
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Anim. Sci. , 23 February 2023
Sec. Animal Nutrition
Volume 4 - 2023 | https://doi.org/10.3389/fanim.2023.1066388
This article is part of the Research Topic Ingredients for Poultry View all 6 articles
 Caven Mguvane Mnisi1,2*
Caven Mguvane Mnisi1,2* Chika Ethelbert Oyeagu3Eunice Amaka Akuru4
Chika Ethelbert Oyeagu3Eunice Amaka Akuru4 Oziniel Ruzvidzo5Francis Bayo Lewu3
Oziniel Ruzvidzo5Francis Bayo Lewu3Diversification and expansion of the poultry industry with fast-growing and highly prolific birds such as the quail (Coturnix coturnix), could contribute significantly in achieving global food and nutrition security. However, sustainable intensification of the quail relies on the cost of dietary ingredients used during feed formulations. The use of non-conventional energy sources such as sorghum, millet, and cassava in lieu of expensive energy sources such as maize, could ensure sustainable quail businesses. Generally, alternative feedstuffs should be cost-effective and possess comparable nutritional qualities as maize. In tropical countries such as South Africa, the use of sorghum, millet, and cassava in quail diets can serve as ideal alternatives because they have relatively comparable energy values as maize. Furthermore, these alternatives are largely available and easily accessible in many farming areas of South Africa. However, the presence of antinutritional factors such as tannins, cyanides and phytic acid, among others, as well as their high fiber levels may limit their utilization in quail nutrition. Nevertheless, attempts have been made to develop improved varieties with low antinutrient compositions, for instance, low-tannin sorghum varieties are increasingly being fed to poultry birds. Furthermore, there is growing evidence that certain processing techniques such as sun-drying, soaking, boiling and fermentation, among others, can lower the concentrations of antinutrients in these alternative feedstuffs, thus increasing their feeding value. To this end, nutritional feeding trials on the positive effects of sorghum, millet and cassava in quail nutrition are inconsistent, mainly due to differences in cultivar type, harvesting site or environmental conditions, sampling, and handling methods amongst other factors. Thus, the present review aimed to discuss the potential of substituting maize with sorghum, millet, and cassava in quail diets.
Worldwide production of poultry products (meat and eggs) and their consumption has consistently increased over the years and is expected to continue to increase to meet the nutritional demand of the rapidly growing human population (Mardoia, 2016). According to Thea et al. (2018), the increased consumption of poultry products is because they are readily accessible and affordable sources of animal protein with low-fat content. In developing countries, poultry farming is classified into commercial and traditional sub-sectors. The commercial sub-sector is focused on large-scale production of poultry products for nutritional and economic gains, while the traditional sub-sector is largely found in rural settlements, where the livelihood of most households depends on it (Pius et al., 2021). The traditional sub-sector contributes about 80% of the country’s rural poultry flock and is a major source of animal protein as well as a source of ready income for 90% of people residing in rural households (Wong et al., 2017).
The increasing demand for animal protein calls for the use of sustainable alternative feed resources as well as poultry birds that can convert non-conventional feed resources into usable protein for human consumption. The adoption of non-conventional feed ingredients could be a viable strategy through which food and nutritional security can be achieved. The increasing demand for animal protein necessitates the diversification of the poultry industry, which can be accomplished by rearing fast growing and highly adaptable birds such as the quail (Coturnix coturnix). According to Shanaway (2008), quails have several advantages such as early maturity, excellent laying capacity, low feed consumption, low floor space requirements, high immunity against diseases, relatively low meat fat content, and a high nutritive and medicinal value of the eggs.
To ensure sustainable intensification of quails, high feeding costs need to be addressed to reduce the cost of production. The use of less expensive feed ingredients such as low-tannin sorghum, millet, or cassava meal, would reduce the over-reliance on maize, which constitutes the largest proportion in conventional poultry diets. Their use would also reduce competition for maize grain between animals and humans, and thus reducing feed-food competitions and the pressure faced by maize producers. Likewise, the use of inexpensive and locally available alternative energy sources may reduce feed costs and promote food and nutrition security, more especially in tropical countries. Sorghum and millet grains are good sources of energy, vitamins, minerals, phytosterols, and antioxidant phenolic compounds (Okarter and Liu, 2010) while cassava meal is rich in minerals, carbohydrates as well as the B and C vitamins (Odo and Nnadi, 2013). The synergistic effects of these biochemical components could improve the nutritional well-being of birds and their overall quality of meat. Sorghum and millet produce grains that are high in nutrients and can tolerate drought conditions (Hossain et al., 2022). Some authors have reported desirable results when sorghum, millet and cassava products were incorporated in diets of other poultry species (Odo and Nnadi, 2013; Almeida et al., 2020). Unfortunately, their feed value is largely unknown for the quail. Thus, the present review discusses the potential of using sorghum, millet, and cassava products as alternative energy sources in quail production. It further discusses the effects of these alternative energy sources on poultry production, health, and meat quality traits.
Chang et al. (2005) reported over 100 different wild breeds of quails that exists, which are mostly found in North America and Asia. There are two main groups of these breeds, which are the New-World and Old-World quails. Cortunix quails, which are commonly used for meat and egg production, are species of the Old-World quail group that belongs to the family Phasianidae of the order Galliformes, which include chickens, partridges, and pheasants (Brian, 2009). The South African poultry subsector has recently seen the evolution of Jumbo quail (Cortunix sp.), a meat type bird developed for meat production. Many domesticated quail breeds reach maturity at six to eight weeks of age and have the same incubation period of 17 to 19 days (Ainsworth et al., 2010). Quail hens start laying closer to 100 eggs per cycle, amounting to about 300 eggs in their first year of life under proper management (Karrar et al., 2019). Santos et al. (2013) reported that the laying cycle of quails varies between 8 and 12 months for breeders, and between 8 and 14 months for layers, and about 50% of the quails are expected to be productive between 42 and 45 days. Quails attain a market average weight of 250 g at 6 weeks of age (Marareni and Mnisi, 2020) and at the same time, reaching their peak egg production (Bagh et al., 2016). Jumbo quails are highly desirable for their high meat-to-bone ratio and excellent egg laying characteristics (Ortlieb, 2015). They have common features with the Japanese quails, namely plumage that has uniform dark rusty brown-red feathers on the cheek and breast for males (Mizutani, 2003), and pale breast feathers with black sparkling-colored spots on the throat and upper breast for females (Vali, 2008). Jumbo quails are over twice the size of an average Japanese quail, and they require double the space of the common Japanese quails. Although they are mostly found in the wild woody and forests of Europe, Asia, and Africa, their (Jumbo quails) role in contributing to food and nutrition security is gaining rapid global recognition (Mnisi et al., 2021).
Quail production is gaining popularity mostly, in the developing countries, due to its significant role in the economic empowerment of the resource poor segment of the society, bridging the protein malnutrition challenge, and fitting well in the farming systems that are commonly practiced (Mnisi et al., 2021). Quail farming is practiced at levels ranging from small-scale to large-scale operations. Regardless of the many benefits of quail farming, there are some constraints such as bird sensitivity, cannibalism, proper nutrition, access to market, problems of egg production, and low body weight that may hinder a successful high-input production of the birds. Further, because the weight of chicken meat far outweighs that of quail meat, it may discourage farmers from venturing into quail farming.
In South Africa and other developing countries, there are currently no feed mill producers that specialize in quail feed production (Mnisi et al., 2021). Low level of dietary protein can reduce growth and egg production performance of the quail bird. Majority of the farmers are familiar with the use of conventional feed hence they find it difficult to engage in quail farming because quails do not have an established standard feed compared to chickens. Many farmers do not have adequate knowledge of the nutritional profile of feedstuffs and nutritional requirements of quails due to lack of standard or conventional diets available for quails and, therefore, cannot formulate a balanced quail diet by themselves. This situation resulted in farmers offering chicken feeds to their quail birds and this is rather an erroneous practice, which seriously needs to be corrected.
Data from different research studies show significant differences that exists between laying quail and chicken hens with regards to protein and energy requirements (Table 1). The data on the nutritional requirements show a clear difference on the protein requirements of layer quail and chickens. Thus, offering chicken feed to quails over a long period of time, limits their optimum production potential and may also reduce their survival rate (Redoy et al., 2017). Prolonged feeding of chicken diets to quails may further result in suboptimal growth rates, shorter laying duration, and poor productivity. Unfortunately, the use of commercial supplements (e.g., protein and energy sources, and vitamin-mineral premixes) in quail diets incurs extra costs.
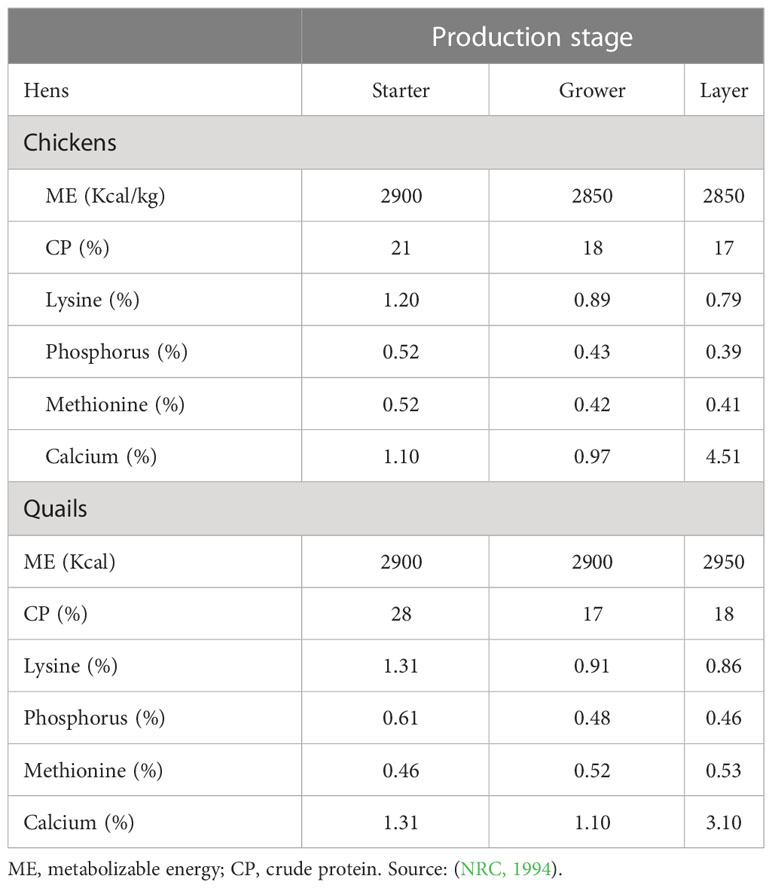
Table 1 Nutritional requirements of quail and chicken hens at different production stages.
The poultry industry is a major portion of the world’s agricultural sector especially in developing countries (Ali et al., 2012). The rapidly growing human population in these countries necessitates the need to establish alternative sources of dietary protein to reduce the risks associated with protein deficiencies. According to Mottet and Tempio (2017), quail farming has the potential to complement animal protein supply through the diversification of poultry production as well as due to their short generation intervals. Ali et al. (2012) reported that quail farming can help diversify, strengthen, and sustain the production of animal protein to close the gap between its demand and supply. Quail farming varies from subsistence to large-scale commercial operations (Kingori, 2011), and is more profitable than any other poultry subsector (Mnisi et al., 2021). Quail farming comes with great benefits due to the faster growth rates, early sexual maturity, and high prolificacy and adaptability (Marareni and Mnisi, 2020).
Quails have a short generation interval and require less feed per adult bird per day and a relatively small floor space of about 150 to 250 cm² compared to broiler chickens (Faitarone et al., 2005). Quails have a more resistant ability against common avian diseases such as the Newcastle disease, infectious bronchitis, avian influenza, and fowl pox (Jatoi et al., 2013). Under proper management and nutritional practices, female quails can lay up to 200 eggs by 50 days of age in their first year of lay (Randall and Bolla, 2008). This is desirable as famers do not have to wait for longer periods before selling their products. In addition, quails can survive harsh environmental conditions, with low disease outbreaks, which suggests a relatively less usage of vaccinations. These birds have a high return rate with low cost of investment, high nutritional and market value as compared to any other poultry industries (Marareni and Mnisi, 2020). As a result of them being small sized birds, a flock of quails can thus be easily raised within a very small area possibly meant for rearing a single adult chicken. Quail farming can create a living for emerging farmers, who are active in increasing the domestic poultry produce through meat and egg production in low- and middle-income countries.
Quail meat and egg play a big role in the diets of humans (consumers). The study reported that quail meat is an ideal food for health-conscious individuals because the meat is all white with a higher concentration of protein and very low in fat and low-density lipoprotein cholesterol content. Moreover, quail eggs and meat contain some nutrients that are important for the physical and mental development of humans; the nutrients include unsaturated fatty acids, vitamins, essential amino acids, and phospholipids. Vanessa (2020) recognized quail meat and eggs as a crucial part of the diets for pregnant mothers, children, and geriatric and convalescent patients. The authors concluded that quail egg and meat constitute an essential part of the diets for the socially and economically marginalized sections of the society. Unlike chicken eggs, quail eggs do not cause allergies or diathesis and they have massive nutritional benefits even though their eggs are 5 – 6 times smaller than the chicken eggs - their nutritional worth is 4 – 5 times higher than the chicken eggs (Savanna, 2020). Quail eggs contain about 2.47% less fat compared to chicken eggs, and up to 13% protein content compared to 11% in chicken eggs (Peralee, 2022).
Additionally, quail eggs contain around 140% of vitamin B1 than 50% in chicken eggs. More so, quail eggs contain potassium and iron as much as five times compared to chicken eggs. Quail eggs provides animal proteins that are not expensive and are good source of essential amino acids, sex hormones, important vitamins and minerals (zinc, iron, nitrogen, etc.) for human health than chicken eggs (Vanessa, 2020). Regular consumption of quail eggs could also help to fight against diseases such as digestive tract disorders, stomach ulcers, among others. Quail eggs also promote memory health, increase brain activity, strengthen the immune system, and stabilize the nervous system (Tunsaringkan et al., 2013). These excellent qualities of quail birds are of interest to many farmers and consumers and such interest has now significantly propelled the gradual proliferation of quail farming to assure food and nutritional security for all.
The essential components of poultry diet must be supplied in adequate amounts to meet up with the bird’s requirements for optimum growth, maintenance, egg production and reproduction (Ferket and Gernat, 2006). Energy is a constituent of energy-yielding nutrients during the period of metabolism when they are oxidized (NRC, 1994). The energy requirement of quails significantly depends on the production stage, reproductive status, ambient temperature, and age of the birds (Shibi et al., 2019). However, there is a gap in literature relating to the nutritional requirements of quail compared to chickens especially in tropical and sub-tropical regions, this is because more research has focused on the nutritional requirements of conventional broiler and layer chickens. Nonetheless, an energy requirement ranging from 2600 to 3000 kcal/kg of the metabolizable energy (ME) in a diet of a growing quail was found to be ideal for the temperate environmental regions, whereas results under the tropical environmental condition, indicated that the ME for a growing and laying quail is 2800 ME kcal/kg and 2550 ME kcal/kg, respectively (Redoy et al., 2017). Bawa (2010) reported that increasing the dietary energy levels from 2600 to 2800 ME kcal/kg increase the body weight gain and significantly influence the efficiency of feed utilization and consumption. The main dietary energy sources are cereal grains, which are also the major ingredients for humans and livestock. Fat sources such as animal tallow, lard and vegetable oils are sometimes added to the diet if high energy is required (Shrivastav, 2000).
To achieve optimal performance in quails, dietary crude protein must be provided with sufficient levels of essential and non-essential amino acids to meet the bird’s metabolic requirements (Mnisi et al., 2021). Protein provides amino acids necessary for egg production and tissue growth. The amino acid and dietary protein requirement of a quail is influenced by growth stage, egg production, dietary metabolizable energy content, overall feed quality, and the age of the birds (Redoy et al., 2017). Table 2 shows the energy, mineral and vitamin requirements of quail production while the protein and amino acid requirements are presented in Table 3. Protein quality is generally based on the availability and composition of amino acids from the feedstuff following digestion (Quaresma et al., 2022). Out of a total of 19 amino acids, only 13 are required by quails, and these are considered as essential because they must be supplied in the diet and cannot be synthesized in the quail’s body. The remaining six are considered non-essential because they are synthesized by the body and therefore, no need for their supply in the diet. As stated by Hou and Wu (2018), the 13 essential amino acids include arginine, glycine, cystine, isoleucine, leucine, lysine, histidine, methionine, phenylalanine, valine, tyrosine threonine and tryptophan.
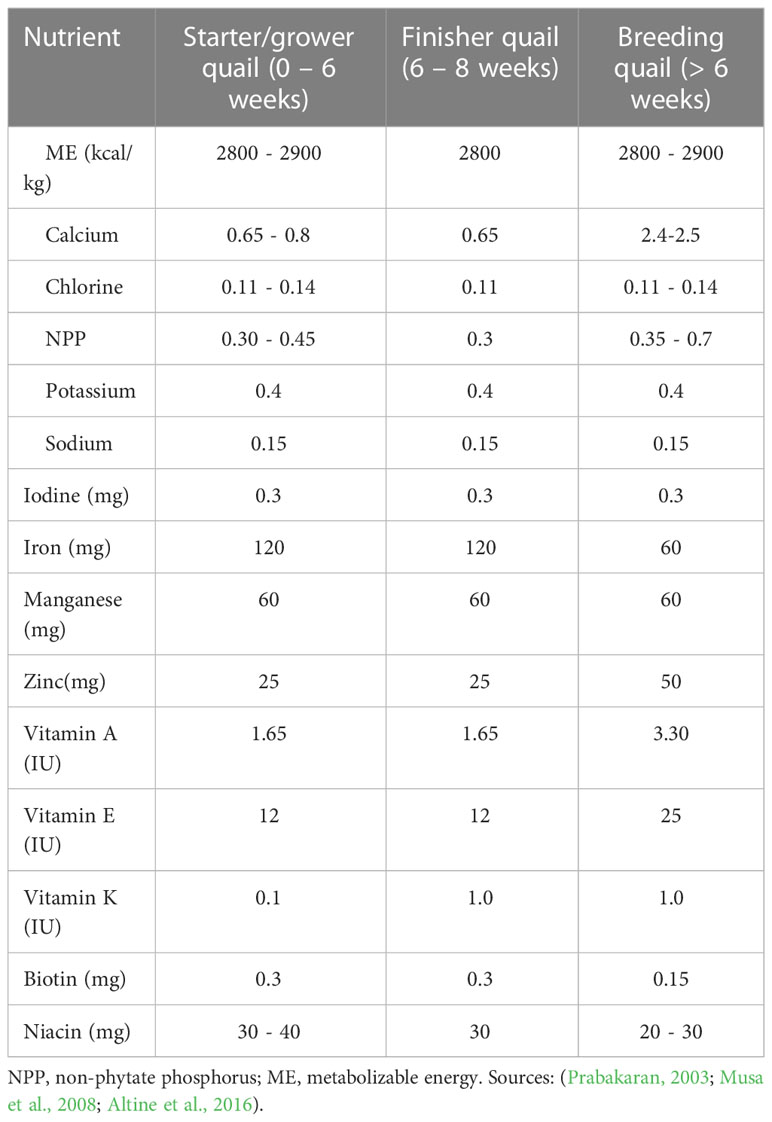
Table 2 Energy, mineral and vitamin requirements of quails (% DM, unless otherwise stated).
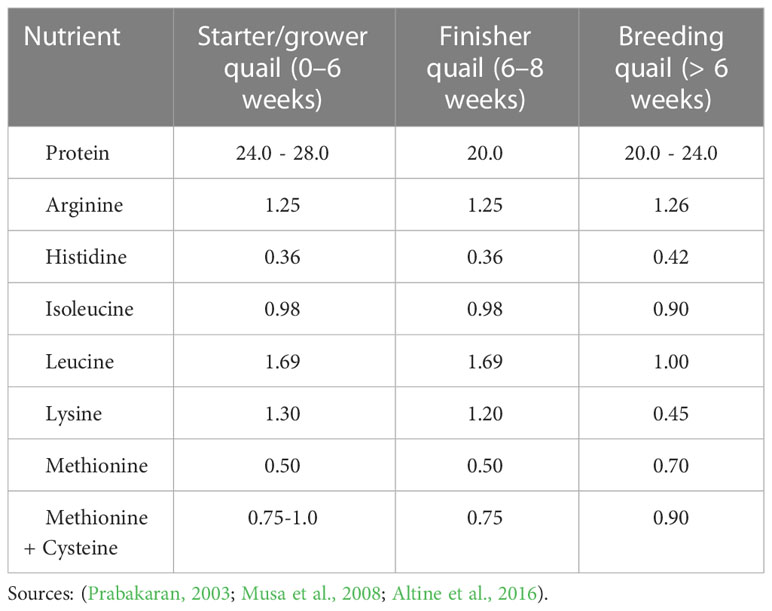
Table 3 Protein and amino acid requirements of quails (% DM, unless otherwise stated).
According to Babatunde et al. (2016), crude protein levels of 20% and 24% are recommended for quails during their production and rearing periods, respectively. Altine et al. (2016) also reported that quails were successfully raised on turkey starter diets containing 25 – 28% crude protein levels. A dietary crude protein of 24% is required in starter diets for quails and may be reduced to 20% when quails are 3 weeks of age (Rafh, 2018). Rabie et al. (2015) reported that quails fed with 26% and 21.6% crude protein levels from day 1 to 21 and from day 22 to 35 of age, respectively, had good performance. Four different crude protein levels (20, 22, 24 and 26%) were evaluated for the Japanese quails and it was found that, after cysteine, methionine and lysine requirements were met, 20% crude protein level from day 1 to 42 of age resulted in best performance (Nwokedi et al., 2010). Thus, this indicates that the protein requirements decrease with age. There was an improved feed conversion and enhanced egg weight when the crude protein level of the diet was raised above 18% (Brando et al., 2021). However, dietary protein and amino acid requirements are influenced by age, stage of production and the ingredients used to formulate the diet.
According to Wenhold and Faber (2009), adequate and fresh clean water may be considered the single most significant nutrient. The amount of water required depends on the relative humidity and environmental temperature, stage of egg production, rate of growth, efficiency of kidney reabsorption of water and the composition of the individual bird’s diet (Al-Saffar and Rose, 2002). Fresh clean water especially under tropical environmental condition should be provided continuously to all birds. According to Altine et al. (2016), quails require water at least twice per day. Quails require more water when the environmental temperatures are high especially during the hot weather or when there are excess salts included in their dietary feed. Quails must have access to fresh and clean drinking water ad libitum because water restriction for a longer period, leads to high mortality rate (Randall and Bolla, 2008). The water requirement of a quail chick changes with the quantity and quality of the feed’s dry matter and age. Quail water-to-feed intake is typically a 3:1 to 4:2 g/g per body weight from day 12-29 of age (Altine et al., 2016).
Cereal grains play a major role as sources of energy and essential nutrients in the everyday human and livestock diet through direct consumption. In African countries including South Africa, the human population largely relies on cereals’ grains as their main energy source (Laskowski et al., 2009). According to Grote et al. (2021), the total crop production during 2016 reached 2577.85 million tons whereas the production of cereal grains, other than wheat and rice, used primarily for animal feed or brewing, reached a total of 1330.02 million tons. The term “cereal” refers to members of the Gramineae family that consist of nine species namely: corn (Zea), wheat (Triticum), barley (Hordeum), oat (Avena), rice (Oryza), rye (Secale), sorghum (Sorghum), and millet (Pennisetum) (Haroon, 2013). The top cereal grains produced worldwide in year 2014, ranked based on million tons (Mt), are corn (1253.6 Mt), rice (949.7 Mt), wheat (854.9 Mt), barley (146.3 Mt), oat (23.2 Mt), and rye (15.8 Mt) (Grote et al., 2021).
Maize (Zea mays L.) is a staple food for most of the population in the Southern African region and the world at large, and a major feed grain for livestock. Approximately 40% of maize is used in the food sector whereas 38% is used in the animal feed industry. Maize is the first most important grain crop in South Africa followed by rice (Oryza sativa L.), barley (Hordeum vulgare L.), wheat (Triticum aestivum L.), sorghum (Sorghum bicolor L.), and pearl millet (Pennisetum miliaceum L.), and is produced throughout the country under diverse environments (Shiferaw et al., 2011). Maize production significantly depends on the agricultural production practices employed, and the application of production inputs that sustain the environment. In developed countries, maize is mainly consumed as a second cycle produce, in the form of dairy, meat and eggs, whereas in developing countries, maize is consumed directly as first cycle produce and serves as a staple food for over 500 million people (Shiferaw et al., 2011). In South Africa, most people regard maize as a breakfast cereal. However, in a processed form, it is also regarded as starch and fuel (ethanol) (Stefan et al., 2009). Starch, in return, is involved in enzymatic conversion into products such as dextrin, sorbitol, lactic and sorbic acid, and appears in household material such as ice cream, beer, shoe polish, syrup, glue, ink, fireworks, batteries, cosmetics, mustard, paint and aspirin (Stefan et al., 2009). Although some studies have demonstrated the potential of millet, cassava, and sorghum to replace maize in conventional poultry diets, it is important to firstly consider their nutritional profiles against the daily nutritional requirements of the quail. This is to ensure that they supply sufficient nutrients that would promote growth performance without compromising their health status.
Mature maize kernel is composed of 4 – 5% oil, 8 – 10% protein and 70 – 75% starch (Duvick and Cassman, 2009). However, there are large variations in the relative concentrations of these components between parts of the kernel. Furthermore, there are two major structures of the kernel namely the germ (embryo) and the endosperm, constituting about 10% and 80% of the mature kernel dry weight, respectively. Maize is an important food crop for humans and a high-energy feed for animals (Klopfenstein et al., 2013). Furthermore, in the human diet, it is a good source of vitamin B1, B5 and C, folate, fiber, manganese, and phosphorus. Maize oil is genetically accepted as high quality, determined by the low linolenic and high linoleic acid contents (Krishnendu et al., 2019). Laurie et al. (2004) reported that the lipid content in maize oil is probably controlled by a multi genetic inheritance of many genes and is also complex.
Sorghum and millet are one of the most important food crops in the arid and semi-arid regions of Asia and Africa (Orr et al., 2016), where they perform and thrive well under harsh climatic and environmental conditions in marginal regions with poor soil fertility. Besides their major use as food crops, the two cereals are also used as bird and cattle feed, fodder, and forage and, recently, as bioenergy crops. During the dry seasons, sorghum stalks are used as feed for livestock, especially in Asian countries whereas in America, sorghum grain is one of the most important sources of animal feed in addition to forage sorghum (Zarei et al., 2022). Millet is a collective term that is commonly used when referring to several small-seeded annual grass grain crops (Lawler, 2009). Millet production is generally limited to fields with poor rainfall and low soil fertility conditions. Prasad and Staggenborg (2010) reported that millet production is only about 2% of the world cereal grain production and is one of the most important staple food crops in semi-arid regions. In the same report, it is stated that Africa and Asia account for about 95% of the total millet production in the world. Furthermore, it is reported that Asia accounts for 40% of millet production. The comparable chemical composition of maize, sorghum and millet is presented in Table 4.
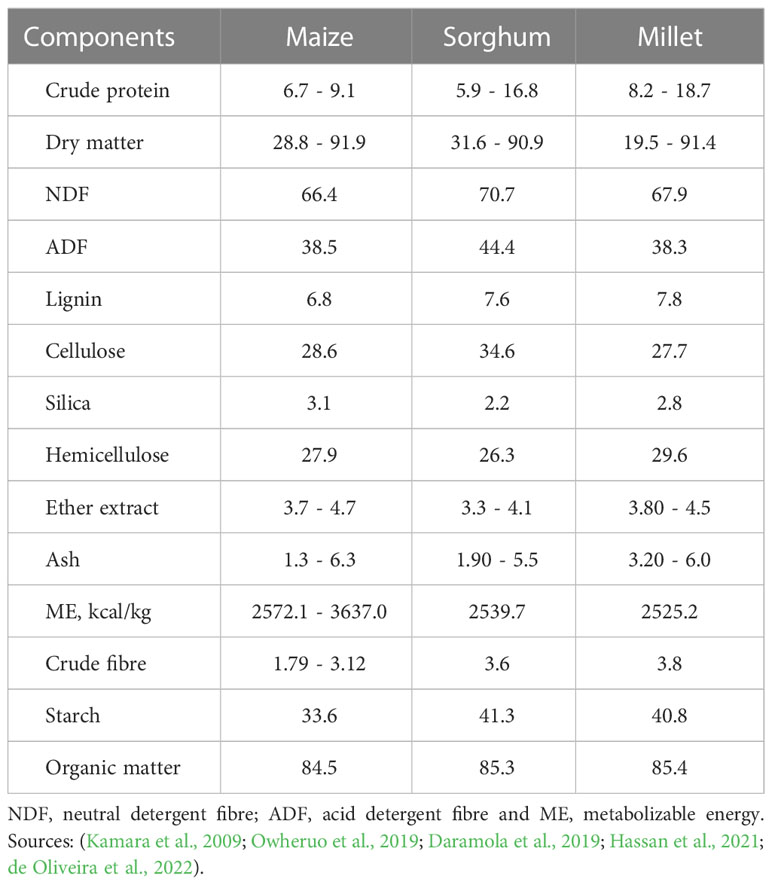
Table 4 Chemical composition of maize, sorghum, and millet (%, dry matter).
The cultivation of sorghum is generally very low and, the cereal is mainly consumed locally, mainly in areas where it is cultivated (Mundia et al., 2019). The use of the old sorghum cultivar in quail diet is so much restricted due to the considerable amount of anti-nutrients such as tannins. These tannins restrict the activities of fibrolytic and proteolytic enzymes, which then reduces nutrient digestibility (Yanza et al., 2021). Meanwhile, the development of a hybrid low-tannin sorghum cultivar has been evaluated in poultry and pig diets. Indeed, the inclusion of 50% low-tannin sorghum did not affect the utilization of nutrients or pose any toxicity to quails (Yanza et al., 2021). A complete substitution of maize with sorghum did not negatively affect the growth traits and efficiency of diet utilization in quails (Masenya et al., 2021). Similarly, complete substitution of maize with low-tannin sorghum had no negative effect on quail and broiler growth traits, gut development, and carcass yield (Farahat et al., 2020).
Millet is a valuable energy source because it has high metabolizable energy (2900 kcal/kg ME), which can be compared favorably to maize (3330 kcal/kg ME) (Farahat et al., 2020). Millets have up to 40% concentration of methionine and lysine than maize (Liu et al., 2021). The nutritional profile of millet shows that it can be easily used to replace maize in quail diet and more so, easily farmed in the tropical conditions because of its drought and heat tolerant attributes. Furthermore, millet has been found to encourage comparable performance without compromising the health status of birds (Hassan et al., 2021). Pearl millet does not negatively change the growth traits, carcass parameters and gut development of quail birds (Liu et al., 2021). The complete replacement of maize with millet in quail and broiler diets enhanced growth and the health status of birds (Masenya et al., 2021).
Cassava (Manihot esculenta) is another alternative energy source for quail birds, and it belongs to the Euphorbiaceae family, indigenous to South America but has become an important part of the diet of many Africans (Almeida et al., 2020). Cassava is a wooden shrub grown on a wide scale in several sub-Saharan African countries and supplies 13 times more energy yield per hectare compared to maize. Cassava is tolerant to harsh tropical climatic conditions and can thrive in drought and poor soil conditions (Okogbenin et al., 2013). A 2016 report showed that the highest producers of cassava were Nigeria, Thailand, Brazil, Indonesia, Ghana, and Democratic Republic of Congo, with a combined total annual production of over 15 million tons (Amelework et al., 2021). Apart from its consumption by humans, cassava is highly demanded for use in the agro-allied and pharmaceutical industries to produce ethanol, plywood, and textile (Dimkpa and Theophilus, 2021). The high demand for cassava by humans, livestock and industries competes with its use in poultry feeding. Nevertheless, the by-products resulting from industrial processing of cassava for food can be channeled into feeding poultry birds, including quails (Morgan and Choct, 2016). The rich carbohydrate content and multifunctional role of cassava makes it a desirable alternative energy source. The chemical compositions of samples of the different parts of cassava are presented in Table 5. The nutrient contents are relatively comparable to those of maize as shown in Table 4 above.
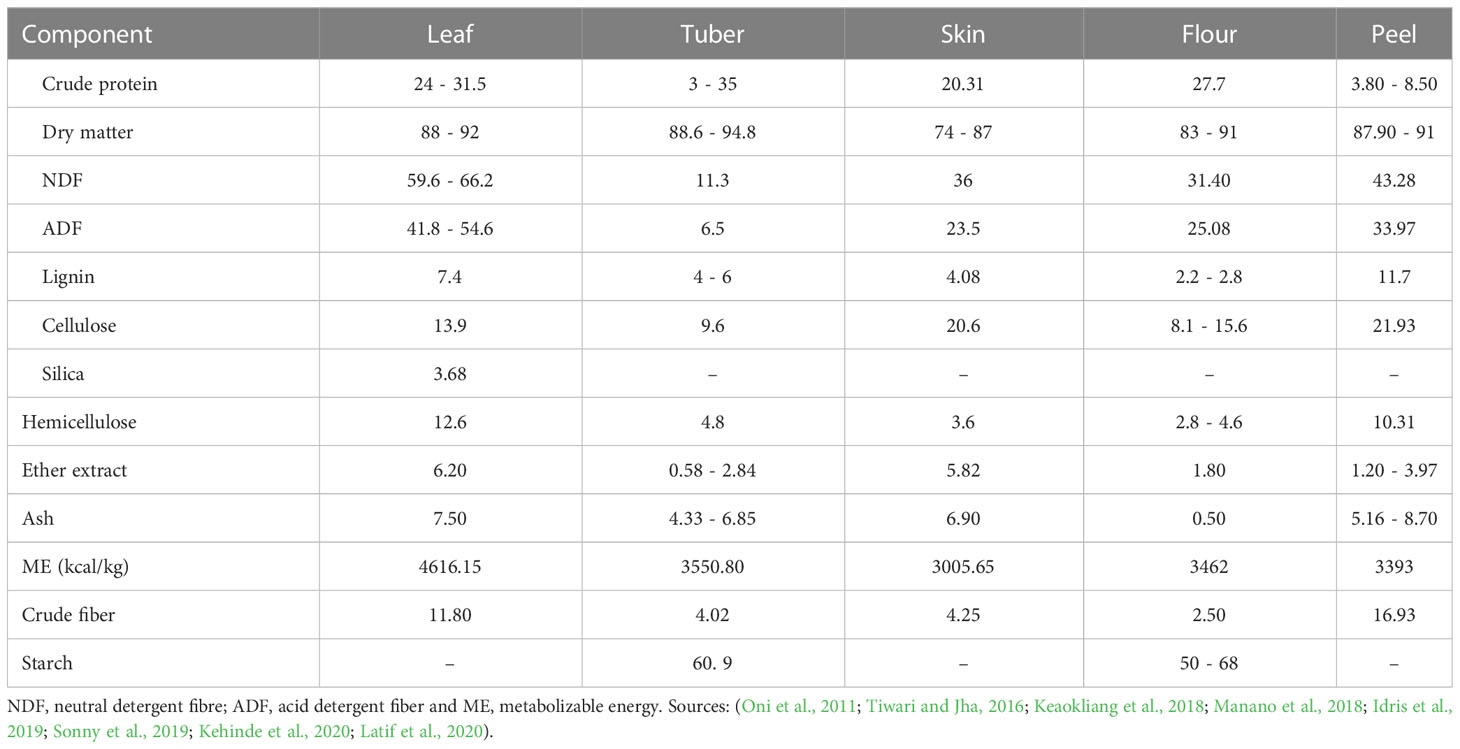
Table 5 Chemical compositions of the different parts of cassava (%, dry matter).
Different parts of cassava such as root, leaves, pulp, foliage, and peels have been found to be suitable for inclusion into poultry diets (Amata, 2014; Diarra, 2018). However, the utilization of cassava in poultry diet is limited by its low protein concentrations and deficiency of fat, minerals, vitamins, and essential amino acids such as methionine (Chauynarong et al., 2009). Cassava also contains an anti-nutrient component called cyanogen, which if not properly processed before consumption, can cause chronic health conditions in humans, including toxicity, neurological disorders amongst others. The two main forms of cyanogen in cassava are linamarin (93 - 95%) and lotaustralin (5 - 7%). Over the years, efforts have been made to employ certain processing methods to lower the high fibre and cyanide contents of cassava, thereby making it easier to incorporate into poultry feeds. Sun-drying and fermentation appeared to be the most common, among the various methods, used to reduce the cyanide content and improve its feeding value (Morgan and Choct, 2016). Other methods such as cooking, soaking, boiling, ensiling, oven-drying, and pelleting, have also been used (Nebiyu and Getachew, 2011; Mbanjo et al., 2021; Baguma et al., 2022), amongst others.
Fermentation methods such as solid-state fermentation, use of non-starch polysaccharide or bacteria proteins, have had varying degrees of effectiveness in lowering down the hydrogen cyanide (HCN) and fiber levels in cassava meal, whilst increasing the crude protein content. A significant HCN reduction of 14 mg/kg from a 400 - 440 mg/kg range was reported in the root meal of cassava by solid-state fermentation (Nambisan, 2010). A 22.1% increase in crude protein content was also recorded in cassava pulp upon fermentation with Aspergillus niger and ammonium nitrate (Oboh et al., 2002; Yafetto, 2018). Sun-drying reduced the HCN in cassava root meal from 416 mg/kg to 42 mg/kg (Morgan and Choct, 2016). The substitution of maize with 50 – 70% fermented cassava peel meal in quail diet enhanced egg production and feed utilization (Oladunjoye and Ojebiyi, 2010). According to Malik et al. (2018a), up to 75% replacement of maize with fermented cassava peel meal did not have any negative effects on the production of eggs and egg quality parameters. The authors concluded that up to 50% replacement of maize with fermented cassava peel meal resulted in optimum growth performance while a 75% fermented cassava peel meal inclusion showed better carcass qualities in growing quails. It was reported that replacement of maize with up to 50% sun-dried cassava peel meal in quail diet, showed optimum growth performance in growing quails, while a 25% replacement produced an optimum egg production at the laying phase (Malik et al., 2018b). Generally, birds could tolerate cassava peel at 50% replacement at the expense of maize with regards to growth traits, egg production, meat quality and nutrient digestibility (Onyimonyi and Ugwu, 2007; and Malik et al., 2018b).
Energy feedstuffs are known to supply body heat, boost growth, provide maintenance needs, and increase production in poultry birds (Inaku et al., 2011). Utilizing alternative energy sources in lieu of maize, which attracts stiff competition from humans and other agro-allied sectors, is a veritable option to sustain the increased production of poultry. Masenya et al. (2021) revealed that replacing maize in the diet of Jumbo quails with pearl millet, enhanced the quail’s body weight gain and feed utilization efficiency. However, the authors noticed that replacing maize with whole sorghum in the diet of Jumbo quails, depressed the feed intake (FI) and growth performance, particularly from day 21 - 35. The poor growth performance indices of the quails could be attributed to the limiting effect of karifin, a major anti-nutrient component in sorghum, which is hydrophobic and thus, poorly utilized by quails (Liu et al., 2015; Truong et al., 2015). Ferulic acid, endosperm cell wall, tannins, and phytate are other anti-nutritional components present in sorghum that reduce its digestibility (Beta and Corke, 2004; Taylor and Emmambux, 2010). There is evidence that sorghum starch is 7.05% less digestible than the starch in maize, mainly because maize starch granules are closer to karifin protein granules than those of sorghum (Truong et al., 2015).
Kanyinji and Moonga (2014) revealed that a 75% replacement of maize by rumen-filtrate fermented cassava meal, enhanced feed intake, and weight gain in Japanese quails. On a complete replacement of maize by fermented cassava meal, it was observed that the quails’ growth performance indices (FI, BW and BWG) were comparable to those of the control groups. Based on these observations, it is apparent that cassava meal could be used to completely replace maize in diets of quails, however, the meal is more potent in enhancing the performance indices of quails at 75% dietary replacement level. The reports of Almeida et al. (2020) also showed that a 10% dietary inclusion of the dried cassava residue did not impair the growth performance of laying quails. At a 60% replacement level, dietary sorghum resulted in increased final body weight and feed intake in Japanese quails, however, the values recorded were comparable to the quails fed with the control diet (Khaleel et al., 2021). Inclusion of the dried cassava residue promoted the development of the gizzard, which then provided an increased grinding ability for nutrient digestion (González-Alvarado et al., 2007). This may be due to the increased levels of fiber in the dried cassava residue that then promote a longer retention time of the nutrient in the organ, hence, promoting nutrient digestion and utilization.
Blood analysis is usually performed on animals subjected to nutritional trials to ascertain their physiological responses to the fed dietary treatments (Kwari et al., 2011). In hematological assays, certain biomarkers are used to assess overall blood count and motility, whereas in serum-biochemical analyses, biomarkers such as alanine aminotransferase, aspartate aminotransferase, among others, are evaluated to determine the effect of the test ingredients on liver and kidney functions (Egbuonu et al., 2013). Feeding sorghum and millet to Japanese laying quails as alternative energy sources to maize showed that the values recorded for the red blood cell count, hemoglobin concentration, mean cell hemoglobin, mean cell volume, and mean cell hemoglobin concentration, were within the recommended range, suggesting normal bone marrow activity and absence of anemia, whether macrocytic, autoimmune, or hypochromic (Aminu et al., 2021). The authors also observed that quails fed with millet and sorghum diets had higher neutrophil values compared to those on maize diet, indicating higher disease fighting abilities for the former than the latter. More so, the groups that received millet and sorghum had comparable values for total protein, glucose, chlorine, albumin, globulins, and calcium, suggesting that both additives could serve as alternative feed resources, capable of providing quails with energy required for normal metabolic and biochemical activities. In addition, the reduced concentrations of alanine aminotransferase and conjugated bilirubin resulting from the feeding of millet was indicative of its positive role in maintaining the normal function of the liver.
Masenya et al. (2021) reported that Jumbo quails fed with whole-grain sorghum diets had higher concentrations of monocytes, suggesting an increased response to disease infection. The authors also observed that low levels of calcium and amylase resulted from the feeding of whole-grain sorghum to quails, indicating that the diet was not properly ingested and utilized due to its large particle size. Moreover, the concentrations of alkaline phosphatase (ALP) were higher in the whole-grain sorghum-fed groups than those that received either maize, crushed sorghum, whole or crushed millet. High level of ALP is suggestive of malnutrition, hence, it is not recommended to feed whole sorghum to quails, for reasons of low disease resistance, and poor feed, energy, and mineral utilization (Masenya et al., 2021). In another study, the inclusion of cassava grit in quail diet increased the red and white blood cells in quails and this might have been due to the influence of the experimental diet on the blood traits, which showed that birds that consumed the experimental diets were healthy (Oyelami and Abu, 2017).
The gastrointestinal mucosa plays an important role in enhancing digestion, absorption, and immunomodulation in animals (Abdulla et al., 2010). Thus, an impairment in its integrity makes it permeable to invasion, exposes animals to increased infection rate and encourages the production of harmful bacteria instead of the positive ones (Lambert, 2009). There is a dearth of literature on the effect of energy sources on gut microbiota in quails. However, the study of Almeida et al. (2020) revealed that the villi height, crypt depth and villi height/crypt depth ratio of quails fed with varying dietary levels (2.5, 5, 7.5, 10%) of dried cassava residue did not differ from those that were fed with the control diet, suggesting that the different dietary levels of the dried cassava residue did not compromise the integrity of the intestinal epithelium in the birds. Nonetheless, future research should investigate the effect of alternative energy sources on gut microbiota of quail and other poultry species.
Carcass traits are important an economic index in quail farming. Hence, reduction in carcass weight and portion sizes will affect the pricing of poultry meat and the total revenue generated. It is thus important that the feed fed to quails is properly utilized to aid muscle gain and deposition of fat. Masenya et al. (2021) reported that total replacement of maize grain with whole sorghum in Jumbo quail diets, resulted in low carcass weight and reduced weights of cut portions such as breasts, thighs, drumsticks, and wings. However, feeding the quail birds with crushed sorghum diet increased the carcass weight and portion size, indicating the higher digestibility and carcass yield potential of the crushed sorghum meal (Bonos et al., 2010). The authors showed that Jumbo quails fed with whole sorghum diet had enhanced gizzard, liver, and large intestine weights, which could have been an anatomical adaptation mechanism to utilize the large particle sizes of whole sorghum grain. In broiler chickens, feeding whole sorghum was reportedly found to improve enzymatic digestive processes and stimulate the development of gizzard muscles (Xu et al., 2015). The findings of Almeida et al. (2020) revealed that inclusion of the dried cassava residue in the diet of laying quails, led to an increase in the relative weights of the gizzard and small intestines. Silva et al. (2017) reported a quadratic increase in carcass yield and gizzard weight when quails were fed with the cassava foliage hay-based diets as replacement for maize. The observed increased gizzard weight was attributed to the fact that at high inclusion level (20%), the cassava foliage was able to stimulate abrasive activities in the gizzard, owing to the fibrous nature of the diet.
Egg quality is of high economic value to the poultry industry. This is because poor egg quality decreases the value, production, and consumer acceptance of eggs, and reduces hatchability, quantity and quality of processed egg and egg products (Abdelqader et al., 2013). According to Sari et al. (2016), nutrition is a major factor that affects egg production and quality, especially the hatchability and fertility of eggs, quality of the chicks hatched and their growth and survival rates (Zaheer, 2015). Malik et al. (2018a) reported that upon feeding the sun-dried cassava meal to the Japanese quails as replacement for maize, there was a marked increase in hen-day egg production, hen-house egg production, yolk height, yolk index, albumen height and egg volume, particularly at the 25% and 75% replacement levels. However, the sun-dried cassava meal had no influence on yolk diameter, yolk weight, eggshell weight and thickness, Haugh unit, and egg shape index, as well as egg length. The study of Ojediran et al. (2019) showed that cassava vinasse meal can replace maize up to 21% in the diet of Japanese quails, without adversely affecting the growth, hematological and meat quality indices of the birds. Japanese quails fed with a 75% rumen-filtrate fermented cassava diet as replacement for maize, had superior egg weights when compared to quails fed with the control diet (Kanyinji and Moonga, 2014). Birds with heavier body weights are known to lay bigger-sized eggs. Thus, the superior egg weights recorded in quails fed with the 75% cassava meal diet could be attributed to their higher body weight compared to quails fed with the other dietary treatments (Oyeagu et al., 2015). Almeida et al. (2020) reported that a 10% dietary inclusion of dried cassava residue enhanced eggshell thickness in laying quails, suggesting an increase in the gizzard weights of the birds, and an ultimate increased endogenous enzyme secretion amongst others.
Freitas et al. (2014) observed that feeding Japanese quails with either white or red sorghum as replacements for maize grain, did not influence egg production, egg mass and egg yolk colour. It was also noted that paler yolks were recorded in the sorghum fed quails, coupled with lack of variation in the response by the white and red sorghum dietary groups to pigment inclusion, which was aimed to augment color deficiencies. The authors also reported that the variety of sorghum used had little or no influence on yolk color of quail eggs, even with the inclusion of pigments. There are reports that sorghum can partially (80%) or completely replace maize in the diets of laying quail or chicken hens, with no harmful effect on growth performance, however, not without the inclusion of pigments to the feed (Faquinello et al., 2004; Freitas et al., 2014). Impaired yolk pigmentation is common when quail birds are fed with sorghum grain, especially at high dietary levels. This is due to the low level of carotenoids present in sorghum than in maize. Hence, to augment the lack in intense yellow yolk color, pigment inclusion is encouraged (Freitas et al., 2014). Aminu et al. (2021) also revealed that the daily egg production of Japanese quail hens was not affected by complete replacement of dietary maize by either millet or sorghum. Abu and Judah (2019) evaluated the comparative effect of millet, sorghum, and cassava flours as alternative energy sources for the Japanese quails in place of maize and found that while egg weight, yolk weight and height were highest in quails fed with the sorghum-based diet, quails fed with millet had the highest yolk and egg shape indices compared to those that had received the maize and cassava-based diets.
The aim of engaging in poultry ventures is to minimize cost and maximize profits. Maize grains are usually expensive due to the high demand by humans for consumption and use in other agro-allied industrial processes (Tenebaum, 2008). Hence, utilizing alternative energy sources such as millet, sorghum, and cassava in the nutrition of quails would promote economic, social, and environmental sustainability (Masenya et al., 2021). According to Abu and Judah (2019), millet is an affordable substitute for maize in the diet of Japanese quails, compared to feeding the quails with either the cassava root meal or sorghum grain. The authors observed that feeding quails with millet resulted in a lowered economic efficiency of 3.17% compared to the 38.61% and 53. 05% recorded for the feeding of sorghum and cassava root flour meal, respectively. Silva et al. (2017) showed that replacing maize with cassava foliage hay in quail diet was cheaper, particularly at a 10% inclusion, as the relative gross margin (102.15) and mean relative return (145.19) were higher than the 100.00 and 143.83, respectively, recorded in the comparative group fed with maize. Malik et al. (2018a) revealed that at higher inclusion levels (beyond 50%), the dietary sun-dried cassava peel meal resulted in decreased profitability index and revenue from the sale of quail eggs. According to the authors, the decreased profit could have had occurred due to reduced feed intake in the quails. Also, the low feed intake could be associated with the increasing fiber level in the diet, coupled with the residue, hydrogen cyanide effect that resulted as the dietary levels of the cassava peel meal increased.
Meat quality describes the overall meat characteristics such as physico-chemical, morphological, biochemical, sensory, technological, hygienic, culinary, and nutritional properties (Fanatico et al., 2007). Accordingly, the most perceptible and important meat features that influence the initial and final quality judgment by consumers before and after purchasing a meat product include appearance, juiciness, texture, tenderness, wateriness, firmness, flavor, and odor (Bhaisare et al., 2014). Based on the visual evaluation, color is a good indicator of freshness and the wholesomeness of meat, and a major criterion for its procurement by consumers. Barbut (2002) reported that the L* value in meat color indicates the lightness of meat, a* value indicates the redness of meat while the b* value indicates the yellowness color. Meat pH is primarily related to the biochemical state of muscles during slaughter and following rigor mortis development. After slaughtering, the muscle transformation and meat complexion occur, this is due to the development of rigor mortis within the muscle and the pH value changes influencing the quality of meat, which occurs within the first few hours after slaughter (Guerrero-Legarreta, 2010). The pH values of the poultry breast muscle and leg muscle are 5.7 - 5.9 and 6.4 - 6.7 respectively (Bhaisare et al., 2014).
Meat color and pH have consistently been reported to be highly inversely correlated, especially when there is an examination of wide ranges of meat color. The lower muscle pH values are associated with lighter meat while the higher muscle pH is associated with dark, dry, and firm meat (Anadon, 2002). Masenya et al. (2021) reported that whole or crushed sorghum and millet fed to Jumbo quails as a complete replacement of the maize grain, enhanced the lightness of the meat. Lighter meat is known to be fresher, and commands increased procurement from consumers (Mancini and Hunt, 2005). Nevertheless, Masenya et al. (2021) reported a high ultimate pH value of 6.20 in the whole sorghum fed quails, and the lowest post-mortem meat pH value of 5.73 in the crushed millet group. The high ultimate pH value recorded in the whole sorghum fed group was attributed to reduced glycogen levels in the meat post-mortem, owing to the decreased feed intake and growth performance recorded in the group. Although it was reported (Jankowiak et al., 2021) that the change in meat pH cannot influence other meat parameters, a previous report had shown that high pH reduces the water-holding capacity of meat and increases the thawing and cooking losses (Watanabe et al., 2018).
The single most critical meat quality factor related with the consumers’ ultimate rejection or acceptance of a meat product is texture. Tang et al. (2009) reported that factors such as age, species, sex, diet, individual skeletal muscle tissue and breed, have some influence on meat tenderness. The tenderer the meat is, the lower the shear force value (Cavitt et al., 2005). According to Fanatico et al. (2007), slow-growing quail birds have narrower fiber diameters than the fast-growing ones. Birds with slower growth rates have firm texture and a more concentrated flavor, and are notably, more popular for these reasons (Castellini et al., 2008). Complete dietary replacement of maize with sorghum or millet grains had no influence on the shear force attribute of the Jumbo quails’ meat. However, quails fed with the whole sorghum grain replacement diet, had the lowest shear force value compared to meat from quails that had received other dietary treatments (Masenya et al., 2021).
About 88 – 95% of water in the muscle is held intracellularly within spaces between the myosin and actin filaments, and the rest located in between the myofibrils (Hooper et al., 2008). After slaughter, lactic acid is produced owing to low oxygen supply, resulting in meat pH decline, accumulated myosin complex action, decrease in water holding capacity (WHC) and loss of space between the myofibrillar proteins amongst others. Masenya et al. (2021) revealed that replacing maize with sorghum or millet in the diet of Jumbo quails, did not have any effect on the WHC, cooking, drip, and thawing losses of meat from the quail birds. However, it was evident that meat from quails fed with the whole and crushed sorghum grain had the highest numerical values for WHC. The cooking loss of meat was lowest in the crushed and whole millet grain groups, whereas thawing loss was lowest in the crushed sorghum grain-fed group. More studies are required to evaluate the effect of cassava by-products on meat quality traits of quails.
Sustainable quail farming as a valuable source of animal protein must be prioritized to ensure food and nutrition security, human health, economic viability, and environmental stewardship. However, large-scale quail intensification is hindered by the over-reliance on conventional energy sources such as maize. Thus, the use of millet, crushed sorghum and cassava meals as potential alternatives to maize would allow for sustainable quail intensification while reducing feed-food competitions and environmental issues that occurs during maize cultivation. However, it is important to establish the optimum inclusion levels of these alternative sources to ensure safe utilization by the quail. Several pre-treatment methods can be employed where higher inclusion levels are desired, but the cost-effectiveness of the methods must be considered to ensure profitable quail production.
CM conceptualized the review. CM, CO, EA, OR, and FL were equally involved in the write-up, editing, and formatting of the review paper. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abdelqader A., Al-Fataftah A. R., Das G. (2013). Effect of dietary bacillus subtilis supplementation on performance, eggshell quality, intestinal morphology and microflora composition of laying hens in the later phase of production. Anim. Feed Sci. Technol. 179, 103–111. doi: 10.1016/j.anifeedsci.2012.11.003
Abdulla Y. A., Mahmoud K. Z., Nusairat B. M., Qudsieh R. I. (2010). Small intestinal histology, production parameters, and meat quality as influenced by dietary supplementation of garlic (Allium sativum) in broiler chicks. Ital. J. Anim. Sci. 9 (4), 80. doi: 10.4081/ijas.2010.e80
Abu O. A., Judah F. I. (2019). Performance and egg quality traits of laying Japanese quail (Coturnix cortunix japonica) fed cereal grains and cassava flour as energy sources. J. Agric. Prod. Technol. 8, 1–9.
Ainsworth S. J., Stanley R. L., Evans D. J. (2010). Developmental stages of the Japanese quail. J. Anat. 216, pp3–p15. doi: 10.1111/j.1469-7580.2009.01173.x
Ali M. A., Hmar L., Devi Li M. P., Lallianchhunga M. C., Tolenkhomba T. C. (2012). Effect of age on the haematological and biochemical profile of Japanese quails, (Coturnix japonica). Int. Multidiscip. Res. J. 2, 32–35.
Almeida A. Z., Eyng C., Garcia R. G., Nunes R. V., Sangalli G. G., Nunes K. C. (2020). Dried cassava residue in laying quail feeding. Braz. J. Poult. Sci. 22 (1), 1–8. doi: 10.1590/1806-9061-2019-1189
Al-Saffar A. A., Rose S. P. (2002). Ambient temperature and the egg laying characteristics of laying fowl. Worlds. Poult. Sci. J. 58 (3), 317–331. doi: 10.1079/WPS20020025
Altine S., Sabo M. N., Muhammad N., Abubakar A., Saulawa L. A. (2016). Basic nutrient requirements of the domestic quails under tropical conditions: A review. World Sci. News. 49 (2), 223–235.
Amata I. A. (2014). The use of non-conventional feed resources (NCFR) for livestock feeding in the tropics: A review. J. Glob. Biosci. 3 (2), 604–613.
Amelework A. B., Bairu M. W., Maema O., Venter S. L., Laing M. (2021). Adoption and promotion of resilient crops for climate risk mitigation and import substitution: A case analysis of cassava for south African agriculture. Front. Sustain. Food Syst. 5. doi: 10.3389/fsufs.2021.617783
Aminu M., Adamu L., Liya A. B., David S. S., Amos D. M., Oluseun A. I., et al. (2021). Millet and sorghum as possible dietary energy sources in the diet of Japanese quail. J. Adv. Vet. Anim. Res. 5 (4), 87–92. doi: 10.11648/j.ijast.20210504.11
Anadon H. L. S. (2002). Biological, nutritional, and processing factors affecting breast meat quality of broilers (USA: Virginia Polytechnic Institute and State University, Blacksburg, VA), 24061.
Babatunde R. O. O., Olatunbosun O., Adebisi F. A., Akinbola D. D., Iyayi E. A. (2016). Crude protein and energy requirements of Japanese quail (Coturnix coturnix japonica) during rearing period. J. World. Poult. Res. 6 (2), 99–104.
Bagh J., Panigrahi B., Panda N., Pradhan C. R., Mallik B. K., Majhi B., et al. (2016). Body weight, egg production, and egg quality traits of gray, brown, and white varieties of Japanese quail (Coturnix coturnix japonica) in coastal climatic condition of odisha. Vet. World. 9 (8), 832–836. doi: 10.14202/vetworld.2016.832-836
Baguma M., Migabo C., Nzabara F., Sami W. L., Akili C. M., Makali Lwamushi S., et al. (2022). Impact of seasonal variation and processing methods on the cassava-derived dietary cyanide poisoning, nutritional status, and konzo appearance in south-kivu, Eastern D.R. Congo. Processes. 10, 337. doi: 10.3390/pr10020337
Barbut S. (2002). “Measuring sensory and functional properties,” in Poultry products processing. an industry guide (Florida: CRC Press), 467–511.
Bawa G. S. (2010). “Response of Japanese quail chicks (Cortunix cortunix japonica) to various dietary energy levels in a tropical environment,” in Fast tracking animal agriculture in a challenged economy, vol. 2010 . Eds. Babayemi O. J., Abu O. A., Ewuola E. O. (Ibadan: Proceedings of the 34th Annual Conference of the Nigerian Society for Animal Production), 381–383.
Beta T., Corke H. (2004). Effect of ferulic acid and catechin on sorghum and maize starch pasting properties. Cereal Chem. 81, 418–422. doi: 10.1094/CCHEM.2004.81.3.418
Bhaisare D. B., Thyagarajan D., Churchill R. R., Punniamurthy N. (2014). Bacterial pathogens in chicken meat. review. Int. J. Life Sci. Res. 2 (3), 1–7.
Bonos E. M., Christaki E. V., Florou-Paneri P. C. (2010). Performance and carcass characteristics of Japanese quail as affected by sex or mannan oligosaccharides and calcium propionate. S. Afr. J. Anim. Sci. 40 (3), 173–184. doi: 10.4314/sajas.v40i3.2
Brando A., Cuaresma H. L., Ermube R. A., Fabros R. D., Lantano J. I., Berdos M. D., et al. (2021). Laying performance of quail (Coturnix coturnix) fed diets formulated based on crude protein restriction. Philipp. J. Vet. Med. 47 (2), 51–58.
Brian H. C. (2009). Galliformes. Handb. Avian Med., 309–334. doi: 10.1016/B978-0-7020-2874-8.00013-4
Castellini C., Berri C., Le Bihan-Duval E., Martino G. (2008). Qualitative attributes and consumer perception of organic and free-range poultry meat. World Poult. Sci. J. 64 (04), 500–512. doi: 10.1017/S0043933908000172
Cavitt L. C., Meullenet J. F., Gandhapuneni R. K., Youm G. W., Owens C. M. (2005). Rigor development and meat quality of large and small broilers and the use of allo-Kramer shear, needle puncture, and razor blade shear to measure texture. Poult. Sci. 84 (1), 113–118. doi: 10.1093/ps/84.1.113
Chang G. B., Chang H., Liu X. P., Xu W., Wang H. Y., Zhao W. M., et al. (2005). Developmental research on the origin and phylogeny of quails. World. Poult. Sci. J. 61 (1), 105–112. doi: 10.1079/WPS200346
Chauynarong N., Elangovan A. V., Iji P. A. (2009). The potential of cassava products in diets for poultry. World. Poult. Sci. J. 65, 23–36. doi: 10.1017/S0043933909000002
Daramola S. T., Sekoni A. A., Omage J. J., Duru S. (2019). Effects of feeding four varieties of sorghum bicolor as replacement for maize on growth performance of broiler chicks in kaduna state. Niger. J. Anim. Sci. 21 (3), 342–351.
de Oliveira L. D. L., de Oliveira G. T., de Alencar E. R., Queiroz V. A. V., de Alencar Figueiredo L. F. (2022). Physical, chemical, and antioxidant analysis of sorghum grain and flour from five hybrids to determine the drivers of liking of gluten-free sorghum breads. LWT –. Food. Sci. Technol. 153, 112407. doi: 10.1016/j.lwt.2021.112407
Diarra S. S. (2018). Peel meals as feed ingredients in poultry diets: Chemical composition, dietary recommendations and prospects. J. Anim. Physiol. Anim. Nutr. 102, 1284–1295. doi: 10.1111/jpn.12954
Dimkpa S. O. N., Theophilus A. J. (2021). Varietal evaluation of the chemical composition, field performance and yield of some improved cassava (Manihot esculenta crantz) varieties in rivers state, Nigeria. Glob. J. Agric. Res. 9 (2), 27–37.
Duvick D. N., Cassman K. G. (2009). Post-green-revolution trends in yield potential of temperate maize in the north-central united states. Crop Sci. 39 (6), 1622–1630. doi: 10.2135/cropsci1999.3961622x
Egbuonu A. C. C., Ezeanyika L. U. S., Ijeh I. I. (2013). Alterations in the liver histology and markers of metabolic syndrome associated with inflammation and liver damage in l-arginine exposed wister albino rats. Pak. J. Biol. Sci. 16, 469–476. doi: 10.3923/pjbs.2013.469.476
Faitarone A. B. G., Pavan A. C., Mori C., Batista L. S., Oliveira R. P., Garcia E. A., et al. (2005). Economic traits and performance of Italian quails reared at different cage stocking densities. Braz. J. Poult. Sci. 7, 19–22. doi: 10.1590/S1516-635X2005000100003
Fanatico A. C., Pillai P. B., Emmert J. L., Owens C. M. (2007). Meat quality of slow- and fast-growing chicken genotypes fed low-nutrient or standard diets and raised indoors or with outdoor access. Poult. Sci. 86 (10), 2245–2255. doi: 10.1093/ps/86.10.2245
Faquinello P., Murakam A. E., Cella P. S. (2004). High tannin sorghum in diets of Japanese quail (Coturnix coturnix japonica). Braz. J. Poult. Sci. 6, 81–84. doi: 10.1590/S1516-635X2004000200002
Farahat M., Badawi M. A., Hussein A. G. (2020). Effect of replacing dietary corn by sorghum on the growth performance, shank skin pigmentation, carcass traits, caecal microflora and nutrient digestibility of broiler chickens. Int. J. Poult. Sci. 19, 424–431. doi: 10.3923/ijps.2020.424.431
Ferket P. R., Gernat A. G. (2006). Factors that affect feed intake of meat birds: A review. Int. J. Poult. Sci. 5 (10), 905–911. doi: 10.3923/ijps.2006.905.911
Freitas E. R., Raquel D. L., Nascimento A. J. N., Watanabe P. H., Lopes I. R. V. (2014). Complete replacement of corn by white or red sorghum in Japanese quail feeds. Braz. J. Poult. Sci. 16 (3), 333–336. doi: 10.1590/1516-633X1603333-336
González-Alvarado J. M., Jiménez-Moreno, Lázaro R., Mateos G. G. (2007). Effect of type of cereal, heat processing of the cereal, and inclusion of fiber in the diet on productive performance and digestive traits of broilers. Poult. Sci. 86 (8), 1705–1715. doi: 10.1093/ps/86.8.1705
Grote U., Fasse A., Nguyen T. T., Erenstein O. (2021). Food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 4. doi: 10.3389/fsufs.2020.617009
Guerrero-Legarreta I. (2010). Handbook of poultry science and technology (New Jersey, USA: John Wiley and Sons, Inc), 293–327.
Haroon S. (2013). The importance of cereals (Poaceae: Gramineae) nutrition in human health: A review. J. Cereals Oilseeds. 4 (3), 32–35. doi: 10.5897/JCO12.023
Hassan Z. M., Sebola N. A., Mabelebele M. (2021). The nutritional use of millet grain for food and feed: a review. Agric. Food. Secur. 10. doi: 10.1186/s40066-020-00282-6
Hooper S. L., Kevin H. H., Thuma J. B. (2008). Invertebrate muscles: thin and thick filament structure; molecular basis of contraction and its regulation, catch and asynchronous muscle. Prog. Neurobiol. 86 (2), 72–127. doi: 10.1016/j.pneurobio.2008.06.004
Hossain M. D., Saddam M. D., NahidulIslam M. D., Mamunur R., Mostofa M. G., Khan A. (2022). Sorghum: A prospective crop for climatic vulnerability, food and nutritional security. J. Agric. Food. Res. 8, 100300. doi: 10.1016/j.jafr.2022.100300
Hou Y., Wu G. (2018). Nutritionally essential amino acids. Adv. Nutr. 9 (6), 849–851. doi: 10.1093/advances/nmy054
Idris S., Rosnah S., Nor M. Z. M., Mokhtar M. N., Abdul Gani S. S. (2019). Physicochemical composition of different parts of cassava (Manihot esculenta crantz) plant. Food Res. 4 (1), 78–84. doi: 10.26656/fr.2017.4(S1).S33
Inaku E. N., Bawa G. S., Olugbemi T. S., Buba W. (2011). “Nutritive value of yam feel meal in broiler diet,” in Value re-orientation in animal production: A key to national food security and sustainable economy, vol. 2011 . Eds. Adeniji A. A., Olatunji E. A., Gana E. S. (Abuja, Nigeria: Proceedings of 36th Annual Conference of Nigerian Society of Animal Production), 481–483.
Jankowiak H., Cebulska A., Bocian M. (2021). The relationship between acidification (pH) and meat quality traits of polish white breed pigs. Eur. Food Res. Technol. 247, 2813–2820. doi: 10.1007/s00217-021-03837-4
Jatoi A. S., Sahota A. W., Akram M., Javed K., Hussain J., Mehmood S., et al. (2013). Response of different body weights on blood serum chemistry values in four close-bred flocks of adult Japanese quails (Coturnix coturnix japonica). J. Anim. Plant Sci. 23 (1), 35–39.
Kamara M. T., Huiming Z., Kexue Z., Amadou I., Tarawalie F. (2009). Comparative study of chemical composition and physicochemical properties of two varieties of defatted foxtail millet flour grown in China. Am. J. Food. Technol. 4, 255–267. doi: 10.3923/ajft.2009.255.267
Kanyinji F., Moonga M. (2014). Effect of replacing maize meal with rumen filtrate-fermented cassava meal on growth and egg production performance in Japanese quails (Cortunix cortunix). J. Adv. Vet. Anim. Res. 1 (3), 100–106. doi: 10.5455/javar.2014.a19
Karrar I. A., Shammari A. L., Batkowska J., Drabik K., Gryzińska M. M. (2019). Time of sexual maturity and early egg quality of Japanese quails affected by in ovo injection of medicinal plants. Arch. Anim. Breed. 62 (2), 423–430. doi: 10.5194/aab-62-423-2019
Keaokliang O., Kawashima T., Angthong W., Suzuki T., Narmseelee R. (2018). Chemical composition and nutritive values of cassava pulp for cattle. Anim. Sci. J. 89, 1120–1128. doi: 10.1111/asj.13039
Kehinde A. S., Babatunde T. O., Kehinde J. O., Babatunde O. O., Adelakun K. M., Fadimu B. O., et al. (2020). Fibre characterization of cassava peel leaf meal and its utilization by broilers. J. Appl. Sci. Environ. Manage. 24, (9) 1529–1533. doi: 10.4314/jasem.v24i9.7
Khaleel A. G., Shaibu I. M., Abdullahi A. Y., Zango M. H., Madaki S., Ibrahim U., et al. (2021). Growth performance and carcass merit of Japanese quails (Cortunix cortunix) fed with sorghum as an energy source substitute for maize in north-western Nigeria. J. Agrobiotechnol. 12 (1), 23–30. doi: 10.37231/jab.2021.12.1.220
Kingori M. A. (2011). Review of factors that influence egg fertility and hatchability in poultry. Int. J. Poult. Sci. 10, 483–492. doi: 10.3923/ijps.2011.483.492
Klopfenstein T. J., Galen E. E., Berger L. (2013). Maize is a critically important source of food, feed, energy and forage in the USA. Field. Crops Res. 153, 5–11. doi: 10.1016/j.fcr.2012.11.006
Krishnendu R., Hirak B., Sudarshan D., Alok K. H., Kaushik M. (2019). Macronutrients influence yield and oil quality of hybrid maize (Zea mays L.). PLoS One 14 (5), e0216939. doi: 10.1371/journal.pone.0216939
Kwari I. D., Igwebuike J. U., Mohammed I. D., Diarra S. S. (2011). Growth, hematology and serum chemistry of broiler chickens fed raw or processed sorrel (Hibiscus sabdariffa) seed meal in a semi-arid environment. Int. J. Nat. Sci. 2, 22–27.
Lambert G. P. (2009). Stress-induced gastrointestinal barrier dysfunction and its inflammatory effects. J. Anim. Sci. 87, E101–E108.
Laskowski W., Hanna G. W., Krystyna R., Maksymilian C., Justyna Z. (2009). How important are cereals and cereal products in the average polish diet? Nutrients. 11, 679. doi: 10.3390/nu11030679
Latif S., Romuli S., Barati Z., Müller J. (2020). CFD assisted investigation of mechanical juice extraction from cassava leaves and characterization of the products. Food. Sci. Nutr. 8, 3089–3098. doi: 10.1002/fsn3.1517
Laurie C. C., Chasalow S. D., LeDeaux J. R., McCarroll R., Bush D., Hauge B., et al. (2004). The genetic architecture of response to long-term artificial selection for oil concentration in the maize kernel. J. Gene. 168, 2141–2155. doi: 10.1534/genetics.104.029686
Lawler A. (2009). Bridging East and West: Millet on the move. Science. 325 (5943), 942–943. doi: 10.1126/science.325_940
Liu S., Cui S., Ying F., Nasar J., Wang Y., Gao Q. (2021). Simultaneous improvement of protein concentration and amino acid balance in maize grains by coordination application of nitrogen and sulphur. J. Cereal. Sci. 99, 103189. doi: 10.1016/j.jcs.2021.103189
Liu S. Y., Fox G., Khoddami A., Neilson K. A., Truong H. H., Moss A. F. (2015). Grain sorghum: A conundrum for chicken-meat production. Agriculture. 5, 1224–1251. doi: 10.3390/agriculture5041224
Malik A. A., Ijaiya A. T., Aremu A., Akinwale M. O. (2018a). Growth response, carcass characteristics and egg laying performance of Japanese quail (Coturnix coturnix japonica) fed diets containing varying levels of fermented cassava (Manihot esculenta) peel meal. Niger. J. Anim Prod. 45 (2), 224–2231. doi: 10.51791/njap.v45i2.492
Malik A. A., Ijaiya A. T., Tsado D. N., Ntaudom N. (2018b). Growth performance and egg production of Japanese quail (Coturnix coturnix japonica) fed diets containing graded levels of sun-dried cassava (Manihot esculenta) peel meal. Niger. J. Anim. Sci. 20 (2), 232–240.
Manano J., Ogwok P., Bazirake G. W. B. (2018). Chemical composition of major cassava varieties in Uganda, targeted for industrialisation. J. Food. Res. 7 (1), 1–9. doi: 10.5539/jfr.v7n1p1
Mancini R. A., Hunt M. C. (2005). Current research in meat color. Meat Sci. 71 (1), 100–121. doi: 10.1016/j.meatsci.2005.03.003
Marareni M., Mnisi C. M. (2020). Growth performance, serum biochemistry and meat quality traits of jumbo quails fed with mopane worm (Imbrasia belina) meal-containing diets. Vet. Anim. Sci. 10, 100141. doi: 10.1016/j.vas.2020.100141
Mardoia M. (2016). Effects of dietary polyphenol-rich grape by-products on growth performance, some physical parameters meat and meat products quality in chickens. Madrid Spain. pp, 1–244.
Masenya T. I., Mlambo V., Mnisi C. M. (2021). Complete replacement of maize grain with sorghum and pearl millet grains in jumbo quail diets: Feed intake, physiological parameters, and meat quality traits. PLoS One 16 (3), e0249371. doi: 10.1371/journal.pone.0249371
Mbanjo E. G. N., Rabbi I. Y., Ferguson M. E., Kayondo S. I., Eng N. H., Tripathi L., et al. (2021). Technological innovations for improving cassava production in Sub-Saharan Africa. Front. Genet. 11. doi: 10.3389/fgene.2020.623736
Mizutani M. (2003). The Japanese quail, laboratory animal research station (Yamanashi, Japan: Nippon Institution for Biological Science), 408–410.
Mnisi C. M., Marareni M., Manyeula F., Madibana M. J. (2021). A way forward for the south African quail sector as a potential contributor to food and nutrition security following the aftermath of COVID-19: a review. Agric. Food Secur 10 (2021), 48. doi: 10.1186/s40066-021-00331-8
Morgan N. K., Choct M. (2016). Cassava: Nutrient composition and nutritive value in poultry diets. Anim. Nutr. 2 (4), 253–261. doi: 10.1016/j.aninu.2016.08.010
Mottet A., Tempio G. (2017). Global poultry production: current state and future outlook and challenges. Worlds. Poult. Sci. J. 73 (2), 1–12. doi: 10.1017/S0043933917000071
Mundia C. W., Silvia S., Kofi A., Wang G. (2019). A regional comparison of factors affecting global sorghum production: The case of north America, Asia and africa’s sahel. Sustainability. 11, 2135. doi: 10.3390/su11072135
Musa U., Haruna E. S., Lombin L. H. (2008). Quails’ production in the tropics (Vom, Nigeria: National veterinary Research Institute press), 158.
Nambisan B. (2010). Strategies for elimination of cyanogens from cassava for reducing toxicity and improving food safety. food and chemical toxicology: an international journal published for the. Br. Ind. Biol. Res. Assoc. 49 (3), 690–693. doi: 10.1016/j.fct.2010.10.035
Nebiyu A., Getachew E. (2011). Soaking and drying of cassava roots reduced cyanogenic potential of three cassava varieties at jima, southwest Ethiopia. Afri. J. Biotechnol. 10 (62), 13465–13469. doi: 10.5897/AJB10.2636
Nwokedi M. N., Sekoni A. A., Alphonsus C. (2010). “Evaluation of optimal energy and protein requirement of Japanese quails (Coturnix coturnix japonica) for growth and carcass characteristics,” in Diversifying Nigerian economy animal production option, vol. 2009 . Eds. Ifut O. J., Inyang U. A., Akpan I. P., Ebeso I. E. (Uyo: Proceedings of the 15th Annual Conference of the Animal Science Association of Nigeria), 503–507.
Oboh G., Afolabi A. A., Oshodi A. A. (2002). Nutrient and anti-nutrient contents of aspergillus niger-fermented cassava products (flour and gari). J. Food Compos. Anal. 15 (5), 617–622. doi: 10.1006/jfca.2002.1065
Odo B. I., Nnadi A. E. (2013). Growth response of quails (Coturnix coturnix japonica) to varying levels of cassava tuber meal (Manihot esculenta) as replacement for maize (Zea mays). Rev. Cie. UDO Agrí. 13 (1), 146–149.
Ojediran T. K., Abioye I., Ajayi A., Emiola A. (2019). Replacement value of cassava vinasse meal for maize on growth performance, haematological parameters and organoleptic properties of Japanese quails (Cortunix cortunix japonica). Acta Fytotechn. Zootechn. 22 (1), 7–12. doi: 10.15414/afz.2019.22.01.7-12
Okarter N., Liu H. R. (2010). Health benefits of whole grain phytochemicals. Crit. Rev. Food. Sci. Nutr. 50, 193–208. doi: 10.1080/10408390802248734
Okogbenin E., Tim L. S., Ferguson M., Mutegi R., Hernan C., Olasanmi B., et al. (2013). Phenotypic approaches to drought in cassava: review. Front. Physiol. 4. doi: 10.3389/fphys.2013.00093
Oladunjoye I. O., Ojebiyi O. O. (2010). Effect of feeding processed cassava (Manihot esculenta crantz) peel meal-based diets on the performance characteristics, egg quality and blood profile of laying chicken. Agric. Trop. Subtrop. 43 (2), 121–123.
Oni A. O., Onwuka C. F. I., Arigbede O. M., Anele U. Y., Oduguwa O. O., Onifade O. S., et al. (2011). Chemical composition and nutritive value of four varieties of cassava leaves grown in south-Western Nigeria. J. Anim. Physiol. Anim. Nutr. 95, 583–590. doi: 10.1111/j.1439-0396.2010.01086.x
Onyimonyi A. E., Ugwu S. O. C. (2007). Bio-economic indices of broiler chicks fed varying ratios of cassava peel to bovine blood. Int. J. Poult. Sci. 3 (7), 463–467. doi: 10.3923/ijps.2007.318.321
Orr A., Mwema C., Gierend A., Nedumaran S. (2016). “Sorghum and millets in Eastern and southern Africa,” in Facts, trends and outlook. working paper series no. 62. ICRISAT research program, markets, institutions and policies. patancheru 502 324, telangana (India: International Crops Research Institute for the Semi-Arid Tropics), 76 pp.
Owheruo J. O., Ifesan B. O. T., Kolawole A. O. (2019). Physicochemical properties of malted finger millet (Eleusine coracana) and pearl millet (Pennisetum glaucum). Food. Sci. Nutr. 7, 476–482. doi: 10.1002/fsn3.816
Oyeagu C. E., Ani A. O., Egbu C. F., Udeh F. U., Omumuabuike J. N. (2015). Comparative performance of nera black and shaver brown hen fed self-compounded and commercial layers’ diet. Asian. J. Sci. Technol. 6 (1), 940–946.
Oyelami B. A., Abu O. A. (2017). Growth, blood indices and carcass characteristics of japanese quail (Coturnix coturnix japonica) fed cassava grit as replacement for maize with or without âglucanase. Niger. J.Anim. Prod. 44 (1), 284–294.
Peralee K. (2022) Quail vs chicken eggs: Are quail eggs better for you. Available at: https://a-z-animals.com/blog/quail-vs-chicken-eggs-are-quail-eggs-better-for-you/ (Accessed 5 August 2022).
Pius L. O., Strausz P., Kusza S. (2021). Overview of poultry management as a key factor for solving food and nutritional security with a special focus on chicken breeding in East African countries. Biology. 10, 810. doi: 10.3390/biology10080810
Prabakaran R. (2003). Good practices in planning and management of integrated commercial poultry production in south Asia food and agricultural organization of the united nations Vol. 159 (Rome: FAO Animal Production and Health Division), 71–82.
Prasad P. V. V., Staggenborg S. (2010). Growth and production of sorghum and millets (U.K: Oxford), 1–27.
Quaresma M. A. G., Antunes I. C., Gil Ferreira B., Parada A., Elias A., Barros M., et al. (2022). The composition of the lipid, protein and mineral fractions of quail breast meat obtained from wild and farmed specimens of common quail (Coturnix coturnix) and farmed Japanese quail (Coturnix japonica domestica). Poult. Sci. 101 (1), 101505. doi: 10.1016/j.psj.2021.101505
Rabie M. H., Hayam M. A., Abo E. M. (2015). Growth performance of Japanese quail as affected by dietary protein level and enzyme supplementation. Asian. J. Anim. Vet. Adv. 10, 74–85. doi: 10.3923/ajava.2015.74.85
Rafh M. T. (2018). Effect of using high protein level in the first week of age on some productive traits of quail. J. Agric. Environ. Vet. Sci. 2 (4), 64–72. doi: 10.26389/AJSRP.R250618
Randall M., Bolla G. (2008). Raising Japanese quail (2nd ed.) Vol. 602 (New South Wales, Australia: PrimeFacts), 1–5.
Redoy M. R. A., Shuvo A. A. S., Al-Mamun M. (2017). A review on present status, problems and prospects of quail farming in Bangladesh. Bangladesh J. Anim. Sci. 46 (2), 109–120. doi: 10.3329/bjas.v46i2.34439
Santos T. C., Murakami A. E., Oliveira C. A. L., Giraldelli N. (2013). Sperm-egg interaction and fertility of Japanese breeder quails from 10 to 61 weeks. Poult. Sci. 92 (1), 205–210. doi: 10.3382/ps.2012-02536
Sari M., Tilki M., Saatci M. (2016). Genetic parameters of egg quality traits in long-term pedigree recorded Japanese quail. Poult. Sci. 95, 1743–1749. doi: 10.10.3382/ps/pew118
Savanna S. (2020) Quail eggs: Nutrition, benefits and precautions. Available at: https://www.healthline.com/nutrition/quail-eggs-benefits (Accessed 5 August 2022).
Shibi K. T., Amutha R., Purushothaman M. R., Jagatheesan P. N., Ezhilvalavan S., Jayalalitha V. (2019). Energy and protein requirements during various stages of production in Japanese quails. Int. J. Sci. Envir. Technol. 8 (4), 790–794.
Shiferaw B., Prasanna B. M., Hellin J., Banziger M. (2011). Crops that feed the world 6. past successes and future challenges to the role played by maize in global food security. Food Security. 3, 307. doi: 10.1007/s12571-011-0140-5
Shrivastav A. K. (2000). Quail nutrition under Indian conditions. India. J. Poult. Sci. 67 (3), 239–241.
Silva J. P. A., Lana S. R. V., Lana G. R. Q., Silva L. C. L., Torres E. C., Ferreira T. S. (2017). Cassava foliage in quail feeding. Acta Veterinária Brasilica. 11, 150–156. doi: 10.21708/avb.2017.11.0.6921
Sonny W., Edi P. S., Achmad R., Made A. (2019). Cellulose nanofibers preparation from cassava peels via mechanical disruption. Fibers. 7, 44. doi: 10.3390/fib7050044
Stefan S., Youngmi K., Eduardo X., Nathan M., Michael L. (2009). Ethanol production from maize. Chapter 23, 347–364. doi: 10.1007/978-3-540-68922-5_23
Tang H., Gong Y. Z., Wu C. X., Jiang J., Wang Y., Li K. (2009). Variation of meat quality traits among five genotypes of chicken. Poult. Sci. 88, 2212–2218. doi: 10.3382/ps.2008-00036
Taylor J. R. N., Emmambux M. N. (2010). Developments in our understanding of sorghum polysaccharides and their health benefits. Cereal Chem. 87, 263–271. doi: 10.1094/CCHEM-87-4-0263
Tenebaum D. J. (2008). Food vs. fuel: diversion of crops could cause more hunger. Environ. Health Perspectives. 116, 254–257. doi: 10.1289/ehp.116-a254
Thea K., Megan O. M., Lesley L. D. (2018). Nanotechnology in the food sector and potential applications for the poultry industry. Trends. Food. Sci. Technol. 72, 62–73. doi: 10.1016/j.tifs.2017.11.015
Tiwari U. P., Jha R. (2016). Nutrient profile and digestibility of tubers and agro-industrial coproducts determined using an in vitro model of swine. Anim. Nutr. 2, 357–360. doi: 10.1016/j.aninu.2016.07.004
Truong H. H., Liu S. Y., Selle P. H. (2015). Starch utilization in chicken-meat production: The foremost influential factors. Anim. Prod. Sci. 56 (5), 797–814. doi: 10.1071/AN15056
Tunsaringkan T., Tungjaroenchai W., Siriwong W. (2013). Nutrient benefits of quails eggs. Int. J. Sci. Res. Pub. 3, 1–8.
Vali N. (2008). The Japanese quail: a review. Int. J. Poult. Sci. 7, 925–931. doi: 10.3923/ijps.2008.925.931
Vanessa J. (2020) Eat your way to good health with quail eggs. Available at: https://www.medindia.net/patients/lifestyleandwellness/eat-your-way-to-good-health-with-quail-eggs.htm (Accessed 5 August 2022).
Watanabe G., Michiyo M., Ikuyo N., Keisuke S. (2018). Relationship between water-holding capacity and intramuscular fat content in Japanese commercial pork loin. Asian-Australas. J. Anim. Sci. 31 (6), 914–918. doi: 10.5713/ajas.17.0640
Wenhold F., Faber M. (2009). Water in nutritional health of individuals and households: an overview. Water SA 35 (1), 61–71.
Wong J. T., de Bruyn J., Bagnol B., Grieve H., Li M., Pym R., et al. (2017). Small-scale poultry and food security in resource-poor settings: A review. Glob. Food Sec. 15, 43–52. doi: 10.1016/j.gfs.2017.04.003
Xu Y., Stark C. R., Ferket P. R., Williams C. M., Brake J. (2015). Effects of feed form and dietary coarse ground corn on broiler live performance, body weight uniformity, relative gizzard weight, excreta nitrogen, and particle size preference behaviors. Poult. Sci. 94 (7), 1549–1556. doi: 10.3382/ps/pev074
Yafetto L. (2018). Protein enrichment of cassava pulp by solid-state fermentation using Aspergillus niger. Stud. Fungi 3, 2. doi: 10.5943/sif/3/1/2
Yanza Y. R., Fitri A., Suwignyo B., Elfahmi Hidayatik N., Kumalasari N. R., Irawan A., et al. (2021). The utilisation of tannin extract as a dietary additive in ruminant nutrition: A meta-analysis. Animals 11, 3317. doi: 10.3390/ani11113317
Zaheer K. (2015). An updated review on chicken eggs: Production, consumption, management aspects and nutritional benefits to human health. Food. Nutr. Sci. 6 (3), 1208–1220. doi: 10.4236/fns.2015.613127
Keywords: energy value, food security, intensive production, quail, sustainability
Citation: Mnisi CM, Oyeagu CE, Akuru EA, Ruzvidzo O and Lewu FB (2023) Sorghum, millet and cassava as alternative dietary energy sources for sustainable quail production – A review. Front. Anim. Sci. 4:1066388. doi: 10.3389/fanim.2023.1066388
Received: 10 October 2022; Accepted: 09 February 2023;
Published: 23 February 2023.
Edited by:
Catarina Stefanello, Federal University of Santa Maria, BrazilReviewed by:
Mürsel Özdoğan, Aydın Adnan Menderes University, TürkiyeCopyright © 2023 Mnisi, Oyeagu, Akuru, Ruzvidzo and Lewu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Caven Mguvane Mnisi, bW5pc2llY21AZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.