
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Cell Dev. Biol., 31 January 2020
Sec. Molecular and Cellular Pathology
Volume 8 - 2020 | https://doi.org/10.3389/fcell.2020.00025
This article is part of the Research TopicPerspectives in Mammary Gland Development and Breast Cancer ResearchView all 13 articles
 Emma H. van Schie1
Emma H. van Schie1 Renée van Amerongen2*
Renée van Amerongen2*WNT signaling is crucial for tissue morphogenesis during development in all multicellular animals. After birth, WNT/CTNNB1 responsive stem cells are responsible for tissue homeostasis in various organs and hyperactive WNT/CTNNB1 signaling is observed in many different human cancers. The first link between WNT signaling and breast cancer was established almost 40 years ago, when Wnt1 was identified as a proto-oncogene capable of driving mammary tumor formation in mice. Since that discovery, there has been a dedicated search for aberrant WNT signaling in human breast cancer. However, much debate and controversy persist regarding the importance of WNT signaling for the initiation, progression or maintenance of different breast cancer subtypes. As the first drugs designed to block functional WNT signaling have entered clinical trials, many questions about the role of aberrant WNT signaling in human breast cancer remain. Here, we discuss three major research gaps in this area. First, we still lack a basic understanding of the function of WNT signaling in normal human breast development and physiology. Second, the overall extent and precise effect of (epi)genetic changes affecting the WNT pathway in different breast cancer subtypes are still unknown. Which underlying molecular and cell biological mechanisms are disrupted as a result also awaits further scrutiny. Third, we survey the current status of targeted therapeutics that are aimed at interfering with the WNT pathway in breast cancer patients and highlight the importance and complexity of selecting the subset of patients that may benefit from treatment.
WNT proteins and their downstream effectors form a highly conserved signaling network that regulates tissue morphogenesis during development and adult tissue homeostasis in virtually all multicellular animals studied to date (van Amerongen and Nusse, 2009; Loh et al., 2016; Schenkelaars et al., 2017). The mammalian genome contains 19 WNT genes, encoding 19 different WNT proteins. These can bind and activate 10 different FZD receptors and a handful of co-receptors, thereby initiating different intracellular signaling cascades. ‘Canonical’ WNT signaling is defined by its use of β-catenin (CTNNB1) as main downstream effector and transcriptional co-activator of TCF/LEF target gene expression (MacDonald et al., 2009; Clevers and Nusse, 2012; Nusse and Clevers, 2017). ‘Non-canonical’ WNT signaling responses do not use CTNNB1, but instead activate different signaling molecules with profound impact on the cytoskeleton and cell migration (Komiya and Habas, 2008; van Amerongen, 2012; VanderVorst et al., 2018).
For both historic and experimental reasons, the intestinal epithelium has become the benchmark against which all other tissues are weighed when it comes to WNT signaling. This has shaped both our thinking and our terminology, with the intestine frequently being referred to as the “typical” example. A large body of literature shows that stem cell self-renewal and differentiation in the intestine and other endodermal derivatives is critically dependent on WNT/CTNNB1 signaling (Sato et al., 2009; Barker et al., 2010; Huch et al., 2013a, b; Clevers et al., 2014; Clevers, 2016). Hyperactive WNT/CTNNB1 signaling is a hallmark of colorectal cancer, both in early stages of polyp formation and at later stages of invasion and metastasis (Zhang and Shay, 2017). In this context, increased WNT/CTNNB1 signaling mainly results from genetic mutations in the APC gene, which encodes a negative regulator of CTNNB1 (Fodde, 2002). The unambiguous genetic evidence from human tumors leaves little doubt about the relevance of aberrant WNT/CTNNB1 signaling in the initiation and progression of colorectal cancer.
The involvement of WNT signaling in breast cancer remains less well understood (Yu et al., 2016; Alexander, 2018). This is surprising, given that the link between WNT signaling and breast cancer is as old as the WNT research field itself (Nusse and Varmus, 2012). In fact, the first mammalian WNT gene (Wnt1, originally identified as int-1) was discovered as a proto-oncogene capable of driving mammary tumor formation in mice (Nusse and Varmus, 1982). Here we review the evidence, highlight current research gaps and indicate future avenues worth exploring to dissect the role of WNT signaling in human breast cancer.
A first major knowledge gap is our lack of a basic understanding of the role of WNT signaling in human breast development and physiology. The mammary gland largely develops after birth and undergoes dynamic tissue remodeling throughout life. The most prominent changes occur in puberty (when the breast tissue develops under the influence of rising levels of estrogen and progesterone), and during pregnancy and lactation (when it differentiates and produces milk to nurture the offspring). Given how critical this tissue has been for our survival as a mammalian species and in view of the prevalence and mortality of breast cancer across different societies in women worldwide, it remains somewhat strange that we still have an incomplete picture of the molecular, cell and tissue biology of the human breast. In fact, one of the most detailed studies of human breast development, and individual variation therein, arguably dates back to 18401.
Most of what we know about WNT signaling in mammary gland biology and breast cancer comes from studies in mice, where both CTNNB1-dependent and -independent signaling are essential for mammary gland development, branching morphogenesis and function during embryogenesis and in postnatal life (Brisken et al., 2000; Chu et al., 2004; Veltmaat et al., 2004; Badders et al., 2009; Roarty et al., 2015; Yu et al., 2016). The mouse was discovered as a useful organism for studying the link between hormones and breast cancer well over a century ago (Lathrop and Loeb, 1916), but it really came to the fore as an experimental model system with the discovery of the fat pad transplantation assay (Deome et al., 1959). This technique remains indispensable for studying the growth, differentiation and regenerative properties of different mammary epithelial cell populations (Faraldo et al., 2015; Wronski et al., 2015). Nowadays, robust protocols allow the prospective isolation of mammary stem cells (capable of forming a new epithelial network upon transplantation) via fluorescence activated cell-sorting (FACS) (Shackleton et al., 2006; Stingl et al., 2006; Prater et al., 2013; Gao et al., 2016). More recently, genetically engineered mouse models have allowed sophisticated lineage tracing approaches, which have been instrumental for studying mammary stem and progenitor cell behavior in situ (van Amerongen, 2015; van de Moosdijk et al., 2017).
Multiple efforts have been made to delineate the mouse mammary epithelial cell hierarchy. The cumulative lineage tracing literature suggests that postnatal mammary gland development, homeostasis and remodeling are mainly driven by unipotent basal and luminal stem cells (Van Keymeulen et al., 2011; Davis et al., 2016; Wuidart et al., 2016, 2018; Scheele et al., 2017), although a rare fraction of bipotent stem cells likely co-exists (Wang et al., 2015). At least some mammary stem cells are WNT/CTNNB1 responsive (Zeng and Nusse, 2010; De Visser et al., 2012; van Amerongen et al., 2012a; Plaks et al., 2013; Wang et al., 2015; Blaas et al., 2016). However, this does not automatically imply that homeostasis and remodeling of the mammary epithelium is as strictly controlled by WNT/CTNNB1 responsive stem cells as appears to be the case for the intestinal epithelium. Moreover, stem cell plasticity can be induced by transplantation (Van Keymeulen et al., 2011; van Amerongen et al., 2012a) or oncogenic mutations (Koren et al., 2015; Van Keymeulen et al., 2015), raising the question if mammary stem and progenitor cells should be forced into a rigid hierarchy to begin with.
How findings from the mouse translate to the human breast remains unclear. In both human and mouse, the mammary gland is comprised of a non-stereotypically branched, ductal network composed of a bilayer of basal and luminal epithelial cells. Yet neither the two tissues, nor the experimental systems available to study each of them, are directly comparable between the two species. Major differences exist in the composition of the stroma, with the mouse mammary gland containing a higher proportion of adipocytes (hence the name ‘fat pad’ for the stromal pocket into which cells can be transplanted) and the human breast containing considerably more collagen. This constitutes a different molecular signaling environment with very different mechanobiological properties. Breast tissue composition changes throughout life and varies between individual women (Sun et al., 2014). Prominent differences in the expression pattern of epithelial cell markers between mouse and human also exist, although these are frequently ignored. For example, KRT14 reliably marks basal cells in the mouse mammary gland but is also expressed in a fraction of luminal cells in the human breast (Santagata et al., 2014; Dontu and Ince, 2015; McNally and Stein, 2017; Gerdur Ísberg et al., 2019).
Unlike in mice, human stem cell activity cannot be readily visualized in vivo. Unraveling the stem and progenitor cell hierarchy in the breast has thus proven difficult, but a recent study managed to use Cytochrome C Oxidase deficiency to identify multi-lineage differentiation in the healthy breast, presumably from stem cells in the luminal layer (Cereser et al., 2018). Experimental systems to study self-renewal and differentiation of human breast epithelial cells are limited to in vitro cell culture assays. Primary mammosphere cultures (in which cells are grown in suspension to enrich for cells with self-renewal properties) are frequently used to evaluate human breast stem cell activity (Shaw et al., 2012). However, this link is indirect and may not reflect the in vivo situation.
Access to healthy human breast tissue for experimental purposes is usually restricted to the leftover material from breast reduction surgeries. FACS protocols have been developed to isolate different cell populations from these specimens, including an ALDH + population with stem/progenitor cell properties as evaluated by multi-lineage differentiation in a 2D clonogenic colony formation assay (Ginestier et al., 2007). Transcriptional profiling of these cells revealed that they express high levels of WNT2 and RSPO3, suggesting an autocrine source of ligands and agonists (Colacino et al., 2018). Mammosphere cultures are typically maintained in the absence of exogenous WNT proteins, but cells in these cultures do express FZD2 (Shaw et al., 2012). Although primary human mammosphere cultures appear to be relatively insensitive to DKK1-mediated inhibition of WNT signaling (Lamb et al., 2013), multiple WNT genes can be induced in these cultures upon stimulation with estrogen or progesterone (Arendt et al., 2014). Comparative transcriptional profiling between mouse and human epithelial cells suggests that active WNT/CTNNB1 signaling in the basal cell population is conserved between the two species (Lim et al., 2010) and long-term maintenance of primary human as well as mouse mammary epithelial cells in Matrigel has been reported in the presence of WNT3A-containing media (Zeng and Nusse, 2010; Sachs et al., 2018).
Summarizing, the human breast likely also uses WNT signaling for growth and differentiation. However, the WNT-secreting and WNT-responsive cells have not been clearly demarcated. Single cell RNA sequencing studies will likely shed more light on the stem and progenitor cell hierarchy in the healthy human breast, and on the position of WNT/CTNNB1 signaling in this hierarchy, in the foreseeable future (Holliday and Speirs, 2011). If and how CTNNB1-dependent and –independent signaling functionally controls proliferation, differentiation and branching morphogenesis of primary human breast epithelial cells is something that can likely only be answered using primary 3D organotypic cultures (Linnemann et al., 2015, 2017).
A second research gap is the lack of specific markers to reliably measure WNT signaling activity in human breast cancer. CTNNB1-independent signaling responses are notorious for their lack of robust readouts in most mammalian cells and tissues. For CTNNB1-dependent signaling, such readouts are available: Reporter constructs with concatemerized TCF/LEF binding sites can be introduced into cells and patient derived xenografts to measure WNT/CTNNB1 signaling (Green et al., 2013; Many and Brown, 2014). However, this approach is unsuitable for monitoring pathway activity in histological specimens, nor does it probe multifactorial signaling in the endogenous chromatin context (Nakamura et al., 2016; Doumpas et al., 2019).
Two of the earliest described WNT/CTNNB1 target genes are CCND1 and MYC (He et al., 1998; Shtutman et al., 1999). Elevated protein levels of CCND1 and MYC are detected in a high proportion of invasive ductal breast carcinomas, but this does not always correlate to CTNNB1 expression levels (Wong et al., 2002; He et al., 2014). Given their general involvement in cell proliferation, upregulation of CCND1 and MYC can be achieved in myriad ways (Lindqvist et al., 2014). So far, AXIN2 appears to be one of the few universal target genes that could be used to reliably measure relative WNT/CTNNB1 signaling activity in human breast cancer (Lustig et al., 2001; Jho et al., 2002).
In the absence of a well-defined, mammary-specific WNT/CTNNB1 target gene expression program and given the preponderance of paraffin embedded tumor specimens, immunohistochemical detection of CTNNB1 protein levels has been used as the most direct way to readout WNT/CTNNB1 signaling. From these analyses it has been known for a long time that elevated intracellular levels of CTNNB1, a hallmark of active WNT/CTNNB1 signaling, can be detected by immunohistochemistry in a significant (13–77%) proportion of all ductal and lobular breast cancer samples (Jonsson et al., 2000; Karayiannakis et al., 2001; Wong et al., 2002; Ozaki et al., 2005; Prasad et al., 2008a; He et al., 2014; Hou et al., 2018). Care should be taken when performing and interpreting these experiments: Dogma dictates that active WNT/CTNNB1 signaling results in increased nuclear CTNNB1 levels, but those with more hands on experience in the field know that changes in CTNNB1 can be quite subtle and even modest (2–5 fold) increases in the levels of intracellular CTNNB1 can be more than sufficient to robustly activate TCF/LEF target gene expression (Jacobsen et al., 2016).
Clinical evidence suggests that WNT/CTNNB1 signaling is elevated across multiple subtypes of human breast cancer. Aggressive triple negative breast carcinomas (TNBC) were found to be enriched for elevated CTNNB1 levels compared to luminal A, luminal B or HER2+ tumors (Khramtsov et al., 2010). Higher levels of intracellular CTNNB1 are associated with a higher tumor grade (Sormunen et al., 1999) and poor prognosis (Lin et al., 2000; Khramtsov et al., 2010). The highest levels of CTNNB1 are found in metaplastic carcinomas and non-metastasizing fibromatosis – two rare subsets of breast cancer (Lacroix-Triki et al., 2010). Here, up to 90% of tumors show increased levels of CTNNB1 and a proportion of these may contain activating genetic mutations in the CTNNB1 gene (Abraham et al., 2002; Hayes et al., 2008; Hennessy et al., 2009). For the most part however, and unlike the situation encountered in colorectal cancer, genetic mutations in APC, AXIN or CTNNB1 are virtually non-existent in human breast tumors (Figure 1A). As first proposed many years ago, this discrepancy can likely be explained by tissue-specific differences in sensitivity to WNT/CTNNB1 signaling (Gaspar and Fodde, 2004; Gaspar et al., 2009).
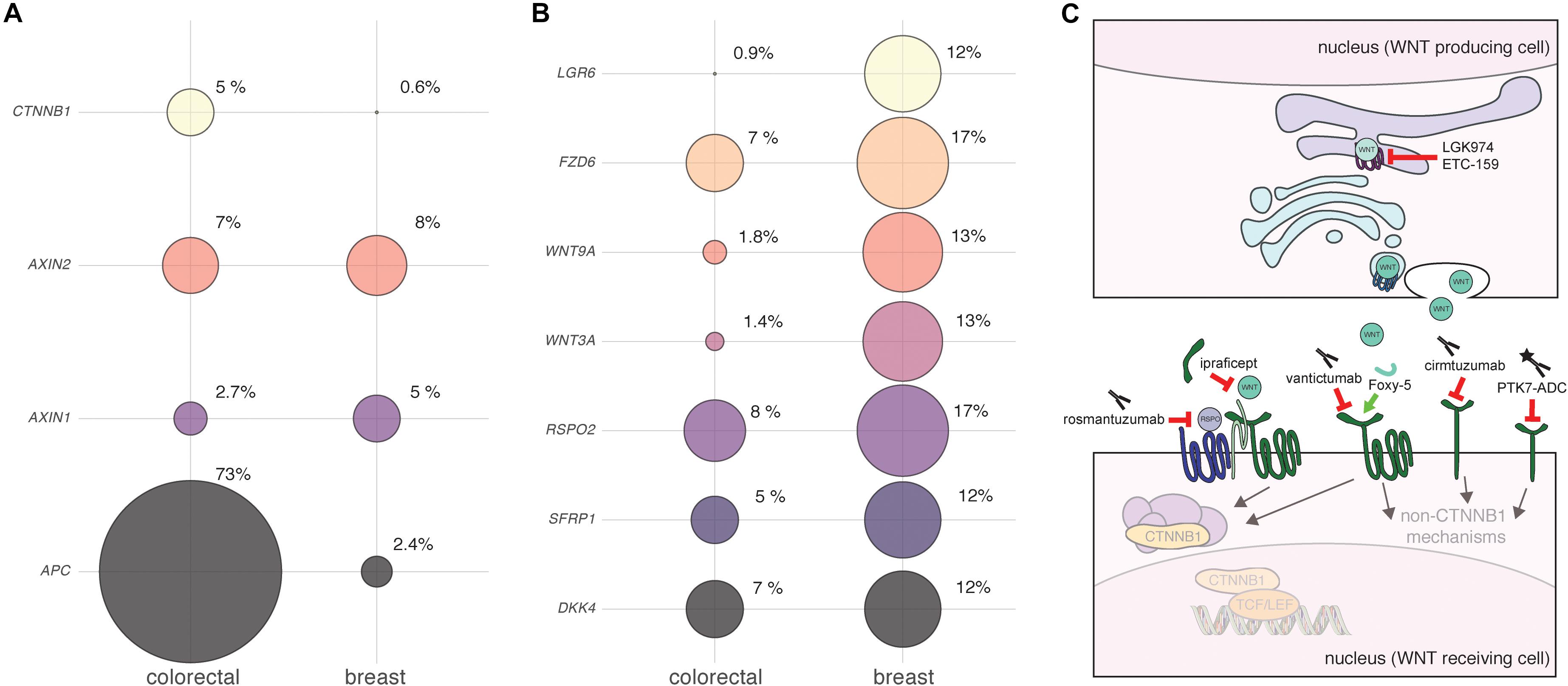
Figure 1. Detecting and targeting aberrant WNT signaling in human breast cancer. (A,B) Bubble plots illustrating the alteration of different WNT pathway components in breast versus colorectal cancer. Plots were generated using data from http://cbioportal.org (accessed on 20 September 2019), using the following datasets: Colorectal Adenocarcinoma (TCGA, Provisional), samples with mutation and copy number alteration data (220 patients/samples). Breast Invasive Carcinoma (TCGA, Provisional), samples with mutation and copy number alteration data (963 patients/samples). Circle sizes reflect the proportion of samples with alterations in each of the genes depicted, with the actual percentages shown. Note that copy number alterations (amplifications + deletions) and mutations (truncations + substitutions) were combined into a single score. No distinction was made between breast cancer subtypes. Data were not corrected for overall differences in mutation rates or genome instability between the different tumor types. No inference can be made about RNA and protein expression level changes based solely on these analyses. (A) APC is the most prominently mutated gene in colorectal cancer. Other endodermal cancers, including hepatocellular carcinoma, also show frequent genetic mutations in WNT/CTNNB1 signaling components (White et al., 2012). Depending on the tissue of origin and tumor subtype, activating mutations in CTNNB1 itself or inactivating mutations in negative regulators like APC or AXIN1 are more or less prevalent (Yanagisawa et al., 1992; Morin, 1997; Ishizaki et al., 2004). In breast cancer, genetic mutations in APC are rare. However, epigenetic changes such as APC promoter hypermethylation have been reported in the literature, with the highest incidence observed in inflammatory breast cancer (Jin et al., 2001; Van Der Auwera et al., 2008; Lindqvist et al., 2014). (B) The top genes that show genetic alterations in breast cancer are implicated to a lesser extent in colorectal cancer. Note that all of these components function at the level of ligand and receptor binding. The top two hits, RSPO2 and FZD6, have both been linked to reduced metastasis free survival, but likely operate via different WNT signaling mechanisms (Corda et al., 2017; Coussy et al., 2017). It should be stressed that in this respect, breast cancer is not unique. As more and more genome-wide expression profiling studies are becoming available, evidence is accumulating that many different cancers likely display changes in WNT/CTNNB1 signaling in the absence of mutations in APC or CTNNB1 (Wiese et al., 2018; Flanagan et al., 2019b). In addition, it was recently demonstrated that FZD7, which functions upstream of APC and CTNNB1, is required for WNT/CTNNB1 signaling in gastric tumors irrespective of their APC status (Flanagan et al., 2019a). This is reminiscent of earlier studies hinting toward a similar phenomenon for other upstream components (Suzuki et al., 2004). Even in colorectal cancer, the situation may thus be far more complex than envisioned, and the local niche may continue to affect signaling levels even when the WNT/CTNNB1 pathway is intrinsically activated through genetic mutations in APC (van Neerven and Vermeulen, 2019). (C) Cartoon showing the points of interception for WNT-pathway targeting drugs that are currently in clinical trials. See text for details.
In the absence of any apparent genetic mutations, what then is the cause of elevated CTNNB1 levels in human breast cancer? In the normal human breast, CTNNB1 is mainly detected in the cell membrane as part of adherens junctions (Hashizume et al., 1996). It cannot be excluded that the increase in CTNNB1 could therefore, at least partially, be due to its release from these junctions upon loss of CDH1, given that this is a frequent event in more advanced and invasive tumors (Prasad et al., 2008b; Zeljko et al., 2011). However, another possibility is that CTNNB1 levels are increased as a direct result of enhanced WNT/CTNNB1 signaling due to changes in the expression levels of upstream WNT pathway components. In large public breast cancer datasets, changes at the level of ligands, (ant)agonists and receptors are readily apparent (Figure 1B). Moreover, the cumulative literature provides ample evidence of changes in the levels of ligands and receptors in primary or metastatic human breast cancer (Table 1). In interpreting these findings, some caution is warranted. First, few of the RNA expression level changes have been shown to affect protein levels. Second, where such follow up is performed, antibody specificity has not always been properly validated.
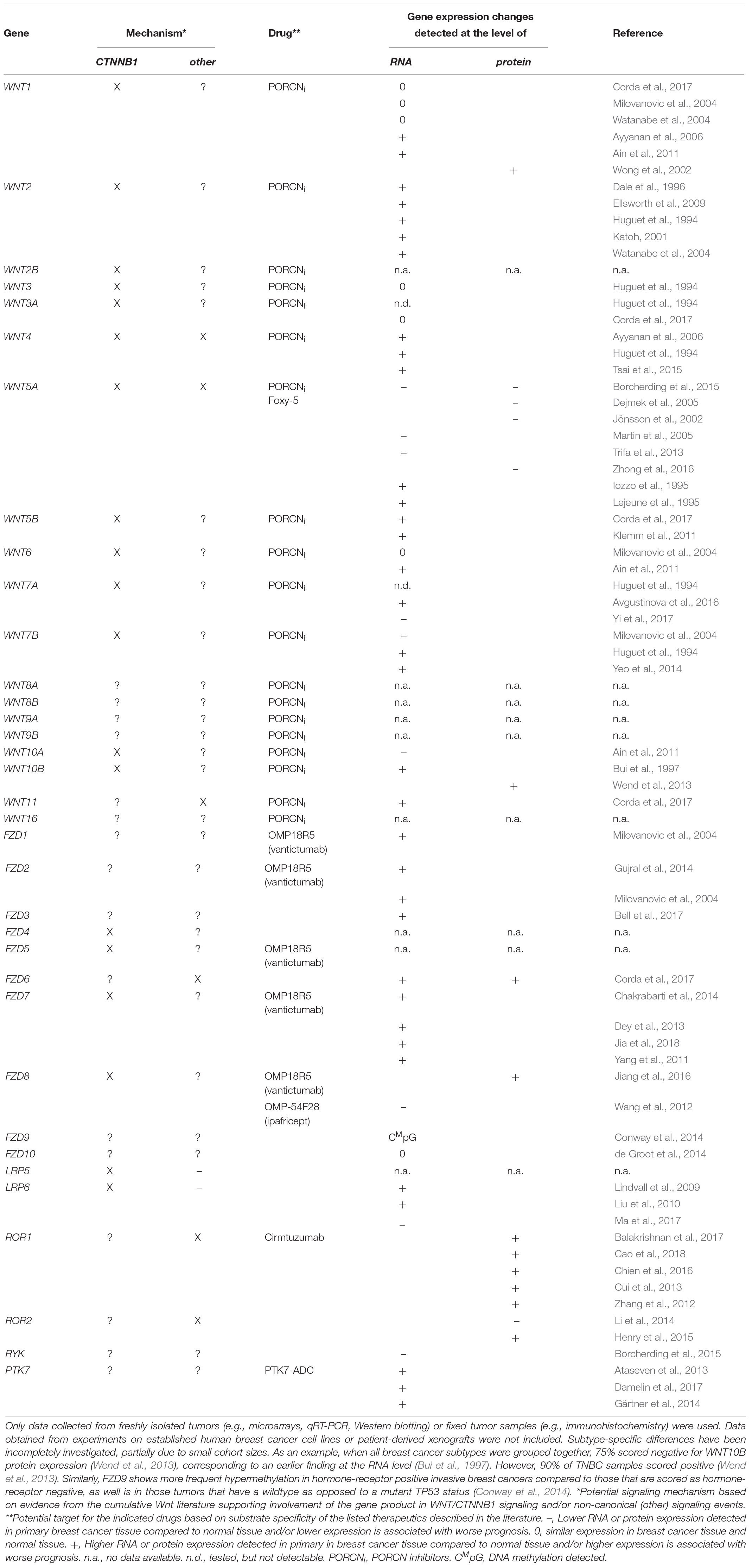
Table 1. Comprehensive overview of ligand (WNT1-16) and receptor (FZD1-10, LRP5-6, ROR1-2, RYK, PTK) genes and their implication in human breast cancer based on a survey of the primary literature.
Since absolutely no inference about cell biological mechanisms can be made solely based on expression level changes, functional follow up is crucial to determine the implications of these alterations. For example, only FZD7 is consistently found to signal through CTNNB1/TCF in human breast cancer cells, thereby affecting cell proliferation (Yang et al., 2011; Chakrabarti et al., 2014; Riley and Day, 2017). In contrast, copy number gain of the FZD6 gene, which can be readily detected in human breast cancer cohorts (Figure 1B) and most predominantly in TNBC, most likely exerts its effects on cell motility and invasion via alternative, non-canonical WNT signaling mechanisms (Corda et al., 2017). For other components, such as RSPO2, RSPO4 and to a lesser extent LGR5 and LGR6, the overexpression of which is enriched in TNBC, the mechanism is more likely to involve amplification of the WNT/CTNNB1 signaling response (Coussy et al., 2017). Importantly, the separation between canonical and non-canonical WNT signaling is not black and white. For instance, WNT5A, still frequently regarded as the “typical” non-canonical WNT ligand, can both repress and activate CTNNB1-dependent signaling, in vitro as well as in vivo (Mikels and Nusse, 2006; van Amerongen et al., 2012b). Especially in the context of cancer, where cellular signaling pathways are invariably deregulated, unexpected signaling activities are likely to be encountered (Grossmann et al., 2013).
Summarizing, more extensive transcriptional and epigenetic profiling of tumor and adjacent normal tissue is needed to reveal the true extent of aberrant WNT signaling in human breast cancer. Early studies reported hypermethylation, and presumably silencing, of genes encoding secreted WNT-pathway inhibitors as a potential mechanism for disrupting the balance in WNT signaling in breast cancer. Examples are widespread and include WIF1 (Wissman et al., 2003; Ai et al., 2006; Veeck et al., 2009), SFRP1 (Ugolini et al., 2001; Veeck et al., 2006; Suzuki et al., 2008), SFRP2 (Suzuki et al., 2008; Lindqvist et al., 2014), SFRP5 (Suzuki et al., 2008; Veeck et al., 2008a; Lindqvist et al., 2014), DKK1 (Forget et al., 2007; Suzuki et al., 2008) and DKK3 (Veeck et al., 2009; Lindqvist et al., 2014; Yamaguchi et al., 2015). Epigenetic analyses, such as those measuring DNA methylation levels, are now becoming part of the standard work flow for large consortia. The first of such analyses indeed revealed extensive changes in WNT signaling components across breast tumors (Koval and Katanaev, 2018). The main challenge still lies ahead as we face the daunting task of properly interpreting these experimental findings. For instance, DKK3 and WIF1 methylation was detected in a similar proportion of breast cancer patients, but only DKK3 methylation was a prognostic marker of survival (Veeck et al., 2009). And while one study reported SFRP2 promoter hypermethylation in more than 80% of breast cancer patients (Veeck et al., 2008b), a recent report suggests that, in contrast, elevated serum levels of SFRP2 may serve as an independent marker for poor prognosis (Huang et al., 2019). Future studies will also have to focus on subtype-specific differences.
Our current lack of understanding which patients are most likely to benefit from treatment with WNT inhibitors is a third major knowledge gap. Several drugs that interfere with the WNT signaling pathway are currently being tested in clinical trials (for recent reviews see Krishnamurthy and Kurzrock, 2018; Ghosh et al., 2019). After decades of ill-fated attempts to block WNT signaling downstream of CTNNB1, the current developmental pipeline is fueled by two different rationales (Figure 1C). The first is the conceptual notion that, even in the absence of apparent mutations, WNT/CTNNB1 plays a central role in the maintenance of multiple adult tissue stem cell populations and, by analogy and extension, in cancer stem cells. This line of reasoning forms the basis for the development of drugs that inhibit WNT protein secretion, such as the PORCN inhibitors LGK974 and ETC-159 (Liu et al., 2013; Madan et al., 2016). The main adverse effects reported for PORCN inhibitors in Phase I clinical trials are related to loss of bone density (Ng et al., 2017; Tan et al., 2018). Somewhat surprisingly, the systemic toxicity of PORCN inhibitors appears to be relatively limited. One potential explanation for this observation comes from experiments conducted in mice. Here, the WNT-secreting intestinal myofibroblasts, which constitute the intestinal stem cell niche, were shown to be intrinsically resistant to xenobiotics, including PORCN inhibitors, because they express a subset of multidrug efflux pumps (Chee et al., 2018). While this opens a therapeutic window, it also leads to the sobering conclusion that tumor cells may likely evolve similar resistance mechanisms upon prolonged treatment. In fact, these same ATP-binding cassette (ABC) transporters have long been implicated in acquired multidrug resistance in cancer, albeit in the context of classical chemotherapeutic agents rather than targeted therapeutics (Robey et al., 2018). In addition, although it is generally assumed that all WNT ligands require PORCN for their secretion, exceptions to this rule may exist (Rao et al., 2018).
The second rationale for designing drugs that interfere with WNT signaling are more focused and evidence based. These efforts are directed toward specific WNT-pathway components that show altered expression in human tumors. Examples include the anti-RSPO3 antibody OMP-131R10/rosmantuzumab and the decoy receptor FZD8-CRD OMP-54F28/ipafricept (Cattaruzza et al., 2015; Le et al., 2015). So far, the most promising results for breast cancer have been obtained with the broad-spectrum anti-FZD antibody OMP-18R5/vantictumab, which blocks FZD1, 2, 5, 7, and 8 (Gurney et al., 2012). In pre-clinical trials, OMP-18R5 was shown to inhibit the outgrowth of patient derived breast cancer xenografts, thus demonstrating potential efficacy against breast cancer (Gurney et al., 2012; Fischer et al., 2017). A phase Ib clinical trial in HER2– breast cancer patients identified a four-gene signature (FBXW2, CCND2, CTBP2, and WIF1) as a potential predictive biomarker for the response to combined treatment with paclitaxel and vantictumab (Zhang et al., 2018). Structure guided design will likely help in generating more specific antibodies that target individual FZD receptors (Raman et al., 2019). Based on the available data, FZD6 and FZD7 seem obvious candidates for therapeutic intervention (Figure 1 and Table 1).
Few WNT-pathway targeting drugs that are currently in clinical trials were explicitly developed with breast cancer in mind. A notable exception is Foxy-5, a peptide mimetic of WNT5A that was designed with the goal of blocking breast cancer metastasis by reconstituting a – presumably non-CTNNB1 driven –WNT5A signaling response in cancers that had lost WNT5A expression (Säfholm et al., 2008). While WNT5A protein expression was found to be low in 75% of TNBC tumors, medium to high expression was detected in 75% of ER+ breast cancer samples (Borcherding et al., 2015). Furthermore, expression levels may change upon treatment, as WNT5A protein levels were significantly higher in 79% of patients after relapse and elevated WNT5A levels were also associated with the induction of multidrug resistance (Hung et al., 2014).
In many cancers, including breast cancer, only a small population of tumor cells, the so-called ‘cancer stem cells,’ may be responsible for driving tumor growth. Human breast cancer stem cells were first identified as tumor initiating cells following transplantation into immunocompromised mice (Al-Hajj et al., 2003) and have been connected to metastasis formation and resistance to therapy. Given the presumed importance of WNT/CTNNB1 signaling in breast cancer stem cell maintenance (Lamb et al., 2013; Jang et al., 2015; Hou et al., 2018), it is somewhat counterintuitive that the non-canonical co-receptor ROR1 is emerging as a potential key mediator of chemoresistance in breast cancer stem cells (Zhang et al., 2019). Overexpression of ROR1 is a prognostic marker in TNBC (Chien et al., 2016) and the anti-ROR1 antibody cirmtuzumab, originally developed for treating chronic lymphocytic B-cell leukemia (Zhang et al., 2013), is therefore also in clinical trials for human breast cancer. Initial interest in ROR1 as a potential therapeutic target arose because of its low expression in healthy adult tissues, although a new antibody against ROR1, specifically designed for immunohistochemistry on FFPE samples, shows higher endogenous ROR1 expression than previously suspected (Shabani et al., 2015; Balakrishnan et al., 2017). Another unexpected candidate for targeting breast cancer stem cells surfaced in the form of PTK7, a WNT receptor whose function is not yet completely elucidated (Damelin et al., 2017). PTK7-ADC, a PTK7-targeting antibody that is conjugated to a cytotoxic drug, has also entered phase I clinical trials for metastatic TNBC (Radovich et al., 2019).
Summarizing, it is still too early to conclude anything about the impact of these drugs on breast cancer patient survival. If these therapeutics continue on to more advanced stages of clinical testing, the main challenge will still be to demonstrate true clinical efficacy by rationally selecting those patients that are most likely to benefit from treatment.
The absence of well-defined genetic mutations complicates our assessment of the functional importance of aberrant WNT signaling in human breast cancer. No definitive or generalized conclusions can be drawn about the role of either WNT/CTNNB1 or CTNNB1-independent WNT signaling at this point. Given their pleiotropic effects, we need a lot more insight into how these different signal transduction routes affect breast cancer initiation and progression. For this, we need to unravel the basic biological mechanisms through which the complex WNT signaling network controls normal human breast development and physiology. These studies will do more than just satisfy scientific curiosity: They will ultimately be critical to determine which breast cancer subtypes or individual patients are most likely to benefit from targeted therapeutics designed to interfere with WNT signaling activity, taking into account the growth promoting and inhibitory activities of individual ligand/receptor pairings in different cellular contexts.
Both patient selection and monitoring of their clinical response will require new assays and biomarkers. Our drug intervention strategies, in turn, need to be fine-tuned in such a way that individual WNT/receptor interactions or downstream signaling responses can be blocked or activated with great precision. For instance, whereas downregulation of DKK1 has been linked to lung metastases, patients with high levels of DKK1 more frequently present with bone metastases (Zhuang et al., 2017). And while the former has been suggested to occur via a non-canonical signaling mechanism, the latter likely occurs through DKK1-mediated inhibition of WNT/CTNNB1 signaling. In either case, the use of a PORCN inhibitor or a pan-FZD antibody would seem ill advised in both of these cases. Moreover, the adverse effects of these pan-WNT inhibitors on bone density will need to be overcome to advance their clinical use (Madan et al., 2018).
Finally, breast cancer is a systemic disease and the involvement of WNT signaling should be considered from this perspective as well. Both in mice and humans, loss of TP53 has recently been associated with the induction of WNT protein production, which may in turn stimulate the immune system to promote metastasis (Kim et al., 2019; Liu et al., 2019; Wellenstein et al., 2019). Likewise, cytokine signaling from the local bone microenvironment may promote metastatic colonization by initiating an autocrine WNT signaling loop in human breast cancer stem cells (Eyre et al., 2019). At present, functional studies almost invariably fall back on the use of established human breast cancer cell lines. It is unlikely that these suffice to unravel the contribution of WNT signaling to human breast cancer. Comparing the results obtained in breast cancer cell lines to those obtained in studies with primary human breast cancer organoids and the analysis of patient-derived xenografts is warranted. Given the (epi)genetic diversity of the human breast cancer landscape, patient-to-patient heterogeneity and the interplay between breast cancer cells and their local and systemic environment, the inclusion of stromal and immune components in these experimental model systems will be essential (Holliday and Speirs, 2011; Stephens et al., 2012; Pereira et al., 2016).
RA contributed to the conception and design of the study and wrote the first draft of the manuscript. ES performed the literature survey that is summarized in Table 1 and wrote sections of the manuscript. ES and RA contributed to acquisition, analysis and interpretation of the literature. All authors contributed to the manuscript revision, read and approved the submitted version.
RA acknowledges funding from the following sources: KWF Kankerbestrijding (Dutch Cancer Society, career development award ANW 2013-6057, project grant 11082/2017-1), NWO (Netherlands Science Foundation, VIDI 864.13.002) and a MacGillavry fellowship from the University of Amsterdam.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank Yorick van de Grift for critical reading and feedback on the manuscript.
Abraham, S. C., Reynolds, C., Lee, J. H., Montgomery, E. A., Baisden, B. L., Krasinskas, A. M., et al. (2002). Fibromatosis of the breast and mutations involving the APC/β-catenin pathway. Hum. Pathol. 33, 39–46. doi: 10.1053/hupa.2002.30196
Ai, L., Tao, Q., Zhong, S., Fields, C. R., Kim, W. J., Lee, M. W., et al. (2006). Inactivation of Wnt inhibitory factor-1 (WIF1) expression by epigenetic silencing is a common event in breast cancer. Carcinogenesis 27, 1341–1348. doi: 10.1093/carcin/bgi379
Ain, Q., Seemab, U., Nawaz, S., and Rashid, S. (2011). Integrative analyses of conserved WNT clusters and their co-operative behaviour in human breast cancer. Bioinformation 7, 339–346. doi: 10.6026/97320630007339
Alexander, C. M. (2018). The Wnt signaling landscape of mammary stem cells and breast tumors. Prog. Mol. Biol. Transl. Sci. 153, 271–298. doi: 10.1016/bs.pmbts.2017.11.020
Al-Hajj, M., Wicha, M. S., Benito-Hernandez, A., Morrison, S. J., and Clarke, M. F. (2003). Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. U.S.A. 100, 3983–3988.
Arendt, L. M., St Laurent, J., Wronski, A., Caballero, S., Lyle, S. R., Naber, S. P., et al. (2014). Human breast progenitor cell numbers are regulated by WNT and TBX3. PLoS One 9:e111442. doi: 10.1371/journal.pone.0111442
Ataseven, B., Angerer, R., Kates, R., Gunesch, A., Knyazev, P., Högel, B., et al. (2013). PTK7 expression in triple-negative breast cancer. Anticancer Res. 33, 3759–3763.
Avgustinova, A., Iravani, M., Robertson, D., Fearns, A., Gao, Q., Klingbeil, P., et al. (2016). Tumour cell-derived WNT7A recruits and activates fibroblasts to promote tumour aggressiveness. Nat. Commun. 7:10305. doi: 10.1038/ncomms10305
Ayyanan, A., Civenni, G., Ciarloni, L., Morel, C., Mueller, N., Lefort, K., et al. (2006). Increased Wnt signaling triggers oncogenic conversion of human breast epithelial cells by a Notch-dependent mechanism. Proc. Natl. Acad. Sci. U.S.A. 103, 3799–3804. doi: 10.1073/pnas.0600065103
Badders, N. M., Goel, S., Clark, R. J., Klos, K. S., Kim, S., Bafico, A., et al. (2009). The Wnt receptor, Lrp5, is expressed by mouse mammary stem cells and is required to maintain the basal lineage. PLoS One 4:e6594. doi: 10.1371/journal.pone.0006594
Balakrishnan, A., Goodpaster, T., Randolph-Habecker, J., Hoffstrom, B. G., Jalikis, F. G., Koch, L. K., et al. (2017). Analysis of ROR1 protein expression in human cancer and normal tissues. Clin. Cancer Res. 23, 3061–3071. doi: 10.1158/1078-0432.CCR-16-2083
Barker, N., Huch, M., Kujala, P., van de Wetering, M., Snippert, H. J., van Es, J. H., et al. (2010). Lgr5+ve stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro. Cell Stem Cell 6, 25–36. doi: 10.1016/j.stem.2009.11.013
Bell, R., Barraclough, R., and Vasieva, O. (2017). Gene expression meta-analysis of potential metastatic breast cancer markers. Curr. Mol. Med. 17, 200–210.
Blaas, L., Pucci, F., Messal, H. A., Andersson, A. B., Ruiz, E. J., Gerling, M., et al. (2016). Lgr6 labels a rare population of mammary gland progenitor cells that are able to originate luminal mammary tumours. Nat. Cell Biol. 18, 1346–1356. doi: 10.1038/ncb3434
Borcherding, N., Kusner, D., Kolb, R., Xie, Q., Li, W., Yuan, F., et al. (2015). Paracrine WNT5A signaling inhibits expansion of tumor-initiating cells. Cancer Res. 75, 1972–1982. doi: 10.1158/0008-5472.CAN-14-2761
Brisken, C., Heineman, A., Chavarria, T., Elenbaas, B., Tan, J., Dey, S. K., et al. (2000). Essential function of Wnt-4 in mammary gland development downstream of progesterone signaling. Genes Dev. 14, 650–654.
Bui, T. D., Rankin, J., Smith, K., Huguet, E. L., Ruben, S., Strachan, T., et al. (1997). A novel human Wnt gene, WNT10B, maps to 12q13 and is expressed in human breast carcinomas. Oncogene 14, 1249–1253. doi: 10.1038/sj.onc.1200936
Cao, J., Wang, X., Dai, T., Wu, Y., Zhang, M., Cao, R., et al. (2018). Twist promotes tumor metastasis in basal-like breast cancer by transcriptionally upregulating ROR1. Theranostics 8, 2739–2751. doi: 10.7150/thno.21477
Cattaruzza, F., Yeung, P., Yen, W.-C., Brunner, A., Wang, M., Liu, Y., et al. (2015). Abstract 4367: discovery and evaluation of pharmacodynamic and predictive biomarkers for anti-RSPO3, a treatment that reduces tumor growth and cancer stem cell frequency in patient derived xenograft tumor models. Cancer Res. 75:4367.
Cereser, B., Jansen, M., Austin, E., Elia, G., McFarlane, T., van Deurzen, C. H. M., et al. (2018). Analysis of clonal expansions through the normal and premalignant human breast epithelium reveals the presence of luminal stem cells. J. Pathol. 244, 61–70. doi: 10.1002/path.4989
Chakrabarti, R., Wei, Y., Hwang, J., Hang, X., Andres Blanco, M., Choudhury, A., et al. (2014). Δnp63 promotes stem cell activity in mammary gland development and basal-like breast cancer by enhancing Fzd7 expression and Wnt signalling. Nat. Cell Biol. 16, 1004–1015. doi: 10.1038/ncb3040
Chee, Y. C., Pahnke, J., Bunte, R., Adsool, V. A., Madan, B., and Virshup, D. M. (2018). Intrinsic xenobiotic resistance of the intestinal stem cell niche. Dev. Cell 46, 681–695.e5. doi: 10.1016/j.devcel.2018.07.023
Chien, H. P., Ueng, S. H., Chen, S. C., Chang, Y. S., Lin, Y. C., Lo, Y. F., et al. (2016). Expression of ROR1 has prognostic significance in triple negative breast cancer. Virchows Arch. 468, 589–595. doi: 10.1007/s00428-016-1911-3
Chu, E. Y., Hens, J., Andl, T., Kairo, A., Yamaguchi, T. P., Brisken, C., et al. (2004). Canonical WNT signaling promotes mammary placode development and is essential for initiation of mammary gland morphogenesis. Development 131, 4819–4829. doi: 10.1242/dev.01347
Clevers, H. (2016). Modeling development and disease with organoids. Cell 165, 1586–1597. doi: 10.1016/j.cell.2016.05.082
Clevers, H., Loh, K. M., and Nusse, R. (2014). An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science 346:1248012. doi: 10.1126/science.1248012
Clevers, H., and Nusse, R. (2012). Wnt/β-catenin signaling and disease. Cell 149, 1192–1205. doi: 10.1016/j.cell.2012.05.012
Colacino, J. A., Azizi, E., Brooks, M. D., Harouaka, R., Fouladdel, S., McDermott, S. P., et al. (2018). Heterogeneity of human breast stem and progenitor cells as revealed by transcriptional profiling. Stem Cell Rep. 10, 1596–1609. doi: 10.1016/j.stemcr.2018.03.001
Conway, K., Edmiston, S. N., May, R., Kuan, P. F., Chu, H., Bryant, C., et al. (2014). DNA methylation profiling in the Carolina Breast Cancer Study defines cancer subclasses differing in clinicopathologic characteristics and survival. Breast Cancer Res. 16:450.
Corda, G., Sala, G., Lattanzio, R., Iezzi, M., Sallese, M., Fragassi, G., et al. (2017). Functional and prognostic significance of the genomic amplification of frizzled 6 (FZD6) in breast cancer. J. Pathol. 241, 350–361. doi: 10.1002/path.4841
Coussy, F., Lallemand, F., Vacher, S., Schnitzler, A., Chemlali, W., Caly, M., et al. (2017). Clinical value of R-spondins in triple-negative and metaplastic breast cancers. Br. J. Cancer 116, 1595–1603. doi: 10.1038/bjc.2017.131
Cui, B., Zhang, S., Chen, L., Yu, J., Widhopf, G. F., Fecteau, J.-F., et al. (2013). Targeting ROR1 inhibits epithelial-mesenchymal transition and metastasis. Cancer Res. 73, 3649–3660.
Dale, T. C., Weber-Hall, S. J., Smith, K., Huguet, E. L., Jayatilake, H., Gusterson, B. A., et al. (1996). Compartment switching of WNT-2 expression in human breast tumors. Cancer Res. 56, 4320–4323.
Damelin, M., Bankovich, A., Bernstein, J., Lucas, J., Chen, L., Williams, S., et al. (2017). A PTK7-targeted antibody-drug conjugate reduces tumor-initiating cells and induces sustained tumor regressions. Sci. Transl. Med. 9:eaag2611. doi: 10.1126/scitranslmed.aag2611
Davis, F. M., Lloyd-Lewis, B., Harris, O. B., Kozar, S., Winton, D. J., Muresan, L., et al. (2016). Single-cell lineage tracing in the mammary gland reveals stochastic clonal dispersion of stem/progenitor cell progeny. Nat. Commun. 7:13053. doi: 10.1038/ncomms13053
de Groot, J. S., Pan, X., Meeldijk, J., van der Wall, E., van Diest, P. J., and Moelans, C. B. (2014). Validation of DNA promoter hypermethylation biomarkers in breast cancer — a short report. Cell. Oncol. 37, 297–303. doi: 10.1007/s13402-014-0189-1
De Visser, K. E., Ciampricotti, M., Michalak, E. M., Tan, D. W. M., Speksnijder, E. N., Hau, C. S., et al. (2012). Developmental stage-specific contribution of LGR5+ cells to basal and luminal epithelial lineages in the postnatal mammary gland. J. Pathol. 228, 300–309. doi: 10.1002/path.4096
Dejmek, J., Leandersson, K., Manjer, J., Bjartell, A., Emdin, S. O., Vogel, W. F., et al. (2005). Expression and signaling activity of Wnt-5a/discoidin domain receptor-1 and Syk plays distinct but decisive roles in breast cancer patient survival. Clin. Cancer Res. 11, 520–528.
Deome, K. B., Faulkin, L. J., Bern, H. A., and Blair, P. B. (1959). Development of mammary tumors from hyperplastic alveolar nodules transplanted into gland-free mammary fat pads of female C3H mice. Cancer Res. 19, 515–520.
Dey, N., Young, B., Abramovitz, M., Bouzyk, M., Barwick, B., De, P., et al. (2013). Differential activation of Wnt-β-catenin pathway in triple negative breast cancer increases MMP7 in a PTEN dependent manner. PLoS One 8:e77425. doi: 10.1371/journal.pone.0077425
Dontu, G., and Ince, T. A. (2015). Of mice and women: a comparative tissue biology perspective of breast stem cells and differentiation. J. Mammary Gland Biol. Neoplasia 20, 51–62. doi: 10.1007/s10911-015-9341-4
Doumpas, N., Lampart, F., Robinson, M. D., Lentini, A., Nestor, C. E., Cantù, C., et al. (2019). TCF/LEF dependent and independent transcriptional regulation of Wnt/β-catenin target genes. EMBO J. 38:e98873.
Ellsworth, R. E., Seebach, J., Field, L. A., Heckman, C., Kane, J., Hooke, J. A., et al. (2009). A gene expression signature that defines breast cancer metastases. Clin. Exp. Metastasis 26, 205–213. doi: 10.1007/s10585-008-9232-9
Eyre, R., Alférez, D. G., Santiago-Gómez, A., Spence, K., McConnell, J. C., Hart, C., et al. (2019). Microenvironmental IL1β promotes breast cancer metastatic colonisation in the bone via activation of Wnt signalling. Nat. Commun. 10:5016.
Faraldo, M. M., Glukhova, M. A., and Deugnier, M.-A. (2015). The transplantation of mouse mammary epithelial cells into cleared mammary fat pads. Methods Mol. Biol. 1293, 161–172. doi: 10.1007/978-1-4939-2519-3_9
Fischer, M. M., Cancilla, B., Yeung, V. P., Cattaruzza, F., Chartier, C., Murriel, C. L., et al. (2017). WNT antagonists exhibit unique combinatorial antitumor activity with taxanes by potentiating mitotic cell death. Sci. Adv. 3:e1700090. doi: 10.1126/sciadv.1700090
Flanagan, D. J., Barker, N., Di Costanzo, N. S., Mason, E. A., Gurney, A., Meniel, V. S., et al. (2019a). Frizzled-7 is required for Wnt signaling in gastric tumors with and without APC mutations. Cancer Res. 79, 970–981. doi: 10.1158/0008-5472.CAN-18-2095
Flanagan, D. J., Vincan, E., and Phesse, T. J. (2019b). Wnt signaling in cancer: not a binary on: off switch. Cancer Res. 79, 5901–5906. doi: 10.1158/0008-5472.CAN-19-1362
Fodde, R. (2002). The APC gene in colorectal cancer. Eur. J. Cancer 38, 867–871. doi: 10.1016/s0959-8049(02)00040-0
Forget, M. A., Turcotte, S., Beauseigle, D., Godin-Ethier, J., Pelletier, S., Martin, J., et al. (2007). The Wnt pathway regulator DKK1 is preferentially expressed in hormone-resistant breast tumours and in some common cancer types. Br. J. Cancer 96, 646–653. doi: 10.1038/sj.bjc.6603579
Gao, H., Dong, Q., Chen, Y., Zhang, F., Wu, A., Shi, Y., et al. (2016). Murine mammary stem/progenitor cell isolation: different method matters? Springerplus 5:140. doi: 10.1186/s40064-016-1787-3
Gärtner, S., Gunesch, A., Knyazeva, T., Wolf, P., Högel, B., Eiermann, W., et al. (2014). PTK 7 is a transforming gene and prognostic marker for breast cancer and nodal metastasis involvement. PLoS One 9:e84472. doi: 10.1371/journal.pone.0084472
Gaspar, C., and Fodde, R. (2004). APC dosage effects in tumorigenesis and stem cell differentiation. Int. J. Dev. Biol. 48, 377–386. doi: 10.1387/ijdb.041807cg
Gaspar, C., Franken, P., Molenaar, L., Breukel, C., van der Valk, M., Smits, R., et al. (2009). A targeted constitutive mutation in the APC tumor suppressor gene underlies mammary but not intestinal tumorigenesis. PLoS Genet. 5:e1000547. doi: 10.1371/journal.pgen.1000547
Gerdur Ísberg, Ó., Kim, J., Fridriksdottir, A. J., Morsing, M., Timmermans-Wielenga, V., Rønnov-Jessen, L., et al. (2019). A CD146 FACS protocol enriches for luminal keratin 14/19 double positive human breast progenitors. Sci. Rep. 9:14843.
Ghosh, N., Hossain, U., Mandal, A., and Sil, P. C. (2019). The Wnt signaling pathway: a potential therapeutic target against cancer. Ann. N. Y. Acad. Sci. 1443, 54–74. doi: 10.1111/nyas.14027
Ginestier, C., Hur, M. H., Charafe-Jauffret, E., Monville, F., Dutcher, J., Brown, M., et al. (2007). ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell 1, 555–567. doi: 10.1016/j.stem.2007.08.014
Green, J. L., La, J., Yum, K. W., Desai, P., Rodewald, L. W., Zhang, X., et al. (2013). Paracrine Wnt signaling both promotes and inhibits human breast tumor growth. Proc. Natl. Acad. Sci. U.S.A. 110, 6991–6996. doi: 10.1073/pnas.1303671110
Grossmann, A. H., Yoo, J. H., Clancy, J., Sorensen, L. K., Sedgwick, A., Tong, Z., et al. (2013). The small GTPase ARF6 stimulates β-catenin transcriptional activity during WNT5A-mediated melanoma invasion and metastasis. Sci. Signal. 6:ra14. doi: 10.1126/scisignal.2003398
Gujral, T. S., Chan, M., Peshkin, L., Sorger, P. K., Kirschner, M. W., and Macbeath, G. (2014). A noncanonical frizzled2 pathway regulates epithelial-mesenchymal transition and metastasis. Cell 159, 844–856. doi: 10.1016/j.cell.2014.10.032
Gurney, A., Axelrod, F., Bond, C. J., Cain, J., Chartier, C., Donigan, L., et al. (2012). Wnt pathway inhibition via the targeting of Frizzled receptors results in decreased growth and tumorigenicity of human tumors. Proc. Natl. Acad. Sci. U.S.A. 109, 11717–11722. doi: 10.1073/pnas.1120068109
Hashizume, R., Koizumi, H., Ihara, A., Ohta, T., and Uchikoshi, T. (1996). Expression of β-catenin in normal breast tissue and breast carcinoma: a comparative study with epithelial cadherin and α-catenin. Histopathology 29, 139–146. doi: 10.1046/j.1365-2559.1996.d01-499.x
Hayes, M. J., Thomas, D., Emmons, A., Giordano, T. J., and Kleer, C. G. (2008). Genetic changes of Wnt pathway genes are common events in metaplastic carcinomas of the breast. Clin. Cancer Res. 14, 4038–4044. doi: 10.1158/1078-0432.CCR-07-4379
He, T. C., Sparks, A. B., Rago, C., Hermeking, H., Zawel, L., Da Costa, L. T., et al. (1998). Identification of c-MYC as a target of the APC pathway. Science 281, 1509–1512. doi: 10.1126/science.281.5382.1509
He, Y., Liu, Z., Qiao, C., Xu, M., Yu, J., and Li, G. (2014). Expression and significance of Wnt signaling components and their target genes in breast carcinoma. Mol. Med. Rep. 9, 137–143. doi: 10.3892/mmr.2013.1774
Hennessy, B. T., Gonzalez-Angulo, A. M., Stemke-Hale, K., Gilcrease, M. Z., Krishnamurthy, S., Lee, J. S., et al. (2009). Characterization of a naturally occurring breast cancer subset enriched in epithelial-to-mesenchymal transition and stem cell characteristics. Cancer Res. 69, 4116–4124. doi: 10.1158/0008-5472.CAN-08-3441
Henry, C., Quadir, A., Hawkins, N. J., Jary, E., Llamosas, E., Kumar, D., et al. (2015). Expression of the novel Wnt receptor ROR2 is increased in breast cancer and may regulate both β-catenin dependent and independent Wnt signalling. J. Cancer Res. Clin. Oncol. 141, 243–254. doi: 10.1007/s00432-014-1824-y
Holliday, D. L., and Speirs, V. (2011). Choosing the right cell line for breast cancer research. Breast Cancer Res. 13:215. doi: 10.1186/bcr2889
Hou, M.-F., Chen, P.-M., and Chu, P.-Y. (2018). LGR5 overexpression confers poor relapse-free survival in breast cancer patients. BMC Cancer 18:219. doi: 10.1186/s12885-018-4018-1
Huang, C., Ye, Z., Wan, J., Liang, J., Liu, M., Xu, X., et al. (2019). Secreted frizzled-related protein 2 is associated with disease progression and poor prognosis in breast cancer. Dis. Markers 2019:6149381.
Huch, M., Bonfanti, P., Boj, S. F., Sato, T., Loomans, C. J. M., van de Wetering, M., et al. (2013a). Unlimited in vitro expansion of adult bi-potent pancreas progenitors through the Lgr5/R-spondin axis. EMBO J. 32, 2708–2721. doi: 10.1038/emboj.2013.204
Huch, M., Dorrell, C., Boj, S. F., Van Es, J. H., Li, V. S. W., Van De Wetering, M., et al. (2013b). In vitro expansion of single Lgr5 + liver stem cells induced by Wnt-driven regeneration. Nature 494, 247–250. doi: 10.1038/nature11826
Huguet, E. L., McMahon, J. A., McMahon, A. P., Bicknell, R., and Harris, A. L. (1994). Differential expression of human Wnt genes 2, 3, 4, and 7B in human breast cell lines and normal and disease states of human breast tissue. Cancer Res. 54, 2615–2621.
Hung, T. H., Hsu, S. C., Cheng, C. Y., Choo, K. B., Tseng, C. P., Chen, T. C., et al. (2014). Wnt5A regulates ABCB1 expression in multidrug-resistant cancer cells through activation of the non-canonical PKA/β-catenin pathway. Oncotarget 5, 12273–12290.
Iozzo, R. V., Eichstetter, I., and Danielson, K. G. (1995). Aberrant expression of the growth factor WntSA in human malignancy. Cancer Res. 55, 3495–3499.
Ishizaki, Y., Ikeda, S., Fujimori, M., Shimizu, Y., Kurihara, T., Itamoto, T., et al. (2004). Immunohistochemical analysis and mutational analyses of beta-catenin, Axin family and APC genes in hepatocellular carcinomas. Int. J. Oncol. 24, 1077–1083.
Jacobsen, A., Heijmans, N., Verkaar, F., Smit, M. J., Heringa, J., van Amerongen, R., et al. (2016). Construction and experimental validation of a Petri net model of Wnt/β-catenin signaling. PLoS One 11:e0155743. doi: 10.1371/journal.pone.0155743
Jang, G.-B., Kim, J.-Y., Cho, S.-D., Park, K.-S., Jung, J.-Y., Lee, H.-Y., et al. (2015). Blockade of Wnt/β-catenin signaling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci. Rep. 5:12465. doi: 10.1038/srep12465
Jho, E.-H., Zhang, T., Domon, C., Joo, C.-K., Freund, J.-N., and Costantini, F. (2002). Wnt/ -catenin/Tcf signaling induces the transcription of Axin2, a negative regulator of the signaling pathway. Mol. Cell Biol. 22, 1172–1183. doi: 10.1128/mcb.22.4.1172-1183.2002
Jia, X. P., Meng, L. L., Fang, J. C., Wang, H. W., Chen, J., Zhou, J., et al. (2018). Aberrant expression of miR-142-3p and its target gene HMGA1 and FZD7 in breast cancer and its clinical significance. Clin. Lab. 64, 915–921. doi: 10.7754/Clin.Lab.2017.171114
Jiang, Q., He, M., Guan, S., Ma, M., Wu, H., Yu, Z., et al. (2016). MicroRNA-100 suppresses the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting Wnt/β-catenin signaling pathway. Tumor Biol. 37, 5001–5011. doi: 10.1007/s13277-015-4342-x
Jin, Z., Tamura, G., Tsuchiya, T., Sakata, K., Kashiwaba, M., Osakabe, M., et al. (2001). Adenomatous polyposis coli (APC) gene promoter hypermethylation in primary breast cancers. Br. J. Cancer 85, 69–73. doi: 10.1054/bjoc.2001.1853
Jonsson, M., Borg, Å., Nilbert, M., and Andersson, T. (2000). Involvement of adenomatous polyposis coli (APC)/β-catenin signalling in human breast cancer. Eur. J. Cancer 36, 242–248. doi: 10.1016/s0959-8049(99)00276-2
Jönsson, M., Dejmek, J., Bendahl, P. O., and Andersson, T. (2002). Loss of Wnt-5a protein is associated with early relapse in invasive ductal breast carcinomas. Cancer Res. 62, 409–416.
Karayiannakis, A. J., Nakopoulou, L., Gakiopoulou, H., Keramopoulos, A., Davaris, P. S., and Pignatelli, M. (2001). Expression patterns of β-catenin in in situ and invasive breast cancer. Eur. J. Surg. Oncol. 27, 31–36. doi: 10.1007/s10549-011-1860-0
Katoh, M. (2001). Frequent up-regulation of WNT2 in primary gastric cancer and colorectal cancer. Int. J. Oncol. 19, 1003–1007.
Khramtsov, A. I., Khramtsova, G. F., Tretiakova, M., Huo, D., Olopade, O. I., and Goss, K. H. (2010). Wnt/β-catenin pathway activation is enriched in basal-like breast cancers and predicts poor outcome. Am. J. Pathol. 176, 2911–2920. doi: 10.2353/ajpath.2010.091125
Kim, S. J., Garcia-Recio, S., Creighton, C. J., Perou, C. M., and Rosen, J. M. (2019). Alterations in Wnt- and/or STAT3 signaling pathways and the immune microenvironment during metastatic progression. Oncogene 38, 5942–5958. doi: 10.1038/s41388-019-0852-0
Klemm, F., Bleckmann, A., Siam, L., Chuang, H. N., Rietkötter, E., Behme, D., et al. (2011). β-catenin-independent WNT signaling in basal-like breast cancer and brain metastasis. Carcinogenesis 32, 434–442. doi: 10.1093/carcin/bgq269
Komiya, Y., and Habas, R. (2008). Wnt signal transduction pathways. Organogenesis 4, 68–75. doi: 10.4161/org.4.2.5851
Koren, S., Reavie, L., Couto, J. P., De Silva, D., Stadler, M. B., Roloff, T., et al. (2015). PIK3CAH1047R induces multipotency and multi-lineage mammary tumours. Nature 525, 114–118. doi: 10.1038/nature14669
Koval, A., and Katanaev, V. L. (2018). Dramatic dysbalancing of the Wnt pathway in breast cancers. Sci. Rep. 8:7329. doi: 10.1038/s41598-018-25672-6
Krishnamurthy, N., and Kurzrock, R. (2018). Targeting the Wnt/beta-catenin pathway in cancer: update on effectors and inhibitors. Cancer Treat. Rev. 62, 50–60. doi: 10.1016/j.ctrv.2017.11.002
Lacroix-Triki, M., Geyer, F. C., Lambros, M. B., Savage, K., Ellis, I. O., Lee, A. H. S., et al. (2010). β-Catenin/Wnt signalling pathway in fibromatosis, metaplastic carcinomas and phyllodes tumours of the breast. Mod. Pathol. 23, 1438–1448. doi: 10.1038/modpathol.2010.141
Lamb, R., Ablett, M. P., Spence, K., Landberg, G., Sims, A. H., and Clarke, R. B. (2013). Wnt pathway activity in breast cancer sub-types and stem-like cells. PLoS One 8:e67811. doi: 10.1371/journal.pone.0067811
Lathrop, A. E. C., and Loeb, L. (1916). On the part played by internal secretion in the spontaneous development of tumors. J. Cancer Res. 1, 1–19.
Le, P. N., McDermott, J. D., and Jimeno, A. (2015). Targeting the Wnt pathway in human cancers: therapeutic targeting with a focus on OMP-54F28. Pharmacol. Ther. 146, 1–11. doi: 10.1016/j.pharmthera.2014.08.005
Lejeune, S., Huguet, E. L., Hamby, A., Poulsom, R., and Harris, A. L. (1995). Wnt5a cloning, expression, and up-regulation in human primary breast cancers. Clin. Cancer Res. 1, 215–222.
Li, L., Ying, J., Tong, X., Zhong, L., Su, X., Xiang, T., et al. (2014). Epigenetic identification of receptor tyrosine kinase-like orphan receptor 2 as a functional tumor suppressor inhibiting β-catenin and AKT signaling but frequently methylated in common carcinomas. Cell. Mol. Life Sci. 71, 2179–2192. doi: 10.1007/s00018-013-1485-z
Lim, E., Wu, D., Pal, B., Bouras, T., Asselin-Labat, M.-L., Vaillant, F., et al. (2010). Transcriptome analyses of mouse and human mammary cell subpopulations reveal multiple conserved genes and pathways. Breast Cancer Res. 12:R21. doi: 10.1186/bcr2560
Lin, S. Y., Xia, W., Wang, J. C., Kwong, K. Y., Spohn, B., Wen, Y., et al. (2000). β-catenin, a novel prognostic marker for breast cancer: its roles in cyclin D1 expression and cancer progression. Proc. Natl. Acad. Sci. U.S.A. 97, 4262–4266. doi: 10.1073/pnas.060025397
Lindqvist, B. M., Wingren, S., Motlagh, P. B., and Nilsson, T. K. (2014). Whole genome DNA methylation signature of HER2-positive breast cancer. Epigenetics 9, 1149–1162. doi: 10.4161/epi.29632
Lindvall, C., Zylstra, C. R., Evans, N., West, R. A., Dykema, K., Furge, K. A., et al. (2009). The Wnt co-receptor Lrp6 is required for normal mouse mammary gland development. PLoS One 4:e5813. doi: 10.1371/journal.pone.0005813
Linnemann, J. R., Meixner, L. K., Miura, H., and Scheel, C. H. (2017). An organotypic 3D assay for primary human mammary epithelial cells that recapitulates branching morphogenesis. Methods Mol. Biol. 1612, 125–137. doi: 10.1007/978-1-4939-7021-6_9
Linnemann, J. R., Miura, H., Meixner, L. K., Irmler, M., Kloos, U. J., Hirschi, B., et al. (2015). Quantification of regenerative potential in primary human mammary epithelial cells. Development 142, 3239–3251. doi: 10.1242/dev.123554
Liu, C.-C., Prior, J., Piwnica-Worms, D., and Bu, G. (2010). LRP6 overexpression defines a class of breast cancer subtype and is a target for therapy. Proc. Natl. Acad. Sci. U.S.A. 107, 5136–5141. doi: 10.1073/pnas.0911220107
Liu, J., Pan, S., Hsieh, M. H., Ng, N., Sun, F., Wang, T., et al. (2013). Targeting Wnt-driven cancer through the inhibition of porcupine by LGK974. Proc. Natl. Acad. Sci. U.S.A. 110, 20224–20229. doi: 10.1073/pnas.1314239110
Liu, Z., Jiang, Z., Gao, Y., Wang, L., Chen, C., and Wang, X. (2019). TP53 mutations promote immunogenic activity in breast cancer. J. Oncol. 2019:5952836. doi: 10.1155/2019/5952836
Loh, K. M., van Amerongen, R., and Nusse, R. (2016). Generating cellular diversity and spatial form: Wnt signaling and the evolution of multicellular animals. Dev. Cell 38, 643–655. doi: 10.1016/j.devcel.2016.08.011
Lustig, B., Jerchow, B., Sachs, M., Weiler, S., Pietsch, T., Rarsten, U., et al. (2001). Negative feedback loop of Wnt signaling through upregulation of conductin/axin2 in colorectal and liver tumors. Langenbecks Arch. Surg. 386:466.
Ma, J., Lu, W., Chen, D., Xu, B., and Li, Y. (2017). Role of Wnt co-receptor LRP6 in triple negative breast cancer cell migration and invasion. J. Cell. Biochem. 118, 2968–2976. doi: 10.1002/jcb.25956
MacDonald, B. T., Tamai, K., and He, X. (2009). Wnt/β-catenin signaling: components, mechanisms, and diseases. Dev. Cell 17, 9–26. doi: 10.1016/j.devcel.2009.06.016
Madan, B., Ke, Z., Harmston, N., Ho, S. Y., Frois, A. O., Alam, J., et al. (2016). Wnt addiction of genetically defined cancers reversed by PORCN inhibition. Oncogene 35, 2197–2207. doi: 10.1038/onc.2015.280
Madan, B., McDonald, M. J., Foxa, G. E., Diegel, C. R., Williams, B. O., and Virshup, D. M. (2018). Bone loss from Wnt inhibition mitigated by concurrent alendronate therapy. Bone Res. 6:17. doi: 10.1038/s41413-018-0017-8
Many, A. M., and Brown, A. M. C. (2014). Both canonical and non-canonical Wnt signaling independently promote stem cell growth in mammospheres. PLoS One 9:e101800. doi: 10.1371/journal.pone.0101800
Martin, T. A., Watkins, G., and Jiang, W. G. (2005). KiSS-1 expression in human breast cancer. Clin. Exp. Metastasis 22, 503–511. doi: 10.1007/s10585-005-4180-0
McNally, S., and Stein, T. (2017). Overview of mammary gland development: a comparison of mouse and human. Methods Mol. Biol. 1501, 1–17. doi: 10.1007/978-1-4939-6475-8_1
Mikels, A. J., and Nusse, R. (2006). Purified Wnt5a protein activates or inhibits β-catenin-TCF signaling depending on receptor context. PLoS Biol. 4:e115. doi: 10.1371/journal.pbio.0040115
Milovanovic, T., Planutis, K., Nguyen, A., Marsh, J. L., Lin, F., Hope, C., et al. (2004). Expression of Wnt genes and frizzled 1 and 2 receptors in normal breast epithelium and infiltrating breast carcinoma. Int. J. Oncol. 25, 1337–1342.
Morin, P. J. (1997). Activation of beta -catenin-Tcf signaling in colon cancer by mutations in beta -catenin or APC. Science 275, 1787–1790. doi: 10.1126/science.275.5307.1787
Nakamura, Y., De Paiva Alves, E., Veenstra, G. J. C., and Hoppler, S. (2016). Tissue-and stage-specific Wnt target gene expression is controlled subsequent to β-catenin recruitment to cis-regulatory modules. Development 143, 1914–1925. doi: 10.1242/dev.131664
Ng, M., Tan, D. S., Subbiah, V., Weekes, C. D., Teneggi, V., Diermayr, V., et al. (2017). First-in-human phase 1 study of ETC-159 an oral PORCN inhbitor in patients with advanced solid tumours. J. Clin. Oncol. 35, 2584–2584. doi: 10.1200/jco.2017.35.15_suppl.2584
Nusse, R., and Clevers, H. (2017). Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell 169, 985–999. doi: 10.1016/j.cell.2017.05.016
Nusse, R., and Varmus, H. (2012). Three decades of Wnts: a personal perspective on how a scientific field developed. EMBO J. 31, 2670–2684. doi: 10.1038/emboj.2012.146
Nusse, R., and Varmus, H. E. (1982). Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 31, 99–109. doi: 10.1016/0092-8674(82)90409-3
Ozaki, S., Ikeda, S., Ishizaki, Y., Kurihara, T., Tokumoto, N., Iseki, M., et al. (2005). Alterations and correlations of the components in the Wnt signaling pathway and its target genes in breast cancer. Oncol. Rep. 14, 1437–1443.
Pereira, B., Chin, S.-F., Rueda, O. M., Vollan, H.-K. M., Provenzano, E., Bardwell, H. A., et al. (2016). The somatic mutation profiles of 2,433 breast cancers refine their genomic and transcriptomic landscapes. Nat. Commun. 7:11479.
Plaks, V., Brenot, A., Lawson, D. A., Linnemann, J. R., Van Kappel, E. C., Wong, K. C., et al. (2013). Lgr5-expressing cells are sufficient and necessary for postnatal mammary gland organogenesis. Cell Rep. 3, 70–78. doi: 10.1016/j.celrep.2012.12.017
Prasad, C. P., Gupta, S. D., Rath, G., and Ralhan, R. (2008a). Wnt signaling pathway in invasive ductal carcinoma of the breast: relationship between β-catenin, disheveled and cyclin D1 expression. Oncology 73, 112–117. doi: 10.1159/000120999
Prasad, C. P., Mirza, S., Sharma, G., Prashad, R., DattaGupta, S., Rath, G., et al. (2008b). Epigenetic alterations of CDH1 and APC genes: relationship with activation of Wnt/β-catenin Pathway in invasive ductal carcinoma of breast. Life Sci. 83, 318–325. doi: 10.1016/j.lfs.2008.06.019
Prater, M., Shehata, M., Watson, C. J., and Stingl, J. (2013). Enzymatic dissociation, flow cytometric analysis, and culture of normal mouse mammary tissue. Methods Mol. Biol. 946, 395–409. doi: 10.1007/978-1-62703-128-8_25
Radovich, M., Solzak, J., Hancock, B., Storniolo, A., Schneider, B., and Miller, K. (2019). Abstract OT3-06-02: an initial safety study of gedatolisib plus PTK7-ADC for metastatic triple-negative breast cancer. Cancer Res. 79(Suppl. 4): OT3-06-02-OT3-06-02.
Raman, S., Beilschmidt, M., To, M., Lin, K., Lui, F., Jmeian, Y., et al. (2019). Structure-guided design fine-tunes pharmacokinetics, tolerability, and antitumor profile of multispecific frizzled antibodies. Proc. Natl. Acad. Sci. U.S.A. 116, 6812–6817. doi: 10.1073/pnas.1817246116
Rao, D. M., Bordeaux, E. K., Yamamoto, T. M., Bitler, B. G., and Sikora, M. J. (2018). WNT4 and WNT3A activate cell autonomous Wnt signaling independent of secretion. bioRxiv [Preprint]. doi: 10.1101/333906
Riley, R. S., and Day, E. S. (2017). Frizzled7 antibody-functionalized nanoshells enable multivalent binding for Wnt signaling inhibition in triple negative breast cancer cells. Small 13:1700544. doi: 10.1002/smll.201700544
Roarty, K., Shore, A. N., Creighton, C. J., and Rosen, J. M. (2015). Ror2 regulates branching, differentiation, and actin-cytoskeletal dynamics within the mammary epithelium. J. Cell Biol. 208, 351–366. doi: 10.1083/jcb.201408058
Robey, R. W., Pluchino, K. M., Hall, M. D., Fojo, A. T., Bates, S. E., and Gottesman, M. M. (2018). Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 18, 452–464. doi: 10.1038/s41568-018-0005-8
Sachs, N., de Ligt, J., Kopper, O., Gogola, E., Bounova, G., Weeber, F., et al. (2018). A living biobank of breast cancer organoids captures disease heterogeneity. Cell 172, 373–386.e10. doi: 10.1016/j.cell.2017.11.010
Säfholm, A., Tuomela, J., Rosenkvist, J., Dejmek, J., Härkönen, P., and Andersson, T. (2008). The Wnt-5a-derived hexapeptide Foxy-5 inhibits breast cancer metastasis in vivo by targeting cell motility. Clin. Cancer Res. 14, 6556–6563. doi: 10.1158/1078-0432.CCR-08-0711
Santagata, S., Thakkar, A., Ergonul, A., Wang, B., Woo, T., Hu, R., et al. (2014). Taxonomy of breast cancer based on normal cell phenotype predicts outcome. J. Clin. Invest. 124, 859–870. doi: 10.1172/JCI70941
Sato, T., Vries, R. G., Snippert, H. J., Van De Wetering, M., Barker, N., Stange, D. E., et al. (2009). Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459, 262–265. doi: 10.1038/nature07935
Scheele, C. L. G. J., Hannezo, E., Muraro, M. J., Zomer, A., Langedijk, N. S. M., Van Oudenaarden, A., et al. (2017). Identity and dynamics of mammary stem cells during branching morphogenesis. Nature 542, 313–317. doi: 10.1038/nature21046
Schenkelaars, Q., Pratlong, M., Kodjabachian, L., Fierro-Constain, L., Vacelet, J., Le Bivic, A., et al. (2017). Animal multicellularity and polarity without Wnt signaling. Sci. Rep. 7:15383. doi: 10.1038/s41598-017-15557-5
Shabani, M., Naseri, J., and Shokri, F. (2015). Receptor tyrosine kinase-like orphan receptor 1: a novel target for cancer immunotherapy. Expert Opin. Ther. Targets 19, 941–955. doi: 10.1517/14728222.2015.1025753
Shackleton, M., Vaillant, F., Simpson, K. J., Stingl, J., Smyth, G. K., Asselin-Labat, M. L., et al. (2006). Generation of a functional mammary gland from a single stem cell. Nature 439, 84–88.
Shaw, F. L., Harrison, H., Spence, K., Ablett, M. P., Simões, B. M., Farnie, G., et al. (2012). A detailed mammosphere assay protocol for the quantification of breast stem cell activity. J. Mammary Gland Biol. Neoplasia 17, 111–117. doi: 10.1007/s10911-012-9255-3
Shtutman, M., Zhurinsky, J., Simcha, I., Albanese, C., D’Amico, M., Pestell, R., et al. (1999). The cyclin D1 gene is a target of the β-catenin/LEF-1 pathway. Proc. Natl. Acad. Sci. U.S.A. 96, 5522–5527. doi: 10.1073/pnas.96.10.5522
Sormunen, R. T., Leong, A. S., Vääräniemi, J. P., Fernando, S. S., and Eskelinen, S. M. (1999). Immunolocalization of the fodrin, E-cadherin, and β-catenin adhesion complex in infiltrating ductal carcinoma of the breast-comparison with an in vitro model. J. Pathol. 187, 416–423. doi: 10.1002/(sici)1096-9896(199903)187:4<416::aid-path255>3.0.co;2-d
Stephens, P. J., Tarpey, P. S., Davies, H., Van Loo, P., Greenman, C., Wedge, D. C., et al. (2012). The landscape of cancer genes and mutational processes in breast cancer. Nature 486, 400–404. doi: 10.1038/nature11017
Stingl, J., Eirew, P., Ricketson, I., Shackleton, M., Vaillant, F., Choi, D., et al. (2006). Purification and unique properties of mammary epithelial stem cells. Nature 439, 993–997. doi: 10.1038/nature04496
Sun, X., Sandhu, R., Figueroa, J. D., Gierach, G. L., Sherman, M. E., and Troester, M. A. (2014). Benign breast tissue composition in breast cancer patients: association with risk factors, clinical variables, and gene expression. Cancer Epidemiol Biomarkers Prev. 23, 2810–2818. doi: 10.1158/1055-9965.EPI-14-0507
Suzuki, H., Toyota, M., Caraway, H., Gabrielson, E., Ohmura, T., Fujikane, T., et al. (2008). Frequent epigenetic inactivation of Wnt antagonist genes in breast cancer. Br. J. Cancer 98, 1147–1156. doi: 10.1038/sj.bjc.6604259
Suzuki, H., Watkins, D. N., Jair, K. W., Schuebel, K. E., Markowitz, S. D., Chen, W. D., et al. (2004). Epigenetic inactivation of SFRP genes allows constitutive WNT signaling in colorectal cancer. Nat. Genet. 36, 417–422. doi: 10.1038/ng1330
Tan, D., Ng, M., Subbiah, V., Messersmith, W., Teneggi, V., Diermayr, V., et al. (2018). 71OPhase I extension study of ETC-159 an oral PORCN inhibitor administered with bone protective treatment, in patients with advanced solid tumours. Ann. Oncol. 29(Suppl._9):mdy430.002.
Trifa, F., Karray-Chouayekh, S., Jmal, E., Jmaa, Z., Ben, Khabir, A., et al. (2013). Loss of WIF-1 and Wnt5a expression is related to aggressiveness of sporadic breast cancer in Tunisian patients. Tumor Biol. 34, 1625–1633. doi: 10.1007/s13277-013-0694-2
Tsai, C. H., Chiu, J. H., Yang, C. W., Wang, J. Y., Tsai, Y. F., Tseng, L. M., et al. (2015). Molecular characteristics of recurrent triple-negative breast cancer. Mol. Med. Rep. 12, 7326–7334. doi: 10.3892/mmr.2015.4360
Ugolini, F., Charafe-Jauffret, E., Bardou, V. J., Geneix, J., Adélaïde, J., Labat-Moleur, F., et al. (2001). WNT pathway and mammary carcinogenesis: loss of expression of candidate tumor suppressor gene SFRP1 in most invasive carcinomas except of the medullary type. Oncogene 20, 5810–5817. doi: 10.1038/sj.onc.1204706
van Amerongen, R. (2012). Alternative Wnt pathways and receptors. Cold Spring Harb. Perspect. Biol. 4:a007914. doi: 10.1101/cshperspect.a007914
van Amerongen, R. (2015). Lineage tracing in the mammary gland using cre/lox technology and fluorescent reporter alleles. Methods Mol. Biol. 1293, 187–211. doi: 10.1007/978-1-4939-2519-3_11
van Amerongen, R., Bowman, A. N. N., Nusse, R., Van Amerongen, R., Bowman, A. N. N., Nusse, R., et al. (2012a). Developmental stage and time dictate the fate of Wnt/β-catenin- responsive stem cells in the mammary gland. Cell Stem Cell 11, 387–400. doi: 10.1016/j.stem.2012.05.023
van Amerongen, R., Fuerer, C., Mizutani, M., and Nusse, R. (2012b). Wnt5a can both activate and repress Wnt/β-catenin signaling during mouse embryonic development. Dev. Biol. 369, 101–114. doi: 10.1016/j.ydbio.2012.06.020
van Amerongen, R., and Nusse, R. (2009). Towards an integrated view of Wnt signaling in development. Development 136, 3205–3214. doi: 10.1242/dev.033910
van de Moosdijk, A. A. A., Fu, N. Y., Rios, A. C., Visvader, J. E., and van Amerongen, R. (2017). Lineage tracing of mammary stem and progenitor cells. Methods Mol. Biol. 1501, 291–308.
Van Der Auwera, I., Van Laere, S. J., Van Den Bosch, S. M., Van Den Eynden, G. G., Trinh, B. X., Van Dam, P. A., et al. (2008). Aberrant methylation of the Adenomatous Polyposis Coli (APC) gene promoter is associated with the inflammatory breast cancer phenotype. Br. J. Cancer 99, 1735–1742. doi: 10.1038/sj.bjc.6604705
Van Keymeulen, A., Lee, M. Y., Ousset, M., Brohée, S., Rorive, S., Giraddi, R. R., et al. (2015). Reactivation of multipotency by oncogenic PIK3CA induces breast tumour heterogeneity. Nature 525, 119–123. doi: 10.1038/nature14665
Van Keymeulen, A., Rocha, A. S., Ousset, M., Beck, B., Bouvencourt, G., Rock, J., et al. (2011). Distinct stem cells contribute to mammary gland development and maintenance. Nature 479, 189–193. doi: 10.1038/nature10573
van Neerven, S. M., and Vermeulen, L. (2019). The interplay between intrinsic and extrinsic Wnt signaling in controlling intestinal transformation. Differentiation 108, 17–23. doi: 10.1016/j.diff.2019.02.002
VanderVorst, K., Hatakeyama, J., Berg, A., Lee, H., and Carraway, K. L. (2018). Cellular and molecular mechanisms underlying planar cell polarity pathway contributions to cancer malignancy. Semin. Cell Dev. Biol. 81, 78–87. doi: 10.1016/j.semcdb.2017.09.026
Veeck, J., Geisler, C., Noetzel, E., Alkaya, S., Hartmann, A., Knüchel, R., et al. (2008a). Epigenetic inactivation of the secreted frizzled-related protein-5 (SFRP5) gene in human breast cancer is associated with unfavorable prognosis. Carcinogenesis 29, 991–998. doi: 10.1093/carcin/bgn076
Veeck, J., Niederacher, D., An, H., Klopocki, E., Wiesmann, F., Betz, B., et al. (2006). Aberrant methylation of the Wnt antagonist SFRP1 in breast cancer is associated with unfavourable prognosis. Oncogene 25, 3479–3488. doi: 10.1038/sj.onc.1209386
Veeck, J., Noetzel, E., Bektas, N., Jost, E., Hartmann, A., Knüchel, R., et al. (2008b). Promoter hypermethylation of the SFRP2 gene is a high-frequent alteration and tumor-specific epigenetic marker in human breast cancer. Mol. Cancer 7:83. doi: 10.1186/1476-4598-7-83
Veeck, J., Wild, P. J., Fuchs, T., Schüffler, P. J., Hartmann, A., Knüchel, R., et al. (2009). Prognostic relevance of Wnt-inhibitory factor-1 (WIF1) and Dickkopf-3 (DKK3) promoter methylation in human breast cancer. BMC Cancer 9:217. doi: 10.1186/1471-2407-9-217
Veltmaat, J. M., Van Veelen, W., Thiery, J. P., and Bellusci, S. (2004). Identification of the mammary line in mouse by Wnt10b expression. Dev. Dyn. 229, 349–356. doi: 10.1002/dvdy.10441
Wang, D., Cai, C., Dong, X., Yu, Q. C., Zhang, X. O., Yang, L., et al. (2015). Identification of multipotent mammary stemcells by protein C receptor expression. Nature 517, 81–84. doi: 10.1038/nature13851
Wang, Y.-Z., Han, Y.-S., Ma, Y.-S., Jiang, J.-J., Chen, Z.-X., Wang, Y.-C., et al. (2012). Differential gene expression of Wnt signaling pathway in benign, premalignant, and malignant human breast epithelial cells. Tumor Biol. 33, 2317–2327. doi: 10.1007/s13277-012-0494-0
Watanabe, O., Imamura, H., Shimizu, T., Kinoshita, J., Okabe, T., Hirano, A., et al. (2004). Expression of twist and Wnt in human breast cancer. Anticancer Res. 24, 3851–3856.
Wellenstein, M. D., Coffelt, S. B., Duits, D. E. M., van Miltenburg, M. H., Slagter, M., de Rink, I., et al. (2019). Loss of p53 triggers WNT-dependent systemic inflammation to drive breast cancer metastasis. Nature 572, 538–542. doi: 10.1038/s41586-019-1450-6
Wend, P., Runke, S., Wend, K., Anchondo, B., Yesayan, M., Jardon, M., et al. (2013). WNT10B/β-catenin signalling induces HMGA2 and proliferation in metastatic triple-negative breast cancer. EMBO Mol. Med. 5, 264–279. doi: 10.1002/emmm.201201320
White, B. D., Chien, A. J., and Dawson, D. W. (2012). Dysregulation of Wnt/β-catenin signaling in gastrointestinal cancers. Gastroenterology 142, 219–232. doi: 10.1053/j.gastro.2011.12.001
Wiese, K. E., Nusse, R., and van Amerongen, R. (2018). Wnt signalling: conquering complexity. Development 145:dev165902. doi: 10.1242/dev.165902
Wissman, C., Wild, P. J., Kaiser, S., Roepcke, S., Stoehr, R., Woenckhaus, M., et al. (2003). WIFI, a component of the Wnt pathway, is down-regulated in prostate, breast, lung, and bladder cancer. J. Pathol. 201, 204–212. doi: 10.1002/path.1449
Wong, S. C. C., Lo, S. F. E., Lee, K. C., Yam, J. W. P., Chan, J. K. C., and Wendy Hsiao, W. L. (2002). Expression of frizzled-related protein and Wnt-signalling molecules in invasive human breast tumours. J. Pathol. 196, 145–153. doi: 10.1002/path.1035
Wronski, A., Arendt, L. M., and Kuperwasser, C. (2015). Humanization of the mouse mammary gland. Methods Mol. Biol. 1293, 173–186. doi: 10.1007/978-1-4939-2519-3_10
Wuidart, A., Ousset, M., Rulands, S., Simons, B. D., Van Keymeulen, A., and Blanpain, C. (2016). Quantitative lineage tracing strategies to resolve multipotency in tissue-specific stem cells. Genes Dev. 30, 1261–1277. doi: 10.1101/gad.280057.116
Wuidart, A., Sifrim, A., Fioramonti, M., Matsumura, S., Brisebarre, A., Brown, D., et al. (2018). Early lineage segregation of multipotent embryonic mammary gland progenitors. Nat. Cell Biol. 20, 666–676. doi: 10.1038/s41556-018-0095-2
Yamaguchi, T., Mukai, H., Yamashita, S., Fujii, S., and Ushijima, T. (2015). Comprehensive DNA methylation and extensive mutation analyses of HER2-positive breast cancer. Oncology 88, 377–384. doi: 10.1159/000369904
Yanagisawa, A., Tsuchiya, E., Kato, Y., Ichii, S., Nagase, H., Ando, H., et al. (1992). Frequent somatic mutations of the APC gene in human pancreatic cancer. Cancer Res. 52, 6696–6698.
Yang, L., Wu, X., Wang, Y., Zhang, K., Wu, J., Yuan, Y. C., et al. (2011). FZD7 has a critical role in cell proliferation in triple negative breast cancer. Oncogene 30, 4437–4446. doi: 10.1038/onc.2011.145
Yeo, E. J., Cassetta, L., Qian, B. Z., Lewkowich, I., Li, J. F., Stefater, J. A., et al. (2014). Myeloid Wnt7b mediates the angiogenic switch and metastasis in breast cancer. Cancer Res. 74, 2962–2973. doi: 10.1158/0008-5472.CAN-13-2421
Yi, K., Min, K. W., Wi, Y. C., Kim, Y., Shin, S. J., Chung, M. S., et al. (2017). Wnt7a deficiency could predict worse disease-free and overall survival in estrogen receptor-positive breast cancer. J. Breast Cancer 20, 361–367. doi: 10.4048/jbc.2017.20.4.361
Yu, Q., Verheyen, E., and Zeng, Y. (2016). Mammary development and breast cancer: a Wnt perspective. Cancers 8:E65. doi: 10.3390/cancers8070065
Zeljko, M., Pecina-Slaus, N., Martic, T. N., Kusec, V., Beros, V., and Tomas, D. (2011). Molecular alterations of E-cadherin and beta-catenin in brain metastases. Front. Biosci. 3, 616–624. doi: 10.2741/e274
Zeng, Y. A., and Nusse, R. (2010). Wnt proteins are self-renewal factors for mammary stem cells and promote their long-term expansion in culture. Cell Stem Cell 6, 568–577. doi: 10.1016/j.stem.2010.03.020
Zhang, C., Henner, W. R., Wang, M., Cattaruzza, F., Yeung, P., O’Young, G., et al. (2018). “Abstract A030: biomarker study of vantictumab plus paclitaxel in HER2- breast cancer patients,” in Proceedings of the AACR-NCI-EORTC International Conference: Molecular Targets and Cancer Therapeutics, Philadelphia, PA.
Zhang, L., and Shay, J. W. (2017). Multiple roles of APC and its therapeutic implications in colorectal cancer. J. Natl. Cancer Inst. 109:djw332.
Zhang, S., Chen, L., Cui, B., Chuang, H. Y., Yu, J., Wang-Rodriguez, J., et al. (2012). ROR1 is expressed in human breast cancer and associated with enhanced tumor-cell growth. PLoS One 7:e31127. doi: 10.1371/journal.pone.0031127
Zhang, S., Wu, C. C. N., Fecteau, J. F., Cui, B., Chen, L., Zhang, L., et al. (2013). Targeting chronic lymphocytic leukemia cells with a humanized monoclonal antibody specific for CD44. Proc. Natl. Acad. Sci. U.S.A. 110, 6127–6132. doi: 10.1073/pnas.1221841110
Zhang, S., Zhang, H., Ghia, E. M., Huang, J., Wu, L., Zhang, J., et al. (2019). Inhibition of chemotherapy resistant breast cancer stem cells by a ROR1 specific antibody. Proc. Natl. Acad. Sci. U.S.A. 116, 1370–1377. doi: 10.1073/pnas.1816262116
Zhong, Z., Shan, M., Wang, J., Liu, T., Shi, Q., and Pang, D. (2016). Decreased Wnt5a expression is a poor prognostic factor in triple-negative breast cancer. Med. Sci. Monit. 22, 1–7. doi: 10.12659/msm.894821
Keywords: canonical Wnt signaling, non-canonical Wnt signaling, beta-catenin, breast cancer, mammary gland, stem cells, cancer stem cells
Citation: van Schie EH and van Amerongen R (2020) Aberrant WNT/CTNNB1 Signaling as a Therapeutic Target in Human Breast Cancer: Weighing the Evidence. Front. Cell Dev. Biol. 8:25. doi: 10.3389/fcell.2020.00025
Received: 30 September 2019; Accepted: 14 January 2020;
Published: 31 January 2020.
Edited by:
Vida Vafaizadeh, University of Basel, SwitzerlandReviewed by:
Jeffrey M. Rosen, Baylor College of Medicine, United StatesCopyright © 2020 van Schie and van Amerongen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renée van Amerongen, ci52YW5hbWVyb25nZW5AdXZhLm5s
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.