 Takumi Yamada1
Takumi Yamada1 Shinya Aoi2*
Shinya Aoi2* Mau Adachi1
Mau Adachi1 Tomoya Kamimura3
Tomoya Kamimura3 Yasuo Higurashi4
Yasuo Higurashi4 Naomi Wada4Kazuo Tsuchiya2
Naomi Wada4Kazuo Tsuchiya2 Fumitoshi Matsuno1
Fumitoshi Matsuno1- 1Department of Mechanical Engineering and Science, Graduate School of Engineering, Kyoto University, Kyoto, Japan
- 2Department of Aeronautics and Astronautics, Graduate School of Engineering, Kyoto University, Kyoto, Japan
- 3Department of Electrical and Mechanical Engineering, Nagoya Institute of Technology, Nagoya, Japan
- 4Laboratory of System Physiology, Joint Faculty of Veterinary Medicine, Yamaguchi University, Yamaguchi, Japan
Horses use the transverse gallop in high-speed running. However, different animals use different gaits, and the gait preference of horses remains largely unclear. Horses have fore-aft asymmetry in their body structure and their center of mass (CoM) is anteriorly located far from the center of the body. Since such a CoM offset affects the running dynamics, we hypothesize that the CoM offset of horses is important in gait selection. In order to verify our hypothesis and clarify the gait selection mechanisms by horses from a dynamic viewpoint, we developed a simple model with CoM offset and investigated its effects on running. Specifically, we numerically obtained periodic solutions and classified these solutions into six types of gaits, including the transverse gallop, based on the footfall pattern. Our results show that the transverse gallop is optimal when the CoM offset is located at the position estimated in horses. Our findings provide useful insight into the gait selection mechanisms in high-speed running of horses.
1 Introduction
Horses use the transverse gallop in high-speed locomotion. This gait has one flight phase in one gait cycle. Specifically, the hind legs first touch the ground, and then the fore legs touch the ground. After that, a flight phase appears. This gait is different from the rotary gallop in cheetahs, which has two flight phases, each of which appears after the touchdowns of the fore legs and those of the hind legs (Bertram and Gutmann, 2008; Biancardi and Minetti, 2012). The gaits of quadrupeds when running at their fastest speeds vary between species, and it remains unclear why horses use the transverse gallop.
Horses have fore-aft asymmetry in their body structure. In particular, they have a long neck, and their center of mass (CoM) is anteriorly located and far from the center of the body (Buchner et al., 1997; Self Davies et al., 2019). Such a CoM offset affects the dynamics of the running motion. For example, when the fore-aft CoM location of dogs was changed by carrying a weight during trotting, which is characterized by the simultaneous touchdown of the diagonal fore and hind legs, the footfall pattern changed (Lee et al., 2004). Specifically, the fore and hind legs came to touch the ground first when the load was applied to the anterior and posterior sides, respectively. In other words, the CoM offset changed the gait. Therefore, we hypothesize that the CoM offset of horses plays an important role in their gait selection.
Since animal locomotion is a complex phenomenon generated through dynamical interactions between the body mechanical system, the nervous system, and the environment, it is difficult to fully understand the mechanisms for gait selection in animals only from observation. Therefore, simple models, which extract essential elements for the running dynamics, have been used to clarify the mechanisms (Tanase et al., 2015; Gan et al., 2016; Chen et al., 2019; Kamimura et al., 2021). Poulakakis et al. (2006) used a simple quadrupedal model and showed the relationship between the pitch angular velocity and the number of flight phases in one gait cycle during bounding gait. In addition, Zou and Schmiedeler (2006) used a model focusing on the vertical and pitch movements and derived a stability condition depending on the CoM offset. However, the model did not incorporate horizontal movement, and the mechanism for the gait selection remains unclear.
In the present study, we investigated the effects of the CoM offset on quadrupedal running in order to verify our hypothesis from a dynamic viewpoint. Specifically, we constructed a bounding model with CoM offset and searched periodic solutions by numerical simulations. We then classified the obtained solutions into six types of gaits depending on the footfall pattern and examined which gait is optimal based on performance criteria. Our findings provide useful insight into the mechanisms for high-speed running in horses.
2 Methods
2.1 Model
We used a horse model composed of a rigid body and two massless springs on the sagittal plane (Figure 1). The springs represent the fore and hind legs (Legs F and H) and are connected to the body by smoothly rotating joints. Here, M and J are the mass and moment of inertia around the CoM of the body, respectively. The distance between the leg joints is 2D. The CoM is located at a distance of αD (−1 ≤ α ≤ 1) from the center C between the leg joints, where α = 0 corresponds to C, and α = 1 and −1 correspond to the joints of the fore and hind legs, respectively. Moreover, x and z are the horizontal and vertical positions of the CoM, respectively, and θ is the pitch angle relative to the horizontal line. The spring constant and neutral length of both the fore and hind legs are K and L0, respectively. When Leg i (i = F, H) is in the air, its length remains L0 and its angle also maintains the touchdown angle
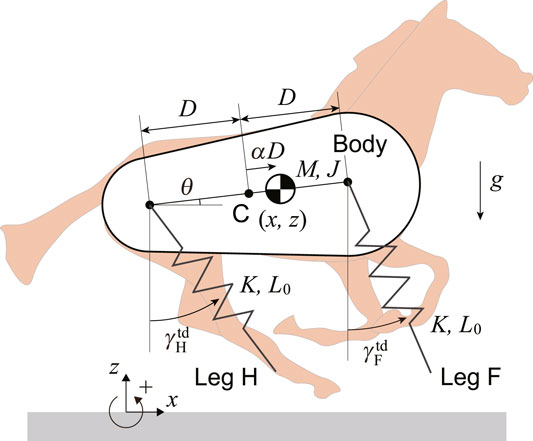
FIGURE 1. Horse model composed of a rigid body with center of mass offset and two massless springs.
The equations of motion of the model are given by
where
and Li and γi (i = F, H) are the length and angle, respectively, of Leg i relative to the vertical line. Moreover, γi is determined by the joint and touchdown positions of Leg i. The touchdown condition
where ɛF = 1 and ɛH = −1.
The physical parameters of the model were determined based on the estimated values of Thoroughbreds (Equus ferus caballus). In particular, we used M = 490 kg and J = 167 kgm2 based on Swanstrom et al. (2005). We used D = 0.48 m from the distance between the shoulder and hip joints and L0 = 1.33 m from the average value of the distances between the shoulder joint and the toe of the fore limb and between the hip joint and the toe of the hind limb based on Grossi and Canals, (2010). We used K = 45.4 kN/m based on Farley et al. (1993).
2.2 Gait
The gait is generally determined based on the order of touchdown and liftoff of the legs. We defined the following four phases: flight (FL), fore leg stance (FS), hind leg stance (HS), and double stance (DS). In FL, both legs are in the air. In FS, only the fore leg is in contact with the ground. In HS, only the hind leg is in contact with the ground. In DS, both legs are in contact with the ground. We investigated motions (periodic solutions) starting from an apex (i.e.,
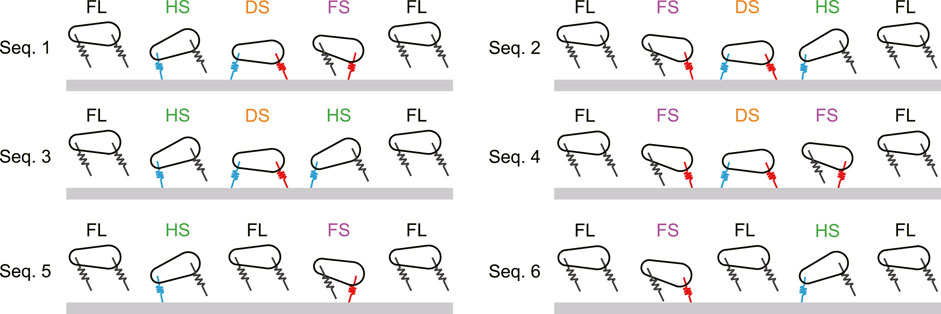
FIGURE 2. Six possible phase transitions (Sequences 1 … 6) from one apex to the next apex. FL, FS, HS, and DS stand for flight, fore leg stance, hind leg stance, and double stance, respectively.
2.3 Search of Solutions
In order to find periodic solutions, we defined the Poincaré section at the apex of the CoM
where qn is the state at the nth intersection with the Poincaré section, and un is the nth parameter set. A periodic solution satisfies
where q∗ is the fixed point on the Poincaré section. We numerically searched fixed points for periodic solutions using the Newton-Raphson method.
2.4 Performance Criteria
In order to evaluate the obtained solutions, we used the gait stability as a performance criterion (Poulakakis et al., 2006; Tanase et al., 2015; Kamimura et al., 2021). In order to analyze the gait stability, we investigated the eigenvalues of the linearized Poincaré map around the fixed points on the Poincaré section. Since our model is energy conservative, the solution is asymptotically stable, when all of the eigenvalues, except for one eigenvalue of 1, are located inside the unit circle on the complex plane. Otherwise, the solution is unstable.
Horses stabilize their gaze during running by preventing the pitch movement of the body from disturbing the head (Dunbar et al., 2008). Therefore, we also used the fluctuation of the pitch movement of the body as another performance criterion, which is obtained from the difference between the maximum and minimum values of θ for one gait cycle.
3 Results
3.1 Effect of Center of Mass Offset on Gait Pattern
First, we set the total energy of our model as E = 20.3 kJ (gravitational potential energy is 0 at the ground level) and the forward speed at the apex as
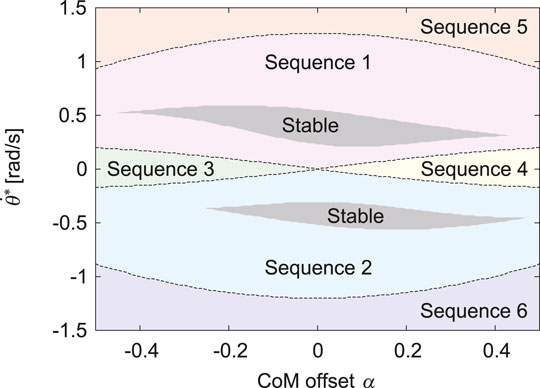
FIGURE 3. Gait classification of obtained periodic solutions for α and
Next, we investigate the time profiles of the periodic solutions in order to clarify the characteristition with α. First, Figure 4A shows the tixme profile of
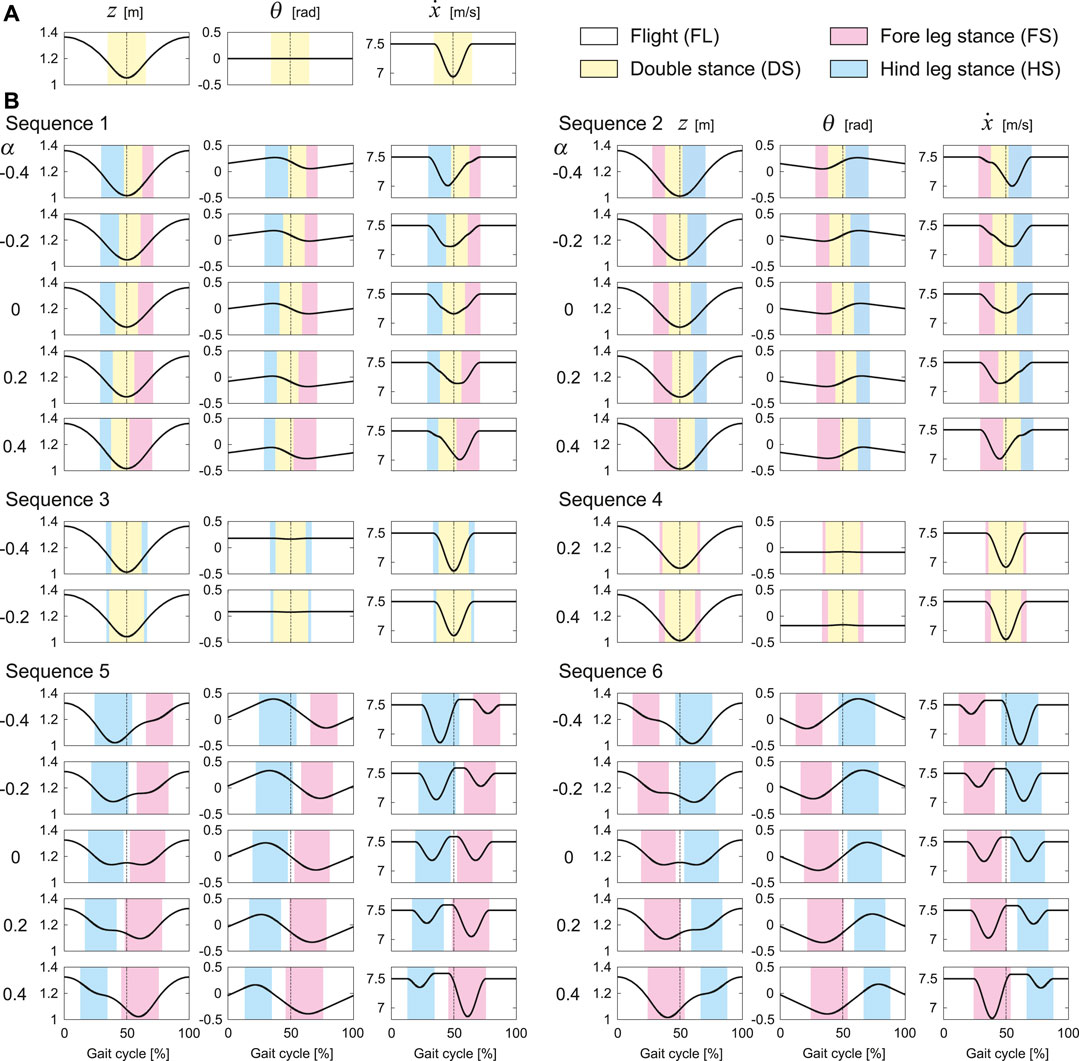
FIGURE 4. Characteristics of time profiles of
Sequences 1 and 2 have solutions for all α = 0, ±0.2, and ±0.4, the trajectories and phases of which are symmetric with respect to 50% of the gait cycle. As α increases, the onset and end of the DS phase are advanced in Sequence 1 and delayed in Sequence 2, whereas those of the FL phase remains almost unchanged. Here, z has a one-peak shape and remains almost unchanged. Although the waveform of θ remains almost unchanged, the mean value decreases. The timing at which
Sequence 3 has solutions only for α = −0.2 and −0.4, and Sequence 4 has solutions only for α = 0.2 and 0.4. These trajectories are identical for the same |α|. In addition, these phases are also identical when the timings of touchdown and liftoff are swapped between the fore and hind legs. Unlike Sequences 1 and 2, the trajectories and the timings of touchdown and liftoff are symmetric with respect to 50% of the gait cycle regardless of α. Here, z has a one-peak shape and remains almost unchanged as α increases. The waveform of θ remains almost unchanged, whereas the mean value decreases. The minimum value of
Sequences 5 and 6 have solutions for all α = 0, ±0.2, and ±0.4, the trajectories and phases of which are symmetric with respect to 50% of the gait cycle. When α = 0, z has a two-peak shape. As |α| increases, one of the two peaks decreases and the two-peak shape changes into a one-peak shape. As α increases, the FS and HS phases are advanced in Sequence 5 and delayed in Sequence 6. Whereas the mean value of θ remain almost unchanged, the peak timings change in accordance with changes in the FS and HS phases. Since Sequences 5 and 6 have two FL phases,
3.2 Effect of Speed on Gait Performance
Although the previous section investigated the effects of the CoM offset α on the gait pattern using the average speed, horses have a wide range of speed for galloping (Hoyt and Taylor, 1981; Minetti et al., 1999). In this section, we investigate the effects of speed on the gait characteristics using the estimated value of α in horses (α = 0.2) (Self Davies et al., 2019) compared with those using α = 0.
The total energy E of our model is explained by the horizontal translational kinetic energy
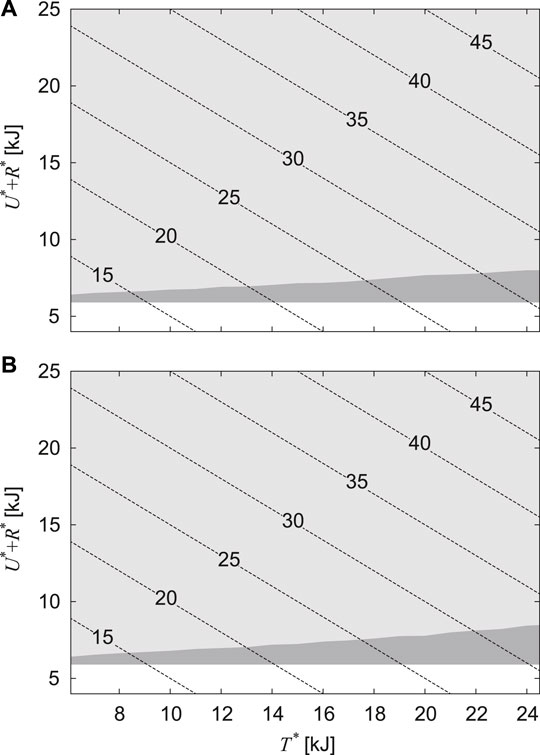
FIGURE 5. Regions in which periodic solutions are found for horizontal translational kinetic energy T* and the sum of gravitational potential energy and rotational kinetic energy U* + R*. (A) α = 0.2 and (B) α = 0. The light and dark gray regions indicate that the solutions are unstable and stable, respectively. The dotted lines indicate the contour of total energy E.
Next, we searched for periodic solutions by using U∗ + R∗ = 6.6 kJ, which corresponds to the value obtained from E = 20.3 kJ and T∗ = 13.8 kJ used in Figure 3, and by changing
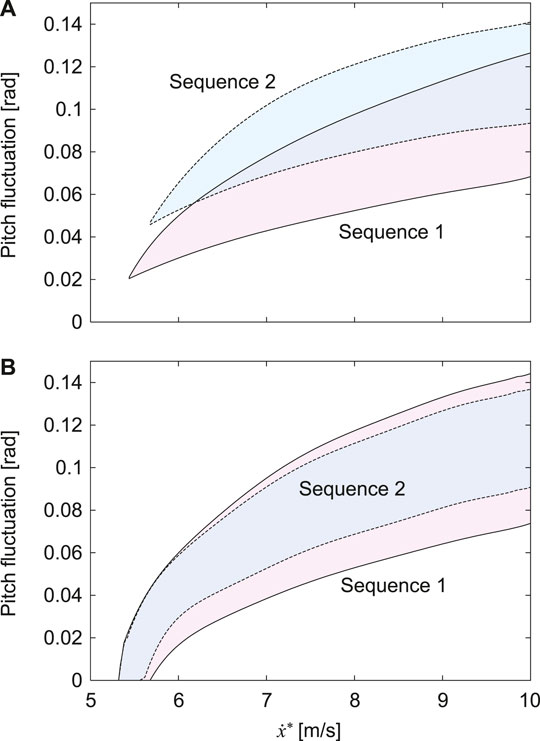
FIGURE 6. Fluctuation of pitch movement of stable solutions for
4 Discussion
4.1 Effect of Center of Mass Offset on Gait
The proposed model has six types of gaits, labeled Sequences 1 through 6 (Figure 2). In Sequences 1, 2, 5, and 6, when a leg first touches the ground, this leg leaves the ground earlier than the other leg. As a result of the search of periodic solutions, we found only these four sequences when the CoM is located at the center (α = 0), as shown in Figure 3. In Sequences 3 and 4, when a leg first touches the ground, this leg leaves the ground later than the other leg (Figure 2). Sequences 3 and 4 appeared only when the CoM is located posteriorly (α < 0) and anteriorly (α > 0), respectively (Figure 3). In other words, the introduction of the CoM offset α to the model allowed Sequences 3 and 4 to appear.
Whereas the trajectories and phases of the periodic solutions were symmetric for α = 0, the solutions became asymmetric as |α| increased (Figure 4). However, the asymmetric tendency depended on the gait. Specifically, as |α| increased, the trajectories and phases showed higher asymmetry in order of Sequences 3 and 4, Sequences 1 and 2, Sequences 5 and 6 (Figure 4). These reasons can be explained from the viewpoint of the dynamics of the body rotation. Specifically, the body rotation is created by the moment of force by the ground reaction forces from the fore and hind legs. Therefore, the periodic solutions require the moment of impulse generated for one gait cycle to be balanced between the fore and hind legs. When α > 0, the distances from the CoM to the joints of the fore and hind legs are short and long, respectively. Therefore, the moment of impulse is balanced by increasing the magnitude of the ground reaction force and stance phase duration of the fore leg and by decreasing those of the hind leg, and vice versa when α < 0. In the DS phase, the net moment applied to the body is reduced by the positive moment from the fore leg and negative moment from the hind leg, which decreases the asymmetry of the body rotation. Since the DS phase duration decreased in order of Sequences 3 and 4, Sequences 1 and 2, Sequences 5 and 6, the asymmetry of the trajectories and phases increased in this order.
This can also explain why Sequences 3 and 4 appear in α < 0 and α > 0, respectively. Specifically, in Sequences 3 and 4, when a leg first touches the ground, this leg leaves the ground later than the other leg (Figure 2). Based on the solution for α = 0 and
4.2 Gait Selection by Horses
Sequence 1 has one flight phase, after which the hind leg first touches the ground (Figure 2), and thus corresponds to the transverse gallop used by horses and gnus (Muybridge, 1957; Pennycuick, 1975; Hildebrand, 1977; Hildebrand, 1989). Sequence 2 has one flight phase, after which the fore leg first touches the ground (Figure 2), and thus corresponds to the transverse gallop used by deer and antelopes (Bigalike, 1972; FitzGibbon and Fanshawe, 1988; Hildebrand, 1989). Sequences 3 and 4 also have one flight phase (Figure 2). When touchdown and liftoff occur almost simultaneously between the fore and hind legs and the pitch fluctuation is small, as obtained in Figure 4B, these gaits correspond to the pronk used by springboks and Thomson’s gazelles (Bigalike, 1972; FitzGibbon and Fanshawe, 1988; Hildebrand, 1989). In contrast, Sequences 5 and 6 have two flight phases and thus correspond to the rotary gallop used by cheetahs and greyhounds (Muybridge, 1957; Hildebrand, 1977; Hildebrand, 1989; Bertram and Gutmann, 2008; Biancardi and Minetti, 2012; Hudson et al., 2012).
When we used the physical parameters estimated in horses, including the CoM offset α = 0.2, only Sequences 1 and 2 had stable solutions (Figures 3, 6). Sequence 1 had a wider speed range (5.5–10 m/s) than Sequence 2 (5.8–10 m/s) (Figure 6) and the speed range of Sequence 1 was closer to that of a galloping horse (5–10 m/s) (Hoyt and Taylor, 1981; Minetti et al., 1999). Furthermore, Sequence 1 had smaller pitch fluctuations (2.2–7.2 deg) than Sequence 2 (3.2–7.9 deg) (Figure 6A), and the amount of the fluctuations of Sequence 1 was closer to that of a galloping horse (2–8 deg) (Dunbar et al., 2008). However, when we used α = 0 instead of α = 0.2 estimated in horses, although Sequences 1 and 2 also had stable solutions, the speed range and pitch fluctuation were not much different between Sequence 1 and 2 (Figure 6B). Our results suggest that Sequence 1, which corresponds to the transverse gallop actually used by horses, is a suitable gait for horses from a dynamical viewpoint.
4.3 Vertical and Pitch Movements
Although stable periodic solutions existed for a large range of the horizontal translational kinetic energy T∗, these solutions existed for a limited range of the sum of the gravitational and rotational kinetic energies U∗ + R∗ (Figure 5). This means that the stability of bounding mainly depends on the vertical and pitch movements of the body. Thus far, simple models focusing on the vertical and pitching movements have been used to investigate the gait stability (Berkemeier, 1998; Zou and Schmiedeler, 2006; De and Koditschek, 2018; Kamimura et al., 2021). In particular, Berkemeier (1998) investigated the stability of bounding (which corresponds to Sequences 5 and 6) using a symmetrical model, which is identical to our model with α = 0, and derived the stability condition as μ < 1, where μ = J/(MD2). Zou and Schmiedeler (2006) improved his model by introducing the CoM offset α, as in the present study, and derived the stability condition for α > 0 as μ < 1 − α2. For the physical parameter μ = 1.48 estimated in horses (Swanstrom et al., 2005), our results showed that all solutions of Sequences 5 and 6 were unstable, regardless of α (Figure 3), which is consistent with their results.
While dogs have a larger CoM offset (α = 0.28) (Ben-Amotz et al., 2020) than horses (α = 0.2) (Self Davies et al., 2019), they use both transverse and rotary gallop depending on the speed (Biancardi and Minetti, 2012). Polet (2021) showed that the pitch moment of inertia plays an important role for the gait determination using a simple model. In addition to the CoM offset, we would like to investigate the contribution of the pitch moment of inertia to the gait selection in the future.
4.4 Limitations and Future Research
Ground reaction forces of animals during fast running show sinusoidal patterns (Alexander et al., 1986; Full and Tu, 1991; Farley et al., 1993). Blickhan (1989) and McMahon and Cheng (1990) introduced a simple spring-mass model to achieve these patterns for the ground reaction forces. This representation of the leg by a linear spring successfully described and predicted animal locomotion (Blickhan and Full, 1993; Farley et al., 1993; Deng et al., 2012; Tanase et al., 2015; Gan et al., 2016). For example, Gan et al. (2016) reproduced three different gaits (walk, trot, and tölt) of horses by using a quadrupedal spring-mass model and suggested that different quadrupedal gaits are interpreted as different elastic oscillations. Moreover, passively stable running allows the controller and sensing to be simple, even when there are disturbances (Poulakakis et al., 2006). Therefore, such a simple passive model is useful to investigate the gait selection mechanisms by animals (Tanase et al., 2015; Kamimura et al., 2021). However, actual animals lose kinetic energy by collisions of their legs with the ground and by dissipation via friction and compensate for this loss by their muscles. Energy efficiency is an important factor for animal gait (Ruina et al., 2005; Chatzakos and Papadopoulos, 2009; Cao and Poulakakis, 2015; Polet and Bertram, 2019; Polet, 2021). We would like to introduce the elements for energy dissipation and generation in order to obtain a deeper understanding of the running mechanism in animals in the future.
In addition to the CoM offset and pitch moment of inertia, different characteristics between the fore and hind legs could also influence the running dynamics. For example, the muscle mass of the hind legs is greater than that of the fore legs in horses, and it has been suggested that the main role of the fore legs is to support the body weight, whereas that of the hind legs is to generate driving forces (Payne et al., 2005; Crook et al., 2008). Therefore, future investigations of the effects of different characteristics of the legs would be useful for a better understanding of the relationship between the body structure and running in animals.
In the present study, we used the physical parameters estimated in horses to discuss the gait selection by horses. Physical parameters, such as body weight, moment of inertia, and leg length, vary between species. Different parameters could influence the gait preference. We would like to investigate gait selection by animals other than horses in order to clarify the mechanisms for difference gaits between species in future studies.
Data Availability Statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Author Contributions
SA and FM contributed to the design of this study. TY conducted the numerical simulation and analysis in consultation with SA, MA, TK, YH, NW, KT, and FM. TY and SA wrote the manuscript, and all of the authors reviewed and approved the manuscript.
Funding
This study was supported in part by JSPS KAKENHI Grant Number JP20H00229 and JST FOREST Program Grant Number JPMJFR2021.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fbioe.2022.825157/full#supplementary-material
References
Alexander, R. M., Bennett, M. B., and Ker, R. F. (1986). Mechanical Properties and Function of the Paw Pads of Some Mammals. J. Zoolog. 209 (3), 405–419. doi:10.1111/j.1469-7998.1986.tb03601.x
Ben-Amotz, R., Dycus, D., Levine, D., Arruda, A. G., Fagan, N., and Marcellin-Little, D. (2020). Stance and Weight Distribution after Tibial Plateau Leveling Osteotomy in Forelimb and Hind Limb Amputee Dogs. BMC Vet. Res. 16, 188. doi:10.1186/s12917-020-02402-7
Berkemeier, M. D. (1998). Modeling the Dynamics of Quadrupedal Running. Int. J. Robotics Res. 17 (9), 971–985. doi:10.1177/027836499801700905
Bertram, J. E. A., and Gutmann, A. (2008). Motions of the Running Horse and Cheetah Revisited: Fundamental Mechanics of the Transverse and Rotary Gallop. J. R. Soc. Interf. 6 (35), 549–559. doi:10.1098/rsif.2008.0328
Biancardi, C. M., and Minetti, A. E. (2012). Biomechanical Determinants of Transverse and Rotary Gallop in Cursorial Mammals. J. Exp. Biol. 215 (23), 4144–4156. doi:10.1242/jeb.073031
Bigalike, R. (1972). Observations on the Behaviour and Feeding Habits of the Springbok, Antidorcas marsupialis. Afr. Zool. 7 (1), 333–359.
Blickhan, R. (1989). The spring-mass Model for Running and Hopping. J. Biomech. 22 (11-12), 1217–1227. doi:10.1016/0021-9290(89)90224-8
Blickhan, R., and Full, R. (1993). Similarity in Multilegged Locomotion: Bouncing like a Monopode. J. Comp. Physiol. A. 173 (5), 509–517. doi:10.1007/bf00197760
Buchner, H. H. F., Savelberg, H. H. C. M., Schamhardt, H. C., and Barneveld, A. (1997). Inertial Properties of Dutch Warmblood Horses. J. Biomech. 30 (6), 653–658. doi:10.1016/s0021-9290(97)00005-5
Cao, Q., and Poulakakis, I. (2015). On the Energetics of Quadrupedal Running: Predicting the Metabolic Cost of Transport via a Flexible-Torso Model. Bioinspir. Biomim. 10 (5), 056008. doi:10.1088/1748-3190/10/5/056008
Chatzakos, P., and Papadopoulos, E. (2009). Bio-inspired Design of Electrically-Driven Bounding Quadrupeds via Parametric Analysis. Mechanism Machine Theor. 44, 559–579. doi:10.1016/j.mechmachtheory.2008.08.007
Chen, D., Gong, C., Xing, F., Zhou, C., Qi, M., and Wang, L. (2019). The Effect of Head Movement on the Bounding Gait of a Quadruped Robot with an Active Spine. Adv. Mech. Eng. 11 (9), 1–13. doi:10.1177/1687814019876184
Crook, T. C., Cruickshank, S. E., McGowan, C. M., Stubbs, N., Wakeling, J. M., Wilson, A. M., et al. (2008). Comparative Anatomy and Muscle Architecture of Selected Hind Limb Muscles in the Quarter Horse and Arab. J. Anat. 212 (2), 144–152. doi:10.1111/j.1469-7580.2007.00848.x
De, A., and Koditschek, D. E. (2018). Vertical Hopper Compositions for Preflexive and Feedback-Stabilized Quadrupedal Bounding, Pacing, Pronking, and Trotting. Int. J. Robotics Res. 37 (7), 743–778. doi:10.1177/0278364918779874
Deng, Q., Wang, S., Xu, W., Mo, J., and Liang, Q. (2012). Quasi Passive Bounding of a Quadruped Model with Articulated Spine. Mechanism Machine Theor. 52, 232–242. doi:10.1016/j.mechmachtheory.2012.02.003
Dunbar, D. C., Macpherson, J. M., Simmons, R. W., and Zarcades, A. (2008). Stabilization and Mobility of the Head, Neck and Trunk in Horses during Overground Locomotion: Comparisons with Humans and Other Primates. J. Exp. Biol. 211 (24), 3889–3907. doi:10.1242/jeb.020578
Farley, C. T., Glasheen, J., and McMahon, T. A. (1993). Running Springs: Speed and Animal Size. J. Exp. Biol. 185, 71–86. doi:10.1242/jeb.185.1.71
FitzGibbon, C. D., and Fanshawe, J. H. (1988). Stotting in Thomson's Gazelles: an Honest Signal of Condition. Behav. Ecol. Sociobiol. 23 (2), 69–74. doi:10.1007/bf00299889
Full, R. J., and Tu, M. S. (1991). Mechanics of a Rapid Running Insect: Two-, Four- and Six-Legged Locomotion. J. Exp. Biol. 156 (1), 215–231. doi:10.1242/jeb.156.1.215
Gan, Z., Wiestner, T., Weishaupt, M. A., Waldern, N. M., and David Remy, C. (2016). Passive Dynamics Explain Quadrupedal Walking, Trotting, and Tölting. J. Comput. Nonlinear Dyn. 11 (2), 0210081–2100812. doi:10.1115/1.4030622
Grossi, B., and Canals, M. (2010). Comparison of the Morphology of the Limbs of Juvenile and Adult Horses (Equus Caballus) and Their Implications on the Locomotor Biomechanics. J. Exp. Zool A. Ecol. Genet. Physiol. 313 (5), 292–300. doi:10.1002/jez.598
Hildebrand, M. (1977). Analysis of Asymmetrical Gaits. J. Mammalogy 58 (2), 131–156. doi:10.2307/1379571
Hildebrand, M. (1989). The Quadrupedal Gaits of Vertebrates. BioScience 39 (11), 766–775. doi:10.2307/1311182
Hoyt, D. F., and Taylor, C. R. (1981). Gait and the Energetics of Locomotion in Horses. Nature 292 (5820), 239–240. doi:10.1038/292239a0
Hudson, P. E., Corr, S. A., and Wilson, A. M. (2012). High Speed Galloping in the Cheetah (Acinonyx Jubatus) and the Racing Greyhound (Canis familiaris): Spatio-Temporal and Kinetic Characteristics. J. Exp. Biol. 215 (14), 2425–2434. doi:10.1242/jeb.066720
Kamimura, T., Aoi, S., Higurashi, Y., Wada, N., Tsuchiya, K., and Matsuno, F. (2021). Dynamical Determinants Enabling Two Different Types of Flight in Cheetah Gallop to Enhance Speed through Spine Movement. Sci. Rep. 11 (1), 9631. doi:10.1038/s41598-021-88879-0
Lee, D. V., Stakebake, E. F., Walter, R. M., and Carrier, D. R. (2004). Effects of Mass Distribution on the Mechanics of Level Trotting in Dogs. J. Exp. Biol. 207 (10), 1715–1728. doi:10.1242/jeb.00947
McMahon, T. A., and Cheng, G. C. (1990). The Mechanics of Running: How Does Stiffness Couple with Speed? J. Biomech. 23 (1), 65–78. doi:10.1016/0021-9290(90)90042-2
Minetti, A. E., Ardigò, L. P., Reinach, E., and Saibene, F. (1999). The Relationship between Mechanical Work and Energy Expenditure of Locomotion in Horses. J. Exp. Biol. 202 (17), 2329–2338. doi:10.1242/jeb.202.17.2329
Payne, R. C., Veenman, P., and Wilson, A. M. (2005). The Role of the Extrinsic Thoracic Limb Muscles in Equine Locomotion. J. Anat. 206 (2), 193–204. doi:10.1111/j.1469-7580.2005.00353.x
Pennycuick, C. J. (1975). On the Running of the Gnu (Connochaetes Taurinus) and Other Animals. J. Exp. Biol. 63, 775–799. doi:10.1242/jeb.63.3.775
Polet, D. T. (2021). The Murphy Number: How Pitch Moment of Inertia Dictates Quadrupedal Walking and Running Energetics. J. Exp. Biol. 224, jeb228296. doi:10.1242/jeb.228296
Polet, D. T., and Bertram, J. E. A. (2019). An Inelastic Quadrupedal Model Discovers Four-Beat Walking, Two-Beat Running, and Pseudo-elastic Actuation as Energetically Optimal. Plos Comput. Biol. 15 (11), e1007444. doi:10.1371/journal.pcbi.1007444
Poulakakis, I., Papadopoulos, E., and Buehler, M. (2006). On the Stability of the Passive Dynamics of Quadrupedal Running with a Bounding Gait. Int. J. Robotics Res. 25 (7), 669–687. doi:10.1177/0278364906066768
Ruina, A., Bertram, J. E. A., and Srinivasan, M. (2005). A Collisional Model of the Energetic Cost of Support Work Qualitatively Explains Leg Sequencing in Walking and Galloping, Pseudo-elastic Leg Behavior in Running and the Walk-To-Run Transition. J. Theor. Biol. 237 (2), 170–192. doi:10.1016/j.jtbi.2005.04.004
Self Davies, Z. T., Spence, A. J., and Wilson, A. M. (2019). Ground Reaction Forces of Overground Galloping in Ridden Thoroughbred Racehorses. J. Exp. Biol. 222 (16), jeb204107. doi:10.1242/jeb.204107
Swanstrom, M. D., Zarucco, L., Hubbard, M., Stover, S. M., and Hawkins, D. A. (2005). Musculoskeletal Modeling and Dynamic Simulation of the Thoroughbred Equine Forelimb during Stance Phase of the Gallop. J. Biomech. Eng. 127 (2), 318–328. doi:10.1115/1.1865196
Tanase, M., Ambe, Y., Aoi, S., and Matsuno, F. (2015). A Galloping Quadruped Model Using Left-Right Asymmetry in Touchdown Angles. J. Biomech. 48 (12), 3383–3389. doi:10.1016/j.jbiomech.2015.06.003
Keywords: horse, transverse gallop, center of mass offset, gait selection, model
Citation: Yamada T, Aoi S, Adachi M, Kamimura T, Higurashi Y, Wada N, Tsuchiya K and Matsuno F (2022) Center of Mass Offset Enhances the Selection of Transverse Gallop in High-Speed Running by Horses: A Modeling Study. Front. Bioeng. Biotechnol. 10:825157. doi: 10.3389/fbioe.2022.825157
Received: 30 November 2021; Accepted: 11 January 2022;
Published: 28 February 2022.
Edited by:
John R. Hutchinson, Royal Veterinary College (RVC), United KingdomReviewed by:
John E. A. Bertram, University of Calgary, CanadaMichael Peterson, University of Kentucky, United States
Copyright © 2022 Yamada, Aoi, Adachi, Kamimura, Higurashi, Wada, Tsuchiya and Matsuno. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shinya Aoi, c2hpbnlhX2FvaUBrdWFlcm8ua3lvdG8tdS5hYy5qcA==