 Paige Allard
Paige Allard Fotini Papazotos
Fotini Papazotos Laurent Potvin-Trottier
Laurent Potvin-Trottier
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Bioeng. Biotechnol. , 12 October 2022
Sec. Nanobiotechnology
Volume 10 - 2022 | https://doi.org/10.3389/fbioe.2022.968342
This article is part of the Research Topic Next-Generation Quantitative and Synthetic Biology: High-sensitivity, High-accuracy, and Digital Approaches View all 9 articles
Cells are inherently dynamic, whether they are responding to environmental conditions or simply at equilibrium, with biomolecules constantly being made and destroyed. Due to their small volumes, the chemical reactions inside cells are stochastic, such that genetically identical cells display heterogeneous behaviors and gene expression profiles. Studying these dynamic processes is challenging, but the development of microfluidic methods enabling the tracking of individual prokaryotic cells with microscopy over long time periods under controlled growth conditions has led to many discoveries. This review focuses on the recent developments of one such microfluidic device nicknamed the mother machine. We overview the original device design, experimental setup, and challenges associated with this platform. We then describe recent methods for analyzing experiments using automated image segmentation and tracking. We further discuss modifications to the experimental setup that allow for time-varying environmental control, replicating batch culture conditions, cell screening based on their dynamic behaviors, and to accommodate a variety of microbial species. Finally, this review highlights the discoveries enabled by this technology in diverse fields, such as cell-size control, genetic mutations, cellular aging, and synthetic biology.
Genetically identical cells can display strikingly different phenotypes within a fixed environment. In multicellular organisms, this is evident through cell differentiation and specialization. Unicellular organisms can also perform specialized behaviors within a group, such as distinct metabolic states (Rosenthal et al., 2018) or discrete roles in biofilm formation (Arnaouteli et al., 2021; Dar et al., 2021). In addition, some species prepare for changing environmental conditions on a population level by keeping a small fraction of the population ready for such changes, a phenomenon referred to as “bet-hedging” (Veening et al., 2008; Morawska et al., 2022). Because biochemical reactions depend on physical interactions between low-abundance molecules, these reactions are inherently stochastic, which results in heterogeneous gene expression between genetically identical cells exposed to the same environment (Elowitz et al., 2002; Paulsson, 2005; Raj and van Oudenaarden, 2008). For these reasons, techniques that can measure single-cell properties rather than population averages have revealed important information about many cellular processes, from cell-size control to differentiation. For example, the ability to measure the mRNA profile of single-cells through techniques such as single-cell RNA-seq (scRNA-seq) has advanced many fields, as evidenced by the breadth and the number of publications in recent years (Karaayvaz et al., 2018; Kinker et al., 2020; Peyrusson et al., 2020). While such techniques have proven to be very useful, they are typically limited to static snapshots and cannot follow gene expression in individual cells over time. Instead, this is typically achieved using single-cell time-lapse fluorescence microscopy, which for microbes has traditionally involved tracking the growth of single bacteria into microcolonies on agar pads. While technically simple and very useful, agar pads only support growth for a short period of time before cells start competing for nutrients, limiting observation to a few cell divisions (Young et al., 2011; Moffitt et al., 2012). Due to limitations in using wide-field fluorescence microscopy (i.e., the point spread function has a long tail), cellular crowding can impact the fluorescence imaging measurements, meaning that the presence of many cells can bias the measured fluorescence of a cell far away (Hardo and Bakshi, 2021). Finally, it can be difficult to change the environmental conditions in this setup, which is important for studying processes such as stress responses. These limitations motivated the development of microfluidic platforms for single-cell analysis that enables single-cell tracking over many generations under precisely controlled conditions.
The first iterations of microfluidic devices for imaging bacteria utilized closed linear trenches (Balaban et al., 2004) or mono-layer chambers (Cookson et al., 2005) to trap cells. The mono-layer chamber devices (Cookson et al., 2005; Ullman et al., 2013) enable the growth of microcolonies while flushing extra cells and continuously providing growth media, and have been particularly useful for studying group behaviors such as quorum sensing (Danino et al., 2010; Prindle et al., 2011; Din et al., 2016; Scott and Hasty, 2016; Miano et al., 2020). Continuous culture devices were then developed to enable long-term imaging of bacterial cells undergoing steady-state growth while facilitating the tracking of single cells without crowding limitations, making them ideal for measuring single-cell gene expression (Wang et al., 2010; Moffitt et al., 2012). A configuration nicknamed the “mother machine” (MM) traps bacteria at the end of single-cell-width dead-end trenches (Wang et al., 2010). Newborn cells are flushed out of the device by the constant flow of growth media, thereby allowing single-cell lineages to be followed for hundreds of generations (Figure 1). The name “mother machine” refers to the fact that the cells trapped at the end of the trenches are tracked growing and dividing throughout an experiment, and are thus referenced as “mother” cells. A similar layout named the chemostat (Moffitt et al., 2012) has trenches open on both sides, thus providing better feeding through convective flow. This device enables long-term time-lapse microscopy while keeping cells that renew both poles and has been used to quantify the maturation time of fluorescent proteins (Balleza et al., 2017).
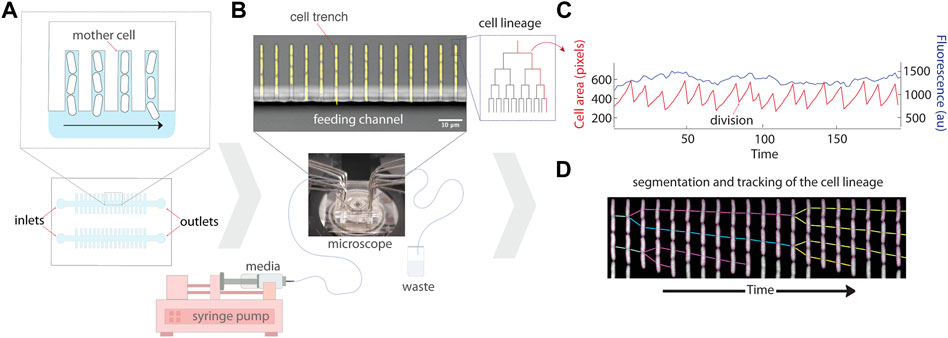
FIGURE 1. Schematic of the experimental setup for the mother machine microfluidic device and data analysis. (A) Schematic representation of the platform which traps single bacterial cells in trenches that are perpendicular to a larger feeding channel. Daughter cells are flushed out of the trenches with flowing media, while mothers remain trapped at the end of the cell trench. (B) A micrograph of the mother machine, with YFP fluorescence showing the cells superimposed on a brightfield image of the device. Media is pumped through the inlet into the main feeding channel by a syringe pump, and then exits through the outlet into a waste beaker. (C) The lineages of growing cells in the trenches can then be followed under precisely controlled environmental conditions using time-lapse microscopy. (D) An example kymograph of a growing cell imaged in fluorescence, illustrating the segmentation and tracking of the lineage.
In this review, we focus on the technical developments of the MM and its applications in the study of bacteria. We start by describing the original design and the constraints that motivated further technical developments. We then overview recent image analysis tools that enable segmentation and tracking of cells in the device using phase contrast images, and modifications to the device that enables the precise control of environmental conditions, screening and isolation of cells, and cultivation of a variety of microbial species. Finally, we highlight how the MM enabled discoveries in a wide range of fields in microbiology, such as cell-size control, genetic mutations, cellular aging, stress responses, cell-fate determination, antibiotic tolerance/persistence, and synthetic biology. For more details, we refer the interested reader to previous reviews that have comprehensively described device fabrication and setup (Weibel et al., 2007; Hol and Dekker, 2014; Eland et al., 2016), discussed challenges associated with single-cell analysis (Yang et al., 2016; Hardo and Bakshi, 2021), and reviewed different microfluidic devices (Weibel et al., 2007; Bennett and Hasty, 2009; Wessel et al., 2013; Vasdekis and Stephanopoulos, 2015; Potvin-Trottier et al., 2018; Scheler et al., 2019).
Performing experiments with the MM requires the following: designing and fabricating a mold for the device, building a microfluidic chip, performing the time-lapse microscopy, and analyzing the images to create time traces. Here, we start with an overview of the original MM design and the challenges that motivated recent technical developments.
The original MM design was developed to study Escherichia coli cells (Wang et al., 2010). Narrow trenches trap bacterial cells perpendicular to a larger feeding channel that allows media to deliver nutrients to the cells and wash away progeny emerging from the trenches (Figures 1A,B). Several constraints must be considered in the design: the width, height, and length of cell trenches, spacing between cell trenches, and dimensions of the feeding channel. The width and height of the trenches are approximately the dimensions of the particular strain of bacteria being cultivated (e.g., 1.2 μm height, 1.3 μm width, and 20 μm length for E. coli MG1655). Proper trench dimensions ensure that the cells are in focus, restrict growth to single file within each trench, and ensure sufficient diffusion of nutrients to all cells in a trench. Too large of dimensions results in cells overlapping each other in the z direction–making segmentation and tracking hard to impossible. Too small of dimensions leads to difficulties loading the cells into the trenches and results in poor nutrient diffusion to cells deep in the trenches, including the mother cells. The chosen length of a cell trench is a trade-off between cell retention over time, as short trenches lose cells more rapidly (e.g., through stochastic filamentation that pulls them out in the feeding channel), and feeding of the mother cell. It is thus important to ensure that the growth rate of the mother cell is the same inside the device and in batch culture for each strain and device combination (Yang et al., 2018). Spacing between cell trenches is a trade-off between the throughput (i.e., number of lineages followed per image) and accuracy of fluorescence measurements. Trenches too close to one another can result in biased fluorescence measurements, particularly if neighboring trenches have very different signal intensities (Kaiser et al., 2018; Hardo and Bakshi, 2021). Finally, the width and height of the feeding channel are chosen to minimize hydraulic resistance to facilitate the flow of growth medium, e.g., with syringe pumps. A single centimetre-sized chip can typically fit multiple channels, each with their own inlet and outlet, to enable simultaneous experiments with multiple strains (Figure 1A).
After the design is finalized, a mask can be drawn using CAD software and ordered through different companies (e.g., Toppan Photomasks, United States). The mold is then built on a silicon wafer in a cleanroom environment using photolithography techniques, where the mask is used to expose photo-sensitive resin to light to define the features (Weibel et al., 2007; Eland et al., 2016). The mold is a negative of the features of the chips (i.e., what is solid on the mold becomes air in the device). The ∼1 µm-sized features are on the lower end of the resolution of these techniques, and smaller features (e.g., Section 2.4) require other fabrication techniques (e.g., electron beam lithography). As such, fabrication protocols typically require fine-tuning to obtain the critical feature size (cell trench width and height) within a ∼0.1 µm range, which can affect the experiments as described above. However, once built these molds can be re-used indefinitely to build microfluidic chips. Alternatively, the mold can be ordered custom-built from companies such as: ConScience (Sweden), Cornell NanoScale Science and Technology Facility (United States), Innopsys (France), Kavli Nanolab Delft (Netherlands), Micro Resist Technology (Germany), Sigatec (Switzerland), and TTP (UK). Molds can also be duplicated in epoxy, which can be an inexpensive option for sharing molds between groups (Kamande et al., 2015).
The MM microfluidic chip is made by pouring and curing polydimethylsiloxane (PDMS) on top of the mold, imprinting the features of the mold onto the chip. Individual chips are then cut out of the PDMS slab, and holes are punched (e.g., with biopsy punchers) at the inlets and outlets to allow the connection of tubing which provides growth medium to the device. The chips are then covalently bonded to a coverglass using plasma treatment. Cells can then be loaded into trenches via centrifugation with a custom adapter, or by simple diffusion by loading a very dense culture. Tubing is connected to the inlets and outlets (e.g., using syringe needles) for flow of fresh growth media into the chip, as well as removal of used media which has passed through the chip (Figure 1B). The media is typically pushed through the device using syringe pumps, initially at a high flow to clear biofilm that may be growing in dead spaces (i.e., regions of low flow) in the inlets and outlets. The formation of biofilms can be limited by pre-coating the chip with bovine serum albumin (BSA) and/or supplementing the growth media with BSA or Pluronic (Ullman et al., 2013; Cabeen and Losick, 2018). The chip is then mounted on an inverted fluorescence microscope for automated time-lapse microscopy of the lineages growing in the trenches. A cage incubator and hardware autofocus are typically required to ensure stability of the focus over multiple days. Detailed protocols for setting up MM experiments have been published (Cabeen and Losick, 2018). After the experiment, the multi-dimensional images (position, fluorescence channels, time) can be processed to track the properties of single lineages growing in the device (Figure 1C; Section 2.2).
The original design of the device enabled time-lapse microscopy of single E. coli cells under controlled growth conditions, leading to many biological discoveries. There are however some challenges associated with the experimental setup, which have led to new technical developments discussed below. For example, automated image processing traditionally required constitutive expression of a fluorescent protein (FP) at intermediate levels specifically for that purpose. This limited the number of simultaneous reporters that could be used in an experiment, while also preventing the study of bacteria not expressing exogenous FPs. Section 2.2 describes the recent tools developed for segmentation and tracking of cells using phase contrast images. In addition, the original setup of the MM makes switching conditions within an experiment challenging. With this design, growth media can be changed using a Y-junction close to the chip, although this creates a short (and imprecise) delay in the switch as the medium between the Y-junction and the chip is being replaced. Creating complex environmental conditions within the chip, i.e., by flowing a growing culture, can be challenging due to the introduction of air bubbles into the feeding channel. Such challenges have inspired the development of more sophisticated fluidic and environmental control strategies, which are detailed in Section 2.3. Another limitation of the original design is the inability to genotype or isolate cells within the device, which can be useful when imaging libraries of cells. While phenotyping pooled genetic libraries is possible in the device, isolating single cells of interest from the conventional MM platform has been impractical due to the inevitable formation of biofilms in the inlets and outlets, which additionally limit the total duration of an experiment. While cells could potentially be genotyped on-chip through fluorescence in situ hybridization (FISH) techniques, diffusion of probes through the cell trenches makes this process inefficient. These limitations have led to the development of techniques for single-cell screening detailed in Section 2.4. Finally, the original device was limited to culturing E. coli or other similarly sized microbes. Modifications to trench dimensions and the addition of other features have now allowed the growth and imaging of a variety of other microbes. The cultivation of some species requires additional modifications, which are further described in Section 2.5.
While the experimental techniques for using the MM have become increasingly accessible, the image analysis pipeline to convert time-lapse images into single-cell traces through segmentation and tracking has lagged. For many years, laboratories using the MM have developed their own customized data analysis pipelines. While tools have been developed for tracking the growth of microbes on agar pads using phase contrast (Paintdakhi et al., 2016), the large features of the MM PDMS chip can confound such image analysis tools. Recently, multiple open-source software packages have been published specifically for MM experiments with the capability of performing the segmentation and tracking on phase contrast images (listed in Table 1).
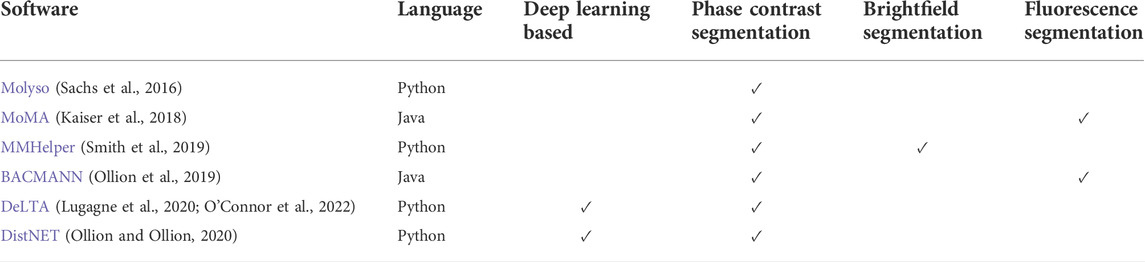
TABLE 1. Overview of open-source software packages developed for mother machine segmentation and lineage tracking. The applicability to specific imaging modalities indicates which ones have been demonstrated. All programs are available on Github or Gitlab.
Most of these image analysis methods share a common overall workflow: pre-processing of the images, segmentation of the cells, and tracking the lineages. In the pre-processing step, the image time series are first registered to correct for drift and jitters of the stage, for example, using cross-correlation between successive images. The images are rotated to align the micro-channels vertically, which simplifies further analyses. The micro-channels are then identified and segmented. Accurately segmenting individual cells is typically the most challenging task, as the cells are small and in contact with each other, and strategies vary between implementations. Tracking is then performed to create time traces of individual cells, where cells from each time point are connected to the cells in the next one, and cell divisions are identified (Figure 1C). Properties such as cell size and fluorescence intensity are extracted along these single-cell time traces. Finally, a manual curation pipeline is typically available, as even rare segmentation and tracking errors can have large effects on sensitive measurements such as the variance.
In Molyso (Sachs et al., 2016), the cell segmentation is done in one dimension with cells identified by bounding rectangles instead of cell contour. The tracking is done by solving an optimization problem, where a cost is imputed for cell displacement between time points and cell division events. MoMA generalized the optimization problem for both the segmentation and tracking, overpredicting possible cell segmentation and performing the tracking simultaneously (Kaiser et al., 2018). This 1D segmentation works well when the cells are perfectly aligned with the channels. However, it can result in errors when the cell width is smaller than the channel width, resulting in tilted cells, or if the cells are not perfectly rod-shaped (e.g., mutants or other bacteria with different morphologies). BACMANN enables 2D segmentation (i.e., cell contour) through watershed-based image processing techniques, while the tracking is based on the position with respect to the top of the trench (Ollion et al., 2019). BACMANN also incorporates a spot-tracking algorithm in its pipeline. MMHelper was developed using similar segmentation approaches to also segment using bright-field instead of phase contrast (Smith et al., 2019). DeLTA (Lugagne et al., 2020) utilizes 3 U-net convolution neural networks (Ronneberger et al., 2015) to perform channel identification, cell segmentation, and tracking. DistNET incorporates a self-attention layer into the U-net architecture to provide information about the whole channel to the neural network, and performs segmentation and tracking through one deep neural network (Ollion and Ollion, 2020). Comparison of BACMANN, DeLTA, and DistNET on the same dataset showed that they could achieve <1% combined segmentation and tracking error rates (Ollion and Ollion, 2020). While BACMANN’s tracking performed better than DeLTA, DeLTA’s segmentation performed better. DistNET’s self-attention layer mainly improved the tracking performance of DeLTA. Benchmarking the techniques against data from different experimental conditions showed good performance but an increased error rate, suggesting a need for more training data or for changing the analysis parameters (Ollion and Ollion, 2020).
When choosing a program, we encourage users to consider the following criteria: accuracy of segmentation and tracking, need for manual curation, speed of analysis, quality of documentation, readability of code, ease of use, flexibility to specific experimental needs, and an actively maintained code-base. Deep-learning methods can be fast and very accurate while requiring training on a relatively small set of manually analyzed data. However, they often lose accuracy when the experimental conditions are different from the training set (i.e., cell size, trench width, etc.). Conventional methods typically require changing analysis parameters to accommodate these kinds of changes, but that can be less tedious than manual segmentation of many images required to re-train a deep-learning algorithm. Generating synthetic data to train the neural networks would greatly alleviate their main shortcomings.
One key strength of the MM is the precise control over the growth conditions. However, switching between conditions within an experiment and flowing mixed media are challenging using the original design. A rapid and precise switch between growth media can be obtained by modifying the device design to include two inlets for each feeding channel. To flow a mixture of media, it is necessary to introduce a serpentine channel between the inlet and the feeding channel to overcome the mixing limitation of the low Reynolds number environment. The dual input mother machine (DIMM, Figure 2A) utilized this strategy to study the induction of the lac operon while switching from glucose to lactose (Kaiser et al., 2018). The authors could track the lag in growth of single cells exposed to this transition and found that the distribution of the growth lag was multi-modal. By quantifying the number of LacY/Z (“sensor”) molecules in single cells in the device, this multi-modality was subsequently attributed to a fraction of the population expressing zero LacY/Z molecules, relying instead on stochastic leaky expression for induction of the operon (Julou et al., 2020). However, any number of expressed lacY/Z molecules was sufficient for fast induction of the operon, making it a single-molecule trigger.
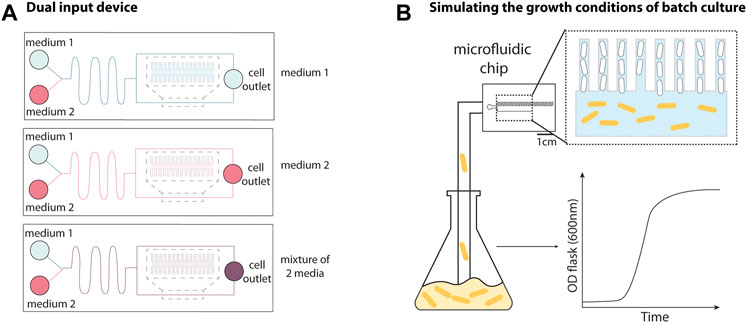
FIGURE 2. Adaptations to the mother machine architecture for improved fluidic and environmental control. (A) The dual input mother machine (DIMM) has two media inlets followed by a serpentine channel that fluid passes through prior to reaching cell trenches to facilitate mixing and/or rapid switching between different environmental conditions. Schematic representation inspired from Kaiser et al. (2018). (B) The growth curve platform allows batch culture to be fed into the device to recapitulate batch culture conditions. This allows for observation of cells entering and exiting the stationary phase by switching between nutrient depleted culture and fresh media. Schematic representation inspired from Bakshi et al. (2021).
An experimental setup was developed to mimic the conditions of batch culture (Figure 2B), where a growing culture is flowed directly into the microfluidic device, enabling the study of cells transitioning between different growth phases (Bakshi et al., 2021). This was achieved using peristaltic pumps to flow a culture growing in a separate shaker-incubator into the device, and bubble traps to prevent air bubbles from entering the device. By alternating between flowing fresh media and batch culture into the MM, the authors monitored cells after multiple rounds entering and exiting the stationary phase and found that the cell-size regulation strategy changed throughout phases of the growth curve (discussed in Section 3.1).
Genetic screens have been instrumental in biology for assigning function to molecular components and generally linking genotypes to phenotypes. Many powerful screening platforms have been developed, but they have been mostly limited to distinguishing static phenotypes in the population. Recent technical developments have transformed the MM into a powerful platform that combines dynamic phenotype screening with genotyping or isolation capabilities. This enables screening based on dynamic and/or spatially resolved phenotypes, such as oscillations in gene expression, cell-size control mechanisms, response to changes in environmental conditions, and intracellular localization of proteins. Notably, even static phenotypes could be isolated more precisely because cells can be quantified over many generations, and thus genetic and non-genetic heterogeneity could be distinguished.
Two techniques have been developed to date for screening cells in the MM. The first technique, named “dynamic µ-fluidic microscopy-based phenotyping of a library before in situ genotyping” (DuMPLING) (Lawson et al., 2017), enables dynamic phenotyping of pooled libraries. After characterization of the library in the MM, the cells of the barcoded library are fixed and identified via fluorescence in situ hybridization (FISH), connecting genotypes to dynamic phenotypes (Figure 3A). A 300 nm gap at the dead-end of the trench is connected to a back-end channel, which generates convective flow that facilitates efficient movement of probes and media over the cells. This convective flow also facilitates feeding of the mother cell therefore reducing the diffusion limitations related to the dimensions of the cell trenches discussed in Section 2.1.1. The DuMPLING platform was used to identify the effects of a CRISPR interference-mediated gene knockdown library on the coordination of replication and division by tracking chromosome replication forks throughout cell division (Camsund et al., 2020).
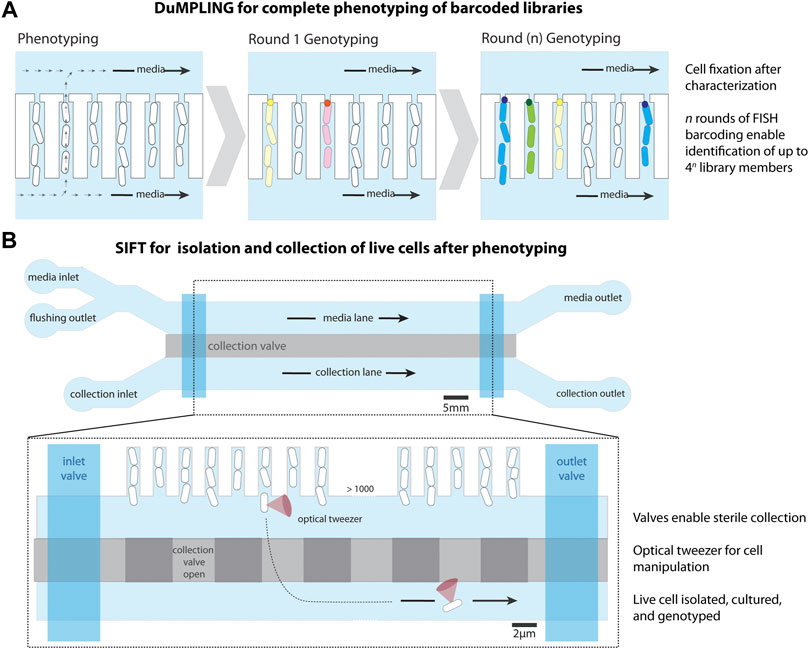
FIGURE 3. Modifications to the mother machine to enable cell screening. (A) Dynamic u-fluidic microscopy-based phenotyping of a library before in situ genotyping” (DuMPLING), has a 300 nm gap at the end of the cell trench, allowing media to flow through the cell channels. Rounds of barcoding through FISH enable genotyping of the pooled library. Schematic representation inspired from Lawson et al. (2017). (B) Single-cell isolation following time-lapse microscopy (SIFT) uses a modified microfluidic chip containing an additional lane for cell isolation below the cell trenches, separated by a pressurized valve system (collection valve). A second set of valves (inlet and outlet) allows for the lane to be sealed for inlet cleaning and restricting media flow after cell loading. An optical tweezer moves cells of interest from their trench to a collection trap, where they are isolated and removed from the device to be cultured and sequenced. Schematic representation inspired from Luro et al. (2020).
The second technique, named single-cell isolation following time-lapse microscopy (SIFT) (Luro et al., 2020), uses a modified microfluidic chip containing an additional media channel used for cell isolation (Figure 3B). The device has a system of pressurized valves that separates the cell trenches from the collection channels, temporarily closes the inlets and outlets for sterilization, and closes the media channel to stop the liquid flow. This enables an optical tweezer to move cells of interest from their growth trench to a collection trap where they are isolated, cultured, and sequenced. SIFT was used to screen two libraries of synthetic genetic oscillators based on the periodicity and precision of oscillations, showcasing its strength in isolating dynamic phenotypes.
Both of these screening techniques have advantages and disadvantages. While DuMPLING enables the in situ genotyping of entire libraries and allows genotype-phenotype mapping across a large number of cells, it requires a barcoded library and cell fixation prior to hybridization, thereby eliminating any possible downstream growth and analysis (Lawson et al., 2017). Conversely, SIFT does not require barcode labeling, enabling the screening of unmodified libraries and natural populations, the isolation of live cells, and downstream analysis of isolates (Luro et al., 2020). However, only isolated cells can be genotyped, limiting the scale of phenotype-to-genotype mapping throughput. Additionally, the optical trapping mechanism in this technique requires an extensive platform. These extended capabilities for screening and isolating cells within MM-like devices have the potential to enable discoveries in diverse fields of microbiology.
Adaptations to the MM have enabled single-cell studies of a variety of microbes. In principle, adaptation of the MM design to other symmetrically dividing organisms should only be a matter of adapting the trench size, although other minor modifications may be necessary to maintain species-specific optimal growth conditions. Such devices have been fabricated for cultivation of Corynebacterium glutamicum (Sachs et al., 2016) and Bacillus subtilis (Norman et al., 2013; Cabeen and Losick, 2018). As B. subtilis stochastically forms long multicellular chains which would be pulled out of the trenches, an adaptation of the classic MM device incorporated an increased trench length of 75 μm (Norman et al., 2013). This two-layer device included shallower feeding channels surrounding the cell trenches to ensure sufficient feeding of cells at the end of these long trenches (Figure 4A).
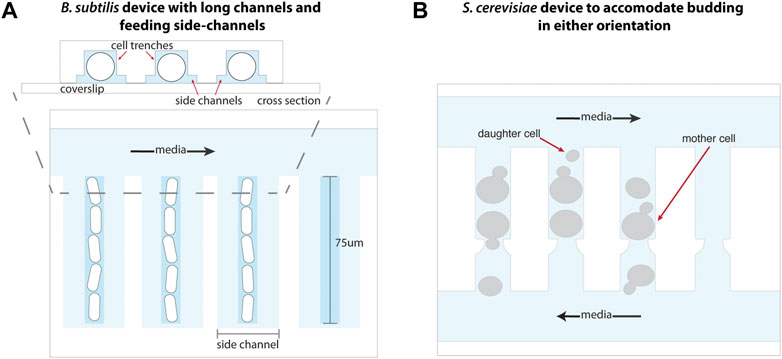
FIGURE 4. Adaptations to the mother machine architecture to optimize growth of other organisms. (A) The mother machine design adapted for B. subtilis growth includes elongated trenches 75 μm in length to accommodate its multicellular, chained state, as well as side channels that enable uniform nutrient availability throughout the trenches. Schematic representation inspired from Norman et al. (2013). (B) Maintaining an opening at either end of each cell trench in a modified MM device enables removal of S. cerevisiae daughter cells produced from budding in either orientation into perpendicular media channels. Schematic representation inspired from Li et al. (2017).
The classic MM architecture has been used to study archaea and symmetrically-dividing yeast organisms such as Halobacterium salinarum (Darnell et al., 2020) and Schizosaccharomyces pombe (Nakaoka and Wakamoto, 2017), respectively, by scaling up device dimensions. The ability of the budding yeast Saccharomyces cerevisiae to switch budding orientation over the course of its lifespan necessitated the trapping and retention of mother cells in trenches open on both ends to flowing media (Li et al., 2017). This accommodates the removal of daughter cells produced by budding in either orientation (Figure 4B). Alternatively, the “yeast jail” design forgoes trenches altogether and instead employs a microfluidic device with an array of jail units (Ryley and Pereira-Smith, 2006). Each jail is composed of three PDMS posts (among other designs) that behave as “jail bars” to retain a single mother cell, while daughter cells produced by budding are washed away with the flow of media. Similarly, A Long-term Culturing And TRApping System (ALCATRAS) (Crane et al., 2014), High-throughput Yeast Aging Analysis chip (HYAA) (Jo et al., 2015), and slipstreaming MM (Durán et al., 2020) devices comprise an array of PDMS trapping units, many of which may fit into a single field of view during imaging, to enable tracking of individual mother cells over their lifespan. In the latter, S. cerevisiae cells are loaded through the outlet of the device such that media flow reversal enables the trapping of mother cells in a low-pressure zone behind PDMS pillars. This method is ideal for studying cell replicative life span as it does not place cells under mechanical pressure during trapping, and the effect of such stress on aging is not yet known. An alternative method utilizes channels or PDMS structures of optimal heights for trapping mother cells underneath them, while permitting smaller daughter cells to be flushed away (Lee et al., 2012; Xie et al., 2012; Zhang et al., 2012).
Several features of the MM have facilitated important discoveries about a variety of cellular processes. The tracking of single cells under controlled and tunable growth conditions with high throughput has shed light on how cellular processes are affected by stochastic gene expression, and how such heterogeneity can affect the cell’s phenotype. The ability to track the same cells for many generations has facilitated studies in cellular aging, and an inherent lack of competition between cells in the device has proven useful in studying genetic mutations. Here we will highlight discoveries about cell-size control mechanisms, genetic mutations, cellular aging, stochastic pulsing of gene expression, phenotypic states in Bacillus subtilis, antibiotic resistance and persistence, and synthetic biology that have been enabled by the MM platform. Instead of a detailed description of each study, we highlight how the device has enabled such discoveries.
Most prokaryotes divide via binary fission, yielding two daughter cells of nearly identical size and volume (Angert, 2005). If left unchecked, small fluctuations in cell size at cell division can result in significant size divergence between cells of a population over successive generations. Cell-size homeostasis could in principle be achieved through different control mechanisms (Facchetti et al., 2017). Several studies have shown that E. coli, B. subtilis and Caulobacter crescentus maintain cell-size homeostasis by behaving primarily as “adders”, adding on average a constant length between birth and division, rather than as “sizers” or “timers”, for which cell division is triggered upon cells growing to a threshold size or for a fixed time, respectively (Voorn and Koppes, 1998; Amir, 2014; Campos et al., 2014; Wallden et al., 2016). As time-lapse microscopy on agar pads is limited to a few cell divisions where the growth conditions can change, microfluidic approaches have been important in studying cell-size regulation. The ability to image hundreds of single cells under constant growth conditions while precisely measuring their size throughout the cell cycle (e.g., size at birth and added during the cell cycle) has made the MM a particularly useful platform in elucidating the specific mechanisms underlying cell-size regulation (Taheri-Araghi et al., 2015; Sauls et al., 2019; Si et al., 2019; Witz et al., 2019; Nieto et al., 2020).
Several models have been suggested to explain the mechanism underlying adder behavior (Ho and Amir, 2015; Micali et al., 2018; Si et al., 2019; Witz et al., 2019; Nieto et al., 2020). While a detailed description of the different models is outside the scope of this review, a recent analysis by Le Treut et al. has found the authors’ model, the independent double adder, to be the most consistent with current data (Le Treut et al., 2021). This model proposes that the processes of cell division and initiation of DNA replication are controlled by distinct (independent) adders (Si et al., 2019). It further suggests that the mechanism underlying this type of cell-size control relies on accumulation of specific initiator proteins for independent regulation of cell division and DNA replication to a certain threshold and that they are produced at a rate proportional to cell growth without being actively degraded. This hypothesis was supported by experiments that used the MM to measure the cell size added between cell divisions, and the cell size added between DNA replication initiation events (tracked with DnaN-YPet) (Si et al., 2019), which were both found to be independent of initial cell size (i.e., adders). Perturbing the production of each initiator protein (FtsZ and DnaA respectively) only dysregulated cell-size control for its respective process.
Some studies carried out in the MM have shown deviations from the adder principle under slow growth conditions (Wallden et al., 2016; Si et al., 2019; Nieto et al., 2020). Si et al. (2019) suggest that the adder principle can be broken if the underlying mechanisms (threshold and constant production) are affected. They suggest that active degradation of the division initiatior (FtsZ) could play that role, and elimination of degradation through clpX repression restored the adder phenotype under that growth condition (Si et al., 2019). A recent study used a variation of the MM setup which flows batch culture into the device as a means of replicating conditions within that culture on-chip to monitor E. coli and B. subtilis growth, and revealed that cells alter their size regulation strategy as they progress through different growth phases (Bakshi et al., 2021). Cells switched to mixed “adder-timers” while they entered the stationary phase and behaved as “sizers” while exiting the stationary phase, suggesting that different strategies might be used to respond to changes in environmental conditions. These results also highlight the importance of using the MM for studying this process, as other single cell imaging techniques such as agar pads do not offer precise control over environmental conditions. Developments in the control of growth conditions as well as screening capabilities will thus facilitate the development of more complete models that can capture cell-size control strategies under varying environmental conditions.
Mutations and other DNA damage events such as double-stranded breaks (DSBs) have the potential to drastically affect cell survival and fitness if left unrepaired (Gordo et al., 2011). To counter such mutations, E. coli employs effective DNA repair pathways to recognize and repair genetic perturbations before they are propagated to future generations. The ability to track thousands of lineages in parallel (without growth competition between individuals) makes the MM an ideal platform to study rare events such as the emergence of DNA mutations. Its capacity to capture heterogeneity in number and types of mutations emerging between isogenic cells of a population further highlights the utility of such single-cell, time-lapse studies. To this end, the MM has been used to visualize copy number variations (Tomanek et al., 2020), DNA mismatch error (Robert et al., 2018, 2019), DSBs (Wiktor et al., 2021), and alkylation damage (Uphoff et al., 2016; Uphoff, 2018; Vincent and Uphoff, 2021).
The timing of mutations as well as their impact on the fitness of the cells are important variables in the process of evolution. The rate of chromosomal gene copy number mutations in E. coli was estimated at ∼3 × 10−3 per cell per generation in the MM (Tomanek et al., 2020). This was achieved by expressing several copies of a chromosomally integrated fluorescent reporter and measuring changes in fluorescence over time as an indicator for gene copy number (because gene expression is roughly proportional to gene copy number). Robert et al. leveraged the lack of competition in the MM to visualize the appearance of DNA replication errors and their fitness impact (Robert et al., 2018, 2019). To measure the fitness effect of the mutations, they measured the decrease in growth rate (i.e., fitness) of the individual lineages through the accumulation of mutations (Robert et al., 2018). Instantaneous single-cell growth rate can be calculated in the MM using the relative change in cell size between successive time points (i.e., doubling rate). Although the entire distribution of fitness effects (DFE, which has proven challenging to measure in the field of evolution modeling) was still not directly estimated, the authors were successfully able to infer all the moments of DFE (i.e. average, variance, skewness, etc.) - thanks to the ample statistics enabled by the MM. The analysis indicated the underlying distribution to be long-tailed, with most mutations having little-to-no cost on cell fitness: mean fitness cost was only ∼0.3 %, which was apparently overestimated by the previous studies. In contrast, 1% of the mutations were found to be lethal, directly measured by observing cell death in the MM. Mutations were detected by tracking the appearance of YFP-MutL foci, and occurred at a constant rate over time during steady state exponential growth, as expected from a memoryless Poisson process. However, another MM study showed that a subset of cells within a population exhibited a period of elevated mutation rates in response to DNA alkylation damage, due to delayed activation of the Ada DNA damage response regulon (Uphoff, 2018; Vincent and Uphoff, 2021). These periods of high mutation rate lasted several generations, since many cells had no sensor-activator Ada molecules present, and had to wait for stochastic expression to induce the response (Uphoff et al., 2016). Finally, the response to DSB has also been characterized using an adapted MM with multiple inlet ports connected to the feeding channel via a short junction, enabling rapid switching between different media (Wiktor et al., 2021). Switching between a growth medium and an induction medium enabled short induction of Cas9, which created a targeted DSB in the chromosome. The chromosomes could be tracked using fluorescently-tagged proteins (ParB and MalI) with DNA-binding sites localized close to the DSB, with the foci disappearing during the DSB. Repairs were rapid (∼15 min), homogeneous, robust (∼95% of cells repaired the damage), and had low impact on the fitness of the cells. This is impressive given that the DSB needs to find/colocalize with its repair template on the sister chromosome. The RecA-single stranded DNA complex was observed to extend in a long filament spanning the length of the cell. This could facilitate the homology search for the repair template by eliminating the need to search along the length of the cell in the z direction, thereby reducing it from a 3D problem to a 2D one and making the search process 100 times faster.
The phenomenon of aging in unicellular organisms is broadly described in terms of senescence, or the progressive loss of fitness over time (Moger-Reischer and Lennon, 2019). This loss of fitness can be due to a decrease in growth rate and/or an increase in death rate. Defining the ‘age’ of a unicellular organism that lacks replicative asymmetry can be challenging, but generally takes into account the asymmetric segregation of damage factors during cell division, which creates effectively ‘older’ and ‘younger’ progeny cells. For example, in budding yeast asymmetric division creates a finite replicative lifespan by partitioning detrimental cellular factors such as misfolded protein aggregates to older ‘mother’ cells, while preserving the daughter lineage (Knorre et al., 2018). The mother exhibits senescence over successive generations and eventual cell death. While E. coli divides symmetrically, there is still an intrinsic asymmetry in the process: one pole is created during division (the ‘new’ pole), and one is left intact (the ‘old’ pole) (Proenca et al., 2018). Asymmetries in partitioning of cellular contents have also been observed in E. coli. For example, the main efflux pump (AcrAB-TolC) was shown to be partitioned with a bias for the old pole cell, leading to elevated efflux activity in ‘older’ cells (Bergmiller et al., 2017). Non-random segregation of sister chromatids has also been observed, with the ancestral strand being partitioned preferentially in the old pole cell (Mäkelä et al., 2021). The MM is uniquely suited to study aging in symmetrically dividing bacteria as it retains an old pole cell at the end of each trench over an entire experiment, thus facilitating monitoring of the old pole lineage for many generations. Here, we present findings from several studies monitoring the aging process in E. coli in the MM, where the age of a cell is defined as the age of the old pole.
Both decreased growth rate and increased death rates have been associated with aging in E. coli (Lindner et al., 2008; Wang et al., 2010; Proenca et al., 2019; Łapińska et al., 2019). The first report of the MM studied the aging of the mother cell over consecutive generations (Wang et al., 2010). They found that the growth rate of the mother cell was stable for more than a hundred generations, but observed that the filamentation rate increased until 50 generations (Wang et al., 2010). Note that in 50 generations of exponential growth, 1 cell would divide into ∼1015 cells, making such studies intractable without the use of devices like the MM. Subsequent studies tracked the growth rate of both the mother cell and its immediate progeny, the ‘daughter’ cell. This showed that these cells reached different equilibrium growth rates, with the mother cell stabilizing at a growth rate slightly slower than the daughter cell (Proenca et al., 2018, 2019). This suggests that damaged molecules divided asymmetrically, with a preference to the old pole cell. The nature of this damage and the mechanisms underlying asymmetrical partitioning of cellular components are under investigation. Misfolded proteins are a prime suspect, as a chaperone fusion (IbpA-YFP) was shown to preferentially localize to the old pole as a foci (Lindner et al., 2008; Proenca et al., 2019), while protein stress such as phototoxic stress affected the asymmetry (Proenca et al., 2019). However, FP fusions such as YFP have been shown to create artifactual foci due to their oligomeric properties (Landgraf et al., 2012). Therefore, the use of FP fusions that do not cause aberrant foci (e.g., ClpB-msfGFP) will be informative in tracking protein aggregates (Govers et al., 2018). In contrast, another study has shown no asymmetries in misfolded protein aggregates using brightfield imaging for inclusion bodies and ThT dyes to visualize protein aggregates (Łapińska et al., 2019). Asymmetric retention of protein aggregates at the old pole has also been observed in the symmetrically dividing fission yeast S. pombe in a MM-like device (Nakaoka and Wakamoto, 2017). These aggregates were not associated with increased division times and were eventually transferred to a new daughter, thereby rejuvenating the old lineage. While it is clear that even cells dividing “almost symmetrically” like E. coli exhibit aging, the elucidation of the molecular mechanisms and the agents causing this phenomenon will shed light on the universal properties of cellular senescence, and the MM provides an ideal platform with which to study it.
Stochastic fluctuations in gene expression can drive phenotypic heterogeneity among clonal bacterial populations, such that genetically identical cells display distinct behaviors. The use of FP reporters for gene expression have been invaluable in quantifying this heterogeneity. In principle, this heterogeneity can be generated on different timescales. For example, cells can stochastically express genes at different levels for long periods of time (e.g., stable epigenetic state), or they can fluctuate rapidly between these different expression levels (e.g., rapid pulses). Long-term time-lapse microscopy using microfluidic devices such as the MM enables tracking the expression dynamics in thousands of individual cells under controlled growth conditions and can elucidate the timescales of such fluctuations. Employing this strategy, it was shown that the promoters controlling flagellar biosynthesis genes in E. coli activate in stochastic pulses even if expression of their master regulator was constant (Kim et al., 2020). Measuring expression of a gene (bolA) dependent on the E. coli general stress response factor, RpoS, revealed heterogeneous expression in liquid culture, in the MM, and in another microfluidic device (Patange et al., 2018). The promoter activity (production rate of the reporter) can be calculated using the derivative of the fluorescence intensity while accounting for dilution of the FP present (Locke et al., 2011). The promoter activity revealed stochastic pulses of expression which coincided with periods of slower growth in the device (Patange et al., 2018). The ability to rapidly change environmental conditions and measure single-cell properties prior to and after the change enabled them to test whether these periods of slow growth led to increased resistance to stress. They observed that cells with higher RpoS activity and slower growth immediately prior to the stress were more likely to survive a hydrogen peroxide treatment. Another study measured the expression of multiple stress response genes in the MM and found additional RpoS-dependent promoters exhibiting stochastic pulses that negatively correlated with growth rate (Sampaio et al., 2022). Genes from the SOS regulon also displayed pulsatile activity, but these were not correlated with the growth rate. Cells undergoing pulses of genes from both these groups prior to a short treatment of the antibiotic ciprofloxacin in the MM had increased likelihood of survival (Sampaio et al., 2022). Pulses in the SOS regulon have also been observed in another study using the MM, and have been attributed to variability in the degradation of its regulator, LexA (Jones and Uphoff, 2021).
In B. subtilis, several sigma factors have been reported to exhibit stochastic, pulsatile bursts of expression (Locke et al., 2011; Cabeen et al., 2017; Park et al., 2018), which can be similarly tracked in the MM. Sigma factors in B. subtilis promote the production of their own operons, which also encodes their anti-sigma factors, creating positive and negative feedback that can cause pulses of gene expression. Molecular “time-sharing” was proposed as a mechanism in which alternative sigma factors competing for RNA polymerase binding opportunities are able to share such core resources over time (Park et al., 2018). Stochastic bursts of sigma factor activity also seem to play a role in stress response in B. subtilis: heterogeneous response to lysozyme stress was observed between individual cells grown in the MM (Schwall et al., 2021). Pulsatile expression of the sigV operon preceding exposure to lysozyme stress led to increased survival probability.
Early work with B. subtilis in the MM revealed the existence of a cell fate switch controlling whether B. subtilis exists in a free-living, motile state or as a sessile member of a multicellular chain associated with biofilm formation (Norman et al., 2013). A modified device was used, with side-channels to ensure even nutrient availability in the long channels that accommodate the chain phenotype. By tracking the fates using fluorescent reporters over hundreds of cell generations, it was found that the transition from the motile to the sessile state happened at a constant rate over time (i.e., is a memoryless process), but the cells spent a precise amount of time in the sessile state. A simple network of three proteins could recapitulate all the properties and the modularity of the switch and the commitment to the chained state. Remarkably, the circuit was reconstituted in evolutionarily distant E. coli, showing that this simple network is sufficient to drive cell-fate decision making (Lord et al., 2019). The stochastic entry into another cell fate–sporulation–was also studied using the MM, and was shown to occur at a constant rate over time after adaptation to the sporulation-inducing conditions (Russell et al., 2017).
In another study, a clonal B. subtilis population diverged into subpopulations of distinct metabolic specialists, each characterized by differential expression of metabolic genes (Rosenthal et al., 2018). Cells with stochastically upregulated sucC expression in mid to late exponential phase were associated with the production of acetate, while a subpopulation expressing alsS in early stationary phase was linked to production of acetoin. Cells could be observed stochastically switching in and out of such states with fluorescent reporters in the MM. As acetoin can neutralize low pH conditions caused by acetate accumulation, the slow-growing alsS-expressing subpopulation enabled growth and expansion of an alsS- subpopulation which benefited from the neutralization of acetate in an agar pad microenvironment. This showcases how stochastic gene expression can help populations of genetically identical cells achieve cooperative behaviors. Therefore, bacteria have shown the ability to harness molecular fluctuations through simple circuits of a handful of proteins to establish heterogeneous phenotypic states that can be advantageous to the bulk isogenic population.
The rise of antibiotic resistance combined with the lack of new antibiotics is an alarming threat to public health (Aslam et al., 2018). Persister cells can survive antibiotic treatment by remaining in a temporary state of dormancy throughout antibiotic exposure, without being genetically resistant (Balaban et al., 2019). The switch to this state can happen spontaneously or be induced by stress such as starvation. Microfluidic devices are particularly well-suited for the study of this non-genetic heterogeneity since growth conditions can be precisely controlled and single lineages tracked over time. One of the first applications of a MM-like device was to establish the persister state as a phenotypic switch (Balaban et al., 2004). Persisters can be identified in such microfluidic devices as cells that are not growing prior to antibiotic exposure, but resume growth at some later time point following removal of the antibiotic. With an increase in throughput of the MM and microscopy (e.g., more trenches per chip and faster microscope imaging with larger field of view), it was possible to observe hundreds of E. coli persisters without mutations that increase their typically low frequency of approximately 1 in 1000 cells (Bakshi et al., 2021). The molecular mechanisms underlying this phenotypic switch are still under investigation (Goode et al., 2021b; Manuse et al., 2021). Studies using the MM enabled the characterization of these persisters and have shown that they have smaller size (Bakshi et al., 2021), lower ATP levels (Manuse et al., 2021), and are more likely to contain protein aggregates (Goode et al., 2021a). Recent developments in microscopy throughput, simulating batch culture conditions, and screening in the MM will likely help us to understand the molecular mechanisms behind bacterial persisters.
The MM has also been used to study more broadly the response of bacteria to antibiotics. The device with back-channels (Figure 3A) was used to rapidly load cells into the device for fast antibiotic susceptibility testing (∼30 min) of clinical samples (Baltekin et al., 2017), by directly visualizing growth or death of the bacteria during antibiotic exposure through microscopy. A study looked at the accumulation of the antibiotic ofloxacin inside E. coli in the MM and has shown that stationary phase cells appeared to absorb the antibiotic more slowly than exponentially growing cells (Cama et al., 2020).
It has become increasingly clear that cellular circuits must contend with stochastic gene expression and that this noise can have an important impact. Therefore, it is valuable to have the ability to quantify such variability for the engineering of cells for synthetic biology applications. Indeed, microfluidic devices have been instrumental in the development of synthetic gene circuits with dynamic properties (Cookson et al., 2005; Stricker et al., 2008; Danino et al., 2010; Din et al., 2016; Lezia et al., 2022). The use of the MM and insights from theory of stochastic gene expression enabled the re-engineering of the repressilator - the iconic synthetic oscillator that helped kick-start the field of synthetic biology—to achieve a precision that approaches natural oscillators (Potvin-Trottier et al., 2016; Luro et al., 2020). The MM was instrumental in enabling the precise characterization of the oscillators, identifying factors that disrupted oscillations, characterizing redesigned iterations of the circuit, and subsequent screening of pooled libraries. While a handful of studies have used the MM to evaluate (Niederholtmeyer et al., 2015; Zhang et al., 2022) or control (Lugagne et al., 2017) synthetic gene circuits, the broader use of the MM throughout the design process could lead to a new generation of precise and robust synthetic circuits.
Single-cell microfluidic platforms have facilitated important discoveries in a variety of fields in biology by enabling the quantification of dynamic and heterogenous processes. Recent technical developments of the MM, including the ability to phenotype or screen pooled libraries based on their dynamics, achieve better control over the growth conditions, and to culture additional organisms should continue to expand the applications of these devices to new fields. Further developments could facilitate the study of species-species interactions, as has been done in a few studies (Din et al., 2016; Cooper et al., 2017). The majority of studies using the MM have focused on E. coli and B. subtilis, and applications of the device to other organisms (e.g., microbes important in the clinic or in the gut microbiota) would broaden its scope. Outside the microbial realm, two studies so far have used the device with non-adherent mammalian cells (Pearl Mizrahi et al., 2016; Seita et al., 2021). More studies could shed light on dynamic and heterogeneous processes, such as phenotypic resistance to cancer treatment and the differentiation of hematopoietic stem cells (Huang, 2009; Jolly et al., 2018; Reyes and Lahav, 2018). Adaptation of the phenotyping and screening platforms to mammalian cells could provide an alternative to the single-cell screening techniques that have been developed (Stuart and Satija, 2019; Chandrasekaran et al., 2020).
Future studies using the MM will likely continue to advance studies of the heterogeneous processes discussed above. By showcasing discoveries in diverse fields, we hope to inspire the readers to implement such devices to explore questions in new fields and expand their possible applications. For example, the timing of other epigenetic processes in bacteria, such as prions (Yuan and Hochschild, 2017; Fleming et al., 2019), could be elucidated. While many studies have quantified the heterogeneity in protein production, few studies have examined how the degradation of proteins can create heterogeneity (Wong et al., 2007; Jones and Uphoff, 2021). Although most bacteria in nature are in stationary phase (Gefen et al., 2014), the majority of studies have focused on exponentially-growing bacteria. The technical developments of the MM enabling the observation of cells in different growth phases could fill this gap in the literature and generate insights into the stationary phase. Adopting the MM setup has become increasingly accessible, with molds available through different companies, detailed protocols published, and open-source data analysis software now available. Ultimately, because these microfluidic devices provide a new quantitative way to look at cells, they have the potential to continue to contribute to discoveries in diverse areas in biology.
All authors reviewed the articles, wrote the review, and approved it for publication.
This work was supported by a NSERC Discovery grant (RGPIN-2019-07002), a FRQNT Établissement de la relève professorale grant (2020-NC-268622), and a CFI John R. Evans Leader Fund (38290). PA and FP received fellowship support from the NSERC-CREATE (511601-2018)-supported SynBioApps program at Concordia University. FP also received support from an NSERC Canada Graduate Scholarship for the Master’s program, FRQNT (Fonds de recherche du Québec–Nature et technologies) doctoral scholarship, and a Miriam Aaron Roland graduate fellowship.
We would like to thank K. Jager for help with the figures, and G. McCallum, F. Perez, M. Orozco-Hidalgo and K. Jager for comments about the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Amir, A. (2014). Cell size regulation in bacteria. Phys. Rev. Lett. 112, 208102. doi:10.1103/physrevlett.112.208102
Angert, E. R. (2005). Alternatives to binary fission in bacteria. Nat. Rev. Microbiol. 3, 214–224. doi:10.1038/nrmicro1096
Arnaouteli, S., Bamford, N. C., Stanley-Wall, N. R., and Kovács, Á. T. (2021). Bacillus subtilis biofilm formation and social interactions. Nat. Rev. Microbiol. 19, 600–614. doi:10.1038/s41579-021-00540-9
Aslam, B., Wang, W., Arshad, M. I., Khurshid, M., Muzammil, S., Rasool, M. H., et al. (2018). Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 11, 1645–1658. doi:10.2147/idr.s173867
Bakshi, S., Leoncini, E., Baker, C., Cañas-Duarte, S. J., Okumus, B., and Paulsson, J. (2021). Tracking bacterial lineages in complex and dynamic environments with applications for growth control and persistence. Nat. Microbiol. 6, 783–791. doi:10.1038/s41564-021-00900-4
Balaban, N. Q., Helaine, S., Lewis, K., Ackermann, M., Aldridge, B., Andersson, D. I., et al. (2019). Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 17, 441–448. doi:10.1038/s41579-019-0196-3
Balaban, N. Q., Merrin, J., Chait, R., Kowalik, L., and Leibler, S. (2004). Bacterial persistence as a phenotypic switch. Science 305, 1622–1625. doi:10.1126/science.1099390
Balleza, E., Kim, J. M., and Cluzel, P. (2017). Systematic characterization of maturation time of fluorescent proteins in living cells. Nat. Methods 15, 47–51. doi:10.1038/nmeth.4509
Baltekin, Ö., Boucharin, A., Tano, E., Andersson, D. I., and Elf, J. (2017). Antibiotic susceptibility testing in less than 30 min using direct single-cell imaging. Proc. Natl. Acad. Sci. U. S. A. 114, 9170–9175. doi:10.1073/pnas.1708558114
Bennett, M. R., and Hasty, J. (2009). Microfluidic devices for measuring gene network dynamics in single cells. Nat. Rev. Genet. 10, 628–638. doi:10.1038/nrg2625
Bergmiller, T., Andersson, A. M. C., Tomasek, K., Balleza, E., Kiviet, D. J., Hauschild, R., et al. (2017). Biased partitioning of the multidrug efflux pump AcrAB-TolC underlies long-lived phenotypic heterogeneity. Science 356, 311–315. doi:10.1126/science.aaf4762
Cabeen, M. T., and Losick, R. (2018). Single-cell microfluidic analysis of Bacillus subtilis. J. Vis. Exp. doi:10.3791/56901
Cabeen, M. T., Russell, J. R., Paulsson, J., and Losick, R. (2017). Use of a microfluidic platform to uncover basic features of energy and environmental stress responses in individual cells of Bacillus subtilis. PLoS Genet. 13 (131), e1006901. doi:10.1371/journal.pgen.1006901
Cama, J., Voliotis, M., Metz, J., Smith, A., Iannucci, J., Keyser, U. F., et al. (2020). Single-cell microfluidics facilitates the rapid quantification of antibiotic accumulation in Gram-negative bacteria. Lab. Chip 20, 2765–2775. doi:10.1039/d0lc00242a
Campos, M., Surovtsev, I. V., Kato, S., Paintdakhi, A., Beltran, B., Ebmeier, S. E., et al. (2014). A constant size extension drives bacterial cell size homeostasis. Cell 159, 1433–1446. doi:10.1016/j.cell.2014.11.022
Camsund, D., Lawson, M. J., Larsson, J., Jones, D., Zikrin, S., Fange, D., et al. (2020). Time-resolved imaging-based CRISPRi screening. Nat. Methods 17, 86–92. doi:10.1038/s41592-019-0629-y
Chandrasekaran, S. N., Ceulemans, H., Boyd, J. D., and Carpenter, A. E. (2020). Image-based profiling for drug discovery: Due for a machine-learning upgrade? Nat. Rev. Drug Discov. 20, 145–159. doi:10.1038/s41573-020-00117-w
Cookson, S., Ostroff, N., Pang, W. L., Volfson, D., and Hasty, J. (2005). Monitoring dynamics of single-cell gene expression over multiple cell cycles. Mol. Syst. Biol. 1, 2005.0024. doi:10.1038/msb4100032.2005.0024
Cooper, R. M., Tsimring, L., and Hasty, J. (2017). Inter-species population dynamics enhance microbial horizontal gene transfer and spread of antibiotic resistance. Elife 6, 259500–e26026. doi:10.7554/elife.25950
Crane, M. M., Clark, I. B. N., Bakker, E., Smith, S., and Swain, P. S. (2014). A microfluidic system for studying ageing and dynamic single-cell responses in budding yeast. PLoS One 9, e100042. doi:10.1371/journal.pone.0100042
Danino, T., Mondragón-Palomino, O., Tsimring, L., and Hasty, J. (2010). A synchronized quorum of genetic clocks. Nature 463, 326–330. doi:10.1038/nature08753
Dar, D., Dar, N., Cai, L., and Newman, D. K. (2021). Spatial transcriptomics of planktonic and sessile bacterial populations at single-cell resolution. Science 373, eabi4882. doi:10.1126/science.abi4882
Darnell, C. L., Zheng, J., Wilson, S., Bertoli, R. M., Bisson-Filho, A. W., Garner, E. C., et al. (2020). The ribbon-helix-helix domain protein CdrS regulates the tubulin homolog ftsZ2 to control cell division in archaea. MBio 11, e01007–e01020. doi:10.1128/mbio.01007-20
Din, M. O., Danino, T., Prindle, A., Skalak, M., Selimkhanov, J., Allen, K., et al. (2016). Synchronized cycles of bacterial lysis for in vivo delivery. Nature 536, 81–85. doi:10.1038/nature18930
Durán, D. C., Hernández, C. A., Suesca, E., Acevedo, R., Acosta, I. M., Forero, D. A., et al. (2020). Slipstreaming mother machine: A microfluidic device for single-cell dynamic imaging of yeast. Micromachines (Basel) 12, 4. doi:10.3390/mi12010004
Eland, L. E., Wipat, A., Lee, S., Park, S., and Wu, L. J. (2016). “Chapter 3 - microfluidics for bacterial imaging,” in Methods in microbiology. Editors C. Harwood, and G. J. Jensen (Academic Press), 69–111.
Elowitz, M. B., Levine, A. J., Siggia, E. D., and Swain, P. S. (2002). Stochastic gene expression in a single cell. Science 297, 1183–1186. doi:10.1126/science.1070919
Facchetti, G., Chang, F., and Howard, M. (2017). Controlling cell size through sizer mechanisms. Curr. Opin. Syst. Biol. 5, 86–92. doi:10.1016/j.coisb.2017.08.010
Fleming, E., Yuan, A. H., Heller, D. M., and Hochschild, A. (2019). A bacteria-based genetic assay detects prion formation. Proc. Natl. Acad. Sci. U. S. A. 116, 4605–4610. doi:10.1073/pnas.1817711116
Gefen, O., Fridman, O., Ronin, I., and Balaban, N. Q. (2014). Direct observation of single stationary-phase bacteria reveals a surprisingly long period of constant protein production activity. Proc. Natl. Acad. Sci. U. S. A. 111, 556–561. doi:10.1073/pnas.1314114111
Goode, O., Smith, A., Łapińska, U., Bamford, R., Kahveci, Z., Glover, G., et al. (2021a). Heterologous protein expression favors the formation of protein aggregates in persister and viable but nonculturable bacteria. ACS Infect. Dis. 7, 1848–1858. doi:10.1021/acsinfecdis.1c00154
Goode, O., Smith, A., Zarkan, A., Cama, J., Invergo, B. M., Belgami, D., et al. (2021b). Persister Escherichia coli cells have a lower intracellular pH than susceptible cells but maintain their pH in response to antibiotic treatment. MBio 12, e0090921. doi:10.1128/mbio.00909-21
Gordo, I., Perfeito, L., and Sousa, A. (2011). Fitness effects of mutations in bacteria. Microb. Physiol. 21, 20–35. doi:10.1159/000332747
Govers, S. K., Mortier, J., Adam, A., and Aertsen, A. (2018). Protein aggregates encode epigenetic memory of stressful encounters in individual Escherichia coli cells. PLoS Biol. 16, e2003853. doi:10.1371/journal.pbio.2003853
Hardo, G., and Bakshi, S. (2021). Challenges of analysing stochastic gene expression in bacteria using single-cell time-lapse experiments. Essays Biochem. 65, 67–79. doi:10.1042/ebc20200015
Ho, P.-Y., and Amir, A. (2015). Simultaneous regulation of cell size and chromosome replication in bacteria. Front. Microbiol. 6, 662. doi:10.3389/fmicb.2015.00662
Hol, F. J. H., and Dekker, C. (2014). Zooming in to see the bigger picture: Microfluidic and nanofabrication tools to study bacteria. Science 346, 1251821. doi:10.1126/science.1251821
Huang, S. (2009). Non-genetic heterogeneity of cells in development: More than just noise. Development 136, 3853–3862. doi:10.1242/dev.035139
Jo, M. C., Liu, W., Gu, L., Dang, W., and Qin, L. (2015). High-throughput analysis of yeast replicative aging using a microfluidic system. Proc. Natl. Acad. Sci. U. S. A. 112, 9364–9369. doi:10.1073/pnas.1510328112
Jolly, M. K., Kulkarni, P., Weninger, K., Orban, J., and Levine, H. (2018). Phenotypic plasticity, bet-hedging, and androgen independence in prostate cancer: Role of non-genetic heterogeneity. Front. Oncol. 8, 50. doi:10.3389/fonc.2018.00050
Jones, E. C., and Uphoff, S. (2021). Single-molecule imaging of LexA degradation in Escherichia coli elucidates regulatory mechanisms and heterogeneity of the SOS response. Nat. Microbiol. 6, 981–990. doi:10.1038/s41564-021-00930-y
Julou, T., Zweifel, L., Blank, D., Fiori, A., and van Nimwegen, E. (2020). Subpopulations of sensorless bacteria drive fitness in fluctuating environments. PLoS Biol. 18, e3000952.
Kaiser, M., Jug, F., Julou, T., Deshpande, S., Pfohl, T., Silander, O. K., et al. (2018). Monitoring single-cell gene regulation under dynamically controllable conditions with integrated microfluidics and software. Nat. Commun. 9, 212. doi:10.1038/s41467-017-02505-0
Kamande, J. W., Wang, Y., and Taylor, A. M. (2015). Cloning SU8 silicon masters using epoxy resins to increase feature replicability and production for cell culture devices. Biomicrofluidics 9, 036502. doi:10.1063/1.4922962
Karaayvaz, M., Cristea, S., Gillespie, S. M., Patel, A. P., Mylvaganam, R., Luo, C. C., et al. (2018). Unravelling subclonal heterogeneity and aggressive disease states in TNBC through single-cell RNA-seq. Nat. Commun. 9, 3588. doi:10.1038/s41467-018-06052-0
Kim, J. M., Garcia-Alcala, M., Balleza, E., and Cluzel, P. (2020). Stochastic transcriptional pulses orchestrate flagellar biosynthesis in Escherichia coli. Sci. Adv. 6, eaax0947. doi:10.1126/sciadv.aax0947
Kinker, G. S., Greenwald, A. C., Tal, R., Orlova, Z., Cuoco, M. S., McFarland, J. M., et al. (2020). Pan-cancer single-cell RNA-seq identifies recurring programs of cellular heterogeneity. Nat. Genet. 52, 1208–1218. doi:10.1038/s41588-020-00726-6
Knorre, D. A., Azbarova, A. V., Galkina, K. V., Feniouk, B. A., and Severin, F. F. (2018). Replicative aging as a source of cell heterogeneity in budding yeast. Mech. Ageing Dev. 176, 24–31. doi:10.1016/j.mad.2018.09.001
Landgraf, D., Okumus, B., Chien, P., Baker, T. A., and Paulsson, J. (2012). Segregation of molecules at cell division reveals native protein localization. Nat. Methods 9, 480–482. doi:10.1038/nmeth.1955
Łapińska, U., Glover, G., Capilla-Lasheras, P., Young, A. J., and Pagliara, S. (2019). Bacterial ageing in the absence of external stressors. Phil. Trans. R. Soc. B 374, 20180442. doi:10.1098/rstb.2018.0442
Lawson, M. J., Camsund, D., Larsson, J., Baltekin, Ö., Fange, D., and Elf, J. (2017). In situ genotyping of a pooled strain library after characterizing complex phenotypes. Mol. Syst. Biol. 13, 947. doi:10.15252/msb.20177951
Le Treut, G., Si, F., Li, D., and Jun, S. (2021). Quantitative examination of five stochastic cell-cycle and cell-size control models for Escherichia coli and Bacillus subtilis. Front. Microbiol. 12, 721899. doi:10.3389/fmicb.2021.721899
Lee, S. S., Avalos Vizcarra, I., Huberts, D. H. E. W., Lee, L. P., and Heinemann, M. (2012). Whole lifespan microscopic observation of budding yeast aging through a microfluidic dissection platform. Proc. Natl. Acad. Sci. U. S. A. 109, 4916–4920. doi:10.1073/pnas.1113505109
Lezia, A., Csicsery, N., and Hasty, J. (2022). Design, mutate, screen: Multiplexed creation and arrayed screening of synchronized genetic clocks. Cell Syst. 13, 365–375.e5. e5. doi:10.1016/j.cels.2022.02.005
Li, Y., Jin, M., O’Laughlin, R., Bittihn, P., Tsimring, L. S., Pillus, L., et al. (2017). Multigenerational silencing dynamics control cell aging. Proc. Natl. Acad. Sci. U. S. A. 114, 11253–11258. doi:10.1073/pnas.1703379114
Lindner, A. B., Madden, R., Demarez, A., Stewart, E. J., and Taddei, F. (2008). Asymmetric segregation of protein aggregates is associated with cellular aging and rejuvenation. Proc. Natl. Acad. Sci. U. S. A. 105, 3076–3081. doi:10.1073/pnas.0708931105
Locke, J. C. W., Young, J. W., Fontes, M., Hernández Jiménez, M. J., and Elowitz, M. B. (2011). Stochastic pulse regulation in bacterial stress response. Science 334, 366–369. doi:10.1126/science.1208144
Lord, N. D., Norman, T. M., Yuan, R., Bakshi, S., Losick, R., and Paulsson, J. (2019). Stochastic antagonism between two proteins governs a bacterial cell fate switch. Science 366, 116–120. doi:10.1126/science.aaw4506
Lugagne, J.-B., Carillo, S. S., Kirch, M., Köhler, A., Batt, G., and Hersen, P. (2017). Balancing a genetic toggle switch by real-time feedback control and periodic forcing. Nat. Commun. 8, 1671–1677. doi:10.1038/s41467-017-01498-0
Lugagne, J.-B., Lin, H., and Dunlop, M. J. (2020). DeLTA: Automated cell segmentation, tracking, and lineage reconstruction using deep learning. PLoS Comput. Biol. 16, e1007673. doi:10.1371/journal.pcbi.1007673
Luro, S., Potvin-Trottier, L., Okumus, B., and Paulsson, J. (2020). Isolating live cells after high-throughput, long-term, time-lapse microscopy. Nat. Methods 17, 93–100. doi:10.1038/s41592-019-0620-7
Mäkelä, J., Uphoff, S., and Sherratt, D. J. (2021). Nonrandom segregation of sister chromosomes by Escherichia coli MukBEF. Proc. Natl. Acad. Sci. U. S. A. 118, e2022078118. doi:10.1073/pnas.2022078118
Manuse, S., Shan, Y., Canas-Duarte, S. J., Bakshi, S., Sun, W.-S., Mori, H., et al. (2021). Bacterial persisters are a stochastically formed subpopulation of low-energy cells. PLoS Biol. 19, e3001194. doi:10.1371/journal.pbio.3001194
Miano, A., Liao, M. J., and Hasty, J. (2020). Inducible cell-to-cell signaling for tunable dynamics in microbial communities. Nat. Commun. 11, 1193–1198. doi:10.1038/s41467-020-15056-8
Micali, G., Grilli, J., Osella, M., and Lagomarsino, M. C. (2018). Concurrent processes set E. coli cell division. Sci. Adv. 4, eaau3324. doi:10.1126/sciadv.aau3324
Moffitt, J. R., Lee, J. B., and Cluzel, P. (2012). The single-cell chemostat: An agarose-based, microfluidic device for high-throughput, single-cell studies of bacteria and bacterial communities. Lab. Chip 12, 1487–1494. doi:10.1039/c2lc00009a
Moger-Reischer, R. Z., and Lennon, J. T. (2019). Publisher correction: Microbial ageing and longevity. Nat. Rev. Microbiol. 17, 716. doi:10.1038/s41579-019-0279-1
Morawska, L. P., Hernandez-Valdes, J. A., and Kuipers, O. P. (2022). Diversity of bet-hedging strategies in microbial communities-Recent cases and insights. WIREs Mech. Dis. 14, e1544. doi:10.1002/wsbm.1544
Nakaoka, H., and Wakamoto, Y. (2017). Aging, mortality, and the fast growth trade-off of Schizosaccharomyces pombe. PLoS Biol. 15, e2001109. doi:10.1371/journal.pbio.2001109
Niederholtmeyer, H., Sun, Z. Z., Hori, Y., Yeung, E., Verpoorte, A., Murray, R. M., et al. (2015). Rapid cell-free forward engineering of novel genetic ring oscillators. Elife 4, 097711–e9818. doi:10.7554/elife.09771
Nieto, C., Arias-Castro, J., Sánchez, C., Vargas-García, C., and Pedraza, J. M. (2020). Unification of cell division control strategies through continuous rate models. Phys. Rev. E 101, 022401. doi:10.1103/physreve.101.022401
Norman, T. M., Lord, N. D., Paulsson, J., and Losick, R. (2013). Memory and modularity in cell-fate decision making. Nature 503, 481–486. doi:10.1038/nature12804
O’Connor, O. M., Alnahhas, R. N., Lugagne, J.-B., and Dunlop, M. J. (2022). DeLTA 2.0: A deep learning pipeline for quantifying single-cell spatial and temporal dynamics. PLoS Comput. Biol. 18, e1009797. doi:10.1371/journal.pcbi.1009797
Ollion, J., Elez, M., and Robert, L. (2019). High-throughput detection and tracking of cells and intracellular spots in mother machine experiments. Nat. Protoc. 14, 3144–3161. doi:10.1038/s41596-019-0216-9
Ollion, J., and Ollion, C. (2020). DistNet: Deep tracking by displacement regression: Application to bacteria growing in the mother machine. Med. Image Comput. Comput. Assist. Interv. - MICCAI 2020 12265, 215–225.
Paintdakhi, A., Parry, B., Campos, M., Irnov, I., Elf, J., Surovtsev, I., et al. (2016). Oufti: An integrated software package for high-accuracy, high-throughput quantitative microscopy analysis. Mol. Microbiol. 99, 767–777. doi:10.1111/mmi.13264
Park, J., Dies, M., Lin, Y., Hormoz, S., Smith-Unna, S. E., Quinodoz, S., et al. (2018). Molecular time sharing through dynamic pulsing in single cells. Cell Syst. 6, 216–229.e15. e15. doi:10.1016/j.cels.2018.01.011
Patange, O., Schwall, C., Jones, M., Villava, C., Griffith, D. A., Phillips, A., et al. (2018). Escherichia coli can survive stress by noisy growth modulation. Nat. Commun. 9, 5333. doi:10.1038/s41467-018-07702-z
Paulsson, J. (2005). Models of stochastic gene expression. Phys. Life Rev. 2, 157–175. doi:10.1016/j.plrev.2005.03.003
Pearl Mizrahi, S., Gefen, O., Simon, I., and Balaban, N. Q. (2016). Persistence to anti-cancer treatments in the stationary to proliferating transition. Cell Cycle 15, 3442–3453. doi:10.1080/15384101.2016.1248006
Peyrusson, F., Varet, H., Nguyen, T. K., Legendre, R., Sismeiro, O., Coppée, J.-Y., et al. (2020). Intracellular Staphylococcus aureus persisters upon antibiotic exposure. Nat. Commun. 11, 2200. doi:10.1038/s41467-020-15966-7
Potvin-Trottier, L., Lord, N. D., Vinnicombe, G., and Paulsson, J. (2016). Synchronous long-term oscillations in a synthetic gene circuit. Nature 538, 514–517. doi:10.1038/nature19841
Potvin-Trottier, L., Luro, S., and Paulsson, J. (2018). Microfluidics and single-cell microscopy to study stochastic processes in bacteria. Curr. Opin. Microbiol. 43, 186–192. doi:10.1016/j.mib.2017.12.004
Prindle, A., Samayoa, P., Razinkov, I., Danino, T., Tsimring, L. S., and Hasty, J. (2011). A sensing array of radically coupled genetic ‘biopixels. Nature 481, 39–44. doi:10.1038/nature10722
Proenca, A. M., Rang, C. U., Buetz, C., Shi, C., and Chao, L. (2018). Age structure landscapes emerge from the equilibrium between aging and rejuvenation in bacterial populations. Nat. Commun. 9, 3722. doi:10.1038/s41467-018-06154-9
Proenca, A. M., Rang, C. U., Qiu, A., Shi, C., and Chao, L. (2019). Cell aging preserves cellular immortality in the presence of lethal levels of damage. PLoS Biol. 17, e3000266. doi:10.1371/journal.pbio.3000266
Raj, A., and van Oudenaarden, A. (2008). Nature, nurture, or chance: Stochastic gene expression and its consequences. Cell 135, 216–226. doi:10.1016/j.cell.2008.09.050
Reyes, J., and Lahav, G. (2018). Leveraging and coping with uncertainty in the response of individual cells to therapy. Curr. Opin. Biotechnol. 51, 109–115. doi:10.1016/j.copbio.2017.12.007
Robert, L., Ollion, J., and Elez, M. (2019). Real-time visualization of mutations and their fitness effects in single bacteria. Nat. Protoc. 14, 3126–3143. doi:10.1038/s41596-019-0215-x
Robert, L., Ollion, J., Robert, J., Song, X., Matic, I., and Elez, M. (2018). Mutation dynamics and fitness effects followed in single cells. Science 359, 1283–1286. doi:10.1126/science.aan0797
Ronneberger, O., Fischer, P., and Brox, T. (2015). U-Net: Convolutional networks for biomedical image segmentation. Med. Image Comput. Computer-Assisted Intervention MICCAI 9351, 234–241.
Rosenthal, A. Z., Qi, Y., Hormoz, S., Park, J., Li, S. H.-J., and Elowitz, M. B. (2018). Metabolic interactions between dynamic bacterial subpopulations. Elife 7, e33099. doi:10.7554/elife.33099
Russell, J. R., Cabeen, M. T., Wiggins, P. A., Paulsson, J., and Losick, R. (2017). Noise in a phosphorelay drives stochastic entry into sporulation in Bacillus subtilis. EMBO J. 36, 2856–2869. doi:10.15252/embj.201796988
Ryley, J., and Pereira-Smith, O. M. (2006). Microfluidics device for single cell gene expression analysis in Saccharomyces cerevisiae. Yeast 23, 1065–1073. doi:10.1002/yea.1412
Sachs, C. C., Grünberger, A., Helfrich, S., Probst, C., Wiechert, W., Kohlheyer, D., et al. (2016). Image-based single cell profiling: High-throughput processing of mother machine experiments. PLoS One 11, e0163453. doi:10.1371/journal.pone.0163453
Sampaio, N. M. V., Blassick, C. M., Andreani, V., Lugagne, J.-B., and Dunlop, M. J. (2022). Dynamic gene expression and growth underlie cell-to-cell heterogeneity in Escherichia coli stress response. Proc. Natl. Acad. Sci. U. S. A. 119, e2115032119. doi:10.1073/pnas.2115032119
Sauls, J. T., Cox, S. E., Do, Q., Castillo, V., Ghulam-Jelani, Z., and Jun, S. (2019). Control of Bacillus subtilis replication initiation during physiological transitions and perturbations. MBio 10, e02205–e02219. doi:10.1128/mbio.02205-19
Scheler, O., Postek, W., and Garstecki, P. (2019). Recent developments of microfluidics as a tool for biotechnology and microbiology. Curr. Opin. Biotechnol. 55, 60–67. doi:10.1016/j.copbio.2018.08.004
Schwall, C. P., Loman, T. E., Martins, B. M. C., Cortijo, S., Villava, C., and Kusmartsev, V. (2021a). Tunable phenotypic variability through an autoregulatory alternative sigma factor circuit. Mol. Syst. Biol. 17, e9832.
Scott, S. R., and Hasty, J. (2016). Quorum sensing communication modules for microbial consortia. ACS Synth. Biol. 5, 969–977. doi:10.1021/acssynbio.5b00286
Seita, A., Nakaoka, H., Okura, R., and Wakamoto, Y. (2021). Intrinsic growth heterogeneity of mouse leukemia cells underlies differential susceptibility to a growth-inhibiting anticancer drug. PLoS One 16, e0236534. doi:10.1371/journal.pone.0236534
Si, F., Le Treut, G., Sauls, J. T., Vadia, S., Levin, P. A., and Jun, S. (2019). Mechanistic origin of cell-size control and homeostasis in bacteria. Curr. Biol. 29, 1760–1770.e7. e7. doi:10.1016/j.cub.2019.04.062
Smith, A., Metz, J., and Pagliara, S. (2019). MMHelper: An automated framework for the analysis of microscopy images acquired with the mother machine. Sci. Rep. 9, 10123. doi:10.1038/s41598-019-46567-0
Stricker, J., Cookson, S., Bennett, M. R., Mather, W. H., Tsimring, L. S., and Hasty, J. (2008). A fast, robust and tunable synthetic gene oscillator. Nature 456, 516–519. doi:10.1038/nature07389
Stuart, T., and Satija, R. (2019). Integrative single-cell analysis. Nat. Rev. Genet. 20, 257–272. doi:10.1038/s41576-019-0093-7
Taheri-Araghi, S., Bradde, S., Sauls, J. T., Hill, N. S., Levin, P. A., Paulsson, J., et al. (2015). Cell-size control and homeostasis in bacteria. Curr. Biol. 25, 385–391. doi:10.1016/j.cub.2014.12.009
Tomanek, I., Grah, R., Lagator, M., Andersson, A. M. C., Bollback, J. P., Tkačik, G., et al. (2020). Gene amplification as a form of population-level gene expression regulation. Nat. Ecol. Evol. 4, 612–625. doi:10.1038/s41559-020-1132-7
Ullman, G., Wallden, M., Marklund, E. G., Mahmutovic, A., Razinkov, I., and Elf, J. (2013). High-throughput gene expression analysis at the level of single proteins using a microfluidic turbidostat and automated cell tracking. Phil. Trans. R. Soc. B 368, 20120025. doi:10.1098/rstb.2012.0025
Uphoff, S., Lord, N. D., Okumus, B., Potvin-Trottier, L., Sherratt, D. J., and Paulsson, J. (2016). Stochastic activation of a DNA damage response causes cell-to-cell mutation rate variation. Science 351, 1094–1097. doi:10.1126/science.aac9786
Uphoff, S. (2018). Real-time dynamics of mutagenesis reveal the chronology of DNA repair and damage tolerance responses in single cells. Proc. Natl. Acad. Sci. U. S. A. 115, E6516–E6525. doi:10.1073/pnas.1801101115
Vasdekis, A. E., and Stephanopoulos, G. (2015). Review of methods to probe single cell metabolism and bioenergetics. Metab. Eng. 27, 115–135. doi:10.1016/j.ymben.2014.09.007
Veening, J.-W., Smits, W. K., and Kuipers, O. P. (2008). Bistability, epigenetics, and bet-hedging in bacteria. Annu. Rev. Microbiol. 62, 193–210. doi:10.1146/annurev.micro.62.081307.163002
Vincent, M. S., and Uphoff, S. (2021). Cellular heterogeneity in DNA alkylation repair increases population genetic plasticity. Nucleic Acids Res. 49, 12320–12331. doi:10.1093/nar/gkab1143
Voorn, W. J., and Koppes, L. J. (1998). Skew or third moment of bacterial generation times. Arch. Microbiol. 169, 43–51. doi:10.1007/s002030050539
Wallden, M., Fange, D., Lundius, E. G., Baltekin, Ö., and Elf, J. (2016). The synchronization of replication and division cycles in individual E. coli cells. Cell 166, 729–739. doi:10.1016/j.cell.2016.06.052
Wang, P., Robert, L., Pelletier, J., Dang, W. L., Taddei, F., Wright, A., et al. (2010). Robust growth of Escherichia coli. Curr. Biol. 20, 1099–1103. doi:10.1016/j.cub.2010.04.045
Weibel, D. B., Diluzio, W. R., and Whitesides, G. M. (2007). Microfabrication meets microbiology. Nat. Rev. Microbiol. 5, 209–218. doi:10.1038/nrmicro1616
Wessel, A. K., Hmelo, L., Parsek, M. R., and Whiteley, M. (2013). Going local: Technologies for exploring bacterial microenvironments. Nat. Rev. Microbiol. 11, 337–348. doi:10.1038/nrmicro3010
Wiktor, J., Gynnå, A. H., Leroy, P., Larsson, J., Coceano, G., Testa, I., et al. (2021). RecA finds homologous DNA by reduced dimensionality search. Nature 597, 426–429. doi:10.1038/s41586-021-03877-6
Witz, G., van Nimwegen, E., and Julou, T. (2019). Initiation of chromosome replication controls both division and replication cycles in E. coli through a double-adder mechanism. Elife 8, e48063. doi:10.7554/elife.48063
Wong, W. W., Tsai, T. Y., and Liao, J. C. (2007). Single-cell zeroth-order protein degradation enhances the robustness of synthetic oscillator. Mol. Syst. Biol. 3, 130. doi:10.1038/msb4100172
Xie, Z., Zhang, Y., Zou, K., Brandman, O., Luo, C., Ouyang, Q., et al. (2012). Molecular phenotyping of aging in single yeast cells using a novel microfluidic device. Aging Cell 11, 599–606. doi:10.1111/j.1474-9726.2012.00821.x
Yang, D., Jennings, A. D., Borrego, E., Retterer, S. T., and Männik, J. (2018). Analysis of factors limiting bacterial growth in PDMS mother machine devices. Front. Microbiol. 9, 871. doi:10.3389/fmicb.2018.00871
Yang, Y., Song, X., and Lindner, A. B. (2016). “Chapter 2 - time-lapse microscopy and image analysis of Escherichia coli cells in mother machines,” in Methods in microbiology. Editors C. Harwood, and G. J. Jensen (Academic Press), 49–68.
Young, J. W., Locke, J. C. W., Altinok, A., Rosenfeld, N., Bacarian, T., Swain, P. S., et al. (2011). Measuring single-cell gene expression dynamics in bacteria using fluorescence time-lapse microscopy. Nat. Protoc. 7, 80–88. doi:10.1038/nprot.2011.432
Yuan, A. H., and Hochschild, A. (2017). A bacterial global regulator forms a prion. Science 355, 198–201. doi:10.1126/science.aai7776
Zhang, F., Sun, Y., Zhang, Y., Shen, W., Wang, S., Ouyang, Q., et al. (2022). Independent control of amplitude and period in a synthetic oscillator circuit with modified repressilator. Commun. Biol. 5, 23. doi:10.1038/s42003-021-02987-1
Keywords: microfluidics, time-lapse microscopy, cell screening, single-cell analysis, phenotypic heterogeneity, cellular dynamics
Citation: Allard P, Papazotos F and Potvin-Trottier L (2022) Microfluidics for long-term single-cell time-lapse microscopy: Advances and applications. Front. Bioeng. Biotechnol. 10:968342. doi: 10.3389/fbioe.2022.968342
Received: 13 June 2022; Accepted: 21 September 2022;
Published: 12 October 2022.
Edited by:
Burak Okumus, XCellCure, United StatesReviewed by:
Sumitra Debina Mitra, Independent researcher, Amsterdam, NetherlandsCopyright © 2022 Allard, Papazotos and Potvin-Trottier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laurent Potvin-Trottier, bGF1cmVudC5wb3R2aW5AY29uY29yZGlhLmNh
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.