 Timo Z. Nazari-Shafti1,2,3
Timo Z. Nazari-Shafti1,2,3 Vasileios Exarchos1,4Héctor Rodriguez Cetina Biefer1,5
Vasileios Exarchos1,4Héctor Rodriguez Cetina Biefer1,5 Nikola Cesarovic1,4Heike Meyborg1Volkmar Falk1,3,4,5
Nikola Cesarovic1,4Heike Meyborg1Volkmar Falk1,3,4,5 Maximilian Y. Emmert1,3,5,6,7*
Maximilian Y. Emmert1,3,5,6,7*- 1Department for Cardiovascular and Thoracic Surgery, German Heart Center Berlin, Berlin, Germany
- 2Berlin Institute of Health, Berlin, Germany
- 3Deutsches Zentrum für Herz-und Kreislauferkrankungen, Berlin, Germany
- 4Department of Health Sciences and Technology, ETH Zürich, Zurich, Switzerland
- 5Clinic for Cardiovascular Surgery, Charité Universitätsmedizin Berlin, Berlin, Germany
- 6Institute for Regenerative Medicine, University of Zurich, Zurich, Switzerland
- 7Wyss Zurich, University of Zurich and ETH Zurich, Zurich, Switzerland
In the past 20 years, there have been several approaches to achieve cardioprotection or cardiac regeneration using a vast variety of cell therapies and remote ischemic pre-conditioning (RIPC). To date, substantial proof that either cell therapy or RIPC has the potential for clinically relevant cardiac repair or regeneration of cardiac tissue is still pending. Preclinical trials indicate that the secretome of cells in situ (during RIPC) as well as of transplanted cells may exhibit cardioprotective properties in the acute setting of cardiac injury. The secretome generally consists of cell-specific cytokines and extracellular vesicles (EVs) containing microRNAs (miRNAs). It is currently hypothesized that a subset of known miRNAs play a crucial part in the facilitation of cardioprotective effects. miRNAs are small non-coding RNA molecules that inhibit post-transcriptional translation of messenger RNAs (mRNAs) and play an important role in gene translation regulation. It is also known that one miRNAs usually targets multiple mRNAs. This makes predictability of pharmacokinetics and mechanism of action very difficult and could in part explain the inferior performance of various progenitor cells in clinical studies. Identification of miRNAs involved in cardioprotection and remodeling, the composition of miRNA profiles, and the exact mechanism of action are important to the design of future cell-based but also cell-free cardioprotective therapeutics. This review will give a description of miRNA with cardioprotective properties and a current overview on known mechanism of action and potential missing links. Additionally, we will give an outlook on the potential for clinical translation of miRNAs in the setting of myocardial infarction and heart failure.
Introduction
Cardiovascular disease remains one of the most important challenges clinicians face today. Despite huge efforts in prevention and undeniable progress in acute and short-term survival, disease progression over long-term still burdens our health care system without a real curative approach. The reason: regenerative capacity of an adult human heart is quite limited due to the low turnover rate of cardiomyocytes and lack of a sufficient pool of tissue resident progenitors (Tzahor and Poss, 2017). This renders the human heart very susceptible to any form of acute or chronic injury with a critical loss of cardiomyocytes or their function, ultimately leading to the clinical manifestation of heart failure. For these facts, cardiac medicine has been identified as a promising field for application of regeneration technologies. In the past 20 years, cell-based therapies have aimed to either induce ‘de novo’ generation or stimulate tissue dormant progenitors to differentiate into mature cardiomyocytes (Behfar et al., 2014). The promising results from in vitro studies and pre-clinical trials have led to a large number of clinical trials that for the most part investigated the therapeutic effect of different bone marrow, adipose- or neonatal tissue derived progenitor cells (first generation cell therapy). With limited pre-clinical data available in hindsight, a rush into first clinical trials in the early and mid 2000s has largely failed to demonstrate any meaningful results for patients with cardiac disease. More recently, myocardium resident cardiac progenitor cells, iPSCs, and conditioned progenitor cells have also been investigated in that context (second generation cell therapy) (Cambria et al., 2017). However, they as well failed to reach respective primary endpoints in most clinical trials; that is to say reduction in scar size and stabilization or improvement of cardiac function (Moyé, 2014; Gyöngyösi et al., 2015). As a result, the idea of a regenerating heart has been abandoned by many scientists. Luckily, the failure of such clinical studies has also led to post hoc analyses of the mechanism of action by which various cell types exhibit cardioprotection in the injured heart. Today, there is consensus that paracrine mediators, released by transplanted cells upon injury signals, mediate protection and limit adverse myocardial remodeling (Madonna et al., 2019). There is evidence emerging, that mediators can limit the extend of cardiomyocyte loss during acute injury and positively impact adverse myocardial remodeling in the chronic setting. Within the past 10 years, microRNAs (miRNAs) have come into focus as the next generation “cell” therapy studies have demonstrated that the paracrine secretion of nano- and macrovesicles containing miRNAs are mainly responsible for the cardioprotective effect of cellular therapies. Preclinical trials were able to demonstrate that miRNAs or extracellular vesicles (EVs) containing miRNAs were capable to reproduce the cellular effects of cardioprotection (Behfar and Terzic, 2019; Madonna et al., 2019; Maring et al., 2019). Within the scope of this review, we intend to provide a general overview of the potential role of miRNAs in cardioprotection and elaborate on potential use of miRNAs as a therapeutic agent.
Micro RNAs – One Shoe Fits All?
MicroRNAs play an important role in the inhibition of messenger RNA (mRNA) translation in the cytoplasm but have been also identified to regulate transcription in the nuclear compartment of mammalian cells (Bartel, 2004). The biosynthesis was long believed to be linear and universal for all miRNAs. However, recent functional studies demonstrated, that a multitude of alternate miRNA-specific biosynthesis pathways exist and that they require a plethora of regulatory mechanisms – many of which still need to be identified (Lee et al., 1993; Bartel, 2004) (Figure 1). miRNA are encoded within the entire genome and are usually arranged in clusters (Rodriguez et al., 2004). Most miRNA genes are located in the non-coding areas of the genome. In some cases, miRNAs are located within the introns of protein coding genes such as miR-103 which is located within in the intron of pantothenate kinase 1, 2, and 3 together with miR-107 (Rodriguez et al., 2004; Lin et al., 2006). miRNAs are transcribed by RNA Polymerases II and III into a primary miRNA (pri-miRNA). The pri-miRNA consists of a terminal loop region, a stem and two single-stranded flanking RNA regions up- and downstream of the hairpin (Cai et al., 2004; Lee et al., 2004). For the canonical pathway of miRNA maturation, the terminal single stranded RNA region is then spliced by the complex of Drosha and DGCR8 protein (DiGeorge critical region 8) (Lee et al., 2003). The DGCR8 protein has a binding and proof-reading domain that ensures the correct binding and identification of the cleavage site for the Drosha protein, a RNase III enzyme. For some miRNAs additional factors may be required for the correct splicing of the hairpin precursor (Guil and Cáceres, 2007; Davis et al., 2008; Michlewski et al., 2008) before the precursor miRNA (pre-miRNA) is transported from the nucleus into the cytoplasm by Exportin-5 (Yi et al., 2003). Exportin-5 not only acts as a transporter but also as an additional “proof-reader” only transporting correctly processed pre-miRNA (Zeng and Cullen, 2004). In the cytoplasm, the pre-miRNA – still consisting of the terminal loop and the double stranded stem – binds to the RNA-induced silencing complex (RISC) (Gregory et al., 2005). This protein complex contains RNase Dicer, double stranded RNA binding domain proteins (Tar RNA binding protein), PACT (protein activator of PKR), and Argonaute-2 (MacRae et al., 2008). The latter is the key component that mediates the miRNA inhibition of transcription of mRNAs (Diederichs and Haber, 2007). The terminal loop of pre-miRNAs is spliced off, whereas for some of the miRNAs both the guide and passenger strand can serve as individual mature miRNAs (Matranga et al., 2005). In general, the passenger strand is degraded after unwinding of the double stranded pre-miRNA by helicases (Leuschner et al., 2006). Similar to the processing of miRNAs in the nucleus some miRNAs are dependent on additional co-factors for maturation (Hutvágner et al., 2001; Diederichs et al., 2008). This highlights again, that miRNA biosynthesis is subject to an integrate regulatory machinery which we only just begin to understand. Upon maturation, miRNAs as part of the RISC complex either inhibit the translation of their target mRNA or they are packaged into EVs contained in multivesicular bodies to be released in the extracellular space (Turchinovich et al., 2012). The encapsulation of miRNAs does not happen at random and is also a highly regulated process (Turchinovich et al., 2012; Villarroya-Beltri et al., 2013). This allows for the dynamic response of cells in regard to which miRNAs are released upon microenvironmental cues. A study by Villarroya-Beltri et al. (2013) has shown that specific motifs within the sequence of miRNA determine their localization in either the cytoplasm or EVs. They have also identified heterogeneous nuclear ribonucleoproteins such as hnRNPA2B1 and hnRNPA1 that specifically bind to these motifs and are also present in EVs. Independent of their origin, all miRNAs have in common to alter gene expression via inhibitory mechanisms of posttranscriptional modification of mRNAs. The mechanism of inhibition depends on the complementarity of the miRNA sequence to its target mRNA. miRNAs with high complementarity direct the RISC to the mRNA and initiate the degradation. Lower complementarity can lead to inhibition of ribosomal translation of the target mRNA. It is overall hypothesized that the specificity of miRNAs is determined by the quality and stability of base pairing to their respective targets. Furthermore, miRNAs have also been identified to perform transcriptional silencing by targeting promotor regions within the heterochromatin. The aforementioned regulation of biosynthesis, maturation, and biological effects of numerous miRNA can be influenced by changes in the cellular microenvironment in a non-linear manner – posing a major challenge for scientists to predict possible effects of miRNA in vivo (Lee et al., 1993; Liu B. et al., 2014).
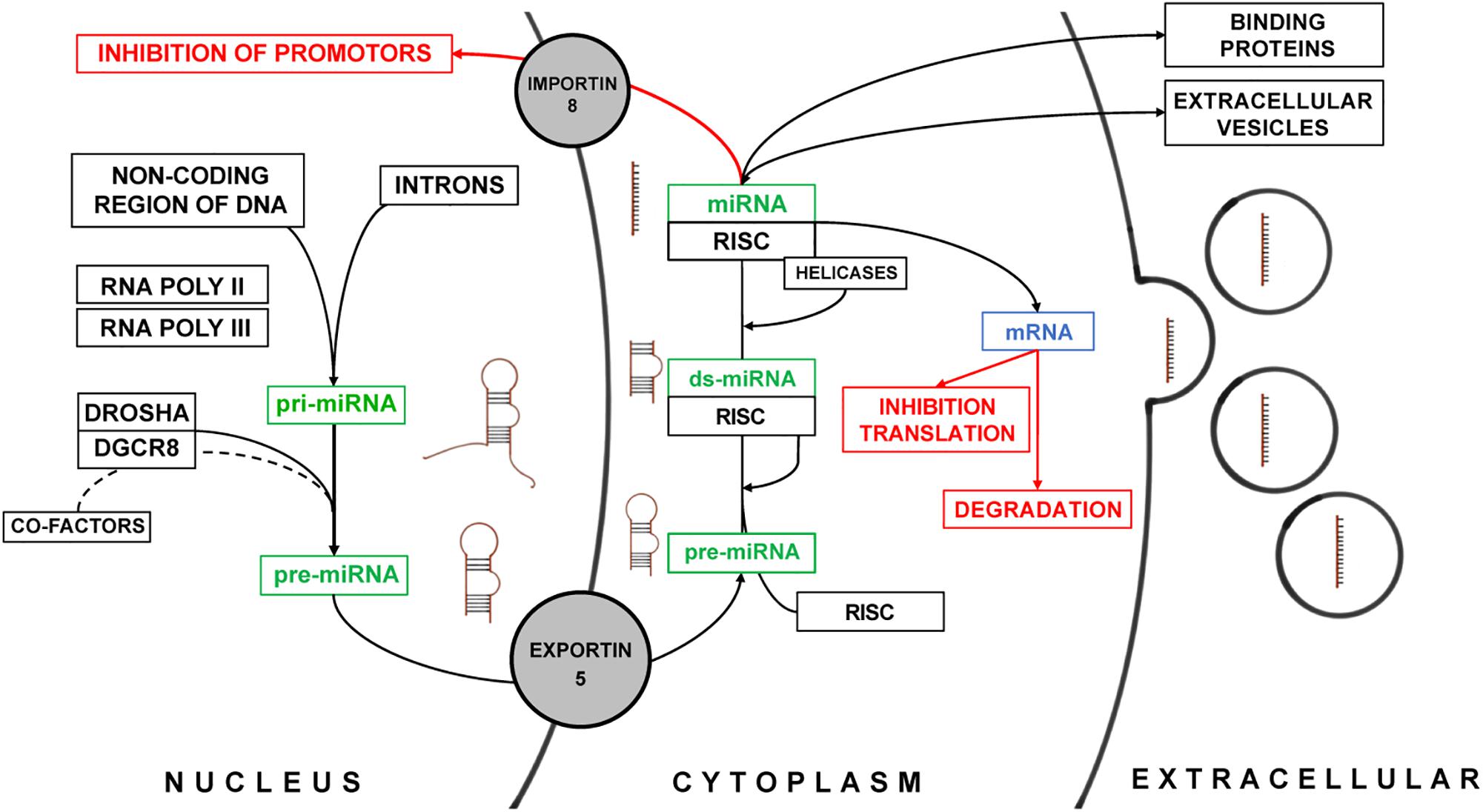
Figure 1. Summary of the biosynthesis and the biological effect of miRNAs. The different stages of miRNA are schematically depicted next to the green description of miRNA. The biological effect of miRNAs is marked in red.
Cardioprotective miRNAs – a Two-Sided Sword
For some time now, it has been hypothesized that factors released by cells under stress can induce a protective effect in neighboring or remote tissues (Kharbanda et al., 2002; Chen et al., 2008; Hausenloy et al., 2015). We now know that the key player in transmitting these signals are miRNAs. In the extracellular compartment, miRNAs are usually transported via EVs or binding proteins to protect them from degradation by nucleases (Li et al., 2012; Boon and Dimmeler, 2015). Similarly, the effect of therapeutic cell preparations exhibits their protective effect via the transmission of EVs loaded with miRNAs. Over the course of the past 10 years the sera of patients undergoing an ischemic event as well as the secretory profile of most cells utilized for cardioprotective purposes have been characterized (Baglio et al., 2015; Barile et al., 2016; Barile and Vassalli, 2017; Shao et al., 2017; Bellin et al., 2019). Each year, an increasing number of pathways are identified that either hint toward protection or damage of the myocardium (Barile et al., 2014, 2016; Gallet et al., 2016; Ciullo et al., 2019). Many functional studies have preceded these in-depth analyses of miRNA as cardioprotective agents, remote ischemic pre-conditioning (RIPC) being one of the most prominent examples (Kharbanda et al., 2002; Hausenloy et al., 2015). Here, a different organ or the heart itself is exposed to brief, non-fatal ischemia/reperfusion. Pre-clinical models show that RIPC can increase the survival of cardiomyocytes upon injury and positively impacts the myocardial remodeling (Konstantinov et al., 2005; Wei et al., 2011). In RIPC, circulating miRNA and EVs play an important role (Frey et al., 2018; Spannbauer et al., 2019). Even though circulating miRNAs and EVs have been identified as key mediators of that cardioprotective effect, it is a rather crude and a non-targeted therapeutic approach. Most when RIPC was investigated in clinical trials, none of the primary endpoints predicted by preclinical studies were met (Brevoord et al., 2012). Similar to experience from clinical trials investigating cell therapies, the exact mechanisms of action of RIPC were not fully understood and may explain the failed translation. However, the data collected on miRNAs from these studies laid the groundwork for many functional studies investigating the cardioprotective effects of miRNAs.
In recent years, a plethora of mechanistical studies for the downstream effect of various miRNA were conducted. Here, miRNAs were either investigated as diagnostic markers or as potential targets for therapies (Barile et al., 2016). The in-depth analysis of regulation of the identified miRNAs and their downstream targets revealed that some miRNAs had contradictory effects in the myocardium (Table 1). Furthermore, to the best of our knowledge no study performed any concurrent analysis to identify potential targets that are not related to a cardioprotective effect. In this next section, we highlight a selection of miRNAs that have been associated with a cardioprotective potential but also bear the risk of adverse or off-target effects. Most of these miRNAs were identified either in EVs of therapeutic cell products or as biomarkers during acute and chronic myocardial injury. The knowledge of their exact mechanism of action is therefore highly recommended. The following miRNAs are only a small example of miRNAs that are commonly identified but not limited to their cardioprotective potential. The selection of miRNA should not be seen as a comprehensive summary of all known cardioprotective miRNAs which would be beyond the scope of this review. Others have provided more detailed lists of cardioprotective miRNAs (Varga et al., 2015; Wendt et al., 2018). The pre-clinical experience with the following miRNAs should highlight the potential and pitfalls we as scientist may face when designing therapeutic strategies with miRNAs.
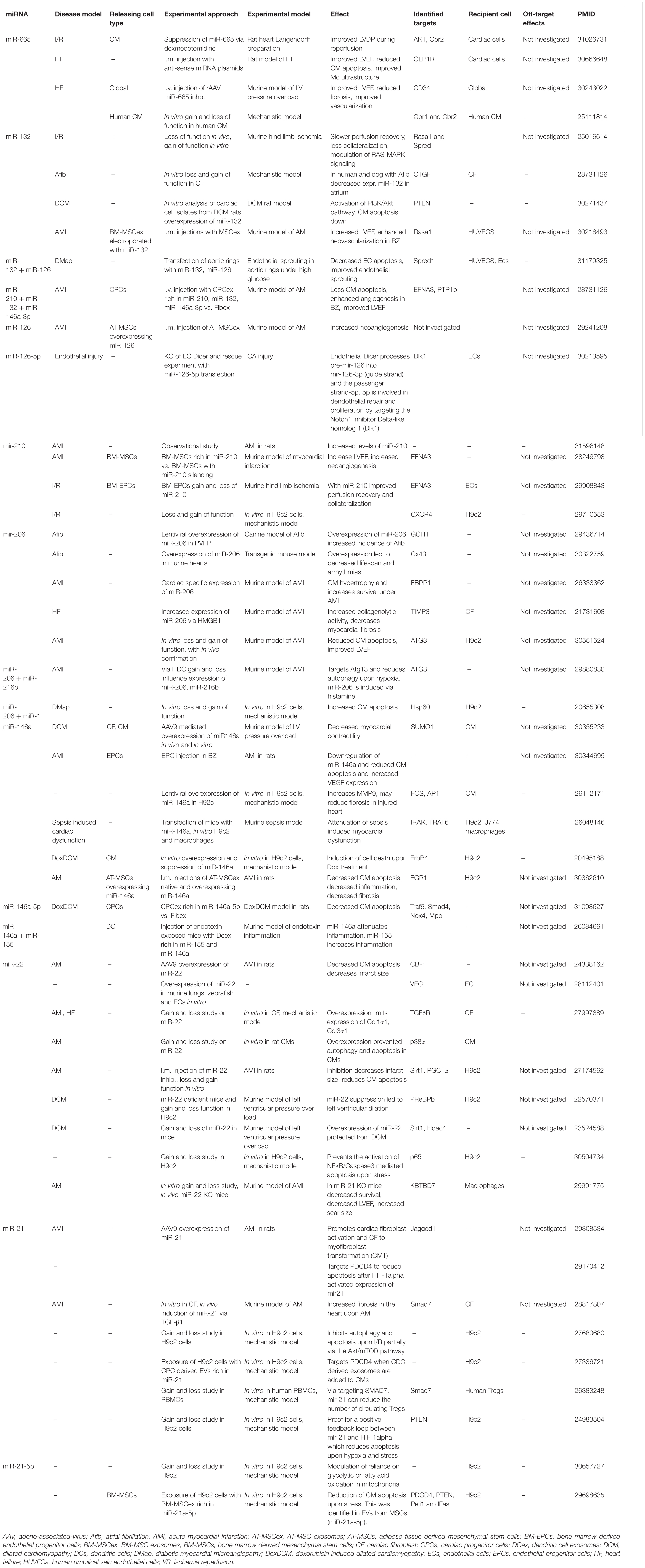
Table 1. Summarizes a selection of miRNAs that have been identified with either a harmful or cardioprotective property in cardiovascular disease.
MicroRNAs – The Good, the Bad, and the Ugly
Off-Target Effects of miRNAs
Especially studies investigating RIPC have identified clusters of cardioprotective miRNAs (Varga et al., 2015). In these studies as well as in those that used EVs, an undefined cocktail of miRNAs was systemically applied. In both cases, defining the mode of action is virtually impossible. Usually, hundreds of miRNAs can be identified in RIPC and even in EV preparations numerous miRNAs can be found, some of which taken by themselves have been identified as damaging to the myocardium. As an example, miR-665 has been identified in sera and serum exosomes of patients with heart failure (Li et al., 2016; Fan et al., 2019). MiR-665 directly targets the cannabinoid receptor 2 (CbR2) and adenylate kinase 1 (AK1) (Möhnle et al., 2014; Lin et al., 2019). In a rat heart Langendorff preparation, inhibition of CbR2 and AK1 leads to the upregulation of pro-apoptotic genes such as B cell lymphoma 2 (Bcl-2) and Bcl-2-associated X protein (Bax) and caspase-3 in H9c2 cells, a commonly used rat cardiomyocyte cell line (Yu et al., 2019). In a murine model of myocardial infarction, AAV9 transfection with miR-665 antisense miRNA led to improvement of cardiac function (Fan et al., 2018). Here, the group has identified glucagon-like peptide-1 receptor (GLP1R) as the target for miR-665. Inhibition of miR-665 expression led to increased cAMP signaling in the heart via the promotor GLP1R and reduced apoptotic events upon ischemia/reperfusion injury in the heart. Identification of harmful miRNAs like miRNA-665 in autologous exosome preparations from patients with heart failure could serve as quality markers and help prevent potential off-target effects. In some cases, off target effects or cardioprotective miRNAs have already been identified. miRNA-206, for example, has been associated with both cardioprotective, but also damaging effects upon overexpression (Table 1). Transcription of miR-206 can be induced via histamine release upon myocardial stress in mice (Ding et al., 2018). In vitro, miR-206 can prevent apoptosis in hypoxic conditions by targeting autophagy related protein 2 (ATG3) (Kong et al., 2019). In a murine model of acute myocardial infarction, this prevented ubiquitination of cytosomal proteins and apoptosis (Ding et al., 2018). By cardiac-specific overexpression of miR-206 is has been shown that forkhead box protein P1 induces hypertrophy of cardiomyocytes which during stress can prevent cardiomyocyte apoptosis (Yang et al., 2015). By inhibiting metalloproteinase inhibitor 3 (TIMP3), miR-206 has also been shown to attenuate cardiac fibrosis in the setting of chronic heart failure (Limana et al., 2011). However, two independent groups have also uncovered two mechanisms by which miR-206 increases the risk of cardiac arrhythmias. miR-206 targets connexin 43 (Cx43), an important component of gap junctions in the myocardium in a transgenic mouse model (Roell et al., 2007). Low expression of Cx43 has been associated with cardiac arrhythmias such as atrial fibrillation. Additionally, Wei et al. (2018) uncovered that miR-206 binds to GTP cyclohydrolase I (GCH1) in a canine model of atrial fibrillation (Afib). GCH1 is the rate limiting enzyme in de novo synthesis of tetrahydrobiopterin (BH4). Decreased expression of BH4 was associated with shortened refractory times in atrial cardiomyocytes in humans, which led to Afib (Wei et al., 2018). In this example, different groups were able to demonstrate a cardioprotective effect of miR-206. At the same time potential off target effects were uncovered that could impede the translation of important signal transduction proteins in the myocardium. These off-target effects were never investigated in the murine models that investigated the cardioprotective effects for this miRNA. It has also been shown that miRNA have species specific effects. A recently published large animal study demonstrated that the delivery of miR-199 via adenoviral transfer to the myocardium of pigs resulted in improved contractility and myocardial mass in the short term (Gabisonia et al., 2019). After 1 month, however, the pigs died of arrhythmias. Histological analysis revealed that the myocardium was infiltrated with proliferating cells displaying a poorly differentiated myoblastic phenotype. This off-target effect of miR-199 was not identified in small animal studies. Results like these raise the question whether results from murine models or even porcine models are really translatable into clinical applications without additional safeguards that can predict these off-target and adverse effects.
Neoangiogenesis – Good for Cardioprotection, Bad for Cancer Progression
Neoangiogenesis plays an important role in protecting the myocardium in the border zones from infarcts from myocardial remodeling. Therefore, many miRNAs such as miR-132, miR-126, and miR-210 have been investigated for their angiogenic potential (Table 1). The transcription of most of these miRNAs is dependent on the expression VEGF (Lei et al., 2015; Chodari et al., 2019). For instance, miR-132 expression is dependent in the promotor cAMP response element-binding protein as the transcription factor, which is induced by VEGF stimulation (Chodari et al., 2019). EVs from cardiac progenitor cells (CPCs) rich in miR-132 inhibit the translation of Ras GTPase activating protein (p120RasGAP) which promotes proliferation and sprouting of endothelial cells thus improving neovascularization in vivo and in vitro (Barile et al., 2014). In a murine model of acute myocardial infarction this led to improved left ventricular ejection fraction on a functional level and improved vascularization in the infarct border zone on a histological level (Barile et al., 2014). Similarly, EVs from adipose-derived mesenchymal stem cells (AT-MSCs) that overexpressed miR-126, also improved cardiac function and resulted in a denser microvasculature in the infarct border zone in rats (Luo et al., 2017). Functional studies have shown that miR-126 is highly depended on the activation of endothelial dicer RNA Polymerase III. Only the passenger strand miR-126-5p can bind to the Notch1 inhibitor Delta-like homolog 1 (Dlk1) (Zhou Z. et al., 2018). The notch signaling pathway is crucial to endothelial cell differentiation and endothelial sprouting (Mack and Iruela-Arispe, 2018). While there is no direct evidence linking the cardioprotective effect of miR-132 to cardiomyocytes, there is evidence that miR-132 can also reduce fibrosis in by targeting connective tissue growth factor (Zhang C.-J. et al., 2018). Furthermore, miR-132 has also been identified in a rat model of DCM to target phosphatase and tensin homolog (PTEN) (Ma et al., 2018). Suppression of PTEN activates the phosphoinositide 3-kinases/protein kinase B pathway (PI3K/Akt-pathway) which facilitates cardiomyocyte and endothelial proliferation alike (Zhang C.-J. et al., 2018). miR-210 has also been shown to promote angiogenesis in the myocardium (Mutharasan et al., 2011). In contrast to miR-126 and miR132, the expression of miR-210 depends on HIF-1alpha, which is also released under hypoxic stress (Barile et al., 2014). In both murine models of acute myocardial infarction and hind limb ischemia it has been shown that miR-210 encapsulated by EVs promotes angiogenesis in endothelial cells as well as suppresses apoptosis in cardiomyocytes by targeting Ephrin A3 (Barile et al., 2014; Wang et al., 2017; Besnier et al., 2018). While the upregulation or substitution of all aforementioned miRNAs are associated with a pro-angiogenic profile in the setting of myocardial infarction, they have also been identified in cancer biogenesis and metastasis formation. Also, in the field of cancer biology the data for these miRNAs is heterogeneous and in depending on cancer type they are both associated as a positive and negative prognostic marker. The heterogeneity of these results and their role in tumor progression could, however, pose an obstacle for their use as a cardioprotective agent. As a solution, patients susceptible to certain cancers that depend on overexpression of these miRNA need to be identified to prevent adverse effects from a hypothetical therapeutic miRNA.
How to Deal With Contradictory Results
miR-146a is part of a negative feedback loop in the canonical pathway of NFkB activation. miR-146a binds to the mRNA encoding for interleukin-1 receptor associated kinase 1 (IRAK1) and tumor necrosis factor receptor-associated receptor 6 (TRAF6). Both of this receptor bound factors are essential for the IL-1 and TNFalpha activation of NFkB (Fish and Cybulsky, 2015; Gao et al., 2015; Milano et al., 2019). As part of that negative feedback loop miR-146a is highly expressed in atherosclerotic plaques. However, the overexpression of this miRNA can also attenuate the inflammatory response as shown in gain and loss studies in ApoE deficient mice, where miRNA-146a plays an important part in the attenuation of atherosclerotic plaques (Fish and Cybulsky, 2015). In a rat model of AMI, EVs from AT-MSCs overexpressing miR-146a targeted the early growth response protein 1 in cardiomyocytes decreasing cardiomyocyte apoptosis, cardiac fibrosis and ultimately improving the heart function (Pan et al., 2019). Milano et al. (2019) demonstrated that the passenger strand miR-146a-5p reduced the inflammatory signaling pathways by directly targeting TRAF6, SMAD4, IRAK1, NADPH oxidase 4 (NOX4), and myeloperoxidase (MPO) in a model of doxorubicin/trastuzumab induced cardiomyopathy. Interestingly, another group showed that with treatment of doxorubicin alone an upregulation of miR146a occurs, targeting the receptor tyrosine-protein kinase erbB-4 (ErbB4). Here, a negative correlation between miR-146a and cardiac function was confirmed and explained by the suppression of the ErbB4 dependent neuregulin1/ErbB pathway, which is essential for adult cardiac function (Horie et al., 2010). It is, however, important to note that these results were only obtained from in vitro experiments. Both miR-21 and -22 have been associated with cardioprotective properties by reducing cardiomyocyte apoptosis and have been found in most EVs from therapeutic cell product isolates to date (Barile et al., 2016; Gallet et al., 2016). While there is evidence, that miR-22 also exhibits an anti-fibrotic effect during myocardial remodeling, overexpression of miR-21 is clearly associated with promoting cardiac fibrosis (Table 1). miR-21 directly targets jagged1 and SMAD7 in rat hearts when overexpressed via AAV9 (Zhou X.-L. et al., 2018). In the aforementioned experiment, jagged1 suppression activates cardiac fibroblast proliferation and facilitates cardiac fibroblast to myofibroblast transformation. In a murine model of AMI, suppression of SMAD7 via miR-21 led to an increased expression of Collagen 1 alpha, alpha-smooth muscle actin (alpha-SMA) and F-actin. In a similar model of AMI in rats, AVV9 mediated overexpression of miR-22 let to the inhibition of CBP-associated factor AP1 (Yang et al., 2014). Downregulation of this promotor leads to the activation of MMP9, which in turn can reduce cardiac fibrosis. Additionally, miR-22 binds to the mRNA coding for the TGFbeta receptor 1 (Hong et al., 2016). In mice, silencing of miR-22 led to increased expression of Collagen 1 alpha 1 and 3alpha1 and an overall increased amount of cardiac fibrosis after myocardial infarction. Regarding the anti-apoptotic effect of miR-22, there are contradictory results published to date. Gurha et al. (2012) and Du et al. (2016) both demonstrated that SIRT1 and PGC1alpha are both targeted by miR-22 in cardiomyocytes. In both experiments, overexpression of miR-22 lead to increased cell death and reduction in myocardial mass whereas in the studies published by Huang et al. (2013) and Hong et al. (2016) cardiac mass increased upon miR-22 overexpression and ischemia/reperfusion injury. Huang et al. (2013) also demonstrate the targeting of SIRT1 but in their hands, cardiomyocyte hypertrophy was the predominant finding. Hong et al. (2016) demonstrated decreased apoptosis levels in the myocardium and linked that effect to the inhibition of CREB. For miR-21 the anti-apoptotic effects is shown more robustly between different research teams (Table 1). miR-21 expression is under direct control of HIF1alpha but can also influence the expression of HIF1alpha itself in a positive feedback loop (Liu Y. et al., 2014). The miRNA targets programmed cell death protein 4 (PCDP4) and reduces cell apoptosis upon hypoxic stress. This effect has been demonstrated with EVs containing high levels of miR-21 from different cell types such as CPCs and MSCs (Xiao et al., 2016; Luther et al., 2018). Both miR-21 and miR-22 exemplify the various outcomes that can be seen when working with miRNAs in pre-clinical models. Even within one species there are variations and sometime contradictory results that can be elaborated by other groups on both ends of the aisle. Especially miRNAs that can promote fibrosis can have severe and unwanted effects in the injured heart and may lead to increase in scar mass. Here, patient screening and good patient selection may help to prevent these contradictory results. This can only be achieved by understanding all pathways that can be altered by each respective miRNA and tools to identify patients that may be susceptible to treatments with certain miRNAs.
Summary
The data that has been collected on miRNAs targeting cardioprotection so far exemplifies the importance of knowing the relevant targets of miRNAs, since introduction of foreign miRNAs via exosomal transfer or other clinical relevant approaches may have some severe side effects. In this context, it is also worth noting that all of the aforementioned miRNAs are also involved in tumor biology. Especially miRNAs that impact neoangiogenesis in myocardial repair are important factors in tumor angiogenesis as well. In addition, most of the mechanistical studies have focused on the interaction of one miRNA on multiple targets. Upon ischemia/reperfusion or transfer of exogenous EVs the interaction of numerous miRNAs on multiple potential targets needs to be taken into account. With current methods, experimental data usually depicts a single linear arm in a complex matrix of interactions between miRNAs, mRNAs and transcription factors. Our current understanding of these complex matrices is only rudimental at best. And it may also be one of the main reasons why none of the clinical trials with cell-based therapies or RIPC have delivered the expected results we anticipated from pre-clinical experience. As repeatedly highlighted by numerous experts in the field, understanding the pathways by which a single- or a collection of miRNAs in an exosome, facilitate cardioprotection will be crucial for successful clinical translation (Madonna et al., 2016, 2019; Behfar and Terzic, 2019). Combined effort of computational models and artificial intelligence in merging and interpreting the acquired data, might help us in the future to achieve this goal. Identifying such ‘pathway’ matrices will be detrimental in defining quality standards for therapeutic exosome or single miRNA-based products. The past experience from cell-based therapies have taught us that preclinical data in the field of cardiac regeneration or cardioprotection does not necessarily translate into therapeutic success in the clinical setting. Going forward, we will have to deepen our understanding of miRNA interactions by strengthen our efforts to collaborate with bioinformaticians for more sophisticated predictive algorithms.
Author Contributions
TN-S and ME contributed to the conception and design of this review. VE, HR, and HM helped to organize the article database and supported the literature review. TN-S wrote the first draft of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
TN-S is a participant in the BIH Charité Clinician Scientist Program funded by the Charité -Universitätsmedizin Berlin and the Berlin Institute of Health.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Baglio, S. R., Rooijers, K., Koppers-Lalic, D., Verweij, F. J., Pérez Lanzón, M., Zini, N., et al. (2015). Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 6:127. doi: 10.1186/s13287-015-0116-z
Barile, L., Lionetti, V., Cervio, E., Matteucci, M., Gherghiceanu, M., Popescu, L. M., et al. (2014). Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovasc. Res. 103, 530–541. doi: 10.1093/cvr/cvu167
Barile, L., Moccetti, T., Marbán, E., and Vassalli, G. (2016). Roles of exosomes in cardioprotection. Eur. Heart J. 38, 1372–1379. doi: 10.1093/eurheartj/ehw304
Barile, L., and Vassalli, G. (2017). Exosomes: therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 174, 63–78. doi: 10.1016/j.pharmthera.2017.02.020
Bartel, D. P. (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281–297. doi: 10.1016/S0092-8674(04)00045-5
Behfar, A., Crespo-Diaz, R., Terzic, A., and Gersh, B. J. (2014). Cell therapy for cardiac repair–lessons from clinical trials. Nat. Rev. Cardiol. 11, 232–246. doi: 10.1038/nrcardio.2014.9
Behfar, A., and Terzic, A. (2019). Regeneration for all: an Odyssey in biotherapy. Eur. Heart J. 40, 1033–1035. doi: 10.1093/eurheartj/ehz095
Bellin, G., Gardin, C., Ferroni, L., Chachques, J., Rogante, M., Mitrečić, D., et al. (2019). Exosome in cardiovascular diseases: a complex world full of hope. Cells 8:166. doi: 10.3390/cells8020166
Besnier, M., Gasparino, S., Vono, R., Sangalli, E., Facoetti, A., Bollati, V., et al. (2018). miR-210 enhances the therapeutic potential of bone-marrow-derived circulating proangiogenic cells in the setting of limb ischemia. Mol. Ther. 26, 1694–1705. doi: 10.1016/j.ymthe.2018.06.003
Boon, R. A., and Dimmeler, S. (2015). MicroRNAs in myocardial infarction. Nat. Rev. Cardiol. 12, 135–142. doi: 10.1038/nrcardio.2014.207
Brevoord, D., Kranke, P., Kuijpers, M., Weber, N., Hollmann, M., and Preckel, B. (2012). Remote ischemic conditioning to protect against ischemia-reperfusion injury: a systematic review and meta-analysis. PLoS One 7:e42179. doi: 10.1371/journal.pone.0042179
Cai, X., Hagedorn, C. H., and Cullen, B. R. (2004). Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 10, 1957–1966. doi: 10.1261/rna.7135204
Cambria, E., Pasqualini, F. S., Wolint, P., Günter, J., Steiger, J., Bopp, A., et al. (2017). Translational cardiac stem cell therapy: advancing from first-generation to next-generation cell types. npj Regen. Med. 2:17. doi: 10.1038/s41536-017-0024-1
Chen, L., Tredget, E. E., Wu, P. Y. G., Wu, Y., and Wu, Y. (2008). Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS One 3:e1886. doi: 10.1371/journal.pone.0001886
Chodari, L., Dariushnejad, H., and Ghorbanzadeh, V. (2019). Voluntary wheel running and testosterone replacement increases heart angiogenesis through miR-132 in castrated diabetic rats. Physiol. Int. 106, 48–58. doi: 10.1556/2060.106.2019.06
Ciullo, A., Biemmi, V., Milano, G., Bolis, S., Cervio, E., Fertig, E. T., et al. (2019). Exosomal expression of CXCR4 targets cardioprotective vesicles to myocardial infarction and improves outcome after systemic administration. Int. J. Mol. Sci. 20:468. doi: 10.3390/ijms20030468
Davis, B. N., Hilyard, A. C., Lagna, G., and Hata, A. (2008). SMAD proteins control DROSHA-mediated microRNA maturation. Nature 454, 56–61. doi: 10.1038/nature07086
Diederichs, S., and Haber, D. A. (2007). Dual role for argonautes in microRNA processing and posttranscriptional regulation of microRNA expression. Cell 131, 1097–1108. doi: 10.1016/j.cell.2007.10.032
Diederichs, S., Jung, S., Rothenberg, S. M., Smolen, G. A., Mlody, B. G., and Haber, D. A. (2008). Coexpression of Argonaute-2 enhances RNA interference toward perfect match binding sites. Proc. Natl. Acad. Sci. U.S.A. 105, 9284–9289. doi: 10.1073/pnas.0800803105
Ding, S., Abudupataer, M., Zhou, Z., Chen, J., Li, H., Xu, L., et al. (2018). Histamine deficiency aggravates cardiac injury through miR-206/216b-Atg13 axis-mediated autophagic-dependant apoptosis. Cell Death Dis. 9:694. doi: 10.1038/s41419-018-0723-6
Du, J.-K., Cong, B.-H., Yu, Q., Wang, H., Wang, L., Wang, C.-N., et al. (2016). Upregulation of microRNA-22 contributes to myocardial ischemia-reperfusion injury by interfering with the mitochondrial function. Free Radic. Biol. Med. 96, 406–417. doi: 10.1016/j.freeradbiomed.2016.05.006
Fan, J., Li, H., Nie, X., Yin, Z., Zhao, Y., Zhang, X., et al. (2018). MiR-665 aggravates heart failure via suppressing CD34-mediated coronary microvessel angiogenesis. Aging (Albany NY) 10, 2459–2479. doi: 10.18632/aging.101562
Fan, J., Zhang, X., Nie, X., Li, H., Yuan, S., and Dai, B., et al., (2019). Nuclear miR-665 aggravates heart failure via suppressing phosphatase and tensin homolog transcription. Sci. China Life Sci. 1–13. doi: 10.1007/s11427-018-9515-1
Fish, J. E., and Cybulsky, M. I. (2015). ApoE attenuates atherosclerosis via miR-146a. Circ. Res. 117, 3–6. doi: 10.1161/CIRCRESAHA.115.306733
Frey, U. H., Klaassen, M., Ochsenfarth, C., Murke, F., Thielmann, M., Kottenberg, E., et al. (2018). Remote ischaemic preconditioning increases serum extracellular vesicle concentrations with altered micro−RNA signature in CABG patients. Acta Anaesthesiol. Scand. 63, 483–492. doi: 10.1111/aas.13296
Gabisonia, K., Prosdocimo, G., Aquaro, G. D., Carlucci, L., Zentilin, L., Secco, I., et al. (2019). MicroRNA therapy stimulates uncontrolled cardiac repair after myocardial infarction in pigs. Nature 569, 418–422. doi: 10.1038/s41586-019-1191-6
Gallet, R., Dawkins, J., Valle, J., Simsolo, E., de Couto, G., Middleton, R., et al. (2016). Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 38, 201–211. doi: 10.1093/eurheartj/ehw240
Gao, M., Wang, X., Zhang, X., Ha, T., Ma, H., Liu, L., et al. (2015). Attenuation of cardiac dysfunction in polymicrobial sepsis by MicroRNA-146a is mediated via targeting of IRAK1 and TRAF6 expression. J. Immunol. 195, 672–682. doi: 10.4049/jimmunol.1403155
Gregory, R. I., Chendrimada, T. P., Cooch, N., and Shiekhattar, R. (2005). Human RISC couples microRNA biogenesis and posttranscriptional gene silencing. Cell 123, 631–640. doi: 10.1016/j.cell.2005.10.022
Guil, S., and Cáceres, J. F. (2007). The multifunctional RNA-binding protein hnRNP A1 is required for processing of miR-18a. Nat. Struct. Mol. Biol. 14, 591–596. doi: 10.1038/nsmb1250
Gurha, P., Abreu-Goodger, C., Wang, T., Ramirez, M. O., Drumond, A. L., Van Dongen, S., et al. (2012). Targeted deletion of MicroRNA-22 promotes stress-induced cardiac dilation and contractile dysfunction. Circulation 125, 2751–2761. doi: 10.1161/CIRCULATIONAHA.111.044354
Gyöngyösi, M., Wojakowski, W., Lemarchand, P., Lunde, K., Tendera, M., Bartunek, J., et al. (2015). Meta-analysis of cell-based CaRdiac stUdiEs (ACCRUE) in patients with acute myocardial infarction based on individual patient data. Circ. Res. 116, 1346–1360. doi: 10.1161/CIRCRESAHA.116.304346
Hausenloy, D. J., Candilio, L., Evans, R., Ariti, C., Jenkins, D. P., Kolvekar, S., et al. (2015). Remote ischemic preconditioning and outcomes of cardiac surgery. N. Engl. J. Med. 373, 1408–1417. doi: 10.1056/NEJMoa1413534
Hong, Y., Cao, H., Wang, Q., Ye, J., Sui, L., Feng, J., et al. (2016). MiR-22 may suppress fibrogenesis by targeting TGFβR I in cardiac fibroblasts. Cell. Physiol. Biochem. 40, 1345–1353. doi: 10.1159/000453187
Horie, T., Ono, K., Nishi, H., Nagao, K., Kinoshita, M., Watanabe, S., et al. (2010). Acute doxorubicin cardiotoxicity is associated with miR-146a-induced inhibition of the neuregulin-ErbB pathway. Cardiovasc. Res. 87, 656–664. doi: 10.1093/cvr/cvq148
Huang, Z.-P., Chen, J., Seok, H. Y., Zhang, Z., Kataoka, M., Hu, X., et al. (2013). MicroRNA-22 regulates cardiac hypertrophy and remodeling in response to stress. Circ. Res. 112, 1234–1243. doi: 10.1161/CIRCRESAHA.112.300682
Hutvágner, G., McLachlan, J., Pasquinelli, A. E., Bálint, É, Tuschl, T., and Zamore, P. D. (2001). A cellular function for the RNA-interference enzyme dicer in the maturation of the let-7 small temporal RNA. Science 293, 834–838. doi: 10.1126/science.1062961
Kharbanda, R. K., Mortensen, U. M., White, P. A., Kristiansen, S. B., Schmidt, M. R., Hoschtitzky, J. A., et al. (2002). Transient limb ischemia induces remote ischemic preconditioning in vivo. Circulation 106, 2881–2883. doi: 10.1161/01.CIR.0000043806.51912.9B
Kong, F., Jin, J., Lv, X., Han, Y., Liang, X., Gao, Y., et al. (2019). Long noncoding RNA RMRP upregulation aggravates myocardial ischemia-reperfusion injury by sponging miR-206 to target ATG3 expression. Biomed. Pharmacother. 109, 716–725. doi: 10.1016/j.biopha.2018.10.079
Konstantinov, I. E., Arab, S., Li, J., Coles, J. G., Boscarino, C., Mori, A., et al. (2005). The remote ischemic preconditioning stimulus modifies gene expression in mouse myocardium. J. Thorac. Cardiovasc. Surg. 130, 1326–1332. doi: 10.1016/j.jtcvs.2005.03.050
Lee, R. C., Feinbaum, R. L., and Ambros, V. (1993). The C. elegans heterochronic gene lin-4 encodes small rnas with antisense complementarity to lin-14. Cell 75, 843–854. doi: 10.1016/0092-8674(93)90529-Y
Lee, Y., Ahn, C., Han, J., Choi, H., Kim, J., Yim, J., et al. (2003). The nuclear RNase III Drosha initiates microRNA processing. Nature 425, 415–419. doi: 10.1038/nature01957
Lee, Y., Kim, M., Han, J., Yeom, K. H., Lee, S., Baek, S. H., et al. (2004). MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 23, 4051–4060. doi: 10.1038/sj.emboj.7600385
Lei, Z., van Mil, A., Brandt, M. M., Grundmann, S., Hoefer, I., Smits, M., et al. (2015). MicroRNA-132/212 family enhances arteriogenesis after hindlimb ischaemia through modulation of the Ras-MAPK pathway. J. Cell. Mol. Med. 19, 1994–2005. doi: 10.1111/jcmm.12586
Leuschner, P. J. F., Ameres, S. L., Kueng, S., and Martinez, J. (2006). Cleavage of the siRNA passenger strand during RISC assembly in human cells. EMBO Rep. 7, 314–320. doi: 10.1038/sj.embor.7400637
Li, H., Fan, J., Yin, Z., Wang, F., Chen, C., and Wang, D. W. (2016). Identification of cardiac-related circulating microRNA profile in human chronic heart failure. Oncotarget 7, 33–45. doi: 10.18632/ONCOTARGET.6631
Li, L., Zhu, D., Huang, L., Zhang, J., Bian, Z., Chen, X., et al. (2012). Argonaute 2 complexes selectively protect the circulating MicroRNAs in cell-secreted microvesicles. PLoS One 7:e46957. doi: 10.1371/journal.pone.0046957
Limana, F., Esposito, G., D’Arcangelo, D., Di Carlo, A., Romani, S., Melillo, G., et al. (2011). HMGB1 attenuates cardiac remodelling in the failing heart via enhanced cardiac regeneration and miR-206-mediated inhibition of TIMP-3. PLoS One 6:e19845. doi: 10.1371/journal.pone.0019845
Lin, B., Feng, D. G., and Xu, J. (2019). microRNA-665 silencing improves cardiac function in rats with heart failure through activation of the cAMP signaling pathway. J. Cell. Physiol. 234, 13169–13181. doi: 10.1002/jcp.27987
Lin, S. L., Miller, J. D., and Ying, S. Y. (2006). Intronic microRNA (miRNA). J. Biomed. Biotechnol 2006:26818. doi: 10.1155/JBB/2006/26818
Liu, B., Li, J., and Cairns, M. J. (2014). Identifying miRNAs, targets and functions. Brief. Bioinform. 15, 1–19. doi: 10.1093/bib/bbs075
Liu, Y., Nie, H., Zhang, K., Ma, D., Yang, G., Zheng, Z., et al. (2014). A feedback regulatory loop between HIF-1α and miR-21 in response to hypoxia in cardiomyocytes. FEBS Lett. 588, 3137–3146. doi: 10.1016/j.febslet.2014.05.067
Luo, Q., Guo, D., Liu, G., Chen, G., Hang, M., and Jin, M. (2017). Exosomes from MiR-126-overexpressing Adscs Are therapeutic in relieving acute myocardial ischaemic injury. Cell. Physiol. Biochem. 44, 2105–2116. doi: 10.1159/000485949
Luther, K. M., Haar, L., McGuinness, M., Wang, Y., Lynch Iv, T. L., Phan, A., et al. (2018). Exosomal miR-21a-5p mediates cardioprotection by mesenchymal stem cells. J. Mol. Cell. Cardiol. 119, 125–137. doi: 10.1016/j.yjmcc.2018.04.012
Ma, T., Chen, Y., Chen, Y., Meng, Q., Sun, J., Shao, L., et al. (2018). MicroRNA-132, delivered by mesenchymal stem cell-derived exosomes, promote angiogenesis in myocardial infarction. Stem Cells Int. 2018:3290372. doi: 10.1155/2018/3290372
Mack, J. J., and Iruela-Arispe, M. L. (2018). NOTCH regulation of the endothelial cell phenotype. Curr. Opin. Hematol. 25, 212–218. doi: 10.1097/MOH.0000000000000425
MacRae, I. J., Ma, E., Zhou, M., Robinson, C. V., and Doudna, J. A. (2008). In vitro reconstitution of the human RISC-loading complex. Proc. Natl. Acad. Sci. U.S.A. 105, 512–517. doi: 10.1073/pnas.0710869105
Madonna, R., Van Laake, L. W., Botker, H. E., Davidson, S. M., De Caterina, R., Engel, F. B., et al. (2019). ESC working group on cellular biology of the heart: position paper for cardiovascular research: tissue engineering strategies combined with cell therapies for cardiac repair in ischaemic heart disease and heart failure. Cardiovasc. Res. 115, 488–500. doi: 10.1093/cvr/cvz010
Madonna, R., Van Laake, L. W., Davidson, S. M., Engel, F. B., Hausenloy, D. J., Lecour, S., et al. (2016). Position paper of the European society of cardiology working group cellular biology of the heart: cell-based therapies for myocardial repair and regeneration in ischemic heart disease and heart failure. Eur. Heart J. 37, 1789–1798. doi: 10.1093/eurheartj/ehw113
Maring, J. A., Lodder, K., Mol, E., Verhage, V., Wiesmeijer, K. C., Dingenouts, C. K. E., et al. (2019). Cardiac progenitor cell–derived extracellular vesicles reduce infarct size and associate with increased cardiovascular cell proliferation. J. Cardiovasc. Transl. Res. 12, 5–17. doi: 10.1007/s12265-018-9842-9
Matranga, C., Tomari, Y., Shin, C., Bartel, D. P., and Zamore, P. D. (2005). Passenger-strand cleavage facilitates assembly of siRNA into Ago2-containing RNAi enzyme complexes. Cell 123, 607–620. doi: 10.1016/j.cell.2005.08.044
Michlewski, G., Guil, S., Semple, C. A., and Cáceres, J. F. (2008). Posttranscriptional regulation of miRNAs harboring conserved terminal loops. Mol. Cell 32, 383–393. doi: 10.1016/j.molcel.2008.10.013
Milano, G., Biemmi, V., Lazzarini, E., Balbi, C., Ciullo, A., Bolis, S., et al. (2019). Intravenous administration of cardiac progenitor cell-derived exosomes protects against doxorubicin/trastuzumab-induced cardiac toxicity. Cardiovasc. Res. 116, 383–392. doi: 10.1093/cvr/cvz108
Möhnle, P., Schütz, S. V., Schmidt, M., Hinske, C., Hübner, M., Heyn, J., et al. (2014). MicroRNA-665 is involved in the regulation of the expression of the cardioprotective cannabinoid receptor CB2 in patients with severe heart failure. Biochem. Biophys. Res. Commun. 451, 516–521. doi: 10.1016/j.bbrc.2014.08.008
Moyé, L. (2014). DAMASCENE and meta-ecological research: a bridge too far. Circ. Res. 115, 484–487. doi: 10.1161/CIRCRESAHA.114.304767
Mutharasan, R. K., Nagpal, V., Ichikawa, Y., and Ardehali, H. (2011). microRNA-210 is upregulated in hypoxic cardiomyocytes through Akt- and p53-dependent pathways and exerts cytoprotective effects. Am. J. Physiol. Circ. Physiol. 301, H1519–H1530. doi: 10.1152/ajpheart.01080.2010
Pan, J., Alimujiang, M., Chen, Q., Shi, H., and Luo, X. (2019). Exosomes derived from miR-146a-modified adipose-derived stem cells attenuate acute myocardial infarction-induced myocardial damage via downregulation of early growth response factor 1. J. Cell. Biochem. 120, 4433–4443. doi: 10.1002/jcb.27731
Rodriguez, A., Griffiths-Jones, S., Ashurst, J. L., and Bradley, A. (2004). Identification of mammalian microRNA host genes and transcription units. Genome Res. 14, 1902–1910. doi: 10.1101/gr.2722704
Roell, W., Lewalter, T., Sasse, P., Tallini, Y. N., Choi, B.-R., Breitbach, M., et al. (2007). Engraftment of connexin 43-expressing cells prevents post-infarct arrhythmia. Nature 450, 819–824. doi: 10.1038/nature06321
Shao, L., Zhang, Y., Lan, B., Wang, J., Zhang, Z., Zhang, L., et al. (2017). MiRNA-sequence indicates that mesenchymal stem cells and exosomes have similar mechanism to enhance cardiac repair. Biomed Res. Int. 2017:4150705. doi: 10.1155/2017/4150705
Spannbauer, A., Traxler, D., Lukovic, D., Zlabinger, K., Winkler, J., Gugerell, A., et al. (2019). Effect of ischemic preconditioning and postconditioning on exosome-rich fraction microRNA levels, in relation with electrophysiological parameters and ventricular arrhythmia in experimental closed-chest reperfused myocardial infarction. Int. J. Mol. Sci. 20:2140. doi: 10.3390/ijms20092140
Turchinovich, A., Weiz, L., and Burwinkel, B. (2012). Extracellular miRNAs: the mystery of their origin and function. Trends Biochem. Sci. 37, 460–465. doi: 10.1016/j.tibs.2012.08.003
Tzahor, E., and Poss, K. D. (2017). Cardiac regeneration strategies: staying young at heart. Science 356, 1035–1039. doi: 10.1126/science.aam5894
Varga, Z. V., Giricz, Z., Bencsik, P., Madonna, R., Gyongyosi, M., and Schulz, R., et al., (2015). Functional genomics of cardioprotection by ischemic conditioning and the influence of comorbid conditions: implications in target identification. Curr. Drug Targets 16, 904–911. doi: 10.2174/1389450116666150427154203
Villarroya-Beltri, C., Gutiérrez-Vázquez, C., Sánchez-Cabo, F., Pérez-Hernández, D., Vázquez, J., Martin-Cofreces, N., et al. (2013). Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 4:2980. doi: 10.1038/ncomms3980
Wang, N., Chen, C., Yang, D., Liao, Q., Luo, H., Wang, X., et al. (2017). Mesenchymal stem cells-derived extracellular vesicles, via miR-210, improve infarcted cardiac function by promotion of angiogenesis. Biochim. Biophys. Acta Mol. Basis Dis. 1863, 2085–2092. doi: 10.1016/j.bbadis.2017.02.023
Wei, J., Zhang, Y., Li, Z., Wang, X., Chen, L., Du, J., et al. (2018). GCH1 attenuates cardiac autonomic nervous remodeling in canines with atrial-tachypacing via tetrahydrobiopterin pathway regulated by microRNA-206. Pacing Clin. Electrophysiol. 41, 459–471. doi: 10.1111/pace.13289
Wei, M., Xin, P., Li, S., Tao, J., Li, Y., Li, J., et al. (2011). Repeated remote ischemic postconditioning protects against adverse left ventricular remodeling and improves survival in a rat model of myocardial infarction. Circ. Res. 108, 1220–1225. doi: 10.1161/CIRCRESAHA.110.236190
Wendt, S., Goetzenich, A., Goettsch, C., Stoppe, C., Bleilevens, C., Kraemer, S., et al. (2018). Evaluation of the cardioprotective potential of extracellular vesicles – a systematic review and meta-analysis. Sci. Rep. 8:15702. doi: 10.1038/s41598-018-33862-5
Xiao, J., Pan, Y., Li, X. H., Yang, X. Y., Feng, Y. L., Tan, H. H., et al. (2016). Cardiac progenitor cell-derived exosomes prevent cardiomyocytes apoptosis through exosomal miR-21 by targeting PDCD4. Cell Death Dis. 7:e2277. doi: 10.1038/cddis.2016.181
Yang, J., Yang, J., Chen, L., Ding, J., Li, S., Wu, H., et al. (2014). MicroRNA-22 targeting CBP protects against myocardial ischemia-reperfusion injury through anti-apoptosis in rats. Mol. Biol. Rep. 41, 555–561. doi: 10.1007/s11033-013-2891-x
Yang, Y., Del Re, D. P., Nakano, N., Sciarretta, S., Zhai, P., Park, J., et al. (2015). miR-206 mediates YAP-induced cardiac hypertrophy and survival. Circ. Res. 117, 891–904. doi: 10.1161/CIRCRESAHA.115.306624
Yi, R., Qin, Y., Macara, I. G., and Cullen, B. R. (2003). Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev. 17, 3011–3016. doi: 10.1101/gad.1158803
Yu, J., Yang, W., Wang, W., Wang, Z., Pu, Y., Chen, H., et al. (2019). Involvement of miR-665 in protection effect of dexmedetomidine against oxidative stress injury in myocardial cells via CB2 and CK1. Biomed. Pharmacother. 115:108894. doi: 10.1016/j.biopha.2019.108894
Zeng, Y., and Cullen, B. R. (2004). Structural requirements for pre-microRNA binding and nuclear export by Exportin 5. Nucleic Acids Res. 32, 4776–4785. doi: 10.1093/nar/gkh824
Zhang, C.-J., Huang, Y., Lu, J.-D., Lin, J., Ge, Z.-R., and Huang, H. (2018). Upregulated microRNA-132 rescues cardiac fibrosis and restores cardiocyte proliferation in dilated cardiomyopathy through the phosphatase and tensin homolog-mediated PI3K/Akt signal transduction pathway. J. Cell. Biochem. 120, 1232–1244. doi: 10.1002/jcb.27081
Zhou, X.-L., Xu, H., Liu, Z.-B., Wu, Q.-C., Zhu, R.-R., and Liu, J.-C. (2018). miR-21 promotes cardiac fibroblast-to-myofibroblast transformation and myocardial fibrosis by targeting Jagged1. J. Cell. Mol. Med. 22, 3816–3824. doi: 10.1111/jcmm.13654
Keywords: microRNA, extracellular vesicles, second generation cell therapies, translation, cardioprotection, secretome
Citation: Nazari-Shafti TZ, Exarchos V, Rodriguez Cetina Biefer H, Cesarovic N, Meyborg H, Falk V and Emmert MY (2020) MicroRNA Mediated Cardioprotection – Is There a Path to Clinical Translation? Front. Bioeng. Biotechnol. 8:149. doi: 10.3389/fbioe.2020.00149
Received: 21 October 2019; Accepted: 13 February 2020;
Published: 20 March 2020.
Edited by:
Monica Boffito, Politecnico di Torino, ItalyReviewed by:
Zoltan Varga, Semmelweis University, HungaryMara Brancaccio, University of Turin, Italy
Copyright © 2020 Nazari-Shafti, Exarchos, Rodriguez Cetina Biefer, Cesarovic, Meyborg, Falk and Emmert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maximilian Y. Emmert, TWF4aW1pbGlhbi5FbW1lcnRAaXJlbS51emguY2g=; bWF4aW1pbGlhbi5lbW1lcnRAdXpoLmNo