 Qiang Gao1†
Qiang Gao1† Hao Yang2†Chi Wang2†Xin-Ying Xie2Kai-Xuan Liu2Ying Lin2
Hao Yang2†Chi Wang2†Xin-Ying Xie2Kai-Xuan Liu2Ying Lin2 Shuang-Yan Han2
Shuang-Yan Han2 Mingjun Zhu2
Mingjun Zhu2 Markus Neureiter3*Yina Lin2*
Markus Neureiter3*Yina Lin2* Jian-Wen Ye2*
Jian-Wen Ye2*- 1Key Laboratory of Plateau Ecology and Agriculture, Qinghai University, Xining, QH, China
- 2School of Biology and Biological Engineering, South China University of Technology, Guangzhou, China
- 3Institute for Environmental Biotechnology, Department of Agrobiotechnology, University of Natural Resources and Life Sciences, Tulln, Austria
With the rapid development of synthetic biology, a variety of biopolymers can be obtained by recombinant microorganisms. Polyhydroxyalkanoates (PHA) is one of the most popular one with promising material properties, such as biodegradability and biocompatibility against the petrol-based plastics. This study reviews the recent studies focusing on the microbial synthesis of PHA, including chassis engineering, pathways engineering for various substrates utilization and PHA monomer synthesis, and PHA synthase modification. In particular, advances in metabolic engineering of dominant workhorses, for example Halomonas, Ralstonia eutropha, Escherichia coli and Pseudomonas, with outstanding PHA accumulation capability, were summarized and discussed, providing a full landscape of diverse PHA biosynthesis. Meanwhile, we also introduced the recent efforts focusing on structural analysis and mutagenesis of PHA synthase, which significantly determines the polymerization activity of varied monomer structures and PHA molecular weight. Besides, perspectives and solutions were thus proposed for achieving scale-up PHA of low cost with customized material property in the coming future.
Introduction
Polyhydroxyalkanoates (PHAs) is a series of polyesters synthesized by different microbes (Steinbüchel, 2001), which have been widely used as bio-plastics for replacing petrol-based plastic due to their outstanding biodegradability and biocompatibility. Accordingly, PHA can be divided into three categories (Sudesh et al., 2000) including short-, medium- and long- chain-length PHAs, namely SCL-, MCL- and LCL-PHA, respectively. Of which, the monomers of SCL-, MCL- and LCL-PHA generally contain 2–5, 6–14 and over 15 carbon atoms, respectively. Because of the competitive material properties, PHA has attracted growing attentions of commercial interests in different application areas, such as medical implant (Chen and Wu, 2005), cosmetic beads (Choi et al., 2020), packaging (Chen and Patel, 2012), agricultural film (Chen, 2009), textile (Chen, 2009), feeding additives (Chen, 2009) and so on. In the past decades, intensive efforts have been made to generate various PHA productions consisting of diverse polymerized units with different carbon-chain-length and structures by genetically modified bacterial (Chen and Jiang, 2017), such as Halomonas spp. (Tan et al., 2011; Fu X. Z. et al., 2014), Ralstonia eutropha (Antonio et al., 2000; Raberg et al., 2018; Xiong et al., 2018), Escherichia coli (Park et al., 2001; Linares-Pastén et al., 2015; Sudo et al., 2020), Pseudomonas spp (Chanasit et al., 2016; Liang et al., 2020; Li M. et al., 2021) and so on (Hyakutake et al., 2014; Tariq et al., 2015). Therefore, over 150 types of PHAs have been obtained including homopolymers (PHB, poly-3-hydroxybutyrate) (Tan et al., 2011), random- and/or block- copolymers such as poly(3-hydroxybutyrate-co-4-hydroxybutyrate) (P34HB), poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV) (Fu X. Z. et al., 2014), poly(3-hydroxybutyrate-co-3-hydroxyhexonate) (PHBHHx) (Park et al., 2001), etc. (Li M. et al., 2021). To date, many building blocks, including rational designed enzymes (Chek et al., 2019; Lim et al., 2021), fine-tuned metabolic pathways towards monomer synthesis (Pacholak et al., 2021) and genetically engineered chassis of predominant PHA accumulation performance (Liang et al., 2020; Ye and Chen, 2021), have been developed for sufficient PHA synthesis using a variate of substrates.
In particular, scale-up industrial production lines for various PHA manufacturing have been recently launched or established by several companies, for example, MedPHA (operating production line of 1,000 ton/year PHB and/or P34HB, China) (Obruča et al., 2022), PhaBuilder (10,000 ton/year, under construction, China) (Yang et al., 2010), Tianan (3,000 ton/year PHBV, China) (Modi et al., 2011), Tepha (P4HB for medical uses, United States) (Martin and Williams, 2003), Danimer Scientific (6,000 ton/year PHBHHx, United State) (Mehrpouya et al., 2021), Keneka (5,000 ton/year PHBHHx, Japan) (Tanaka et al., 2021). However, the production cost of PHA still challenges for wide range commercial uses. Therefore, many solutions have been proposed and developed to reduce the industrial cost of PHA, including high cell density fermentation based on optimized feeding solution (Silva et al., 2017), non-sterile open fermentation process based on recombinant halophiles (Tan et al., 2011), cell factory engineering for effective utilization of low-cost carbon sources (Murugan et al., 2017; Panich et al., 2021), carbon fixation engineering for the improved conversion rate from glucose to PHA (Salehizadeh et al., 2020), co-production of PHA and value-added chemicals (Lan et al., 2016; Li et al., 2016) and so on.
Therefore, this study summarized recent advances of various PHA production and industrial trends thereof. Additionally, major building blocks, including representative workhorses, metabolic pathways and critical enzymes, for PHA synthesis have been reviewed and discussed. This study provides an entire landscape of PHA productions powered by synthetic biology, as well as perspectives focusing on cost-effective PHA manufacturing in the coming future.
Workhorses for PHA production
Halomonas bluephagenesis TD01
Halomonas bluephagenesis TD01 (H. bluephagenesis), a natural PHB producer isolated from salt lake (Tan et al., 2011), has been recently developed as a versatile chassis for PHA productions and value added chemicals, which exemplifies a cost-effective biomanufacturing paradigm based on next generation industrial biotechnology (NGIB) enabling non-sterile open fermentation process under high salt and high pH condition (Ye and Chen, 2021). Currently, the genetically reprogrammed H. bluephagenesis can produce various PHA polymers, including PHB (Tan et al., 2011), PHBV (Fu X. Z. et al., 2014), P34HB (Chen et al., 2017) and PHBP (poly-3-hydroxybutyrate-co-3-hydroxypropionate) (Jiang et al., 2021) using glucose, starch, gluconate and structural related carbon sources for corresponding monomer synthesis whenever necessary, for example, 4HB from γ-butyrolactone (GBL), 3HP from 1,3-propanediol, 3HV from propionate, etc. Notably, pilot-scale production of PHB and P34HB have succeeded in a 5,000-L bioreactor, yielding up to 100 g/L dry cell mass (DCM) containing 60–70 wt% PHA content with over 30% cost reduction (Ye et al., 2018). Besides, engineering electron transport system could significantly improve the supplementation of NADH (Ling et al., 2018), overexpression of Vitreoscilla hemoglobin (VHb) protein led to improved oxygen uptake efficiency (Ouyang et al., 2018), deficiency of outer membrane synthesis enabled sufficient production yield of PHA from glucose and simplified cell lysis (Wang Z. et al., 2021), manipulation of cell morphology also resulted in self-flocculation separation process (Ling et al., 2019). Moreover, different genetic parts and tools have been established allowing for rational engineering of H. bluephagenesis (Zhang et al., 2020). These efforts have proved successful in building a high-performing workhorse for PHA production based on NGIB. Additionally, many other Halomonas strains were also successfully developed for PHA synthesis, such as Halomonas campanesis LS21 (Yue et al., 2014), Halomonas elongate DSM2581 (Ilham et al., 2014), Halomonas pacifica ASL10 (Abd El-malek et al., 2020) and so on, illustrating the great potential of halophiles used as PHA producers.
Ralstonia eutropha
Ralstonia eutropha H16 (Cupriavidus necator) is a well-studied PHA producer from glucose, glycerol, palm oil and other fatty acids (FAs) (Murugan et al., 2017). In addition to short chain length PHA synthesis, R. eutropha H16 has been engineered to produce varied copolymers consisting of SCL-monomer (3HB) and MCL-monomers, such as 3HHx, 3HO (3-hydroxyoctanoate), 3HDD (3-hydroxydodecanoate) and so on (Antonio et al., 2000). In previous studies, genetic editing tools for chromosomal engineering was established based on CRISPR/Cas9 system and Cre/LoxP integrase system (Park et al., 2001). An electroporation approach was developed in recombinant R. eutropha H16 allowing for sufficient and high-through clone construction (Xiong et al., 2018). More importantly, over 200 g/L DCM with over 70 wt% PHA accumulation can be obtained by R. eutropha H16 and its derivates during fed-batch fermentation conducted in the lab- (<10-L) and/or pilot- (>100-L) scale bioreactors under strictly sterilized conditions (Ryu et al., 1997). Moreover, industrial productions of PHB, PHBV and PHBHHx based on recombinant R. eutropha H6 have been achieved by several companies. Therefore, R. eutropha H16 is expected to be a prominent chassis for PHA productions, especially for PHBHHx, however, high production cost remains challenging (Raberg et al., 2018).
Escherichia coli
Escherichia coli (E. coli), such BL21, JM109, etc., are well-studied model chassis that have clear genetic background and effective genetic tools for cell factory engineering of varied purposes, such as PHA biosynthesis. Even though E. coli is not a natural PHA producer, the heterogonous expression of phaCAB gene cluster from R. eutropha could efficiently boost carbon flux from pyruvate towards PHB synthesis. Therefore, intensive studies focusing on CO2 fixation (Lee et al., 2021), pathway engineering (Chen and Jiang, 2017) and feeding solution design of fed-batch fermentation (Yang et al., 2014) have been performed to generate enhanced production yield of PHB. Besides, E. coli is an ideal workhorse for studying the novel-type PHA synthesis, such as copolymers of 3HB and lactate, glycolic acid, 4-hydroxybutyrate, 5-hydroxyvalerate and other monomers with functional groups (Scheel et al., 2021). Specifically, the DCM and PHA content reached up to 194 g/L and 73 wt% by recombinant E. coli grown in fed-batch fermentation condition (JONG-IL CHOI, 1998), which shows promising performance in PHA accumulation.
Pseudomonas
Pseudomonas, including P. putida KT2440, P. entomophila, etc. have been recently engineered to be dominant producers of PHA copolymers consist of 3HB and MCL- and LCL-3HAs due to their strong FAs metabolism involved in β-oxidation cycle and de novo FAs synthesis pathways. Currently, PHA copolymers are composed of 3HB, 4HB, 3HV, 3HHx, 3HHp (3-hydroxyheptanoate), 3HO (3-hydroxyoctanoate), 3HD (3-hydroxydecanoate), etc. could be obtained by metabolically engineered Pseudomonas strains (Prieto et al., 2016). Many PHA synthases able to polymerize MCL- and LCL-3HA into polymers were thus identified from different Pseudomonas strains (Chung et al., 2011; Li et al., 2019; Tan et al., 2020; Li M. et al., 2021). Notably, an effective platform was developed for producing full spectrum of PHAs, which contain SCl-, MCL, LCL- 3HAs and monomers carrying carbon-carbon double bones, with over 90% increase in production yield based on recombinant P. entomophila (Li M. et al., 2021). Moreover, higher DCM, reaching over 70 g/L, was also achieved by Pseudomonas leveraging fed-batch fermentation process optimization (Cerrone et al., 2014). These efforts demonstrate proven success in scalable tailor-made PHA synthesis of varied functions by reprogrammed Pseudomonas.
Additionally, various attempts have been carried out to achieve PHA synthesis based on different hosts, such as Alcaligenes (H W Ryu 1996), Bacillus (Sathiyanarayanan et al., 2013), Burkholderia (Miranda De Sousa Dias et al., 2017), microalgae (Costa et al., 2019), Salinivibrio (Van Thuoc et al., 2019; Van Thuoc et al., 2020), Marinobacterium (Wang et al., 2022), Vibrio alginolyticus (Li H. F. et al., 2021) and so on, using diverse carbon sources including sucrose, propionate, carbon dioxide, volatile fatty acids, etc. It is important to note that the highest resultant DCM reached up to 281 g/L with 232 g/L PHB accumulation by Alcaligenes eutrophus, a natural PHB producer of high cell density growth and effective PHA accumulation, during a 74 h fed-batch fermentation conducted in a 60-L bioreactor (H W Ryu 1996).
Metabolic pathways for PHA synthesis
The biosynthesis pathways of most PHA monomers from varied carbon sources like glucose, fatty acids, etc. are mainly related to essential carbon metabolic pathways, such as glycolysis, β-oxidation and de novo fatty acid synthesis (Figure 1). Besides, using structurally related carbon sources as precursors is an alternative strategy to generate diverse PHA copolymers consist of different monomers, including 4HB from γ-butyrolactone (GBL)/1,4-butanediol (BDO), 3HV from propionate, 3HP from 1,3-propionediol (PDO), middle- and long-chain length 3HA from different fatty acids with corresponding carbon atoms and so on, which have significant impact on the material property (Chen et al., 2016). Therefore, a wide variety of PHA homo- and co-polymers can be obtained by engineered microbes by feeding customized feedstocks (Figure 1).
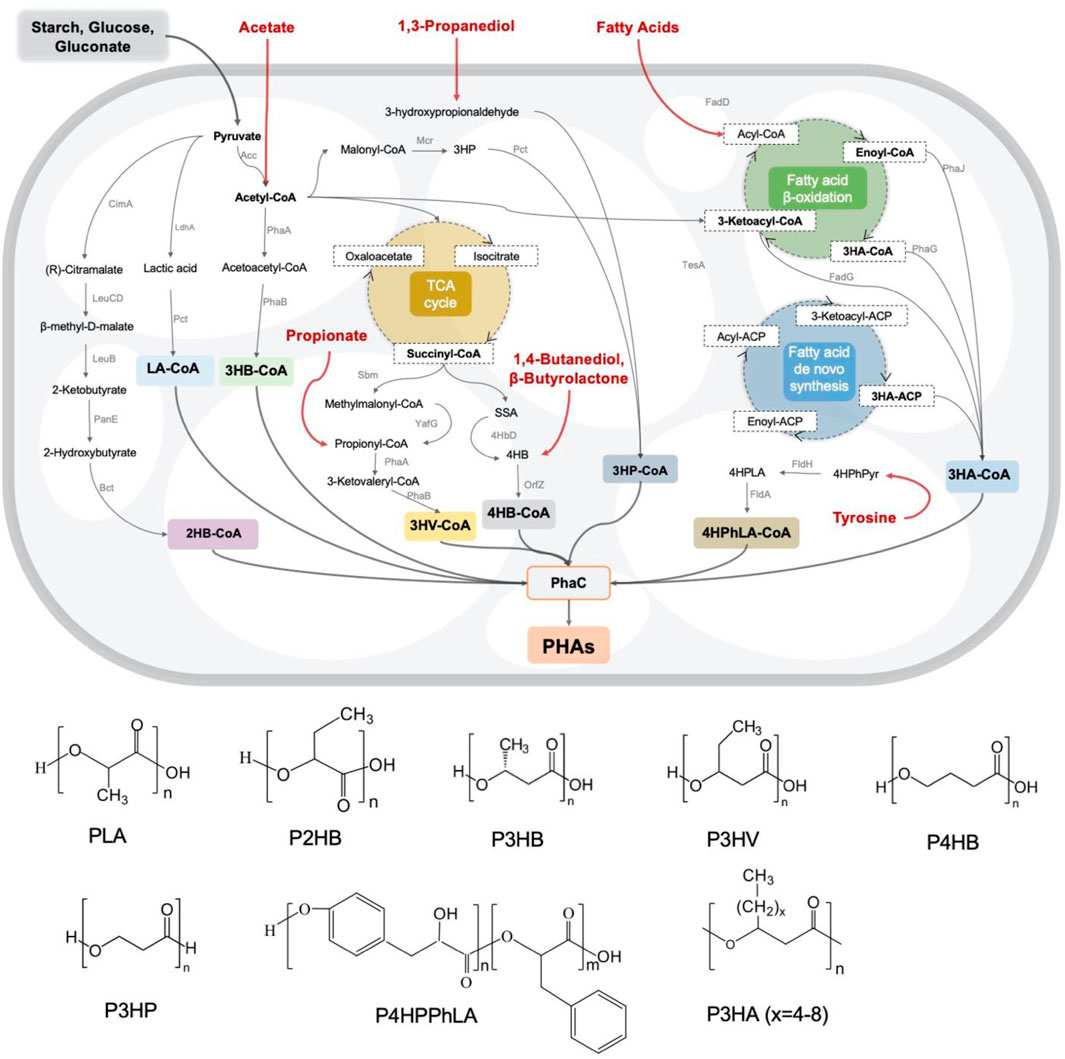
FIGURE 1. Metabolic pathways and monomer structures of different microbial synthesized PHAs.
PHA from structure-unrelated carbon sources
Glucose is a widely used feedstock in biomanufacturing. Similarly, intensive studies have been carried out for generating different PHA using glucose as the sole carbon source by metabolically engineered microorganisms. To date, many metabolic pathways have been mined and refined as significant building blocks for rewiring glucose-derived fluxes towards various monomers, such as converting pyruvate into2HB-CoA (Park et al., 2012c) and LA-CoA (Park et al., 2012b), acetyl-CoA into 3HB-CoA and 3HP-CoA (Meng et al., 2015), succinyl-CoA into 3HV-CoA (Bhatia et al., 2015) and 4HB-CoA (Lv et al., 2015), respectively. Interestingly, starch (Yang et al., 2020), volatile fatty acids like acetate (Yang et al., 2019), waste gluconate (Ciesielski et al., 2010), the byproduct of glucose processing, were also used to culture engineered Halomonas and Pseudomonas to achieve cost-effective PHA productions (Figure 1).
In addition to glucose, building blocks for many other carbon sources metabolism, such as glycerol, sucrose, xylose, C1 compounds, etc., have been constructed to synthesize PHA. Specifically, the highest PHA accumulation, reaching 38.9 wt% with 0.34 g/L/h of productivity have been achieved by engineered P. putida KT2440 (Borrero-de Acuna et al., 2021). Fu et al. produced MCL-PHA also could be obtained by grown on chemical-grade glycerol (PG) and biodiesel-derived waste glycerol (WG) as sole carbon sources (Fu J. et al., 2014). Moreover, recombinant strains including P. putida S12 and R. eutropha harboring expression vessel containing isomerase (XylA) and xylulokinase (XylB) have been constructed by Meijnen et al. (2008) and Kim et al. (2017), respectively, to produce PHA using xylose as sole carbon source (Meijnen et al., 2008). Similarly, a sucrose-favored P. putida strain was also developed for PHA synthesis from sucrose only (Hobmeier et al., 2020). More importantly, due to the growing interests of global carbon neutral, many bacterial like P. furiosus and R. eutropha B8562 were engineered to produce PHA polymers containing 3HP and 3HB units, respectively, using CO2 as carbon source (Volova et al., 2006; Keller et al., 2013). Besides, biosynthesis pathways for PHA synthesis from CH4 were also established based on many hydrogen-oxidizing bacteria (Khosravi-Darani et al., 2013). In summary, metabolic engineering of microbes is able to achieve targeted PHA synthesis from different structure-unrelated carbon sources.
PHA from structure-related carbon sources
For most MCL- and LCL-PHA synthesis, supplementation of structure-related fatty acids in the medium is a commonly used strategy to grow recombinant cells with defected β-oxidation cycle or reprogramed de novo fatty acids synthesis pathways (Gutierrez-Gomez et al., 2019). For instance, a wide range of PHA copolymers composed of 3HB and MCL-/LCL-3HA units containing carbon atoms numbered from 6 to 18, even with carbon-carbon double bone, have been achieved by engineered Pseudomonas, yielding over 100% increase of production titer (Yao J, 1999). Besides, many short chain length (SCL) PHA units were also produced from structure-related carbon sources used as precursors, such as 4HB synthesis from 1,4-propanediol (PDO) and β-butyrolactone (GBL) (Cavalheiro et al., 2012), 3HP synthesis from 1,3-propionediol (PDO) (Zhou et al., 2011), 5HV synthesis from 1,5-pentanediol (Yan et al., 2022), as well as functional group monomer like 4HPhLA synthesized from tyrosine (Yang et al., 2018), etc. Recently, high production yield of P34HB with 4HB molar ratio from 5 mol% to 26 mol% has been achieved by recombinant H. bluephagenesis based on NGIB platform, which also demonstrated the success in scale-up production of low cost conducted in 5-to-200 m³ fermenters (Ling et al., 2018). Notably, Lee et al. (2021) used engineered Escherichia coli to synthesize aromatic polyester, P(3HB-co-D-phenylacetate), from tyrosine, of which the molar ratio of D-phenylacetate monomer reaches up to 47.7 mol% (Yang et al., 2018). Moreover, tailor-made copolymers, as well as block copolymers, consisting of two, three and even more units could be easily obtained by designing the supplementation formula of target precursors and feeding strategy thereof (Yu et al., 2020).
Engineering tools and strategies for sufficient PHA synthesis
In addition to the biosynthesis pathway construction for diverse PHA productions, many metabolic engineering tools including high resolution gene expression tuning (Ye et al., 2020), high throughput library construction (Zhou et al., 2015; Young et al., 2018), constitutive and inducible promoter design (Shen et al., 2018; Ma et al., 2020) and so on have been developed for constructing effective PHA producing strains. Moreover, a carbon fixation of CO2 was established in E. coli to generate an increased bioconversion rate of glucose towards PHB (Lin et al., 2015). Modulating the NADH levels and its regeneration pathways could also show proven effects on PHA accumulation in both E. coli and Halomonas strains with PHA content increased up to 90 wt% (Ling et al., 2018). Interestingly, cell morphology control is an efficient strategy to obtain enhanced PHA accumulation with significantly improved substrate conversion rate (Wang X. et al., 2021). Manipulation of PHA granule size also demonstrated strong significance for downstream processing, which dramatically reduce the energy consumption of cell separation and PHA purification (Kourmentza et al., 2017). Therefore, the downstream-inspired engineering of microbes also displays great significance in cost-reduction for industrial PHA biomanufacturing.
PHA synthase
PHA synthase (PhaC) is an important building block for PHA synthesis. Generally, there are four major types of PhaC, namely Class-I/II/III/IV (Możejko-Ciesielska and Kiewisz, 2016), which have been identified from different PHA producing strains (Table 1). Of which, Class-I, -III and IV generally show higher activity on short-chain-length (SCL) monomers (C3-C5) polymerization, while Class-II has higher specificity to medium- and long- chain-length (M/LCL) monomers containing 6–18 carbon atoms, namely C6-C18 (Chek et al., 2017). Specifically, most Class-I PHA synthases, such as PhaCs from R. eutropha (Ushimaru et al., 2014), Alcaligenes latus (Park et al., 2012a), Aeromonas Caviae and Chromobacterium sp. (Choi et al., 2020), not only show effective activity on SCL PHA accumulation including 3HB, 3HP, 4HB and 3HV units, but also display polymerization capability of MCL PHA like 3HHx (Antonio et al., 2000).
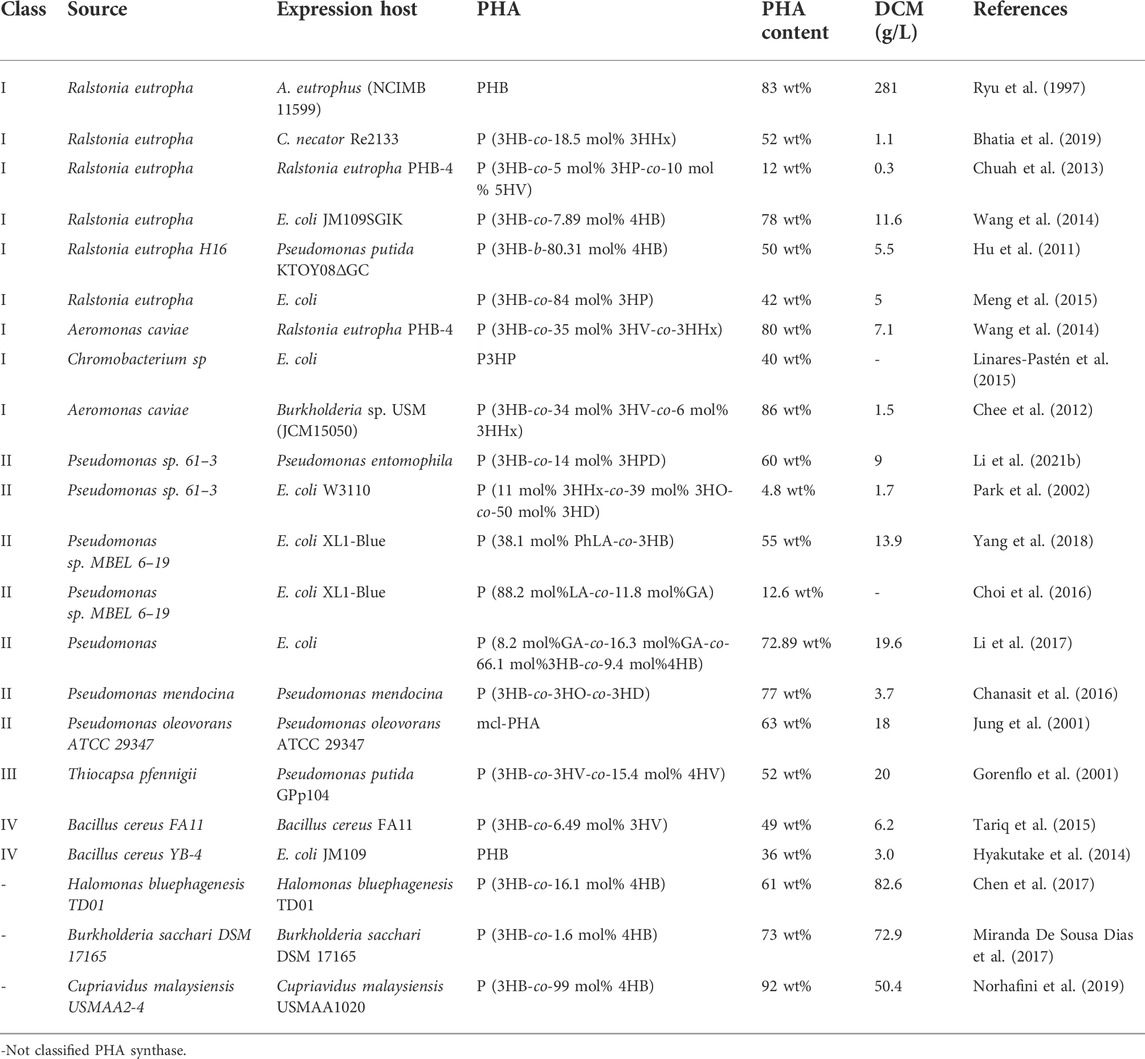
TABLE 1. Different PHA synthases identified from natural PHA producing strains.
Recently, many efforts have been made to modified the polymerization activity of PHA synthase, including protein structure analysis (Chek et al., 2017), mutagenesis (Zou et al., 2017) and fusion of functional domains from different PhaCs (Matsumoto et al., 2009), to generate high-performing PHA synthase. For instance, Kim et al. (2017) report the first crystal structure of Ralstonia eutropha PHA synthase at 1.8 Å resolution and structure-based mechanisms for PHA polymerization, RePhaC1 contains two distinct domains, the N-terminal (RePhaC1ND) and C-terminal domains (RePhaC1CD), and exists as a dimer (Kim et al., 2017). Furthermore, site-directed mutation was employed to generate PhaC mutants, namely PhaC61-3 and PhaC1437, based on PHA synthases from Pseudomonas sp. 61–3 and Pseudomonas sp. MBEL 6–19, respectively, which show wide substrate specificity to both SCL and M/LCL monomers (Yang et al., 2010), as well as monomers with a particular structure like benzene ring (Mizuno et al., 2018). Moreover, an artificial PHA synthase, PhaCAR, was constructed by hybridizing the C-terminal of PhaCAC from Aermonas Caviae and N-terminal of PhaCRE from R. eutropha (Cupriavidus necator), enabling effective accumulation for block copolymers containing 2-hydroxybutyrate (2HB) (Sudo et al., 2020);
Conclusion and perspective
In this study, we highlighted the global trends of industrial PHA productions reported by different companies and start-up teams, and briefly summarized and discussed the advances of different building blocks focusing on PHA synthase, biosynthesis pathways of SCL-, MCL- and LCL-PHA, dominant PHA workhorses of industrial potential and optimization strategies for effective PHA synthesis. This study provides an overview of PHA biosynthesis from enzyme engineering, cell factory design, towards scale-up bio-manufacturing. However, more attempts are still required to achieve further cost-reduction and improved material properties of tailor-made PHAs against the petrol-based plastics.
Author contributions
QG, HY, and CW contributed equally in this study. X-YX, K-XL, YgL, S-YH, MN, YaL, and J-WY wrote the manuscript, J-WY, YaL, and MN proposed the idea and revised the manuscript.
Funding
This research is supported by National Natural Science Foundation of China (Grant No. 32001029), National Natural Science Foundation of Qinghai Province (Grant No. 2020-ZJ-759), Guangdong Basic and Applied Basic Research Foundation (Grant No. 2020A1515111079), State Key Laboratory of Plateau Ecology and Agriculture, Qinghai University (Grant No. 2021-KF-09), and Tsinghua University-INDITEX Sustainable Development Fund (Grant No. TISD201907).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abd El-Malek, F., Farag, A., Omar, S., and Khairy, H. (2020). Polyhydroxyalkanoates (PHA) from Halomonas pacifica ASL10 and Halomonas salifodiane ASL11 isolated from Mariout salt lakes. Int. J. Biol. Macromol. 161, 1318–1328. doi:10.1016/j.ijbiomac.2020.07.258
Antonio, R. V., Steinbüchel, A., and Rehm, B. H. A. (2000). Analysis of in vivo substrate specificity of the PHA synthase from Ralstonia eutropha: Formation of novel copolyesters in recombinant Escherichia coli. FEMS Microbiol. Lett. 182, 111–117. doi:10.1111/j.1574-6968.2000.tb08883.x
Bhatia, S. K., Gurav, R., Choi, T.-R., Jung, H.-R., Yang, S.-Y., Song, H.-S., et al. (2019). Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) production from engineered Ralstonia eutropha using synthetic and anaerobically digested food waste derived volatile fatty acids. Int. J. Biol. Macromol. 133, 1–10. doi:10.1016/j.ijbiomac.2019.04.083
Bhatia, S. K., Yi, D. H., Kim, H. J., Jeon, J. M., Kim, Y. H., Sathiyanarayanan, G., et al. (2015). Overexpression of succinyl-CoA synthase for poly (3-hydroxybutyrate-co-3-hydroxyvalerate) production in engineered Escherichia coli BL21(DE3). J. Appl. Microbiol. 119, 724–735. doi:10.1111/jam.12880
Borrero-De Acuna, J. M., Rohde, M., Saldias, C., and Poblete-Castro, I. (2021). Fed-batch mcl- polyhydroxyalkanoates production in Pseudomonas putida KT2440 and DeltaphaZ mutant on biodiesel-derived crude glycerol. Fron t Bioeng. Biotechnol. 9, 642023. doi:10.3389/fbioe.2021.642023
Cavalheiro, J. M., De Almeida, M. C., Da Fonseca, M. M., and De Carvalho, C. C. (2012). Adaptation of Cupriavidus necator to conditions favoring polyhydroxyalkanoate production. J. Biotechnol. 164, 309–317. doi:10.1016/j.jbiotec.2013.01.009
Cerrone, F., Duane, G., Casey, E., Davis, R., Belton, I., Kenny, S. T., et al. (2014). Fed-batch strategies using butyrate for high cell density cultivation of Pseudomonas putida and its use as a biocatalyst. Appl. Microbiol. Biotechnol. 98, 9217–9228. doi:10.1007/s00253-014-5989-8
Chanasit, W., Hodgson, B., Sudesh, K., and Umsakul, K. (2016). Efficient production of polyhydroxyalkanoates (PHAs) from Pseudomonas mendocina PSU using a biodiesel liquid waste (BLW) as the sole carbon source. Biosci. Biotechnol. Biochem. 80, 1440–1450. doi:10.1080/09168451.2016.1158628
Chee, J.-Y., Lau, N.-S., Samian, M.-R., Tsuge, T., and Sudesh, K. (2012). Expression of Aeromonas caviae polyhydroxyalkanoate synthase gene in Burkholderia sp. USM (JCM15050) enables the biosynthesis of SCL-MCL PHA from palm oil products. J. Appl. Microbiol. 112, 45–54. doi:10.1111/j.1365-2672.2011.05189.x
Chek, M. F., Hiroe, A., Hakoshima, T., Sudesh, K., and Taguchi, S. (2019). PHA synthase (PhaC): Interpreting the functions of bioplastic-producing enzyme from a structural perspective. Appl. Microbiol. Biotechnol. 103, 1131–1141. doi:10.1007/s00253-018-9538-8
Chek, M. F., Kim, S. Y., Mori, T., Arsad, H., Samian, M. R., Sudesh, K., et al. (2017). Structure of polyhydroxyalkanoate (PHA) synthase PhaC from Chromobacterium sp. USM2, producing biodegradable plastics. Sci. Rep. 7, 5312. doi:10.1038/s41598-017-05509-4
Chen, G.-Q., and Patel, M. K. (2012). Plastics derived from biological sources: Present and future: A technical and environmental review. Chem. Rev. 112, 2082–2099. doi:10.1021/cr200162d
Chen, G.-Q., and Wu, Q. (2005). The application of polyhydroxyalkanoates as tissue engineering materials. Biomaterials 26, 6565–6578. doi:10.1016/j.biomaterials.2005.04.036
Chen, G. Q. (2009). A microbial polyhydroxyalkanoates (PHA) based bio- and materials industry. Chem. Soc. Rev. 38, 2434. doi:10.1039/b812677c
Chen, G. Q., and Jiang, X. R. (2017). Engineering bacteria for enhanced polyhydroxyalkanoates (PHA) biosynthesis. Synth. Syst. Biotechnol. 2, 192–197. doi:10.1016/j.synbio.2017.09.001
Chen, G. Q., Jiang, X. R., and Guo, Y. (2016). Synthetic biology of microbes synthesizing polyhydroxyalkanoates (PHA). Synth. Syst. Biotechnol. 1, 236–242. doi:10.1016/j.synbio.2016.09.006
Chen, X., Yin, J., Ye, J., Zhang, H., Che, X., Ma, Y., et al. (2017). Engineering Halomonas bluephagenesis TD01 for non-sterile production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate). Bioresour. Technol. 244, 534–541. doi:10.1016/j.biortech.2017.07.149
Choi, S. Y., Cho, I. J., Lee, Y., Kim, Y.-J., Kim, K.-J., Lee, S. Y., et al. (2020). Bacterial polyesters: Microbial polyhydroxyalkanoates and nonnatural polyesters (adv. Mater. 35/2020). Adv. Mat. 32, 2070264. doi:10.1002/adma.202070264
Choi, S. Y., Park, S. J., Kim, W. J., Yang, J. E., Lee, H., Shin, J., et al. (2016). One-step fermentative production of poly(lactate-co-glycolate) from carbohydrates in Escherichia coli. Nat. Biotechnol. 34, 435–440. doi:10.1038/nbt.3485
Chuah, J.-A., Yamada, M., Taguchi, S., Sudesh, K., Doi, Y., Numata, K., et al. (2013). Biosynthesis and characterization of polyhydroxyalkanoate containing 5-hydroxyvalerate units: Effects of 5HV units on biodegradability, cytotoxicity, mechanical and thermal properties. Polym. Degrad. Stab. 98, 331–338. doi:10.1016/j.polymdegradstab.2012.09.008
Chung, A. L., Jin, H. L., Huang, L. J., Ye, H. M., Chen, J. C., Wu, Q., et al. (2011). Biosynthesis and characterization of poly(3-hydroxydodecanoate) by beta-oxidation inhibited mutant of Pseudomonas entomophila L48. Biomacromolecules 12, 3559–3566. doi:10.1021/bm200770m
Ciesielski, S., Mozejko, J., and Przybylek, G. (2010). The influence of nitrogen limitation on mcl-PHA synthesis by two newly isolated strains of Pseudomonas sp. J. Ind. Microbiol. Biotechnol. 37, 511–520. doi:10.1007/s10295-010-0698-5
Costa, S. S., Miranda, A. L., De Morais, M. G., Costa, J. a. V., and Druzian, J. I. (2019). Microalgae as source of polyhydroxyalkanoates (PHAs) - a review. Int. J. Biol. Macromol. 131, 536–547. doi:10.1016/j.ijbiomac.2019.03.099
Fu, J., Sharma, U., Sparling, R., Cicek, N., and Levin, D. B. (2014a). Evaluation of medium-chain-length polyhydroxyalkanoate production by Pseudomonas putida LS46 using biodiesel by-product streams. Can. J. Microbiol. 60, 461–468. doi:10.1139/cjm-2014-0108
Fu, X. Z., Tan, D., Aibaidula, G., Wu, Q., Chen, J. C., Chen, G. Q., et al. (2014b). Development of Halomonas TD01 as a host for open production of chemicals. Metab. Eng. 23, 78–91. doi:10.1016/j.ymben.2014.02.006
Gorenflo, V., Schmack, G., Vogel, R., and Steinbüchel, A. (2001). Development of a process for the biotechnological large-scale production of 4-hydroxyvalerate-containing polyesters and characterization of their physical and mechanical properties. Biomacromolecules 2, 45–57. doi:10.1021/bm0000992
Gutierrez-Gomez, U., Servin-Gonzalez, L., and Soberon-Chavez, G. (2019). Role of beta-oxidation and de novo fatty acid synthesis in the production of rhamnolipids and polyhydroxyalkanoates by Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 103, 3753–3760. doi:10.1007/s00253-019-09734-x
Hobmeier, K., Lowe, H., Liefeldt, S., Kremling, A., and Pfluger-Grau, K. (2020). A nitrate-blind P. Putida strain boosts PHA production in a synthetic mixed culture. Front. Bioeng. Biotechnol. 8, 486. doi:10.3389/fbioe.2020.00486
Hu, D., Chung, A.-L., Wu, L.-P., Zhang, X., Wu, Q., Chen, J.-C., et al. (2011). Biosynthesis and characterization of polyhydroxyalkanoate block copolymer P3HB-b-P4HB. Biomacromolecules 12, 3166–3173. doi:10.1021/bm200660k
Hyakutake, M., Tomizawa, S., Mizuno, K., Abe, H., and Tsuge, T. (2014). Alcoholytic cleavage of polyhydroxyalkanoate chains by class IV synthases induced by endogenous and exogenous ethanol. Appl. Environ. Microbiol. 80, 1421–1429. doi:10.1128/aem.03576-13
Ilham, M., Nakanomori, S., Kihara, T., Hokamura, A., Matsusaki, H., Tsuge, T., et al. (2014). Characterization of polyhydroxyalkanoate synthases from Halomonas sp. O-1 and Halomonas elongata DSM2581: Site-directed mutagenesis and recombinant expression. Polym. Degrad. Stab. 109, 416–423. doi:10.1016/j.polymdegradstab.2014.04.024
Jiang, X. R., Yan, X., Yu, L. P., Liu, X. Y., and Chen, G. Q. (2021). Hyperproduction of 3-hydroxypropionate by Halomonas bluephagenesis. Nat. Commun. 12, 1513. doi:10.1038/s41467-021-21632-3
Jong-Il Choi, S. Y. L., Lee, S. Y., and Han, K. (1998). Kyuboem HanCloning of the Alcaligenes latus polyhydroxyalkanoate biosynthesis genes and use of these genes for enhanced production of poly(3-hydroxybutyrate) in Escherichia coli. Appl. Environ. Microbiol. 1, 4897–4903. doi:10.1128/aem.64.12.4897-4903.1998
Jung, K., Hazenberg, W., Prieto, M., and Witholt, B. (2001). Two-stage continuous process development for the production of medium-chain-length poly(3-hydroxyalkanoates). Biotechnol. Bioeng. 72, 19–24. doi:10.1002/1097-0290(20010105)72:1<19::aid-bit3>3.0.co;2-b
Keller, M. W., Schut, G. J., Lipscomb, G. L., Menon, A. L., Iwuchukwu, I. J., Leuko, T. T., et al. (2013). Exploiting microbial hyperthermophilicity to produce an industrial chemical, using hydrogen and carbon dioxide. Proc. Natl. Acad. Sci. U. S. A. 110, 5840–5845. doi:10.1073/pnas.1222607110
Khosravi-Darani, K., Mokhtari, Z. B., Amai, T., and Tanaka, K. (2013). Microbial production of poly(hydroxybutyrate) from C(1) carbon sources. Appl. Microbiol. Biotechnol. 97, 1407–1424. doi:10.1007/s00253-012-4649-0
Kim, J., Kim, Y. J., Choi, S. Y., Lee, S. Y., and Kim, K. J. (2017). Crystal structure of Ralstonia eutropha polyhydroxyalkanoate synthase C-terminal domain and reaction mechanisms. Biotechnol. J. 1, 1600648. doi:10.1002/biot.201600648
Kourmentza, C., Plácido, J., Venetsaneas, N., Burniol-Figols, A., Varrone, C., Gavala, H. N., et al. (2017). Recent advances and challenges towards sustainable polyhydroxyalkanoate (PHA) production. Bioengineering 4, 55. doi:10.3390/bioengineering4020055
Lan, L.-H., Zhao, H., Chen, J.-C., and Chen, G.-Q. (2016). Engineering Halomonas spp. as A low-cost production host for production of bio-surfactant protein PhaP. Biotechnol. J. 11, 1595–1604. doi:10.1002/biot.201600459
Lee, J., Park, H. J., Moon, M., Lee, J.-S., and Min, K. (2021). Recent progress and challenges in microbial polyhydroxybutyrate (PHB) production from CO2 as a sustainable feedstock: A state-of-the-art review. Bioresour. Technol. 339, 125616. doi:10.1016/j.biortech.2021.125616
Li, H. F., Wang, M. R., Tian, L. Y., and Li, Z. J. (2021a). Production of polyhydroxyalkanoates (PHAs) by Vibrio alginolyticus strains isolated from salt fields. Molecules 26 (20), 6283. doi:10.3390/molecules26206283
Li, M., Chen, X., Che, X., Zhang, H., Wu, L. P., Du, H., et al. (2019). Engineering Pseudomonas entomophila for synthesis of copolymers with defined fractions of 3-hydroxybutyrate and medium-chain-length 3-hydroxyalkanoates. Metab. Eng. 52, 253–262. doi:10.1016/j.ymben.2018.12.007
Li, M., Ma, Y., Zhang, X., Zhang, L., Chen, X., Ye, J. W., et al. (2021b). Tailor-made polyhydroxyalkanoates by reconstructing Pseudomonas entomophila. Adv. Mat. 33, e2102766. doi:10.1002/adma.202102766
Li, T., Guo, Y.-Y., Qiao, G.-Q., and Chen, G.-Q. (2016). Microbial synthesis of 5-aminolevulinic acid and its coproduction with polyhydroxybutyrate. ACS Synth. Biol. 5, 1264–1274. doi:10.1021/acssynbio.6b00105
Li, Z. J., Qiao, K., Che, X. M., and Stephanopoulos, G. (2017). Metabolic engineering of Escherichia coli for the synthesis of the quadripolymer poly(glycolate-co-lactate-co-3-hydroxybutyrate-co-4-hydroxybutyrate) from glucose. Metab. Eng. 44, 38–44. doi:10.1016/j.ymben.2017.09.003
Liang, P., Zhang, Y., Xu, B., Zhao, Y., Liu, X., Gao, W., et al. (2020). Deletion of genomic islands in the Pseudomonas putida KT2440 genome can create an optimal chassis for synthetic biology applications. Microb. Cell. Fact. 19, 70. doi:10.1186/s12934-020-01329-w
Lim, H., Chuah, J.-A., Chek, M. F., Tan, H. T., Hakoshima, T., Sudesh, K., et al. (2021). Identification of regions affecting enzyme activity, substrate binding, dimer stabilization and polyhydroxyalkanoate (PHA) granule morphology in the PHA synthase of Aquitalea sp. USM4. Int. J. Biol. Macromol. 186, 414–423. doi:10.1016/j.ijbiomac.2021.07.041
Lin, Z., Zhang, Y., Yuan, Q., Liu, Q., Li, Y., Wang, Z., et al. (2015). Metabolic engineering of Escherichia coli for poly (3-hydroxybutyrate) production via threonine bypass. Microb. Cell. Fact. 14, 185. doi:10.1186/s12934-015-0369-3
Linares-Pastén, J. A., Sabet-Azad, R., Pessina, L., Sardari, R. R. R., Ibrahim, M. H. A., Hatti-Kaul, R., et al. (2015). Efficient poly(3-hydroxypropionate) production from glycerol using Lactobacillus reuteri and recombinant Escherichia coli harboring L. reuteri propionaldehyde dehydrogenase and Chromobacterium sp. PHA synthase genes. Bioresour. Technol. 180, 172–176. doi:10.1016/j.biortech.2014.12.099
Ling, C., Qiao, G.-Q., Shuai, B.-W., Song, K.-N., Yao, W.-X., Jiang, X.-R., et al. (2019). Engineering self-flocculating Halomonas campaniensis for wastewaterless open and continuous fermentation. Biotechnol. Bioeng. 116, 805–815. doi:10.1002/bit.26897
Ling, C., Qiao, G. Q., Shuai, B. W., Olavarria, K., Yin, J., Xiang, R. J., et al. (2018). Engineering NADH/NAD(+) ratio in Halomonas bluephagenesis for enhanced production of polyhydroxyalkanoates (PHA). Metab. Eng. 49, 275–286. doi:10.1016/j.ymben.2018.09.007
Lv, L., Ren, Y. L., Chen, J. C., Wu, Q., and Chen, G. Q. (2015). Application of CRISPRi for prokaryotic metabolic engineering involving multiple genes, a case study: Controllable P(3HB-co-4HB) biosynthesis. Metab. Eng. 29, 160–168. doi:10.1016/j.ymben.2015.03.013
Ma, H., Zhao, Y., Huang, W., Zhang, L., Wu, F., Ye, J., et al. (2020). Rational flux-tuning of Halomonas bluephagenesis for co-production of bioplastic PHB and ectoine. Nat. Commun. 11, 3313. doi:10.1038/s41467-020-17223-3
Martin, D. P., and Williams, S. F. (2003). Medical applications of poly-4-hydroxybutyrate: A strong flexible absorbable biomaterial. Biochem. Eng. J. 16, 97–105. doi:10.1016/s1369-703x(03)00040-8
Matsumoto, K. I., Takase, K., Yamamoto, Y., Doi, Y., and Taguchi, S. (2009). Chimeric enzyme composed of polyhydroxyalkanoate (PHA) synthases from Ralstonia eutropha and Aeromonas caviae enhances production of PHAs in recombinant Escherichia coli. Biomacromolecules 10, 682–685. doi:10.1021/bm801386j
Mehrpouya, M., Vahabi, H., Barletta, M., Laheurte, P., and Langlois, V. (2021). Additive manufacturing of polyhydroxyalkanoates (PHAs) biopolymers: Materials, printing techniques, and applications. Mater. Sci. Eng. C 127, 112216. doi:10.1016/j.msec.2021.112216
Meijnen, J. P., De Winde, J. H., and Ruijssenaars, H. J. (2008). Engineering Pseudomonas putida S12 for efficient utilization of D-xylose and L-arabinose. Appl. Environ. Microbiol. 74, 5031–5037. doi:10.1128/aem.00924-08
Meng, D. C., Wang, Y., Wu, L. P., Shen, R., Chen, J. C., Wu, Q., et al. (2015). Production of poly(3-hydroxypropionate) and poly(3-hydroxybutyrate-co-3-hydroxypropionate) from glucose by engineering Escherichia coli. Metab. Eng. 29, 189–195. doi:10.1016/j.ymben.2015.03.015
Miranda De Sousa Dias, M., Koller, M., Puppi, D., Morelli, A., Chiellini, F., and Braunegg, G. (2017). Fed-batch synthesis of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-4-hydroxybutyrate) from sucrose and 4-hydroxybutyrate precursors by Burkholderia sacchari strain DSM 17165. Bioengineering 4, 36. doi:10.3390/bioengineering4020036
Mizuno, S., Enda, Y., Saika, A., Hiroe, A., and Tsuge, T. (2018). Biosynthesis of polyhydroxyalkanoates containing 2-hydroxy-4-methylvalerate and 2-hydroxy-3-phenylpropionate units from a related or unrelated carbon source. J. Biosci. Bioeng. 125, 295–300. doi:10.1016/j.jbiosc.2017.10.010
Modi, S., Koelling, K., and Vodovotz, Y. (2011). Assessment of PHB with varying hydroxyvalerate content for potential packaging applications. Eur. Polym. J. 47, 179–186. doi:10.1016/j.eurpolymj.2010.11.010
Możejko-Ciesielska, J., and Kiewisz, R. (2016). Bacterial polyhydroxyalkanoates: Still fabulous? Microbiol. Res. 192, 271–282. doi:10.1016/j.micres.2016.07.010
Murugan, P., Gan, C.-Y., and Sudesh, K. (2017). Biosynthesis of P(3HB-co-3HHx) with improved molecular weights from a mixture of palm olein and fructose by Cupriavidus necator Re2058/pCB113. Int. J. Biol. Macromol. 102, 1112–1119. doi:10.1016/j.ijbiomac.2017.05.006
Norhafini, H., Huong, K.-H., and Amirul, A. A. (2019). High PHA density fed-batch cultivation strategies for 4HB-rich P(3HB-co-4HB) copolymer production by transformant Cupriavidus malaysiensis USMAA1020. Int. J. Biol. Macromol. 125, 1024–1032. doi:10.1016/j.ijbiomac.2018.12.121
Obruča, S., Dvořák, P., Sedláček, P., Koller, M., Sedlář, K., Pernicová, I., et al. (202210790). Polyhydroxyalkanoates synthesis by halophiles and thermophiles: Towards sustainable production of microbial bioplastics. Biotechnol. Adv. 58, 107906. doi:10.1016/j.biotechadv.2022.107906
Ouyang, P., Wang, H., Hajnal, I., Wu, Q., Guo, Y., Chen, G. Q., et al. (2018). Increasing oxygen availability for improving poly(3-hydroxybutyrate) production by Halomonas. Metab. Eng. 45, 20–31. doi:10.1016/j.ymben.2017.11.006
Pacholak, A., Gao, Z.-L., Gong, X.-Y., Kaczorek, E., and Cui, Y.-W. (2021). The metabolic pathways of polyhydroxyalkanoates and exopolysaccharides synthesized by Haloferax mediterranei in response to elevated salinity. J. Proteomics 232, 104065. doi:10.1016/j.jprot.2020.104065
Panich, J., Fong, B., and Singer, S. W. (2021). Metabolic engineering of cupriavidus necator H16 for sustainable biofuels from CO2. Trends Biotechnol. 39, 412–424. doi:10.1016/j.tibtech.2021.01.001
Park, S. J., Ahn, W. S., Green, P. R., and Lee, S. Y. (2001). Production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) by metabolically engineered Escherichia coli strains. Biomacromolecules 2, 248–254. doi:10.1021/bm000105u
Park, S. J., Kim, T. W., Kim, M. K., Lee, S. Y., and Lim, S.-C. (2012a). Advanced bacterial polyhydroxyalkanoates: Towards a versatile and sustainable platform for unnatural tailor-made polyesters. Biotechnol. Adv. 30, 1196–1206. doi:10.1016/j.biotechadv.2011.11.007
Park, S. J., Lee, S. Y., Kim, T. W., Jung, Y. K., and Yang, T. H. (2012b). Biosynthesis of lactate-containing polyesters by metabolically engineered bacteria. Biotechnol. J. 7, 199–212. doi:10.1002/biot.201100070
Park, S. J., Lee, T. W., Lim, S. C., Kim, T. W., Lee, H., Kim, M. K., et al. (2012c). Biosynthesis of polyhydroxyalkanoates containing 2-hydroxybutyrate from unrelated carbon source by metabolically engineered Escherichia coli. Appl. Microbiol. Biotechnol. 93, 273–283. doi:10.1007/s00253-011-3530-x
Park, S. J., Park, J. P., and Lee, S. Y. (2002). Metabolic engineering of Escherichia coli for the production of medium-chain-length polyhydroxyalkanoates rich in specific monomers. FEMS Microbiol. Lett. 214, 217–222. doi:10.1111/j.1574-6968.2002.tb11350.x
Prieto, A., Escapa, I. F., Martinez, V., Dinjaski, N., Herencias, C., De La Pena, F., et al. (2016). A holistic view of polyhydroxyalkanoate metabolism in Pseudomonas putida. Environ. Microbiol. 18, 341–357. doi:10.1111/1462-2920.12760
Raberg, M., Volodina, E., Lin, K., and Steinbuchel, A. (2018). Ralstonia eutropha H16 in progress: Applications beside PHAs and establishment as production platform by advanced genetic tools. Crit. Rev. Biotechnol. 38, 494–510. doi:10.1080/07388551.2017.1369933
Ryu, H. W., S, K. H., Chang, Y. K., and Chang, H. N. (1996). Production of poly(3-hydroxybutyrate) by high cell density fed-batch culture of Alcaligenes eutrophus with phospate limitation. Biotechnol. Bioeng. 55 (51), 28–32. doi:10.1002/(sici)1097-0290(19970705)55:1<28::aid-bit4>3.0.co;2-z
Ryu, H. W., Hahn, S. K., Chang, Y. K., and Chang, H. N. (1997). Production of poly(3‐hydroxybutyrate) by high cell density fed‐batch culture of Alcaligenes eutrophus with phospate limitation. Biotechnol. Bioeng. 55, 28–32. doi:10.1002/(sici)1097-0290(19970705)55:1<28::aid-bit4>3.0.co;2-z
Salehizadeh, H., Yan, N., and Farnood, R. (2020). Recent advances in microbial CO2 fixation and conversion to value-added products. Chem. Eng. J. 390, 124584. doi:10.1016/j.cej.2020.124584
Sathiyanarayanan, G., Kiran, G. S., Selvin, J., and Saibaba, G. (2013). Optimization of polyhydroxybutyrate production by marine Bacillus megaterium MSBN04 under solid state culture. Int. J. Biol. Macromol. 60, 253–261. doi:10.1016/j.ijbiomac.2013.05.031
Scheel, R. A., Ho, T., Kageyama, Y., Masisak, J., Mckenney, S., Lundgren, B. R., et al. (2021). Optimizing a fed-batch high-density fermentation process for medium chain-length poly(3-hydroxyalkanoates) in Escherichia coli. Front. Bioeng. Biotechnol. 9, 618259. doi:10.3389/fbioe.2021.618259
Shen, R., Yin, J., Ye, J.-W., Xiang, R.-J., Ning, Z.-Y., Huang, W.-Z., et al. (2018). Promoter engineering for enhanced P (3HB-co-4HB) production by Halomonas bluephagenesis. ACS Synth. Biol. 7, 1897–1906. doi:10.1021/acssynbio.8b00102
Silva, F., Campanari, S., Matteo, S., Valentino, F., Majone, M., Villano, M., et al. (2017). Impact of nitrogen feeding regulation on polyhydroxyalkanoates production by mixed microbial cultures. N. Biotechnol. 37, 90–98. doi:10.1016/j.nbt.2016.07.013
Steinbüchel, A. (2001). Perspectives for biotechnological production and utilization of biopolymers: Metabolic engineering of polyhydroxyalkanoate biosynthesis pathways as a successful example. Macromol. Biosci. 1, 1–24. doi:10.1002/1616-5195(200101)1:1<1::aid-mabi1>3.0.co;2-b
Sudesh, K., Abe, H., and Doi, Y. (2000). Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 25, 1503–1555. doi:10.1016/s0079-6700(00)00035-6
Sudo, M., Hori, C., Ooi, T., Mizuno, S., Tsuge, T., Matsumoto, K. I., et al. (2020). Synergy of valine and threonine supplementation on poly(2-hydroxybutyrate-block-3-hydroxybutyrate) synthesis in engineered Escherichia coli expressing chimeric polyhydroxyalkanoate synthase. J. Biosci. Bioeng. 129, 302–306. doi:10.1016/j.jbiosc.2019.09.018
Tan, D., Xue, Y. S., Aibaidula, G., and Chen, G. Q. (2011). Unsterile and continuous production of polyhydroxybutyrate by Halomonas TD01. Bioresour. Technol. 102, 8130–8136. doi:10.1016/j.biortech.2011.05.068
Tan, I. K. P., Foong, C. P., Tan, H. T., Lim, H., Zain, N. A., Tan, Y. C., et al. (2020). Polyhydroxyalkanoate (PHA) synthase genes and PHA-associated gene clusters in Pseudomonas spp. and Janthinobacterium spp. isolated from Antarctica. J. Biotechnol. 313, 18–28. doi:10.1016/j.jbiotec.2020.03.006
Tanaka, K., Yoshida, K., Orita, I., and Fukui, T. (2021). Biosynthesis of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a recombinant cupriavidusnecator. Bioengineering 8, 179. doi:10.3390/bioengineering8110179
Tariq, A., Hameed, A., Bokhari, H., and Masood, F. (2015). Is atomic rearrangement of type IV PHA synthases responsible for increased PHA production? J. Biomol. Struct. Dyn. 33, 1225–1238. doi:10.1080/07391102.2014.941401
Ushimaru, K., Motoda, Y., Numata, K., and Tsuge, T. (2014). Phasin proteins activate Aeromonas caviae polyhydroxyalkanoate (PHA) synthase but not Ralstonia eutropha PHA synthase. Appl. Environ. Microbiol. 80, 2867–2873. doi:10.1128/aem.04179-13
Van Thuoc, D., Loan, T. T., Trung, T. A., Van Quyen, N., Tung, Q. N., Tien, P. Q., et al. (2020). Genome mining reveals the biosynthetic pathways of polyhydroxyalkanoate and ectoines of the halophilic strain Salinivibrio proteolyticus M318 isolated from fermented shrimp paste. Mar. Biotechnol. (NY). 22, 651–660. doi:10.1007/s10126-020-09986-z
Van Thuoc, D., My, D. N., Loan, T. T., and Sudesh, K. (2019). Utilization of waste fish oil and glycerol as carbon sources for polyhydroxyalkanoate production by Salinivibrio sp. M318. Int. J. Biol. Macromol. 141, 885–892. doi:10.1016/j.ijbiomac.2019.09.063
Volova, T., Trusova, M., Kalacheva, G., and Kozhevnicov, I. (2006). Physiological–biochemical properties and the ability to synthesize polyhydroxyalkanoates of the glucose-utilizing strain of the hydrogen bacterium Ralstonia eutropha B8562. Appl. Microbiol. Biotechnol. 73, 429–433. doi:10.1007/s00253-006-0460-0
Wang, M. R., Li, H. F., Yi, J. J., Tao, S. Y., and Li, Z. J. (2022). Production of polyhydroxyalkanoates by three novel species of Marinobacterium. Int. J. Biol. Macromol. 195, 255–263. doi:10.1016/j.ijbiomac.2021.12.019
Wang, X., Han, J.-N., Zhang, X., Ma, Y.-Y., Lin, Y., Wang, H., et al. (2021a). Reversible thermal regulation for bifunctional dynamic control of gene expression in Escherichia coli. Nat. Commun. 12, 1411. doi:10.1038/s41467-021-21654-x
Wang, Y., Wu, H., Jiang, X., and Chen, G.-Q. (2014). Engineering Escherichia coli for enhanced production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) in larger cellular space. Metab. Eng. 25, 183–193. doi:10.1016/j.ymben.2014.07.010
Wang, Z., Qin, Q., Zheng, Y., Li, F., Zhao, Y., Chen, G.-Q., et al. (2021b). Engineering the permeability of Halomonas bluephagenesis enhanced its chassis properties. Metab. Eng. 67, 53–66. doi:10.1016/j.ymben.2021.05.010
Xiong, B., Li, Z., Liu, L., Zhao, D., Zhang, X., Bi, C., et al. (2018). Genome editing of Ralstonia eutropha using an electroporation-based CRISPR-Cas9 technique. Biotechnol. Biofuels 11, 172. doi:10.1186/s13068-018-1170-4
Yan, X., Liu, X., Yu, L. P., Wu, F., Jiang, X. R., Chen, G. Q., et al. (2022). Biosynthesis of diverse α, ω-diol-derived polyhydroxyalkanoates by engineered Halomonas bluephagenesis. Metab. Eng. 72, 275–288. doi:10.1016/j.ymben.2022.04.001
Yang, J. E., Choi, Y. J., Lee, S. J., Kang, K. H., Lee, H., Oh, Y. H., et al. (2014). Metabolic engineering of Escherichia coli for biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from glucose. Appl. Microbiol. Biotechnol. 98, 95–104. doi:10.1007/s00253-013-5285-z
Yang, J. E., Park, S. J., Kim, W. J., Kim, H. J., Kim, B. J., Lee, H., et al. (2018). One-step fermentative production of aromatic polyesters from glucose by metabolically engineered Escherichia coli strains. Nat. Commun. 9, 79. doi:10.1038/s41467-017-02498-w
Yang, S., Li, S., and Jia, X. (2019). Production of medium chain length polyhydroxyalkanoate from acetate by engineered Pseudomonas putida KT2440. J. Ind. Microbiol. Biotechnol. 46, 793–800. doi:10.1007/s10295-019-02159-5
Yang, T. H., Kim, T. W., Kang, H. O., Lee, S. H., Lee, E. J., Lim, S. C., et al. (2010). Biosynthesis of polylactic acid and its copolymers using evolved propionate CoA transferase and PHA synthase. Biotechnol. Bioeng. 105, 150–160. doi:10.1002/bit.22547
Yang, X., Li, S., and Jia, X. (2020). A four-microorganism three-step fermentation process for producing medium-chain-length polyhydroxyalkanoate from starch. Biotech 10, 352. doi:10.1007/s13205-020-02347-6
Yao J, Z., Wu, Q., Chen, G., and Zhang, R. (1999). Production of polyhydroxyalkanoates by Pseudomonas nitroreducens. Antonie Leeuwenhoek 75 (74), 345–349. doi:10.1023/a:1002082303615
Ye, J., Hu, D., Yin, J., Huang, W., Xiang, R., Zhang, L., et al. (2020). Stimulus response-based fine-tuning of polyhydroxyalkanoate pathway in Halomonas. Metab. Eng. 57, 85–95. doi:10.1016/j.ymben.2019.10.007
Ye, J., Huang, W., Wang, D., Chen, F., Yin, J., Li, T., et al. (2018). Pilot scale-up of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) production by Halomonas bluephagenesis via cell growth adapted optimization process. Biotechnol. J. 13, 1800074. doi:10.1002/biot.201800074
Ye, J. W., and Chen, G. Q. (2021). Halomonas as a chassis. Essays Biochem. 65, 393–403. doi:10.1042/ebc20200159
Young, E. M., Zhao, Z., Gielesen, B. E., Wu, L., Gordon, D. B., Roubos, J. A., et al. (2018). Iterative algorithm-guided design of massive strain libraries, applied to itaconic acid production in yeast. Metab. Eng. 48, 33–43. doi:10.1016/j.ymben.2018.05.002
Yu, L. P., Yan, X., Zhang, X., Chen, X. B., Wu, Q., Jiang, X. R., et al. (2020). Biosynthesis of functional polyhydroxyalkanoates by engineered Halomonas bluephagenesis. Metab. Eng. 59, 119–130. doi:10.1016/j.ymben.2020.02.005
Yue, H., Ling, C., Yang, T., Chen, X., Chen, Y., Deng, H., et al. (2014). A seawater-based open and continuous process for polyhydroxyalkanoates production by recombinant Halomonas campaniensis LS21 grown in mixed substrates. Biotechnol. Biofuels 7, 108. doi:10.1186/1754-6834-7-108
Zhang, X., Lin, Y., Wu, Q., Wang, Y., and Chen, G. Q. (2020). Synthetic biology and genome-editing tools for improving PHA metabolic engineering. Trends Biotechnol. 38, 689–700. doi:10.1016/j.tibtech.2019.10.006
Zhou, H., Vonk, B., Roubos, J. A., Bovenberg, R. A., and Voigt, C. A. (2015). Algorithmic co-optimization of genetic constructs and growth conditions: Application to 6-ACA, a potential nylon-6 precursor. Nucleic Acids Res. 43, 10560–10570. doi:10.1093/nar/gkv1071
Zhou, Q., Shi, Z. Y., Meng, D. C., Wu, Q., Chen, J. C., Chen, G. Q., et al. (2011). Production of 3-hydroxypropionate homopolymer and poly(3-hydroxypropionate-co-4-hydroxybutyrate) copolymer by recombinant Escherichia coli. Metab. Eng. 13, 777–785. doi:10.1016/j.ymben.2011.10.002
Keywords: polyhydroxyalkanoates, synthetic pathway, metabolic engineering, PHA synthase, microbial production
Citation: Gao Q, Yang H, Wang C, Xie X-Y, Liu K-X, Lin Y, Han S-Y, Zhu M, Neureiter M, Lin Y and Ye J-W (2022) Advances and trends in microbial production of polyhydroxyalkanoates and their building blocks. Front. Bioeng. Biotechnol. 10:966598. doi: 10.3389/fbioe.2022.966598
Received: 11 June 2022; Accepted: 01 July 2022;
Published: 19 July 2022.
Edited by:
Xinjun Feng, Qingdao Institute of Bioenergy and Bioprocess Technology (CAS), ChinaReviewed by:
Zheng-Jun Li, Beijing University of Chemical Technology, ChinaDan Tan, Xi’an Jiaotong University, China
Copyright © 2022 Gao, Yang, Wang, Xie, Liu, Lin, Han, Zhu, Neureiter, Lin and Ye. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Markus Neureiter, bWFya3VzLm5ldXJlaXRlckBib2t1LmFjLmF0; Yina Lin, bGlueWluYTIwMjFAMTYzLmNvbQ==; Jian-Wen Ye, eWVqaWFud2VuQHNjdXQuZWR1LmNu
†These authors have contributed equally to this work