 Konshau W. Duman
Konshau W. Duman Thomas P. Hahn
Thomas P. Hahn- Animal Behavior Graduate Group, Neurobiology, Physiology, and Behavior Department, University of California Davis, Davis, CA, United States
The Evening Grosbeak (Coccothraustes vesperitinus Cooper, 1825) is a species with multiple vocally distinct recognized groups known as call types. The range boundaries and degrees of overlap between these groups remain an area of continuing research. We report sympatric breeding occurrence of Evening Grosbeaks of call type 1 and call type 4 in the Jackson Hole area of Wyoming, and we show that these two call types have been overlapping in this region for at least 24 years. We also note a major increase in the local abundance of Evening Grosbeaks in 2023 compared to past years. These findings suggest that the range boundary between these two call types is relatively consistent at this short time scale, but it may fluctuate somewhat given changes in the proportions of the two types over this period. We detected both call types continuously through the 2023 breeding period in the Jackson Hole area, and we had breeding season detections of both types in the area in 2000, 2005, 2006, 2007, 2008, 2010, and 2012. In 2023, a majority were the Pacific Northwest-centered type 1 while roughly a quarter were type 4 which is known from the Central Rockies. In 2023, we generally saw type 4 individuals alongside type 1 individuals, and both call types tended to cluster in the same areas though type 1 birds were frequently detected without any type 4s nearby. We most frequently encountered both types in Douglas Fir (Pseudotsuga menziesii Mirb. Franco) or Subalpine Fir (Abies lasiocarpa (Hook.) Nutt.) dominated forests through the June–July breeding season, and both were abundant in areas with Chokecherry (Prunus virginiana L.) or Serviceberry (Amelanchier alnifolia (Nutt.) Nutt. Ex M.Roem.) after the breeding season. Despite considerable survey effort, we could not detect either type north of Moran, WY. Describing this close overlap of call types with uncertain taxonomic status paves the way for further research investigating the potential for ecological and genetic isolation of the two groups in areas where mixed parentage is most possible.
1 Introduction
Like other nomadic finches, Evening Grosbeaks present a challenge to mapping spatial distributions of subspecies that are essentially impossible to identify visually. Variable movements and changes in breeding density from year to year (Gillihan and Byers, 2001; Robinson et al., 2022; this study) prevent clear definition of ranges and abundances, and this irregular distribution combined with the cryptic habits of birds in the breeding season (Bekoff et al., 1989) makes it difficult to study breeding distribution and behaviors. This challenge has prevented clear geographic definition of the ranges and overlap zones of the subspecies and call types of Evening Grosbeak, a critical step for assessing taxonomic status of these groups (James, 2010). It is also necessary to assess assortative mating rates and cohesiveness of evolutionary lineages of these forms in such overlap zones to determine their taxonomic status. Discovery that the subspecies of Evening Grosbeaks may correspond to readily field-identifiable call types (Sewall et al., 2004; Robinson et al., 2024; this study) presents the possibility of greatly improved range mapping and assessment of assortative mating (or lack thereof) in areas of overlap.
In the earliest description of infraspecific variation in the Evening Grosbeak, Ridgway defined subspecies C.v. montanus from Mexico (Baird et al., 1874). This was later extended to include all Western North America after Mearns (1890) examined additional specimens from British Columbia, Washington, Oregon, California, Utah, Arizona, New Mexico, the Valley of Mexico, and Veracruz. Grinnell (1917) subsequently split the Western forms into 4 subspecies according to differences in bill proportions, overall darkness of plumage, and width of the yellow frontal bar in the male. In this treatment, the nominate subspecies was assumed to breed in the boreal forest east of the main divide of the Rockies, C.v. montanus in Mexico and extreme SE Arizona, C.v. brooksi west of the divide in British Columbia and Washington, C.v. californicus in the Sierra Nevada and Oregon Cascades, and C.v. warreni in the Rockies of Colorado, New Mexico, and N. Arizona. The latter three of these later appeared lumped together on the AOU Checklist (Wetmore et al., 1957), leaving three subspecies recognized today with their ranges roughly in Mexico, the Western USA and Canada, and the Eastern USA and Canada.
The flight calls of Evening Grosbeaks (produced perched or in flight and with a clear up–down quality relative to their other calls) fall into 5 vocally distinct groups, referred to as call types, whose ranges may correspond with Grinnell’s 5 subspecies (Sewall et al., 2004), including the two now unrecognized subspecies C.v. warreni and C.v. californicus. Despite the highly nomadic tendencies of types 1 and 3 in the non-breeding season and some degree of variable breeding distribution, the core breeding ranges of these call types appear to have remained consistent for at least the past twenty years (eBird, 2023; this study). Call type 1 (possibly C.v. brooksi sensu stricto) appears to occur in the breeding season across the Pacific Northwest and Northern Rockies, with its southern limits in the Cascades and Central Rockies still poorly understood. Type 1 is also an irregular migrant throughout the western United States in the non-breeding season, and it is recorded farther from its breeding range than the other western call types. Call type 2 (possibly the former C.v. californicus) primarily breeds in the Sierra Nevada of California, with some breeding as far north as the Cascades of Central Oregon, and it is recorded occasionally outside this range in winter, mostly within California. Call type 3 (C.v. vespertinus) breeds in the eastern boreal forest, with its western limits in Alberta unknown. It irregularly moves south across most of the eastern United States. Call type 4 (possibly the former C.v. warreni) has its core range in the Central Rockies. The northern limit of call type 4 and the southern limit of call type 1 remain ambiguous, and both limits were thought to lie somewhere in Southern Wyoming (see Sewall et al., 2004 – Figure 2), with the extent of overlap remaining unknown. The Jackson Hole area (Southern Teton County) in Western Wyoming represents the site of the northern-most recordings of call type 4 in Sewall et al. (2004), all but one of which were in the post-breeding season (August–September).
The Evening Grosbeak lacks an obvious song. It is somewhat unusual among passerines in that it produces the same few short calls across the entire annual cycle without any long or melodious vocalization at any time of year. However, emerging observational evidence suggests that the flight call may be used as a sort of dawn song during the breeding season, when it is sometimes produced in a continuous repetitive pattern from a prominent perch (M. A. Young pers. comm.; pers. obs.). This could result in the call types being reproductively isolated by breeding behaviors coordinated by song as well as by flocking and mate choice based on flight calls. The fact that these calls appear to remain discrete in overlap zones such as Western Wyoming despite apparently overlapping ecology presents an interesting system to assess the potential for vocal groups to lead to cryptic diversity. Rapid behavioral evolution of vocalizations has been shown to track bursts of speciation in both oscines and suboscines, with oscines showing faster radiations in vocal phenotype and a high potential for rapid evolution driven by plasticity in vocalizations (Mason et al., 2017). Songs in particular can act as barriers to gene flow between populations (Lipshutz et al., 2017), so the potential dual function of the Evening Grosbeak flight call could lead to behavioral isolation in multiple contexts. It is thus critical to study vocal behavior after the reunion of formerly allopatric groups to determine whether breeding isolation has arisen and if it is causing pre-mating barriers to hybridization via fixation of vocal phenotypes (Grant and Grant, 1996). This mode of vocal evolution may apply to Evening Grosbeak call types since they have core ranges where exclusively one call type occurs in the breeding season and overlap areas at the margins of these core ranges, with calls remaining similar across these ranges. This pattern of occurrence is suggestive of historic allopatry and subsequent contact at the range margins, not unlike other western mountain species (cf. range and mapped glacial refugia for subspecies of Steller’s Jay (Cyanocitta stelleri Gmelin, 1788) (Cicero et al., 2022)).
The Evening Grosbeak system provides an excellent opportunity to examine the relationship between vocal groups and taxonomic groups. This is because the call types may align well with subspecies previously described, the core ranges of the call types are relatively consistent, and the calls that differ between the call types are likely to be involved in breeding behaviors and mate selection in addition to flocking behaviors, with no separate song for these purposes. This connection has been elusive in other cardueline finches which generally use flight calls for flocking and individual recognition (Mundinger, 1970). The Red Crossbill (Loxia curvirostra Linnaeus, 1758) system, for example, has numerous forms which differ in having discrete differences in flight calls (Groth, 1993). However, the nomadic tendencies of Crossbills with breeding occurring in widely separated regions from year to year (Newton, 2006) has prevented sufficient study of breeding isolation except in the most sedentary call type, which was found to have very high rates of assortative mating and is currently considered a separate species, Loxia sinesciurus Benkman, 2009 (Benkman et al., 2009; Chesser et al., 2017). The Red Crossbill also has highly complex and varied songs which are used when they initiate breeding, so the link between the flight calls that differ between types and their breeding isolation by assortative flocking may be less direct for them if song types are shared between call types. Since breeding distributions of Evening Grosbeak call types are far more stable than for Red Crossbills, sympatric breeding may be easier to study and could provide different insights into the role of call types in breeding isolation as well as the possible dual role of flight calls in breeding and flocking behaviors in this system.
2 Methods
2023 Surveys: We conducted 1–5 surveys daily throughout Teton County, WY during the summer of 2023 from June 12 to July 27 and from August 18 to September 4 (n=198). We conducted surveys to maximize aural detections of nomadic finches (mainly surveying for crossbills) by surveying through the whole day and covering as many distinct survey locations as possible. Surveys were mostly done walking (n= 174) on trails or dirt roads with an average length of 2.87 km and average duration of 126 minutes, with some stationary counts (n=17) with average duration of 23 minutes, and a few slow driving counts with windows down (nearly all detections are initially aural) (n=7) with an average duration of 87 minutes and distance of 4.13 km. We made roughly equal numbers of visits to the major local forest types: Subalpine Forest with Subalpine Fir (Abies lasiocarpa (Hook.) Nutt.) and Engelmann Spruce (Picea engelmannii Engelm.), Douglas Fir (Pseudotsuga menziesii Mirb. Franco), Lodgepole Pine (Pinus contorta Douglas ex Loudon), Aspen (Populus trumuloides Michx.), Cottonwood (primarily Populus angustifolia E.James) Riparian, and Blue Spruce (Picea pungens Engelm.) Riparian (Table 1). Cover of different tree species was estimated visually as a percentage of total tree cover within a 300m radius of the route then binned into 4 even levels with zero percent as a fifth level. We identified Evening Grosbeak types by ear in the field and took recordings with a Marantz PMD661 Solid State Recorder and Sennheiser Short Shotgun Microphone for audio spectrographic analysis to confirm aural identifications (Figure 1). We referenced existing work describing the call types in this species (Sewall et al., 2004) along with additional unpublished recordings to make our identifications to type. We followed birds and recorded feeding and behavior of birds when possible. We used our eBird lists to record survey routes and counts and recorded occurrence and behavior notes in field notebooks.
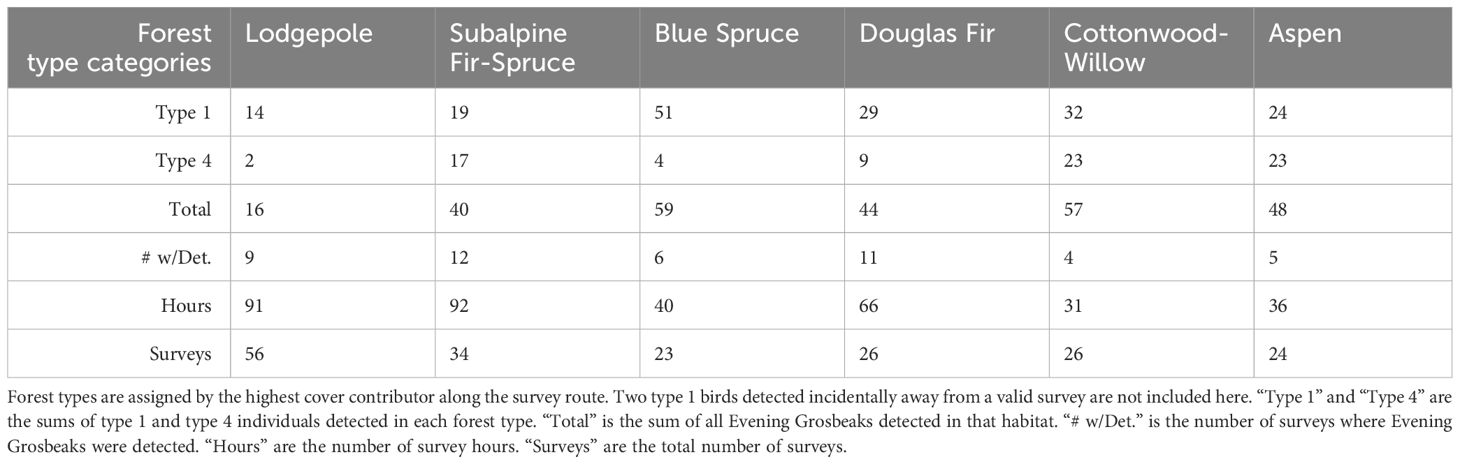
Table 1 Effort and total detected birds according to habitat category for the 2023 field season.
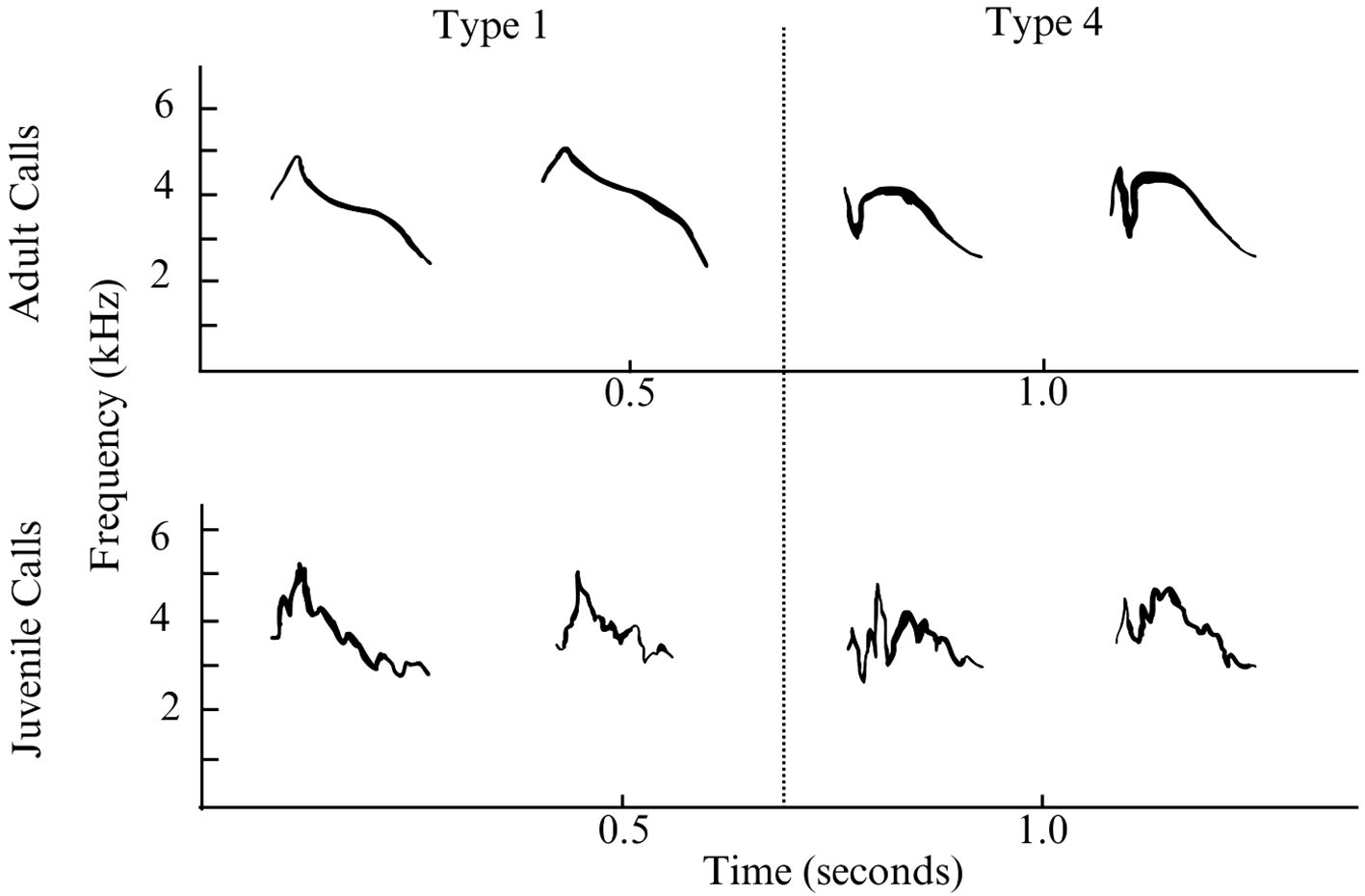
Figure 1 Spectrograms of adult and juvenile type 1 and type 4 Evening Grosbeaks taken during the 2023 field season in Teton County, Wyoming. Top, from left to right: Type 1, August 21, Jackson; Type 1, August 21, Jackson; Type 4, July 22, Cache Creek; Type 4, July 25, String Lake. Bottom, from left to right: Juvenile Evening Grosbeak, probably Type 1, August 19, Wilson; Juvenile Evening Grosbeak, probably Type 1, August 19, Wilson; Juvenile Evening Grosbeak, probably Type 4, August 21, Jackson; Juvenile Evening Grosbeak, probably Type 4, August 21, Jackson.
2000–2016 surveys: We recorded observations of Evening Grosbeak during field work predominantly on crossbills throughout the Jackson Hole region, including many of the same survey areas visited in 2023, from 2000–2016. This field work included numerous point counts as well as walking surveys along trails, and some opportunistic notations of Evening Grosbeaks encountered during other activities. Evening Grosbeaks were identified to call type by ear, but were often also recorded (e.g., point counts were recorded digitally). During over 900 hours of field time across these years we encountered Evening Grosbeaks infrequently, and many of these sporadic encounters were outside the June–July breeding season. For this paper, we separate detections during times of year comparable to field surveys in 2023 from those at other times of the year (Table 2).
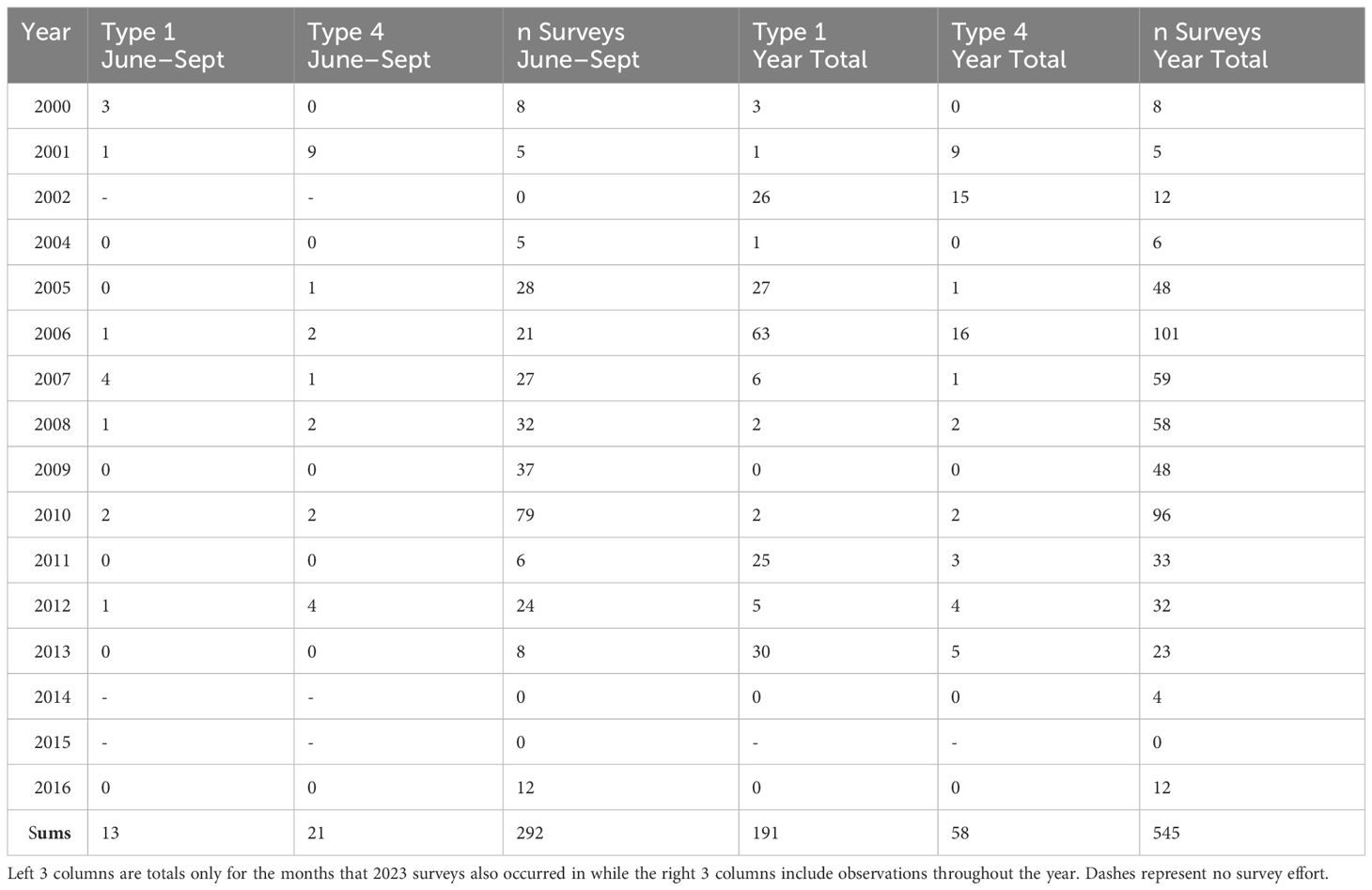
Table 2 2000–2016 Evening Grosbeak totals and number of surveys.
2023 field work was conducted by KD while 2000–2016 field work was conducted by TPH. All analyses were done in R Version 4.3.2 (R Core Team, 2023). We did mixed effects logistic regressions in package lme4 (Bates et al., 2015) with binned percent cover of tree species (5 levels) and time × (1 + distance) survey effort as a random effect with 5 binned levels. We present model outputs only for the two tree species with a significant fixed effect level (Table 3), and models incorporating multiple tree species produced no significant estimates due to loss of degrees of freedom and were not reported. We also included two-proportion z-tests to contrast proportions of the two types between 2023 and past surveys, and we used Fisher’s Exact Test to compare observed vs expected association between the two call types in 2023 and the past as well as to compare the observed vs expected associations with tree species presence. Of the tree species, only Douglas Fir yielded an observed proportion significantly greater than expected, so that is reported in results.
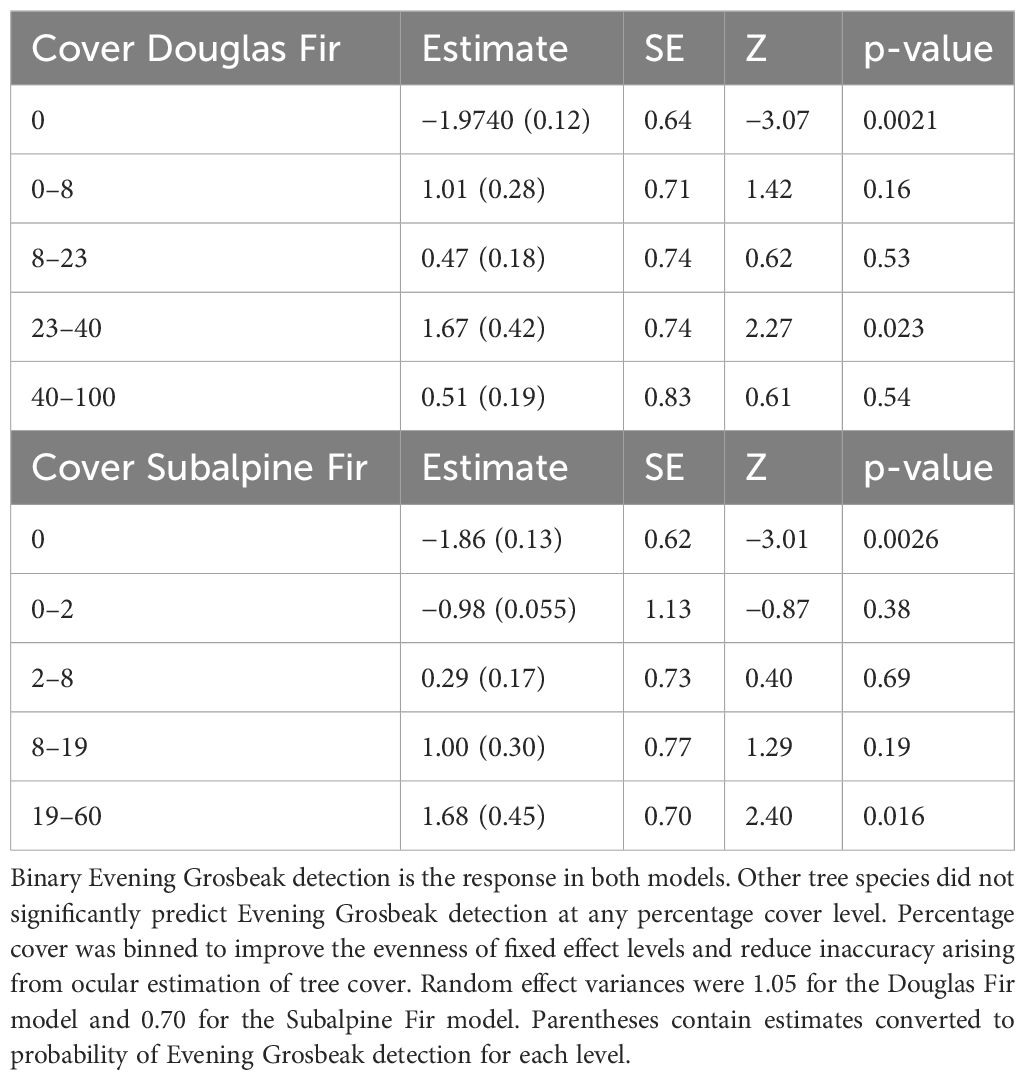
Table 3 Logistic regression estimates of the model with Douglas Fir cover categories as a predictor and effort as a random effect and of the model with Subalpine Fir cover categories as a predictor and effort as a random effect.
3 Results
3.1 Occurrence of the two types in 2023
We detected a total of 266 Evening Grosbeaks over the course of the summer of 2023 (double counting limited by visiting different sites on consecutive days). Of those, 78 were type 4, 171 were type 1, and 17 were not identified to call type. Evening Grosbeaks were somewhat common, and we detected them on 24% of surveys. The initial burst of detections was of exclusively type 1 birds from June 11 to 16 (Figure 2) followed by the first type 4 birds on June 25. Both types were thinly spread during the breeding season (late June–early July), with groups no larger than 3 detected between June 13 and July 22. We detected larger groups before and especially after this period, and mixed-type flocks of 10–28 with many juveniles present became common in late August and early September. These post-breeding flocks were all encountered in brushy aspen or cottonwood woodlands or suburban areas away from coniferous forest and near crops of Chokecherry and/or Serviceberry fruits, the seeds of which they feed on extensively during this time. These post breeding flocks all consisted of both call types and had slightly more type 4 birds than were encountered during the breeding season (Figure 2). During the breeding season, most feeding observations were on budworms (Choristoneura occidentalis Freeman, 1967 and/or Dioryctria reniculelloides Mutuura & Munroe, 1973 and/or D. pseudotsugella Munroe, 1959), which they captured on Subalpine Fir and Douglas Fir. There were no obvious differences in the forest composition (Table 1) or elevational association (type 1 breeding season average = 2076 m sd = 169 m, type 4 breeding season average = 2108 m sd = 233 m) between the two types. Both types associated with mixed conifer forests through the breeding season, especially those with Douglas Fir. Douglas Fir was the only tree species whose presence had a significant positive association with Evening Grosbeak detection during the breeding season (Fisher’s Exact Test p-value = 0.01 for both types and higher than expected proportion with detection and Douglas Fir present), and Subalpine Fir and Douglas Fir were the only tree species that significantly predicted higher Evening Grosbeak breeding season detection (Table 3). Detections of the two call types were significantly positively correlated with each other in the 2023 surveys (Fisher’s Exact Test p-value = 9.336e-08, odds ratio = 18.65 with 99% CI 3.88-139.53) and in the 2000–2016 surveys (Fisher’s Exact Test p-value = 6.236e-06), with the associations remaining the same when only breeding season surveys were included. We did not detect Evening Grosbeaks north of Moran, WY in the 39 surveys at that latitude or higher in the county in 2023 (Figure 3) or on the several surveys done there in years past. This pattern is also reflected in eBird submissions, with the percentage of complete and historical checklists recording Evening Grosbeak being an order of magnitude greater in Teton County south of Moran than the part of the County north of Moran (3.7% n=24,374 vs 0.38% n=15,943). This distribution pattern could be due to any of several factors relating to the shift in forest type towards vast and continuous stands of Rocky Mountain Lodgepole Pine forest (USGS, 2023), including lower budworm availability, more closed canopy, lower overstory diversity, or less Douglas Fir cover. Generally fewer Evening Grosbeaks were detected in Lodgepole forest across all surveys (Table 1). For example, no more than two Evening Grosbeaks were detected on any surveys where we estimated Lodgepole Pine percentage of tree cover to be over 60%.
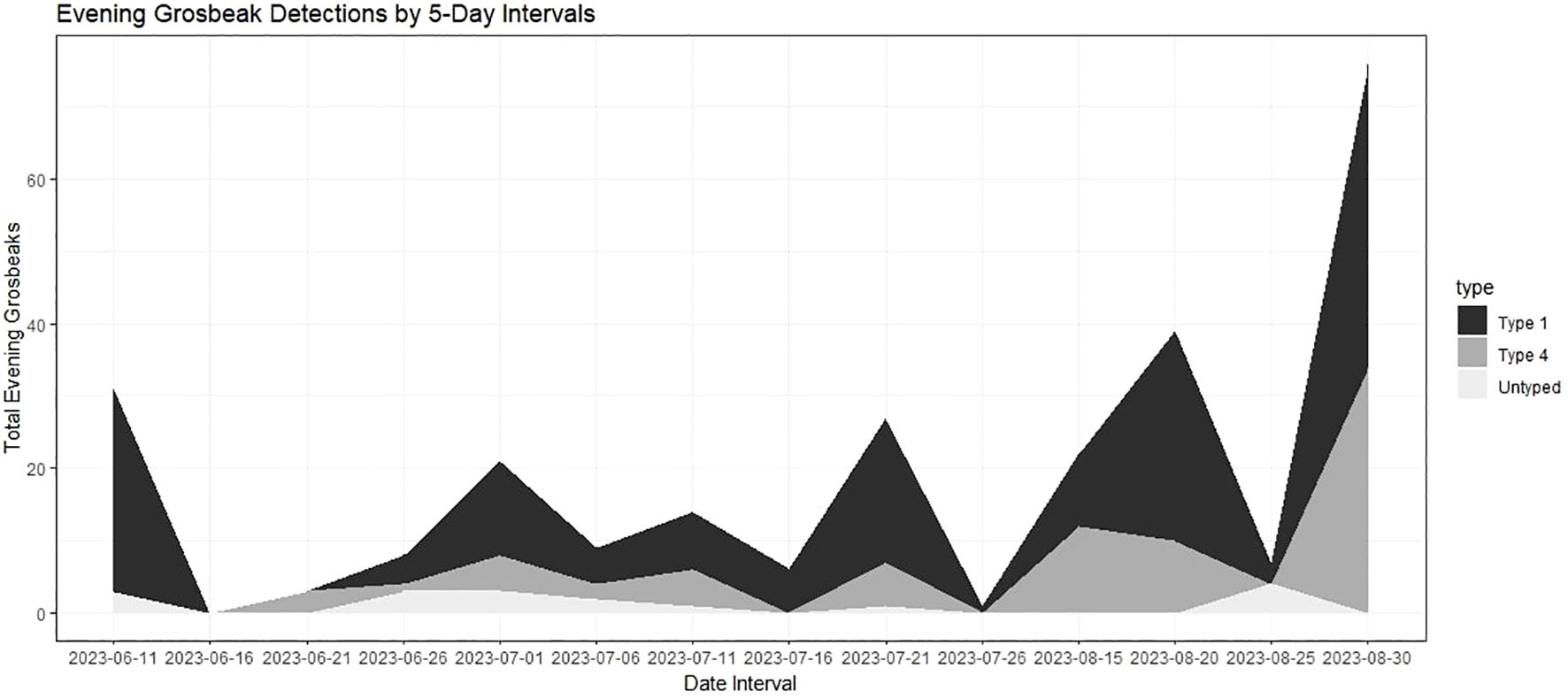
Figure 2 Total Evening Grosbeak detections per 5-day interval of surveys in 2023.
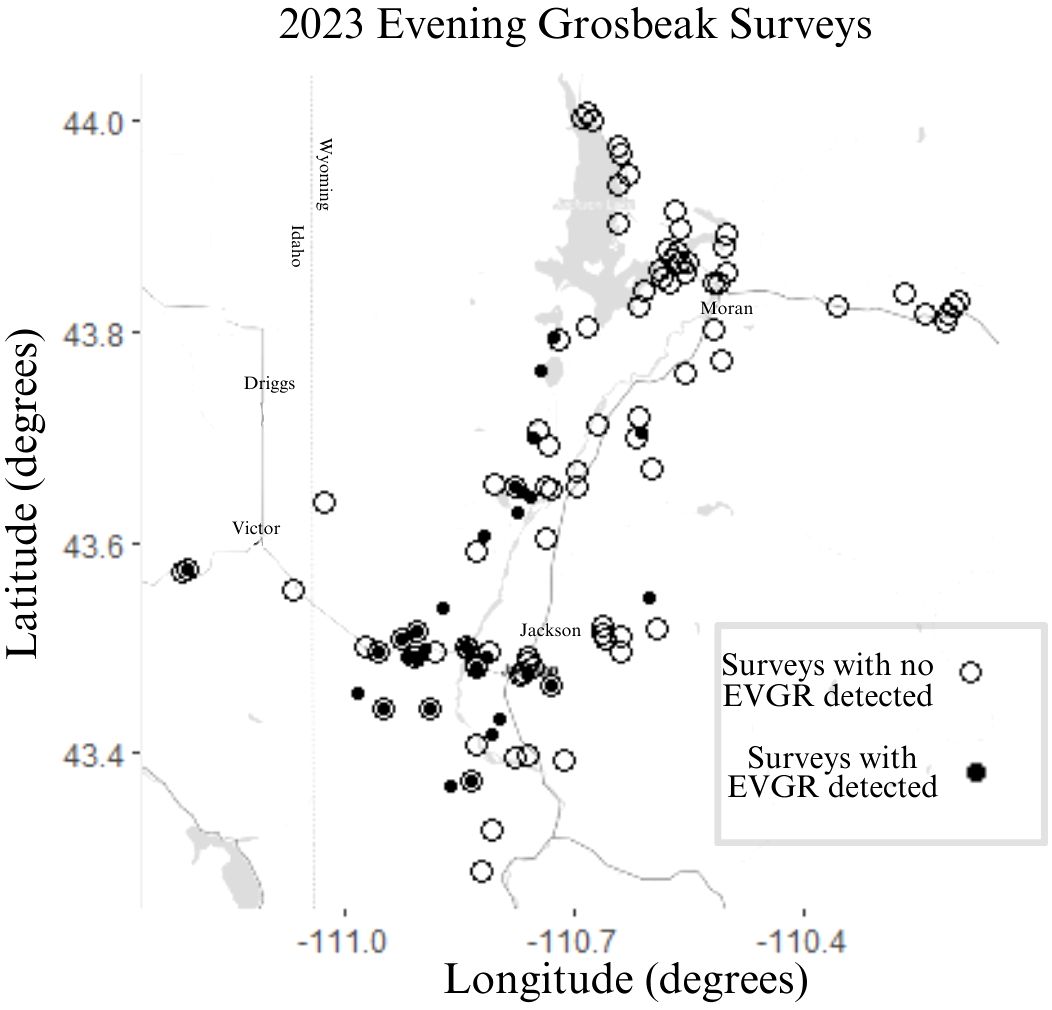
Figure 3 2023 survey locations. Open points indicate survey locations with no Evening Grosbeak (EVGR) detections while closed points indicate at least one Evening Grosbeak detected. Closed point inside an open point indicates that some surveys at that location had detections and others did not.
3.2 Past abundance and budworm fluctuation
On surveys conducted from 2000 to 2016 in the Jackson Hole area, Evening Grosbeaks were generally uncommon, but a few surveys in 2001, 2002, 2011, and 2013 had higher detection rates (Table 2). We detected a total of 191 type 1 birds, 58 type 4 birds, and 24 un-typed birds in 933 survey hours across 561 surveys (including non-targeted efforts where Evening Grosbeaks could have been detected) in multiple seasons during that 17-year period (Table 2). At no point during these or the 2023 surveys did we detect call types other than 1 and 4. On the 292 surveys (125 survey days) that overlapped with the 2023 survey period (June 10–September 4), we only detected 13 type 1 and 21 type 4 Evening Grosbeaks. We cannot determine if the proportion of type 1 to type 4 has changed between these periods (two-proportion z-test of “surveys with type 4” to “surveys with type 4 or type 1” compared in the June–September across 2000–2016 vs 2023 yielded p-value = 0.11 for proportions 0.69:0.41with two-sided Ha), but the proportions of the sums of the two types appear to have shifted towards more type 1 birds (two-proportion z-test of “sum of type 1” to “sum of both types” compared in June–September across 2000–2016 vs 2023 yielded p-value = .0011 for proportions 0.62:0.32). Extreme fluctuations in density at the local level have been documented before (Bekoff, pers. comm.), and some such population changes have been attributed to budworm outbreaks (Blais and Parks, 1964). In our case it seems likely that budworm availability had an effect because we observed qualitatively more budworm damage in 2022 and 2023 visits than on any of the 2000–2016 visits. We did not conduct routine budworm surveys until the 2023 season, but both authors noted the broad and severe scale of defoliation in late summer of 2022 (which recurred in 2023), suggesting that such defoliation would have been recorded in past field notes. Moreover, we recorded 4 isolated occurrences of insect defoliation in 2012 surveys suggesting that our past years’ notes would have reflected levels of defoliation as severe as those in 2023.
3.3 Local abundance increase in 2023
The increase in local abundance of Evening Grosbeaks in 2023 relative to decades past is conspicuous at a qualitative level, with far more Evening Grosbeaks of both call types (but especially type 1) detected per approximate survey effort (Tables 1, 2). It is further reflected by higher counts on surveys done in the same locations and along the same routes in 2023 as in multiple past years (Table 4). The increased counts seem to be mostly of type 1 birds though type 4 was also detected more broadly in 2023 than in past years, suggesting an increase for them as well. Though the differences in survey methods and variable effort prevents direct comparison of the counts or rates of detection, the expansion in the number of sites where Evening Grosbeaks were detected and the increase in counts at sites where survey effort was comparable (Table 4) supports our qualitative observation of much higher numbers in the region.
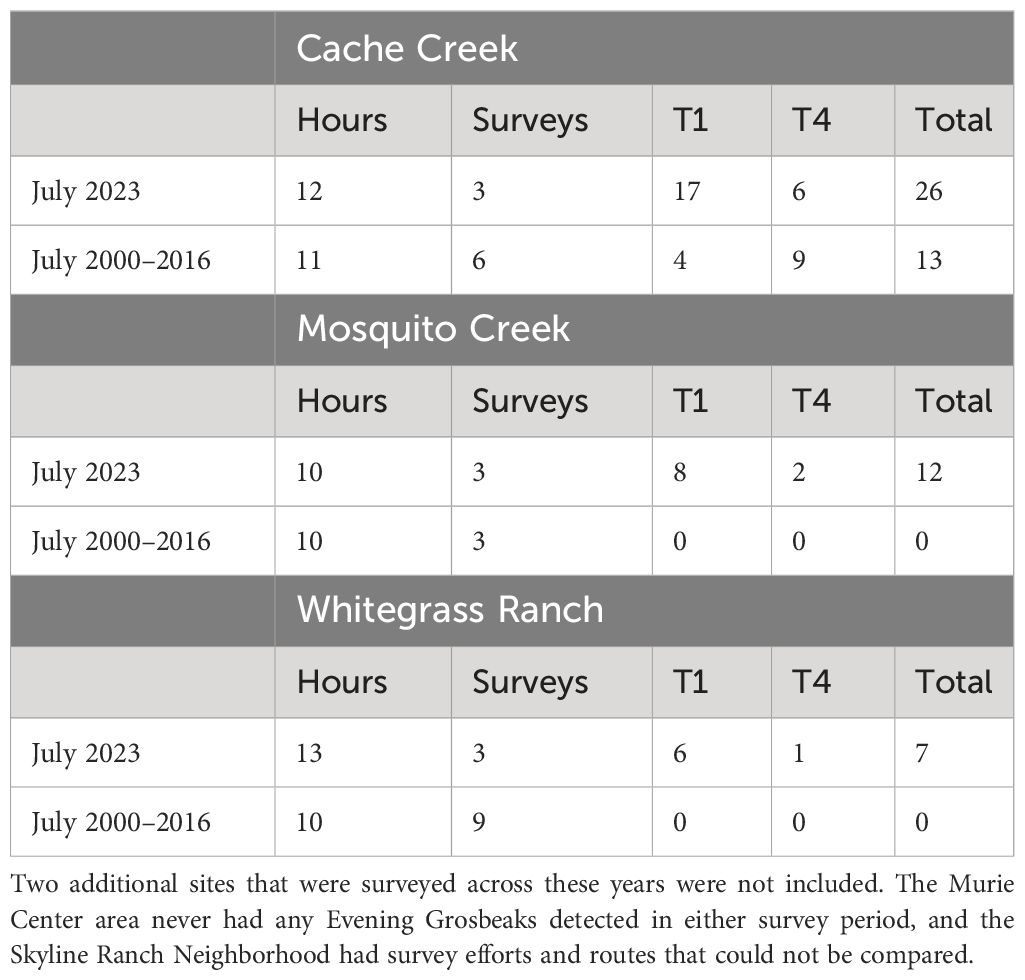
Table 4 July survey hours, number of July surveys, and Evening Grosbeak detections by type at three locations in the Jackson hole area that were repeatedly surveyed in July across 2000–2016 period and in 2023 following the same route over similar distances.
3.4 Breeding evidence for both types
Both call types 1 and 4 bred in the Jackson Hole area during the 2023 season, and we detected both types in the area through this entire period (Figure 2). We found two type 1 nests in different locations, one with both male and female feeding young and producing type 1 flight calls and another with only a female that sat on the nest and was heard calling. No type 4 nest was found, but a female type 4 was feeding a fledged juvenile on one survey and calling juveniles of type 4 (and type 1) were present in the post-breeding flocks. We also detected 2 potential type 4 pairs around Teton Pass by doubled calls (two flight call notes produced within 250 milliseconds of each other) on July 14. Type 1 pairs produced these doubled calls as well on 6 survey occasions, including one where we visually confirmed a male and a female.
4 Discussion
Our findings show that these two call types (and no others as far as we can tell) reside and breed in the same areas in the Jackson Hole area, and that they have both been in this area for at least 24 years. This consistency of the two types’ occurrences suggests a somewhat stable range boundary. This is notable because Evening Grosbeaks are well known for rapid range shifts and major population responses to budworm outbreaks in the eastern part of their range (Bolgiano, 2004), and this study reflects that such population fluctuations also occur in the west without any appreciable shift in range boundaries of call types at the scale we surveyed. Such a stable overlap zone with frequent mixed groups and no evidence of intermediate adult call notes suggests that discrete call types could be arising from early learning in their natal groups, without subsequent imitation or learning in flocks post-fledging. This would be more like the pattern in Red Crossbills (Sewall, 2009; Sewall, 2010) and less like that of several other Carduelines (Mundinger, 1979). Alternatively, if there is extensive flexibility in adult call learning or matching, vocal distinctness of the two call types could be maintained by not forming cross-type flock or pair associations despite apparent proximity. Such assortative flocking and pairing would be especially significant given the apparently limited ecological differences between these two call types, with no clear differences in habitat and frequent co-occurrence. If bill sizes of the types align with Grinnell’s subspecies, then type 4 could have a somewhat narrower bill which may provide some subtle degree of ecological distinctness.
Research on overlap zones between vocal groups is critical for establishing species limits and causes of ecological or behavioral differentiation (e.g. Toews and Irwin, 2008). Further work will be necessary to establish if type 1 and type 4 Evening Grosbeaks in this region are forming mixed-type pairs or not. Given the extent of overlap that we documented, with type 4 rarely occurring away from type 1, this area would be a strong candidate region for assessing levels of introgression. However, we did not detect any mixed-type pairs or groups until July 25, well after the main breeding period. This lack of mixed groups could also be due to the very small group sizes during the breeding season and the difficulty in detecting pairs. Though many juvenile calls had the overall pitch profile of type 1 or type 4, the more irregular and modulated quality (Figure 1) of juvenile calls prevented confident intermediate-type identifications of many juvenile birds.
The absence of both types in the part of the county north of Moran in 2023 is interesting given that it is forested and similar in elevation to other areas surveyed. Evening Grosbeaks have been shown to be less abundant in dense even-aged stands in Oregon (Hagar et al., 1996; Hayes et al., 2003), and these attributes are hallmarks of the Rocky Mountain Lodgepole Pine Forest system (NatureServe, 2023). Budworm availability could be another factor causing this difference since we counted far fewer budworms per bud tip north of Moran, even on the same tree species, and the soft-coned conifers with the most budworms on them are less dominant in that region. Within Teton County, WY, Moran appears to be the northern bound of type 4’s range. Further survey efforts in Eastern Idaho and Fremont and Park Counties of Wyoming will be necessary to establish if this latitude truly represents a northern bound to type 4’s breeding range. Other records farther north include a summer and a possible spring record in Gallatin County, Montana; a late summer record from Idaho County, Idaho, and an early July record from the Bighorn Mountains in Wyoming (eBird, 2023; Xeno Canto, 2023). Occasional occurrence of type 4 far north of the normal range limit would not be surprising; type 2 Evening Grosbeaks drop off rapidly in abundance north of Mount Shasta, CA, yet a few have been detected as far north as the southern Washington Cascades (pers. obs.).
Defining the ranges of these call types is an area of research that birders and other community scientists can contribute to greatly due to the minimal equipment required to sufficiently record flight calls and the vast geographic scope involved. Despite 5,279 recordings in the Macaulay Library and 269 in Xeno Canto (eBird, 2023; Xeno Canto, 2023), large areas of the Evening Grosbeak breeding range still lack any available recordings of their calls. This is especially true for some of the regions where two call types may overlap such as Alberta, Wyoming, and the Cascades of Oregon. For example, only 8 eBird checklists in the state of Wyoming had recordings of Evening Grosbeaks on them prior to our field 2023 season despite there being 2,688 checklists that reported Evening Grosbeak in the state.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because solely observational work with no audio playback, physical contact, or disturbance was used. This does not necessitate IACUC approval.
Author contributions
KD: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. TH: Data curation, Investigation, Methodology, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding for this field work was provided by the University of Wyoming- National Parks Service Research Grant (PTE Federal Award Number: P22AC00531, Subaward#: 1005851B-UCD_Duman) and the Western Field Ornithologists Student Research Grant.
Acknowledgments
We would like to thank Mary Lohuis of Jackson, WY for providing housing and good company during the 2023 field season and Matthew Young at the Finch Research Network for discussing these ideas with us.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Baird S. F., Brewer T. M., Ridgway R. (1874). A history of North American birds: land birds Vol. 1 (Boston, MA: Little Brown). doi: 10.5962/bhl.title.7493
Bates D., Mächler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Software 67, 1–48. doi: 10.18637/jss.v067.i01
Bekoff M., Scott A. C., Conner D. A. (1989). Ecological analyses of nesting success in evening grosbeaks. Oecologia 81, 67–74. doi: 10.1007/BF00377012
Benkman C. W., Smith J. W., Keenan P. C., Parchman T. L., Santisteban L. (2009). A new species of the Red Crossbill (Fringillidae: Loxia) from Idaho. Condor 111, 169–176. doi: 10.1525/cond.2009.080042
Blais J. R., Parks G. H. (1964). Interaction of evening grosbeak (Hesperiphona vespertina) and spruce budworm (Choristoneura fumiferana (Clem.)) in a localized budworm outbreak treated with DDT in Quebec. Can. J. Zool. 42, 1017–1024. doi: 10.1139/z64-099
Bolgiano N. C. (2004). Changes in boreal bird irruptions in eastern North America relative to the 1970s spruce budworm infestation. Am. Birds 54, 26–33.
Chesser R. T., Burns K. J., Cicero C., Dunn J. L., Kratter A. W., Lovette I. J., et al (2017). Fifty-eighth supplement to the American Ornithological Society's check-list of North American birds. The Auk: Ornithological Advances. 134 (3), 751–773.
Cicero C., Mason N. A., Oong Z., Title P. O., Morales M. E., Feldheim K. A., et al. (2022). Deep ecomorphological and genetic divergence in Steller’s Jays (Cyanocitta stelleri, Aves: Corvidae). Ecol. Evol. 12, e9517. doi: 10.1002/ece3.9517
eBird (2023) eBird: An online database of bird distribution and abundance. eBird, Cornell Lab of Ornithology, Ithaca, New York. Available online at: http://www.ebird.org (Accessed November 11, 2023).
Gillihan S. W., Byers B. (2001)Evening Grosbeak: Coccothraustes vespertinus. In: Birds of The World. Available online at: https://birdsoftheworld.org/bow/species/evegro/cur/introduction (Accessed October 25, 2023).
Grant B. R., Grant P. R. (1996). Cultural inheritance of song and its role in the evolution of Darwin's finches. Evolution. 50 (6), 2471–2487.
Grinnell J. (1917). The subspecies of Hesperiphona vesperitina. Condor 19, 17–22. doi: 10.2307/1362452
Groth J. G. (1993). Evolutionary differentiation in morphology, vocalizations, and allozymes among nomadic sibling species in the North American Red Crossbill (Loxia curvirostra) complex (Berkeley, CA, USA: University of California Publication in Zoology).
Hagar J. C., McComb W. C., Emmingham W. H. (1996). Bird communities in commercially thinned and unthinned Douglas-fir stands of western Oregon (Wildlife Society Bulletin), 353–366.
Hayes J. P., Weikel J. M., Huso M. M. (2003). Response of birds to thinning young Douglas-fir forests. Ecol. Appl. 13, 1222–1232. doi: 10.1890/02-5068
James F. C. (2010). Avian subspecies: introduction. Ornithol. Monogr. 67, 1–5. doi: 10.1525/om.2010.67.1.1
Lipshutz S. E., Overcast I. A., Hickerson M. J., Brumfield R. T., Derryberry E. P. (2017). Behavioural response to song and genetic divergence in two subspecies of white-crowned sparrows (Zonotrichia leucophrys). Mol. Ecol. 26, 3011–3027. doi: 10.1111/mec.14002
Mason N. A., Burns K. J., Tobias J. A., Claramunt S., Seddon N., Derryberry E. P. (2017). Song evolution, speciation, and vocal learning in passerine birds. Evolution 71, 786–796. doi: 10.1111/evo.13159
Mearns E. A. (1890). Descriptions of a new species and three new subspecies of birds from Arizona. Auk 7, 243–251. doi: 10.2307/4067983
Mundinger P. C. (1970). Vocal imitation and individual recognition of finch calls. Science 168, 480–482. doi: 10.1126/science.168.3930.480
Mundinger P. C. (1979). Call learning in the Carduelinae: ethological and systematic considerations. System. Biol. 28, 270–283. doi: 10.1093/sysbio/28.3.270
NatureServe. (2023). NatureServe Network Biodiversity Location Data accessed through NatureServe Explorer [web application]. NatureServe, Arlington, Virginia. Available https://explorer.natureserve.org/. (Accessed November 11, 2023).
R Core Team (2023). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Robinson W. D., Greer J., Masseloux J., Hallman T. A., Curtis J. R. (2022). Dramatic declines of evening grosbeak numbers at a spring migration stop-over site. Diversity 14, 496. doi: 10.3390/d14060496
Robinson W. D., Nanau M., Kirsch W., Centanni C. T., Clements N. M. (2024). Flight calls and trills of Evening Grosbeaks can be used to map movements and ranges of call types 1 and 2. Frontiers in Bird Science. 3, 1340750.
Sewall K. B. (2009). Limited adult vocal learning maintains call dialects but permits pair-distinctive calls in red crossbills. Anim. Behav. 77, 1303–1311. doi: 10.1016/j.anbehav.2009.01.033
Sewall K. B. (2010). Early learning of discrete call variants in red crossbills: implications for reliable signaling. Behav. Ecol. Sociobiol. 65, 157–166. doi: 10.1007/s00265-010-1022-0
Sewall K., Kelsey R., Hahn T. P. (2004). Discrete variants of Evening Grosbeak flight calls. Condor 106, 161–165. doi: 10.1093/condor/106.1.161
Toews D. P., Irwin D. E. (2008). Cryptic speciation in a Holarctic passerine revealed by genetic and bioacoustic analyses. Mol. Ecol. 17, 2691–2705. doi: 10.1111/j.1365-294X.2008.03769.x
U.S. Geological Survey GAP Program (2023) National Land Cover Data – Wyoming. Available online at: http://gap.uidaho.edu/index.php/gap-home/Northwest-GAP (Accessed October 25, 2023).
Wetmore A., Friedmann H., Amadon D., Lincoln F. C., Lowery G. H. Jr., Miller A. H., et al. (1957). Check-List of North American Birds. 5th Edition (Lord Baltimore Press, Inc), 557.
Xeno Canto (2023) Xeno-canto: Sharing wildlife sounds from around the world. Available online at: https://xeno-canto.org (Accessed November 11, 2023).
Keywords: call type, range overlap, Evening Grosbeak, sympatric breeding, vocal group
Citation: Duman KW and Hahn TP (2024) Sympatric breeding occurrence of two call types of Evening Grosbeak in Western Wyoming. Front. Bird Sci. 3:1371155. doi: 10.3389/fbirs.2024.1371155
Received: 15 January 2024; Accepted: 21 March 2024;
Published: 19 April 2024.
Edited by:
Scott Rush, Mississippi State University, United StatesReviewed by:
Herbert Hoi, University of Veterinary Medicine Vienna, AustriaJesús Domínguez, University of Santiago de Compostela, Spain
Copyright © 2024 Duman and Hahn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Konshau W. Duman, a2R1bWFuQHVjZGF2aXMuZWR1