 Markian S. Bahniuk
Markian S. Bahniuk Fawad Alidina
Fawad Alidina Xiaoli Tan
Xiaoli Tan Larry D. Unsworth
Larry D. Unsworth- Department of Chemical and Materials Engineering, University of Alberta, Edmonton, AB, Canada
The generation of kaolin-containing wastewater is an inevitable consequence in a number of industries including mining, wastewater treatment, and bitumen processing. In some cases, the production of kaolin tailings waste during the production of bitumen or phosphate is as high as 3 times greater than the actual produced product. The existing inventory of nearly five billion barrels of oil sands tailings alone represents a massive storage and reclamation challenge, as well as a significant economic and environmental liability. Current reclamation options like inorganic coagulants and organic synthetic polymers may settle kaolin effectively, but may themselves pose an additional environmental hazard. Bioflocculants are an emerging alternative, given the inherent safety and biodegradability of their bio-based compositions. This review summarizes the different research attempts towards a better bioflocculant of kaolin, with a focus on the bioflocculant source, composition, and effective flocculating conditions. Bacillus bacteria were the most prevalent single species for bioflocculant production, with wastewater also hosting a large number of bioflocculant-producing microorganisms while serving as an inexpensive nutrient. Effective kaolin flocculation could be obtained over a broad range of pH values (1–12) and temperatures (5–95°C). Uronic acid and glutamic acid were predominant sugars and amino acids, respectively, in a number of effective bioflocculants, potentially due to their structural and charge similarities to effective synthetic polymers like polyacrylamide. Overall, these results demonstrate that bioflocculants can be produced from a wide range of microorganisms, can be composed of polysaccharides, protein or glycoproteins and can serve as effective treatment options for kaolin. In some cases, the next obstacle to their wide-spread application is scaling to industrially relevant volumes and their deployment strategies.
1 Introduction
The production of clay-containing wastewater tailings is an inevitable consequence of a wide variety of industries including mining and mineral processing (Wang et al., 2014; Shaikh et al., 2017; Liu et al., 2020). Four to six tons of phosphogypsum, phosphate tailings, are produced per ton of fertilizer: 100–280 million tons of phosphate tailings are estimated to be produced annually (Anagnostopoulas et al., 2022). Similarly, bitumen production from oil sands results in 3 times as much tailings as product, with a current daily production of ∼3 million barrels of tailings being added to the existing inventory of ∼4.8 billion barrels of mature fine tailings (Masliyah et al., 2011; Parajulee and Wania, 2014; Schindler, 2014). Tailings from phosphates, coal, and oil sands extraction are primarily kaolinite and are the impetus for kaolin-focused clay flocculation studies (Sabah et al., 2004; Ofori et al., 2011; Wang et al., 2014). Kaolin fine fluid tailings are particularly difficult to treat due to the fact that clay particles are highly charged and can remain colloidally stable for centuries, particularly as the particle size decreases (Hocking et al., 1999; Bergaya and Lagaly, 2006; Wang et al., 2014). Management of these tailings requires massive storage solutions, sequesters significant volumes of water along with the kaolin, and represents significant potential fiscal and environmental liabilities.
Coagulants and flocculants have been applied to try to solve the technical and environmental challenges posed by the management of tailings. Multiple flocculating mechanisms have been described for destabilizing colloidal clay dispersions: electrical double layer compression, charge neutralization and polymer bridging, sweep flocculation, and the electrostatic patch mechanism (Hocking et al., 1999; Shaikh et al., 2017). Furthermore, these flocculation mechanisms and the specific interactions between different types of flocculants and kaolin particles have both been extensively documented in established literature. (Hocking et al., 1999; Shaikh et al., 2017; Liu et al., 2020). While inorganic metal salt coagulants and synthetic polymers exhibit strong flocculating activity (FA) to settle suspended clays, there are additional complications associated with their application. Inorganic metal salts like ferric chloride, lime, alum, and polyaluminum chloride do not form strong flocs nor easily dewater and can be highly corrosive (Hocking et al., 1999; Pan et al., 1999; Salehizadeh and Shojaosadati, 2001; Salehizadeh and Shojaosadati, 2002; More et al., 2012; Busi et al., 2017). Polymers like polyacrylamide have been shown to have strong FA but multiple mechanisms for their degradation have been established, with iron, oxygen, or photolytic based free radical-mediated chain scission being the most relevant for tailings applications (Xiong et al., 2018). The environmental risk associated with monomeric acrylamide may be attenuated, however, given that acrylamide may also be further degraded by a range of soil and water microbes (Smith et al., 1997; Kay-Shoemake et al., 1998; Shukor et al., 2009; Labahn et al., 2010).
Bioflocculants are an emerging class of flocculant that have the potential to combine the efficacy of conventional synthetic polymer flocculants with the inherent biodegradability and long-term safety of bio-derived materials. Bioflocculants are composed of a range of biological building blocks: polysaccharides, proteins, nucleic acids, lipids, and combinations thereof (Yokoi et al., 1997; Zhao et al., 2013; Busi et al., 2017). Bioflocculants can be produced by a wide range of microorganisms including bacteria, fungi, and algae, typically as extracellular bio-polymeric substances, though there are studies characterizing intracellular and plant-based bioflocculants (Yim et al., 2007; Bala Subramanian et al., 2010; Liu et al., 2010; Pu et al., 2014; Choy et al., 2015). Current hurdles for the widespread adoption of bioflocculants include purported high production cost of purified flocculants and relatively low FA where
and uses the control (B) and sample (A) optical density. (Kurane et al., 1986) Herein, the different research attempts from the last 25 years towards a better bioflocculant of kaolin are summarized. Flocculation parameters leading to an effective (FA ≥80%) settling of kaolin, including temperature, pH, metal-ion supplements, and bioflocculant concentration, are reported. Studies will be generally organized based on the bacterial genera producing the bioflocculant, or environment in which the bioflocculant-producing microorganism is found.
2 Bioflocculant-producing organisms and sources
2.1 Bacillus
Bacillus bacteria are the most frequently studied bioflocculant-producing microorganism. Bacillus are well-characterized, Gram-positive, aerobic or facultatively anaerobic, endospore-forming bacteria (Baron, 1996). Bacillus occur ubiquitously in soil, with species capable of existing in extreme environments including at high temperatures, salt concentrations, and pH conditions (Liu et al., 2014a; Robinson, 2014). Bacillus are well suited for bioflocculant production as they produce and excrete considerable amounts of enzymes, metabolites, and antibiotics and are employed in a host of industrial and pharmaceutical processes (Takami and Horikoshi, 2000).
The majority of the Bacillus-derived bioflocculants (Table 1) are primarily (>50%) polysaccharides, though there are 5 studies which examined protein bioflocculants. Of those, three were found to be polyglutamic acid, a known component of the Bacillus capsule (Lee et al., 2014; Lee et al., 2017; Ajayeoba et al., 2019). Bacillus-derived bioflocculants proved to be effective in an extremely wide pH range, with individual studies showing optimal performance at pH values between 1 and 12. This versatility allows for the use of these bioflocculants in industrial processes at acidic or alkaline conditions without requiring neutralization, streamlining treatment processes and costs.
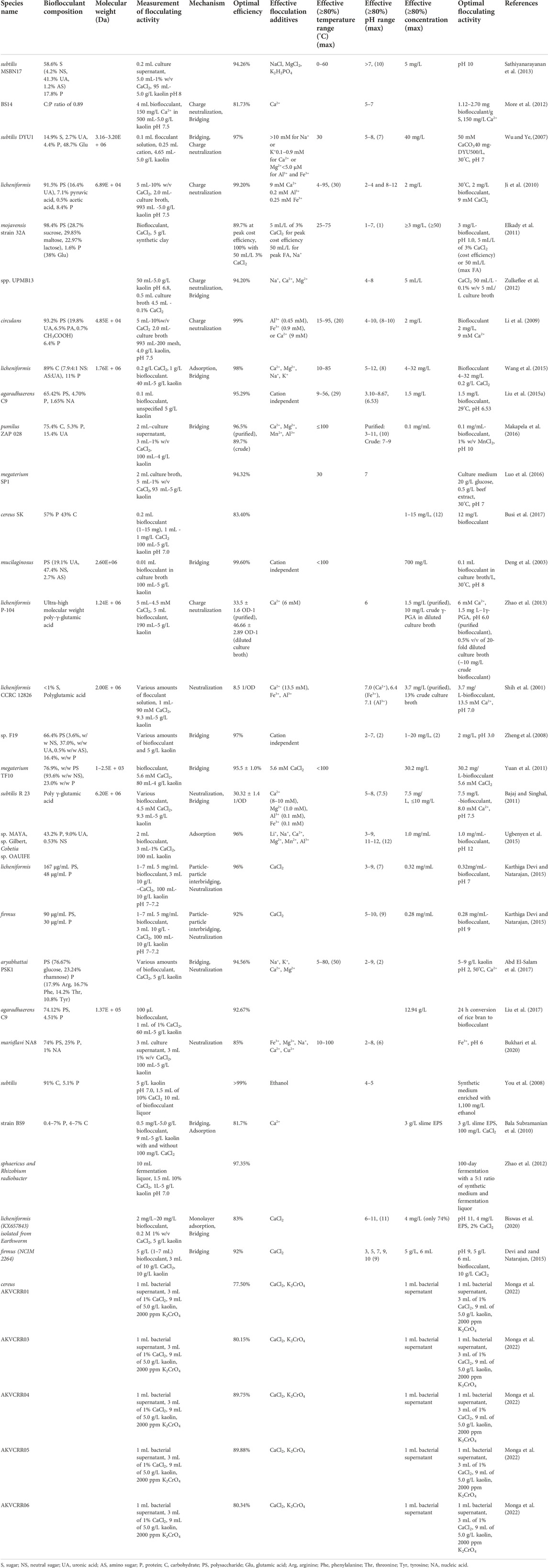
TABLE 1. Bacillus genera and their bioflocculant performance.
In addition to their general acceptance in industrial processes, these bioflocculants exhibit a very high flocculating activity (FA). For instance, Bacillus mucilaginosus produced a bioflocculant with 99.6% FA using 0.1 mL culture broth/L at pH 8 and 30°C, while retaining good FA at temperatures as high as 100°C (Deng et al., 2003). Bacillus circulans produced a bioflocculant with an optimal FA of 99% using 2 mg/L bioflocculant and 9 mM Ca2+, while retaining good FA at temperatures as high as 95°C (Li et al., 2009) Bacillus subtilis produced a bioflocculant capable of 99.7% FA using a synthetic medium enriched with 1,100 mg/L ethanol (You et al., 2008).
2.2 Pseudomonas
Pseudomonas are rod-shaped, aerobic Gram-negative bacteria with a polar flagella for motility that are also commonly used for bioflocculant production (Madigan et al., 2008). As a result of their metabolic versatility, they can be found in soil, water and vegetation (Baron, 1996). This versatility allows for the use of a variety of organic compounds as nutrients, as well as survival in a broad range of temperatures and salts and even in the presence of some weak antiseptics and antibiotics (Baron, 1996). Pseudomonas bacteria are known to produce biofilms by excreting polysaccharides, a quality which is directly applicable for bioflocculation (Hassett et al., 2002).
The studies on Pseudomonas-derived bioflocculants (Table 2) all used kaolin test solutions of 4–5 g/L, allowing straightforward comparisons with Bacillus results. Calcium was the most commonly used cation, though two studies showed that monovalent cations could also be used for effective flocculation (Gomaa, 2012; Farag et al., 2014). As with Bacillus bioflocculants, polysaccharides were the most prominent structural element, though most were composed of some protein as well. Of note is the bioflocculant produced by Pseudomonas Sp. HP2 because of the large proportion of proline, glutamic, and aspartic acids found in the considerable proteinaceous component (Qi et al., 2019). Glutamic and aspartic acid sidechains differ by a single methylene group, and polyglutamic acids were found to be effective bioflocculants produced by Bacillus bacteria. The frequency of proline (25.5% of all amino acids) is very high and may influence the protein secondary structure away from conventional α-helices and β-sheets and toward intrinsically disordered structures and/or polyproline helices (Morgan and Rubenstein, 2013; Bahniuk et al., 2020). Three of the Pseudomonas studies attempt to address the economic concerns associated with bioflocculant production by utilizing alternative nutrient sources, driving down production costs while maintaining FA >90% (Liu et al., 2016; Guo et al., 2018a; Qi et al., 2019).
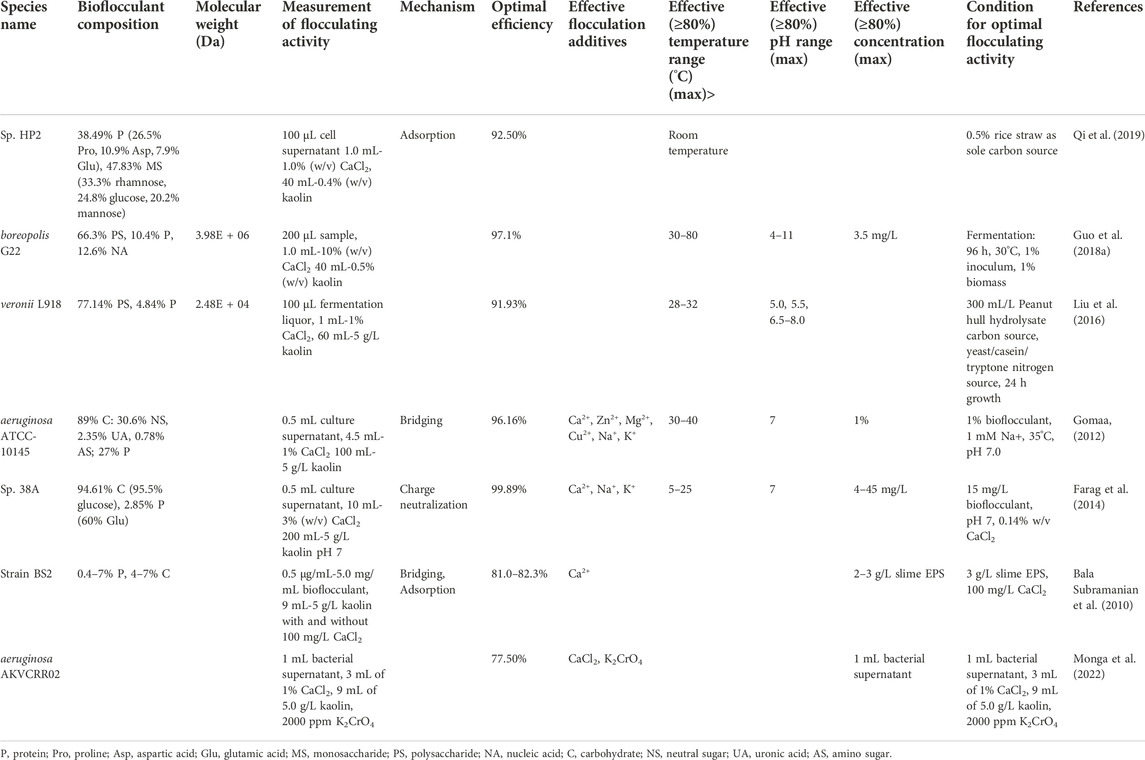
TABLE 2. Pseudomonas genera and their bioflocculation performance.
Pseudomonas boreopolis G22 produced a bioflocculant capable of an optimal FA of 97.1% at 60°C after 4 days of fermentation using wood dust as an inexpensive carbon source (Guo et al., 2018a). Pseudomonas Sp. 38A produced a bioflocculant with an optimal FA of 99.89% at pH 7.0 when 15 mg/L bioflocculant and 0.14% w/v CaCl2 were used (Farag et al., 2014). Pseudomonas aeruginosa produced a bioflocculant with an optimal FA of 96.16% when monovalent Na+ ions were included (Eman Zakaria, 2012).
2.3 Wastewater bacteria
Wastewater contains a diverse population of bacteria that can serve as a source for the discovery of novel bacteria capable of efficient bioflocculation, as well as an inexpensive and ubiquitous source of nutrition for bioflocculant-producing bacterial cultures and a host of potential pollutants including suspended wastewater solids, estrogen, arsinite, dyes, cadmium and other heavy metals (Liu et al., 2009; Zhong et al., 2014a; Cao et al., 2015; Chen et al., 2016). The use of kaolin to screen and initially assess bioflocculant performance is warranted (Table 3), given the zeta potential values for wastewater suspended solids is comparable to that of kaolin at ∼ -32 mV (Nguyen et al., 2016). Table 3 summarizes the physical characteristics and flocculation performance of bioflocculants produced by wastewater-derived bacteria.
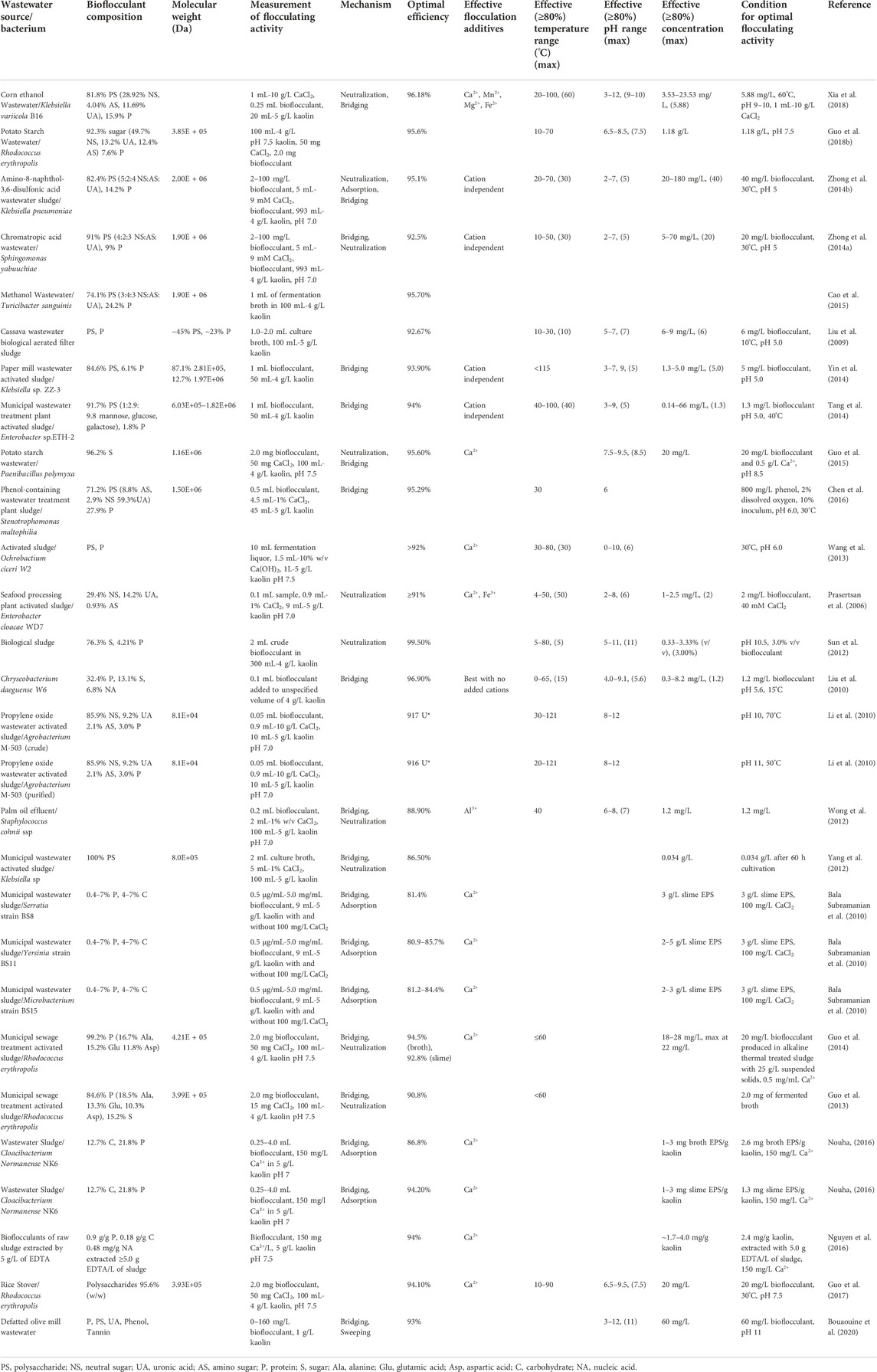
TABLE 3. Bacterium from wastewater and their flocculation performance.
A wide range of wastewater bacteria were studied, with Rhodococcus producing four bioflocculants (Guo et al., 2013; Guo et al., 2014; Guo et al., 2017; Guo et al., 2018b). Most of the bioflocculants summarized in the table performed well at relatively low concentrations. The majority were stable up to 60°C due to being primarily polysaccharide-based, though some were found to be primarily composed of protein (Liu et al., 2010; Guo et al., 2013; Guo et al., 2014) or lipids and/or nucleic acids (Bala Subramanian et al., 2010; Nouha, 2016). The localization of bioflocculant molecules was examined, in some instances, by quantifying the different FA of slime and capsular culture fractions to determine whether the bioflocculants themselves were extracellular or cell-bound, respectively, or identified as being an intracellular protein (Bala Subramanian et al., 2010; Liu et al., 2010; Guo et al., 2013; Guo et al., 2014; Nouha, 2016).
Some of the most efficient bioflocculation results came from the use of a biological sludge-derived bioflocculant, yielding an FA of 99.5% when the suspension was maintained at pH 10.5 and dosed with a concentration of bioflocculant at 3.0% v/v (Sun et al., 2012). Chryseobacterium daeguense W6 produced a bioflocculant with an optimal FA of 96.9% by maintaining the suspension at pH 5.6 and 15°C, and then dosed with 1.2 mg/L bioflocculant (Liu et al., 2010). Finally, corn ethanol wastewater was a low cost substrate to produce a bioflocculant with optimal FA of 96.18%, when suspension was maintained at pH 8.1 and 30°C (Xia et al., 2018).
2.4 Plant-derived bioflocculants
Bacteria are not the only source of bioflocculating materials. Plants can be employed directly as bioflocculants (Table 4), in addition to serving as nutrients for bacteria. Ancient African, Indian, and Chinese cultures were documented practicing plant-mediated bioflocculation ∼2000 years ago, particularly for the purification of drinking water (Asrafuzzaman et al., 2011; Choy et al., 2015). The opportunity to use readily available, local florae is advantageous relative to bacteria in that no significant manufacturing infrastructure, beyond farmland, is necessary to produce plant-based bioflocculants. The availability of these materials for water purification is critical in rural locations, and municipalities where local infrastructure is lacking or overwhelmed.
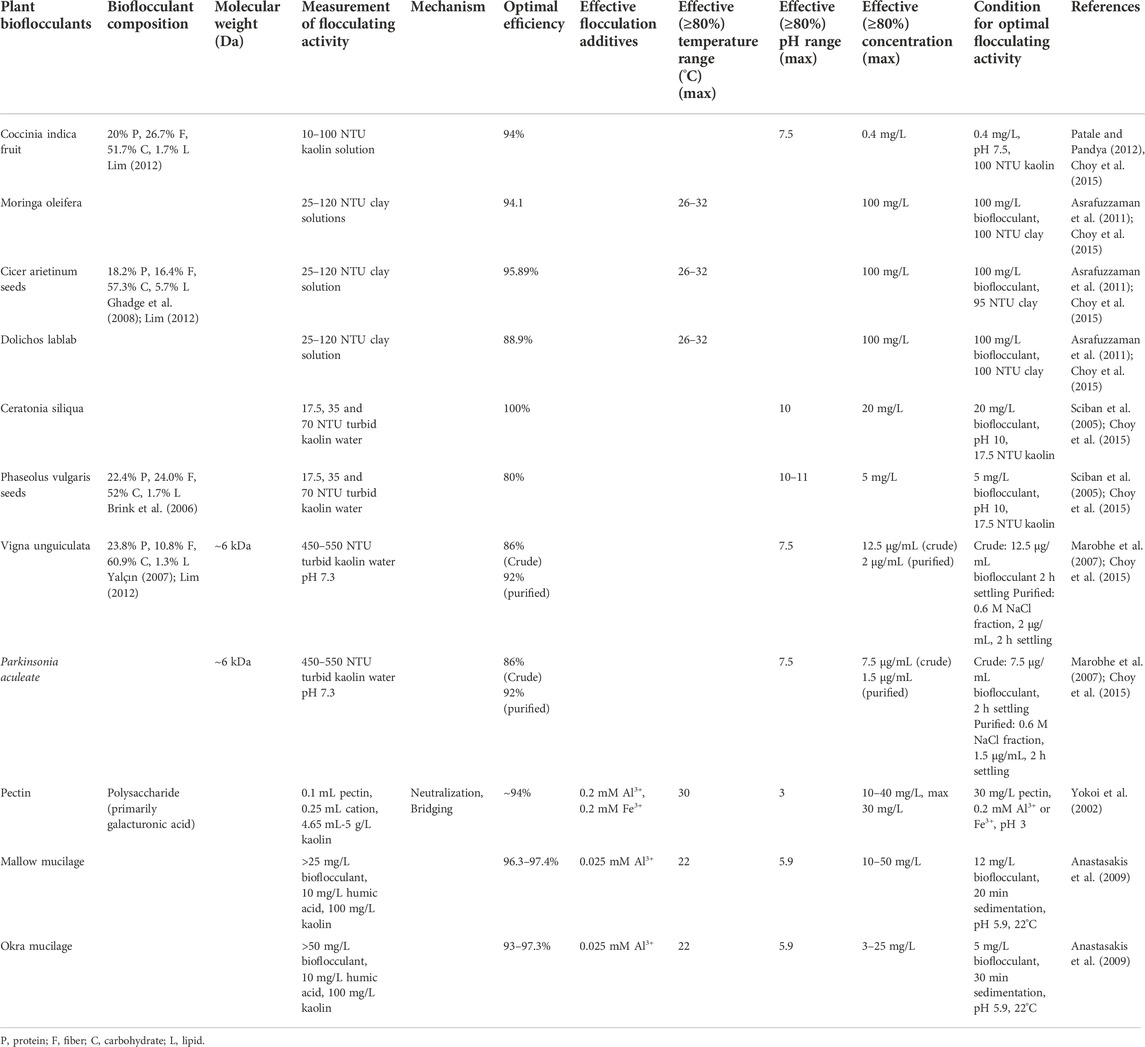
TABLE 4. Plant-derived bioflocculants.
The studies summarized herein focus more on preliminary confirmation of bioflocculation rather than FA optimization or purification of specific entities from the bulk plant material. Additionally, some of these studies utilized various concentrations of suspended clays, based upon nephelometric turbidity units (NTUs) rather than weight percentage; some instances of optimal activity depend upon the initial water NTUs, as detailed below. FA was calculated in numerous ways, including Eq. 1, a modified Lee’s equation, as well as a method proposed by Toeda (Toeda and Kurane, 1991; Lee et al., 1995). The modified Lee’s equation to calculate FA is
where AS represents the sample absorbance, AC represents the control absorbance, t0 is the time immediately after sample homogenization and t1 is a specific time after homogenization (Lee et al., 1995; Marobhe et al., 2007). Toeda’s method is
where A and B represent the sample and reference optical densities, respectively. (Toeda and Kurane, 1991) These results have been converted to percentages when necessary, to allow for more meaningful comparisons with results based on Eq. 1.
Numerous plant-derived bioflocculants showed very strong FA. Mallow mucilage had an optimal FA range of 96.3%–97.4% in synthetic water, which contained 100 mg/L kaolin and 10 mg/L humic acid. This was achieved using 12–26 mg/L of mucilage at pH 7 (Anastasakis et al., 2009). Okra mucilage had an optimal FA range of 93–97.3% under the same conditions but required one-half to one-fifth the dosage (Anastasakis et al., 2009). Widely available pectin was found to have an FA of ∼94% when 12 mg/L was combined with clay and 0.2 mM Al3+ or Fe3+ at pH 3.0 (Yokoi et al., 2002).
2.5 Chemically modified bioflocculants
For the purposes of this review, chemically modified bioflocculants refer to two types of materials: 1) Bioflocculants hybridized with conventional organic flocculants, and 2) Bioflocculants chemically treated to polymerize them without the addition of any other materials. These hybrid materials attempt to combine the safety and biodegradability of bioflocculants with the performance and increased solubility of conventional flocculants. Hybridization can bestow significant cationic charge to bioflocculants via quaternary ammonium groups, like 3-chloro-2-hydroxypropyl trimethyl ammonium chloride (Lu et al., 2011). This hybrid approach is not without its drawbacks, given the increased processing costs associated with chemical modification and the potential decrease in environmental safety with the inclusion of potentially harmful polymers like polyacrylamide.
All of the reviewed studies (Table 5) utilized polysaccharides as their base material due to the copious reactive sites available. Clay solutions utilized were of lower concentration than the Bacillus, Pseudomonas, and wastewater studies and were more in line with the plant-based bioflocculation tests, with some studies focusing on NTU-based analysis (Lu et al., 2011). Kaolin clay was not the only type of clay employed, with one study using oppositely charged hematite as well as kaolin in order to demonstrate the importance of chemically modified bioflocculant charge on FA (Li et al., 2015). In addition to examining the clay flocculation performance of chemically modified bioflocculants, some of the articles attempt to optimize the material FA by varying graft ratios and degrees of substitution (Ghimici et al., 2010; Lu et al., 2011; Li et al., 2015).
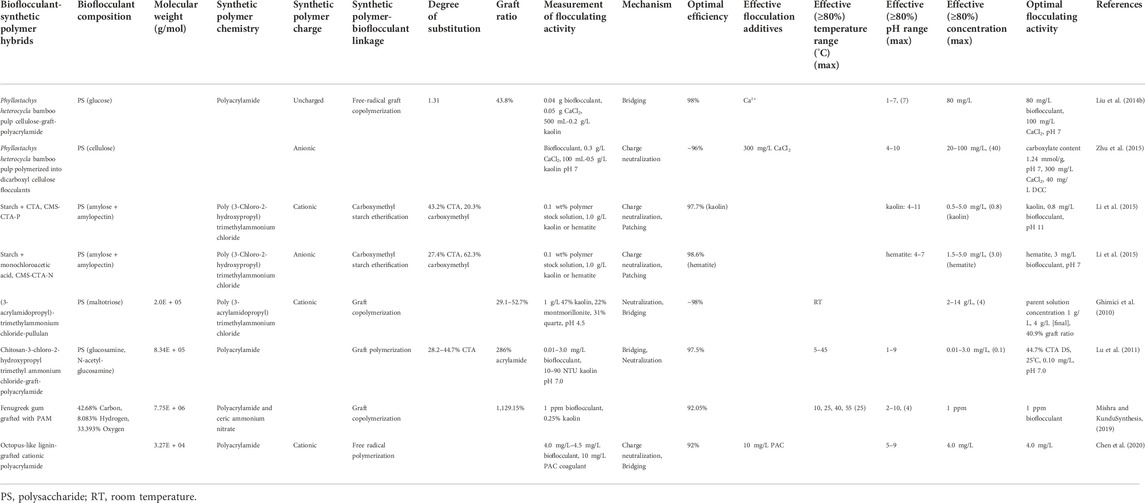
TABLE 5. Chemically modified bioflocculants and their flocculation performance.
Bamboo pulp cellulose-grafted polyacrylamide was found to have a very high FA of 98%. Suspension was dosed with 0.04 g of bioflocculant and 0.05 g CaCl2 at pH 7.0 (Liu et al., 2014b). When (3-acrylamidopropyl)-trimethylammonium chloride and pullulan were grafted, ∼98% FA was achieved at the highest graft ratio (Ghimici et al., 2010). Artificially polymerized bamboo pulp cellulose had an FA of ∼96% at 40 mg/L DCC with 300 mg/L of CaCl2 at pH 7 (Zhu et al., 2015).
2.6 Soil bacteria and fungi
Bacillus bacteria are not the only soil bacteria known to produce effective bioflocculants. There are numerous other bacterial genera, as well as some fungi, currently being studied. This diversity in the source of bioflocculant-producing microorganisms will naturally lead to a greater variety of bioflocculant molecules, production conditions, and ideal flocculating environments suitable for a range of industrial processes. Another bioflocculation study with broad industrial potential involved supplementing Agrobacterium tumefaciens cultures with a quorum sensing signal molecule to directly induce greater bioflocculant production without increasing the bacterial population density (Yang et al., 2016). This unusual but effective approach could be broadly applicable to various large-scale fermentation processes to increase yields and drive down costs.
The majority of the bioflocculants observed for these groups were primarily polysaccharides (Table 6). Only one bioflocculant composed of >50% protein, produced by an Arthrobacter, was found (Mabinya et al., 2012). Unlike the majority of the plant-derived bioflocculation tests, kaolin test solutions were of higher concentration, in the standard range of 4–5 g/L. The effective and optimal pH ranges for bioflocculants from soil bacteria and fungi tended to be acidic, in the range of 5-7.
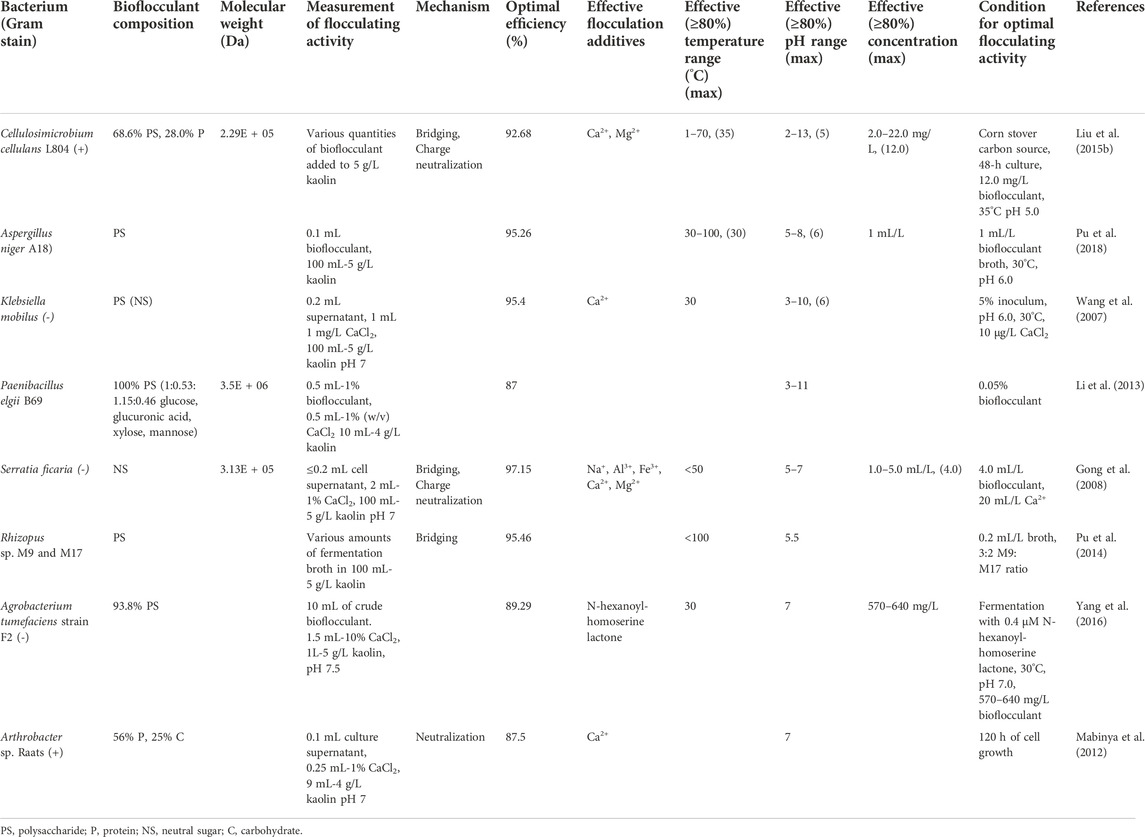
TABLE 6. Soil bacteria and fungi and their flocculation performance.
Soil bacteria and fungi yielded some very high performing bioflocculants. Serratia ficaria produced a bioflocculant with an optimal FA of 97.15% when cultivated after 72 h with lactose as the carbon source (Gong et al., 2008). A coculture of two strains of Rhizopus sp., M9 and M17, produced a bioflocculant having an optimal FA of 95.46% when dosed with 0.2 mL fermentation broth/L kaolin suspension (Pu et al., 2014). Klebsiella mobilus produced a bioflocculant with a peak FA of 95.4% with an initial inoculum volume of 5% was used and the flocculation was carried out at 30°C, pH 6.0 in the presence of 10 μg/L CaCl2 (Wang et al., 2007).
2.7 Aquatic bacteria and algae
Novel bioflocculant-producing microorganisms are not limited to those found in soil. Numerous studies have identified aquatic bacteria and algae capable of producing bioflocculants with considerable flocculating activity. The significant variations in the natural environment of aquatic bacteria compared to those found in soil further broaden the diversity of microorganisms and molecules capable of efficient bioflocculation. The use of algae for bioflocculant production, demonstrated using Gyrodinium impudicum, exemplifies the benefits of diversification as there may be economic benefits by simultaneously using the algae for the production of bioflocculants and algae-related food, oil or pigments (Xia et al., 2018). The inclusion of two Halmonas species also exemplifies the benefits of diversification as these deep-sea bacteria are low temperature and salt tolerant, meaning a wider range of culture and effective bioflocculating conditions can be considered, avoiding potential desalinization and reducing the demands for fresh water (He et al., 2010; Okaiyeto et al., 2013).
All of the bioflocculants produced by aquatic bacteria or algae were primarily polysaccharides, with the algal bioflocculant being highly sulfated (Table 7): a result not seen in any of the other studies reviewed herein (Yim et al., 2007). Kaolin (4–5 g/L) was the test solution for all of these studies, allowing for a direct comparison with the majority of the other results in this review. Cations were necessary in 60% of the studies, with the remaining 40% showing enhanced flocculation in the presence of cations. The effective pH range for flocculation ranged from 4-8.
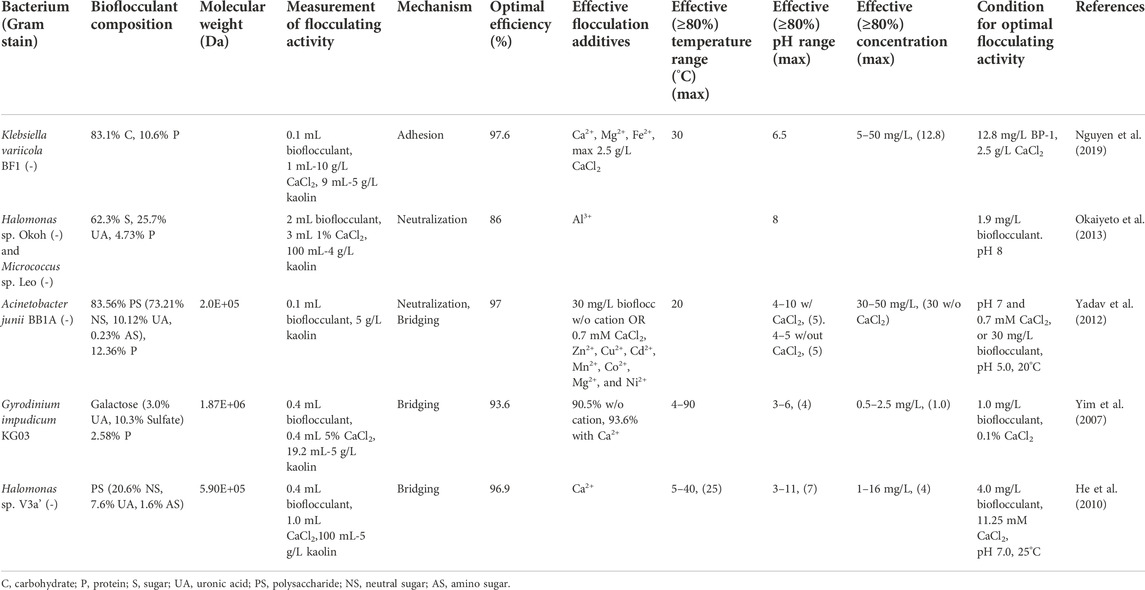
TABLE 7. Aquatic bacteria and algae and their flocculation performance.
Klebsiella variicola BF1 produced a bioflocculant having an optimal FA of 97.6% when suspension was dosed with 12.8 mg/L of bioflocculant and 2.5 g/L of CaCl2 (Nguyen et al., 2019). Acinetobacter junii BB1A produced a bioflocculant having an optimal FA of 97% when suspension was maintained at pH 7 and dosed with 30 mg/L of bioflocculant or 0.7 mM CaCl2 (Yadav et al., 2012). Halomonass sp. V3a’ produced a bioflocculant having an optimal FA of 96.9% when suspension was maintained at pH 7 and 25°C with 11.25 mM Ca2+ and 4 mg of bioflocculant (He et al., 2010).
3 Future directions and conclusion
This review summarizing the different research attempts towards a better bioflocculant of kaolin demonstrates that a wide range of potent bioflocculants and sources have been found, and this distribution of high FA sources and materials limits the opportunity for the formation of a general consensus or direction in the field. These works are still very exploratory, with few studies iterating and improving upon previous works, and/or focusing on kaolin flocculation specifically. In most instances, kaolin solutions are used as a simple screening test to show FA, then the bioflocculants are applied to other sludge or wastewater solutions. This ultimately leaves the field of kaolin bioflocculation underdeveloped, despite the immense quantities of kaolin tailings requiring remediation, including ∼4.8 billion barrels of mature fine tailings from oil sands production (Schindler, 2014; Wang et al., 2014; Liu et al., 2020; Anagnostopoulas et al., 2022). The intention of this review is to highlight some common sources and characteristics of effective kaolin bioflocculants, with a mind towards future technically and economically viable applications of these systems.
Taken as a whole, the studies summarized herein demonstrate that bioflocculants can have significant kaolin FA over a wide range of pH and temperature conditions, alleviating some of the need to bring kaolin solutions to standard pH and/or temperature ranges for efficient flocculation. In fact, optimal FA was observed at pH values ranging from 1–12 and temperatures from 5–80°C. Bacillus-derived bioflocculants tended to show optimal FA over the broadest range of pH values including extremely acidic and alkaline conditions, with wastewater and plant-derived bioflocculants also showing some optimal activity under a greater range of alkaline conditions. Most optimal FA conditions were at temperatures ranging from 20–35°C, but both Bacillus and wastewater-derived bioflocculant categories contained at least five studies where effective FA (≥80%) was achieved at 0–10°C, with Cassava wastewater biological aerated filter sludge and biological sludge exhibiting optimal FAs of 92.67% and 99.5% at 10°C and 5°C, respectively (Liu et al., 2009; Sun et al., 2012). Seven Bacillus-derived bioflocculants were shown to have effective FA (≥80%) at temperatures from 60–95°C, with Bacillus aryabhattai PSK1 having an optimal FA of 94.56% at 50°C (Abd El-Salam et al., 2017). Multiple wastewater-derived Klebsiella, Rhodococcus and Enterobacter bioflocculants also showed effective FAs at high temperatures, with Enterobacter cloacae WD7 having optimal FA of ≥91% at 50°C and Klebsiella variicola B16 peaking at 96.18% FA at 60°C (Prasertsan et al., 2006; Xia et al., 2018).
Of the 73 studies which quantified the composition of the bioflocculants, 53 were primarily polysaccharide, 10 were protein, and 10 were composed of significant proportions of both polysaccharide and protein. The compositions of the polysaccharide portions of the bioflocculants varied, with a range of proportions of neutral sugars, amino sugars and uronic acids. The uronic acid content in particular shifted considerably, from 1.2–59.3% of the total polysaccharide (Sathiyanarayanan et al., 2013; Chen et al., 2016). The uronic acid content is noteworthy because these are sugar derivatives containing carboxylic acid functional groups which are know to participate in kaolin particle attachment (Xia et al., 2018). Carboxylic acid chemistries also make up the sidechains of glutamic acid and aspartic acid, amino acids that were enriched in most of the protein components of the bioflocculants reviewed herein. Three studies found that polyglutamic acids, proteins composed almost exclusively of glutamic acid, were responsible for kaolin bioflocculation (Shih et al., 2001; Bajaj and Singhal, 2011; Zhao et al., 2013). Carboxylic acid functional groups are replete throughout common synthetic polymer flocculants including polyacrylamide, and the structural and charge similarities of acrylamide monomers and uronic acid as well as the amino acids glutamic acid and aspartic acid cannot be overlooked as these chemical similarities may be one of the main driving forces of bioflocculant efficacy (Liu et al., 2020).
While these studies demonstrate the range of bacteria capable of producing effective kaolin bioflocculants, the limitations of these studies must also be addressed. Complete treatment of kaolin tailings requires the dewatering of the settled material, a topic which was only examined in 7 of the manuscripts, primarily regarding the dewatering of sludge rather than kaolin. Dewatering releases bound water, reduces the tailings volume and improves the mechanical strength of the clay, factors necessary for the proper disposal of these materials. Dewatering efficiency can be impacted by the choice of flocculant with, for instance, aluminum-polyacrylamide hybrid polymers leading to rapid dewatering (Wang et al., 2014). The kaolin concentrations used to assess bioflocculation were limited almost exclusively to 5 g/L or less, conditions forty to sixty times more dilute than oil sands tailings. As such, to accurately gauge bioflocculant FA for applications such as treating oil sands tailings, further testing will be necessary as the composition of oil sands tailings is much more complex than just kaolin. Another line of inquiry relevant for successful industrial deployment of bioflocculants is the in situ stability of the flocculating molecules. While some studies did examine bioflocculant pH and temperature stability, the results only demonstrate the retention of FA over a short period of time. Bioflocculant stability in situ over longer periods of time would more accurately reflect real-world applications, and may impact the overall FA and dewatering outcomes. Safety is another limitation that has not been fully addressed. While bioflocculants can be safer alternatives to conventional inorganic and organic polymeric flocculants, the hazards arising from the microorganisms producing the bioflocculants must be kept in mind. For instance, the use of Enterobacter and Pseudomonas aeruginosa for bioflocculant production poses a serious hazard due to their pathogenicity in humans. Furthermore, large scale growth of Gyrodinium or similar algae blooms can lead to toxic conditions towards marine and mammalian life forms, including humans, in addition to potentially inhibiting flocculation (Sano et al., 2011).
Even as studies like those reviewed herein advance the scientific understanding of the effective compositions for bioflocculants, optimization of bioflocculant production, range of bioflocculant-producing microorganisms, and FA, the widespread deployment of bioflocculants needs to overcome the economic hurdles accompanying their use. The vast volumes of kaolin tailings requiring treatment, ∼4.8 billion barrels worth from oil sands production alone dictates that any reasonable treatment solution must be economically, as well as technically, viable. Even the addition of a small amount of CaCl2 represents a significant cost when scaled to industrial proportions, and in the case of recycled water applications can reduce the efficiency of bitumen extraction. Every effort must be made to drive bioflocculant production costs down. Some of the reviewed studies omitted bioflocculant purification while retaining considerable FA, a strategy that minimizes sample preparation and the costs associated with it. Another potential option not well explored in the literature insofar would be to culture the bioflocculant-producing microorganisms in the tailings they are meant to treat. This would eliminate and transportation costs and the need for a large-scale offsite culturing facility, and any nutrients found in the tailings would offset at least a portion of the costs of the culture broth. While the current studies optimize bioflocculant production and performance and begin to consider minimizing purification and nutrient requirements, the use of more advanced bioengineering techniques common to fields like biopharmaceutical production should also be considered to further enhance bioflocculant production and performance to minimize costs. Two such strategies, each seen in only one bioflocculation study, the enhanced induction of exopolysaccharide bioflocculant production using quorum sensing-associated N-acyl homoserine lactones and the directed evolution of bioflocculant-producing microorganisms, have been reported to enhance bioflocculant production and performance (He et al., 2010; Yang et al., 2016). These are powerful methods which must be applied to bioflocculation further. Metabolic engineering has been shown to enhance exopolysaccharide and polyglutamic acid production for other applications, and must be employed to enhance bioflocculant production (Levander et al., 2002; Henry and Durocher, 2011; Feng et al., 2015; Cai et al., 2018). The bioflocculant design principles gleaned from this review could also be used for the intelligent design and recombinant production of repetitive proteins for use as bioflocculants (Pickens et al., 2011; Bahniuk et al., 2016). Biopanning strategies are well suited for high-throughput screening of bioflocculant-producing microbes and their mutants produced using any of the preceding bioengineering methods, using adhesion to kaolin particles as the separation method (Sano et al., 2011). Naturally, the costs associated with these methods would need to scale appropriately with the scale of the bioflocculant production and treatment, and these methods have the potential to scale very well with cost, as, unlike direct media supplementation, mutations and recombinant plasmids tend to be naturally replicated and maintained in the cell culture as it propagates and scales.
The production of bioflocculants may face significant barriers to their application on an industrial scale. Perhaps the first major barrier to the application of bioflocculants revolves around safety; systems that use microorganisms as bioflocculants need to be designed so as to prevent unwanted release, in tandem with microbe selection to reduce any potentially harmful effects of release. These issues must include the input from a broad group of experts, including governmental safety leaders, biochemists, and application engineers to ensure public safety is maintained. Moreover, should the bioflocculant need to be produced by the microorganisms and purified, without properly assessing production yields the technology would be uneconomical. Although there are potential risks to the application of microorganisms as bioflocculants, one should remember that there may be significant benefits that could be realized as well. For example, if a microorganism can be a good bioflocculant but also fix carbon within the tailings environment a significant carbon capture strategy could be developed to reduce greenhouse gases. Moreover, the economic burden of deploying microorganisms could be offset through production of a high-value commodity, and using the ‘waste’ microorganisms as the bioflocculant. After 25 years of research it is evident that we have only started to apply biochemical techniques to the significant environmental liabilities that kaolin tailings present.
Author contributions
MB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Writing—Original draft preparation, Writing—review and editing. FA: Data curation, Investigation, Writing—Original draft preparation. XT: Supervision, Validation, Writing—review and editing. LU: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing—review and editing.
Funding
This work was supported by the Natural Sciences and Engineering Research Council (Canada)-Collaborative Research and Development Program, Canada’s Oil Sands Innovation Alliance and the Institute for Oil Sands Innovation. The funding sources were not involved in the study design, collection, analysis and interpretation of data, or writing of the report. The funding sources did review the article before submission, but only for potential intellectual property disclosures, as opposed to academic matters.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
FA, flocculating activity; NTUs, nephelometric turbidity units.
References
Abd El-Salam, A. E., Abd-El-Haleem, D., Youssef, A. S., Zaki, S., Abu-Elreesh, G., and El-Assar, S. A. (2017). Isolation, characterization, optimization, immobilization and batch fermentation of bioflocculant produced by Bacillus aryabhattai strain PSK1. J. Genet. Eng. Biotechnol. 15, 335–344. doi:10.1016/j.jgeb.2017.07.002
Ajayeoba, T. A., Dula, S., and Ijabadeniyi, O. A. (2019). Properties of poly-γ-glutamic acid producing-bacillus species isolated from Ogi Liquor and Lemon-Ogi Liquor. Front. Microbiol. 10, 771. doi:10.3389/fmicb.2019.00771
Anagnostopoulas, A., Navarro, M., Ahmad, A., Ding, Y., and Gaidajis, G. (2022). Valorization of phosphogypsum as a thermal energy storage material for low temperature applications. J. Clean. Prod. 342, 130839. doi:10.1016/j.jclepro.2022.130839
Anastasakis, K., Kalderis, D., and Diamadopoulos, E. (2009). Flocculation behavior of mallow and okra mucilage in treating wastewater. Desalination 249, 786–791. doi:10.1016/j.desal.2008.09.013
Asrafuzzaman, M., Fakhruddin, A. N. M., and Hossain, M. A. (2011). Reduction of turbidity of water using locally available natural coagulants. ISRN Microbiol. 2011, 1–6. doi:10.5402/2011/632189
Bahniuk, M. S., Alshememry, A. K., and Unsworth, L. D. (2016). High-yield recombinant expression and purification of marginally soluble, short elastin-like polypeptides. BioTechniques 61, 297–304. doi:10.2144/000114482
Bahniuk, M. S., Alshememry, A. K., and Unsworth, L. D. (2020). Human plasma protein adsorption to elastin-like polypeptide nanoparticles. Biointerphases 15, 021007. doi:10.1116/6.0000027
Bajaj, I. B., and Singhal, R. S. (2011). Flocculation properties of poly(γ-glutamic acid) produced from Bacillus subtilis isolate. Food bioproc. Tech. 4, 745–752. doi:10.1007/s11947-009-0186-y
Bala Subramanian, S., Yan, S., Tyagi, R. D., and Surampalli, R. Y. (2010). Extracellular polymeric substances (EPS) producing bacterial strains of municipal wastewater sludge: Isolation, molecular identification, EPS characterization and performance for sludge settling and dewatering. Water Res. 44, 2253–2266. doi:10.1016/j.watres.2009.12.046
Baron, S. (1996). Leptospira--Medical microbiology. Galveston, TX, USA: University of Texas Medical Branch at Galveston.
Bergaya, F., and Lagaly, G. (2006). General introduction: Clays, clay minerals, and clay science. Dev. clay Sci. 1, 1–18.
Biswas, J. K., Banerjee, A., Sarkar, B., Sarkar, D., Sarkar, S. K., Rai, M., et al. (2020). Exploration of an extracellular polymeric substance from earthworm gut bacterium (Bacillus licheniformis) for bioflocculation and heavy metal removal potential. Appl. Sci. 10, 349. doi:10.3390/app10010349
Bouaouine, O., Bourven, I., Khalil, F., and Baudu, M. (2020). Reuse of olive mill wastewater as a bioflocculant for water treatment processes. J. Clean. Prod. 246, 119031. doi:10.1016/j.jclepro.2019.119031
Brink, M., Belay, G., and De Wet, J. (2006). Plant resources of tropical africa 1: Cereals and pulses. The Netherlands: PROTA Foundation Wageningen.
Bukhari, N. A., Loh, S. K., Nasrin, A. B., and Jahim, J. M. (2020). Enzymatic hydrolysate of palm oil mill effluent as potential substrate for bioflocculant BM-8 production. Waste Biomass Valorization 11, 17–29. doi:10.1007/s12649-018-0421-8
Busi, S., Karuganti, S., Rajkumari, J., Paramanandham, P., and Pattnaik, S. (2017). Sludge settling and algal flocculating activity of extracellular polymeric substance (EPS) derived from bacillus cereus SK. Water Environ. J. 31, 97–104. doi:10.1111/wej.12215
Cai, D., Chen, Y., He, P., Wang, S., Mo, F., Li, X., et al. (2018). Enhanced production of poly-γ-glutamic acid by improving ATP supply in metabolically engineered Bacillus licheniformis. Biotechnol. Bioeng. 115, 2541–2553. doi:10.1002/bit.26774
Cao, G., Zhang, Y., Chen, L., Liu, J., Mao, K., Li, K., et al. (2015). Production of a bioflocculant from methanol wastewater and its application in arsenite removal. Chemosphere 141, 274–281. doi:10.1016/j.chemosphere.2015.08.009
Chen, H., Zhong, C., Berkhouse, H., Zhang, Y., Lv, Y., Lu, W., et al. (2016). Removal of cadmium by bioflocculant produced by Stenotrophomonas maltophilia using phenol-containing wastewater. Chemosphere 155, 163–169. doi:10.1016/j.chemosphere.2016.04.044
Chen, N., Liu, W., Huang, J., and Qiu, X. (2020). Preparation of octopus-like lignin-grafted cationic polyacrylamide flocculant and its application for water flocculation. Int. J. Biol. Macromol. 146, 9–17. doi:10.1016/j.ijbiomac.2019.12.245
Choy, S. Y., Prasad, K. M. N., Wu, T. Y., and Ramanan, R. N. (2015). A review on common vegetables and legumes as promising plant-based natural coagulants in water clarification. Int. J. Environ. Sci. Technol. (Tehran). 12, 367–390. doi:10.1007/s13762-013-0446-2
Deng, S. B., Bai, R. B., Hu, X. M., and Luo, Q. (2003). Characteristics of a bioflocculant produced by Bacillus mucilaginosus and its use in starch wastewater treatment. Appl. Microbiol. Biotechnol. 60, 588–593. doi:10.1007/s00253-002-1159-5
Devi, K., and zand Natarajan, K. A. (2015). Production and characterization of bioflocculants for mineral processing applications. Int. J. Min. Process. 137, 15–25. doi:10.1016/j.minpro.2015.02.007
Elkady, M. F., Farag, S., Zaki, S., Abu-Elreesh, G., and Abd-El-Haleem, D. (2011). Bacillus mojavensis strain 32A, a bioflocculant-producing bacterium isolated from an Egyptian salt production pond. Bioresour. Technol. 102, 8143–8151. doi:10.1016/j.biortech.2011.05.090
Eman Zakaria, G. (2012). Production and characteristics of a heavy metals removing bioflocculant produced by Pseudomonas aeruginosa. Pol. J. Microbiol. 61, 281–289. doi:10.33073/pjm-2012-038
Farag, S., Zaki, S., Elkady, M., and Abd-El-Haleem, D. (2014). Production and characteristics of a bioflocculant produced by Pseudomonas sp. strain 38A. J. Adv. Biol. 4, 286.
Feng, J., Gu, Y., Quan, Y., Cao, M., Gao, W., Zhang, W., et al. (2015). Improved poly-γ-glutamic acid production in Bacillus amyloliquefaciens by modular pathway engineering. Metab. Eng. 32, 106–115. doi:10.1016/j.ymben.2015.09.011
Ghadge, P., Vairagar, P., and Prasad, K. (2008). Some physical properties of chick pea split (Cicer arietinum L.) Agric. Eng. Int. CIGR J.
Ghimici, L., Constantin, M., and Fundueanu, G. (2010). Novel biodegradable flocculanting agents based on pullulan. J. Hazard. Mater. 181, 351–358. doi:10.1016/j.jhazmat.2010.05.017
Gomaa, E. Z. (2012). Production and characteristics of a heavy metals removing bioflocculant produced by Pseudomonas aeruginosa. Pol. J. Microbiol. 61, 281–289. doi:10.33073/pjm-2012-038
Gong, W. X., Wang, S. G., Sun, X. F., Liu, X. W., Yue, Q. Y., and Gao, B. Y. (2008). Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour. Technol. 99, 4668–4674. doi:10.1016/j.biortech.2007.09.077
Guo, J., Yang, C., and Zeng, G. (2013). Treatment of swine wastewater using chemically modified zeolite and bioflocculant from activated sludge. Bioresour. Technol. 143, 289–297. doi:10.1016/j.biortech.2013.06.003
Guo, J., Yang, C., and Peng, L. (2014). Preparation and characteristics of bacterial polymer using pre-treated sludge from swine wastewater treatment plant. Bioresour. Technol. 152, 490–498. doi:10.1016/j.biortech.2013.11.037
Guo, J., Lau, A. K., Zhang, Y., and Zhao, J. (2015). Characterization and flocculation mechanism of a bioflocculant from potato starch wastewater. Appl. Microbiol. Biotechnol. 99, 5855–5861. doi:10.1007/s00253-015-6589-y
Guo, J., Du, J., Chen, P., Tan, X., Huang, X., Gan, P., et al. (2017). Enhanced efficiencies of sludge dewatering and domestic wastewater treatment by using the bioflocculant from rice stover. Water Environ. J. 31, 120–126. doi:10.1111/wej.12221
Guo, H., Hong, C., Zhang, C., Zheng, B., Jiang, D., and Qin, W. (2018). Bioflocculants’ production from a cellulase-free xylanase-producing Pseudomonas boreopolis G22 by degrading biomass and its application in cost-effective harvest of microalgae. Bioresour. Technol. 255, 171–179. doi:10.1016/j.biortech.2018.01.082
Guo, J., Liu, J., Yang, Y., Zhou, Y., Jiang, S., and Chen, C. (2018). Fermentation and kinetics characteristics of a bioflocculant from potato starch wastewater and its application. Sci. Rep. 8, 3631–3711. doi:10.1038/s41598-018-21796-x
Hassett, D. J., Cuppoletti, J., Trapnell, B., Lymar, S. V., Rowe, J. J., Yoon, S. S., et al. (2002). Anaerobic metabolism and quorum sensing by Pseudomonas aeruginosa biofilms in chronically infected cystic fibrosis airways: Rethinking antibiotic treatment strategies and drug targets. Adv. drug Deliv. Rev. 54, 1425–1443. doi:10.1016/s0169-409x(02)00152-7
He, J., Zou, J., Shao, Z., Zhang, J., Liu, Z., and Yu, Z. (2010). Characteristics and flocculating mechanism of a novel bioflocculant HBF-3 produced by deep-sea bacterium mutant Halomonas sp. V3a. World J. Microbiol. Biotechnol. 26, 1135–1141. doi:10.1007/s11274-009-0281-2
Henry, O., and Durocher, Y. (2011). Enhanced glycoprotein production in HEK-293 cells expressing pyruvate carboxylase. Metab. Eng. 13, 499–507. doi:10.1016/j.ymben.2011.05.004
Hocking, M., Klimchuk, K., and Lowen, S. (1999). Polymeric flocculants and flocculation. J. Macromol. Sci. Part C Polym. Rev. 39, 177–203. doi:10.1081/mc-100101419
Ji, B., Zhang, X. Y., Li, Z., Xie, H. Q., Xiao, X. M., and Fan, G. J. (2010). Flocculation properties of a bioflocculant produced by Bacillus licheniformis. Water Sci. Technol. 62, 1907–1913. doi:10.2166/wst.2010.478
Karthiga Devi, K., and Natarajan, K. A. (2015). Production and characterization of bioflocculants for mineral processing applications. Int. J. Mineral Process. 137, 15–25. doi:10.1016/j.minpro.2015.02.007
Kay-Shoemake, J. L., Watwood, M. E., Lentz, R. D., and Sojka, R. E. (1998). Polyacrylamide as an organic nitrogen source for soil microorganisms with potential effects on inorganic soil nitrogen in agricultural soil. Soil Biol. Biochem. 30, 1045–1052. doi:10.1016/s0038-0717(97)00250-2
Kurane, R., Takeda, K., and Suzuki, T. (1986). Screening for and characteristics of microbial flocculants. Agric. Biol. Chem. 50, 2301–2307. doi:10.1080/00021369.1986.10867746
Labahn, S. K., Fisher, J. C., Robleto, E. A., Young, M. H., and Moser, P. (2010). Microbially mediated aerobic and anaerobic degradation of acrylamide in a Western United States irrigation canal. J. Environ. Qual. 39, 1563–1569. doi:10.2134/jeq2009.0318
Lee, S. H., Lee, S. O., Jang, K. L., and Lee, T. H. (1995). Microbial flocculant from arcuadendron sp. TS-49. Biotechnol. Lett. 17, 95–100. doi:10.1007/bf00134203
Lee, N.-R., Go, T.-H., Lee, S.-M., Jeong, S.-Y., Park, G.-T., Hong, C.-O., et al. (2014). In vitro evaluation of new functional properties of poly-γ-glutamic acid produced by Bacillus subtilis D7. Saudi J. Biol. Sci. 21, 153–158. doi:10.1016/j.sjbs.2013.09.004
Lee, H.-R., Jeon, J. H., and Rhie, G.-E. (2017). The poly-γ-D-glutamic acid capsule of Bacillus licheniformis, a surrogate of Bacillus anthracis capsule induces interferon-gamma production in NK cells through interactions with macrophages. J. Microbiol. Biotechnol. 27, 1032–1037. doi:10.4014/jmb.1612.12043
Levander, F., Svensson, M., and Rådström, P. (2002). Enhanced exopolysaccharide production by metabolic engineering of Streptococcus thermophilus. Appl. Environ. Microbiol. 68, 784–790. doi:10.1128/aem.68.2.784-790.2002
Li, Z., Chen, R. W., Lei, H. Y., Shan, Z., Bai, T., Yu, Q., et al. (2009). Characterization and flocculating properties of a novel bioflocculant produced by Bacillus circulans. World J. Microbiol. Biotechnol. 25, 745–752. doi:10.1007/s11274-008-9943-8
Li, Q., Liu, H. l., Qi, Q. s., Wang, F. s., and Zhang, Y. z. (2010). Isolation and characterization of temperature and alkaline stable bioflocculant from Agrobacterium sp. M-503. New Biotechnol. 27, 789–794. doi:10.1016/j.nbt.2010.09.002
Li, O., Lu, C., Liu, A., Zhu, L., Wang, P. M., Qian, C. D., et al. (2013). Optimization and characterization of polysaccharide-based bioflocculant produced by Paenibacillus elgii B69 and its application in wastewater treatment. Bioresour. Technol. 134, 87–93. doi:10.1016/j.biortech.2013.02.013
Li, H., Cai, T., Yuan, B., Li, R., Yang, H., and Li, A. (2015). Flocculation of both kaolin and hematite suspensions using the starch-based flocculants and their floc properties. Ind. Eng. Chem. Res. 54, 59–67. doi:10.1021/ie503606y
Lim, T. K. (2012). Edible medicinal and non-medicinal plants. Berlin/Heidelberg, Germany: Springer, 1.
Liu, W. J., Yuan, H. L., Yang, J. S., and Li, B. Z. (2009). Characterization of bioflocculants from biologically aerated filter backwashed sludge and its application in dying wastewater treatment. Bioresour. Technol. 100, 2629–2632. doi:10.1016/j.biortech.2008.12.017
Liu, W. J., Wang, K., Li, B. Z., Yuan, H. L., and Yang, J. S. (2010). Production and characterization of an intracellular bioflocculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresour. Technol. 101, 1044–1048. doi:10.1016/j.biortech.2009.08.108
Liu, Y., Zhang, T., Zhang, Z., Sun, T., Wang, J., and Lu, F. (2014). Improvement of cold adaptation of Bacillus alcalophilus alkaline protease by directed evolution. J. Mol. Catal. B Enzym. 106, 117–123. doi:10.1016/j.molcatb.2014.05.005
Liu, H., Yang, X., Zhang, Y., Zhu, H., and Yao, J. (2014). Flocculation characteristics of polyacrylamide grafted cellulose from phyllostachys heterocycla: Anefficient and eco-friendly flocculant. Water Res. 59, 165–171. doi:10.1016/j.watres.2014.04.022
Liu, C., Wang, K., Jiang, J.-H., Liu, W.-J., and Wang, J.-Y. (2015). A novel bioflocculant produced by a salt-tolerant, alkaliphilic and biofilm-forming strain Bacillus agaradhaerens C9 and its application in harvesting Chlorella minutissima UTEX2341. Biochem. Eng. J. 93, 166–172. doi:10.1016/j.bej.2014.10.006
Liu, W., Zhao, C., Jiang, J., Lu, Q., Hao, Y., Wang, L., et al. (2015). Bioflocculant production from untreated corn stover using Cellulosimicrobium cellulans L804 isolate and its application to harvesting microalgae. Biotechnol. Biofuels 8, 170–213. doi:10.1186/s13068-015-0354-4
Liu, W., Hao, Y., Jiang, J., Zhu, A., Zhu, J., and Dong, Z. (2016). Production of a bioflocculant from Pseudomonas veronii L918 using the hydrolyzate of peanut hull and its application in the treatment of ash-flushing wastewater generated from coal fired power plant. Bioresour. Technol. 218, 318–325. doi:10.1016/j.biortech.2016.06.108
Liu, C., Hao, Y., Jiang, J., and Liu, W. (2017). Valorization of untreated rice bran towards bioflocculant using a lignocellulose-degrading strain and its use in microalgal biomass harvest. Biotechnol. Biofuels 10, 90–12. doi:10.1186/s13068-017-0780-6
Liu, D., Edraki, M., Fawell, P., and Berry, L. (2020). Improved water recovery: A review of clay-rich tailings and saline water interactions. Powder Technol. 364, 604–621. doi:10.1016/j.powtec.2020.01.039
Lu, Y., Shang, Y., Huang, X., Chen, A., Yang, Z., Jiang, Y., et al. (2011). Preparation of strong cationic chitosan- graft -polyacrylamide flocculants and their flocculating properties. Ind. Eng. Chem. Res. 50, 7141–7149. doi:10.1021/ie2000665
Luo, L., Zhao, Z., Huang, X., Du, X., Wang, C. a., Li, J., et al. (2016). Isolation, identification, and optimization of culture conditions of a bioflocculant-producing bacteriumBacillus megateriumSP1 and its application in aquaculture wastewater treatment. BioMed Res. Int. 2016, 1–9. doi:10.1155/2016/2758168
Mabinya, L. V., Cosa, S., Nwodo, U., and Okoh, A. I. (2012). Studies on bioflocculant production by arthrobacter sp. raats, a freshwater bacteria isolated from tyume river, South Africa. Int. J. Mol. Sci. 13, 1054–1065. doi:10.3390/ijms13011054
Madigan, M. T., Martinko, J. M., Dunlap, P. V., and Clark, D. P. (2008). Brock biology of microorganisms 12th edn. Int. Microbiol. 11, 65–73.
Makapela, B., Okaiyeto, K., Ntozonke, N., Nwodo, U. U., Green, E., Mabinya, L. V., et al. (2016). Assessment of bacillus pumilus isolated from fresh water milieu for bioflocculant production, 6.Appl. Sci. Switz.
Marobhe, N. J., Dalhammar, G., and Gunaratna, K. R. (2007). Simple and rapid methods for purification and characterization of active coagulants from the seeds of vigna unguiculata and parkinsonia aculeata. Environ. Technol. 28, 671–681. doi:10.1080/09593332808618827
Masliyah, J. H., Xu, Z., and Czarnecki, J. A. (2011). Handbook on theory and practice of bitumen recovery from athabasca oil sands: Theoretical basis. Pampanga, Philippines: Kingsley Knowledge Pub.
Mishra, S., and KunduSynthesis, K. (2019). Synthesis, characterization and applications of polyacrylamide grafted fenugreek gum (FG-g-PAM) as flocculant: Microwave vs thermal synthesis approach. Int. J. Biol. Macromol. 141, 792–808. doi:10.1016/j.ijbiomac.2019.09.033
Monga, A., Fulke, A. B., Gaud, A., Sharma, A., Ram, A., and Dasgupta, B. (2022). Isolation and identification of novel chromium tolerant bacterial strains from a heavy metal polluted urban creek: An assessment of bioremediation efficiency and flocculant production. Thalassas. doi:10.1007/s41208-022-00458-w
More, T. T., Yan, S., John, R. P., Tyagi, R. D., and Surampalli, R. Y. (2012). Biochemical diversity of the bacterial strains and their biopolymer producing capabilities in wastewater sludge. Bioresour. Technol. 121, 304–311. doi:10.1016/j.biortech.2012.06.103
Morgan, A. A., and Rubenstein, E. P (2013). Proline: The distribution, frequency, positioning, and common functional roles of proline and polyproline sequences in the human proteome. PloS one 8, e53785. doi:10.1371/journal.pone.0053785
Nguyen, N. T., Phan, T. H. M., Tran, T. N., Velmurugan, B. K., and Kiefer, R. (2019). Production of Novel Bio-Flocculants from Klebsiella variicola BF1 using Cassava Starch Wastewater and its Application. Curr. Sci. 117, 121. doi:10.18520/cs/v117/i1/121-129
Nguyen, V. H., Klai, N., Nguyen, T. D., and Tyagi, R. D. (2016). Impact of extraction methods on bio-flocculants recovered from backwashed sludge of bio-filtration unit. J. Environ. Manag. 180, 344–350. doi:10.1016/j.jenvman.2016.05.051
Nouha, K. (2016). Characterization of extracellular polymeric substances (eps) produced by cloacibacterium normanense isolated from wastewater sludge for sludge settling and dewatering. J. Civ. Environ. Eng. 05. doi:10.4172/2165-784x.1000191
Ofori, P., Nguyen, A. V., Firth, B., McNally, C., and Ozdemir, O. (2011). Shear-induced floc structure changes for enhanced dewatering of coal preparation plant tailings. Chem. Eng. J. 172, 914–923. doi:10.1016/j.cej.2011.06.082
Okaiyeto, K., Nwodo, U. U., Mabinya, L. V., and Okoh, A. I. (2013). Characterization of a bioflocculant produced by a consortium of halomonas sp. okoh and micrococcus sp. leo. Int. J. Environ. Res. Public Health 10, 5097–5110. doi:10.3390/ijerph10105097
Pan, J. R., Huang, C., Chuang, Y.-C., and Wu, C.-C. (1999). Dewatering characteristics of algae-containing alum sludge. Colloids Surfaces A Physicochem. Eng. Aspects 150, 185–190. doi:10.1016/s0927-7757(98)00868-1
Parajulee, A., and Wania, F. (2014). Evaluating officially reported polycyclic aromatic hydrocarbon emissions in the Athabasca oil sands region with a multimedia fate model. Proc. Natl. Acad. Sci. U. S. A. 111, 3344–3349. doi:10.1073/pnas.1319780111
Patale, V., and Pandya, J. (2012). Mucilage extract of Coccinia indica fruit as coagulant-flocculent for turbid water treatment. Asian J. Plant Sci. Res. 2, 442–445.
Pickens, L. B., Tang, Y., and Chooi, Y.-H. (2011). Metabolic engineering for the production of natural products. Annu. Rev. Chem. Biomol. Eng. 2, 211–236. doi:10.1146/annurev-chembioeng-061010-114209
Prasertsan, P., Dermlim, W., Doelle, H., and Kennedy, J. F. (2006). Screening, characterization and flocculating property of carbohydrate polymer from newly isolated Enterobacter cloacae WD7. Carbohydr. Polym. 66, 289–297. doi:10.1016/j.carbpol.2006.03.011
Pu, S. y., Qin, L. l., Che, J. p., Zhang, B. r., and Xu, M. (2014). Preparation and application of a novel bioflocculant by two strains of Rhizopus sp. using potato starch wastewater as nutrilite. Bioresour. Technol. 162, 184–191. doi:10.1016/j.biortech.2014.03.124
Pu, S., Ma, H., Deng, D., Xue, S., Zhu, R., Zhou, Y., et al. (2018). Isolation, identification, and characterization of an Aspergillus Niger bioflocculant-producing strain using potato starch wastewater as nutrilite and its application. PLoS One 13, e0190236. doi:10.1371/journal.pone.0190236
Qi, Z., Zhu, Y., Guo, H., Chen, Y., Zhao, Y., Zhou, Y., et al. (2019). Production of glycoprotein bioflocculant from untreated rice straw by a CAZyme-rich bacterium, Pseudomonas sp. HP2. J. Biotechnol. 306, 185–192. doi:10.1016/j.jbiotec.2019.10.011
Robinson, R. K. (2014). Encyclopedia of food microbiology. Cambridge, Massachusetts, USA: Academic Press.
Sabah, E., Yüzer, H., and Celik, M. (2004). Characterization and dewatering of fine coal tailings by dual-flocculant systems. Int. J. Mineral Process. 74, 303–315. doi:10.1016/j.minpro.2004.03.001
Salehizadeh, H., and Shojaosadati, S. (2001). Extracellular biopolymeric flocculants: Recent trends and biotechnological importance. Biotechnol. Adv. 19, 371–385. doi:10.1016/s0734-9750(01)00071-4
Salehizadeh, H., and Shojaosadati, S. (2002). Isolation and characterisation of a bioflocculant produced by Bacillus firmus. Biotechnol. Lett. 24, 35–40. doi:10.1023/a:1013853115624
Sano, D., Ishifuji, S., Sato, Y., Imae, Y., Takaara, T., Masago, Y., et al. (2011). Identification and characterization of coagulation inhibitor proteins derived from cyanobacterium Microcystis aeruginosa. Chemosphere 82, 1096–1102. doi:10.1016/j.chemosphere.2010.12.005
Sathiyanarayanan, G., Seghal Kiran, G., and Selvin, J. (2013). Synthesis of silver nanoparticles by polysaccharide bioflocculant produced from marine Bacillus subtilis MSBN17. Colloids Surfaces B Biointerfaces 102, 13–20. doi:10.1016/j.colsurfb.2012.07.032
Schindler, D. W. (2014). Unravelling the complexity of pollution by the oil sands industry. Proc. Natl. Acad. Sci. U. S. A. 111, 3209–3210. doi:10.1073/pnas.1400511111
Sciban, M., Klasnja, M., and Stojimirovic, J. (2005). Investigation of coagulation activity of natural coagulants from seeds of different leguminose species. Acta Period. Technol. 266, 81–90. doi:10.2298/apt0536081s
Shaikh, S. M., Nasser, M. S., Hussein, I., Benamor, A., Onaizi, S. A., and Qiblawey, H. (2017). Influence of polyelectrolytes and other polymer complexes on the flocculation and rheological behaviors of clay minerals: A comprehensive review. Sep. Purif. Technol. 187, 137–161. doi:10.1016/j.seppur.2017.06.050
Shih, I. L., Van, Y. T., Yeh, L. C., Lin, H. G., and Chang, Y. N. (2001). Production of a biopolymer flocculant from Bacillus licheniformis and its flocculation properties. Bioresour. Technol. 78, 267–272. doi:10.1016/s0960-8524(01)00027-x
Shukor, M., Gusmanizar, N., Azmi, N., Hamid, M., Ramli, J., Shamaan, N., et al. (2009). Isolation and characterization of an acrylamide-degrading Bacillus cereus. J. Environ. Biol. 30, 57–64.
Smith, E. A., Prues, S. L., and Oehme, F. W. (1997). Environmental degradation of polyacrylamides. Ecotoxicol. Environ. Saf. 37, 76–91. doi:10.1006/eesa.1997.1527
Sun, J., Zhang, X., Miao, X., and Zhou, J. (2012). Preparation and characteristics of bioflocculants from excess biological sludge. Bioresour. Technol. 126, 362–366. doi:10.1016/j.biortech.2012.08.042
Takami, H., and Horikoshi, K. (2000). Analysis of the genome of an alkaliphilic Bacillus strain from an industrial point of view. Extremophiles 4, 99–108. doi:10.1007/s007920050143
Tang, W., Song, L., Li, D., Qiao, J., Zhao, T., and Zhao, H. (2014). Production, characterization, and flocculation mechanism of cation independent, pH tolerant, and thermally stable bioflocculant from Enterobacter sp. ETH-2. PLoS ONE 9, 114591. doi:10.1371/journal.pone.0114591
Toeda, K., and Kurane, R. (1991). Microbial flocculant from Alcaligenes cupidus KT201. Agric. Biol. Chem. 55, 2793–2799. doi:10.1080/00021369.1991.10857749
Ugbenyen, A. M., Vine, N., Simonis, J. J., Basson, A. K., and Okoh, A. I. (2015). Characterization of a bioflocculant produced from the consortium of three marine bacteria of the genera cobetia and bacillus and its application for wastewater treatment. J. Water, Sanitation Hyg. Dev. 5, 81–88. doi:10.2166/washdev.2014.181
Wang, S. G., Gong, W. X., Liu, X. W., Tian, L., Yue, Q. Y., and Gao, B. Y. (2007). Production of a novel bioflocculant by culture of Klebsiella mobilis using dairy wastewater. Biochem. Eng. J. 36, 81–86. doi:10.1016/j.bej.2007.02.003
Wang, L., Ma, F., Lee, D. J., Wang, A., and Ren, N. (2013). Bioflocculants from hydrolysates of corn stover using isolated strain Ochrobactium ciceri W2. Bioresour. Technol. 145, 259–263. doi:10.1016/j.biortech.2012.11.020
Wang, C., Harbottle, D., Liu, Q., and Xu, Z. (2014). Current state of fine mineral tailings treatment: A critical review on theory and practice. Miner. Eng. 58, 113–131. doi:10.1016/j.mineng.2014.01.018
Wang, Z., Shen, L., Zhuang, X., Shi, J., Wang, Y., He, N., et al. (2015). Flocculation characterization of a bioflocculant from bacillus licheniformis. Ind. Eng. Chem. Res. 54, 2894–2901. doi:10.1021/ie5050204
Wong, Y. S., Ong, S. A., Teng, T. T., Aminah, L. N., and Kumaran, K. (2012). Production of bioflocculant by staphylococcus cohnii ssp. from palm oil mill effluent (POME). Water Air Soil Pollut. 223, 3775–3781. doi:10.1007/s11270-012-1147-z
Wu, J. Y., and Ye, H. F. (2007). Characterization and flocculating properties of an extracellular biopolymer produced from a Bacillus subtilis DYU1 isolate. Process Biochem. 42, 1114–1123. doi:10.1016/j.procbio.2007.05.006
Xia, X., Liang, Y., Lan, S., Li, X., Xie, Y., and Yuan, W. (2018). Production and flocculating properties of a compound biopolymer flocculant from corn ethanol wastewater. Bioresour. Technol. 247, 924–929. doi:10.1016/j.biortech.2017.10.003
Xiong, B., Loss, R. D., Shields, D., Pawlik, T., Hochreiter, R., Zydney, A. L., et al. (2018). Polyacrylamide degradation and its implications in environmental systems. NPJ Clean. Water 1, 17–19. doi:10.1038/s41545-018-0016-8
Yadav, K. K., Mandal, A. K., Sen, I. K., Chakraborti, S., Islam, S. S., and Chakraborty, R. (2012). Flocculating property of extracellular polymeric substances produced by a biofilm-forming bacterium acinetobacter junii BB1A. Appl. Biochem. Biotechnol. 168, 1621–1634. doi:10.1007/s12010-012-9883-5
Yalçın, İ. (2007). Physical properties of cowpea (Vigna sinensis L.) seed. J. Food Eng. 79, 57–62. doi:10.1016/j.jfoodeng.2006.01.026
Yang, Q., Luo, K., Liao, D. X., Li, X. M., Wang, D. B., Liu, X., et al. (2012). A novel bioflocculant produced by Klebsiella sp. and its application to sludge dewatering. Water Environ. J. 26, 560–566. doi:10.1111/j.1747-6593.2012.00319.x
Yang, J., Wu, D., Li, A., Guo, H., Chen, H., Pi, S., et al. (2016). The addition of N-Hexanoyl-Homoserine lactone to improve the microbial flocculant production of Agrobacterium tumefaciens strain F2, an exopolysaccharide bioflocculant-producing bacterium. Appl. Biochem. Biotechnol. 179, 728–739. doi:10.1007/s12010-016-2027-6
Yim, J. H., Kim, S. J., Ahn, S. H., and Lee, H. K. (2007). Characterization of a novel bioflocculant, p-KG03, from a marine dinoflagellate, Gyrodinium impudicum KG03. Bioresour. Technol. 98, 361–367. doi:10.1016/j.biortech.2005.12.021
Yin, Y. J., Tian, Z. M., Tang, W., Li, L., Song, L. Y., and McElmurry, S. P. (2014). Production and characterization of high efficiency bioflocculant isolated from Klebsiella sp. ZZ-3. Bioresour. Technol. 171, 336–342. doi:10.1016/j.biortech.2014.08.094
Yokoi, H., Yoshida, T., Mori, S., Hirose, J., Hayashi, S., and Takasaki, Y. (1997). Biopolymer flocculant produced by an Enterobacter sp. Biotechnol. Lett. 19, 569–573. doi:10.1023/a:1018301807009
Yokoi, H., Obita, T., Hirose, J., Hayashi, S., and Takasaki, Y. (2002). Flocculation properties of pectin in various suspensions. Bioresour. Technol. 84, 287–290. doi:10.1016/s0960-8524(02)00023-8
You, Y., Ren, N., Wang, A., Ma, F., Gao, L., Peng, Y., et al. (2008). Use of waste fermenting liquor to produce bioflocculants with isolated strains. Int. J. Hydrogen Energy 33, 3295–3301. doi:10.1016/j.ijhydene.2008.03.049
Yuan, S. J., Sun, M., Sheng, G. P., Li, Y., Li, W. W., Yao, R. S., et al. (2011). Identification of key constituents and structure of the extracellular polymeric substances excreted by bacillus megaterium TF10 for their flocculation capacity. Environ. Sci. Technol. 45, 1152–1157. doi:10.1021/es1030905
Zhao, G., Ma, F., Wei, L., and Chua, H. (2012). Using rice straw fermentation liquor to produce bioflocculants during an anaerobic dry fermentation process. Bioresour. Technol. 113, 83–88. doi:10.1016/j.biortech.2011.11.040
Zhao, C., Zhang, Y., Wei, X., Hu, Z., Zhu, F., Xu, L., et al. (2013). Production of ultra-high molecular weight poly-γ-glutamic acid with bacillus licheniformis P-104 and characterization of its flocculation properties. Appl. Biochem. Biotechnol. 170, 562–572. doi:10.1007/s12010-013-0214-2
Zheng, Y., Ye, Z. L., Fang, X. L., Li, Y. H., and Cai, W. M. (2008). Production and characteristics of a bioflocculant produced by Bacillus sp. F19. Bioresour. Technol. 99, 7686–7691. doi:10.1016/j.biortech.2008.01.068
Zhong, C., Xu, A., Chen, L., Yang, X., Yang, B., Hong, W., et al. (2014). Production of a bioflocculant from chromotropic acid waste water and its application in steroid estrogen removal. Colloids Surfaces B Biointerfaces 122, 729–737. doi:10.1016/j.colsurfb.2014.08.006
Zhong, C., Xu, A., Wang, B., Yang, X., Hong, W., Yang, B., et al. (2014). Production of a value added compound from the H-acid waste water-Bioflocculants by Klebsiella pneumoniae. Colloids Surfaces B Biointerfaces 122, 583–590. doi:10.1016/j.colsurfb.2014.07.036
Zhu, H., Zhang, Y., Yang, X., Liu, H., Shao, L., Zhang, X., et al. (2015). One-step green synthesis of non-hazardous dicarboxyl cellulose flocculant and its flocculation activity evaluation. J. Hazard. Mater. 296, 1–8. doi:10.1016/j.jhazmat.2015.04.029
Keywords: Bacteria, bioflocculant, clay, polysaccharide, protein, tailings pond, wastewater
Citation: Bahniuk MS, Alidina F, Tan X and Unsworth LD (2022) The last 25 years of research on bioflocculants for kaolin flocculation with recent trends and technical challenges for the future. Front. Bioeng. Biotechnol. 10:1048755. doi: 10.3389/fbioe.2022.1048755
Received: 19 September 2022; Accepted: 09 November 2022;
Published: 24 November 2022.
Edited by:
Anuj Kumar, Yeungnam University, South KoreaReviewed by:
Bridgid Lai Fui Chin, Curtin University Malaysia, MalaysiaAtul Dev, University of California, Davis, United States
Copyright © 2022 Bahniuk, Alidina, Tan and Unsworth. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Markian S. Bahniuk, bWJhaG5pdWtAdWFsYmVydGEuY2E=; Larry D. Unsworth, bHVuc3dvcnRAdWFsYmVydGEuY2E=