
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Bioeng. Biotechnol., 06 March 2023
Sec. Bioprocess Engineering
Volume 11 - 2023 | https://doi.org/10.3389/fbioe.2023.1143255
 Marco Silvano1,2
Marco Silvano1,2 Nikolaus Virgolini1,2
Nikolaus Virgolini1,2 Ricardo Correia1,2
Ricardo Correia1,2 Colin Clarke3,4
Colin Clarke3,4 Inês A. Isidro1,2
Inês A. Isidro1,2 Paula M. Alves1,2
Paula M. Alves1,2 António Roldão1,2*
António Roldão1,2*The insect cell-baculovirus expression vector system (IC-BEVS) has been widely used to produce recombinant protein at high titers, including complex virus-like particles (VPLs). However, cell-to-cell variability upon infection is yet one of the least understood phenomena in virology, and little is known about its impact on production of therapeutic proteins. This study aimed at dissecting insect cell population heterogeneity during production of influenza VLPs in IC-BEVS using single-cell RNA-seq (scRNA-seq). High Five cell population was shown to be heterogeneous even before infection, with cell cycle being one of the factors contributing for this variation. In addition, infected insect cells were clustered according to the timing and level of baculovirus genes expression, with each cluster reporting similar influenza VLPs transgenes (i.e., hemagglutinin and M1) transcript counts. Trajectory analysis enabled to track infection progression throughout pseudotime. Specific pathways such as translation machinery, protein folding, sorting and degradation, endocytosis and energy metabolism were identified as being those which vary the most during insect cell infection and production of Influenza VLPs. Overall, this study lays the ground for the application of scRNA-seq in IC-BEVS processes to isolate relevant biological mechanisms during recombinant protein expression towards its further optimization.
The insect cells (IC) and baculovirus expression vector system (BEVS) constitute an attractive alternative to mammalian cells for manufacturing of heterologous gene products, including recombinant proteins as vaccine candidates and viral vectors for gene therapy (Drugmand et al., 2012). Recent advances in next-generation sequencing technologies have enabled a considerable improvement of our understanding of the IC-BEVS. For example, RNA-seq has been used to assess the transcriptional changes of alphanodavirus-free High Five cells upon infection by Autographa californica multiple nucleopolyhedrovirus (AcMNPV) (Chen et al., 2013; Chen et al., 2014), providing a global picture of the AcMNPV transcription regulation throughout the infection cycle. As the knowledge of the IC-BEVS grows, potential engineering targets to increase recombinant protein production are being identified (Silvano et al., 2022).
In recent years, technological advances in areas such as cell isolation methods using microfluidics or microwell devices, preparation of next-generation sequencing libraries from ultra-low quantities of nucleic acids, and innovative labelling strategies for MS-based proteomics have enabled the characterization of DNA, RNA and proteins at single-cell resolution (Lee et al., 2020). Using different single-cell omics profiling strategies as building blocks, we can now build a multi-omics profile of the same cell. These multi-omics methods will play an important role in many diverse fields, and their applications are rapidly expanding, including delineating cellular diversity (Gulati et al., 2020), lineage tracing (Wagner and Klein, 2020), identifying new cell types (Ianevski et al., 2022), and deciphering the regulatory mechanisms between omics (Cao and Gao, 2022). Single-cell analysis has unambiguously demonstrated that cell populations are often heterogeneous (Goldman et al., 2019). This heterogeneity not only applies to different cell types in a tissue (Karlsson et al., 2021) but also to clonal cell population (Stockholm et al., 2007).
Single-cell RNA sequencing (scRNA-seq) has just recently be applied to virus-based processes. The power of scRNA-seq lies in the simultaneous delivery of snapshots of virus and host transcriptomes, and allows to compare host transcriptome between cells with low and high viral loads (Suomalainen and Greber, 2021). The high-resolution dissection of viral and host cell gene expression patterns reveals that the transcriptional responses of individual infected cells can be divergent, as the interplay between underlying cellular heterogeneity and viral population diversity influences the fate of infection (Sun et al., 2020). For example, cell-to-cell variation in viral transcription has been observed during influenza virus infection in mammalian (A549, MDCK and HEK293) cells (Russell et al., 2018).
To date, the understanding of the IC-BEVS transcriptome has been mostly relying on bulk RNA-seq analysis (Nguyen et al., 2013; Chen et al., 2014; Silvano et al., 2022; Virgolini et al., 2022). For instance, we have previously assessed whole transcriptome changes in High Five insect cells during expression of influenza HA-displaying virus-like particles (HA-VLPs) using IC-BEVS, which enabled to identify key biological processes impacted by virus infection (Silvano et al., 2022). Although these studies uncover transcriptional changes in insect cell response to baculovirus infection, they only provide rough models of the host cell response. Understanding IC-BEVS at the single-cell level could elucidate better the mechanisms of viral infection and potentially enable to identify, within a potentially heterogenous cell population and infection process, the characteristics of cells associated with a more efficient progression of infection and production of heterologous proteins.
In this study, we used scRNA-seq for the characterization of the High Five insect cell line during production of influenza HA-VLPs using IC-BEVS. The transcriptomics pipeline here described allowed to study, at a single-cell level, High Five cell population heterogeneity (prior and during infection), host cell response to virus infection, and progression of infection (expression of virus genes and transgenes encoding HA-VLPs).
High Five insect cells (Invitrogen) were routinely sub-cultured to .3–.5×106 cell mL−1 every 2–3 days when cell concentration reached 2–3×106 cell mL−1 in serum-free Insect-XPRESS™ medium (Sartorius) using 125–500 mL shake flasks with a 10% working volume, and maintained at 27°C in a Inova 44R shaking incubator (orbital motion diameter of 2.54 cm—Eppendorf) operating at 100 RPM.
Recombinant baculoviruses carrying influenza capsid M1 from A/California/06/2009 H1N1 strain and hemagglutinin (HA) from A/Brisbane/59/2007 strain genes were kindly provided by Redbiotec AG (Schlieren, Switzerland). Baculovirus amplification was performed as described elsewhere (Vieira et al., 2005).
HA-VLPs production was carried out in a 0.5 L stirred tank bioreactor (BIOSTAT Qplus–Sartorius) as specified elsewhere (Correia et al., 2020). Cells were expanded in 500–2000 mL shake flasks with a 10% working volume as described above. Infection experiments were performed in bioreactor at cell concentration at the time of infection (CCI) of 2×106 cell mL−1 and multiplicity of infection (MOI) of 1 pfu.cell−1. Medium exchange was performed at the time of infection by centrifugation at 200 g at room temperature for 10 min. Samples were taken every 24 h for the assessment of cell concentration and viability, and detection of M1 and HA proteins; for scRNA-seq, samples were taken before infection, and at 8 and 22 h post-infection (hpi).
Culture bulk from bioreactor run was harvested 3 days post-infection and centrifuged at (first) 4°C, 200 g, for 10 min and (second) four°C, 2000 g, for 20 min. The supernatant was filtered using a .22 µm Stericup (Millipore), and the HA-VLPs were purified using a SartoBind Q capsule (Sartorius Stedim Biotech) according to manufacturer’s instructions. Purified material was formulated in 50 mM HEPES, 300 mM NaCl, 15% (w/v) trehalose, pH 7.4, and stored at −80 or four°C.
Cell concentration was determined in a Fuchs-Rosenthal hemocytometer chamber (Brand) and cell viability assessed by trypan blue exclusion method (J R Tennant, 1964).
Baculovirus titers were determined using the MTT assay as described elsewhere (Mena et al., 2003; Roldão et al., 2009).
Identification and relative quantification of M1 and HA in culture supernatant were performed as reported elsewhere (Correia et al., 2020).
Negative staining transmission electron microscopy was used to assess the conformation and size of HA-VLPs. Briefly, 5 μl of purified VLP sample was fixed for 2 min in a copper grid coated with Formvar-carbon (Electron Microscopy Sciences, Hatfield). Grids were washed with H2O and then stained with 2% (v/v) uranyl acetate for 5 min and left to air dry. Samples were then observed in a Hitachi H-7650 Transmission Electron Microscope (JEOL, United States).
For single-cell gene expression profiling, ≈6000 cells (at 0 hpi and 8 hpi) or ≈8000 cells (at 22 hpi) were loaded into a BD Rhapsody cartridge (BD Biosciences) and libraries were generated according to BD Rhapsody™ System mRNA Whole Transcriptome Analysis (WTA). Upon confirming the quality of the resulting libraries using a Bioanalyser, the quantity of each library was determined using Qubit. ScRNA-seq libraries were sequenced using an Illumina NovaSeq (Illumina) configured to yield 150 bp paired end reads.
The cellular barcodes were pre-processed and demultiplexed by the BD Rhapsody WTA bioinformatic workflow (BD Biosciences) on the Seven Bridges Genomics (SBG) cloud platform using default parameters, as reported elsewhere (Tzani et al., 2021). STAR indexes were generated from the Trichoplusia ni (Tnl) reference genome (RefSeq assembly accession: GCF_003590095.1) and from AcMNPV (RefSeq assembly accession: GCF_000838,485.1, ViralProj14023) (Dobin et al., 2013). Specifically, a hybrid reference genome was used for RNA-seq read mapping using transgenes (M1 and HA) sequences and mtDNA sequence of the Tnl reference genome (GenBank accession No. MK714850.1).
The cell/gene matrices output from the SBG pipeline were imported into the R-4.2.1 Statistical Software Environment and merged to form a single matrix for further analysis. The proportion of unique molecular indexes (UMIs) originating from mtDNA was also determined for each cellular barcode, and cells with >5% mitochondrial UMIs counts were considered of low-quality and thus removed from further analysis.
Seurat v4 was used to apply a graph-based clustering approach (Hao et al., 2021). These methods embed cells in a graph structure with edges drawn between cells with similar feature expression patterns, and then attempt to partition this graph into highly interconnected “communities” (Xu and Su, 2015). To evaluate cell heterogeneity, data sets (0, 8, and 22 hpi) were merged prior to global scaling normalization method. Normalized and merged samples were scaled and variations caused by different total UMIs per cell were regressed out. The most variable features were considered for principal component analysis, and 20 principal components were used to perform cluster analysis. The Uniform Manifold Approximation and Projection (UMAP) technique was used to run non-linear dimensional reduction and to visualize and explore the datasets (McInnes et al., 2018). Monocle three was run to conduct trajectory analysis (Trapnell et al., 2014) and its function graph_test was used to identify genes that change as function of pseudotime. Genes with an average expression change of ≥ .5 and p-value <.05 were considered significant.
Cell cycle scoring function in Seurat v4 was used to determine the likelihood of cells being in either S or G2/M phase, based on reference genes known to play a role in distinct phases of the cell cycle. To conduct this procedure, we mapped the mouse gene list to the Tnl genome to carry out the classification and draw a list of Tnl cell-cycle genes. The resulting scores for S, G2/M and G1 phases were used to regress out the effect of cell cycle in downstream analysis.
For gene annotation, the amino acid sequence of protein-coding genes was used as a query. Blastp search was applied in the NCBI nr protein database using Blast2GO OmicsBox (Götz et al., 2008). No taxonomy filter was applied, and the E-value cutoff was set to 1 × 10−3. The over-representation of pathways within gene lists found to have a statistically significant association with pseudotime were identified using the Fisher’s exact test. Pathway terms with a False Discovery Rate of <.05 were considered significant (Benjaminit and Hochberg, 1995).
High Five cells were infected at CCI of 2×106 cell mL−1 with a MOI of 1 pfu.cell−1, and infection kinetics were assessed throughout (Figure 1A). M1 and HA proteins were identified by Western blot (Figure 1B), and particles resembling influenza HA-VLPs, both in size and morphology, detected by TEM (Figure 1C). These results are in line with those previously reported (Silvano et al., 2022) and demonstrate that HA-VLPs were successfully produced.
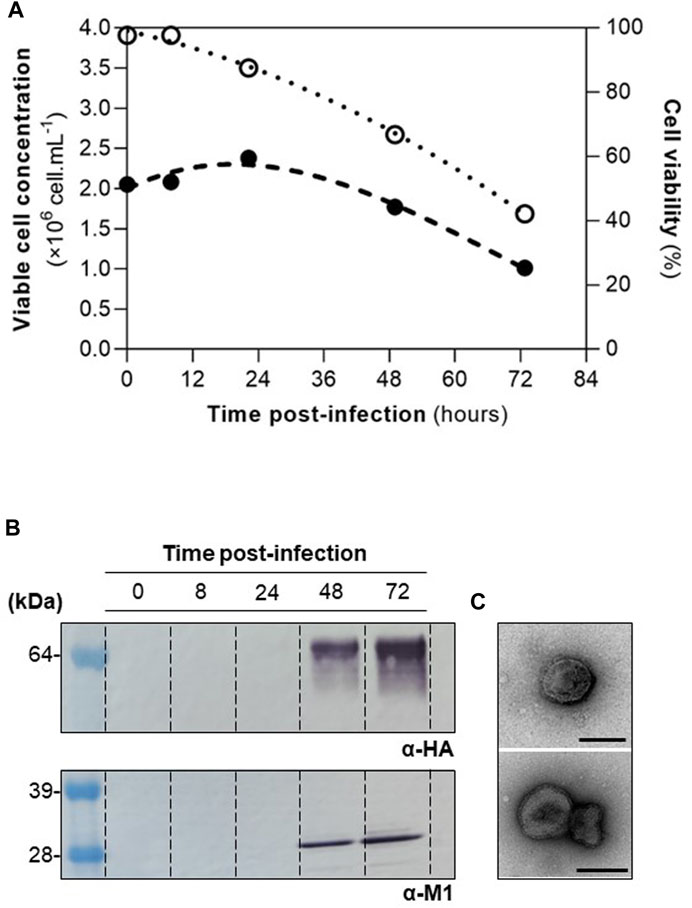
FIGURE 1. Production of influenza HA-VLPs. (A) Cell growth kinetics upon infection. Viable cell concentration and cell viability in full and empty circles, respectively. (B) Identification of HA and M1 proteins by Western blot. (C) TEM of purified HA-VLPs; scale bar represents 100 nm.
For scRNA-seq, ∼6,000–8,000 cells were collected at each time point (>90% cell viability). After sequencing, an average of 391 million 150 bp-reads were acquired for each sample. Following the completion of this initial pre-processing stage, approx. 4% of the sequenced RNA-seq reads were removed from further analysis due to insufficient read length, low base quality and/or high single nucleotide frequency; an average of ≈377 million reads per time point remained valid for further analysis (Supplementary Table S1). From the reads that passed quality control, 87% of the reads were successfully assigned to cell barcodes following demultiplexing. Mapping to the reference genome resulted in a unique alignment rate of ∼71%. Upon collapsing to UMIs and application of the RSEC algorithm, between 4,496 and 5,408 unique cell barcodes were identified for the three samples taken throughout the process. The mean number of reads and mRNA molecules detected per cell in this experiment were 41,166 and 28,787, respectively, with an average of ∼3,372 genes detected in each cell (Supplementary Table S1).
To ensure that only high-quality genes were retained for further analysis, the UMI count matrix was filtered to remove data that might have originated from non-viable cells. An average of 8% of cells contained >5% of detected UMIs originating from mitochondrial genes and thus were eliminated from further analysis (Figure 2A). The number of genes per cell and baculovirus UMIs (Figure 2B) as well as the amount of total UMIs per cell (Supplementary Figure S1) were assessed; while at 0 and 8 hpi the average number of genes identified per cell is 4000–5000, this number decreases significantly at later stages of infection (i.e., 22 hpi) concomitantly with an increase in the percentage of baculovirus UMIs.
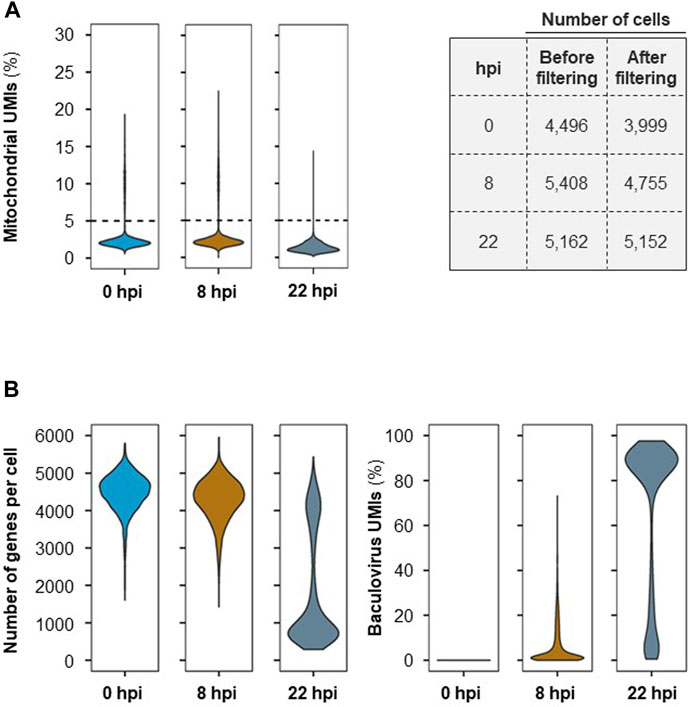
FIGURE 2. Single-cell RNA-seq quality control. (A) Mitochondrial UMIs per cell (on the left) and number of cells before and after filtering per UMIs originating from mitochondrial genes (>5%) (on the right). (B) Number of genes identified per cell and percentage (%) of cells containing UMIs originating from baculovirus genes at each time point after cell filtering.
To identify cell populations existing across the three single-cell datasets (0, 8, and 22 hpi), merged analysis was performed using Seurat v4. A total of 10 clusters were drawn from this analysis (Figure 3A), with a noticeable cluster re-organization being observed throughout infection progression. Importantly, five of those clusters were already present before infection (0 hpi) suggesting that High Five cells population was heterogeneous. Cell cycle is known to contribute to heterogeneity in scRNA-seq datasets (McDavid et al., 2016); to ascertain this in our study, cell cycle covariate was estimated using the Cell Cycle Scoring method in the Seurat package (Tirosh et al., 2016). While heterogeneity is clear at 0 hpi, with cells being associated to different cell cycle stages in a proportion of ≈1:7:5 (S:G2M:G1), at 22 hpi most cells have been identified as being in G1 phase (74%) (Supplementary Figure S2A,B). However, at this later timepoint, cell cycle association seems to be misassigned as exemplified by the overall low expression of the G1 cell cycle-related genes ccnd3, ccne1, and cdk6 in Supplementary Figure S2C. This could be a consequence of overexpression of baculovirus UMIs (hence lower expression of host cell genes), impairing correct cell cycle identification. Thus, the cell cycle regression was not further applied for the merged dataset.
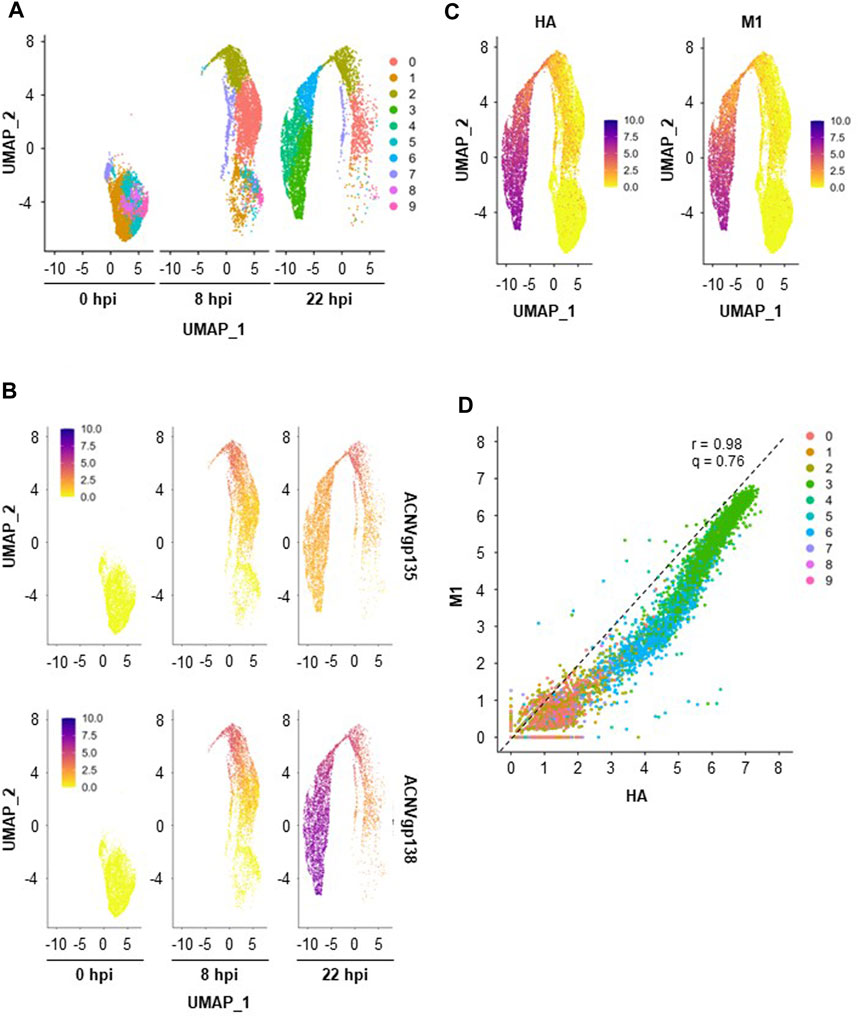
FIGURE 3. Insect cells clustering and transgenes expression. (A) Merged scRNA-seq dataset obtained with UMAP. (B) Relative expression of early (ACNVgp135) and late (ACNVgp138) baculovirus genes. (C) Relative expression of transgenes HA and M1. A color gradient scale was used to show the relative gene expression per cell. (D) Relative HA and M expression per cell, with dashed line representing the best linear fit to the data (r—Pearson’s correlation coefficient, q–angular coefficient). A color code was used to identify each cell cluster.
Cluster re-organization throughout infection seems to be correlated with the expression of baculovirus genes as infection progresses (Figure 3B); for example, expression of an early baculovirus gene (ACNVgp135) was higher in clusters denoting a transitory stage (i.e., cluster #2) whereas a late baculovirus gene (ACNVgp138) was more expressed in clusters (e.g., clusters #3, #4, and #6) furthest from those identified at 0 hpi. As seen for ACNVgp138, expression of transgenes M1 and HA, both under the regulation of the late expression promoter polyhedrin, was mainly identified in clusters #3, #4, and #6 (Figure 3C), with similar expression levels of HA and M1 genes being observed regardless of the cell cluster (r = .98, q = .76, Figure 3D).
To assess cell population evolution during infection, pseudo-temporal ordering (i.e., trajectory analysis) was applied. To perform this analysis, 13,906 cells from the merged Seurat analysis were used as input of Monocle 3. The cell cluster with the lowest percentage of HA (cluster #5, see Figure 3A) was selected as root state of the trajectory since it was the one most closely resembling the non-infected cell population; the pseudotime variable was then ordered accordingly (Figure 4A).
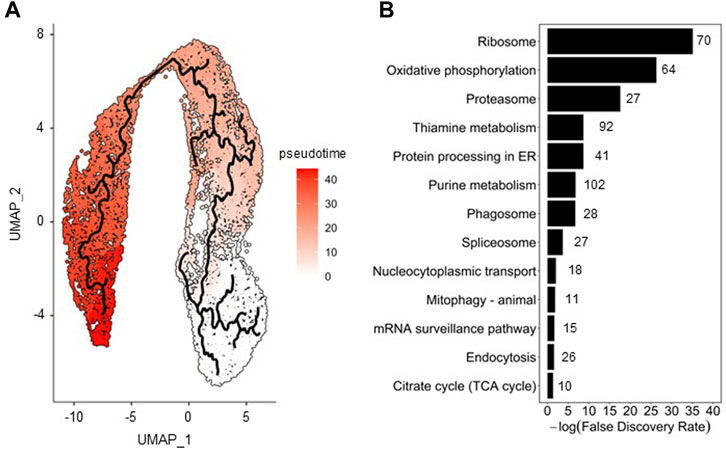
FIGURE 4. Pseudotime analysis of insect cells along infection. (A) Trajectory analysis on merged cell dataset (0, 8, and 22 hpi); a color gradient scale was used to visualize the pseudotime. (B) Pathway enrichment analysis performed with genes identified as the most changing in function of pseudotime using the Fisher’s exact test; the number of genes involved and found in the enrichment analysis are shown next to the bars.
Transcriptomic changes characterizing the progression of cells along HA production were assessed through identification of genes correlated with pseudotime. Overall, 921 host cell genes were found to be significantly (q value <.05) associated with pseudotime. Pathway enrichment analysis allowed to identify biological processes varying most along infection, of which those associated to translation machinery, energy metabolism, protein folding and endocytosis (Figure 4B) are some examples, in good agreement to what we have previously found in bulk RNA sequencing analysis (Silvano et al., 2022; Virgolini et al., 2022). The relative expression of a selected number of genes involved in these pathways is presented in Supplementary Figure S3, illustrating the significant transcriptional changes in cells upon infection.
In this work, we used scRNA-seq sequencing to analyze, at the single-cell level, alterations in the transcriptome of a High Five insect cell line infected with baculovirus during the production of influenza HA-VLPs.
Stable and high foreign gene expression levels are important criteria during the development of producer cell lines for pharmaceutical applications (Tripathi and Shrivastava, 2019), and understanding the mechanisms behind gene expression variation of genetically identical cells is the first step of this process. Cell population heterogeneity can influence parameters such as cell growth rate, genetic stability and productivity (Schmitz et al., 2019), and thus tracking it is key to avoid process failure and guarantee reproducibility (Olsson et al., 2022). Most studies on this topic focus on mammalian cells (Lewis et al., 2016; Samoudi et al., 2021), with none to date exploring insect cells. In our study, we could observe heterogeneity in High Five cell population before infection (as demonstrated by the identification of five cell clusters), largely resulting from the phase of the cell cycle that the cells are in, which was further amplified upon infection (6 more cell clusters were identified) as consequence of viral DNA replication and gene expression (Du and Thiem, 1997) and cell cycle arrest (Braunagel et al., 1998).
Baculovirus genes are known to be transcribed temporally, a process highly regulated by infection-derived mechanisms and mediated by host and viral protein expression (Nguyen et al., 2013). The timing and level of baculovirus gene expression were herein identified as the main factors driving clustering of infected insect cells. Trajectory analysis allowed us to confirm this, in which a clear path along pseudotime is observed although cells separate and order across multiple branches spanning the transcriptomic space.
Biological mechanisms associated with baculovirus infection and transgenes (in our case those coding for influenza HA-VLPs) expression can be identified by correlating changes in gene(s) expression to progression of cells along the infection trajectory. Through pathway enrichment analysis, we found the endocytosis pathway as being one of the most significantly enriched biological processes during infection, which derives from viruses exploiting cellular structures towards endocytosis-mediated viral nucleocapsid transport to the nucleus (Monteiro et al., 2012). In addition, entry of baculovirus is found dependent on clathrin-mediated endocytosis (Long et al., 2006), which was herein corroborated by the upregulation of clathrin cltc and actin-related arpc5 and capza1 proteins at early infection stages.
Among the cellular defense responses to environmental and pharmacological stresses, the activation of heat shock response (HSR) is one of the most important. It leads to rapid and robust expression of members of the chaperone family of heat shock proteins (HSPs) in order to protect the cell from proteotoxic stresses and to maintain protein homeostasis (Fujimoto and Nakai, 2010). Interestingly, viruses can exploit HSR as an infection strategy, making use of HSPs such as HSP70 and HSP90 for regulation of viral gene expression and capsid assembly/disassembly (Mayer and Bukau, 2005; Xiao et al., 2010; Nagy et al., 2011). Our data corroborates this, with the expression of hsp70 found significantly upregulated early in infection. In addition, the proteasome pathway was found enriched, in agreement with the reported evidence of close collaboration between HSPs and ubiquitin-proteasome system during the baculovirus replicative cycle (Katsuma et al., 2011).
Baculovirus infection induces an important metabolic burden on insect cells, enhancing the fluxes through the major catabolic pathways including the tricarboxylic acid cycle (TCA) (Bernal et al., 2009). Within the TCA cycle, the citrate synthase cs gene is involved in aerobic energy production and metabolic interconversions in mitochondria (Holloszy et al., 1970); in our analysis, the expression of cs was found significantly increased at the onset of infection, suggesting that this gene plays a key role as a first-line response to infection.
Overall, the enrichment analysis allowed to identify several pathways (e.g., ribosome, spliceosome, oxidative phosphorylation) that were common to those previously identified in our bulk RNA sequencing study (Silvano et al., 2022), demonstrating the robustness and replicability of the data. Importantly, single-cell RNA sequencing allowed to evaluate single cells at different states of infection within the same sample and capture the transcriptional changes associated with the infection process (not possible with the bulk RNA sequencing approach), thus elevating the importance of single-cell omics analysis in the IC-BEVS system.
Single-cell transcriptomics enabled us to study host cell and baculovirus gene expression patterns at a resolution previously unobtainable in a bulk approach, allowing to isolate traces of different stages of infection progression. Such understanding can be further applied through genetic engineering approaches for overexpression/knock-out of specific genes, an approach that opens possibilities such as developing cell lines specialized for either virus replication or foreign protein expression, establish inducible systems, and even stimulate infection synchronization across all cells in culture towards a more controlled, homogeneous production process. Notwithstanding, using scRNA-seq to study additional IC-BEVS processes (i.e., comprising different products of interest, infection conditions, among others) is crucial for a more broader understanding of the transcriptome footprints of this expression system.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA911494. The sensitive nature of some of the reagents used in this study (e.g., cell lines, plasmids, baculoviruses, and antibodies) means that they are only readily available internally to the author’s institutions staff for the R&D purposes. For external researchers, approval of reagents request may be obtained via e-mail addressed to the corresponding author.
Conceptualization, MS, AR, II, and NV; Investigation, MS, RC, and NV; data curation, MS; data analysis, MS, RC, NV, and AR; supervision, AR, II, PA, and CC; funding acquisition, PA, AR, and II; writing—original draft, MS; writing—review and editing, MS, II, CC, AR, PA, RC, and NV.
This work has received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Sklodowska-Curie grant agreement No. 813453 and the grant agreement N° 730964, and by Fundação para a Ciência e a Tecnologia/Ministério da Ciência, Tecnologia e Ensino Superior (FCT/MCTES, Portugal) through the following initiatives: “Investigador FCT” Program (IF/01704/2014), Exploratory Research and Development Project (IF/01704/2014/CP1229/CT0001), PhD fellowships (SFRH/BD/134107/2017), iNOVA4Health (UIDB/04462/2020 and UIDP/04462/2020) and the Associate Laboratory LS4FUTURE (LA/P/0087/2020). Funding from INTERFACE Programme, through the Innovation, Technology, and Circular Economy Fund (FITEC), is gratefully acknowledged.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fbioe.2023.1143255/full#supplementary-material
Supplementary Figure S1 | Number of total UMIs per cell at each time point after cell filtering.
Supplementary Figure S2 | Cell cycle association to non-infected and infected insect cells. (A) Cell cycle score assignment for merged scRNA-seq dataset. (B) Proportion of cells assigned to each cell cycle phase. Pink, green and blue are the colors assigned to cells in G2M, S, G1, respectively. (C) Relative expression of genes involved in G1 phase: Cyclin D3 (ccdn3), Cyclin D1 (ccne1), and Cyclin-dependent kinase 6 (cdk6).
Supplementary Figure S3 | Relative expression of genes associated to pathways found enriched in pseudotime analysis. Cltc, arpc5, and capza1 are genes involved in the endocytosis pathway; hsp70, psmb1, and cs are genes involved in the protein processing in ER, proteasome and citrate cycle (TCA cycle), respectively. A color gradient scale was used to show the relative gene expression per cell.
Benjaminit, Y., and Hochberg, Y. (1995). Controlling the False Discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 57, 289–300. doi:10.1111/j.2517-6161.1995.tb02031.x
Bernal, V., Carinhas, N., Yokomizo, A. Y., Carrondo, M. J. T., and Alves, P. M. (2009). Cell density effect in the baculovirus-insect cells system: A quantitative analysis of energetic metabolism. Biotechnol. Bioeng. 104 (1), 162–180. doi:10.1002/bit.22364
Braunagel, S. C., Parr, R., Belyavskyi, M., and Summers, M. D. (1998). Autographa californica nucleopolyhedrovirus infection results in Sf9 cell cycle arrest at G 2/M phase. Virology 244, 195–211. doi:10.1006/viro.1998.9097
Cao, Z. J., and Gao, G. (2022). Multi-omics single-cell data integration and regulatory inference with graph-linked embedding. Nat. Biotechnol. 40, 1458–1466. doi:10.1038/s41587-022-01284-4
Chen, Y.-R., Zhong, S., Fei, Z., Gao, S., Zhang, S., Li, Z., et al. (2014). Transcriptome responses of the host Trichoplusia ni to infection by the baculovirus Autographa californica multiple nucleopolyhedrovirus. J. Virology 88 (23), 13781–13797. doi:10.1128/jvi.02243-14
Chen, Y.-R., Zhong, S., Fei, Z., Hashimoto, Y., Xiang, J. Z., Zhang, S., et al. (2013). The transcriptome of the baculovirus Autographa californica multiple nucleopolyhedrovirus in Trichoplusia ni cells. J. Virology 87 (11), 6391–6405. doi:10.1128/jvi.00194-13
Correia, R., Fernandes, B., Alves, P. M., Carrondo, M. J. T., and Roldão, A. (2020). Improving influenza HA-VLPs production in insect high five cells via adaptive laboratory evolution. Vaccines 8 (4), 589–615. doi:10.3390/vaccines8040589
Dobin, A., Davis, C. A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., et al. (2013). Star: Ultrafast universal RNA-seq aligner. Bioinformatics 29 (1), 15–21. doi:10.1093/bioinformatics/bts635
Drugmand, J. C., Schneider, Y. J., and Agathos, S. N. (2012). Insect cells as factories for biomanufacturing. Biotechnol. Adv. 30 (5), 1140–1157. doi:10.1016/j.biotechadv.2011.09.014
Du, X., and Thiem, S. M. (1997). Responses of insect cells to baculovirus infection: Protein synthesis shutdown and apoptosis. J. VIROLOGY 71 (10), 7866–7872. doi:10.1128/jvi.71.10.7866-7872.1997
Fujimoto, M., and Nakai, A. (2010). The heat shock factor family and adaptation to proteotoxic stress. FEBS J. 277 (20), 4112–4125. doi:10.1111/j.1742-4658.2010.07827.x
Goldman, S. L., MacKay, M., Afshinnekoo, E., Melnick, A. M., Wu, S., and Mason, C. E. (2019). The impact of heterogeneity on single-cell sequencing. Front. Genet. 10. doi:10.3389/fgene.2019.00008
Götz, S., García-Gómez, J. M., Terol, J., Williams, T. D., Nagaraj, S. H., Nueda, M. J., et al. (2008). High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 36 (10), 3420–3435. doi:10.1093/nar/gkn176
Gulati, G. S., Sikandar, S. S., Wesche, D. J., Manjunath, A., Bharadwaj, A., Berger, M. J., et al. (2020). Single-cell transcriptional diversity is a hallmark of developmental potential. Available at: http://science.sciencemag.org/.
Hao, Y., Hao, S., Andersen-Nissen, E., Mauck, W. M., Zheng, S., Butler, A., et al. (2021). Integrated analysis of multimodal single-cell data. Cell 184 (13), 3573–3587.e29. doi:10.1016/j.cell.2021.04.048
Holloszy, J. 0, Oscai, L. B., Don, I. J., and Mo, P. A. (1970). Mitochondrial citric acid cycle and related enzymes: Adaptive response to exercise. Biochem. biophysical Res. Commun. 40 (6), 1368–1373. doi:10.1016/0006-291X(70)90017-3
Ianevski, A., Giri, A. K., and Aittokallio, T. (2022). Fully-automated and ultra-fast cell-type identification using specific marker combinations from single-cell transcriptomic data. Nat. Commun. 13 (1), 1246. doi:10.1038/s41467-022-28803-w
Tennant, (1964). Evaluation of the trypan blue technique for determination of cell viability. Transplantation 2, 685–694. doi:10.1097/00007890-196411000-00001
Karlsson, M., Zhang, C., Méar, L., Zhong, W., Digre, A., Katona, B., et al. (2021). A single-cell type transcriptomics map of human tissues. Sci. Adv. 7, eabh2169. doi:10.1126/sciadv.abh2169
Katsuma, S., Tsuchida, A., Matsuda-Imai, N., Kang, W. K., and Shimada, T. (2011). Role of the ubiquitin-proteasome system in Bombyx mori nucleopolyhedrovirus infection. J. General Virology 92 (3), 699–705. doi:10.1099/vir.0.027573-0
Lee, J., Hyeon, D. Y., and Hwang, D. (2020). Single-cell multiomics: Technologies and data analysis methods. Exp. Mol. Med. 52 (9), 1428–1442. doi:10.1038/s12276-020-0420-2
Lewis, A. M., Abu-Absi, N. R., Borys, M. C., and Li, J. (2016). The use of ’omics technology to rationally improve industrial mammalian cell line performance. Biotechnol. Bioeng. 113, 26–38. doi:10.1002/bit.25673
Long, G., Pan, X., Kormelink, R., and Vlak, J. M. (2006). Functional entry of baculovirus into insect and mammalian cells is dependent on clathrin-mediated endocytosis. J. Virology 80 (17), 8830–8833. doi:10.1128/jvi.00880-06
Mayer, M. P., and Bukau, B. (2005). “Hsp70 chaperones: Cellular functions and molecular mechanism,” in Cellular and molecular life Sciences, 62, 670–684. doi:10.1007/s00018-004-4464-66
McDavid, A., Finak, G., and Gottardo, R. (2016). The contribution of cell cycle to heterogeneity in single-cell RNA-seq data. Nat. Biotechnol. 34 (6), 591–593. doi:10.1038/nbt.3498
McInnes, L., Healy, J., and Melville, J. (2018). Umap: Uniform Manifold approximation and projection for dimension reduction. Available at: http://arxiv.org/abs/1802.03426.
Mena, J. A., Ramírez, O. T., and Palomares, L. A. (2003). Titration of non-occluded baculovirus using a cell viability assay. BioTechniques 34 (2), 260–264. doi:10.2144/03342bm05
Monteiro, F., Carinhas, N., Carrondo, M. J. T., Bernal, V., and Alves, P. M. (2012). Toward system-level understanding of baculovirus-host cell interactions: From molecular fundamental studies to large-scale proteomics approaches. Front. Microbiol. 3. doi:10.3389/fmicb.2012.00391
Nagy, P. D., Wang, R. Y., Pogany, J., Hafren, A., and Makinen, K. (2011). Emerging picture of host chaperone and cyclophilin roles in RNA virus replication. Virology 411 (2), 374–382. doi:10.1016/j.virol.2010.12.061
Nguyen, Q., Nielsen, L. K., and Reid, S. (2013). Genome scale transcriptomics of baculovirus-insect interactions. Viruses 5 (11), 2721–2747. doi:10.3390/v5112721
Olsson, L., Rugbjerg, P., Torello Pianale, L., and Trivellin, C. (2022). “Robustness: Linking strain design to viable bioprocesses,” in Trends in Biotechnology. Elsevier Ltd. 40 (8), 918–931. doi:10.1016/j.tibtech.2022.01.004
Roldão, A., Oliveira, R., Carrondo, M. J. T., and Alves, P. M. (2009). Error assessment in recombinant baculovirus titration: Evaluation of different methods. J. Virological Methods 159 (1), 69–80. doi:10.1016/j.jviromet.2009.03.007
Russell, A. B., Trapnell, C., and Bloom, J. D. (2018). Extreme heterogeneity of influenza virus infection in single cells. doi:10.7554/eLife.32303.001
Samoudi, M., Masson, H. O., Kuo, C. C., Robinson, C. M., and Lewis, N. E. (2021). “From omics to cellular mechanisms in mammalian cell factory development,” in Current Opinion in Chemical Engineering. Elsevier Ltd. 32. doi:10.1016/j.coche.2021.100688
Schmitz, J., Noll, T., and Grünberger, A. (2019). “Heterogeneity studies of mammalian cells for bioproduction: From tools to application,” in Trends in Biotechnology. Elsevier Ltd. 37 (6), 645–660. doi:10.1016/j.tibtech.2018.11.007
Silvano, M., Correia, R., Virgolini, N., Clarke, C., Alves, P. M., Isidro, I. A., et al. (2022). Gene expression analysis of adapted insect cells during influenza VLP production using RNA-sequencing. Viruses 14 (10), 2238. doi:10.3390/v14102238
Stockholm, D., Benchaouir, R., Picot, J., Rameau, P., Neildez, T. M. A., Landini, G., et al. (2007). The origin of phenotypic heterogeneity in a clonal cell population in vitro. PLoS ONE 2 (4), e394. doi:10.1371/journal.pone.0000394
Sun, J., Vera, J. C., Drnevich, J., Lin, Y. T., Ke, R., and Brooke, C. B. (2020). Single cell heterogeneity in influenza A virus gene expression shapes the innate antiviral response to infection. PLoS Pathog. 16 (7), e1008671. doi:10.1371/journal.ppat.1008671
Suomalainen, M., and Greber, U. F. (2021). Virus infection variability by single-cell profiling. Viruses 13 (8), 1568. doi:10.3390/v13081568
Tirosh, I., Izar, B., Prakadan, S. M., Wadsworth, M. H., Treacy, D., Trombetta, J. J., et al. (2016). Dissecting the multicellular ecosystem of metastatic melanoma by single-cell RNA-seq. Available at: http://science.sciencemag.org/.
Trapnell, C., Cacchiarelli, D., Grimsby, J., Pokharel, P., Li, S., Morse, M., et al. (2014). The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 32 (4), 381–386. doi:10.1038/nbt.2859
Tripathi, N. K., and Shrivastava, A. (2019). “Recent developments in bioprocessing of recombinant proteins: Expression hosts and process development,” in Frontiers in Bioengineering and Biotechnology. Frontiers Media S.A. 7. doi:10.3389/fbioe.2019.00420
Tzani, I., Herrmann, N., Carillo, S., Spargo, C. A., Hagan, R., Barron, N., et al. (2021). Tracing production instability in a clonally derived CHO cell line using single-cell transcriptomics. Biotechnol. Bioeng. 118 (5), 2016–2030. doi:10.1002/bit.27715
Vieira, H. L. A., Estêvão, C., Roldão, A., Peixoto, C. C., Sousa, M. F. Q., Cruz, P. E., et al. (2005). Triple layered rotavirus VLP production: Kinetics of vector replication, mRNA stability and recombinant protein production. J. Biotechnol. 120 (1), 72–82. doi:10.1016/j.jbiotec.2005.03.026
Virgolini, N., Hagan, R., Correia, R., Silvano, M., Fernandes, S., Alves, P. M., et al. (2022). Transcriptome analysis of Sf9 insect cells during production of recombinant Adeno-associated virus. Biotechnol. J. 18, 2200466. doi:10.1002/biot.202200466
Wagner, D. E., and Klein, A. M. (2020). Lineage tracing meets single-cell omics: Opportunities and challenges. Nat. Rev. Genet. 21 (7), 410–427. doi:10.1038/s41576-020-0223-2
Xiao, A., Wong, J., and Luo, H. (2010). Viral interaction with molecular chaperones: Role in regulating viral infection. Archives Virology 155 (7), 1021–1031. doi:10.1007/s00705-010-0691-3
Keywords: IC-BEVS, influenza VLP, single-cell RNA sequencing, cell heterogeneity, pathway analysis
Citation: Silvano M, Virgolini N, Correia R, Clarke C, Isidro IA, Alves PM and Roldão A (2023) Dissecting insect cell heterogeneity during influenza VLP production using single-cell transcriptomics. Front. Bioeng. Biotechnol. 11:1143255. doi: 10.3389/fbioe.2023.1143255
Received: 12 January 2023; Accepted: 20 February 2023;
Published: 06 March 2023.
Edited by:
Frank Delvigne, University of Liège, BelgiumReviewed by:
Qingnan Liang, Baylor College of Medicine, United StatesCopyright © 2023 Silvano, Virgolini, Correia, Clarke, Isidro, Alves and Roldão. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: António Roldão, YXJvbGRhb0BpYmV0LnB0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.