 Lamya Rhayat1*Gaël Even2,3Tadele G. Kiros1Géraldine Kuhn1Ségolène Lebrun-Ruer2,3Christophe Audebert2,3Julie Schulthess1*
Lamya Rhayat1*Gaël Even2,3Tadele G. Kiros1Géraldine Kuhn1Ségolène Lebrun-Ruer2,3Christophe Audebert2,3Julie Schulthess1*- 1Phileo by Lesaffre, Rue de Menin, Marquettes-lez-Lille, France
- 2GD Biotech - Gènes Diffusion, Douai, France
- 3Plateforme d'Expertises Génomiques Appliquées aux Sciences Expérimentales (PEGASE)-Biosciences, Institut Pasteur de Lille, 1 Rue du Professeur Calmette, Lille, France
Introduction: Chronic diarrhoea is one of the most recurrent health issues in pig herds. Among Q9 the causative agents of diarrhoea, Lawsonia intracellularis is quite frequent with worldwide distribution leading to detrimental economical losses, as a result of the poor growth rate of the infected pigs. A growing body of evidence has shown that branched yeast b-glucans are excellent inducers of innate immunity memory in humans and in pigs.
Methods: In this study, yeast branched b-glucans were evaluated as a potential solution to control infection by L. intracellularis in pigs by reducing the shedding of L. intracellularis in the faeces of infected animals. To address this hypothesis, naturally infected grow finishing pigs were divided into two treatment groups (n=80) and fed either with a basal diet (Control) or the same basal diet supplemented with 250 g/ton b-glucans (BG-fed group).
Results: Our results have shown that BG-supplemented pigs during the growing and finishing period had significantly increased body weight (BW) by 4.22 kg on average on day 97 and ADG up to 5.6% over the whole experiment (p<5.0 x 10-2) with no significant increase in ADFI. Similarly, supplementation of pigs with BG during the grow-finish phase has significantly reduced (p<4.0x10-4) faecal shedding of the bacteria in infected pigs on day 97. Furthermore, our results indicate that microbial a-diversity was negatively correlated with the presence of L. intracellularis. Animals fed with yeast BG show a reduction of the two genera Sarcina and Catenibacterium and a deeper analysis indicated that the ratio of Fibrobacter to Lacnospiraceae_AC2044 group could be a potential marker for L. Intracellularis susceptibility.
Conclusion: This study demonstrated the beneficial effect of adding branched yeast BG in the diet of grow-finish pigs in reducing the dissemination of L. intracellularis between pigs in a given herd.
1 Introduction
Diarrhoea associated with enteric diseases caused by infectious bacterial and viral pathogens is a common problem in grow-finishing pigs, even on farms with excellent herd management practices (Karuppannan and Opriessnig, 2018). These infectious pathogens affect the health and production efficacy of pigs, leading to negative economic impacts on swine production. Among bacterial pathogens, the most important and prevalent pathogen worldwide is Lawsonia intracellularis (L. intracellularis) occurring mainly in post-weaned and growing pigs (Lawson and Gebhart, 2000). A study from Sweden showed that up to 67% of faecal samples collected from pigs with diarrhoea were positive for L. intracellularis (Jacobson et al., 2003). Similarly, up to 31% of enteric infections in diarrhoea in pig herds in Brazilian pig farms were also caused by L. intracellularis (Viott et al., 2013).L. intracellularis is an anaerobic obligate intracellular bacterium infecting actively dividing cells in the crypt of the small intestine of pigs and is responsible for the porcine proliferative enteropathy (PPE), also referred to as ileitis. The disease occurs either as an acute or chronic infection. The acute form, also known as proliferative haemorrhagic enteropathy (PHE), is characterized by bloody diarrhoea due to severe damage to the small intestine. For reasons not yet clear, the acute form of PPE is common in healthy herds, resulting in the sudden death of healthy adult pigs (Gilts and Boars). On the other hand, the chronic form of PPE, also known as Porcine Intestinal Adenomatosis (PIA), is characterized by weight loss and chronic diarrhoea. The diarrhoea is greyish in colour due to death and sloughing off the proliferating intestinal cells. During postmortem, the typical findings are thickened mucosa in the distal ileum and loss of goblet cells (Lawson and Gebhart, 2000; Kroll et al., 2005).
The economic impact of ileitis could be important, particularly in grow-finish pigs, resulting in a reduction in daily weight gain of approximately 3% to 19%, depending on the severity of the disease, and a decrease in refeed efficiency up to 7%. In acute cases, the disease may also kill 1,3% to 6,7% of the pigs within infected herds, leading to fewer and lighter finishing pigs. In addition to production loss, the cost incurred for treatment and prevention is considerable for farmers (Fourchon and Chouet, 2000; Holtkamp, 2019). In general, the monetary value of the diseases is estimated to be around $2.00 to $3.00 per pig in mild and subclinical cases, and as high as $15.00 per pig in severe cases (McOrist et al., 1997). Due to the ban of antibiotics in many countries and the limitations that currently available vaccines have to fully control ileitis, the swine industry is looking for alternative products to control infection or increase the potency of vaccines.
Recently, it was shown that the innate immune system could build innate immune memory responses after exposure to pathogens and, therefore, developed stronger and enhanced immune responses during secondary exposure. Innate immune memory, also called trained immunity, is non-specific and can mount protection against a cognate antigen or an unrelated pathogen (Netea et al., 2011; Netea et al., 2016). B-glucans (BG), an abundant branched polysaccharide located in the yeast cell wall, has been described as a strong inducer of trained immunity. In vitro studies using porcine monocytes exposed to BG showed stronger immune responses after exposure to microbial pathogen-associated molecular patterns (PAMPs), such as lipopolysaccharides (LPS) (Byrne et al., 2020). Furthermore, in vivo mouse studies using BG have demonstrated that macrophages develop better protection against Staphylococcus aureus (Quintin et al., 2012). Intestinal homeostasis is crucial for good performance of animals and is achieved by fine regulation of the intestinal barrier to reduce the risk of infectious or inflammatory conditions. Intestinal macrophages, which can be infected by L. intracellularis (Pereira et al., 2020), play an important role in host defence against invading pathogens (Bain and Schridde, 2018). Therefore, taking advantage of the ability of BG to induce trained immunity in intestinal macrophages could be an interesting strategy to reduce the damage caused by pathogens.
Additionally, crosstalk between the microbiota, epithelial cells, and immune system is key for the development and regulation of intestinal immune processes. (Rajput et al., 2022).- Therefore, L. intracellularis infection may result in microbial dysbiosis and decrease the immune response of the host to pathogens, thereby exposing the host to other enteric infections, such as Salmonella (Leite et al., 2018). We hypothesized that by stimulating the innate immune response (trained immunity) and through its ability to modify the gut microbiota of pigs, yeast-extracted BG could mitigate the negative impact of L. intracellularis on pig health and performance. In addition to demonstrating the efficacy of yeast BG in decreasing the negative impact of L. intracellularis on performance parameters and its positive effect in reducing faecal shedding of the pathogen in fattening pigs protective immunity induced by BG, we aimed to outline the specific changes in faecal microbiota composition related to yeast-derived BG supplementation during L. intracellularis infection.
2 Materials and methods
2.1 Ethical statement
The study was performed and documented according to appropriate quality standards. The experimental procedures used in this research were approved by the Animal Ethics Committee of RDN and were in compliance with the Directive 2010/63/EU of the European Parliament and Council, and with the Spanish Guidelines for the Care and Use of Animals in Research (Boletín Oficial del Estado, 2013) This trial followed all the recommendations of the Spanish Royal Decree 1135/2002 related to animal welfare in pig farms’.
2.2 Animals
A total of 160 healthy male and female pigs Danbred x Duroc (10 weeks of age) were sourced from a commercial farm (Explotación Porcina Vaqueril, Malpaca de Tajo, Spain). The study was then carried out at farm Miguel de la Pena Sanchez, San Bartolome de las Abiertas, Toledo, Spain. All the pigs entered the study at ten weeks old, at a weight ranging from 18 to 27 kg and ear-tagged on arrival to the experimental facility. They were then randomly assigned to one of the two treatment groups with 8 pens per treatment and 10 pigs per pen (n = 80). All the pens (3x3.2 m in size) were equipped with an individual feeder and a nipple drinker. The two experimental groups started with a similar average body weight (BW) of 21.5 kg and with equal number of gilts and barrows, providing 0.96 m2 per pig at 110 kg of BW.
2.3 Diets
Two dietary treatment groups were considered and consisted of 1) a control diet (with no BG supplementation), and 2) a BG supplemented diet at 250 ppm (Safglucan®, Phileo by Lesaffre) according to manufacturer recommendation. The diet used was a wheat/barley/soybean meal-based diet and nutrients levels were calculated to meet requirements of the animals (Table 1). The feeding program consisted of 4 feeding phases: adaptation (from arrival to 30 kg of BW), grower 1 (from 30 kg to 70 kg of BW), grower 2 (from 70 kg to 90 kg of BW), and finisher (from 90 kg of BW to slaughter) (Supplementary Figure 1). The diet formulation was provided by RDN and all the feeds were in pellet form. Diets were manufactured by Piensos COSAD (La Pueblanueva, Toledo, Spain). Feed and water were available ad libitum throughout the trial.
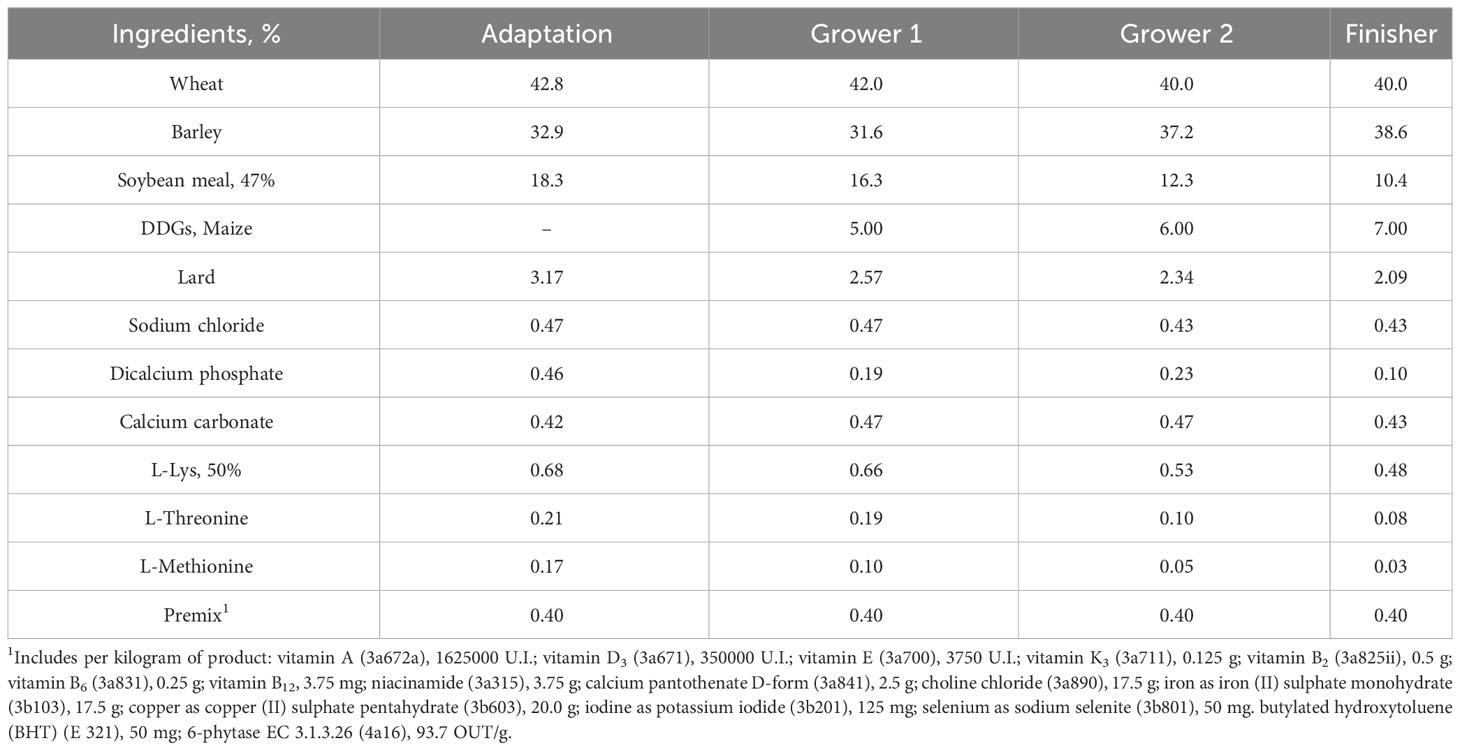
Table 1 Pigs diet composition.
2.4 Zootechnical data and faeces sample collection
Pigs were weighed individually, and feed disappearance was determined per pen at day 0, 63, 81 and 97 of the trial. From these data, average daily gain (ADG), average daily feed intake (ADFI) and feed conversion ratio (FCR) were calculated. Dead or culled pigs were weighed, and their estimated feed consumption was considered for FCR calculation (Supplementary Table 1). Faecal samples were also collected individually on the above indicated days via rectal massage from 5 pigs per pen. The same 5 pigs within each pen were sampled at each time point. Faecal samples were then stored at -20°C until analysis.
2.5 Lawsonia intracellularis detection by quantitative PCR
Bacterial DNA from each faeces sample was extracted using MagMaxCore (Thermofisher) according to instructions from the manufacturer (Qiagen, CA, USA). Extracted DNA was quantified for L. intracellularis by qPCR technique following manufacture instruction (EXOone Lawsonia intracellularis oneMIX, EXOPOL).Bacterial load (BL) classes were defined according to the L. intracellularis PCR quantification with a load > 200 copies/g of faeces for High BL, and< 200 copies/g of faeces for Low BL.
2.6 DNA extraction for microbiota analysis
Each sample previously stored at -20°C was placed 5 min at room temperature. The extraction was performed by the Nucleospin® 96 Soil kit (Macherey Nagel) under aseptic conditions at room temperature. First, 700 µL of lysis buffer (suitable for samples with organic carbon) was added to rehydrate the sample for 5 min. The extraction was performed by adding the Enhancer buffer in accordance with the supplier’s instructions. The tubes were then subjected to intense 30 Hz agitation for 2 min with bead mills (Retsch, Haan) to mechanically lyse the sample. The protocol followed the supplier’s recommendations until the elution.
2.7 Library preparation
The sequencing library is based on the principle of dual-indexing thanks to paired-end sequencing. Two PCRs were successively applied: from 2 µL of the extracted DNA diluted to 1/200, a first PCR in final volume of 50 µL, 1 U of Phanta Max Supplementaryer-Fidelity DNA Polymerase (Vazyme, each primer had a final concentration of 500 nM. For this first PCR, forward and reverse primers were designed with a 5’-Tag sequence respectively TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG for forward primer and GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG for reverse and a 16S RNAr gene specific sequence respectively CCTACGGGNGGCWGCAG for forward primer and GACTACHVGGGTATCTAATCC for reverse. According to E. coli 16S RNAr sequence gene specific primers targeted a locus between position 341 and 785, resulting in the amplification of a locus of 445 bp. The amplification conditions were 3 min at 95 °C, 25 cycles of 15 s at 94 °C for denaturation, 15 s at 51 °C for primers annealing and 45 s at 72°C for extension, followed by an incubation at 72°C for 1 min. At the end of this first PCR, amplification products were purified with NucleoFast® 96 PCR (Macherey Nagel) according to supplier recommendations except for the last step for which 30 µL of TE 1X preheated at 70°C were used for elution. From 5 µL of the previously purified DNA, a second PCR was performed in a final volume of 50 µL, 1 U of Phanta Max Supplementaryer-Fidelity DNA Polymerase (Vazyme). Each primer had a final concentration of 500 nM. The amplification conditions were the same as those of the previous one except the number of cycles reduced to 8. In addition to the Tag sequences, these PCR2 primers contained a locus to index the samples (barcode sequence) and a locus sequence adapter suitable for the Illumina sequencing technology. A NucleoFast® purification step identical to the one presented above was performed followed by a Quant-iT PicoGreen ds DNA quantification (Life Technologies). An equimolar pool of the library was produced.
2.8 Sequencing
Sequencing library was paired-end sequenced on MiSeq platform (Illumina) with MiSeq Reagent Kit v3 allowing 600 sequencing cycles to be performed. At the end of the sequencing a quality control by FastQC was carried out.
2.9 Primary analysis
Raw metagenomic reads were processed using QIIME2 (Bolyen et al., 2019) microbiome analysis package Data were first imported in Fastq format paired-end sequence. After a quality check, DADA2 (Callahan et al., 2016) was used as the main process of selecting sequence variants for amplicon analysis.
DADA2 used a statistical error correction model and builds ASVs (Amplicon sequence variants). In order to link each ASVs to the closest organism, SILVA database v138 (Quast et al., 2013) was used as the reference database for identifying the sequences.
De novo phylogenetic trees used in downstream measures of diversity was also built using default QIIME2 provided methods phylogeny align-to-tree-mafft-fastree (MAFFT multiple sequence alignment program) (Price et al., 2010; Katoh and Standley, 2013). The ASVs file and associated tree and classification files were then used for diversity analysis.
2.10 Statistical analysis
2.10.1 Zootechny, qPCR
A Mann-Whitney nonparametric test or a Kruskal-Wallis test (one-way Anova analysis) were used to compare significance between groups according to the data set. Graph and statistical analyses were performed using Prism version7 software (GraphPad, LaJolla, CA). A p<5.0x10-2 was considered statistically significant.
2.10.2 Microbiota analysis
Alpha-diversity was measured using Faith’s Phylogenetic Diversity (PD) (Faith, 1992) indices, using rarefied counts (i.e. subsampled to the same sequencing depth across samples). Comparing to others diversity indices such as Shannon or observed OTUs, Faith-PD takes into account the phylogenetic relationships between species in a community and tends to be less sensitive to undersampling or variations in sequencing depth. Alpha-diversity measures were done within QIIME2, with between group statistical differences determined using the one-way analysis of variance (ANOVA) tests. Betadiversity (across-sample) carried out using rarefied counts (as above) was measured using the Weighted Unifrac dissimilarity measures that takes into account phylogenetic distances between taxa. Differential Analysis of Compositions of Microbiomes with Bias Correction were done with ANCOM-BC (Lin and Peddada, 2020) for single time point and linDA (Zhou et al., 2022) for longitudinal test, which are both methodologies of differential abundance (DA) analysis for microbial absolute abundances. ANCOM-BC estimates the unknown sampling fractions, corrects the bias induced (Sex, Treatment or L. intracellularis qPCR status) by their differences through a log linear regression model including the estimated sampling fraction as an offset term, and identifies taxa that are differentially abundant according to the variable of interest. In ANCOM-BC, False Discovery Rate (FDR) correction was applied to adjust the p-values, ensuring a more stringent control of Type I errors.
In order to detect a microbial signature that are predictive of our phenotypes of interest, selbal method was used. selbal is a CoDA (compositional data analysis) methods for variable selection. Unlike approaches that define biomarker signatures as a linear combination of individual markers, the microbial signature from selbal has the form of a balance between two groups of microbial taxa.
The optimal balance between two groups relies on identifying taxa in either the first or second group of interest. The selbal algorithm first evaluates all possible taxon pairs to find the one most associated with the response. Then, it performs forward selection by adding new taxa to improve balance in either group, based on criteria like AUC, explained deviance, or mean squared error. The algorithm stops when no variable improves the criterion or when the maximum balance components are reached via cross-validation. selbal results help interpret microbial balances in microbiome studies. selbal is distributed as a R package. https://malucalle.github.io/Microbiome-Variable-Selection/selbal.html.
To explore interaction with zootechnical variables, FactomineR (Le et al., 2008) and Factoextra R (Kassambara & Mundt, 2021) packages were used specifically for Multiple Factor analysis (MFA). MFA is used to identify the relationships between multiple variables or factors in a dataset. It is similar to principal component analysis (PCA), but unlike PCA, which is used to identify the underlying structure of the data, MFA is used to identify the relationships between the variables or factors.
3 Results
3.1 Yeast BG improve the performances of the fattening pigs
Upon their arrival, 160 pigs were randomly assigned in 2 groups in a way that no significative difference in weight was observed between the two groups (Figure 1A). Then, one of the two groups was randomly assigned to the supplemented yeast BG diet over the growing and the fattening periods while the second group remained with a control diet. Pigs were individually weighted on days 63, 81 and 97. At D63 and D81, no significant difference (p=8.2x10-1 and 3.4x10-1, respectively) was observed between the groups (Figure 1B). However, at D97, the group fed with yeast BG showed a significant increase (p=4.5x10-20) in body weight which were 4.22 kg heavier than the pigs fed with the control diet (Figures 1B, C).
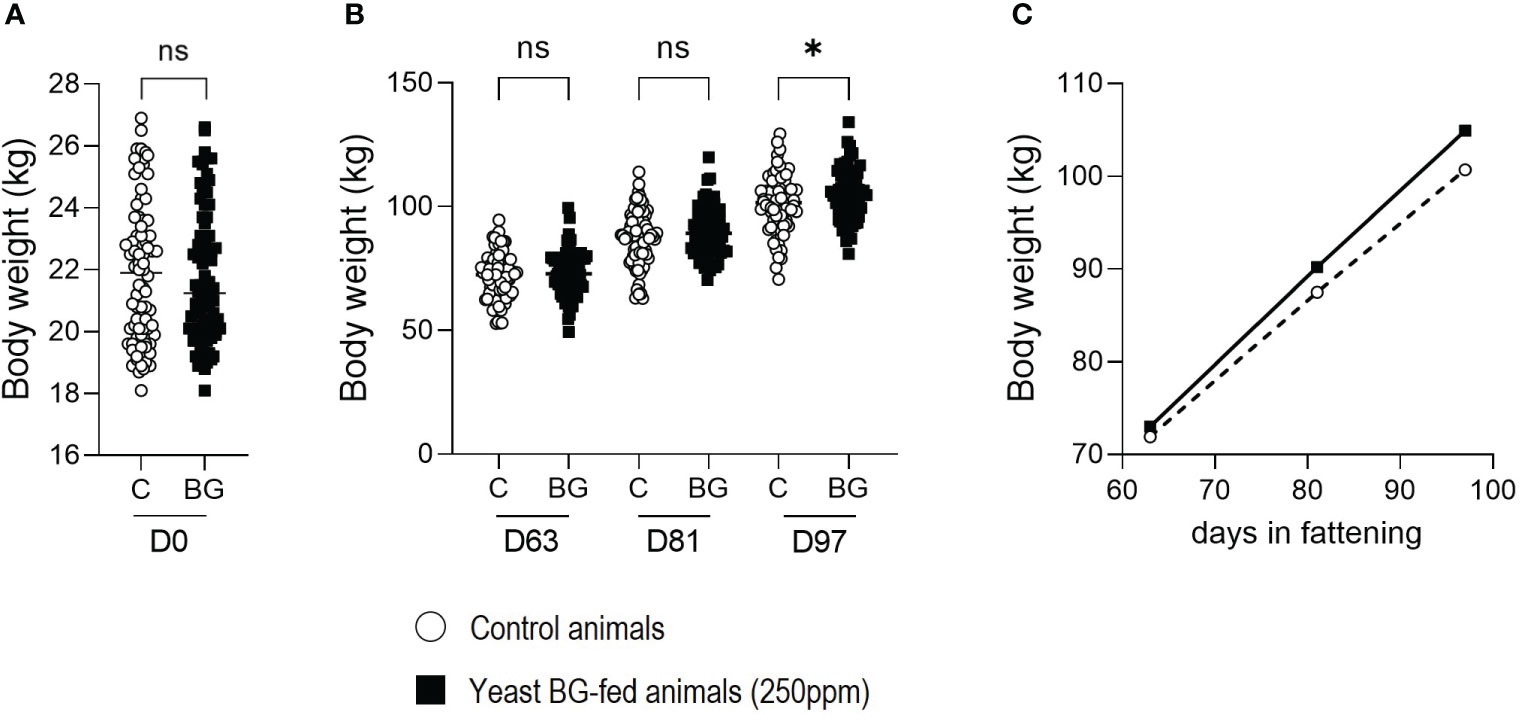
Figure 1 Supplementation in yeast BG increases the average body weight (BW) of fattening pigs. (A) BW in kg of the pigs on their arrival at the fattening facility day 0 (B) BW in kg of the pigs on day 63, 81 and 97, (C) BW of the pigs expressed over the time in fattening facility. N = 80 pigs per groups, white open round represents the control, black square represents yeast BG-fed animals. * p< 5.0x10-2, ns: not significative.
As the overall weight of the fattening pigs has increased in the yeast BG-fed group, the average daily gain (ADG) also increased significantly (p=4.6x10-2) in the yeast BG-fed group from D63 to D81 and from D81 to D97 (p=2.0x10-4) by 7.3% and 15.8%, respectively, compared to the control group (Figure 2A). The ADG of the animals during the period D63-D97, which is critical for the growth of the animals, is strongly and significatively enhanced (+10.6%, p=1.0x10-4) in the yeast BG-fed group compared to the control group (Figure 2B). Overall, from D0 to D97, yeast BG group showed a significantly (p=2.2x10-2) higher ADG with an increase of 5.6% as compared to the control group. (Figure 2C).
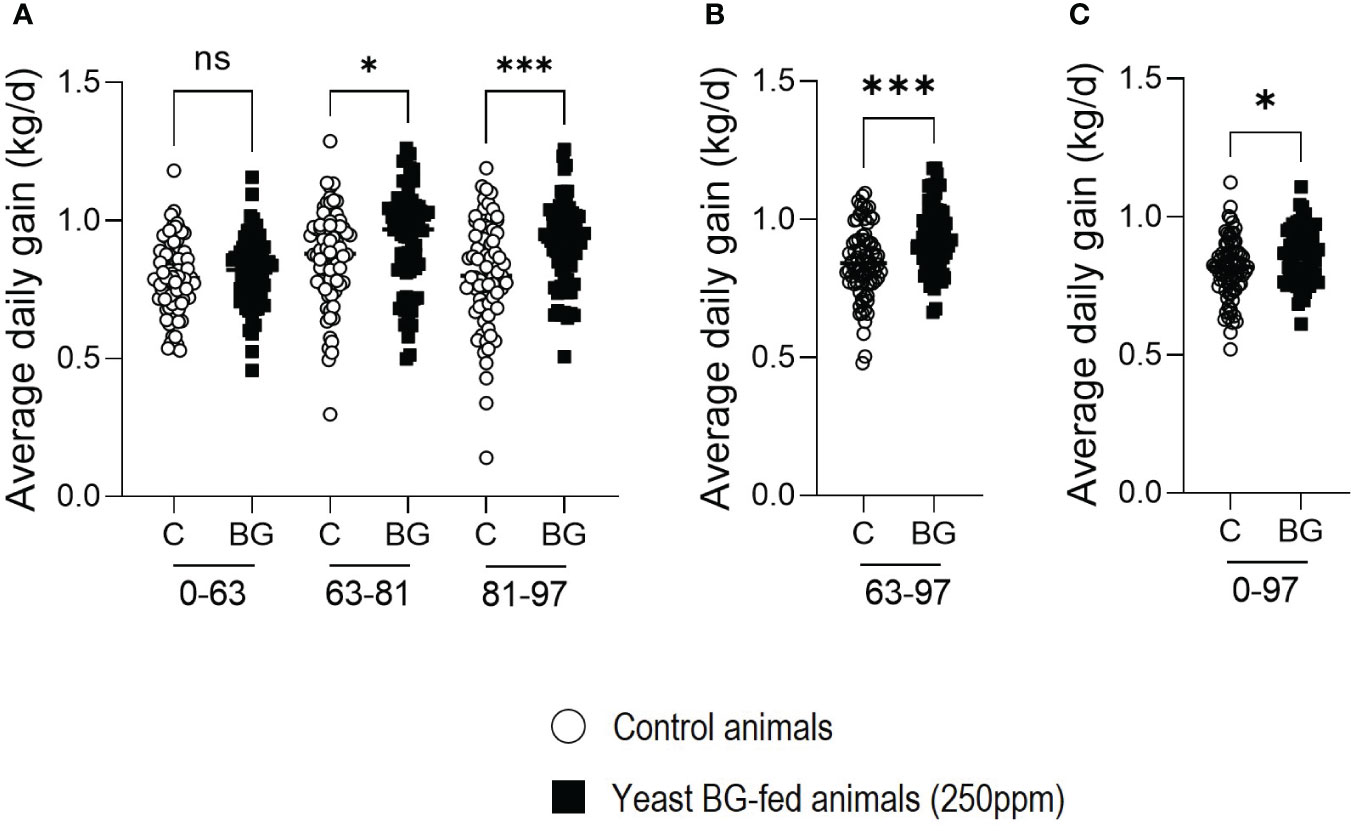
Figure 2 Average daily gain (ADG) of fattening pigs is increased by the supplementation in yeast BG. Average daily gain of the pigs over period of time (A) D0-63, D63-81 and D81-93, (B) period from D63 to D97, (C) the complete period of the fattening from D0 to D97. N=80 pigs per group, white open round represents the control, black square represents yeast BG-fed animals. * p< 5.0x10-2, *** p< 1.0x10-3; ns, not significative.
Similarly, the average daily feed intake (ADFI) significantly increased in yeast BG-fed group by +8,9% (p=2.4x10-2) and +9% (p=2.0x10-2) in the last two periods of time (D63-D81 and D81-D97, respectively) compared to the control group (Figure 3A). However, over the whole period of the trial (D0 to D97), no significant difference (p=1,7x10-1) was observed between the two groups (Figure 3B).
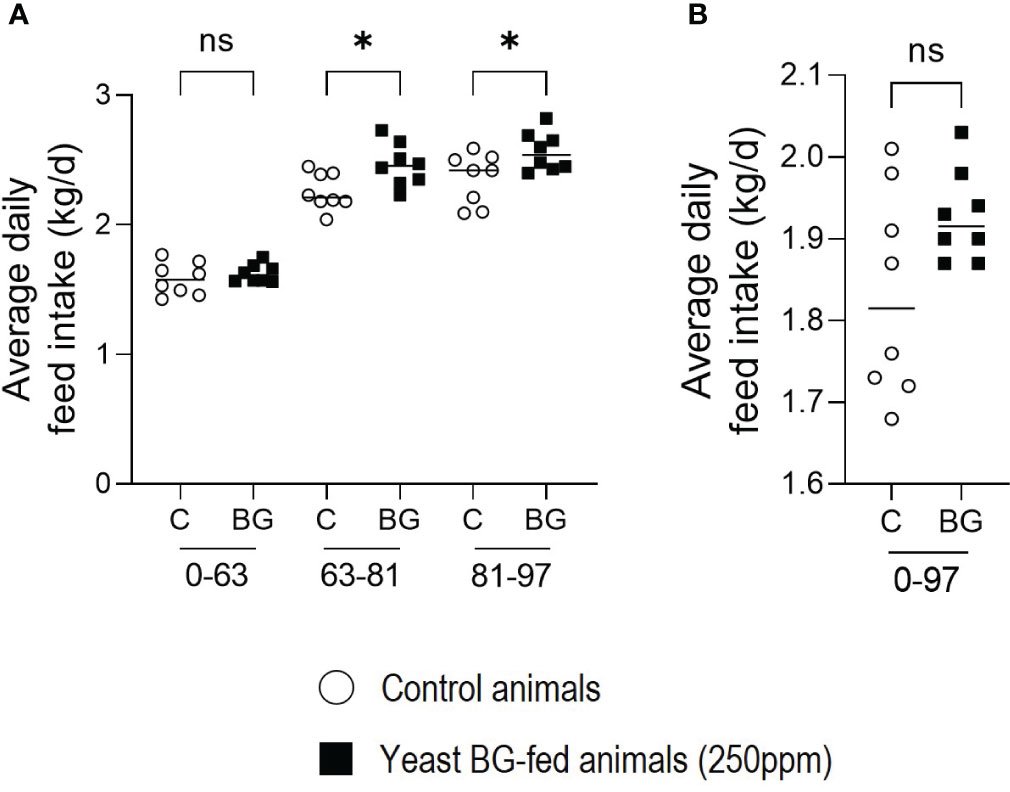
Figure 3 Average daily feed intake (ADFI) is increased by the supplementation in yeast BG in fattening pigs. (A) Average daily feed intake of the pigs over period of time (A) D0-63, D63-81 and D81-93, (B) period from D0 to D97, n=8 pens per group (of 10 animals each), white open round represents the control, black square represents yeast BG-supplemented animals. * p< 5.0x10-2; ns, not significative.
Finally, the feed conversion ratio (FCR) was evaluated per pen. No significant difference was observed for the FCR between the two groups on the overall period, despite a reduction of 2.7%, from D81 to D97 in the BG-fed group (Figures 4A, B). All the zootechnical data have been summarized in Supplementary Table 1.
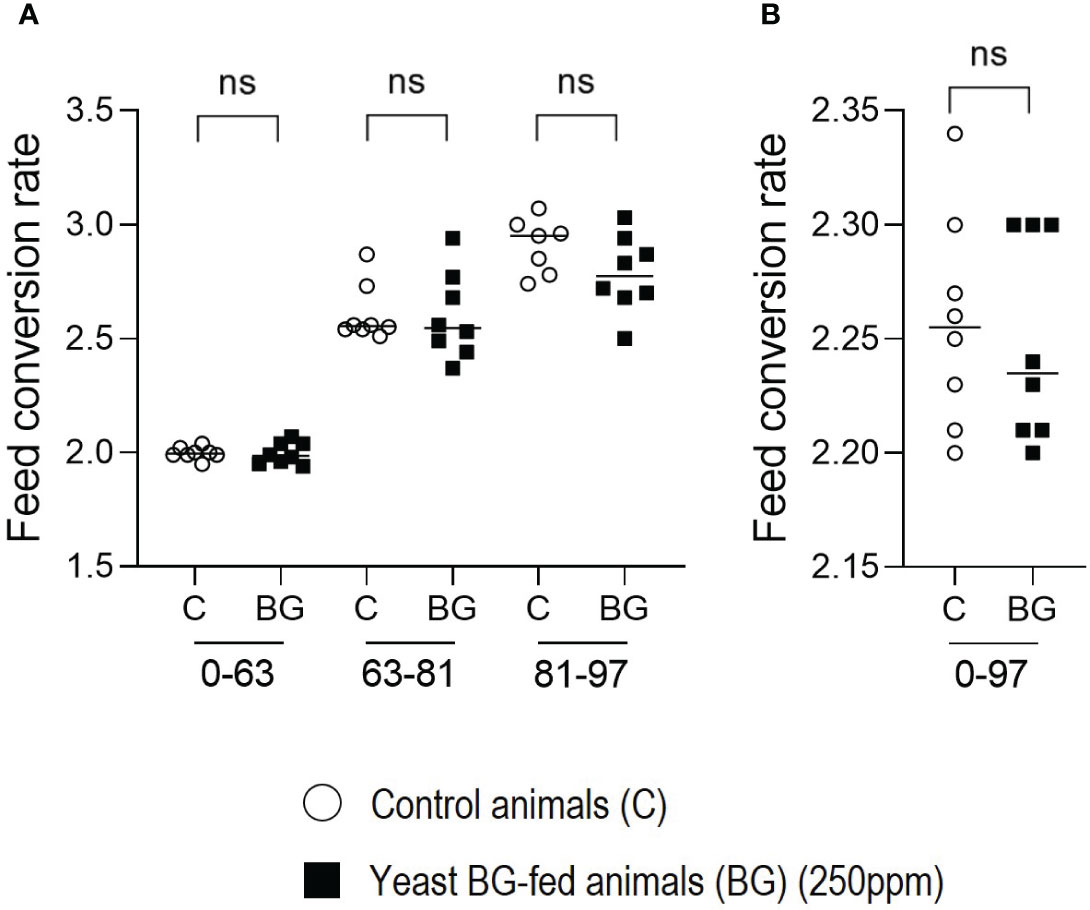
Figure 4 Feed conversion rate (FCR) is not modulated by the supplementation in yeast BG in fattening pigs. (A) Feed conversion rate of the pigs over period of time (A) D0-63, D63-81 and D81-93, (B) period from D0 to D97, n=8 pens per group (of 80 animals each), white open round represents the control, black square represents yeast BG-fed animals. ns, not significative.
3.2 Faecal shedding of Lawsonia intracellularis is reduced in the fattening pigs fed with yeast BG
Quantitative PCR were performed on the faeces of 40 animals per group at D63, D81 and D97 (Figures 5A–C) to evaluate the presence of L. intracellularis. At D63 and D81, no significant difference was observed between the groups even though there was a numerical decrease in the BG group (p=2.3x10-1 and p=1.4x10-1 respectively). However, the load of L. intracellularis in the faeces of BG-fed animals decreased significantly compared to the control group (3.7 versus 5.1 log10 copy/g of faeces (p=4.0x10-4) at D97. This agrees with the number of animals detected positive for L. intracellularis (Figure 5D) at D81 with 82% in control group compared to 70.2% in the BG group and at D97 with 65.8% of positive animals in the control group compared to 30.8% in the BG-supplemented group, resulting close to 50% reduction in the number of positive animals in the BG group compared to the control group.
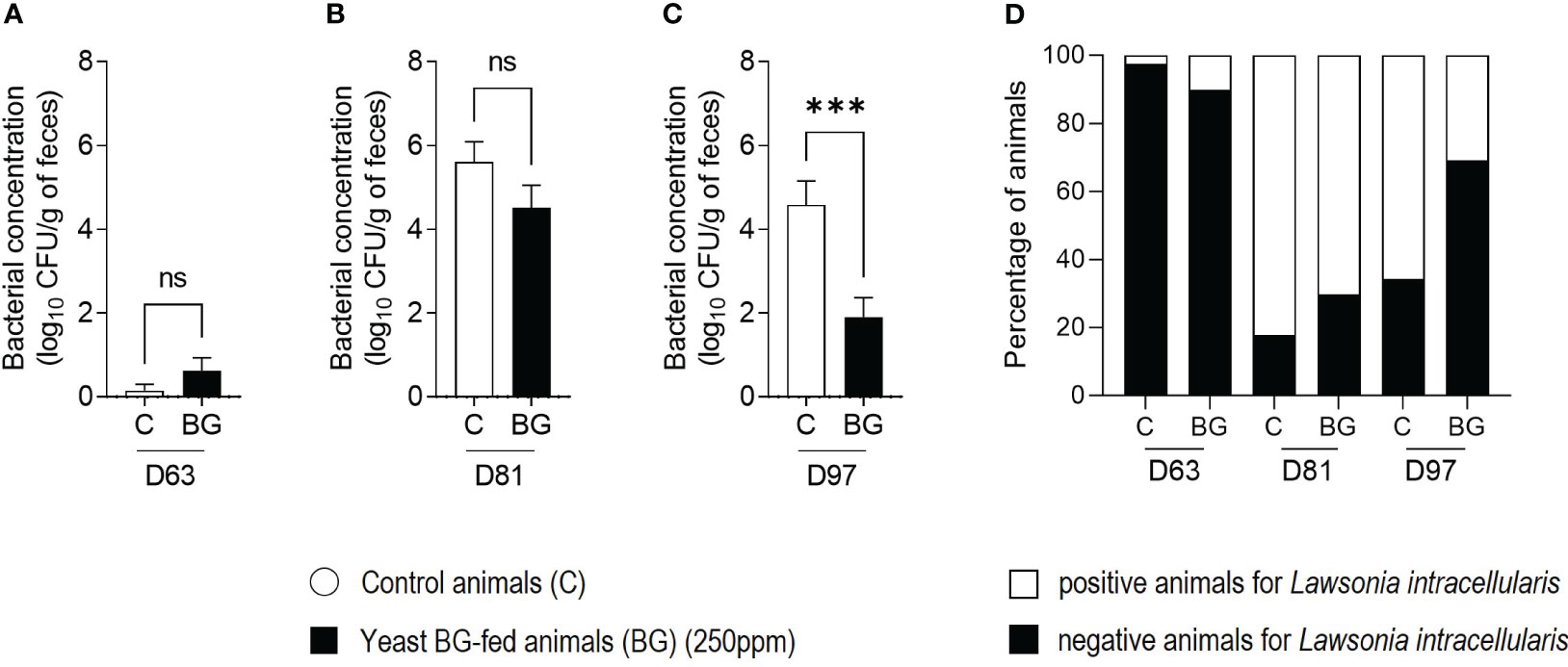
Figure 5 The shedding of Lawsonia intracellularis is reduced in the group of yeast BG-fed pigs. Quantitative PCR targeting L. intracellularis in the faeces of 37 to 40 animals per group at D63 (A), at D81 (B) and D97 (C). White bars represent the control, black bars represent yeast BG-fed animals. *** p< 1.0x10-3; ns, not significative. (D) Percentage of animals negative or positive for L. intracellularis at each time point. Bottom black bars represent the negative animals, top white bars represent the positive animals. N=37-40 pigs per group.
We wanted to assess here the impact of L. intracellularis infection on the faecal microbiome of the pigs. In the last decade, growing evidence demonstrate the influence of environmental conditions including infections on the composition of the microbiota and thus, as a mediator of pig health, on zootechnical parameters. BW or ADG can also be influenced by the ingestion of BG. In a context of L. intracellularis infection, it seems relevant to elucidate the modalities of BG actions and to consider a possible link to the modification of the faecal microbiota.
3.3 Association between Lawsonia intracellularis and the faecal microbiota diversities
3.3.1 Moderate negative association between α-diversity and Lawsonia intracellularis at D81
It appears that no significant differences were observed between L. intracellularis positive and negative animals at any of the time points concerning the α-diversity of the studied microbiota. However, a relationship seems to be drawn between α-diversity and L. intracellularis burden as a significant difference (p=2.1x10-2) between the two has been observed at day 81 (Figure 6). Indeed, the more the burden increases the more the α-diversity decreases. No significant difference was observed between negative samples and samples with a low L. intracellularis load (LOW BL), certainly due to the lower number of samples in this negative group. However, α-diversity from samples with a higher L. intracellularis load (HIGH BL was significantly reduced compared to the previous ones (p=8.1x10-3), suggesting that the presence of L. intracellularis can negatively impact microbiota α-diversity according to its load.
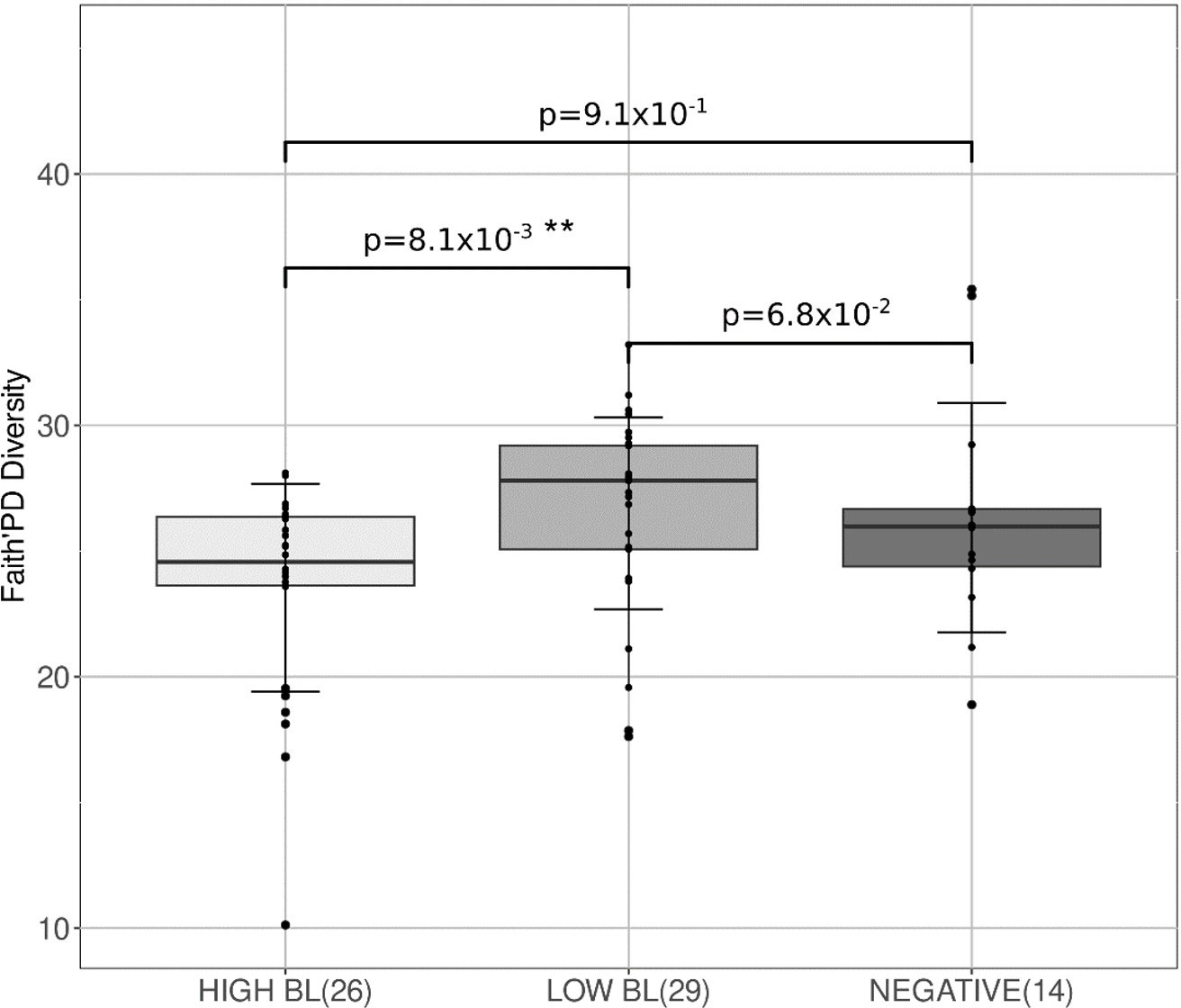
Figure 6 Box-plots associated with α-diversity at day 81 according to L. intracellularis burden. Bacterial loads (BL) classes were defined according to the L. intracellularis PCR quantification with a load > 200 copies/g of faeces for High BL, and< 200 copies/g of faeces for Low BL. 3 groups Anova test, p-value= 2.1x10-2. ** p< 1.0x10-2.
3.3.2 Lawsonia intracellularis impacts microbiota at taxonomic level
No significant relationship was observed until the last day of the study. A significant difference in terms of microbiota composition between L. intracellularis-positive (n=34) and negative (n=37) individuals was observed (Supplementary Table 2). The genera Sarcina and Catenibacterium were found in higher proportions (0.227% for positives and 0.016% for negatives for Sarcina; 0.251% for positives and 0.049% for negatives for Catenibacterium) in animals carrying L. intracellularis (p-value.fdr =2.5x10-2, p-value.fdr =2.8x10-2 respectively). Interestingly, a slight relative proportion of L. intracellularis was observed at the genus level among the taxa constituting the microbiota patterns of the animals in the control group (n=24, 0.12%), whereas this taxon was not detected (n=10, 0%) in the BG-supplemented animals (p =4.0x10-2).
3.3.3 Potential biomarker to predict Lawsonia intracellularis infection
As displayed in Figure 7, a simple ratio of two taxa might identify animals whose microbiota would be conducive to a later infection or would be an early indicator of a L. intracellularis infection not yet detectable by qPCR. Our dataset analysis led to the ratio constituted by Fibrobacter (as numerator), and Lachnospiraceae_AC2044 (as denominator), the increase of which was linked to animals infected by L. intracellularis in later times, with a good discrimination power (AUC>0.84). Thus, a higher proportion of Fibrobacter would be preferentially associated with a faecal microbiota that would be more susceptible to L intracellularis, as opposed to a higher proportion of Lachnospiraceae_AC2044_group, which would be associated with a greater resistance to future infection.
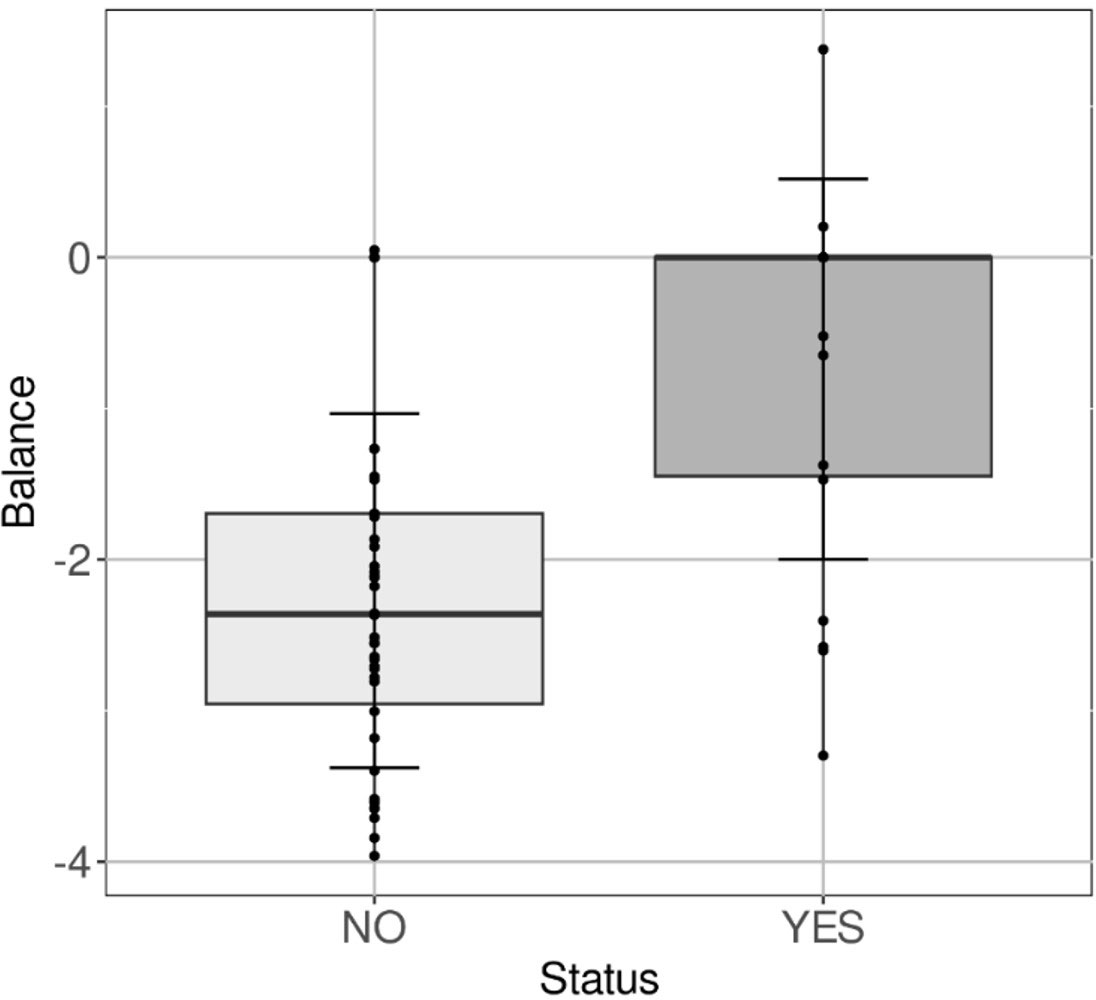
Figure 7 Ratio predicting Lawsonia contamination. Boxplot of the ratio FibrobacterLachnospiraceae AC2044 group FibrobacterLachnospiraceae AC2044 group according to negative animals in qPCR at day 63, which will become positive (YES) thereafter compared to animals that will remain negative (NO).
3.4 Microbiota is influenced by the yeast BG diet in fattening pigs
Comparing the controls (n=36) to the BG-fed animals (n=38), a higher α-diversity (Shannon index 5.4 for BG-fed vs 5.03 for controls, p=3.0x10-2) was observed only at day 63.
However, a longitudinal analysis at days 63, 81 and 97 revealed significant differences in microbiota bacterial composition. At the genus level, Sarcina, Lachnospiraceae UCG-007, Butyricicoccus, Subdoligranulum, Catenibacterium, Roseburia, Faecalibacterium, [Ruminococcus] gauvreauiigroup, Dorea, Intestinibacter, [Eubacterium] ruminantium group, [Eubacterium] eligens group, Colidextribacter, Oribacterium, Streptococcus, Olsenella, Solobacterium, Holdemanella, Prevotella were more abundant in the control group, while in the BG-fed group Prevotellaceae UCG-004 UCG-008 and Megasphaera were in higher proportion (Figure 8, Supplementary Table 3).
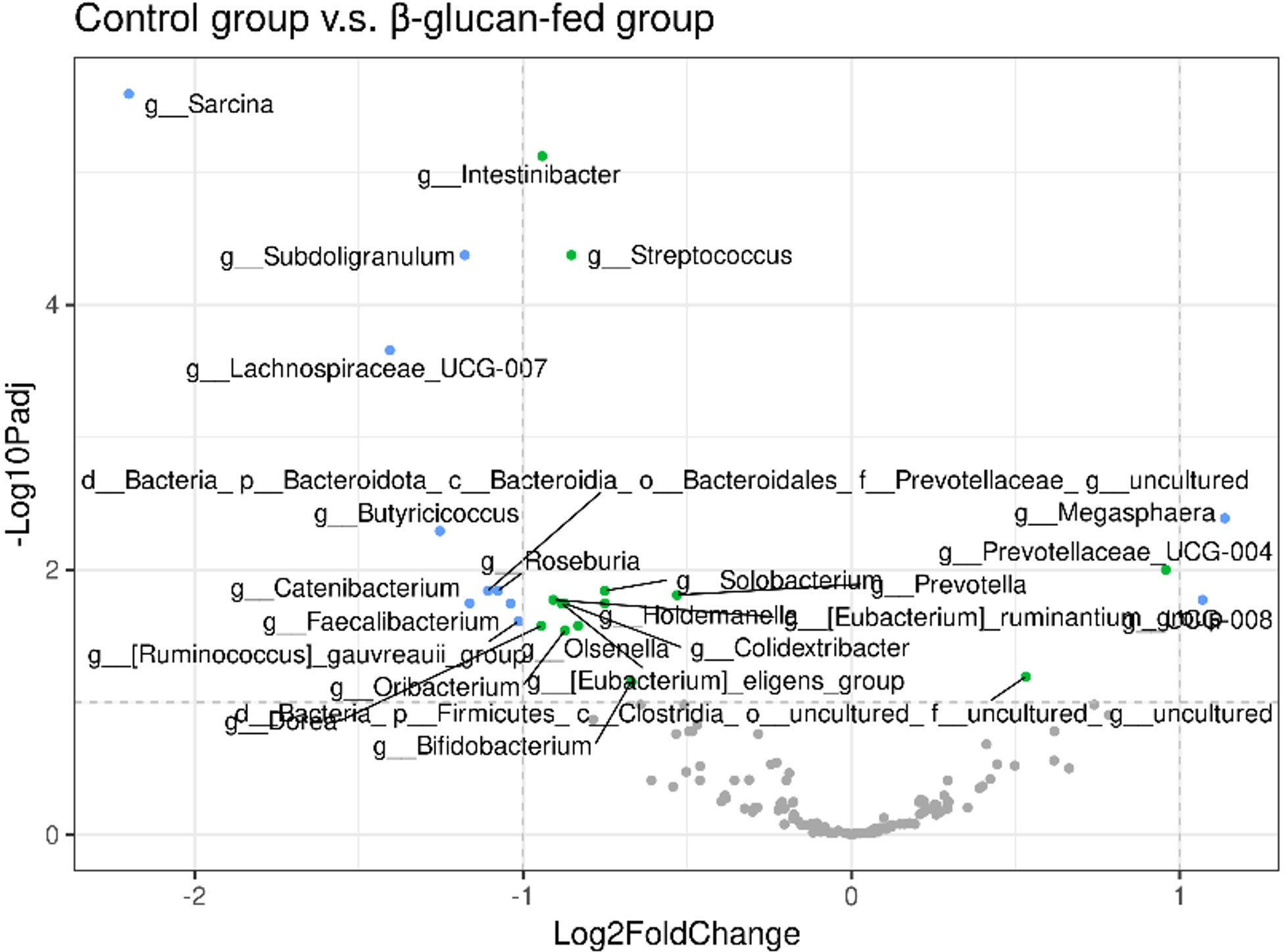
Figure 8 Volcano plot of longitudinal follow-up from day 63 to day 97, corrected for the presence of L. intracellularis. Plots are displayed in grey for adjusted p-value (P-adj)> 1.0x10-1 and log fold change (LFC), in green for P-adj ≤ 1.0x10-1 and LFC ≤ 1, in blue for P-adj ≤ 1.0x10-1 and LFC > 1.
Looking in more details at the taxonomic level at the last day of the study, when the two experimental groups were more contrasted, significant differences have been observed (Table 2). The bacterial taxa impacted by BG are mainly taxa whose contribution to the microbiota fingerprint is rather low. Compared with Figure 8, showing results following longitudinal follow-up, the genera Sarcina and Lachnospiraceae_UCG-007 remain particularly significant at this latest stage of the study with P-value.fdr respectively of 1.3x10-18 and 1.4x10-9.
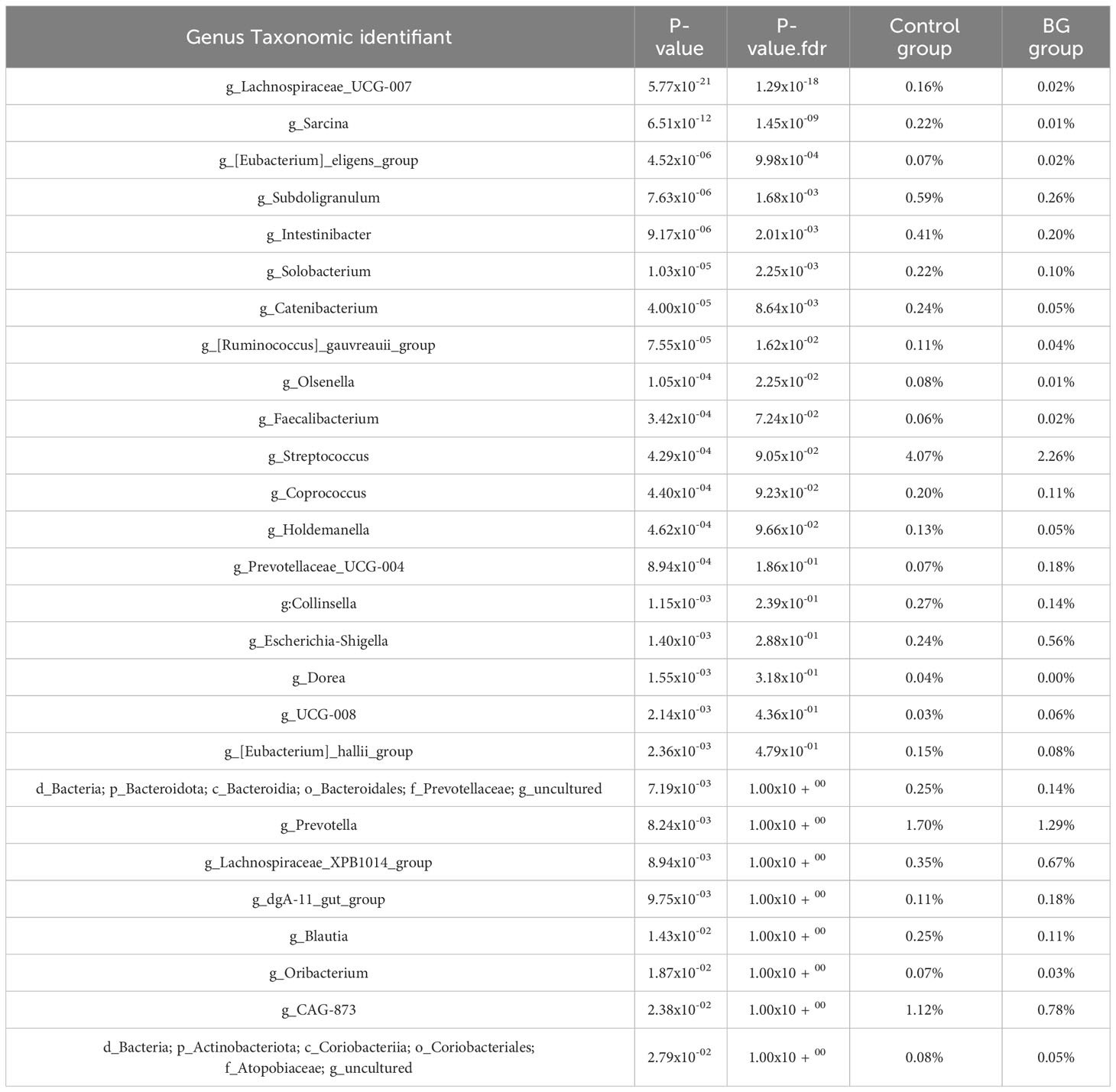
Table 2 Genera relative abundance significantly associated with BG-fed status at day 97.
Another ratio has been defined at the genus level, allowing to discriminate the BG-fed animals from those belonging to the control group with an accuracy > 0.8 (Figure 9):
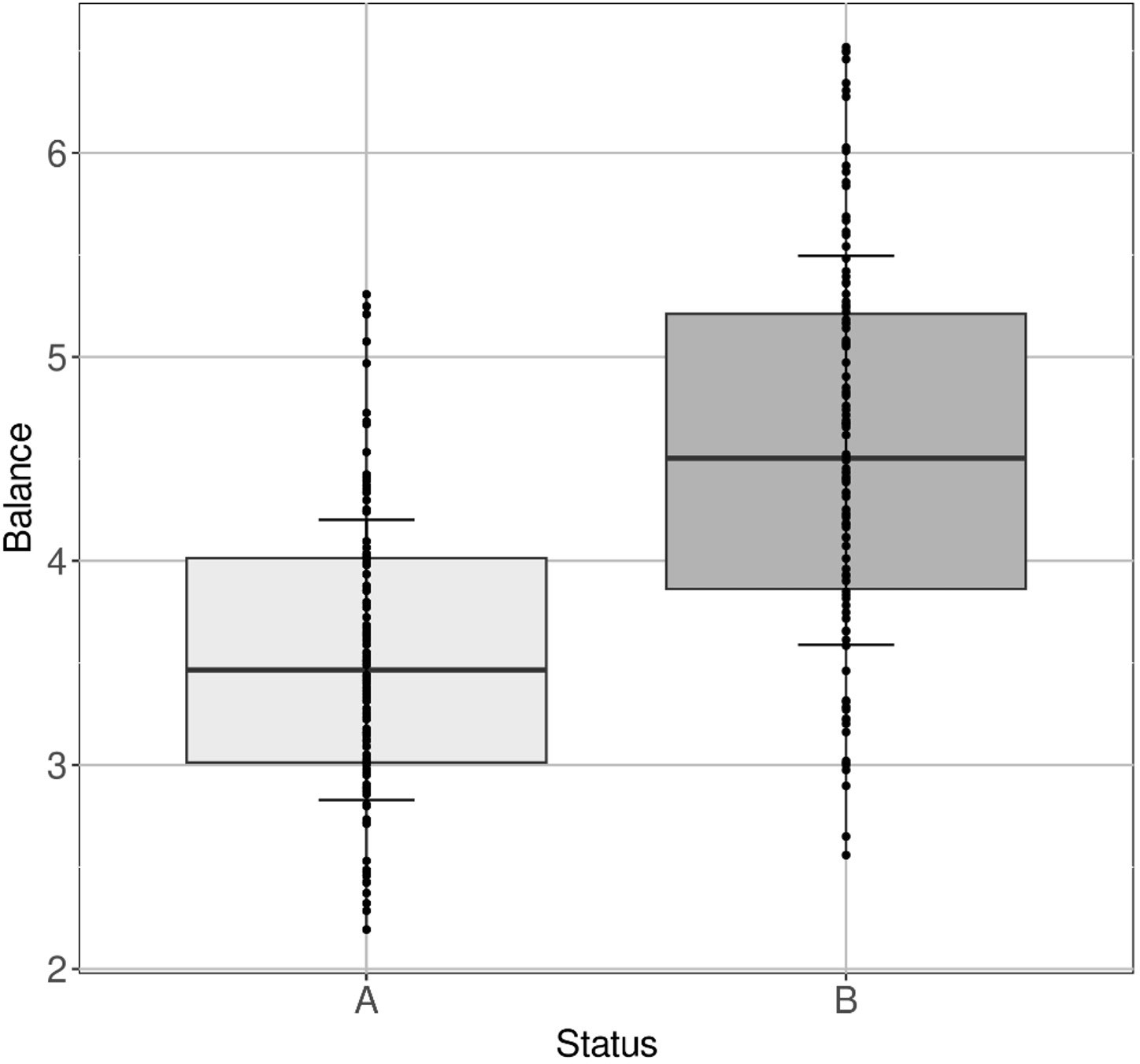
Figure 9 Ratio Clostridia UCG−014 + TerrisporobacterSarcina + Lachnospiraceae UCG−007 + Subdoligranulum + StreptococcusClostridia UCG−014 + TerrisporobacterSarcina + Lachnospiraceae UCG−007 + Subdoligranulum + Streptococcus associated with BG intake. Boxplot of the ratio for (A) control group and for (B) BG-fed group accumulating all time points except day 0.
Not surprisingly, in the denominator of the ratio constituting this biomarker, the genera Sarcina, Lachnospiraceae UCG-007, Subdoligranulum, Streptococcus were found.
3.5 Microbiota pattern associated with zootechnical performances
Figure 10 shows the results of the multifactorial analysis on the first two principal components, considering the following quantitative variables: BW at D97, L. intracellularis qPCR Ct values and normalised taxons constituting the biomarker previously described. The status of the fed group was also taken into account as qualitative variable.
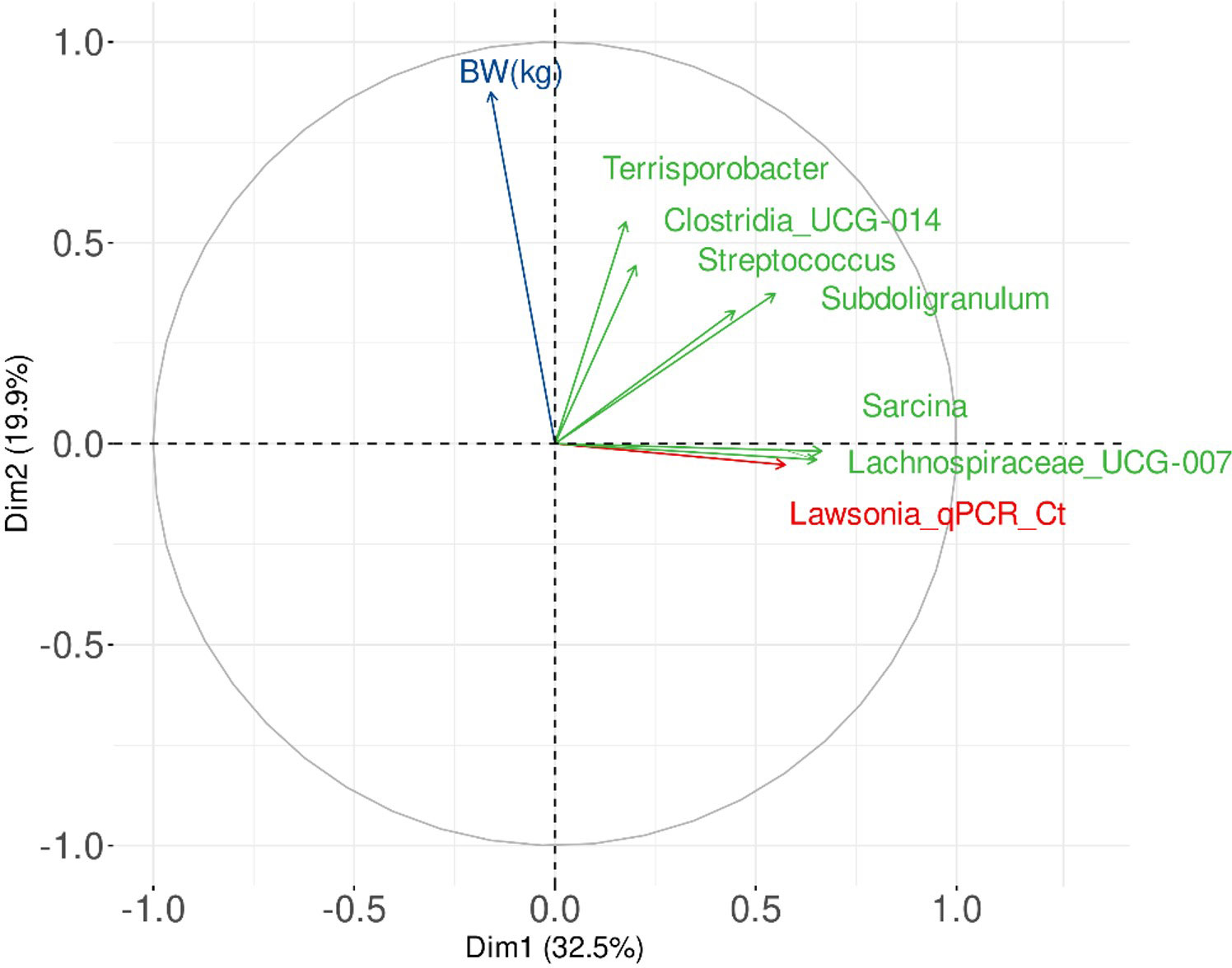
Figure 10 Graphical representation of a Multiple Factor Analysis (MFA) linking variables associated with the faecal microbiota fingerprints (themselves related to the experimental groups), BW and L. intracellularis burden.
These results suggest that it exists a positive trend between BW at D97, Terrisporobacter, and Clostridia UCG-014. These two taxons were also linked to the group of BG-fed animals. Among the four taxons linked to the control group, there was no clear link between BW and Subdoligranulum and Streptococcus, but it seems to exist a slight negative trend between BW and Lachnospiraceae UCG-007 and Sarcina, in parallel of a clear positive trend between L. intracellularis load and of Lachnospiraceae UCG-007 and Sarcina.
4 Discussion
In this study, we reported that yeast-extracted BG ameliorates the negative impact of L. intracellularis infection on the health and performance of grow-finishing pigs. Previous studies in different animal species have shown that BG were able to enhance growth and immunity in Nile Tilapia (Selim and Reda, 2014) and pigs (Vetvicka and Oliveira, 2014), as well as to decrease lesions in Eimeria-challenged broiler (Cox et al., 2010). In agreement with all these previous findings, our current study has also shown significant improvement of zootechnical performances of pigs under L. intracellularis pressure when supplemented with BG. Overall, this study showed that the administration of yeast BG resulted in an increase of BW and ADG, compared to the animals from the control group.
The performances improvement can be explained by the decrease of L.intracellularis shedding, itself explained by the known effect of BG on inducing the innate immune memory. Indeed, we observed a lower proportion of positive animals for L.intracellularis among those having received BG, at D81 as well as at D97, compared to the control group. In addition to this, our results have also shown a significant decrease in bacterial L.intracellularis shedding in the faeces of BG-supplemented pigs as compared to control. Those results are in line with a recent finding that reported a limited translocation of Salmonella Typhimurium to lymph nodes and decrease bacterial shedding in Salmonella challenged nursery piglets fed with BG diet (Bearson et al., 2023). As L. intracellularis infects macrophages, one of the most represented innate immune cells of the intestinal tract (Pereira et al., 2020), BG would reinforce this macrophage line of defence helping the animals to eliminate more efficiently the pathogen. These could be explained by the impact of BG in stimulating innate immune cells (through immune training) or their role in modifying the gut microbiota of piglets towards beneficial bacteria. To support this last hypothesis, we are discussing here below the positive impact of yeast BG in modifying the gut microbiota of pigs in the context of L. intracellularis infection.
Gut microbiota α-diversity decreases with increasing L. intracellularis burden. This observation is supported by the one made by Muwonge and collaborators who observed a difference in the α-diversity of the luminal ileum microbiota (Muwonge et al., 2021) according to the presence of L. intracellularis, and an increased α-diversity associated with reduced histopathology lesions and faecal shedding. In our study, the reduction in α-diversity was mainly observed for animals with the highest L. intracellularis burden (i.e. low copy number per mg of faeces). In addition, at D97, according to the infectious status of the animals, a slight modulation of the faecal microbiota has been observed. Sarcina and Catenibacterium were found in higher proportions in animals carrying L. intracellularis. Sarcina species are fastidious Gram-positive anaerobic bacteria that occur in cubic packs of eight or more cells. In 2006 and 2007, they were associated with cases of acute abomasal bloat in young lambs and calves (Edwards et al., 2008), whereas Catenibacterium, a genus of Gram-positive and obligate anaerobe bacteria, are mainly involved in the biosynthesis and metabolism of fatty acids (Choudhury et al., 2021; Loo et al., 2022).
Although, L.intracellularis has little impact on the faecal microbiota, some taxa can modulate the outcome of the infection. The biomarker defined by the ratio Fibrobacter/Lachnospiraceae AC2044 group at a time when L. intracellularis infection is not yet detectable by qPCR, could be a predictor of future infection and/or a marker of susceptibility to the infection. Indeed, before the infection occurs, this ratio was higher than in the ‘future’ positive animals’ group. Interestingly, a previous study by Leite (Leite et al., 2018) noted that a challenge with L. intracellularis decreased the relative proportion of Fibrobacter. On the contrary, Lachnospiraceae AC2044 group could have a preventive character in agreement with the positive correlation found between Lachnospiraceae AC2044 group and the contents of SCFAs, fat, and crude ash, as well as the negative correlation with amino acids in different gut locations of pigs (Song et al., 2022). Our results seem to be in line with these study as when the infection is declared, this ratio decreased in the positive animals throughout the trial until reaching the same level as that of the negative animals (Supplementary Data…).
BG used in this study also influenced the relative proportion of several taxa. The genus Prevotellaceae UCG-004 and Megasphaera, which are positively modulated in animals receiving BG, are known as polysaccharides degraders and SCFAs producers (Heinritz et al., 2016). In contrast, animals in the control group exhibited a much higher relative abundance in Sarcina, Lachnospiraceae UCG-007, Subdoligranulum, three taxa belonging to the Clostridia class, and to a lesser degree Streptococcus and Intestinibacter.
Here, the data led to the identification of a potential biomarker allowing to discriminate the microbiota pattern of BG-fed animals from the one associated with the control group. Although this potential gut microbiota pattern displayed the sum of Sarcina, Lachnospiraceae UCG-007, Subdoligranulum and Streptococcus as the denominator, as expected, these are Terrisporobacter and Clostridia UCG-014 that appeared in the numerator. The positive correlation between their relative abundance and the BW highlighted at the end of the trial is in accordance with previous studies. Interestingly, these two genera are considered to be plant fibre degrading bacteria (Li et al., 2021). In addition, other studies showed that relative abundance of Terrisporobacter is also positively correlated to feed efficiency in young pigs as well as amino acids metabolism, including tryptophan, a metabolite of which (indoleacetic acid) that can positively modulate intestinal homeostasis and inflammatory response (McCormack et al., 2019; Yan et al., 2021; Shen et al., 2022).
Even though it could be tricky to distinguish between the consequence from the cause in correlation studies between phenotype and microbiota, we can nevertheless make a hypothesis considering the nature of the additive. β-1,3/1,6-glucans are essential components of a prebiotic-rich diet that promotes growth and metabolism of the gastrointestinal microbiota (Metzler-Zebeli et al., 2011). Indeed, in the hypothesis of a prebiotic-type treatment favouring an increase in the relative abundance of certain taxa at the expense of others, BG used in this study would favour the taxa Terrisporobacter and Clostridia UCG-014, which are themselves associated with a higher weight gain, decreasing consequently Sarcina, Lachnospiraceae UCG-007, Streptococcus and Subdoligranulum relative abundance. In our study, Sarcina and Lachnospiraceae UCG-007 were relatively independent of body weight, although Sarcina has been associated with poorer performance in terms of weight gain in previous studies as well as Lachnospiraceae (Levesque et al., 2014; Bergamaschi et al., 2020; Quan et al., 2020; Holanda and Kim, 2022). But, interestingly, these two taxa were strongly associated with the level of L. intracellularis contamination. Two other genera, Streptococcus and Subdoligranulum, do not appear to have a clear association with the BW or burden of L. intracellularis.
5 Conclusion
In conclusion, some specific yeast branched BG were able to improve health, by reducing bacteria shedding and by increasing body weight in pigs. Regarding biomarker and differential analysis, this study showed that taxa, such as Clostridia UCG-014 and Terrisporobacter, are preferentially related to animals fed with BG and with an improved BW whereas the taxa Subdoligranulum, Streptococcus, Sarcina and Lachnospiraceae UCG-007 are more present in animals in the control group and are partly related to the L. intracellularis load (Figure 11). This dual analytical approach led to the establishment of a ratio with these taxa, that should be validated in future studies, allowing to distinguish the faecal microbiota of an animals fed or not with BG. Those findings open up also the possibility of having microbial biomarkers to evaluate high performing and L. intracellularis-resistant animals.

Figure 11 Schematic representation of the effects related to the intake of BG in presence of L. intracellularis. The figure shows on the left the control group and on the right, the yeast BG group. The bacteria that are the most representative for each condition are written on top of the figure. * means p ≤5.0x10-2; ** means p≤ 1.0x10-2..
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Files, further inquiries can be directed to the corresponding author/s.
Ethics statement
The animal study was approved by spanish Royall Decree 1135/2002. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
GE, CA performed the microbiota analysis and analysed the data. LR coordinated the microbiota analysis. GE, CA, SL-R, LR, TK, JS wrote the manuscript. JS designed the study, organised and conducted the study, analysed the performance data. GK found the CRO and dealed with the administrative work. All authors contributed and approved the submitted version.
Conflict of interest
LR, TK, GK and JS are employees of the funding agency Phileo by Lesaffre, manufacturer of the BG mentioned in this study.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2023.1245061/full#supplementary-material
References
Bain C. C., Schridde A. (2018). Origin, differentiation, and function of intestinal macrophages. Front. Immunol. 9. doi: 10.3389/fimmu.2018.02733
Bearson S. M. D., Trachsel J. M., Bearson B. L., Loving C. L., Kerr B. J., Shippy D. C., et al. (2023). Effects of β-glucan on Salmonella enterica serovar Typhimurium swine colonization and microbiota alterations. Porcine Health Manag 9, 7. doi: 10.1186/s40813-023-00302-4
Bergamaschi M., Tiezzi F., Howard J., Huang Y. J., Gray K. A., Schillebeeckx C., et al. (2020). Gut microbiome composition differences among breeds impact feed efficiency in swine. Microbiome. 8, 110. doi: 10.1186/s40168-020-00888-9
Boletın Oficial del Estado. (2013). Royal Decree 53/2013, of February 1, which establishes the basic rules applicable for the protection of animals used in experimentation and other scientific purposes, including teaching. p. 11370-11421. Available at: https://www.boe.es/eli/es/rd/2013/02/01/53.
Bolyen E., Rideout J. R., Dillon M. R., Bokulich N. A., Abnet C. C., Al-Ghalith G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Byrne A. K., Tuggle C. K., Loving C. L. (2020). Differential induction of innate memory on porcine monocytes by β-glucans or bacillus Calmette-Guerin. Innate Immunity. 27, 6. doi: 10.1177/1753425920951607
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 3, 581–583. doi: 10.1038/nmeth.3869
Choudhury R., Middelkoop A., Boekhorst J., Gerrits W. J. J., Kemp B., Bolhuis J. E., et al. (2021). Early life feeding accelerates gut microbiome maturation and suppresses acute post-weaning stress in piglets. Environ. Microbiol. 23, 7201–7213. doi: 10.1111/1462-2920.15791
Cox C. M., Sumners L. H., Kim S., McElroy A. P., Bedford M. R., Dalloul R. A. (2010). Immune responses to dietary beta-glucan in broiler chicks during an Eimeria challenge. Poult Sci. 89, 2597–2607. doi: 10.3382/ps.2010-00987
Edwards G. T., Woodger N. G. A., Barlow A. M., Bell S. J., Harwood D. G., Otter A., et al. (2008). Sarcina-like bacteria associated with bloat in young lambs and calves. Vet. Rec 63, 391–393. doi: 10.1136/vr.163.13.391
Faith D. P. (1992). Conservation evaluation and phylogenetic diversity. Biol. Conserv. 61, 1–10. doi: 10.1016/0006-3207(92)91201-3
Fourchon A., Chouet S. (2000). Technical results of swine herds and serological results on pigs for Lawsonia intracellularis. Proc. IPVS 16, 62. doi: 10.1111/j.1751-0813.2010.00558.x
Heinritz S. N., Weiss E., Eklund M., Aumiller T., Louis S., Rings A., et al. (2016). Intestinal microbiota and microbial metabolites are changed in a pig model fed a high-fat/low-fiber or a low-fat/high-fiber diet. PloS One 11. doi: 10.1371/journal.pone.0154329
Holanda D. M., Kim S. W. (2022). Impacts of weaning weights and mycotoxin challenges on jejunal mucosa-associated microbiota, intestinal and systemic health, and growth performance of nursery pigs. J. Anim. Sci. Biotechnol. 13, 43. doi: 10.1186/s40104-022-00691-6
Holtkamp D. (2019). Economic losses associated with ileitis; MSD animal health: kenilworth, NJ, USA. PloS One.
Jacobson M., Hård af Segerstad C., Gunnarsson A., Fellström C., de Verdier Klingenberg K., Wallgren P., et al. (2003). Diarrhoea in the growing pig – a comparison of clinical, morphological and microbial findings between animals from good and poor performance herds. Res. Vet. Sci. 74, 163–169. doi: 10.1016/S0034-5288(02)00187-X
Karuppannan A. K., Opriessnig T. (2018). Lawsonia intracellularis: revisiting the disease ecology and control of this fastidious pathogen in pigs. Front. Vet. Sci. 5. doi: 10.3389/fvets.2018.00181
Kassambara A., Mundt F. (2021) Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R package version 1.0.7. Available at: https://CRAN.R-project.org/package=factoextra.
Katoh K., Standley D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 4, 772–780. doi: 10.1093/molbev/mst010
Kroll J. J., Roof M. B., Hoffman L. J., Dickson J. S., Harris D. L. (2005). Proliferative enteropathy: A global enteric disease of pigs caused by Lawsonia intracellularis. Anim. Health Res. Rev. 6, 173–119. doi: 10.1079/ahr2005109
Lawson G., Gebhart C. (2000). Proliferative enteropathy. J. Comp. Pathol. 122, 77–100. doi: 10.1053/jcpa.1999.0347
Le S., Josse J., Husson F. (2008). FactoMineR: An R package for multivariate analysis. J. Stat. Software 25, 1–18. doi: 10.18637/jss.v025.i01
Leite F. L. L., Singer R. S., Ward T., Gebhart C. J., Isaacson R. E. (2018). Vaccination Against Lawsonia intracellularis Decreases Shedding of Salmonella enterica serovar Typhimurium in Co-Infected Pigs and Alters the Gut Microbiome. Sci. Rep. 8, 2857. doi: 10.1038/s41598-018-21255-7
Levesque C. L., Hooda S., Swanson K. S., de Lange K. (2014). Alterations in ileal mucosa bacteria related to diet complexity and growth performance in young pigs. PloS One 9, 9e. doi: 10.1371/journal.pone.0108472
Li H., Mas L., Li Z., Yin J., Tan B., Chen J., et al. (2021). Evolution of the gut microbiota and its fermentation characteristics of ningxiang pigs at the young stage. Animals 11, 638. doi: 10.3390/ani11030638
Lin H., Peddada S. D. (2020). Analysis of compositions of microbiomes with bias correction. Nat. Commun. 11, 3514. doi: 10.1038/s41467-020-17041-7
Loo Y. T., Howell K., Suleria H., Zhang P., Gu C., Ng K. (2022). Sugarcane polyphenol and fiber to affect production of short-chain fatty acids and microbiota composition using in vitro digestion and pig faecal fermentation model. Food. Chem. 385, 132665. doi: 10.1016/j.foodchem.2022.132665
McCormack U. M., Curião T., Metzler-Zebeli B. U., Wilkinson T., Reyer H., Crispie F., et al. (2019). Improvement of Feed Efficiency in Pigs through Microbial Modulation via Fecal Microbiota Transplantation in Sows and Dietary Supplementaryplementation of Inulin in Offspring. Appl. And Env. Microbiol. 85, 22. doi: 10.1128/AEM.01255-19
McOrist S., Smith S. H., Green L. E. (1997). Estimate of direct financial losses due to porcine proliferative enteropathy. Veterinary Rec. 140 (22), 579–581. doi: 10.1136/vr.140.22.579
Metzler-Zebeli B. U., Zijlstra R. T., Mosenthin R., Gänzle M. G. (2011). Dietary calcium phosphate content and oat β-glucans influence gastrointestinal microbiota, butyrate-producing bacteria and butyrate fermentation in weaned pigs. FEMS Microbiol. Ecol. 75, 402–413. doi: 10.1111/j.1574-6941.2010.01017.x
Muwonge A., Karuppannan A. K., Opriessnig T. (2021). Probiotics mediated gut microbiota diversity shifts are associated with reduction in histopathology and shedding of Lawsonia intracellularis. Anim. Microbiome 3, 22. doi: 10.1186/s42523-021-00084-6
Netea M. G., Joosten L. A. B., Latz E., Mills K. H. G., Natoli G., Stunnenberg H. G., et al. (2016). Trained immunity: A program of innate immune memory in health and disease. Science. 352, 6284. doi: 10.1126/science.aaf1098
Netea M. G., Quitin J., van der Meer J. W. M. (2011). Trained immunity: A memory for innate host defence. Cell Host Microbe 9, 355–361. doi: 10.1016/j.chom.2011.04.006
Pereira C. E. R., Resende T. P., Armién A. G., Laub R. P., Vannucci F. A., Santos R. L., et al. (2020). Survival of Lawsonia intracellularis in porcine peripheral blood monocytes-derived macrophages. PloS One 15, 7. doi: 10.1371/journal.pone.0236887
Price M. N., Dehal P. S., Arkin A. P. (2010). FastTree 2–approximately maximum-likelihood trees for large alignments. PloS One 5, 3. doi: 10.1371/journal.pone.0009490
Quan J., Wu Z., Ye Y., Peng L., Wu J., Ruan D., et al. (2020). Metagenomic characterization of intestinal regions in pigs with contrasting feed efficiency. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.00032
Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, 90–96. doi: 10.1093/nar/gks1219
Quintin J., Saeed S., Martens J. H. A., Giamarellos-Bourboulis E. J., Ifrim D. C., Logie C., et al. (2012). Candida albicans Infection Affords Protection against Reinfection via Functional Reprogramming of Monocytes. Cell Host Microbe 12, 223–232. doi: 10.1016/j.chom.2012.06.006
Rajput M., Momin T., Singh A., Banerjee S., Villasenor A., Sheldon J., et al. (2022). Determining the association between gut microbiota and its metabolites with higher intestinal Immunoglobulin A response. Vet. Anim. Sci. 19, 100279. doi: 10.1016/j.vas.2022.100279
Selim K. M., Reda R. M. (2014). Beta-glucans and mannan oligosaccharides enhance growth and immunity in nile tilapia. North Am. J. Aquaculture 77, 1. doi: 10.1080/15222055.2014.951812
Shen J., Yang L., You K., Chen T., Su Z., Cui Z., et al. (2022). Indole-3-acetic acid alters intestinal microbiota and alleviates ankylosing spondylitis in mice. Front. Immunol. 13. doi: 10.3389/fimmu.2022.762580
Song Y., Chen K., Lv L., Xiang Y., Du X., Zhao G., et al. (2022). Uncovering the biogeography of the microbial community and its association with nutrient metabolism in the intestinal tract using a pig model. Front. Nutr. 9. doi: 10.3389/fnut.2022.1003763
Vetvicka V., Oliveira C. (2014). β(1-3)(1-6)-D-glucans modulate immune status in pigs: potential importance for efficiency of commercial farming. Ann. Transl. Med. 2, 16. doi: 10.3978/j.issn.2305-5839.2014.01.04
Viott A. M., Lage A. P., Cruz Junior E. C. C., Guedes R. M. C. (2013). The prevalence of swine enteropathogens in Brazilian grower and finish herds. Braz. J. Microbiol. 44, 1. doi: 10.1590/S1517-83822013005000033
Yan H., Wie W., Hu L., Zhang Y., Zhang H., Liu J. (2021). Reduced feeding frequency improves feed efficiency associated with altered fecal microbiota and bile acid composition in pigs. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.761210
Keywords: lawsonia intracellularis, yeast β-glucans, finishing pigs, body weight, microbiota, trained immunity
Citation: Rhayat L, Even G, Kiros TG, Kuhn G, Lebrun-Ruer S, Audebert C and Schulthess J (2023) The effect of yeast-derived β-glucans in reducing the adverse outcome of Lawsonia intracellularis in finishing pigs. Front. Anim. Sci. 4:1245061. doi: 10.3389/fanim.2023.1245061
Received: 23 June 2023; Accepted: 15 November 2023;
Published: 01 December 2023.
Edited by:
David L. Harmon, University of Kentucky, United StatesReviewed by:
Tao Ran, Lanzhou University, ChinaMichael D. Flythe, United States Department of Agriculture, United States
Copyright © 2023 Rhayat, Even, Kiros, Kuhn, Lebrun-Ruer, Audebert and Schulthess. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lamya Rhayat, l.rhayat@phileo.lesaffre.com; Julie Schulthess, j.schulthess@phileo.lesaffre.com