solid manner allowed the high-throughput assessment of homogenous organoids in 3D culture (Brandenberg et al., 2020). The most exciting aspect for organoid use in clinics is the implementation of PDOs for personalized medicine but this requires that pure PDO cultures can be established, which is not always the case. For example, prostate cancer organoids can only be generated from metastases because normal prostate epithelial cells overgrow cancer cells (Gao et al., 2014). Additionally, the majority of organoids derived from intrapulmonary tumors were overgrown by normal airway organoids (Dijkstra et al., 2020), hampering their use for preclinical studies. Nonetheless, evidence of divergence from primary tumors emerged over time with a decreased abundance of populations from the TME coupled with lower expression of immune-related genes in PDOs (Jacob et al., 2020). Future studies are needed to improve this issue and to maintain the immune compartment, notably for relevant testing of immunotherapies in PDO biobanks.
Microfluidics require refined technical innovations to enable scaling up for HTS. Integration of organotypic models, spheroids, organoids or PDOs, with simulated physiology in microfluidic platforms could represent one of the most relevant in vitro models. Two very exciting studies recently reported a near complete body-on-a-chip system. One described an eight organ-chip model linked via vascular endothelial-lined compartments: gut, liver, heart, kidney, lung, heart, brain, blood-brain barrier, and skin (Novak et al., 2020). Using the same approach, intravenously administered cisplatin via an arteriovenous reservoir, provided clinically relevant results when compared to in vivo behavior (Herland et al., 2020). In this regard, the microfluidic field is still maturing, with a need for regulatory guidelines among the scientific community, specifically for the validation of organ-on-a-chip technology for pharmacological drug testing.
Concluding Remarks
Understanding tumors, now considered as heterogenous abnormal organs, is insufficient if the tumor cells are studied individually. Methods that are more inclusive are needed that integrate the cellular, genomic, microenvironmental and spatial features of cancers to be able to understand and overcome their numerous resistance mechanisms. Increasing the complexity of the used models lead to the development of many organotypic cancer models that are physiologically relevant and allow in-depth understanding of the interactions that take place within a tumor. Moreover, future studies are needed to standardize organoid culture methods across the scientific community, as this is very heterogenous at the moment. It is also needed to enhance such cultures by adding stromal and immune compartments to organoid culture to better mimic the tumor microenvironment. This is important because patient-derived organoids represent a very promising approach for personalized medicine, as they retain patient and tumor identity and mirror drug response, thus allowing the use of tailored medicine and avoiding the use of unnecessary treatments. Such organoids, cryo-preserved and collected to form biobanks, should they be available to the scientific community, may replace conventional drug screening assays because they fit the requirements of automated high-throughput screenings. More sophisticated organotypic models, fruits of the collaboration between biologists and engineers, could represent the future of cancer research. Multi-organoid systems also referred to as “body-on-a-chip” will enable the development of biologically complex systems, where organoids derived from different tissues are brought together and allowed to integrate, mimicking organ function and allowing disease modeling.
Author Contributions
OM conceived the review outline. MH wrote the manuscript and made the figures with support from OM. CN and CV contributed to the final manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Sylvie Rodrigues-Ferreira for the discussions and comments. We also thank the Foundation Janssen Horizon for supporting MH’s Ph.D. thesis.
Footnotes
References
Alimperti, S., Mirabella, T., Bajaj, V., Polacheck, W., Pirone, D. M., Duffield, J., et al. (2017). Three-dimensional biomimetic vascular model reveals a RhoA, Rac1, and N-cadherin balance in mural cell–endothelial cell-regulated barrier function. Proc. Natl. Acad. Sci. U.S.A. 114, 8758–8763. doi: 10.1073/pnas.1618333114
Alizadeh, D., Katsanis, E., and Larmonier, N. (2013). The Multifaceted Role of Th17 lymphocytes and their associated cytokines in cancer. Clin. Dev. Immunol. 2013, 1–11. doi: 10.1155/2013/957878
Andrique, L., Recher, G., Alessandri, K., Pujol, N., Feyeux, M., Bon, P., et al. (2019). A model of guided cell self-organization for rapid and spontaneous formation of functional vessels. Sci. Adv. 5:eaau6562. doi: 10.1126/sciadv.aau6562
Asakawa, N., Shimizu, T., Tsuda, Y., Sekiya, S., Sasagawa, T., Yamato, M., et al. (2010). Pre-vascularization of in vitro three-dimensional tissues created by cell sheet engineering. Biomaterials 31, 3903–3909. doi: 10.1016/j.biomaterials.2010.01.105
Avgustinova, A., Iravani, M., Robertson, D., Fearns, A., Gao, Q., Klingbeil, P., et al. (2016). Tumour cell-derived Wnt7a recruits and activates fibroblasts to promote tumour aggressiveness. Nat. Commun. 7:10305.
Barcellos-Hoff, M. H., and Ravani, S. A. (2000). Irradiated mammary gland stroma promotes the expression of tumorigenic potential by unirradiated epithelial cells. Cancer Res. 60, 1254–1260.
Barretina, J., Caponigro, G., Stransky, N., Venkatesan, K., Margolin, A. A., Kim, S., et al. (2012). The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 483, 603–607.
Bartfeld, S., Bayram, T., van de Wetering, M., Huch, M., Begthel, H., Kujala, P., et al. (2015). In vitro expansion of human gastric epithelial stem cells and their responses to bacterial infection. Gastroenterology 148:126-36.e6.
Beckwitt, C. H., Clark, A. M., Wheeler, S., Taylor, D. L., Stolz, D. B., Griffith, L., et al. (2018). Liver ‘organ on a chip’. Exp. Cell Res. 363, 15–25.
Ben-David, U., Ha, G., Tseng, Y.-Y., Greenwald, N. F., Oh, C., Shih, J., et al. (2017). Patient-derived xenografts undergo mouse-specific tumor evolution. Nat. Genet. 49, 1567–1575. doi: 10.1038/ng.3967
Bersini, S., Jeon, J. S., Dubini, G., Arrigoni, C., Chung, S., Charest, J. L., et al. (2014). A microfluidic 3D in vitro model for specificity of breast cancer metastasis to bone. Biomaterials 35, 2454–2461. doi: 10.1016/j.biomaterials.2013.11.050
Bhatia, S. N., and Ingber, D. E. (2014). Microfluidic organs-on-chips. Nat. Biotechnol. 32, 760–772. doi: 10.1038/nbt.2989
Bonnans, C., Chou, J., and Werb, Z. (2014). Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 15, 786–801. doi: 10.1038/nrm3904
Brancato, V., Gioiella, F., Imparato, G., Guarnieri, D., Urciuolo, F., and Netti, P. A. (2018). 3D breast cancer microtissue reveals the role of tumor microenvironment on the transport and efficacy of free-doxorubicin in vitro. Acta Biomaterialia 75, 200–212. doi: 10.1016/j.actbio.2018.05.055
Brandenberg, N., Hoehnel, S., Kuttler, F., Homicsko, K., Ceroni, C., Ringel, T., et al. (2020). High-throughput automated organoid culture via stem-cell aggregation in microcavity arrays. Nat. Biomed. Eng. 4, 863–874. doi: 10.1038/s41551-020-0565-2
Buckley, C. D., Pilling, D., Lord, J. M., Akbar, A. N., Scheel-Toellner, D., and Salmon, M. (2001). Fibroblasts regulate the switch from acute resolving to chronic persistent inflammation. Trends Immunol. 22, 199–204. doi: 10.1016/s1471-4906(01)01863-4
Calandrini, C., Schutgens, F., Oka, R., Margaritis, T., Candelli, T., Mathijsen, L., et al. (2020). An organoid biobank for childhood kidney cancers that captures disease and tissue heterogeneity. Nat. Commun. 11:1310.
Camp, J. G., Sekine, K., Gerber, T., Loeffler-Wirth, H., Binder, H., Gac, M., et al. (2017). Multilineage communication regulates human liver bud development from pluripotency. Nature 546, 533–538. doi: 10.1038/nature22796
Cardin, D. B., Goff, L., Li, C.-I., Shyr, Y., Winkler, C., DeVore, R., et al. (2014). Phase II trial of sorafenib and erlotinib in advanced pancreatic cancer. Cancer Med. 3, 572–579. doi: 10.1002/cam4.208
Chambers, A. F., Groom, A. C., and MacDonald, I. C. (2002). Dissemination, and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2, 563–572. doi: 10.1038/nrc865
Chen, A. A., Underhill, G. H., and Bhatia, S. N. (2010). Multiplexed, high-throughput analysis of 3D microtissue suspensions. Integr. Biol. 2, 517–527. doi: 10.1039/c0ib00054j
Chen, H. J., Wei, Z., Sun, J., Bhattacharya, A., Savage, D. J., Serda, R., et al. (2016). A recellularized human colon model identifies cancer driver genes. Nat. Biotechnol. 34, 845–851. doi: 10.1038/nbt.3586
Chen, M. B., Lamar, J. M., Li, R., Hynes, R. O., and Kamm, R. D. (2016). Elucidation of the roles of tumor integrin β1 in the extravasation stage of the metastasis cascade. Cancer Res. 76:2513. doi: 10.1158/0008-5472.can-15-1325
Chen, M. B., Whisler, J. A., Fröse, J., Yu, C., Shin, Y., and Kamm, R. D. (2017). On-chip human microvasculature assay for visualization and quantification of tumor cell extravasation dynamics. Nat. Protoc. 12, 865–880. doi: 10.1038/nprot.2017.018
Chen, S., Giannakou, A., Wyman, S., Gruzas, J., Golas, J., Zhong, W., et al. (2018). Cancer-associated fibroblasts suppress SOX2-induced dysplasia in a lung squamous cancer coculture. Proc. Natl. Acad. Sci. U.S.A. 115, E11671–E11680.
Choi, J., Cha, Y. J., and Koo, J. S. (2018). Adipocyte biology in breast cancer: from silent bystander to active facilitator. Progr. Lipid Res. 69, 11–20. doi: 10.1016/j.plipres.2017.11.002
Choi, Y., Hyun, E., Seo, J., Blundell, C., Kim, H. C., Lee, E., et al. (2015). A microengineered pathophysiological model of early-stage breast cancer. Lab Chip. 15, 3350–3357. doi: 10.1039/c5lc00514k
Clohessy, J. G., and Pandolfi, P. P. (2015). Mouse hospital and co-clinical trial project—from bench to bedside. Nat. Rev. Clin. Oncol. 12, 491–498. doi: 10.1038/nrclinonc.2015.62
Co, J. Y., Margalef-Català, M., Li, X., Mah, A. T., Kuo, C. J., Monack, D. M., et al. (2019). Controlling epithelial polarity: a human enteroid model for host-pathogen interactions. Cell Rep. 26, 2509–2520.e4. doi: 10.1016/j.celrep.2019.01.108
Coleman, S. J. (2014). Pancreatic cancer organotypics: high throughput, preclinical models for pharmacological agent evaluation. World J. Gastroenterol. 20, 8471–8481. doi: 10.3748/wjg.v20.i26.8471
Condeelis, J., and Pollard, J. W. (2006). Macrophages: obligate partners for tumor cell migration, invasion, and metastasis. Cell 124, 263–266. doi: 10.1016/j.cell.2006.01.007
Conklin, M. W., Eickhoff, J. C., Riching, K. M., Pehlke, C. A., Eliceiri, K. W., Provenzano, P. P., et al. (2011). Aligned collagen is a prognostic signature for survival in human breast carcinoma. Am. J. Pathol. 178, 1221–1232. doi: 10.1016/j.ajpath.2010.11.076
Day, C.-P., Merlino, G., and Van Dyke, T. (2015). Preclinical mouse cancer models: a maze of opportunities and challenges. Cell 163, 39–53. doi: 10.1016/j.cell.2015.08.068
De La Rochere, P., Guil-Luna, S., Decaudin, D., Azar, G., Sidhu, S. S., and Piaggio, E. (2018). Humanized mice for the study of immuno-oncology. Trends Immunol. 39, 748–763. doi: 10.1016/j.it.2018.07.001
Dekkers, J. F., Whittle, J. R., Vaillant, F., Chen, H.-R., Dawson, C., Liu, K., et al. (2020). Modeling Breast Cancer Using CRISPR-Cas9–mediated engineering of human breast organoids. JNCI 112, 540–544. doi: 10.1093/jnci/djz196
del Pozo Martin, Y., Park, D., Ramachandran, A., Ombrato, L., Calvo, F., Chakravarty, P., et al. (2015). Mesenchymal cancer cell-stroma crosstalk promotes niche activation, epithelial reversion, and metastatic colonization. Cell Rep. 13, 2456–2469. doi: 10.1016/j.celrep.2015.11.025
Deng, T., Lyon, C. J., Bergin, S., Caligiuri, M. A., and Hsueh, W. A. (2016). Obesity, inflammation, and cancer. Ann. Rev. Pathol. 11, 421–449.
Dijkstra, K. K., Cattaneo, C. M., Weeber, F., Chalabi, M., van de Haar, J., Fanchi, L. F., et al. (2018). Generation of tumor-reactive T cells by co-culture of peripheral blood lymphocytes and tumor organoids. Cell 174:1586-98.e12.
Dijkstra, K. K., Monkhorst, K., Schipper, L. J., Hartemink, K. J., Smit, E. F., Kaing, S., et al. (2020). Challenges in establishing pure lung cancer organoids limit their utility for personalized medicine. Cell Rep. 31:107588. doi: 10.1016/j.celrep.2020.107588
Dirat, B., Bochet, L., Dabek, M., Daviaud, D., Dauvillier, S., Majed, B., et al. (2011). Cancer-associated adipocytes exhibit an activated phenotype and contribute to breast cancer invasion. Cancer Res. 71, 2455–2465. doi: 10.1158/0008-5472.can-10-3323
Drost, J., van Boxtel, R., Blokzijl, F., Mizutani, T., Sasaki, N., Sasselli, V., et al. (2017). Use of CRISPR-modified human stem cell organoids to study the origin of mutational signatures in cancer. Science 358, 234–238. doi: 10.1126/science.aao3130
Dutta, D., Heo, I., and Clevers, H. (2017). Disease Modeling in Stem Cell-Derived 3D Organoid Systems. Trends Mol. Med. 23, 393–410. doi: 10.1016/j.molmed.2017.02.007
Duval, K., Grover, H., Han, L.-H., Mou, Y., Pegoraro, A. F., Fredberg, J., et al. (2017). Modeling physiological events in 2D vs. 3D cell culture. Physiology 32, 266–277. doi: 10.1152/physiol.00036.2016
Dye, B. R., Hill, D. R., Ferguson, M. A., Tsai, Y.-H., Nagy, M. S., Dyal, R., et al. (2015). In vitro generation of human pluripotent stem cell derived lung organoids. eLife 4:e05098.
Ehsan, S. M., Welch-Reardon, K. M., Waterman, M. L., Hughes, C. C. W., and George, S. C. (2014). A three-dimensional in vitro model of tumor cell intravasation. Integr. Biol. 6, 603–610. doi: 10.1039/c3ib40170g
Erler, J. T., Bennewith, K. L., Cox, T. R., Lang, G., Bird, D., Koong, A., et al. (2009). Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the premetastatic niche. Cancer Cell 15, 35–44. doi: 10.1016/j.ccr.2008.11.012
Etzerodt, A., Moulin, M., Doktor, T. K., Delfini, M., Mossadegh-Keller, N., Bajenoff, M., et al. (2020). Tissue-resident macrophages in omentum promote metastatic spread of ovarian cancer. J. Exp. Med. 217:e20191869.
Fearon, E. R., and Vogelstein, B. (1990). A genetic model for colorectal tumorigenesis. Cell 61, 759–767. doi: 10.1016/0092-8674(90)90186-i
Fernández-Periáñez, R., Molina-Privado, I., Rojo, F., Guijarro-Muñoz, I., Alonso-Camino, V., Zazo, S., et al. (2013). Basement membrane-rich organoids with functional human blood vessels are permissive niches for human breast cancer metastasis. PLoS One 8:e72957. doi: 10.1371/journal.pone.0072957
Ferreira, L. P., Gaspar, V. M., and Mano, J. F. (2018). Design of spherically structured 3D in vitro tumor models -Advances and prospects. Acta Biomaterialia 75, 11–34. doi: 10.1016/j.actbio.2018.05.034
Folkman, J. (1985). “Tumor Angiogenesis,” in Advances in Cancer Research, eds G. Klein and S. Weinhouse (Cambridge: Academic Press), 175–203.
Folkman, J., Watson, K., Ingber, D., and Hanahan, D. (1989). Induction of angiogenesis during the transition from hyperplasia to neoplasia. Nature 339, 58–61. doi: 10.1038/339058a0
Fowler, J. L., Ang, L. T., and Loh, K. M. (2020). A critical look: challenges in differentiating human pluripotent stem cells into desired cell types and organoids. WIREs Dev. Biol. 9:e368.
Fridman, W. H., Pagès, F., Sautès-Fridman, C., and Galon, J. (2012). The immune contexture in human tumours: impact on clinical outcome. Nat. Rev. Cancer 12, 298–306. doi: 10.1038/nrc3245
Friedl, P., Locker, J., Sahai, E., and Segall, J. E. (2012). Classifying collective cancer cell invasion. Nat. Cell Biol. 14, 777–783. doi: 10.1038/ncb2548
Friedrich, J., Seidel, C., Ebner, R., and Kunz-Schughart, L. A. (2009). Spheroid-based drug screen: considerations and practical approach. Nat. Protoc. 4, 309–324. doi: 10.1038/nprot.2008.226
Fuchs, E. (1990). Epidermal differentiation: the bare essentials. J. Cell Biol. 111, 2807–2814. doi: 10.1083/jcb.111.6.2807
Fujii, M., and Sato, T. (2020). Somatic cell-derived organoids as prototypes of human epithelial tissues and diseases. Nat. Mater. doi: 10.1038/s41563-020-0754-0 [Epub ahead of print].
Fujii, M., Shimokawa, M., Date, S., Takano, A., Matano, M., Nanki, K., et al. (2016). A colorectal tumor organoid library demonstrates progressive loss of niche factor requirements during tumorigenesis. Cell Stem Cell 18, 827–838. doi: 10.1016/j.stem.2016.04.003
Fukuda, A., Wang Sam, C., Morris John, P., Folias Alexandra, E., Liou, A., Kim Grace, E., et al. (2011). Stat3 and MMP7 contribute to pancreatic ductal adenocarcinoma initiation and progression. Cancer Cell 19, 441–455. doi: 10.1016/j.ccr.2011.03.002
Fukumura, D., Xavier, R., Sugiura, T., Chen, Y., Park, E.-C., Lu, N., et al. (1998). Tumor Induction of VEGF promoter activity in stromal cells. Cell 94, 715–725. doi: 10.1016/s0092-8674(00)81731-6
Fumagalli, A., Drost, J., Suijkerbuijk, S. J. E., van Boxtel, R., de Ligt, J., Offerhaus, G. J., et al. (2017). Genetic dissection of colorectal cancer progression by orthotopic transplantation of engineered cancer organoids. Proc. Natl. Acad. Sci. U.S.A. 114, E2357–E2364.
Ganesh, K., Wu, C., O’Rourke, K. P., Szeglin, B. C., Zheng, Y., Sauvé, C.-E. G., et al. (2019). A rectal cancer organoid platform to study individual responses to chemoradiation. Nat. Med. 25, 1607–1614.
Gao, D., Vela, I., Sboner, A., Iaquinta Phillip, J., Karthaus Wouter, R., Gopalan, A., et al. (2014). Organoid cultures derived from patients with advanced prostate cancer. Cell 159, 176–187.
Garcez, P. P., Loiola, E. C., Madeiro da Costa, R., Higa, L. M., Trindade, P., Delvecchio, R., et al. (2016). Zika virus impairs growth in human neurospheres and brain organoids. Science 352:40780.
Gatault, S., Delbeke, M., Driss, V., Sarazin, A., Dendooven, A., Kahn, J.-E., et al. (2015). IL-18 Is Involved in Eosinophil-Mediated Tumoricidal Activity against a Colon Carcinoma Cell Line by Upregulating LFA-1 and ICAM-1. J. Immunol. 195, 2483–2492. doi: 10.4049/jimmunol.1402914
Gentles, A. J., Newman, A. M., Liu, C. L., Bratman, S. V., Feng, W., Kim, D., et al. (2015). The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 21, 938–945. doi: 10.1038/nm.3909
Georgess, D., Padmanaban, V., Sirka, O. K., Coutinho, K., Choi, A., Frid, G., et al. (2020). Twist1-induced epithelial dissemination requires Prkd1 signaling. Cancer Res. 80, 204–218. doi: 10.1158/0008-5472.can-18-3241
Gillet, J.-P., Calcagno, A. M., Varma, S., Marino, M., Green, L. J., Vora, M. I., et al. (2011). Redefining the relevance of established cancer cell lines to the study of mechanisms of clinical anti-cancer drug resistance. Proc. Natl. Acad. Sci. U.S.A. 108, 18708–18713. doi: 10.1073/pnas.1111840108
Gjorevski, N., Sachs, N., Manfrin, A., Giger, S., Bragina, M. E., Ordóñez-Morán, P., et al. (2016). Designer matrices for intestinal stem cell and organoid culture. Nature 539, 560–564. doi: 10.1038/nature20168
Gocheva, V., Wang, H.-W., Gadea, B. B., Shree, T., Hunter, K. E., Garfall, A. L., et al. (2010). IL-4 induces cathepsin protease activity in tumor-associated macrophages to promote cancer growth and invasion. Genes Dev. 24, 241–255. doi: 10.1101/gad.1874010
Goswami, S., Sahai, E., Wyckoff, J. B., Cammer, M., Cox, D., Pixley, F. J., et al. (2005). Macrophages Promote the Invasion of Breast Carcinoma Cells via a Colony-Stimulating Factor-1/Epidermal Growth Factor Paracrine Loop. Cancer Res. 65, 5278–5283. doi: 10.1158/0008-5472.can-04-1853
Goto, H., Shimono, Y., Funakoshi, Y., Imamura, Y., Toyoda, M., Kiyota, N., et al. (2019). Adipose-derived stem cells enhance human breast cancer growth and cancer stem cell-like properties through adipsin. Oncogene 38, 767–779. doi: 10.1038/s41388-018-0477-8
Grisaru-Tal, S., Itan, M., Klion, A. D., and Munitz, A. (2020). A new dawn for eosinophils in the tumour microenvironment. Nat. Rev. Cancer 20, 594–607. doi: 10.1038/s41568-020-0283-9
Hammel, P., Huguet, F., van Laethem, J.-L., Goldstein, D., Glimelius, B., Artru, P., et al. (2016). Effect of Chemoradiotherapy vs Chemotherapy on Survival in Patients With Locally Advanced Pancreatic Cancer Controlled After 4 Months of Gemcitabine With or Without Erlotinib: the LAP07 Randomized Clinical Trial. JAMA 315, 1844–1853. doi: 10.1001/jama.2016.4324
Han, K., Pierce, S. E., Li, A., Spees, K., Anderson, G. R., Seoane, J. A., et al. (2020). CRISPR screens in cancer spheroids identify 3D growth-specific vulnerabilities. Nature 580, 136–141. doi: 10.1038/s41586-020-2099-x
Hanahan, D., and Coussens, L. M. (2012). Accessories to the crime: functions of cells recruited to the tumor microenvironment. Cancer Cell 21, 309–322. doi: 10.1016/j.ccr.2012.02.022
Hanahan, D., and Weinberg, R. A. (2011). Hallmarks of cancer: the next generation. Cell 144, 646–674. doi: 10.1016/j.cell.2011.02.013
Harney, A. S., Arwert, E. N., Entenberg, D., Wang, Y., Guo, P., Qian, B.-Z., et al. (2015). Real-time imaging reveals local, transient vascular permeability, and tumor cell intravasation stimulated by TIE2hi macrophage-derived VEGFA. Cancer Discov. 5, 932–943. doi: 10.1158/2159-8290.cd-15-0012
Hassell, B. A., Goyal, G., Lee, E., Sontheimer-Phelps, A., Levy, O., Chen, C. S., et al. (2017). Human organ chip models recapitulate orthotopic lung cancer growth. therapeutic responses, and tumor dormancy in vitro. Cell Rep. 21, 508–516. doi: 10.1016/j.celrep.2017.09.043
Herbst, R. S., Soria, J.-C., Kowanetz, M., Fine, G. D., Hamid, O., Gordon, M. S., et al. (2014). Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature 515, 563–567.
Herland, A., Maoz, B. M., Das, D., Somayaji, M. R., Prantil-Baun, R., Novak, R., et al. (2020). Quantitative prediction of human pharmacokinetic responses to drugs via fluidically coupled vascularized organ chips. Nat. Biomed. Eng. 4, 421–436. doi: 10.1038/s41551-019-0498-9
Heyder, C., Gloria-Maercker, E., Entschladen, F., Hatzmann, W., Niggemann, B., Zänker, K., et al. (2002). Realtime visualization of tumor cell/endothelial cell interactions during transmigration across the endothelial barrier. J. Cancer Res. Clin. Oncol. 128, 533–538. doi: 10.1007/s00432-002-0377-7
Hida, K., Akiyama, K., Ohga, N., Maishi, N., and Hida, Y. (2013). Tumour endothelial cells acquire drug resistance in a tumour microenvironment. J. Biochem. 153, 243–249. doi: 10.1093/jb/mvs152
Hinshaw, D. C., and Shevde, L. A. (2019). The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res. 79, 4557–4566. doi: 10.1158/0008-5472.can-18-3962
Hohwieler, M., Illing, A., Hermann, P. C., Mayer, T., Stockmann, M., Perkhofer, L., et al. (2017). Human pluripotent stem cell-derived acinar/ductal organoids generate human pancreas upon orthotopic transplantation and allow disease modelling. Gut 66, 473–486. doi: 10.1136/gutjnl-2016-312423
Hsu, T.-H., Xiao, J.-L., Tsao, Y.-W., Kao, Y.-L., Huang, S.-H., Liao, W.-Y., et al. (2011). Analysis of the paracrine loop between cancer cells and fibroblasts using a microfluidic chip. Lab Chip. 11, 1808–1814. doi: 10.1039/c1lc20090a
Hu, B., Castillo, E., Harewood, L., Ostano, P., Reymond, A., Dummer, R., et al. (2012). Multifocal epithelial tumors and field cancerization from loss of mesenchymal CSL signaling. Cell 149, 1207–1220. doi: 10.1016/j.cell.2012.03.048
Huang, A., Garraway, L. A., Ashworth, A., and Weber, B. (2020). Synthetic lethality as an engine for cancer drug. (target). discovery. Nat. Rev. Drug Discov. 19, 23–38. doi: 10.1038/s41573-019-0046-z
Huang, L., Bockorny, B., Paul, I., Akshinthala, D., Frappart, P.-O., Gandarilla, O., et al. (2020). PDX-derived organoids model in vivo drug response and secrete biomarkers. JCI Insight 5:e135544.
Huang, S., Boda, B., Vernaz, J., Ferreira, E., Wiszniewski, L., and Constant, S. (2017). Establishment and characterization of an in vitro human small airway model (SmallAirTM). Eur. J. Pharmaceut. Biopharmaceut. 118, 68–72. doi: 10.1016/j.ejpb.2016.12.006
Huang, S.-M., Lin, C., Lin, H.-Y., Chiu, C.-M., Fang, C.-W., Liao, K.-F., et al. (2015). Brain-derived neurotrophic factor regulates cell motility in human colon cancer. Endocrine Relat. Cancer 22, 455–464. doi: 10.1530/erc-15-0007
Huh, D., Leslie, D. C., Matthews, B. D., Fraser, J. P., Jurek, S., Hamilton, G. A., et al. (2012). A human disease model of drug toxicity–induced pulmonary edema in a lung-on-a-chip microdevice. Sci. Transl. Med. 4:159ra147. doi: 10.1126/scitranslmed.3004249
Huh, D., Matthews, B. D., Mammoto, A., Montoya-Zavala, M., Hsin, H. Y., and Ingber, D. E. (2010). Reconstituting organ-level lung functions on a chip. Science 328, 1662–1668. doi: 10.1126/science.1188302
Hynes, R. O., and Naba, A. (2012). Overview of the matrisome–an inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 4:a004903. doi: 10.1101/cshperspect.a004903
Jacob, F., Salinas, R. D., Zhang, D. Y., Nguyen, P. T. T., Schnoll, J. G., Wong, S. Z. H., et al. (2020). A patient-derived glioblastoma organoid model and biobank recapitulates inter- and intra-tumoral heterogeneity. Cell 180, 188-204.e22.
Jang, K.-J., Mehr, A. P., Hamilton, G. A., McPartlin, L. A., Chung, S., Suh, K.-Y., et al. (2013). Human kidney proximal tubule-on-a-chip for drug transport and nephrotoxicity assessment. Integr. Biol. 5, 1119–1129. doi: 10.1039/c3ib40049b
Jenkins, R. W., Aref, A. R., Lizotte, P. H., Ivanova, E., Stinson, S., Zhou, C. W., et al. (2018). Ex Vivo Profiling of PD-1 blockade using organotypic tumor spheroids. Cancer Discov. 8, 196–215.
Jeon, J. S., Bersini, S., Gilardi, M., Dubini, G., Charest, J. L., Moretti, M., et al. (2015). Human 3D vascularized organotypic microfluidic assays to study breast cancer cell extravasation. Proc. Natl. Acad. Sci. U.S.A. 112, 214–219. doi: 10.1073/pnas.1417115112
Jeon, J. S., Bersini, S., Whisler, J. A., Chen, M. B., Dubini, G., Charest, J. L., et al. (2014). Generation of 3D functional microvascular networks with human mesenchymal stem cells in microfluidic systems. Integr. Biol. 6, 555–563. doi: 10.1039/c3ib40267c
Jia, X., Gábris, F., Jacobsen, Ó, Bedics, G., Botz, B., Helyes, Z., et al. (2020). Foliate Lymphoid Aggregates as Novel Forms of Serous Lymphocyte Entry Sites of Peritoneal B Cells and High-Grade B Cell Lymphomas. J. Immunol. 204, 23–36. doi: 10.4049/jimmunol.1900851
Jobling, P., Pundavela, J., Oliveira, S. M. R., Roselli, S., Walker, M. M., and Hondermarck, H. (2015). Nerve-cancer cell cross-talk: a novel promoter of tumor progression. Cancer Res. 75, 1777–1781. doi: 10.1158/0008-5472.can-14-3180
Jung, H.-Y., Fattet, L., Tsai, J. H., Kajimoto, T., Chang, Q., Newton, A. C., et al. (2019). Apical–basal polarity inhibits epithelial–mesenchymal transition and tumour metastasis by PAR-complex-mediated SNAI1 degradation. Nat. Cell Biol. 21, 359–371. doi: 10.1038/s41556-019-0291-8
Junttila, M. R., and de Sauvage, F. J. (2013). Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 501, 346–354. doi: 10.1038/nature12626
Kalabis, J., Wong, G. S., Vega, M. E., Natsuizaka, M., Robertson, E. S., Herlyn, M., et al. (2012). Isolation and characterization of mouse and human esophageal epithelial cells in 3D organotypic culture. Nat. Protoc. 7, 235–246. doi: 10.1038/nprot.2011.437
Kalluri, R. (2016). The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 16, 582–598. doi: 10.1038/nrc.2016.73
Kaplan, R. N., Riba, R. D., Zacharoulis, S., Bramley, A. H., Vincent, L., Costa, C., et al. (2005). VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche. Nature 438, 820–827. doi: 10.1038/nature04186
Kessler, M., Hoffmann, K., Brinkmann, V., Thieck, O., Jackisch, S., Toelle, B., et al. (2015). The Notch and Wnt pathways regulate stemness and differentiation in human fallopian tube organoids. Nat. Commun. 6:8989.
Kim, K., Ohashi, K., Utoh, R., Kano, K., and Okano, T. (2012). Preserved liver-specific functions of hepatocytes in 3D co-culture with endothelial cell sheets. Biomaterials 33, 1406–1413. doi: 10.1016/j.biomaterials.2011.10.084
Koh, I., Cha, J., Park, J., Choi, J., Kang, S.-G., and Kim, P. (2018). The mode and dynamics of glioblastoma cell invasion into a decellularized tissue-derived extracellular matrix-based three-dimensional tumor model. Sci. Rep. 8:4608.
Kumar, V., Patel, S., Tcyganov, E., and Gabrilovich, D. I. (2016). The nature of myeloid-derived suppressor cells in the tumor microenvironment. Trends Immunol. 37, 208–220.
Lancaster, M. A., Corsini, N. S., Wolfinger, S., Gustafson, E. H., Phillips, A. W., Burkard, T. R., et al. (2017). Guided self-organization and cortical plate formation in human brain organoids. Nat. Biotechnol. 35, 659–666. doi: 10.1038/nbt.3906
Lancaster, M. A., and Knoblich, J. A. (2014). Organogenesis in a dish: modeling development and disease using organoid technologies. Science 345:1247125. doi: 10.1126/science.1247125
Langhans, S. A. (2018). Three-dimensional in vitro cell culture models in drug discovery and drug repositioning. Front. Pharmacol. 9:6. doi: 10.3389/fphar.2018.00006
Lee, E. Y., Parry, G., and Bissell, M. J. (1984). Modulation of secreted proteins of mouse mammary epithelial cells by the collagenous substrata. J. Cell Biol. 98, 146–155. doi: 10.1083/jcb.98.1.146
Lee, M.-Y., Kumar, R. A., Sukumaran, S. M., Hogg, M. G., Clark, D. S., and Dordick, J. S. (2008). Three-dimensional cellular microarray for high-throughput toxicology assays. Proc. Natl. Acad. Sci. U.S.A. 105, 59–63. doi: 10.1073/pnas.0708756105
Leslie, J. L., Huang, S., Opp, J. S., Nagy, M. S., Kobayashi, M., Young, V. B., et al. (2015). Persistence and toxin production by clostridium difficile within human intestinal organoids result in disruption of epithelial paracellular barrier function. Infect. Immunity 83, 138–145. doi: 10.1128/iai.02561-14
Li, P., Lu, M., Shi, J., Gong, Z., Hua, L., Li, Q., et al. (2020). Lung mesenchymal cells elicit lipid storage in neutrophils that fuel breast cancer lung metastasis. Nat. Immunol. 21, 1444–1455. doi: 10.1038/s41590-020-0783-5
Li, R., Hebert, J. D., Lee, T. A., Xing, H., Boussommier-Calleja, A., Hynes, R. O., et al. (2017). Macrophage-Secreted TNFα and TGFβ1 influence migration speed and persistence of cancer cells in 3D tissue culture via independent pathways. Cancer Res. 77, 279–290. doi: 10.1158/0008-5472.can-16-0442
Li, Y., and Kumacheva, E. (2018). Hydrogel microenvironments for cancer spheroid growth and drug screening. Sci. Adv. 4:eaas8998. doi: 10.1126/sciadv.aas8998
Libanje, F., Raingeaud, J., Luan, R., Thomas, Z., Zajac, O., Veiga, J., et al. (2019). ROCK2 inhibition triggers the collective invasion of colorectal adenocarcinomas. EMBO J. 38:e99299.
Lindau, D., Gielen, P., Kroesen, M., Wesseling, P., and Adema, G. J. (2013). The immunosuppressive tumour network: myeloid-derived suppressor cells, regulatory T cells and natural killer T cells. Immunology 138, 105–115. doi: 10.1111/imm.12036
Liu, T., Han, C., Wang, S., Fang, P., Ma, Z., Xu, L., et al. (2019). Cancer-associated fibroblasts: an emerging target of anti-cancer immunotherapy. J. Hematol. Oncol. 12:86.
Liu, Y., and Cao, X. (2016). Characteristics and significance of the pre-metastatic niche. Cancer Cell 30, 668–681. doi: 10.1016/j.ccell.2016.09.011
Lu, Z., Liang, J., He, Q., Wan, Q., Hou, J., Lian, K., et al. (2019). The serum biomarker chemerin promotes tumorigenesis and metastasis in oral squamous cell carcinoma. Clin. Sci. 133, 681–695. doi: 10.1042/cs20181023
Lucarini, V., Ziccheddu, G., Macchia, I., La Sorsa, V., Peschiaroli, F., Buccione, C., et al. (2017). IL-33 restricts tumor growth and inhibits pulmonary metastasis in melanoma-bearing mice through eosinophils. OncoImmunology 6:e1317420. doi: 10.1080/2162402x.2017.1317420
Madeo, M., Colbert, P. L., Vermeer, D. W., Lucido, C. T., Cain, J. T., Vichaya, E. G., et al. (2018). Cancer exosomes induce tumor innervation. Nat. Commun. 9:4284.
Magnon, C., Hall, S. J., Lin, J., Xue, X., Gerber, L., Freedland, S. J., et al. (2015). Autonomic nerve development contributes to prostate cancer progression. Science 341:1236361. doi: 10.1126/science.1236361
Mani, S. A., Guo, W., Liao, M.-J., Eaton, E. N., Ayyanan, A., Zhou, A. Y., et al. (2008). The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 133, 704–715.
Mantovani, A., and Allavena, P. (2015). The interaction of anticancer therapies with tumor-associated macrophages. J. Exp. Med. 212, 435–445. doi: 10.1084/jem.20150295
Mantovani, A., Allavena, P., Sica, A., and Balkwill, F. (2008). Cancer-related inflammation. Nature 454, 436–444.
Martin, O. C. B., Bergonzini, A., D’Amico, F., Chen, P., Shay, J. W., Dupuy, J., et al. (2019). Infection with genotoxin-producing Salmonella enterica synergises with loss of the tumour suppressor APC in promoting genomic instability via the PI3K pathway in colonic epithelial cells. Cell. Microbiol. 21:e13099. doi: 10.1111/cmi.13099
Matano, M., Date, S., Shimokawa, M., Takano, A., Fujii, M., Ohta, Y., et al. (2015). Modeling colorectal cancer using CRISPR-Cas9–mediated engineering of human intestinal organoids. Nat. Med. 21, 256–262. doi: 10.1038/nm.3802
Mazio, C., Casale, C., Imparato, G., Urciuolo, F., and Netti, P. A. (2018). Recapitulating spatiotemporal tumor heterogeneity in vitro through engineered breast cancer microtissues. Acta Biomaterialia 73, 236–249. doi: 10.1016/j.actbio.2018.04.028
McAleer, C. W., Long, C. J., Elbrecht, D., Sasserath, T., Bridges, L. R., Rumsey, J. W., et al. (2019). Multi-organ system for the evaluation of efficacy and off-target toxicity of anticancer therapeutics. Sci. Transl. Med. 11:eaav1386. doi: 10.1126/scitranslmed.aav1386
McAllister, S. S., and Weinberg, R. A. (2014). The tumour-induced systemic environment as a critical regulator of cancer progression and metastasis. Nat. Cell Biol. 16, 717–727. doi: 10.1038/ncb3015
McCracken, K. W., Catá, E. M., Crawford, C. M., Sinagoga, K. L., Schumacher, M., Rockich, B. E., et al. (2014). Modelling human development and disease in pluripotent stem-cell-derived gastric organoids. Nature 516, 400–404.
McFadden, D. G., Politi, K., Bhutkar, A., Chen, F. K., Song, X., Pirun, M., et al. (2016). Mutational landscape of EGFR-, MYC-, and Kras-driven genetically engineered mouse models of lung adenocarcinoma. Proc. Natl. Acad. Sci. U.S.A. 113:E6409.
Mehlen, P., and Puisieux, A. (2006). Metastasis: a question of life or death. Nat. Rev. Cancer 6, 449–458. doi: 10.1038/nrc1886
Merlos-Suárez, A., Barriga Francisco, M., Jung, P., Iglesias, M., Céspedes María, V., Rossell, D., et al. (2011). The intestinal stem cell signature identifies colorectal cancer stem cells and predicts disease relapse. Cell Stem Cell 8, 511–524. doi: 10.1016/j.stem.2011.02.020
Mhaidly, R., and Mechta-Grigoriou, F. (2020). Fibroblast heterogeneity in tumor micro-environment: role in immunosuppression and new therapies. Semin. Immunol. [Online ahead of print] doi: 10.1016/j.smim.2020.101417
Michels, B. E., Mosa, M. H., Streibl, B. I., Zhan, T., Menche, C., Abou-El-Ardat, K., et al. (2020). Pooled In Vitro and In Vivo CRISPR-Cas9 screening identifies tumor suppressors in human colon organoids. Cell Stem Cell 26, 782-92.e7.
Middleton, G., Fletcher, P., Popat, S., Savage, J., Summers, Y., Greystoke, A., et al. (2020). The National Lung Matrix Trial of personalized therapy in lung cancer. Nature 583, 807–812.
Nam, S. H., Kim, D., Lee, D., Lee, H.-M., Song, D.-G., Jung, J. W., et al. (2018). Lysyl-tRNA synthetase–expressing colon spheroids induce M2 macrophage polarization to promote metastasis. J. Clin. Invest. 128, 5034–5055. doi: 10.1172/jci99806
Namekawa, T., Ikeda, K., Horie-Inoue, K., Suzuki, T., Okamoto, K., Ichikawa, T., et al. (2020). ALDH1A1 in patient-derived bladder cancer spheroids activates retinoic acid signaling leading to TUBB3 overexpression and tumor progression. Int. J. Cancer 146, 1099–1113. doi: 10.1002/ijc.32505
Nashimoto, Y., Okada, R., Hanada, S., Arima, Y., Nishiyama, K., Miura, T., et al. (2020). Vascularized cancer on a chip: the effect of perfusion on growth and drug delivery of tumor spheroid. Biomaterials 229:119547. doi: 10.1016/j.biomaterials.2019.119547
Neal, J. T., Li, X., Zhu, J., Giangarra, V., Grzeskowiak, C. L., Ju, J., et al. (2018). Organoid modeling of the tumor immune microenvironment. Cell 175, 1972-88.e16.
Nguyen, M., De Ninno, A., Mencattini, A., Mermet-Meillon, F., Fornabaio, G., Evans, S. S., et al. (2018). Dissecting effects of anti-cancer drugs and cancer-associated fibroblasts by on-chip reconstitution of immunocompetent tumor microenvironments. Cell Rep. 25, 3884-93.e3.
Nombela-Arrieta, C., Ritz, J., and Silberstein, L. E. (2011). The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol. 12, 126–131. doi: 10.1038/nrm3049
Novak, R., Ingram, M., Marquez, S., Das, D., Delahanty, A., Herland, A., et al. (2020). Robotic fluidic coupling and interrogation of multiple vascularized organ chips. Nat. Biomed. Eng. 4, 407–420. doi: 10.1038/s41551-019-0497-x
Nowell, P. (1976). The clonal evolution of tumor cell populations. Science 194, 23–28. doi: 10.1126/science.959840
Noy, R., and Pollard Jeffrey, W. (2014). Tumor-associated macrophages: from mechanisms to therapy. Immunity 41, 49–61. doi: 10.1016/j.immuni.2014.06.010
Öhlund, D., Handly-Santana, A., Biffi, G., Elyada, E., Almeida, A. S., Ponz-Sarvise, M., et al. (2017). Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J. Exp. Med. 214, 579–596. doi: 10.1084/jem.20162024
Ooft, S. N., Weeber, F., Dijkstra, K. K., McLean, C. M., Kaing, S., van Werkhoven, E., et al. (2019). Patient-derived organoids can predict response to chemotherapy in metastatic colorectal cancer patients. Sci. Transl. Med. 11:eaay2574.
Ortiz, M. L., Kumar, V., Martner, A., Mony, S., Donthireddy, L., Condamine, T., et al. (2015). Immature myeloid cells directly contribute to skin tumor development by recruiting IL-17–producing CD4+ T cells. J. Exp. Med. 212, 351–367. doi: 10.1084/jem.20140835
Palikuqi, B., Nguyen, D.-H. T., Li, G., Schreiner, R., Pellegata, A. F., Liu, Y., et al. (2020). Adaptable haemodynamic endothelial cells for organogenesis and tumorigenesis. Nature 585, 426–432. doi: 10.1038/s41586-020-2712-z
Palumbo, J. S., Talmage, K. E., Massari, J. V., La Jeunesse, C. M., Flick, M. J., Kombrinck, K. W., et al. (2005). Platelets and fibrin(ogen) increase metastatic potential by impeding natural killer cell–mediated elimination of tumor cells. Blood 105, 178–185. doi: 10.1182/blood-2004-06-2272
Papetti, M., and Herman, I. M. (2002). Mechanisms of normal and tumor-derived angiogenesis. Am. J. Physiol. Cell Physiol. 282, C947–C970.
Park, J., Morley, T. S., Kim, M., Clegg, D. J., and Scherer, P. E. (2014). Obesity and cancer—mechanisms underlying tumour progression and recurrence. Nat. Rev. Endocrinol. 10, 455–465. doi: 10.1038/nrendo.2014.94
Pasch, C. A., Favreau, P. F., Yueh, A. E., Babiarz, C. P., Gillette, A. A., Sharick, J. T., et al. (2019). Patient-derived cancer organoid cultures to predict sensitivity to chemotherapy and radiation. Clin. Cancer Res. 25, 5376–5387. doi: 10.1158/1078-0432.ccr-18-3590
Pauli, C., Hopkins, B. D., Prandi, D., Shaw, R., Fedrizzi, T., Sboner, A., et al. (2017). Personalized in vitro and in vivo cancer models to guide precision medicine. Cancer Discov. 7, 462–477.
Pelon, F., Bourachot, B., Kieffer, Y., Magagna, I., Mermet-Meillon, F., Bonnet, I., et al. (2020). Cancer-associated fibroblast heterogeneity in axillary lymph nodes drives metastases in breast cancer through complementary mechanisms. Nat. Commun. 11:404.
Plaks, V., Kong, N., and Werb, Z. (2015). The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell Stem Cell 16, 225–238. doi: 10.1016/j.stem.2015.02.015
Polacheck, W. J., Charest, J. L., and Kamm, R. D. (2011). Interstitial flow influences direction of tumor cell migration through competing mechanisms. Proc. Natl. Acad. Sci. U.S.A. 108, 11115–11120. doi: 10.1073/pnas.1103581108
Prasad, V., Fojo, T., and Brada, M. (2016). Precision oncology: origins, optimism, and potential. Lancet Oncol. 17, e81–e86.
Puca, L., Bareja, R., Prandi, D., Shaw, R., Benelli, M., Karthaus, W. R., et al. (2018). Patient derived organoids to model rare prostate cancer phenotypes. Nat. Commun. 9:2404.
Reymond, N., d’Água, B. B., and Ridley, A. J. (2013). Crossing the endothelial barrier during metastasis. Nat. Rev. Cancer 13, 858–870. doi: 10.1038/nrc3628
Richards, Z., McCray, T., Marsili, J., Zenner, M. L., Manlucu, J. T., Garcia, J., et al. (2019). Prostate stroma increases the viability and maintains the branching phenotype of human prostate organoids. iScience 12, 304–317. doi: 10.1016/j.isci.2019.01.028
Ridky, T. W., Chow, J. M., Wong, D. J., and Khavari, P. A. (2010). Invasive three-dimensional organotypic neoplasia from multiple normal human epithelia. Nat. Med. 16, 1450–1455. doi: 10.1038/nm.2265
Roulis, M., Kaklamanos, A., Schernthanner, M., Bielecki, P., Zhao, J., Kaffe, E., et al. (2020). Paracrine orchestration of intestinal tumorigenesis by a mesenchymal niche. Nature 580, 524–529. doi: 10.1038/s41586-020-2166-3
Ruffell, B., and Coussens Lisa, M. (2015). Macrophages and therapeutic resistance in cancer. Cancer Cell 27, 462–472. doi: 10.1016/j.ccell.2015.02.015
Sano, E., Mori, C., Nashimoto, Y., Yokokawa, R., Kotera, H., and Torisawa, Y-s. (2018). Engineering of vascularized 3D cell constructs to model cellular interactions through a vascular network. Biomicrofluidics 12:042204. doi: 10.1063/1.5027183
Sato, T., Vries, R. G., Snippert, H. J., van de Wetering, M., Barker, N., Stange, D. E., et al. (2009). Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459, 262–265. doi: 10.1038/nature07935
Schumacher, D., Strilic, B., Sivaraj Kishor, K., Wettschureck, N., and Offermanns, S. (2013). Platelet-derived nucleotides promote tumor-cell transendothelial migration and metastasis via P2Y2 receptor. Cancer Cell 24, 130–137. doi: 10.1016/j.ccr.2013.05.008
Seano, G., Chiaverina, G., Gagliardi, P. A., di Blasio, L., Sessa, R., Bussolino, F., et al. (2013). Modeling human tumor angiogenesis in a three-dimensional culture system. Blood 121, e129–e137.
Shirure, V. S., Bi, Y., Curtis, M. B., Lezia, A., Goedegebuure, M. M., Goedegebuure, S. P., et al. (2018). Tumor-on-a-chip platform to investigate progression and drug sensitivity in cell lines and patient-derived organoids. Lab Chip. 18, 3687–3702. doi: 10.1039/c8lc00596f
Shultz, L. D., Brehm, M. A., Garcia-Martinez, J. V., and Greiner, D. L. (2012). Humanized mice for immune system investigation: progress, promise and challenges. Nat. Rev. Immunol. 12, 786–798. doi: 10.1038/nri3311
Silvestri, V. L., Henriet, E., Linville, R. M., Wong, A. D., Searson, P. C., and Ewald, A. J. (2020). A tissue-engineered 3D microvessel model reveals the dynamics of mosaic vessel formation in breast cancer. Cancer Res. 80, 4288–4301. doi: 10.1158/0008-5472.can-19-1564
Smith, S. C., Baras, A. S., Lee, J. K., and Theodorescu, D. (2010). The COXEN Principle: translating signatures of in vitro chemosensitivity into tools for clinical outcome prediction and drug discovery in cancer. Cancer Res. 70, 1753–1758. doi: 10.1158/0008-5472.can-09-3562
Sood, D., Tang-Schomer, M., Pouli, D., Mizzoni, C., Raia, N., Tai, A., et al. (2019). 3D extracellular matrix microenvironment in bioengineered tissue models of primary pediatric and adult brain tumors. Nat. Commun. 10:4529.
Spence, J. R., Mayhew, C. N., Rankin, S. A., Kuhar, M. F., Vallance, J. E., Tolle, K., et al. (2011). Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature 470, 105–109.
Steele, C. W., Karim, S. A., Leach, J. D. G., Bailey, P., Upstill-Goddard, R., Rishi, L., et al. (2016). CXCR2 inhibition profoundly suppresses metastases and augments immunotherapy in pancreatic ductal adenocarcinoma. Cancer Cell 29, 832–845. doi: 10.1016/j.ccell.2016.04.014
Strilic, B., Yang, L., Albarrán-Juárez, J., Wachsmuth, L., Han, K., Müller, U. C., et al. (2016). Tumour-cell-induced endothelial cell necroptosis via death receptor 6 promotes metastasis. Nature 536, 215–218. doi: 10.1038/nature19076
Takasato, M., Er, P. X., Chiu, H. S., Maier, B., Baillie, G. J., Ferguson, C., et al. (2015). Kidney organoids from human iPS cells contain multiple lineages and model human nephrogenesis. Nature 526, 564–568. doi: 10.1038/nature15695
Takeda, H., Kataoka, S., Nakayama, M., Ali, M. A. E., Oshima, H., Yamamoto, D., et al. (2019). CRISPR-Cas9–mediated gene knockout in intestinal tumor organoids provides functional validation for colorectal cancer driver genes. Proc. Natl. Acad. Sci. U.S.A. 116, 15635–15644. doi: 10.1073/pnas.1904714116
Talmadge, J. E., and Gabrilovich, D. I. (2013). History of myeloid-derived suppressor cells. Nat. Rev. Cancer 13, 739–752.
Tang, Y., Soroush, F., Sheffield, J. B., Wang, B., Prabhakarpandian, B., and Kiani, M. F. (2017). A biomimetic microfluidic tumor microenvironment platform mimicking the EPR effect for rapid screening of drug delivery systems. Sci. Rep. 7:9359.
Taniguchi, K., and Karin, M. (2018). NF-κB, inflammation, immunity and cancer: coming of age. Nat. Rev. Immunol. 18, 309–324. doi: 10.1038/nri.2017.142
Taniguchi, S., Elhance, A., Duzer, A. V., Kumar, S., Leitenberger, J. J., and Oshimori, N. (2020). Tumor-initiating cells establish an IL-33–TGF-b niche signaling loop to promote cancer progression. Science 369:eaay1813.
Tanner, K., and Gottesman, M. M. (2015). Beyond 3D culture models of cancer. Sci. Transl. Med. 7:283s9.
Tiriac, H., Belleau, P., Engle, D. D., Plenker, D., Deschênes, A., Somerville, T. D. D., et al. (2018). Organoid profiling identifies common responders to chemotherapy in pancreatic cancer. Cancer Discov. 8, 1112–1129.
Tomasek, J. J., Gabbiani, G., Hinz, B., Chaponnier, C., and Brown, R. A. (2002). Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 3, 349–363. doi: 10.1038/nrm809
Truong, D. D., Kratz, A., Park, J. G., Barrientos, E. S., Saini, H., Nguyen, T., et al. (2019). A Human organotypic microfluidic tumor model permits investigation of the interplay between patient-derived fibroblasts and breast cancer cells. Cancer Res. 79, 3139–3151. doi: 10.1158/0008-5472.can-18-2293
van de Wetering, M., Francies Hayley, E., Francis Joshua, M., Bounova, G., Iorio, F., Pronk, A., et al. (2015). Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell 161, 933–945. doi: 10.1016/j.cell.2015.03.053
van Duinen, V., Trietsch, S. J., Joore, J., Vulto, P., and Hankemeier, T. (2015). Microfluidic 3D cell culture: from tools to tissue models. Curr. Opin. Biotechnol. 35, 118–126. doi: 10.1016/j.copbio.2015.05.002
van Zijl, F., Mair, M., Csiszar, A., Schneller, D., Zulehner, G., Huber, H., et al. (2009). Hepatic tumor–stroma crosstalk guides epithelial to mesenchymal transition at the tumor edge. Oncogene 28, 4022–4033. doi: 10.1038/onc.2009.253
Vlachogiannis, G., Hedayat, S., Vatsiou, A., Jamin, Y., Fernández-Mateos, J., Khan, K., et al. (2018). Patient-derived organoids model treatment response of metastatic gastrointestinal cancers. Science 359, 920–926.
Wang, D., Sun, H., Wei, J., Cen, B., and DuBois, R. N. (2017). CXCL1 is critical for premetastatic niche formation and metastasis in colorectal cancer. Cancer Res. 77, 3655–3665. doi: 10.1158/0008-5472.can-16-3199
Wang, K., Demir, I. E., D’Haese, J. G., Tieftrunk, E., Kujundzic, K., Schorn, S., et al. (2014). The neurotrophic factor neurturin contributes toward an aggressive cancer cell phenotype, neuropathic pain and neuronal plasticity in pancreatic cancer. Carcinogenesis 35, 103–113. doi: 10.1093/carcin/bgt312
Ware, M. J., Keshishian, V., Law, J. J., Ho, J. C., Favela, C. A., Rees, P., et al. (2016). Generation of an in vitro 3D PDAC stroma rich spheroid model. Biomaterials 108, 129–142. doi: 10.1016/j.biomaterials.2016.08.041
Weeber, F., van de Wetering, M., Hoogstraat, M., Dijkstra, K. K., Krijgsman, O., Kuilman, T., et al. (2015). Preserved genetic diversity in organoids cultured from biopsies of human colorectal cancer metastases. Proc. Natl. Acad. Sci. U.S.A. 112, 13308–13311. doi: 10.1073/pnas.1516689112
Weis, S. M., and Cheresh, D. A. (2011). Tumor angiogenesis: molecular pathways and therapeutic targets. Nat. Med. 17, 1359–1370. doi: 10.1038/nm.2537
Wilson, C. L., Jurk, D., Fullard, N., Banks, P., Page, A., Luli, S., et al. (2015). NFκB1 is a suppressor of neutrophil-driven hepatocellular carcinoma. Nat. Commun. 6:6818.
Wisdom, K. M., Adebowale, K., Chang, J., Lee, J. Y., Nam, S., Desai, R., et al. (2018). Matrix mechanical plasticity regulates cancer cell migration through confining microenvironments. Nat. Commun. 9:4144.
Wörsdörfer, P., Dalda, N., Kern, A., Krüger, S., Wagner, N., Kwok, C. K., et al. (2019). Generation of complex human organoid models including vascular networks by incorporation of mesodermal progenitor cells. Sci. Rep. 9:15663.
Xiong, G., Flynn, T. J., Chen, J., Trinkle, C., and Xu, R. (2015). Development of an ex vivo breast cancer lung colonization model utilizing a decellularized lung matrix. Integr. Biol. 7, 1518–1525. doi: 10.1039/c5ib00157a
Yamada, K. M., and Cukierman, E. (2007). Modeling tissue morphogenesis and cancer in 3D. Cell 130, 601–610. doi: 10.1016/j.cell.2007.08.006
Yin, M., Li, X., Tan, S., Zhou, H. J., Ji, W., Bellone, S., et al. (2016). Tumor-associated macrophages drive spheroid formation during early transcoelomic metastasis of ovarian cancer. J. Clin. Invest. 126, 4157–4173. doi: 10.1172/jci87252
Zahalka, A. H., Arnal-Estapé, A., Maryanovich, M., Nakahara, F., Cruz, C. D., Finley, L. W. S., et al. (2017). Adrenergic nerves activate an angio-metabolic switch in prostate cancer. Science 358, 321–326. doi: 10.1126/science.aah5072
Zervantonakis, I. K., Hughes-Alford, S. K., Charest, J. L., Condeelis, J. S., Gertler, F. B., and Kamm, R. D. (2012). Three-dimensional microfluidic model for tumor cell intravasation and endothelial barrier function. Proc. Natl. Acad. Sci. U.S.A. 109, 13515–13520. doi: 10.1073/pnas.1210182109
Zhang, J., Goliwas, K. F., Wang, W., Taufalele, P. V., Bordeleau, F., and Reinhart-King, C. A. (2019). Energetic regulation of coordinated leader–follower dynamics during collective invasion of breast cancer cells. Proc. Natl. Acad. Sci. U.S.A. 116, 7867–7872. doi: 10.1073/pnas.1809964116
Zhang, N., Zhang, W.-J., Cai, H.-Q., Liu, H.-L., Peng, L., Li, C.-H., et al. (2011). Platelet adhesion and fusion to endothelial cell facilitate the metastasis of tumor cell in hypoxia-reoxygenation condition. Clin. Exp. Metastasis 28, 1–12. doi: 10.1007/s10585-010-9353-9
Zhang, Y., Daquinag, A. C., Amaya-Manzanares, F., Sirin, O., Tseng, C., and Kolonin, M. G. (2012). Stromal progenitor cells from endogenous adipose tissue contribute to pericytes and adipocytes that populate the tumor microenvironment. Cancer Res. 72, 5198–5208. doi: 10.1158/0008-5472.can-12-0294
Keywords: tumor microenvironment, cancer, 3D model, therapies, metastasis, tumor dissemination
Citation: Haykal MM, Nahmias C, Varon C and Martin OCB (2020) Organotypic Modeling of the Tumor Landscape. Front. Cell Dev. Biol. 8:606039. doi: 10.3389/fcell.2020.606039
Received: 14 September 2020; Accepted: 03 November 2020;
Published: 24 November 2020.
Edited by:
Hasan Korkaya, Augusta University, United StatesReviewed by:
David Barbie, Dana–Farber Cancer Institute, United StatesJianyu Rao, University of California, Los Angeles, United States
Copyright © 2020 Haykal, Nahmias, Varon and Martin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Océane C. B. Martin, b2NlYW5lLm1hcnRpbkB1LWJvcmRlYXV4LmZy