 Elijah G. Kiarie
Elijah G. Kiarie Samantha Steelman2
Samantha Steelman2 Marco Martinez
Marco Martinez
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Anim. Sci. , 06 September 2022
Sec. Animal Nutrition
Volume 3 - 2022 | https://doi.org/10.3389/fanim.2022.875095
This article is part of the Research Topic Nutrient Regulation of Feed Intake, Digestion, Metabolism, Behavior, and Cognition View all 10 articles
The provision of adequate and balanced nutrients is critical for efficient and profitable animal protein production. However, non-nutritive components in feedstuffs can elicit responses that can negatively impact nutrient utilization efficiency. For example, dietary β-mannans are recognizable by cell surface mannose receptors are pivotal for diverse cellular functions. This review will evaluate the physiological implications of dietary native β-mannans, the utility of supplemental feed β-mannanase in hydrolyzing β-mannans, and subsequent metabolic responses. Dietary native β-mannans have been implicated in inadvertent stimulation of immune response through a phenomenon called the feed-induced immune response (FIIR), that has been associated with intestinal inflammation and depression in animal performance. Supplemental β-mannanase blunted the FIIR by hydrolyzing native β-mannans to smaller fragments with a reduced ability to stimulate the innate immune system as indicated by the modulation of oxidative stress, mucosal permeability, and blood concentration of acute phase proteins and immunoglobulins in broilers and piglet models. Moreover, β-mannanase hydrolysis of native β-mannans to mannooligosaccharides (MOS) impacted gastrointestinal microbial ecology. Indeed, β-mannanase-derived MOS reduced the concentration of pathogenic bacteria such as Escherichia coli and Salmonella and increased the production of short-chain fatty acids in gastrointestinal tracts of various animal models. Consequently, by hydrolyzing native β-mannans, supplemental β-mannanase may have nutritional, metabolic, and microbial ecology benefits. In summary, integrating multi-functional feed additives such as β-mannanase into feeding programs for monogastric animals will be critical for efficient and sustainable animal protein production in the context of evolving challenges such as the mandated elimination of use of antibiotics for growth promotion.
Monogastric farm animals are ranked highly in terms of efficiency of conversion of feed to food products. From production cost perspectives, feed accounts for more than 60% of the variable inputs cost, with dietary energy and protein (amino acids) accounting for a significant portion. Yet, these animals excrete significant amounts of undigested nutrients (Schedle, 2016). Most of the variation in dry matter digestibility in feedstuffs is related to the presence of anti-nutritional factors (ANFs) and indigestible complexes (NRC, 1994; NRC, 2012). The implications of insufficient and variable nutrient digestibility include increased feed costs due to poor feed efficiency, increased risk of enteric pathogens, and increased excretion of nutrients into the environment among others (Kiarie et al., 2013; Kiarie et al., 2016). Moreover, indigestible complexes stimulate growth of visceral organs and, as such, increase partitioning of more energy and nutrients for maintenance at the expense of growth, lactation, and reproduction (Cant et al., 1996). Thus, the basis for the utility of exogenous feed enzymes (FE) is to address the inherent digestive insufficiency of monogastric animals (Bedford, 1996; Bedford and Schulze, 1998; Kiarie, 2020). Therefore, the concepts of commercial and practical application of FE technology in animal nutrition are to target ANF in feedstuffs and to complement endogenous enzymes in animals with immature and/or compromised gut function. Indeed, the utility of FE in enhancing the nutritive value of feedstuffs for monogastric farm animals has received extensive critical appraisals in the last three decades (Bedford and Schulze, 1998; Adeola and Cowieson, 2011; Slominski, 2011; Ravindran, 2012; Dersjant-Li et al., 2015). Arguably, features and functionality of FE are designed and set by the source organism and target substrate. However, the utility in animal nutrition is dependent on the mechanism of action (Kiarie et al., 2013). For example, feed β-mannanase has been demonstrated to benefit growth performance and digestibility in pigs and poultry, as summarized in numerous reviews (Shastak et al., 2015; Torres-Pitarch et al., 2019; Saeed et al., 2019; Kipper et al., 2020; Kiarie et al., 2021). However, native/intact feed β-mannans, the target substrate for β-mannanase, not only have anti-nutrient properties but have also been linked to stimulating immune responses with implications on post-absorptive nutrients utilization. The present review will evaluate physiological implications of dietary native β-mannans, utility of supplemental feed β-mannanase in hydrolyzing β-mannans, and subsequent impact on immunometabolism and gut health.
Plant feedstuffs used in monogastric animal diets are rich in non-starch polysaccharides, with hemicellulose and cellulose as dominating fractions (Bach Knudsen, 1997). Protein feedstuffs are particularly rich in β-mannans, a type of hemicellulose rich in D-mannose units linked in β-(1-4) glycosidic bonds (Chen et al., 2018a). We recently reported the structure, abundance, and nutritional implications of β-mannans in common feedstuffs (Kiarie et al., 2021). In general, the major β-mannans in feedstuffs are glucomannan and galactomannan, and the concentration can be as high as 30% DM in some feedstuffs (Kiarie et al., 2021). Figure 1 illustrates the framework for gastrointestinal and ecology responses to intact feed β-mannans. Unlike other dietary non-starch polysaccharides, β-mannans are similar to carbohydrate moieties present in the cell walls of microorganisms that are recognizable by cell membrane mannose receptors (MR) (Arsenault et al., 2017; Mathiesen et al., 2019). As C-type lectin, MR are expressed by macrophages and dendritic cells and are pivotal for antigen detection and adhesion, pathogen infections, and signal transduction (Taylor et al., 2005; Arsenault et al., 2017). In this context, β-mannans are recognizable as pathogen-associated molecular patterns (PAMP) via several cell MR lining the gastrointestinal tract. It has been postulated that β-mannans bind to intestinal cells or are absorbed into the bloodstream to exert local and systematic immune responses (Zhang and Tizard, 1996; Duncan et al., 2002; Arsenault and Kogut, 2015; Arsenault et al., 2017). Consequently, feed β-mannans are associated with inadvertent immune response stimulation through a phenomenon coined feed-induced immune response (FIIR) leading to intestinal inflammation, poor nutrient utilization, and diminished animal productivity (Daskiran et al., 2004; Gabler and Spurlock, 2008; Arsenault et al., 2017). For example, aloe vera β-mannans are chemically similar to soybean meal β-mannans and were demonstrated to increase synthesis of nitric oxide by activating MR in macrophages (Karaca et al., 1995; Ramamoorthy et al., 1996). As such, the host immune system recognizes β-mannans for invading antigens/pathogens and mounting cellular immune response (Wu et al., 2005; Arsenault et al., 2017). Intact feed β-mannans can also impede normal digestion and absorption processes of nutrients with implications on gastrointestinal microbial ecology. Luminal compounds (dietary and/or endogenous) in the gastrointestinal tract serves as a substrate for the microbiota (Kiarie et al., 2013). Soluble β-mannans increase digesta viscosity of intestinal contents, effectively reducing digestion and absorption of nutrients (Rainbird et al., 1984; Lee et al., 2003; Blackburn and Johnson, 2007; Rainbird et al., 2007). The undigested nutrients increase the risk of proliferation of enteric pathogens such as Clostridium perfringens, enterotoxigenic Escherichia coli (Kiarie et al., 2013). Moreover, increased flow of ileal undigestible protein in the hindgut can result in proteolytic fermentation in the large intestine of pigs and the cecum of poultry that can negatively affect their performance and health (Kiarie et al., 2013).
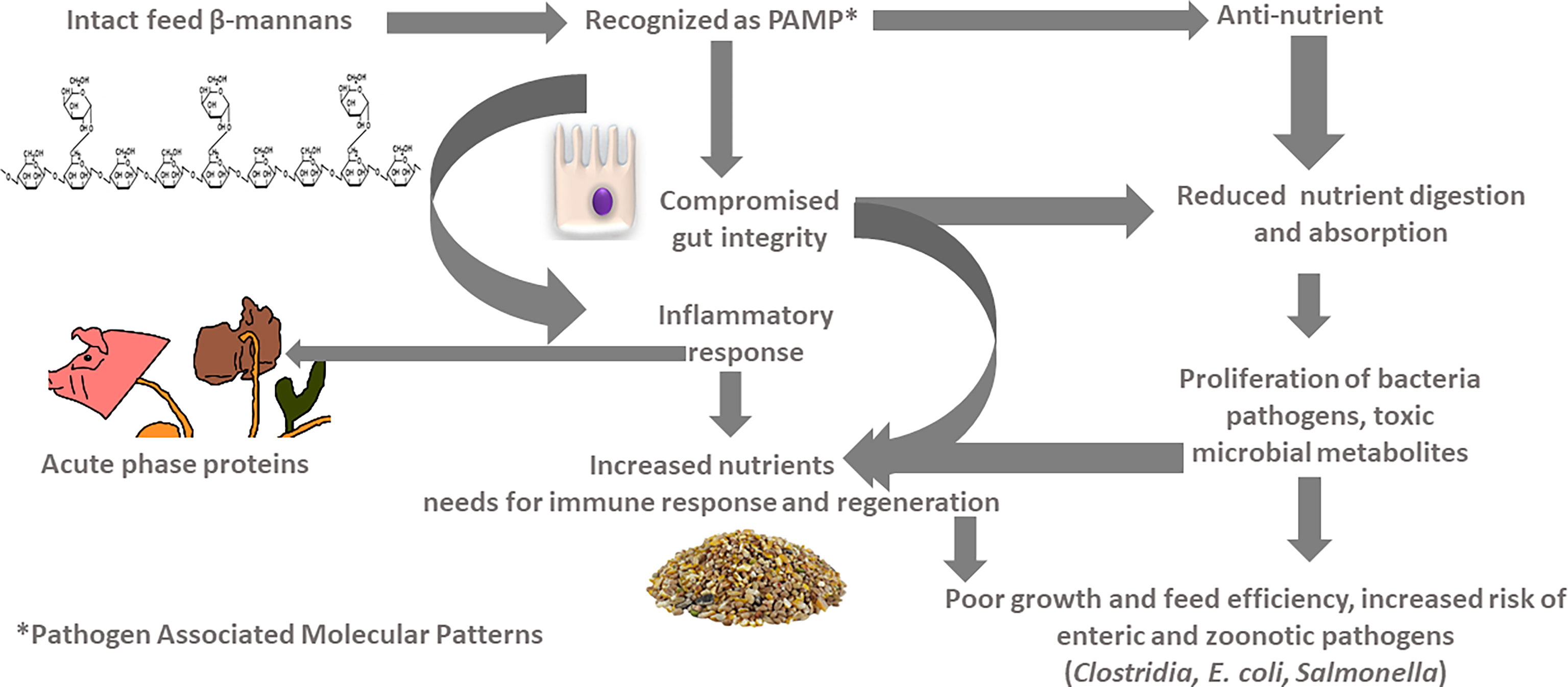
Figure 1 Framework for implications of feed β-mannans on gastrointestinal physiology and ecology.
The supplementation of ß-mannanase has been shown to improve energy and nutrients utilization in broilers (Li et al., 2010), laying hens (Wu et al., 2005), and pigs (Yoon et al., 2010; Kim et al., 2013; Lv et al., 2013). However, the main effect of supplementation of ß-mannanase in monogastric diets has been associated with blunting ß-mannan-induced inflammatory responses and/or releasing prebiotic-like hydrolysis products in the intestinal tract. Various studies have characterized immune system modulation of supplemental ß-mannanase (Klasing, 2007; Arsenault and Kogut, 2015; Arsenault et al., 2017). For example, a positive correlation between the concentration of blood acute phase proteins (APP) and dietary β-mannans has been demonstrated (Arsenault et al., 2017; Huntley et al., 2018). Chicken-specific immunometabolism kinome array was used to characterize jejunal cellular signaling dynamics in broiler chicken fed diets supplemented with β-mannanase (Arsenault et al., 2017). The authors demonstrated that ß-mannan-induced gut inflammation was reduced by supplemental β-mannanase, confirming anti-inflammatory and energy sparing effects of β-mannanase. Specifically, metabolic processes such as insulin, adipocytokine, and mTOR pathway and gut integrity indicators such as tight junctions were modified by supplemental β-mannanase (Arsenault et al., 2017). These observations corroborated earlier studies that showed that mice fed hydrolyzed guar β-mannans upregulated expression of nine genes related to host defense functions (Yasukawa et al., 2012). Moreover, ß-1-4-mannoligosaccharides (MOS) from coffee β-mannans were shown to reduce interleukin-10 production (Ozaki et al., 2007). The native locust bean galactomannan stimulated cells to produce inflammatory cytokine tumor necrosis factor alpha (TNF-α) and β-hexosaminidase secretion (Chen et al., 2018b). These effects were reversed if the native locust bean galactomannan was hydrolyzed with β-mannanase (Chen et al., 2018b). The peculiarity is that β-mannanase hydrolysis of intact native β-mannans results in MOS fragments that can no longer be recognized by toll-like receptors and consequently conserve valuable energy for growth and performance (Anderson and Hsiao, 2006; Arsenault and Kogut, 2015; Arsenault et al., 2017).
The gut immune system has the challenge of responding to pathogenic and non-pathogenic antigens while remaining relatively unresponsive to commensal microflora. Dietary fiber components such as β-mannans are known to influence gastrointestinal microbiota ecology. Thus, hydrolyzing fiber fraction with fiber-degrading enzymes such as β-mannanase can potentially release fiber fragments that can modulate microbial ecology (Kiarie et al., 2013). In vitro studies demonstrated that ß-mannanase released short ß-1,4-mannoligosaccharides (ß-1-4-MOS) from the ß-mannans backbone (McCleary, 1979; Okubo et al., 1994; Mudgil et al., 2018; Mary et al., 2019; Yin et al., 2020; Suryawanshi and Kango, 2021). Further in vitro studies demonstrated that shorter ß-1-4-MOS promoted lactic acid production and inhibited enteropathogenic bacteria such as E. coli and Salmonella in monoculture and co-culture fermentations (Okubo et al., 1994; Mudgil et al., 2018; Mary et al., 2019). These studies suggested that ß-mannans hydrolysis products were preferentially utilized by microorganisms associated with good gut health such as Bifidobacteria and Lactobacilli.
The cell surface molecule structure of most enteric pathogens is critical in the colonization and infection through the host docking sites lining the intestinal mucosa (Giron et al., 2002). For example, D-mannose sugar has been characterized for its competitive inhibition of bacteria adhesion to cellular membranes due to its functional similarity to bacteria type 1 fimbriae (Mirelman et al., 1980; Corrigan et al., 2015). Incubating common feedstuffs with xylanase, cellulases, and ß-mannanase released a range of sugars (Kiarie, 2008). Interestingly, mannose was one of the major sugars released from soybean meal and wheat middlings and was shown to protect against enterotoxigenic E. coli infection in a piglet intestinal model (Kiarie, 2008; Kiarie et al., 2010). Although in vivo monitoring of the production of mannose has not been reported, they could be linked to changes in gut microbiome observed in animals fed diets supplemented with ß-mannanase as presented and discussed later.
Thus, the impact of ß-mannanase supplementation on energy utilization was partly associated with reduced inflammatory response linked to lighter lymphoid organs (thymus, spleen, and bursa) (Li et al., 2010). The addition of ß-mannanase in corn and soybean meal (SBM)-based diets or the removal of SBM reduced the plasma concentration of APP, indicating a lesser immune response (Dale et al., 2008). Studies in broiler chickens and turkeys showed significant improvement in feed conversion concomitant with the reduction in acute phase protein due supplemental β-mannanase in corn–SBM diets (Anderson and Hsiao, 2006). Upon hydrolysis by ß-mannanase, ß-mannans are no longer recognized by the immune system, and more energy is left available to the bird for growth, rather than being consumed by FIIR. Enteric pathogens are of great concern to the poultry industry due to the impact on animal and human health through contamination of poultry products (Williams, 2005; Timbermont et al., 2011). The bird husbandry status is an important consideration in assessing the magnitude of response to ß-mannanase supplementation. For example, ß-mannanase effects might be more pronounced in birds exposed to poor sanitary and health conditions. There are many research investigations that have used major poultry enteric pathogens such as Eimeria and C. perfringens models for evaluating the efficacy of feed additive or dietary strategy (Cooper and Songer, 2010; Peek and Landman, 2011). These in vivo challenge models permit the characterization of the responses of dietary intervention in the context of the presence of enteric pathogens (Kiarie et al., 2019).
The immunological benefits of β-mannanase supplementation in broiler chicken diets were linked to reduction in intestinal damage in necrotic enteritis challenge model (Jackson et al., 2003). ß-Mannanase supplementation reduced the challenge severity as further indicated by improved growth and lower intestinal lesions compared to control (Jackson et al., 2003). Moreover, the performance of birds fed ß-mannanase was commensurate to that of birds fed bacitracin methylene disalicilate and salinomycin. The serum concentration of APP was higher in birds challenged with Eimeria, and the addition of β-mannanase reduced serum APP (Anderson and Hsiao, 2006). Another study showed that supplemental β-mannanase increased serum IgM concentration in broiler chickens linked to the proliferation of T lymphocytes (Zou et al., 2006). ß-Mannanase improvement of growth performance was not seen in a study that used overdose of Eimeria vaccine (Scapini et al., 2019). However, birds fed ß-mannanase had shallow jejunum crypt depth and as such improved their villus to crypt ratio, indicating reduced rate of cell proliferation and thus better intestinal health. Adding ß-mannanase to a corn and SBM fed to broiler chickens resulted in decreased goblet cell number and epithelial thickness and increased villus height to crypt depth ratio in different sections of the small intestine (Mehri et al., 2010). In another study, ß-mannanase fed broiler chickens had improved duodenal villi height and width but increased crypt depth, reduced jejunal and ileal crypt depth, and increased ileal villi length (Karimi and Zhandi, 2015). This corroborated data showing that feeding broiler chickens diets supplemented with β-mannanase-hydrolyzed copra meal improved growth and feed efficiency linked to improved intestinal histomorphology (Ibuki et al., 2014). These changes in the morphological structure of the small intestine have been linked to a reduction in immune challenge. Overall, these studies demonstrated evidence for the efficacy of β-mannanase in immunomodulation.
Fiber-degrading enzymes modulation of gastrointestinal microbial ecology is related to increased digestibility of nutrients, thus reducing the host and microbiota competition for the same and release of prebiotic fiber hydrolysis products (Kiarie et al., 2013). The flow of undigested feed materials in the ceca, particularly those rich in protein fractions, is associated with production of toxic metabolites such as thiols, amines, ammonia, and indoles (Nyachoti et al., 2006). However, something very critical to ceca fermentation of protein-rich materials is the elevation of digesta pH that promotes proliferation of pathogens such as C. perfringens (Williams, 2005; Timbermont et al., 2011). There have been considerable efforts on pre-treating ß-1-4-mannan-rich feedstuffs such as palm kernel meal, guar meal, and copra meal to derive ß-1-4-MOS for feed application. Broiler chickens fed ß-1-4-MOS showed reduced susceptibility to Salmonella enterica infection through higher IgA and inhibition of Salmonella colonization (Agunos et al., 2007). ß-1-4-MOS derived from the hydrolyses of ß-mannan-rich guar, palm kernel, and copra meal inhibited intestinal colonization of Salmonella in broiler and layer chicks (Morikoshi and yokomizo, 2004). Further work demonstrated reduction in Salmonella enteritidis colonization in hens fed corn–SBM plus guar meal diet supplemented with ß-mannanase (Gutierrez et al., 2008). Broiler chickens fed ß-mannanase-pre-treated copra meal exhibited lower E. coli and Salmonella population in excreta relative to broilers fed native copra meal (Khanongnuch et al., 2006).
It has been demonstrated that coccidiosis decreased small intestine pH but increased ceca pH in broiler chickens (Ruff et al., 1974). Thus, it is interesting that fiber-degrading enzymes have been shown to increase concentration of short chain fatty acids such as lactic, acetic, and butyric acids with concomitant reduction in pH in in vitro and in vivo models (Kiarie et al., 2014; Rho et al., 2020). Apajalahti et al. (2004) associated feed-enzyme-induced reduction in coccidiosis intestinal lesion scores in broiler chickens to production of short-chain fatty acids. Acetic acid was demonstrated to have similar effects to Amprolium (a coccidiostat) in reducing negative effects of coccidiosis in broiler chicken growth (Abbas et al., 2011). Such observations suggested that feed enzymes can influence survivability of Eimeria or limit intestinal damage. An attempt was made to investigate the impact of β-mannanase on the gut health and function of broiler chickens challenged with excess coccidiosis vaccine (Bortoluzzi et al., 2019). The data demonstrated that β-mannanase supplementation impacted intestinal microbiota. Specifically, β-mannanase increased abundance of gut-health-associated microbiota (e.g., Lactobacillus, Ruminococcaceae, and Akkermansia) and reduced bacteria associated with poor feed efficiency (Bortoluzzi et al., 2019). Further research showed that broiler chickens fed 0.035% of MOS extracted from copra meal increased lactic acid bacteria (Putri et al., 2017).
Foot pad dermatitis (FPD) is a health, quality, and welfare issue in broiler chicken production, with significant economic effects as chicken feet are a delicacy in many cultures (Chen et al., 2016). The condition is indicated with dermatitis lesions on feed plantar surfaces (Chen et al., 2016). Dietary and non-dietary factors that stimulated higher water consumption and excretion have been linked to incidences and severity of FPD (Swiatkiewicz et al., 2017). Viscous non-starch polysaccharides have been linked to FPD through wet and sticky droppings (Kaukonen et al., 2016). As such supplementation with fiber-degrading enzymes in poultry diets based on viscous feedstuffs such as wheat, rye, and barley has been shown to improve litter quality (Bedford and Schulze, 1998; Slominski, 2011; Swiatkiewicz et al., 2017). It is therefore relevant that supplemental ß-mannanase decreased water consumption per unit of feed consumed in broiler chickens (Daskiran et al., 2004). In another study, ß-mannanase reduced digesta viscosity in broiler chickens, but there was no impact on the incidence and severity of FPD or ammonia volatilization rate (Cengız et al., 2012). However, a commercial trial in two farms in Europe demonstrated that β-mannanase reduced intestinal and excreta moisture content (Grieve et al., 2016). This was associated with flocks fed β-mannanase showing cleaner vents and footpads. Further investigations on the role of β-mannanase on FPD incidences in commercial broiler production applied Health Tracking System (HTSi), a surveillance system for capturing flock-level health and performance information (Kasab-Bachi et al., 2017). Analyses of 44 commercial farms in Europe showed that β-mannanase reduced the incidence and severity of pododermatitis (unpublished, Elanco). Incidences of pendulous crop in turkey production is a serious welfare issue and is indicated by stagnant liquid or semi-liquid content leading to temporary or permanent distension (Wheeler et al., 1960). In severe cases, fatalities may result from the rupture of crops or from starvation (Wheeler et al., 1960). Incidences of pendulous crop have been associated with many factors including genetics, hatching conditions, diet, and management (Wheeler et al., 1960; Almeida et al., 2018). However, Opoku et al. (2015) did not find differences in incidences of pendulous crop and water intake in turkey hens fed diets of diverse composition (wheat, corn, pork meal, SBM, and wheat DDGs) without or with ß-mannanase.
There are many bacterial enteric diseases that afflict pigs that can be modulated with nutrition (Pluske et al., 2002). Moreover, weaning is very stressful due to convergence of multiple factors ranging from management to immature and unstable gut physiology (Pluske, 2016). Therefore, using β-mannanase to prevent the unnecessary energy-demanding pro-inflammatory stimulation of innate immune system by intact β-mannans would be beneficial to weaned pigs. The incubation of corn DDGS with β-mannanase increased the production of butyrate in a simulated pig gastrointestinal tract and increased jejunal concentration of tight junction proteins and tended to reduce mucosal malondialdehyde in piglets fed corn–SBM diet with 15% corn DDGS (Tiwari et al., 2018). However, the authors did not observe β-mannanase benefits on digesta viscosity, pH, small intestine histomorphology (villi height and crypt depth), and plasma or intestinal concentration of tumor necrosis factor-α (Tiwari et al., 2018). β-Mannanase supplementation did not influence diarrhea and fecal concentration of lactobacilli bacteria in piglets but reduced fecal E. coli shedding (Balamuralikrishnan et al., 2018). Similarly, β-mannanase reduced fecal shedding of coliforms in growing pigs fed corn–SBM but had no effects on fecal lactobacilli populations (Upadhaya et al., 2016). These data are indicative that dietary β-mannanase may exert effects at gut level by modulating oxidative stress, mucosal permeability, and microbial ecology in younger pigs presumably mediated by β-mannan hydrolysis products. Serum haptoglobin and interleukin-1-alpha concentrations were decreased in nursery pigs fed corn–SBM diet with 10% soy hulls with β-mannanase (Huntley et al., 2018). However, the maintenance energy requirement was similar between control and β-mannanase fed pigs (Huntley et al., 2018). In another study, the concentrations of acute phase protein, superoxide dismutase, and glutathione peroxidase in nursery pigs fed corn–SBM diet was not impacted by β-mannanase supplementation (Jang et al., 2020a). Increasing dietary β-mannans concentration to (2.8%) by adding 10% copra meal in a corn–SBM diet (0.4% β-mannans) had no effect on the concentration of serum acute phase proteins (C-reactive protein and haptoglobin) (Huntley et al., 2020). Likewise, supplementation of β-mannanase had no impact on the serum concentration of acute phase proteins. β-Mannanase supplementation in a corn–SBM diet did not impact nursery pig serum proinflammatory cytokines, haptoglobin, or mannose binding lectin concentrations. However, serum haptoglobin and IL-1α concentrations were reduced in pigs fed diets supplemented with β-mannanase (Huntley et al., 2018). These studies indicated that β-mannanase can reduce acute phase proteins in pigs with the potential of preventing an energy-demanding immune response in weaned pigs. However, more investigations are required to unravel circumstances under which benefits are more pronounced.
It is weight that increasing dietary fiber levels reduces dressing weight because pigs increase gut volume and weight to sustain critical nutrient intake (Nyachoti et al., 2004; Agyekum et al., 2012). Heavier portal vein drained viscera are associated with greater maintenance energy and nutrient requirements, subsequently reducing allocation for production (Just et al., 1983). It is therefore plausible that supplemental fiber-degrading enzymes will have benefits on aspects of carcass yield and pork quality. Thus, Pettey et al. (2002) demonstrated that pigs fed diets supplemented with β-mannanase exhibited greater fat-free lean index than pigs fed diets containing no enzyme. However, other studies reported no benefits of β-mannanase supplementation in growing and finishing pig diets and had no effects on pork quality indices (dressing percentage, backfat, lean percentage, drip loss, WHC, pH, loin area, and marbling) (Wang et al., 2009; Yoon et al., 2010; Kim et al., 2017; Jang et al., 2020b). Overall, it does not seem plausible that supplemental β-mannanase has an impact on carcass quality in pigs.
Feed cost is and will remain a decisive factor for profitable and sustainable animal protein production. Exogenous FE are widely accepted for managing feed costs through flexible utility of opportunity feed ingredients and abatement of the environmental impact of animal production. However, emerging issues such as the restriction on the use antibiotic use for growth promotion is stimulating new concepts in FE application. The relation between feed enzymes and the host responses is linked to the effects of target substrates on physiology and the modification of these effects by FE to the extent of substrates degradation or modification in the gastrointestinal tract. Thus, the degradation of native dietary β-mannans by supplemental β-mannanase blunt feed induced immune response with implications on improved feed efficiency. The release of mannans hydrolysis products enhanced gut health, as indicated by the reduction in pathogenic bacteria and increased production of short-chain fatty acids. The recognition that FE such as β-mannanase modulate inflammation and gut microbiota will stimulate development of FE capable of benefitting animal health under specific production conditions. However, more research is needed to further elucidate and refine host–microbial ecology–feed β-mannans interactions. For example, there are many published studies that investigated the effects of yeast cell mannans hydrolysis products on poultry and swine performance and health. However, unlike plant ß-1-4-mannans, yeast mannans are characterized with α-1-6-linked mannose chain and side chains of varied features. It is plausible that there could be efficacy variations between plant and yeast hydrolysis products that warrant further studies. Furthermore, differences may be apparent between feedstuffs; for example, ß-1-4-MOS produced from SBM may be quite different from those produced from copra and guar meals.
The original contributions synthesized in the systematic review are included in the article material, further inquiries can be directed to the corresponding author.
EK developed the concepts and had the overall editorial responsibility whereases MM and SS provided critical review of the manuscript. All authors contributed to the article and approved the submitted version.
Natural Sciences and Engineering Research Council of Canada-Discovery program provided base funding for EK research program
Contributions of the current and former students and associates of EK Monogastric Nutrition Research Laboratory at University of Guelph are appreciated.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbas R. Z., Munawar S. H., Manzoor Z., Iqbal Z., Khan M. N., Saleemi M. K., et al. (2011). Anticoccidial effects of acetic acid on performance and pathogenic parameters in broiler chickens challenged with eimeria tenella. Pesqui. Vet. Bras. 31, 99–103. doi: 10.1590/S0100-736X2011000200001
Adeola O., Cowieson A. J. (2011). BOARD-INVITED REVIEW: Opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. J. Anim. Sci. 89 (10), 3189–3218. doi: 10.2527/jas.2010-3715
Agunos A., Ibuki M., Yokomizo F., Mine Y. (2007). Effect of dietary β1–4 mannobiose in the prevention of salmonella enteritidis infection in broilers. Brit Poult. Sci. 48 (3), 331–341. doi: 10.1080/00071660701370442
Agyekum A. K., Slominski B. A., Nyachoti C. M. (2012). Organ weight, intestinal morphology, and fasting whole-body oxygen consumption in growing pigs fed diets containing distillers dried grains with solubles alone or in combination with a multienzyme supplement. J. Anim. Sci. 90 (9), 3032–3040. doi: 10.2527/jas.2011-4380
Almeida E. A., Silva F. H. A., Crowe T. G., Macari M., Furlan R. L. (2018). Influence of rearing temperature and feed format in the development of the pendulous crop in broilers. Poult. Sci. 97 (10), 3556–3563. doi: 10.3382/ps/pey221
Anderson D. M., Hsiao H. Y. (2006). “Effect of ß-mannanase (Hemicell® feed enzyme) on acute phase protein levels in chickens and turkeys,” in Poultry Science Association 95th Annual Meeting No. 85 (Champaign, IL, USA: Poultry Science Association). 130.
Apajalahti J., Kettunen A., Graham H. (2004). Characteristics of the gastrointestinal microbial communities, with special reference to the chicken. World Poult. Sci. J. 60 (2), 223–232. doi: 10.1079/Wps200415
Arsenault R. J., Kogut M. H. (2015). Immunometabolism and the kinome peptide array: A new perspective and tool for the study of gut health. Front. Vet. Sci. 2 (44). doi: 10.3389/fvets.2015.00044
Arsenault R. J., Lee J. T., Latham R., Carter B., Kogut M. H. (2017). Changes in immune and metabolic gut response in broilers fed β-mannanase in β-mannan-containing diets. Poult. Sci. 96 (12), 4307–4316. doi: 10.3382/ps/pex246
Bach Knudsen K. E. (1997). Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed Sci. Technol. 67 (4), 319–338. doi: 10.1016/S0377-8401(97)00009-6
Balamuralikrishnan B., Lee J. H., Kim I. H. (2018). Effects of dietary β-mannanase supplementation of soybean meal on the performance of weanling pigs. Anim. Nutr. Feed Technol. 18 (1), 13–23. doi: 10.5958/0974-181X.2018.00002.1
Bedford M. R. (1996). The effect of enzymes on digestion. J. Appl. Poult. Res. 5 (4), 370–378. doi: 10.1093/japr/5.4.370
Blackburn N. A., Johnson I. T. (2007). The effect of guar gum on the viscosity of the gastrointestinal contents and on glucose uptake from the perfused jejunum in the rat. Brit J Nutr 46 (2), 239–246. doi: 10.1079/BJN19810029
Bedford M. R., Schulze H. (1998). Exogenous enzymes for pigs and poultry. Nutr. Res. Rev. 11 (1), 91–114. doi: 10.1079/NRR19980007
Bortoluzzi C., Scapini L. B., Ribeiro M. V., Pivetta M. R., Buzim R., Fernandes J. I. M. (2019). Effects of β-mannanase supplementation on the intestinal microbiota composition of broiler chickens challenged with a coccidiosis vaccine. Livest Sci. 228, 187–194. doi: 10.1016/j.livsci.2019.09.001
Cant J. P., McBride B. W., Croom W. J. (1996). The regulation of intestinal metabolism and its impact on whole animal energetics. J. Anim. Sci. 74 (10), 2541–2553. doi: 10.2527/1996.74102541x
Cengız Ö., Hess J. B., Bilgili S. F. (2012). Feed enzyme supplementation does not ameliorate foot pad dermatitis in broiler chickens fed on a corn-soyabean diet. Brit Poult. Sci. 53 (4), 401–407. doi: 10.1080/00071668.2012.711467
Chen W.-L., Chen H.-L., Guo G.-w., Huang Y.-C., Chen C.-Y., Tsai Y., et al. (2018b). Locust bean gum galactomannan hydrolyzed by thermostable β-d-mannanase may reduce the secretion of pro-inflammatory factors and the release of granule constituents. Int. J. Biol. Macromol. 114, 181–186. doi: 10.1016/j.ijbiomac.2018.03.097
Chen J., Robb C. S., Unfried F., Kappelmann L., Markert S., Song T., et al. (2018a). Alpha- and beta-mannan utilization by marine bacteroidetes. Environ. Microbiol. 20 (11), 4127–4140. doi: 10.1111/1462-2920.14414
Chen J., Tellez G., Escobar J. (2016). Identification of biomarkers for footpad dermatitis development and wound healing. Front. Cell. Infect. Microbiol. 6. doi: 10.3389/fcimb.2016.00026
Cooper K. K., Songer J. G. (2010). Virulence of clostridium perfringens in an experimental model of poultry necrotic enteritis. Vet. Microbiol. 142 (3), 323–328. doi: 10.1016/j.vetmic.2009.09.065
Corrigan A., de Leeuw M., Penaud-Frezet S., Dimova D., Murphy R. A. (2015). Phylogenetic and functional alterations in bacterial community compositions in broiler ceca as a result of mannan oligosaccharide supplementation. Appl. Environ. Microbiol. 81 (10), 3460–3470. doi: 10.1128/AEM.04194-14
Dale N. M., Anderson D. M., Hsiao H. Y. (2008). “Identification of an inflammatory compound for chicks in soybean meal,” in Poultry Science Association Annual Meeting No. 87, Poultry Science, Niagara Falls, Ontario, Canada (Champaign, IL, USA: Poultry Science Association). 153.
Daskiran M., Teeter R. G., Fodge D., Hsiao H. Y. (2004). An evaluation of endo-β-D-mannanase (Hemicell) effects on broiler performance and energy use in diets varying in β-mannan content1. Poult. Sci. 83 (4), 662–668. doi: 10.1093/ps/83.4.662
Dersjant-Li Y., Awati A., Schulze H., Partridge G. (2015). Phytase in non-ruminant animal nutrition: a critical review on phytase activities in the gastrointestinal tract and influencing factors. J. Sci. Food Agric. 95 (5), 878–896. doi: 10.1002/jsfa.6998
Duncan C. J. G., Pugh N., Pasco D. S., Ross S. A. (2002). Isolation of a galactomannan that enhances macrophage activation from the edible fungus morchella esculenta. J. Agric. Food Chem. 50 (20), 5683–5685. doi: 10.1021/jf020267c
Gabler N. K., Spurlock M. E. (2008). Integrating the immune system with the regulation of growth and efficiency1, 2. J. Anim. Sci. 86 (suppl_14), E64–E74. doi: 10.2527/jas.2007-0466
Giron J. A., Torres A. G., Freer E., Kaper J. B. (2002). The flagella of enteropathogenic escherichia coli mediate adherence to epithelial cells. Mol. Microbiol. 44 (2), 361–379. doi: 10.1046/j.1365-2958.2002.02899.x
Grieve A. M., Cervantes-Pahm S. K., Martinez M. A. (2016). “The impact of β-mannanase enzyme on the intestinal health of poultry under commercial conditions,” in 27th Annual Australian Poultry Science Symposium, Sydney, New South Wales (Camden, Australia: Poultry Research Foundation, University of Sydney). 52–55.
Gutierrez O., Zhang C., Caldwell D. J., Carey J. B., Cartwright A. L., Bailey C. A. (2008). Guar meal diets as an alternative approach to inducing molt and improving salmonella enteritidis resistance in late-phase laying hens. Poult. Sci. 87 (3), 536–540. doi: 10.3382/ps.2007-00337
Huntley N. F., Gould S. A., Patience J. F., Plaizier J. (2020). Evaluation of the effect of β-mannanase supplementation and mannans on nursery pig growth performance and serum acute-phase protein concentrations. Can. J. Anim. Sci. 100 (1), 111–118. doi: 10.1139/cjas-2018-0248
Huntley N. F., Nyachoti C. M., Patience J. F. (2018). Lipopolysaccharide immune stimulation but not β-mannanase supplementation affects maintenance energy requirements in young weaned pigs. J. Anim. Sci. Biotechno. 9 (1), 47. doi: 10.1186/s40104-018-0264-y
Ibuki M., Fukui K., Yamauchi K. (2014). Effect of dietary mannanase-hydrolysed copra meal on growth performance and intestinal histology in broiler chickens. J. Anim. Physiol. Anim. Nutr. 98 (4), 636–642. doi: 10.1111/jpn.12105
Jackson M. E., Anderson D. M., Hsiao H. Y., Mathis G. F., Fodge D. W. (2003). Beneficial effect of β-mannanase feed enzyme on performance of chicks challenged with eimeria sp. and clostridium perfringens. Avian Dis. 47 (3), 759–763. doi: 10.1637/7024
Jang J.-C., Kim K. H., Jang Y. D., Kim Y. Y. (2020a). Effects of dietary β-mannanase supplementation on growth performance, apparent total tract digestibility, intestinal integrity, and immune responses in weaning pigs. Animals 10 (4), 703. doi: 10.3390/ani10040703
Jang J. C., Kim K. H., Kim D. H., Jang S. K., Hong J. S., Heo P. S., et al. (2020b). Effects of increasing levels of palm kernel meal containing β-mannanase to growing-finishing pig diets on growth performance, nutrient digestibility, and pork quality. Livest Sci. 238, 104041. doi: 10.1016/j.livsci.2020.104041
Just A., Fernandez J. A., Jorgensen H., Fernández J., Jørgensen H. (1983). The net energy value of diets for growth in pigs in relation to the fermentative processes in the digestive-tract and the site of absorption of the nutrients. Livest Prod. Sci. 10 (2), 171–186. doi: 10.1016/0301-6226(83)90033-7
Karaca K., Sharma J. M., Nordgren R. (1995). Nitric-oxide production by chicken macrophages activated by acemannan, a complex carbohydrate extracted from aloe-vera. Int. J. Immunopharmacol. 17 (3), 183–188. doi: 10.1016/0192-0561(94)00102-t
Karimi K., Zhandi M. (2015). The effect of β-mannanase and β-glucanase on small intestine morphology in male broilers fed diets containing various levels of metabolizable energy. J. Appl. Anim. Res. 43 (3), 324–329. doi: 10.1080/09712119.2014.978770
Kasab-Bachi H., Arruda A. G., Roberts T. E., Wilson J. B. (2017). The use of large databases to inform the development of an intestinal scoring system for the poultry industry. Prev. Vet. Med. 146, 130–135. doi: 10.1016/j.prevetmed.2017.07.012
Kaukonen E., Norring M., Valros A. (2016). Effect of litter quality on foot pad dermatitis, hock burns and breast blisters in broiler breeders during the production period. Avian Pathol. 45 (6), 667–673. doi: 10.1080/03079457.2016.1197377
Khanongnuch C., Sa-nguansook C., Lumyong S. (2006). Nutritive quality of β-mannanase treated copra meal in broiler diets and effectiveness on some fecal bacteria. Int. J. Poult. Sci. 5, 1087–1091. doi: 10.3923/ijps.2006.1087.1091
Kiarie E. (2008). “Dietary means for enhanced gut health and function in piglets,” in An evaluation of carbohydrase enzymes targeting non-starch polysaccharides (Winnipeg: University of Manitoba).
Kiarie E., Slominski B. A., Nyachoti C. M. (2010). Effect of products derived from hydrolysis of wheat and flaxseed non-starch polysaccharides by carbohydrase enzymes on net absorption in enterotoxigenic Escherichia coli (K88) challenged piglet jejunal segments. Anim. Sci. J. 81: 63–71.
Kiarie E. (2020). “Inherent digestive tract insufficiency in monogastric animals: culpability of the gut microbiome and dietary approaches for optimizing intestinal health,” in Animal Nutrition Conference of Canada (Ottawa, Canada: Animal Nutrition Association of Canada). 85–99.
Kiarie E. G., Leung H., Akbari Moghaddam Kakhki R., Patterson R., Barta J. R. (2019). Utility of feed enzymes and yeast derivatives in ameliorating deleterious effects of coccidiosis on intestinal health and function in broiler chickens. Front. Vet. Sci. 6 (473). doi: 10.3389/fvets.2019.00473
Kiarie E., Romero L. F., Nyachoti C. M. (2013). The role of added feed enzymes in promoting gut health in swine and poultry. Nutr. Res. Rev. 26 (1), 71–88. doi: 10.1017/S0954422413000048
Kiarie E., Romero L. F., Ravindran V. (2014). Growth performance, nutrient utilization, and digesta characteristics in broiler chickens fed corn or wheat diets without or with supplemental xylanase. Poult. Sci. 93 (5), 1186–1196. doi: 10.3382/ps.2013-03715
Kiarie E. G., Steelman S., Martinez M., Livingston K. (2021). Significance of single β-mannanase supplementation on performance and energy utilization in broiler chickens, laying hens, turkeys, sows, and nursery-finish pigs: a meta-analysis and systematic review. Trans. Anim. Sci. 5 (4). doi: 10.1093/tas/txab160
Kiarie E., Walsh M. C., Nyachoti C. M. (2016). Performance, digestive function, and mucosal responses to selected feed additives for pigs. J. Anim. Sci. 94 (supplement3), 169–180. doi: 10.2527/jas.2015-9835
Kim J. S., Ingale S. L., Lee S. H., Kim K. H., Kim J. S., Lee J. H., et al. (2013). Effects of energy levels of diet and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites in growing pigs. Anim. Feed Sci. Technol. 186 (1), 64–70. doi: 10.1016/j.anifeedsci.2013.08.008
Kim H. J., Nam S. O., Jeong J. H., Fang L. H., Yoo H. B., Yoo S. H., et al. (2017). Various levels of copra meal supplementation with β-mannanase on growth performance, blood profile, nutrient digestibility, pork quality and economical analysis in growing-finishing pigs. J. Anim. Sci. Technol. 59, 19–19. doi: 10.1186/s40781-017-0144-6
Kipper M., Andretta I., Quadros V. R., Schroeder B., Pires P. G. S., Franceschina C. S., et al. (2020). Performance responses of broilers and pigs fed diets with β-mannanase. Braz. J. Anim. Sci. Zootecnia. 49, e20180177. doi: 10.37496/rbz4920180177
Klasing K. C. (2007). Nutrition and the immune system. Brit Poult. Sci. 48 (5), 525–537. doi: 10.1080/00071660701671336
Lee J. T., Bailey C. A., Cartwright A. L. (2003). β-Mannanase ameliorates viscosity-associated depression of growth in broiler chickens fed guar germ and hull fractions. Poultry Sci 82 (21), 1925–1931. doi: 10.1093/ps/82.12.1925
Li Y., Chen X., Chen Y., Li Z., Cao Y. (2010). Effects of β-mannanase expressed by pichia pastoris in corn–soybean meal diets on broiler performance, nutrient digestibility, energy utilization and immunoglobulin levels. Anim. Feed Sci. Technol. 159 (1), 59–67. doi: 10.1016/j.anifeedsci.2010.05.001
Lv J. N., Chen Y. Q., Guo X. J., Piao X. S., Cao Y. H., Dong B. (2013). Effects of supplementation of β-mannanase in corn-soybean meal diets on performance and nutrient digestibility in growing pigs. Asian-Australasian J. Anim. Sci. 26 (4), 579–587. doi: 10.5713/ajas.2012.12612
Mary P. R., Prashanth K. V. H., Vasu P., Kapoor M. (2019). Structural diversity and prebiotic potential of short chain β-manno-oligosaccharides generated from guar gum by endo-β-mannanase (ManB-1601). Carbohydr. Res. 486, 107822. doi: 10.1016/j.carres.2019.107822
Mathiesen R., Eld H. M. S., Sørensen J., Fuglsang E., Lund L. D., Taverniti V., et al. (2019). Mannan enhances IL-12 production by increasing bacterial uptake and endosomal degradation in l. acidophilus and s. aureus stimulated dendritic cells. Front. Immunol. 10. doi: 10.3389/fimmu.2019.02646
McCleary B. V. (1979). Modes of action of β-mannanase enzymes of diverse origin on legume seed galactomannans. Phytochemistry 18 (5), 757–763. doi: 10.1016/0031-9422(79)80009-6
Mehri M., Adibmoradi M., Samie A., Shivazad M. (2010). Effects of -mannanase on broiler performance, gut morphology and immune system. Afr. J. Biotechnol. 9 (37), 6221–6228. doi: 10.5897/AJB09.1371
Mirelman D., Altmann G., Eshdat Y. (1980). Screening of bacterial isolates for mannose-specific lectin activity by agglutination of yeasts. J. Clin. Microbiol. 11 (4), 328–331. doi: 10.1128/jcm.11.4.328-331.1980
Morikoshi T., yokomizo F. (2004). ß-1,4-mannobiose-containing composition (Geneva, Switzerland: The World Intellectual Property Organisation Patent).
Mudgil D., Barak S., Patel A., Shah N. (2018). Partially hydrolyzed guar gum as a potential prebiotic source. Int. J. Biol. Macromol. 112, 207–210. doi: 10.1016/j.ijbiomac.2018.01.164
NRC (1994). Nutrient requirements of poultry. 9th revision ed (Washington, D.C: National Academy of Sciences Press).
NRC (2012). Nutrient requirements of swine. 11th revision ed (Washington, D.C: National Academy of Sciences Press).
Nyachoti C. M., Omogbenigun F. O., Rademacher M., Blank G. (2006). Performance responses and indicators of gastrointestinal health in early-weaned pigs fed low-protein amino acid-supplemented diets. J. Anim. Sci. 84 (1), 125–134. doi: 10.2527/2006.841125x
Nyachoti C. M., Zijlstra R. T., de Lange C. F. M., Patience J. F. (2004). Voluntary feed intake in growing-finishing pigs: A review of the main determining factors and potential approaches for accurate predictions. Can. J. Anim. Sci. 84 (4), 549–566. doi: 10.4141/A04-001
Okubo T., Ishihara N., Takahashi H., Fujisawa T., Kim M., Yamamoto T., et al. (1994). Effects of partially hydrolyzed guar gum intake on human intestinal microflora and its metabolism. Biosci. Biotechnol. Biochem. 58 (8), 1364–1369. doi: 10.1271/bbb.58.1364
Opoku E. Y., Classen H. L., Scott T. A. (2015). Evaluation of inclusion level of wheat distillers dried grains with solubles with and without protease or β-mannanase on performance and water intake of turkey hens. Poultry Sci 94(7): 1600–1610. doi: 10.3382/ps/pev088
Ozaki K., Fujii S., Hayashi M. (2007). Effect of dietary mannooligosaccharides on the immune system of ovalbumin-sensitized mice. J. Health Sci. 53 (6), 766–770. doi: 10.1248/jhs.53.766
Peek H. W., Landman W. J. M. (2011). Coccidiosis in poultry: anticoccidial products, vaccines and other prevention strategies. Vet. Q. 31 (3), 143–161. doi: 10.1080/01652176.2011.605247
Pettey L. A., Carter S. D., Senne B. W., Shriver J. A. (2002). Effects of beta-mannanase addition to corn-soybean meal diets on growth performance, carcass traits, and nutrient digestibility of weanling and growing-finishing pigs2. Journal of Animal Science 80(4): 1012–1019. doi: 10.2527/2002.8041012x
Pluske J. R. (2016). Invited review: Aspects of gastrointestinal tract growth and maturation in the pre- and postweaning period of pigs. J. Anim. Sci. 94, 399–411. doi: 10.2527/jas2015-9767
Pluske J. R., Pethick D. W., Hopwood D. E., Hampson D. J. (2002). Nutritional influences on some major enteric bacterial diseases of pig. Nutr. Res. Rev. 15 (2), 333–371. doi: 10.1079/NRR200242
Putri A. N. S., Sumiati S., Meryandini A. (2017). Effect of dietary mannan-oligosaccharides from copra meal on intestinal microbes and blood profile of broiler chickens. J. Indonesian Trop. Anim. Agric. 42 (2), 109–119. doi: 10.14710/jitaa.42.2.109-119
Rainbird A. L., Low A. G., Zebrowska T. (1984). Effect of guar gum on glucose and water absorption from isolated loops of jejunum in conscious growing pigs. Br J Nutr 52(3): 489–498. doi: 10.1079/bjn19840116
Rainbird A. L., Low A. G., Zebrowska T. (2007). Effect of guar gum on glucose and water absorption from isolated loops of jejunum in conscious growing pigs. Br J Nutr 52(3): 489–498. doi: 10.1079/bjn19840116
Ramamoorthy L., Kemp M. C., Tizard I. R. (1996). Acemannan, a beta-(1,4)-acetylated mannan, induces nitric oxide production in macrophage cell line RAW 264.7. Mol. Pharmacol. 50 (4), 878–884.
Ravindran V. (2012). Advances and future directions in poultry nutrition: An overview. Korean J. Poult. Sci. 39, 53–62. doi: 10.5536/KJPS.2012.39.1.053
Rho Y., Patterson R., Joye I., Martinez M., Squires J. E., Kiarie E. G. (2020). Fiber degrading enzyme increased monosaccharides release and fermentation in corn distillers dried grains with solubles and wheat middlings steeped without or with protease. Tran. Anim. Sci. 4, 1–16. doi: 10.1093/tas/txaa153
Ruff M. D., Johnson J. K., Dykstra D. D., Reid W. M. (1974). Effects of eimeria acervulina on intestinal pH in conventional and gnotobiotic chickens. Avian Dis. 18 (1), 96–104. doi: 10.2307/1589247
Saeed M., Aya_an T., Alagawany M., El-Hack M., Abdel-Latif M., Patra A. (2019). The role of ß-mannanase (Hemicell) in improving poultry productivity, health and environment. Braz. J. Poult. Sci. 21. doi: 10.1590/1806-9061-2019-1001
Scapini L. B., de Cristo A. B., Schmidt J. M., Buzim R., Nogueira L. K., Palma S. C., et al. (2019). Effect of β-mannanase supplementation in conventional diets on the performance, immune competence and intestinal quality of broilers challenged with eimeria sp. J. Appl. Poult. Res. 28 (4), 1048–1057. doi: 10.3382/japr/pfz066
Schedle K. (2016). Sustainable pig and poultry nutrition by improvement of nutrient utilisation – a review. Die Bodenkultur. 67 (1), 45–60. doi: 10.1515/boku-2016-0005
Shastak Y., Ader P., Feuerstein D., Ruehle R., Matuschek M. (2015). ß-mannan and mannanase in poultry nutrition. World's Poult. Sci. J. 71 (1), 161–174. doi: 10.1017/S0043933915000136
Slominski B. A. (2011). Recent advances in research on enzymes for poultry diets. Poult. Sci. 90 (9), 2013–2023. doi: 10.3382/ps.2011-01372
Suryawanshi R. K., Kango N. (2021). Production of mannooligosaccharides from various mannans and evaluation of their prebiotic potential. Food Chem. 334, 127428. doi: 10.1016/j.foodchem.2020.127428
Swiatkiewicz S., Arczewska-Wlosek A., Jozefiak D. (2017). The nutrition of poultry as a factor affecting litter quality and foot pad dermatitis – an updated review. J. Anim. Physiol. Anim. Nutr. 101 (5), e14–e20. doi: 10.1111/jpn.12630
Taylor P. R., Gordon S., Martinez-Pomares L. (2005). The mannose receptor: linking homeostasis and immunity through sugar recognition. Trends Immunol. 26 (2), 104–110. doi: 10.1016/j.it.2004.12.001
Timbermont L., Haesebrouck F., Ducatelle R., Van Immerseel F. (2011). Necrotic enteritis in broilers: an updated review on the pathogenesis. Avian Pathol. 40 (4), 341–347. doi: 10.1080/03079457.2011.590967
Tiwari U. P., Chen H., Kim S. W., Jha R. (2018). Supplemental effect of xylanase and mannanase on nutrient digestibility and gut health of nursery pigs studied using both in vivo and in vitro models. Anim. Feed Sci. Technol. 245, 77–90. doi: 10.1016/j.anifeedsci.2018.07.002
Torres-Pitarch A., Manzanilla E. G., Gardiner G. E., O’Doherty J. V., Lawlor P. G. (2019). Systematic review and meta-analysis of the effect of feed enzymes on growth and nutrient digestibility in grow-finisher pigs: Effect of enzyme type and cereal source. Anim. Feed Sci. Technol. 251, 153–165. doi: 10.1016/j.anifeedsci.2018.12.007
Upadhaya S. D., Park J. W., Lee J. H., Kim I. H. (2016). Efficacy of β-mannanase supplementation to corn–soya bean meal-based diets on growth performance, nutrient digestibility, blood urea nitrogen, faecal coliform and lactic acid bacteria and faecal noxious gas emission in growing pigs. Arch. Anim. Nutr. 70 (1), 33–43. doi: 10.1080/1745039X.2015.1117697
Wang J. P., Hong S. M., Yan L., Yoo J. S., Lee J. H., Jang H. D., et al. (2009). Effects of single or carbohydrases cocktail in low-nutrient-density diets on growth performance, nutrient digestibility, blood characteristics, and carcass traits in growing–finishing pigs. Livest Sci. 126 (1), 215–220. doi: 10.1016/j.livsci.2009.07.003
Wheeler H. O., Reid B. L., Ferguson T. M., Couch J. R., Rigdon R. H. (1960). Differences in susceptibility of broad breasted bronze and Beltsville small white turkeys to dietary-induced pendulous crop. Poult. Sci. 39 (2), 263–267. doi: 10.3382/ps.0390263
Williams R. B. (2005). Intercurrent coccidiosis and necrotic enteritis of chickens: rational, integrated disease management by maintenance of gut integrity. Avian Pathol. 34 (3), 159–180. doi: 10.1080/03079450500112195
Wu G., Bryant M. M., Voitle R. A., Roland D. A. (2005). Effects of β-mannanase in corn-soy diets on commercial leghorns in second-cycle hens. Poult. Sci. 84 (6), 894–897. doi: 10.1093/ps/84.6.894
Yasukawa Z., Naito Y., Takagi T., Mizushima K., Tokunaga M., Ishihara N., et al. (2012). Partially hydrolyzed guar gum affects the expression of genes involved in host defense functions and cholesterol absorption in colonic mucosa of db/db male mice. J. Clin. Biochem. Nutr. 51 (1), 33–38. doi: 10.3164/jcbn.11-104
Yin J.-Y., Ma L.-Y., Xie M.-Y., Nie S.-P., Wu J.-Y. (2020). Molecular properties and gut health benefits of enzyme-hydrolyzed konjac glucomannans. Carbohydr. Polymers 237, 116117. doi: 10.1016/j.carbpol.2020.116117
Yoon S. Y., Yang Y. X., Shinde P. L., Choi J. Y., Kim J. S., Kim Y. W., et al. (2010). Effects of mannanase and distillers dried grain with solubles on growth performance, nutrient digestibility, and carcass characteristics of grower-finisher pigs1. J. Anim. Sci. 88 (1), 181–191. doi: 10.2527/jas.2008-1741
Zhang L., Tizard I. R. (1996). Activation of a mouse macrophage cell line by acemannan: The major carbohydrate fraction from aloe vera gel. Immunopharmacology 35 (2), 119–128. doi: 10.1016/S0162-3109(96)00135-X
Keywords: β-mannanase, pig and poultry nutrition, feed-induced inflammatory response, immunometabolism and gut health, growth and feed efficiency
Citation: Kiarie EG, Steelman S and Martinez M (2022) Does supplementing β-mannanase modulate the feed-induced immune response and gastrointestinal ecology in poultry and pigs? An appraisal. Front. Anim. Sci. 3:875095. doi: 10.3389/fanim.2022.875095
Received: 13 February 2022; Accepted: 16 August 2022;
Published: 06 September 2022.
Edited by:
Kendall C. Swanson, North Dakota State University, United StatesReviewed by:
Haihan Zhang, Hunan Agricultural University, ChinaCopyright © 2022 Kiarie, Steelman and Martinez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elijah G. Kiarie, ZWtpYXJpZUB1b2d1ZWxwaC5jYQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.