 Rodrigo Malta Vanucci1
Rodrigo Malta Vanucci1 Daphne Wrobel Goldberg2,3
Daphne Wrobel Goldberg2,3 Andrea Maranho4Bruno de Barros Giffoni1
Andrea Maranho4Bruno de Barros Giffoni1 Isabella Cristina da Rocha Boaventura5Renata Balsamo Dias6
Isabella Cristina da Rocha Boaventura5Renata Balsamo Dias6 Simone Baratto Leonardi7Hugo Gallo Neto7Berenice Maria Gomes Silva1Daniel Wagner Rogerio8
Simone Baratto Leonardi7Hugo Gallo Neto7Berenice Maria Gomes Silva1Daniel Wagner Rogerio8 Camila Domit9
Camila Domit9 Andre Silva Barreto10Pedro Volkmer Castilho11Cristiane Koleniskovas12
Andre Silva Barreto10Pedro Volkmer Castilho11Cristiane Koleniskovas12 Henrique Chupil6
Henrique Chupil6 José Henrique Becker1*
José Henrique Becker1*- 1Fundação Projeto Tamar, Ubatuba, Brazil
- 2Instituto de Pesquisas Ambientais Littoralis, Maricá, Brazil
- 3Instituto Albatroz, Santos, Brazil
- 4Instituto GREMAR, Guarujá, Brazil
- 5Instituto Biopesca, Praia Grande, Brazil
- 6Instituto de Pesquisas de Cananéia – IpeC, Cananéia, Brazil
- 7Instituto Argonauta para Conservação Marinha, Ubatuba, Brazil
- 8Fundação Projeto Tamar, Florianópolis, Brazil
- 9Laboratório de Ecologia e Conservação, Centro de Estudos do Mar, Universidade Federal do Paraná, Pontal do Paraná, Brazil
- 10Universidade do Vale do Itajaí, Itajaí, Brazil
- 11Laboratório de Zoologia da Universidade Estadual de Santa Catarina - UDESC, Laguna, Brazil
- 12Associação R3Animal, Florianópolis, Brazil
Strand monitoring is valuable for identifying human-caused threats to sea turtles but has limitations in fully understanding their overall impact. To address this gap, we conducted a survey to examine sea turtle interactions with longline fisheries, focusing on the types of hooks used for different target fish species. The study aimed to determine the prevalence of sea turtles entangled with hooks in these fisheries and to investigate seasonal and spatial variations in strandings. From August 2015 to December 2020, regular surveys were carried out along 2,013 km, on the South and Southeast coasts of Brazil to monitor sea turtle strandings. Data were collected on the presence and types of hooks, as well as the developmental stages of the affected turtles. The study analyzed spatial and seasonal variations and conducted necropsies to determine causes of death and assess internal damage from fishing gear. A total of 4,631 stranded sea turtles were documented: Caretta caretta, n=3,678, w/hooks=96; Lepidochelys olivacea, n=753, w/hooks=20, and Dermochelys coriacea, n=200, w/hooks= 01. Four types of hooks were identified, with a predominance of “J” hooks typically used in the region to capture dolphinfish, Coryphaena hippurus (72/122 hooks). The most affected developmental stages were large juveniles of C. caretta and L. olivacea. The distribution of hooks differed among the states; “J” hooks targeting C. hippurus on the continental shelf were predominantly found in São Paulo state. In contrast, circular hooks for swordfish (Xiphias gladius) and tuna hooks for tuna (Thunnus spp.), used in offshore fisheries, were more common in southern states. The abundance of “J”, hooks increased during the spring months, aligning with the C. hippurus fishing season. “J” hooks were primarily found in the esophagus of stranded turtles (88/122), indicating that their size and shape facilitate ingestion. Necropsies revealed that 43.1% of deaths (50/116) were due to intestinal lesions caused by hooks, fishing lines, and/or cables. Despite the small relative frequency of individuals found with hooks (117/4631), pelagic longline fisheries, especially those targeting C. hippurus, are a serious threat to sea turtle populations and, therefore, require specific monitoring and mitigating measures. Furthermore, the number of affected individuals may be underestimated since stranding does not represent total mortality at sea.
1 Introduction
The incidental capture in fisheries is considered the greatest global threat to sea turtle populations (Wallace et al., 2011, 2013; Guimarães et al., 2017). Bycatch data can be obtained by monitoring fisheries either through onboard observation programs or electronic monitoring, onboard spreadsheets, or interviews with fishers during landings (Sales et al., 2008; Laporta et al., 2013) However, the Brazilian government’s onboard observer program (PROBORDO) was suspended in 2012 (Zamboni, 2020) and has not been continued to date, resulting in specific observer initiatives carried out only by universities and non-governmental organizations (NGOs). In contrast, the continuous and uninterrupted monitoring of marine animal strandings, with detailed analysis of necropsies, helps to evaluate the spatiotemporal distribution of animals and has proven to be effective in identifying causes of death related to interaction with fisheries (Tomás et al., 2008; Monteiro et al., 2016; Phillott and Godfrey, 2019; Chou et al., 2023). In fact, the presence of fishing gear in the carcasses is considered indisputable evidence of the interaction. However, when gear is not found, macroscopic and histopathological changes can be used to confirm the diagnosis of bycatch.
Pelagic longline fishing that targets swordfish (Xiphias gladius), tuna (Thunnus spp.), dolphinfish (Coryphaena hippurus), and several species of sharks, can accidentally capture, kill, or seriously injure thousands of sea turtles annually (Camiñas, 1998; Witzell, 1999; Lewison et al., 2004). In the South Atlantic, the first interactions of these animals with pelagic longline fisheries were reported in 1998 (Achaval et al., 1998; Barata et al., 1998). Currently, incidental capture in these fisheries is considered an important threat to several sea turtle populations (Domingo et al., 2006; Sales et al., 2008; Castilhos et al., 2011; Santos et al., 2011; Caracappa et al., 2018). Mortality due to lethal takes can occur during interaction or after turtles are released alive from fishing gear. The risk of mortality of post-release turtles in longline fisheries is high, especially when the animals are lifted to the vessel by the master line, due to the strong tension applied (Alegre et al., 2006; Valente et al., 2007) or when they are released with hooks attached to long lines that protrude through the gastrointestinal tract (Orós et al., 2004). Improper handling on board can also lead to fatal damages of internal tissues, and the ingestion of fishing lines can generate severe damage to the turtles, including intussusception, pleating, strangulation, and even rupture of the gastrointestinal tract (Di Bello et al., 2013). In addition to causes directly related to fishing gear, the incidental capture of non-target species can trigger systemic stress in bycatch, resulting in elevated corticosterone and glucose levels (Snoddy et al., 2009). The physical effort to escape the fishing gear and reach the surface to breathe can cause metabolic and respiratory acidosis, marked by increased blood lactate (the product of anaerobic metabolism; Harms et al., 2003), increased partial pressures of carbon dioxide and oxygen, and changes in concentrations of sodium, potassium, and chloride ions. Such acidosis leads to a reduction in blood pH and an increase in respiratory rate (Stabenau et al., 1991; Hoopes et al., 2000; Harms et al., 2003; Stabenau and Vietti, 2003). These metabolic alterations may require a period of recovery of the organism, during which the turtles become lethargic and thus, more vulnerable to predators, collisions with vessels (Williard et al., 2015), and even drowning. Interaction with longline fisheries may result in both immediate death and long-term injury. Research conducted by Work and Balazs (2010), as well as Parga (2012), found that turtles that become entangled in longline gear may suffer from severe subacute injuries, resulting in high late mortality rates. In some cases, turtles can remain adrift for weeks to months, debilitated, with limited capacity to feed and escape predators. Factors such as individual characteristics (e.g., previous health condition and intrinsic behavioral characteristics at the time of interaction), place of capture, related injuries, and environmental and climatic conditions can influence the time interval between capture and death.
Despite the growing number of studies on the sublethal effects of sea turtle bycatch (e.g., Parga, 2012; Wilson et al., 2014), accurately extrapolating the mortality of these animals based on strandings alone remains a huge challenge. A combination of factors, including distance from the shore at which the interaction occurred, prevailing and intensity of currents, and wind, can play a major role in whether the sea turtles strand (Hart et al., 2006; Cantor et al., 2020). The aim of this study was to identify potential longline fisheries that interact with sea turtles based on hooks found in live and dead specimens, as well as, to investigate the possible relationship with the cause of death through the evidence found in the necropsies. Data on the occurrence and abundance of sea turtles with hooks were analyzed to understand potential spatial and temporal variations, across states, mesoregions, months, years, and seasons. Our findings may underscore several impacts from fisheries interactions, but also support the development of strategies for reducing the capture and mortality of sea turtles in fisheries.
2 Methods
Study area. - This study was based on the analysis of data from the Santos Basin Beach Monitoring Project (PMP-BS), an environmental licensing requirement of the Brazilian Institute of the Environment and Renewable Natural Resources (IBAMA) for oil and natural gas production and transport by Petrobras, initiated in August 2015. The monitored area extends from the municipality of Laguna (28°29’43.1”S 48°45’39.0”W), state of Santa Catarina (SC), to Saquarema (22°56’08.0”S; 42°29’43.6”W), state of Rio de Janeiro (RJ), totaling 2,013 km of beaches, divided into 15 stretches. These areas are monitored daily, applying a systematic protocol, by institutions of the Aquatic Mammals Stranding Network of the Southeast and South Brazil (REMASE and REMASUL) (Petrobras, 2019) (Figure 1). The PMP-BS is regularly carried out by technicians from the following institutions: Econservation, Instituto Argonauta, Instituto GREMAR, Instituto Biopesca, Instituto de Pesquisas Cananéia (IPeC), Universidade Federal do Paraná (UFPR), Universidade da Região de Joinville (Univille), Universidade do Vale do Itajaí (UNIVALI), Associação R3 Animal, Instituto Australis, and Universidade Estadual de Santa Catarina (UDESC) (Petrobras, 2022). Data on sea turtles undergoing rehabilitation and necropsies were also collected at the Ubatuba (SP) and Florianópolis (SC) units of the Tamar Project Foundation (Petrobras, 2019) (Figure 1).
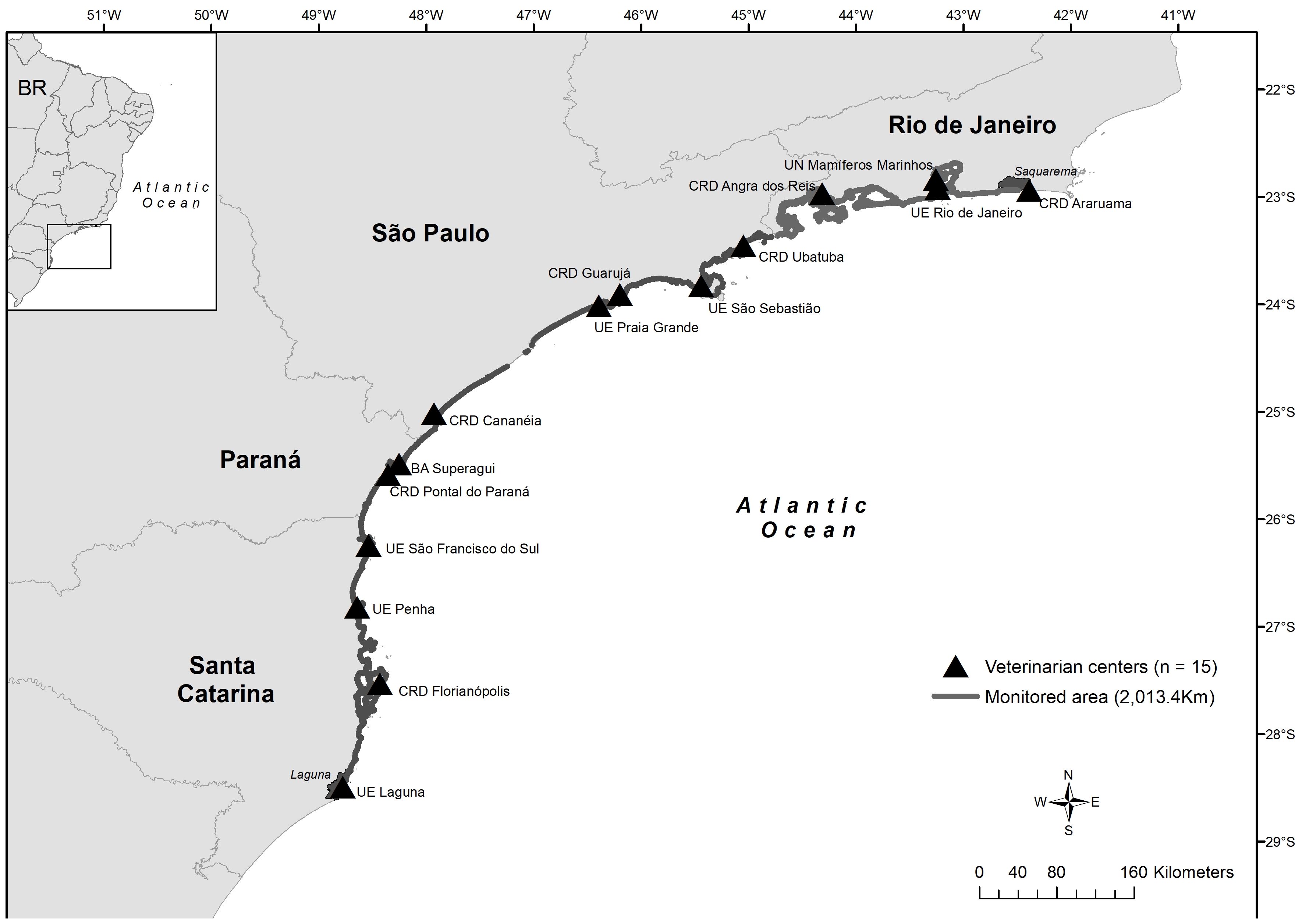
Figure 1 Coverage area of the Santos Basin Beach Monitoring Project (PMP-BS) and location of the marine fauna rehabilitation and stabilization centers. Source: Petrobras, 2022.
The live turtles that were stranded, either injured or sick, on the mentioned beaches were exclusively handled for rehabilitation. Noninvasive procedure or manipulation was performed to generate research data. In cases of rehabilitation success, the turtles were tagged and released back to sea (following standard protocols from Centro TAMAR/ICMBIO, Brazil).
2.1 Data collection and analysis
This study included all records of stranded loggerhead turtles (Caretta caretta), olive ridley turtles (Lepidochelys olivacea), and leatherback turtles (Dermochelys coriacea) found alive or dead, from 2015 to 2020. These species are all known to interact with pelagic longline fisheries in South/Southeast Brazil (Sales et al., 2008) and in other parts of the world (Polovina et al., 2004). Chelonia mydas and Eretmochelys imbricata, which are also found in the region, inhabit more coastal waters (Tagliolatto et al., 2019; Gama et al., 2021; Bezerra et al., 2024), and the interaction with pelagic longlines is rare, and so will not be addressed in this study. In fact, these species are more likely to encounter artisanal and recreational fishing hooks.
When recording stranded animals, dead or alive, species were identified and biometric data was collected, whenever possible, including curved carapace length (CCL) and curved carapace width (CCW), according to Bolten (1999). Individuals were also classified in terms of subjective body condition, when possible, divided into three pre-established categories according to the PMP-BS monitoring protocol, namely: excellent/good, thin, and cachectic. Since the “excellent” category does not apply to sea turtles, as proposed by the PMP-BS protocol, this category was grouped with the “good” classification. The “excellent” or “good” body score was assigned to individuals with straight or convex plastron, firm consistency, suggestive of an animal with developed musculature, a reserve of adipose tissue, and bright and protruding eyes. The “thin” body score was assigned to animals with slightly concave plastron, firm consistency, leaner musculature, little reserve of adipose tissue, and non-protruding eyes. The “cachectic” body score was attributed to animals with concave plastron, carapace softening, prominent bones, exposure of the entoplastron and the supra-occipital crest, and sunken eyes (Thomson et al., 2009).
The animals found alive underwent clinical examination, blood collection for hematological and biochemical analysis, and received proper pharmacological and nutritional care in rehabilitation centers. Hooks lodged on the outside of the body were removed. Individuals with suspected hook ingestion underwent imaging tests (radiography and ultrasonography) to locate the gear and identify associated lesions. Surgical or endoscopic procedures were performed, when possible, to remove hooks, cables, and/or fishing lines.
In case the animals were found dead, the decomposition stage of the carcasses was classified into four categories: code 2 (fresh carcass; recent death), code 3 (moderate decomposition; minimal autolysis), code 4 (advanced decomposition), and code 5 (skeletonized or mummified), adapted from Geraci et al. (2005). Necropsy procedures were exclusively conducted on carcasses classified under codes 2 and 3, highlighting those that presented with either a fresh state of recent death or displayed moderate decomposition with minimal autolysis, as well as on animals that succumbed during the rehabilitation process. During macroscopic examinations, lesions and the type of ingested gear were meticulously recorded based on their specific locations throughout the turtle’s gastrointestinal tract (i.e., oral cavity, esophagus, stomach, small intestine, large intestine, and cloaca). This comprehensive evaluation of both the placement of hooks and lines and their resulting macroscopic injuries facilitated a detailed assessment of the severity of these injuries, allowing for an informed analysis of their potential contribution to the turtles’ mortality. Tissue samples from all organs were then collected and preserved in 10% buffered formalin for further histopathological analysis.
For carcasses in advanced stages of decomposition (i.e., codes 4 and 5), only the presence and location of hooks, cables, and fishing lines were documented. The records of strandings, veterinary care, and necropsy data were stored and made available at the Aquatic Biota Monitoring Information System (SIMBA) (Almeida, 2015).
The reproductive maturity of the individuals was assessed by examining both the macroscopic and histologic features of their gonads. The stage of development of living animals was classified by mean CCL of females nesting on Brazilian beaches, i.e., 70.6 cm (SD=3.4) for Lepidochelys olivacea, ranging between 65 and 79 cm; and 99.9 cm (SD=4.6) for Caretta caretta and ranging between 90 and 114 cm (Soares et al., 2021). The mean CCL provides a more representative measurement of the average size of population by accounting for variations within it, unlike the minimum CCL, which may only indicate the size of the smallest, potentially atypical individuals.
To evaluate the spatial distribution and abundance of individuals with hooks, the number of strandings was examined by comparing each Brazilian federative unit (state) and mesoregion. Mesoregions are distinct geographical areas, which share physical, environmental, and socioeconomic similarities with one another. This comparison helped to gain insights into the spatial occurrence of animals with hooks (IBGE, 1990) (Figure 2). Potential variations in the spatial distribution of different hook types were investigated, categorizing them between those used in oceanic longline fishing and those used for longline fishing over the continental shelf. It was also examined whether the abundance of turtles found with hooks varied seasonally, over months or seasons, potentially correlating with the regional longline fishing seasons.
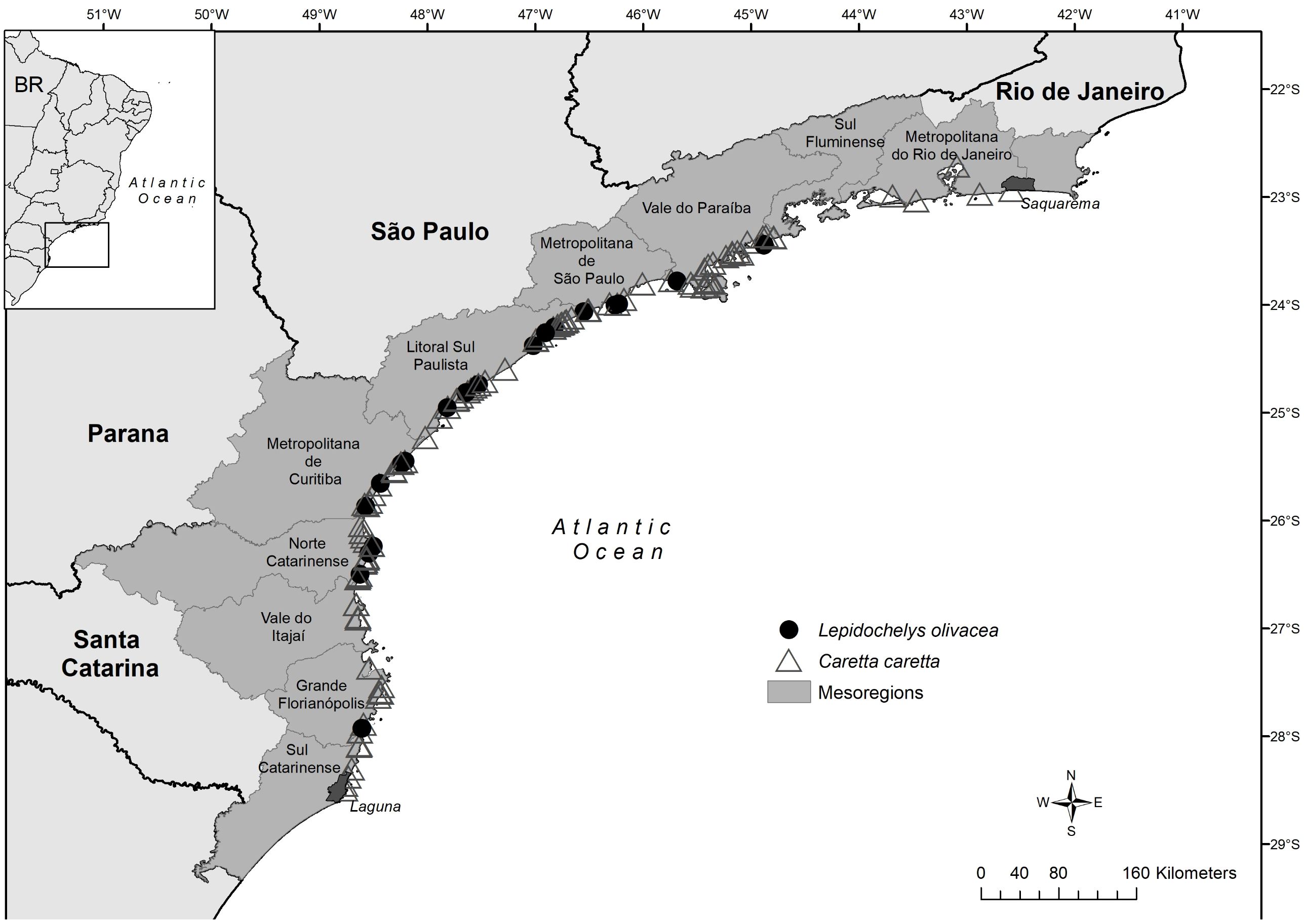
Figure 2 Distribution of sea turtles strandings in the monitored regions (dark shaded areas) with the presence of hooks.
2.2 Occurrence and characterization of hooks
The hooks found were measured and classified using photographs provided by the institutions. Due to the inaccuracy of some photos and scales, the hooks were categorized into size classes with reference to the height (greatest rectilinear length) of each hook. Based on the concept of fishing (Marcovaldi et al., 2006; Giffoni et al., 2017), longline fishing modalities are classified and grouped according to criteria such as target species, hook type, bait type, fishing area, time of year, and other factors. In this regard, the pelagic longlines used in the South/Southeast region of Brazil have typical characteristics such as types and sizes of hooks, method of tying the line, and presence or absence of steel cable in each fishery. This prior knowledge was utilized to categorize the hooks obtained during the study, and additional consultations were made with researchers involved in monitoring these fisheries to address any uncertainties.
The hooks were classified into the following categories: “J-Dolphinfish”, for catching Coryphaena hippurus; “J-Swordfish”, for Xiphias gladius, “Tuna-hook” for catching Thunnus spp., and “Circular” for catching swordfish, tuna, and sharks (Figure 3). The hooks that did not fit in any of the categories, due to the presence of different characteristics or because they were incomplete or fragmented, were grouped in the “unidentified” category. Important features of the hooks, such as the presence of an eye, ring, wire, and/or way to tie the hook on a fishing line, were considered in the classification, as shown in Figure 3.
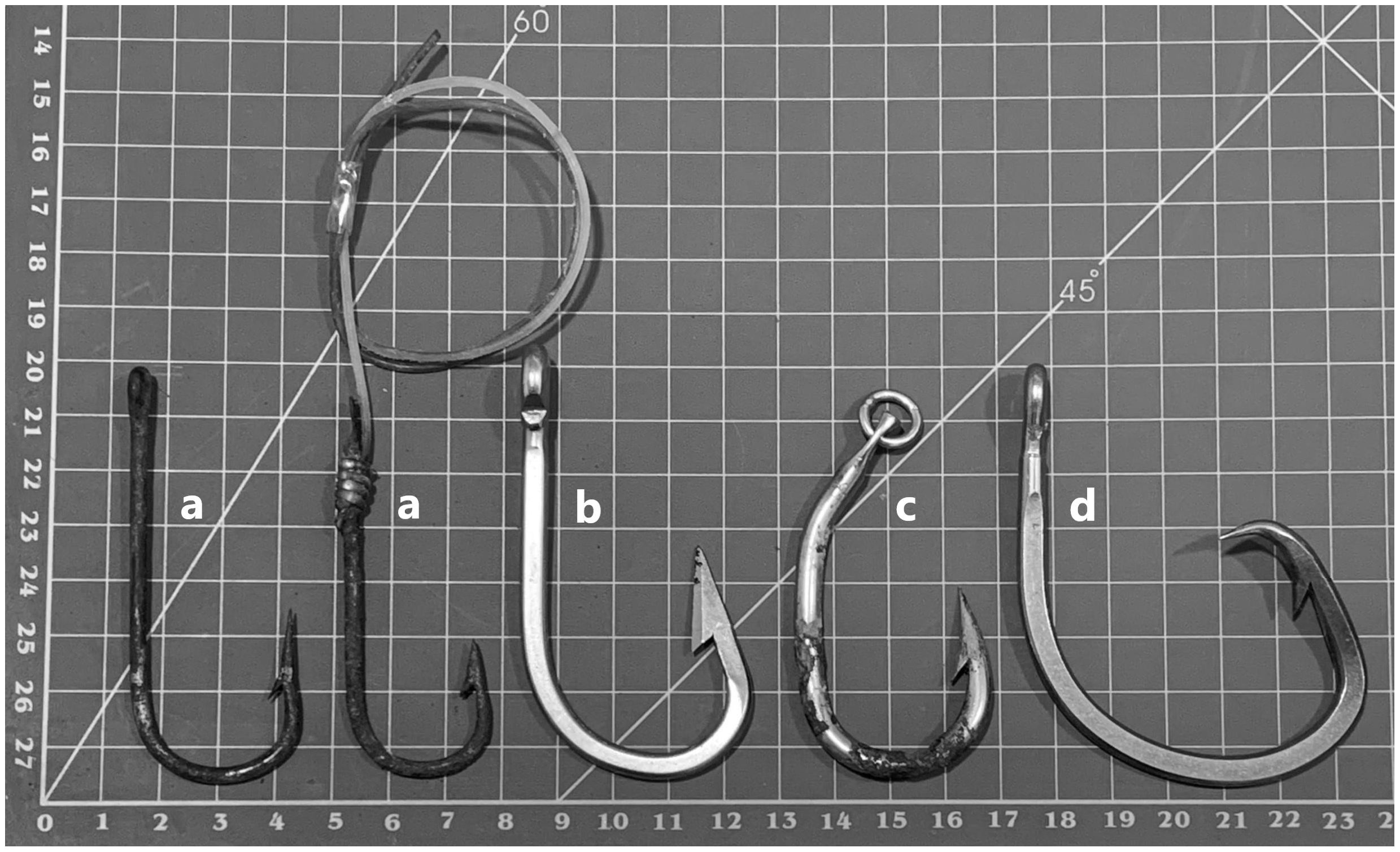
Figure 3 (A) Two different types of “J” Dolphinfish hooks; (B) “J” Swordfish hook, (C) “Tuna-hook”; (D) “Circular” hook. Photo by Fundação Projeto Tamar.
2.3 Background of longline hooks
Hooks used for catching swordfish (Xiphias gladius) are larger and sturdier than hooks used for catching common dolphinfish (Coryphaena hippurus). The national fishing fleet always tie the hooks with a steel cable, which increases the strength for catching sharks. The “J” 9/0 hooks are more commonly used by the industrial fleet, which operates in the Southeast of Brazil. However, this fleet also uses circular hooks and “Tuna-hooks” (Fiedler et al., 2016; Giffoni et al., 2017). The fleet that catches common dolphinfish uses less resistant (and cheaper) hooks than those used to catch swordfish and tuna and does not use steel cable for tying the hooks. The “Tuna-hook”, as the name implies, is mainly used in tuna fishing, and consists of a ring for greater bait mobility. The circular hooks are shaped as their name indicates, with or without a ring, and include a spearhead facing the rod equal to or greater than 14/0. These hooks became mandatory in Brazil in November 2017, with the publication of Interministerial Ordinance No. 74, of the Ministry of Industry, Foreign Trade, and Services and the Ministry of Environment (Brasil, 2017), for fishing tuna (Thunnus albacares, Thunnus alalunga, Thunnus obesus) and for longline fishing aimed at catching swordfish.
2.4 Statistical analysis
Permutational multivariate analysis of variance (PERMANOVA) was used to test the hypotheses involving the records of chelonians with hooks (abundance) in relation to the spatial distribution (states and mesoregions) and temporal distribution (month, year, and season) where the factors were considered as fixed effects, and the number of permutations was 9,999. This analysis was performed in Past software (Hammer et al., 2001), and the similarity matrices were constructed using the Bray-Curtis coefficient (abundance data).
3 Results and discussion
3.1 Species and biometrics
From August 2015 to December 2020, 4,631 stranded sea turtles were recorded, comprised of C. caretta (n=3,678), Lepidochelys olivacea (n=753), and Dermochelys coriacea (n=200). For C. caretta, hooks were found in 96 individuals (2.61%). For L. olivacea, hooks were found in 20 individuals (2.61%). A single individual of D. coriacea was found with a hook (0.5%) (Table 1). Since this was the only occurrence for this species, the analyses were limited in this study. The predominance of C. caretta (82.05%) coincides with previous results of on-board monitoring in pelagic longline fisheries in Brazil (Sales et al., 2008; Giffoni et al., 2017) and in other locations around the world (Lucchetti et al., 2017). However, while the second most commonly encountered species in these studies was the D. coriacea, the second most common species among the stranded individuals observed in this study was L. olivacea (17.09%). These findings corroborate the results from Giffoni et al. (2017), who observed that C. caretta and L. olivacea were the two most frequently observed species of sea turtles in longline fishing for C. hippurus of the Itaipava fleet in Southern and Southeastern Brazil.
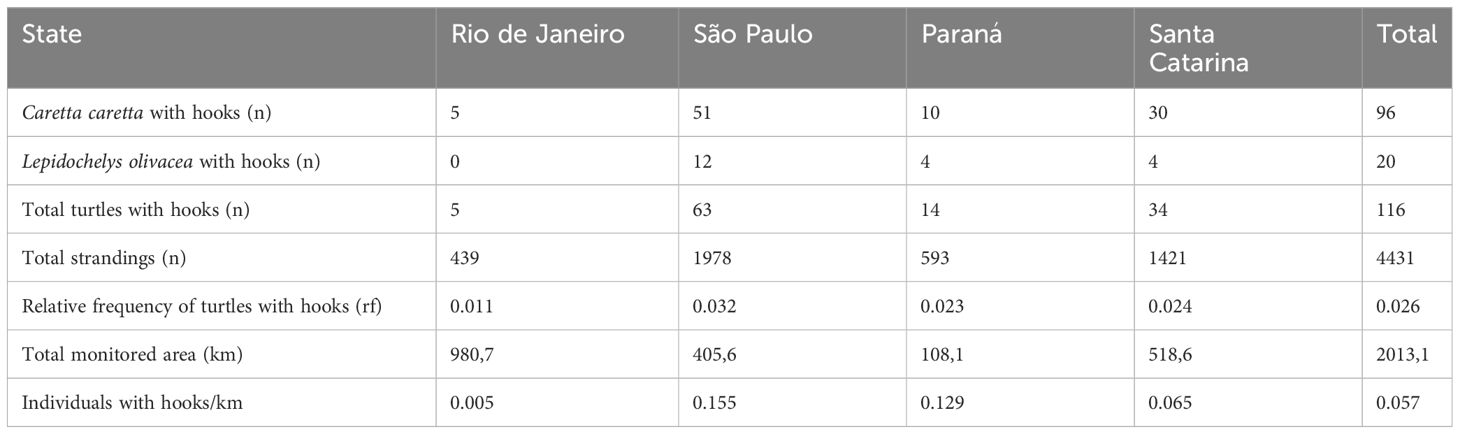
Table 1 Frequency of individuals with hooks in the different states of the study.
The mean curved carapace length of C. caretta individuals that interacted with hooks was 79.6 cm (ranging from 55.8 cm to 116 cm, SD= 10.05; n=85). Of these individuals, 3.52% (n=3) were adults and 96.47% (n=82) were juveniles (see Figure 4B). These findings are in line with previous studies with juvenile C. caretta interacting with longline fisheries (Giffoni et al., 2005, 2017). The individuals of L. olivacea that interacted with hooks had the mean curved carapace length of 66.8 cm (ranging from 58.0 cm to 74.1 cm; SD=4.24, n=15), of which 20,0% (n=3) were adults and 80,0% (n=12) juveniles (Figure 4A), which is similar to the results found in previous studies with longline fisheries of Itaipava fleet (Giffoni et al., 2017). The rate of hook-related impacts on adult turtles of both species may be underestimated. This is due to the minimum sizes of nesting females in Brazil—65 cm for L. olivacea and 90 cm for C. caretta (Soares et al., 2021)—combined with the reality that strandings don’t fully account for all at-sea mortalities. According to Crouse et al. (1987), enhancing the survival rates of large juveniles — those who have already navigated through the most vulnerable years — can significantly increase the likelihood of reaching maturity. This, in turn, magnifies the contribution of adult stages to the population, emphasizing the increased reproductive value that comes with maturity. From this perspective, implementing mitigation measures aimed at reducing late juvenile bycatch is essential.
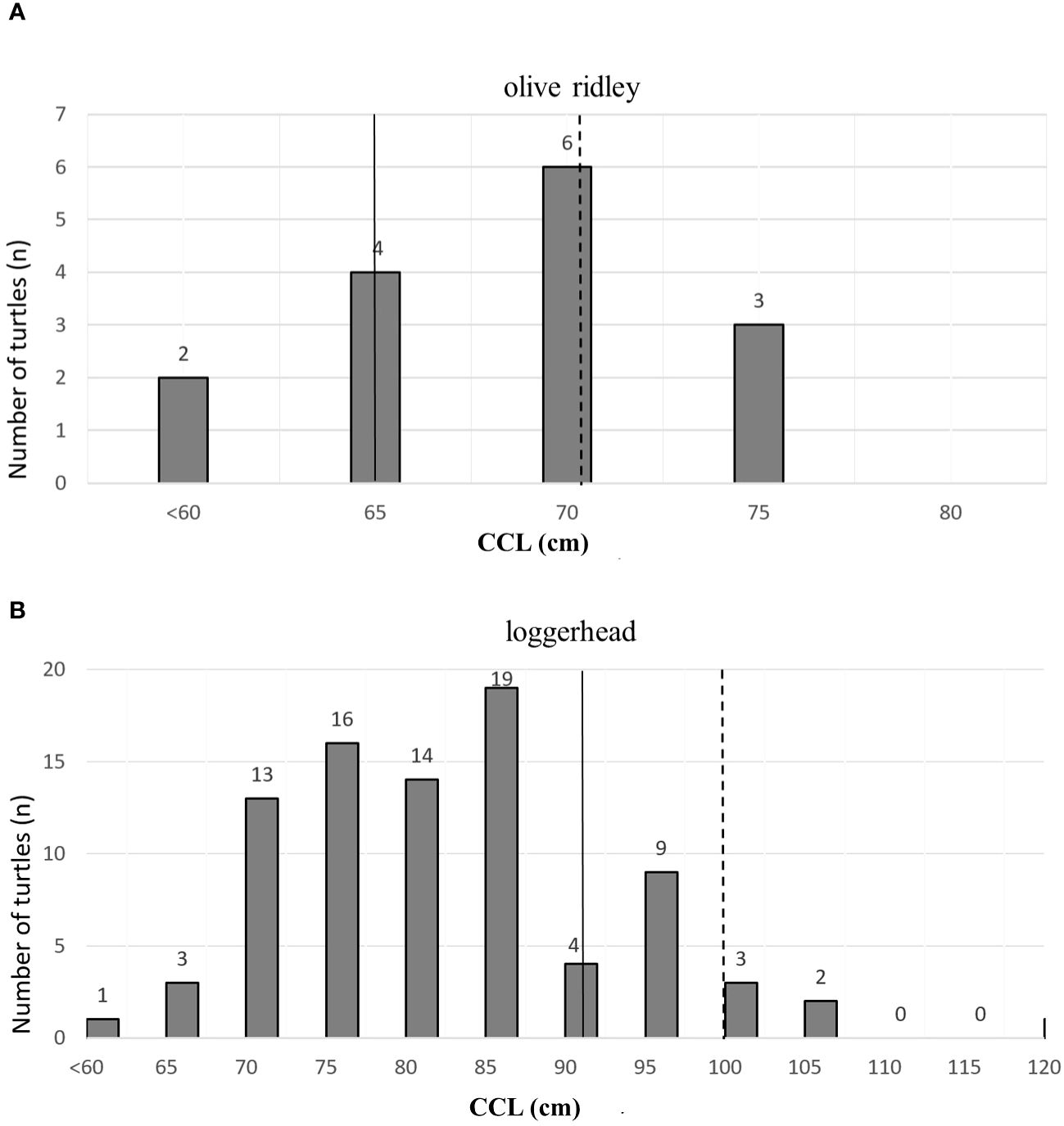
Figure 4 (A) Distribution of curve carapace length classes: (A) olive ridley turtle (L. olivacea); (B) loggerhead turtle (C. caretta). Dashed line indicates the mean CCL of nesting females at Brazilian rookeries: 70,6 cm for olive ridley and 99,9 cm for loggerhead (Soares et al., 2021). Solid lines indicates the minimum CCL of nesting females at the same Brazilian rookeries: 65 cm for Olive ridley and 90 cm for loggerhead (Soares et al., 2021).
3.2 Hooks found: types, seasonal and spatial distribution
Of the 122 hooks found on 116 sea turtles, 88 were identified (Figure 3; Table 2). The “J” hooks were the most frequently used for catching dolphinfish (C. hippurus), representing 59.01% (72/122) of the hooks found. The “J” hooks normally used to catch swordfish (X. gladius) represented 9.02% (11/122) of the hooks found. The least representative hooks in the sample were the “Tuna-hooks” with 1.6% (02/122) and the “Circular” hooks, with 2.45% (03/122). A total of 34 out of 122 hooks tested (27.86%) were not classified due to either the hook being incomplete or because it presented characteristics that were uncommon to the known fishing practices in the region.
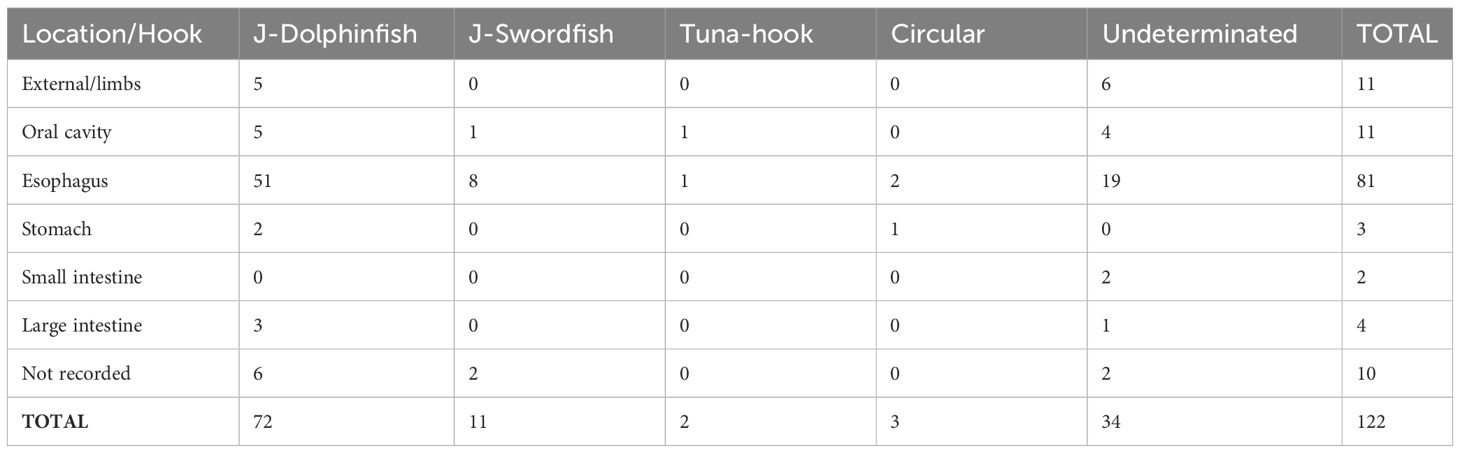
Table 2 Attachment site/use of different hook categories.
The abundance of turtles with hooks varied significantly between states (DF=3, SQ=5.7401, MQ=1.9134, F=0.64133, P=0.0001) and mesorregions (DF=11, SQ=7.8719, MQ=0.71562, F=0.23986, P=0.0002) within the study area. The state of São Paulo (Southeast region) had the highest number of sea turtle strandings (n=1978) for the species Caretta caretta and Lepidochelys olivacea, as well as the highest number of individuals with hooks (n=63), which results in the highest relative frequency (0.032%) of individuals with hooks. Santa Catarina (South region) is the second state in total number of strandings (n=1420), although the relative frequency is similar to its neighboring state, Paraná (South region) (0.023%), which has the smallest monitored beach area of the entire region (area= 90km; stranding records n =593). Rio de Janeiro (Southeast region) has the largest area of monitored beaches and the lowest number of strandings (n=439) and individuals with hooks (n=5) (Table 1). In terms of mesoregions (IBGE, 1990), except for the state of Rio de Janeiro, with rare records of chelonians stranded with hooks, a gradual decrease was observed in the occurrence of “J” hooks for dolphinfish from the coast of the state of São Paulo towards the South of Brazil. This result suggests the greater presence of the longline fleets that use “J” hooks for C. hippurus in the latitudes that cover the state of São Paulo (Southeast), as also found for the Itaipava fleet (Leite et al., 2005; Bugoni et al., 2008).
Longline fishing, especially for C. hippurus, significantly impacts many sea turtles due to the overlap of fishing areas with various species habitats (Gilman et al., 2006, 2008). Dolphinfish are known to inhabit shallower waters (Benetti et al., 1995). Hence, fishing for this species typically occurs closer to the coastline in relatively shallower waters compared to the operations of tuna or swordfish fleets, which often unfold in deeper offshore areas (Lewison and Crowder, 2007). In this study, the predominance of “J” hooks for dolphinfish may be associated with the fishing effort of these fisheries in the continental shelf areas, at latitudes corresponding to the coast of the state of São Paulo (Southeast), which accounts for most strandings (Table 1) (Giffoni et al., 2017; Maçaneiro et al., 2018). Bugoni et al. (2008) reported 15 fishing cruises of this fleet between 2001 and 2006, located between latitudes 18°S and 35°S, and recorded 1.08 catches of turtles of four species for every 1000 hooks. Dallagnolo and Andrade (2008) described the operational area of this fleet between the coast of Paraná and the island of Florianópolis (SC), preferably between the isobaths of 100m and 200m.
The two-factor PERMANOVA revealed a significant variation in the abundance of chelonians with hooks typically used in fisheries on the continental shelf (targeting common dolphinfish) and more oceanic fisheries (targeting tuna and swordfish) among the states DF=3, SQ=6.2081, MQ=2.0694, F=0.7547, P=0.0001) and mesoregions (DF=11, SQ=8.158, MQ=0.74163, F=0.27048, P=0.0001). Few animals with hooks used in oceanic fisheries such as the “J” hook for swordfish and the “Tuna-hook” for tuna were found, likely due to the preference of these fisheries for further offshore areas. This means that carcasses with these types of hooks have a lower chance of stranding. Despite the smaller quantity, “J” hooks for swordfish were found more frequently in the state of Santa Catarina, the southernmost region of the study area, differing significantly from other states and mesoregions. The greater frequency of this hook in Southern Brazil may be related to the proximity of the fleet’s area of operation (Maçaneiro et al., 2018).
Few individuals with circular hooks were found (n=3). At the end of 2017, the use of these hooks became mandatory for pelagic longline fisheries in Brazilian and foreign vessels operating in Brazilian jurisdictional waters or international waters for the capture of tuna and swordfish species (Brasil, 2017). However, this ordinance became valid only in November 2018 and did not include the mandatory use of circular hooks for pelagic longline fisheries targeting the common dolphinfish. Tests carried out between 2004 and 2008 on 229 sets (145,828 hooks) showed satisfactory results in the use of circular hooks to reduce bycatch of C. caretta and D. coriacea (Sales et al., 2010). The lower frequency of turtle strandings with circle hooks could be attributed to oceanic fisheries operating further from the coast than fisheries using “J” type hooks. At the same time, this may also indicate the effectiveness of the hook design, which tends to lodge in the turtles’ mouths, allowing for removal by fishermen. Another point that may explain the low frequency of circular hooks found is that this type of gear was only adopted by the longline fleet in November 2018, through Federal Ordinance No. 74. For this reason, while the other hooks were being used throughout the study period, the circular hooks only began to be used in 2018. Any attempt to relate hook numbers to turtle interaction should be viewed with caution. Furthermore, stranding data associated with other forms of bycatch monitoring should be continuously collected for future comparisons.
The number of turtles with hooks did not vary significantly among years (DF=4, sq=16.13, F=1.418, P=0.1599) and months (DF=11, SQ=16.13, F=0.8675, P=0.6662). However, between seasons there was a significant difference (DF=3, SQ=2.619, F=2.467, P=0.029). An increase in turtles found with hooks during the spring and summer months was observed, contrasting with previous studies that reported no significant seasonal variation in overall strandings (Cantor et al., 2020). The noticeable surge of turtles found stranded with hooks during the spring and summer months could potentially be linked to the start of the dolphinfish (C. hippurus) fishing season in the region. During this season, dolphinfish become a primary target for the area’s pelagic longline fisheries (Dallagnolo and Andrade, 2008). Notably, in 2018, there was a substantial increase in the overall strandings; and while the number of stranded C. caretta remained steady, the count of individuals found with hooks declined in the last four monitored years (Figure 5B). In contrast, for L. olivacea, the number of individuals with hooks increased, while the total number of strandings of this species decreased over the past two years (Figure 5A). For both species, the population tends to grow in Brazil, based on nesting sites (Marcovaldi and Chaloupka, 2007; Silva et al., 2007). In this regard, a gradual increase in the presence and abundance of these species in feeding areas is expected, followed by a consequent increase in interaction with human activities, including fishing. This hypothesis converges with the results observed for L. olivacea but does not explain the reduced presence of hooks for C. caretta in recent years. For the hooks classified as “J-Dolphinfish”, a significantly higher number of strandings was observed from October to December. This period coincides with the time when the Itaipava fleet (from the state of Espírito Santo) migrates to the South/Southeast regions and targets the dolphinfish (C. hippurus) (Fiedler et al., 2015).
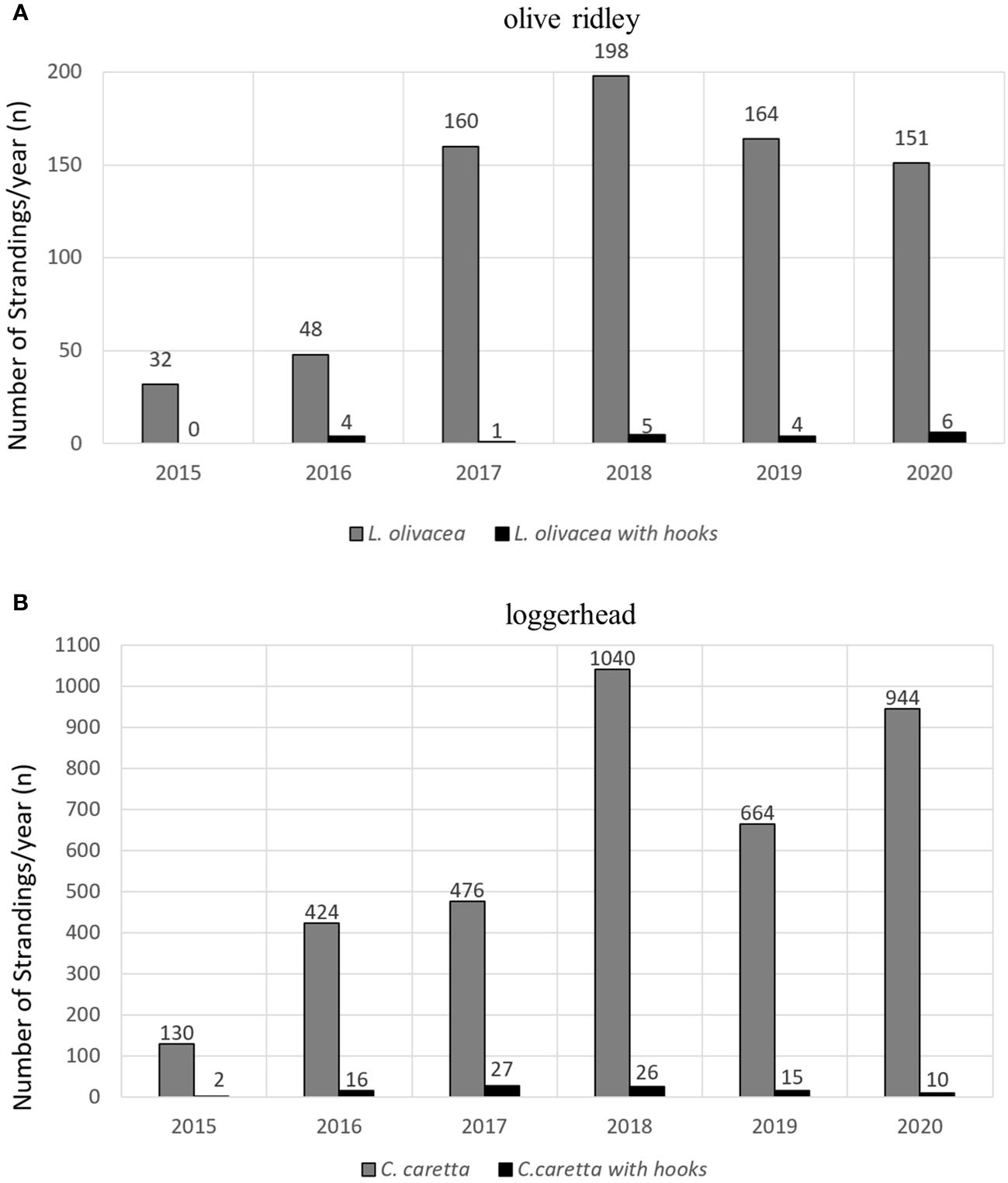
Figure 5 Annual variation in the occurrence of strandings and presence of hooks: (A) olive ridley turtle (L. olivacea); (B) loggerhead turtle (C. caretta).
3.3 Location of hooks and severity of lesions
Out of the 112 hooks recovered, 11 (9.8%) were discovered externally, primarily on the front flippers (Table 3). While these instances are generally classified as low risk (Ryder et al., 2006), complications can arise if these hooks are entangled with fishing lines, potentially leading to strangulation, lacerations, hemorrhages, fractures, avulsions, and even limb loss.
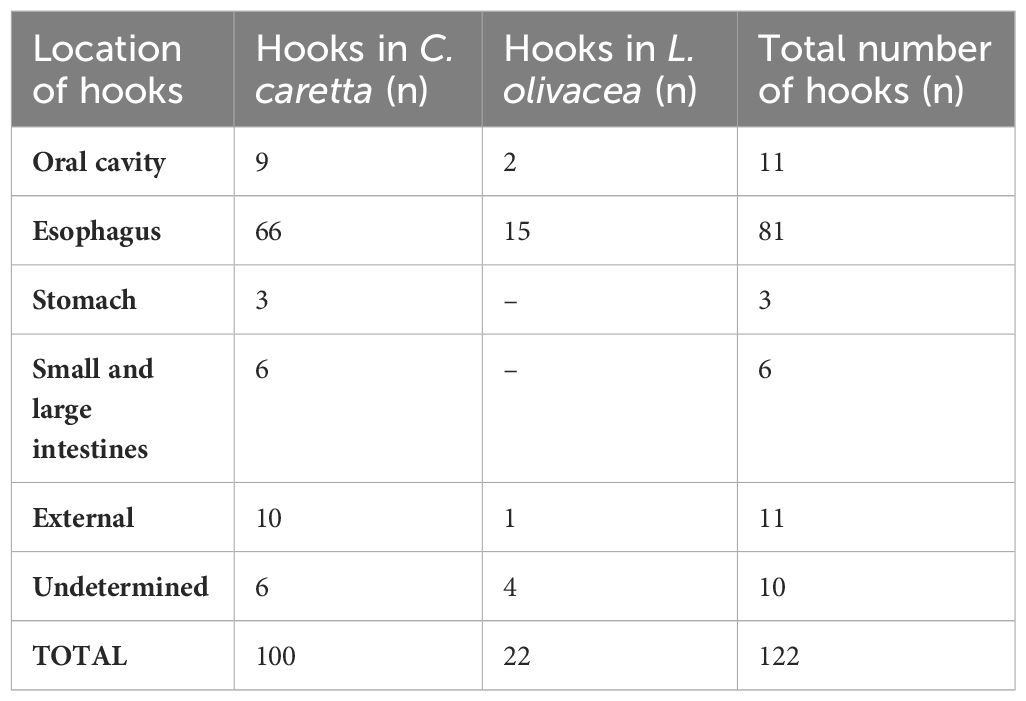
Table 3 Location of the hooks found in the 116 turtles of the study, divided by species (6 individuals had more than one hook).
Although longline fishing is well recognized as a serious threat to several species of sea turtles (Camiñas and de la Serna, 1995; Laurent et al., 1999; Lewison et al., 2004; Camiñas et al., 2006), some authors claim that these animals can tolerate serious injuries after capture and can continue feeding with one or more hooks lodged in the digestive tract (Tomas et al., 2001; Alegre et al., 2006; Caracappa et al., 2018). This fact may explain the good body score of some individuals found with fishing gear in this study. In addition to the potential harm caused by hooks, incidental capture in longline fisheries can cause other serious problems such as gas embolism (i.e., when turtles undergo a rapid decompression, causing nitrogen to form bubbles in supersaturated tissues) (García-Párraga et al., 2018; Crespo-Picazo et al., 2020), capture myopathy (i.e., a syndrome caused by incidental capture, characterized by anaerobic metabolic changes leading to the buildup of large amounts of lactic acid, metabolic acidosis, and secondary muscle necrosis) (Phillips et al., 2015) or trauma during improper handling of animals on board.
The “J” hooks for dolphinfish (C. hippurus) were found in almost all parts of the gastrointestinal tract, with the highest frequency seen in the esophagus. Although the other types of hooks were also found mainly in the esophagus, the sample size was too small to draw conclusions regarding their locations (Table 3).
3.3.1 Oral cavity
Hooks were found in the oral cavity of 9.0% (11/122) of the turtles examined. Injuries from hooks in these regions are typically low risk, unless the hooks embed within critical and sensitive structures such as bones, mandibular joint, palate, tongue, or glottis (Parga, 2012). Necropsies from this study revealed the mild nature of the lesions, characterized by caseous plaques at the hook attachment sites and wire-induced lesions in the buccal commissures (Figures 6A, B). Hooks becoming lodged in the oral cavity, especially those that penetrate bone structures, can cause severe injuries and osteomyelitis (bone infections) (Ninou et al., 2008; Parga et al., 2010). If improperly treated, these infections can spread and potentially lead to life-threatening complications. Damage to the glottis, a delicate hermetically sealed cartilage structure protecting the airway entrance, can result in serious repercussions such as drowning or lung infections. Similarly, large hooks can interfere with a turtle’s ability to eat, leading to chronic malnutrition and cachexia, a wasting syndrome that can ultimately prove fatal over time (Caracappa et al., 2018). Of all injury occurrences, 73.8% were considered severe, with ingested hooks advancing to the digestive tract, beyond the oral cavity. In 66.4% of cases, the hook was lodged in the esophagus (81/122); in 2.45% of cases, in the stomach; in 4.92%, in the small and large intestines (06/122) (Table 3). The ingestion of fishing hooks and/or cables can be life-threatening to animals (Parga, 2012; Caracappa et al., 2018). It can cause severe injuries, such as plication (i.e., folding of intestinal loops) (Bjorndal et al., 1994), which occurs when the hook lodges in some portion of the digestive tract and the line is pulled caudally through peristaltic movements (Franchini et al., 2018). More severe injuries include intussusception, characterized by the penetration of one segment of the gastrointestinal tract into the adjacent segment. Both injuries can provoke intestinal obstruction and ischemia, leading the animal to death.
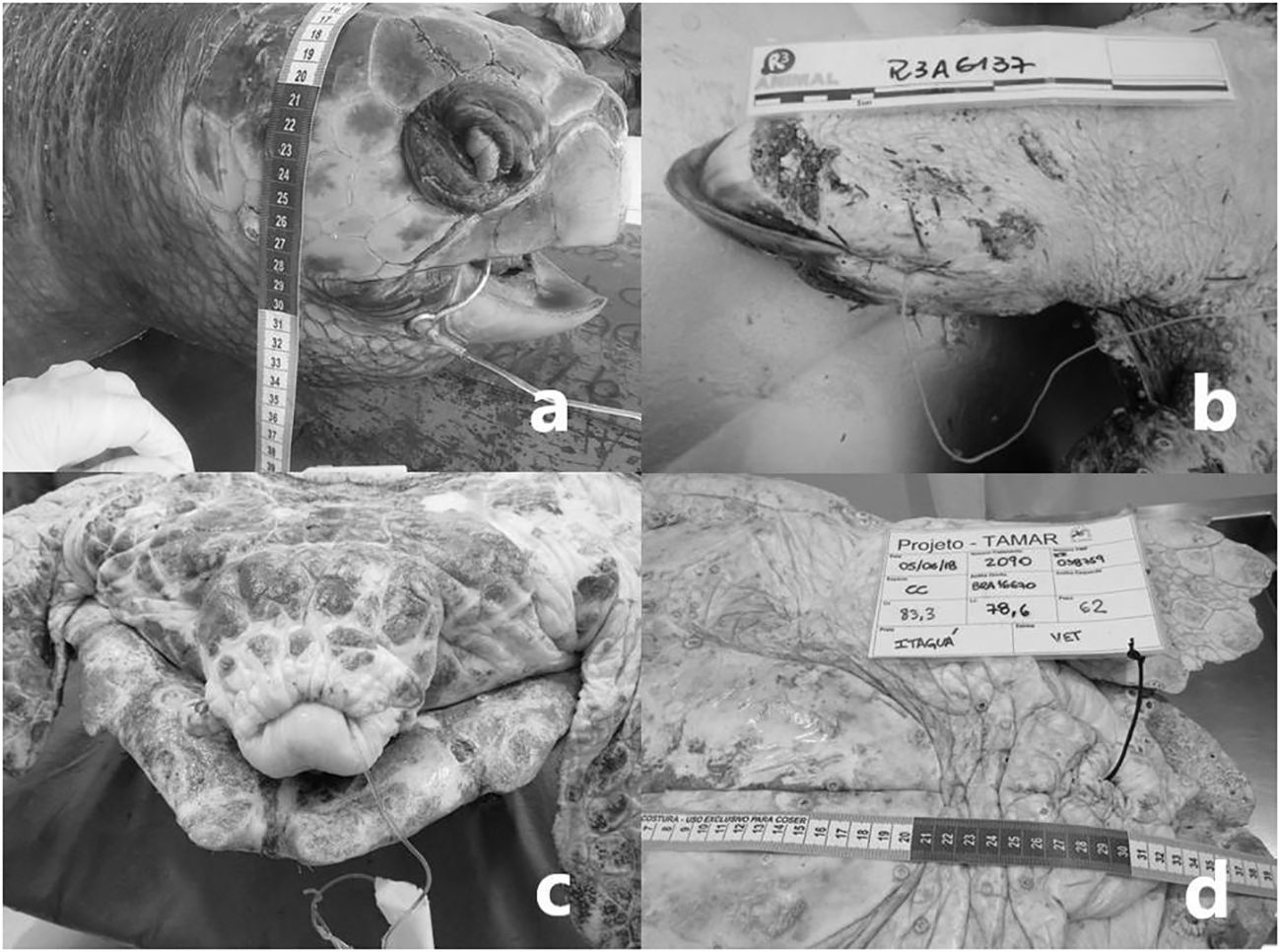
Figure 6 (A) Loggerhead turtle with hook lodged in the oral cavity – Photo by Udesc; (B) Loggerhead turtle with nylon thread coming out of the oral cavity – Photo by R3animal; (C, D) Nylon thread coming out through the cloaca – Photos by Fundação Projeto Tamar.
3.3.2 Esophagus
In this study, 66.4% of the examined chelonians (n=81) exhibited caseous and/or ulcerative lesions in the esophagus resulting from the presence of hooks (Figure 7A). Among this group, fibrous tissue deposits had encapsulated the hooks within the esophageal mucosa in 51% of cases (n=62), as seen in Figure 7B. The esophagus of sea turtles is able to resist injury because of its muscular structure and layers of keratinized squamous epithelium (Díaz-Figueroa and Mitchell, 2006). Despite hooks embedded in their esophageal walls, these animals can survive and continue to feed without experiencing notable nutritional drawbacks, even when fibrous tissue narrows the lumen (Valente et al., 2007). A study by Alegre et al. (2006) tracked 10 turtles with hooks lodged in their esophageal wall over two years, revealing that 50% naturally expelled the hooks without major clinical complications. A subsequent endoscopy of the remaining hooks revealed insignificant injurious effects. However, perforations near the organ’s most caudal portion, close to the heart and primary vessels, pose a heightened risk and can lead to the animal’s immediate death (Ryder et al., 2006; Casale et al., 2008).
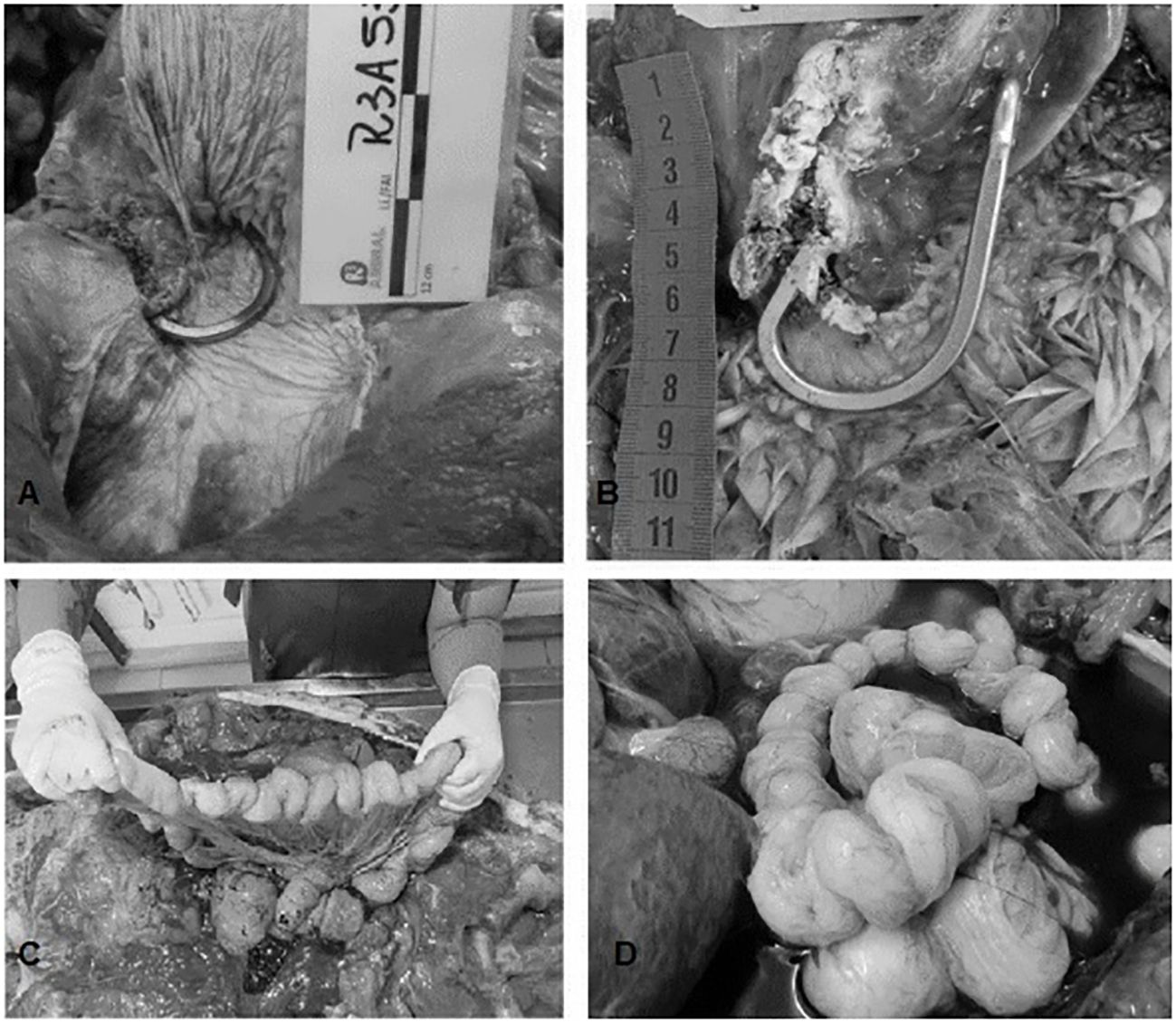
Figure 7 (A) Hook lodged in distal esophagus – Photo by R3animal; (B) Hook attached and encapsulated to the esophagus – Photo by Udesc; (C, D) Necropsies of loggerhead turtles performed at the Fundação Projeto Tamar in Ubatuba, showing pleating of intestinal loops caused by fishing line. Photo by Fundação Projeto Tamar.
3.3.3 Stomach and intestines
In the present study, the occurrence of hooks located in the stomach (3.3%) (n=3) and intestines (4.9%) (n=6) was rare (Table 3); however, mucosal lesions were observed in 77.6% of necropsied animals. These lesions were mainly caused by fishing lines and/or cables attached to the hooks, which when pulled cause characteristic linear lesions in the mucosa of the organ, with subsequent inflammation, ulcers, and caseous plaques. Orós et al. (2004) reported the presence of deep ulcers, necrotic enteritis, coelomitis, and septic shock associated with fishing lines attached to ingested hooks.
Hooks embedded in the stomach pose severe risks as they are prone to pierce this organ’s slender wall. This can yield coelomitis, or an inflammation of the abdominal cavity, potentially culminating in death due to septic shock (Orós et al., 2004; Casale et al., 2008). No living turtles with stomach-trauma from hooks were found in our study. Such injuries typically lead to immediate death, as outlined by Casale et al. (2008). Notably, the turtles discovered with hooks in their stomachs during our study period were already deceased upon beach discovery. Even though rare in our study, intestinal lesions can turn severe. Plications and intussusceptions could trigger ischemia and the intestine’s rupture, damaging the villi, essential for nutrient absorption, which in severe instances can result in critical medical complications. Our study detected injuries of varying severity in 65.6% (n=80) of the animals examined, primarily resulting from the ingestion of fishing lines and/or cables. Enteritis, ulcers, caseous plaques, plication, intussusceptions, and acute gastric volvulus (i.e., stomach rotation around its axis) were most frequently observed; only one turtle presented intestinal rupture (see Figures 7C, D). Following the rehabilitation process, a single live-treated animal expelled the hook lodged in its intestine. Although it is possible for this hook to be spontaneously expelled through a lengthy process, this can worsen the clinical condition of the turtle and should be avoided for the sake of the animal’s wellbeing (Di Bello et al., 2013). In this study, only Caretta caretta turtles were found with hooks lodged in their stomachs and intestines. Comparative studies on the anatomical characteristics of the gastrointestinal tract among the five sea turtle species outlined by Magalhães et al. (2012) suggest that C. caretta possess longer and larger diameter organs compared to other Chelonidae family members. This feature might allow them to ingest larger hooks that L. olivacea could not. In 8.2% of cases, the precise location of the gastrointestinal hooks was unverifiable due to the advanced decomposition state of the carcasses.
3.4 Fishing lines and cables
Fishers often return turtles to the sea with the hook lodged in the gastrointestinal tract, after cutting the fishing line. However, any remaining line fragment associated with the hook can cause injury, as mentioned earlier (Schofield et al., 2000; Di Bello et al., 2007). In this study, 38.7% (45/116) of the turtles presented fragments of fishing lines coming out of the cloaca (Figures 6C, D). Plication was the most important and severe lesion observed in imaging examinations (ultrasonography) and necropsies (Figures 7C, D) and it was observed in 54 of the 86 analyzed individuals (62.8%). In 43.1% of cases (50 out of 116), the animal’s demise was directly attributable to the presence of one or more fishing line fragments in the intestines, causing lethal injuries. This aligns with Di Bello et al. (2013) observations in a study encompassing 126 sea turtles, wherein approximately 20% of mortality was linked to the presence of fishing lines in the digestive tract. These findings support earlier research suggesting that even though turtles may carry hooks internally, they may not necessarily die unless these hooks are attached to fishing lines (Alegre et al., 2006; Valente et al., 2007; Parga, 2012; Caracappa et al., 2018). After ingesting fishing gear that causes gastrointestinal lesions, turtles tend to reduce their food intake and, subsequently, initiate a catabolic state associated with progressive weight loss, anemia, hypoproteinemia, hypoglycemia, dehydration, and bone demineralization. This condition favors the action of opportunistic pathogens resulting from immunosuppression, leading to a gradual decrease in the animals’ body scores.
In this study, body score could be determined in 61.2% of the cases (71/116), showing that body condition of turtles found with hooks was rated as excellent or good for 54.9% of cases, and thin for 45.1% of cases. Only 5.7% were classified as cachectic, or severely underweight. These results suggest that most animals died quickly before any weight loss associated with chronic injuries from fishing could occur. Conversely, the fact that around 45% of turtles were considered thin implies longer-term impacts, likely affecting their ability to forage for food. The body score could be determined for 61.2% (71 out of 116) of the cases in this study.
4 Conclusion
The evidence of hooks, cables, and fishing lines found on stranded sea turtle carcasses clearly demonstrates their interactions with fisheries. However, it’s important to note that these interactions are not always directly responsible for the individual turtles’ deaths. In our study, the “J” hook, primarily used for catching dolphinfish (C. hippurus), was identified as the predominant type. This particular hook is frequently employed by the Itaipava fleet, one of the most vigorously active fleets in the Southeast, where the highest incidences of sea turtle strandings involving hooks have been recorded. This pattern suggests that sea turtles in this region primarily encounter fisheries that utilize this specific hook type. Furthermore, pelagic longline fishing, especially targeting the common dolphinfish, emerges as a significant threat to sea turtle species such as L. olivacea and C. caretta within the study area. Although these interactions represent only a small fraction of the total sea turtle strandings observed during the study period, they pose a notable potential risk. The Itaipava fleet’s activities in the neritic zone, where sea turtles typically forage, combined with the turtles’ opportunistic behavior in consuming bait, suggest an increased likelihood of ongoing interactions between sea turtles and this fishing method.
Fisheries that typically target oceanic species such as swordfish (Xiphias gladius) and tuna (Thunnus spp.), which provide the most reliable identification of hooks, were poorly represented in the area and period of this study, despite the intense fishing effort in the region. The fleet is known to operate extensively and interact with juveniles of C. caretta and adults of D. coriacea in the South/Southeast regions of Brazil. The small representativeness of these hooks in the sample, compared to hooks for catching dolphinfish, may suggest that the carcasses of individuals impacted by typical oceanic longline fisheries do not reach the coast or may strand in other regions.
The high concentration of stranded turtles with J hooks targeting dolphinfish (C. hippurus) indicates that their original habitat was mainly coastal, most likely the continental shelf. Similarly, the low number of “circular” hooks, which have been mandatory in Brazil for longline fisheries since 2017, may be related to its distance from the coast and drift patterns of the carcasses. Moreover, this low number may result from the use of circle hooks as a mitigating measure that reduces the chance of ingestion and facilitates its removal from the oral region of the captured individuals. This measure minimizes the chances of finding evidence of the interaction in cases of eventual deaths that continue to occur during incidental capture.
Most turtles affected by hooking incidents were identified as large juveniles, individuals who have successfully navigated through the most vulnerable stages of their early lives. This observation underscores the critical importance of implementing protection measures for these particular life stages. Concentrating conservation efforts on large juveniles and adults can significantly increase their chances of surviving to maturity. Despite there being fewer adults impacted by hooking, their numbers could be underestimated, since strandings don’t reflect the total mortality at sea.
The high mortality rate of turtles caused by ingestion of hooks or fishing lines, and the detrimental effects such injuries cause in their digestive tracts, emphasizes the need for protective interventions to reduce bycatch. Furthermore, it’s imperative to implement proper handling procedures onboard to minimize the damage inflicted, after the turtles are captured.
The availability of up-to-date, comprehensive data on fishing efforts, including capture per unit effort for the Brazilian fleets, has been significantly affected by the suspension of the Brazilian government’s onboard observer program (PROBORDO) since 2012. Unfortunately, this program has not been reinstated, creating a substantial gap in the continuous collection of such data. In the absence of a government-led observer program, specific initiatives to collect observer data have been limited to those carried out by universities and non-governmental organizations (NGOs). However, these efforts are not ubiquitous and cover only a fraction of the fishing effort, resulting in a lack of accessible, current data on capture per unit effort across fleets. The absence of this vital data stream limits our ability to comprehensively analyze and understand the full scope of bycatch impacts, particularly in the context of stranding results. Strand monitoring, while valuable, does indeed have its limitations in capturing the complete picture of bycatch mortality, especially with respect to at-sea mortalities that do not result in carcasses washing ashore.
In conclusion, the findings of this study underscore the critical need for renewed commitment towards onboard observer programs, highlighting the significant gap in our current knowledge of fisheries’ impacts on marine biodiversity. The absence of comprehensive, up-to-date data significantly hinders our ability to accurately assess the full scale of such impacts. Therefore, it is imperative that collaborative efforts among governmental bodies, academic institutions, and non-profit organizations are strengthened and expanded. Only through such multidisciplinary and intersectoral cooperation can we hope to enhance our understanding of fisheries’ interactions with marine ecosystems and work towards more sustainable practices.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements. The Santos Basin Beach Monitoring Project (PMP-BS) is an environmental licensing requirement of the Brazilian Institute of the Environment and Renewable Natural Resources (IBAMA). The live injured or sick turtles were handled exclusively for rehabilitation purposes. No invasive procedures or manipulations were performed to generate research data. When turtles were successfully rehabilitated, they were tagged and released back into the sea. All samples used in this study were collected as part of the PMP-BS, which is licensed by the Brazilian Institute of the Environment and Renewable Natural Resources (IBAMA) under the Ministry of Environment (ABIO 640/2015 and 1169/2019). The project operates in full compliance with the Biodiversity Information and Authorization System.
Author contributions
RV: Conceptualization, Investigation, Writing – original draft, Writing – review & editing, Data curation. DG: Investigation, Writing – original draft, Supervision, Writing – review & editing, Validation. AM: Investigation, Writing – review & editing. BG: Writing – review & editing, Investigation. IB: Writing – review & editing, Data curation. RD: Writing – review & editing, Investigation. SL: Investigation, Writing – review & editing. BS: Project administration, Writing – review & editing. DR: Investigation, Writing – review & editing. HN: Writing – review & editing, Project administration. CD: Writing – review & editing. AB: Investigation, Writing – review & editing. PC: Writing – review & editing. CK: Investigation, Writing – review & editing. HC: Investigation, Methodology, Writing – review & editing, Formal analysis. JB: Conceptualization, Data curation, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was conducted under the support and funding of the “PMP-BS” (Santos Basin Beach Monitoring Project), which is an integral part of the federal environmental licensing process overseen by IBAMA (Brazilian Institute of Environment and Renewable Natural Resources). The project received funding from Petrobras for the exploration of oil and gas in the Santos Basin pre-salt pole, under the Abio number 640/2015. Petrobras was not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
Acknowledgments
The authors are grateful to the dedicated teams from various institutions involved in the Santos Basin Beach Monitoring Project (PMP-BS) for their efforts in monitoring beaches and collecting data. Special thanks to Mineral Engenharia e Ambiente, Universidade do Vale do Itajaí – UNIVALI, CTA – Serviços em Meio Ambiente, Econservation, Instituto Argonauta, GREMAR, Instituto Biopesca, Instituto de Pesquisas de Cananéia – IPEC, Universidade Federal do Paraná – UFPR; Universidade da Região De Joinville – Univille, Associação R3 Animal, Instituto Australis, and Universidade do Estado de Santa Catarina – UDESC. We also acknowledge Andrei Santo Antonio for organizing the hook collection, and Lucas Rodrigues Ferreira, Icaro Terenciano Ribeiro Gimenez, and Muriell Neves Guerra for their assistance in data research at SIMBA. The PMP-BS, part of the federal environment licensing process conducted by IBAMA for the exploration of oil and gas by Petrobras in the Santos Basin pre-salt pole, were granted permits Abio number 640/2015 with three annual renewals, and Abio number 1169/2019 with one annual renewal.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Achaval F., Marin Y. H., Barea L. C. (1998). Incidental capture of turtles with pelagic longline. Int. Symposium Sea Turtle Biol. Conserv. 18, 83–84.
Alegre F., Parga M., del Castillo C., Pont S. (2006). “Study on the long-term effect of hooks lodged in the mid-esophagus of sea turtles,” in Book of abstracts. 26th annual symposium on sea turtle biology and conservation. Eds. Frick M., Panagopoulu A., Rees A., Williams K. (International Sea Turtle Society, Athens), 234.
Almeida P. (2015). SIMBA – Sistema de Informação de Monitoramento da Biota Aquática: documento de especificação de requisitos, casos de uso, protótipos de telas e diagrama de entidade e relacionamento (Itajaí, Santa Catarina: Geosapiens), 188.
Barata P. C. R., Gallo B. M. G., dos Santos S., Azevedo V. G., Kotas J. E. (1998). “Captura acidental da tartaruga marinha Caretta caretta (Linnaeus 1758) na pesca de espinhel de superfície na ZEE brasileira e em águas internacionais,” in Resumos Expandidos da XI Semana Nacional de Oceanografia (Editora Universitária-UFPe, Pelotas), 579–581.
Benetti D. D., Brill R. W., Kraul S. A. Jr (1995). The standard metabolic rate of dolphin fish. J. Fish Biol. 46, 987–996. doi: 10.1111/j.1095-8649.1995.tb01403.x
Bezerra M., Barrios-Rodriguez C., Rezende C., López-Castro M., Lacerda L. (2024). Trophic ecology of sympatric sea turtles in the tropical Atlantic coast of Brazil. Mar. Environ. Res. 196, 106406. doi: 10.1016/j.marenvres.2024.106406
Bjorndal K. A., Bolten A. B., Lagueux C. J. (1994). Ingestion of marine debris by juvenile sea turtles in coastal Florida habitats. Mar. pollut. Bull. 28, 154–158. doi: 10.1016/0025-326X(94)90391-3
Bolten A. B. (1999). “Techniques for measuring sea turtles,” in Research and Management Techniques for the Conservation of Sea Turtles. Eds. Eckert K. L., Bjorndal K. A., Abreu-Grobois F. A., Donnelly M. (IUCN/SSC Marine Turtle Specialist Group Publication No. 4, Washington), 1–5.
Brasil (2017). “Portaria Interministerial MDIC/MMA n° 74, de 01 de novembro de 2017,” in Estabelece medidas mitigadoras para redução da captura incidental e da mortalidade de Tartarugas marinhas por embarcações pesqueiras que operam na modalidade espinhel horizontal de superfície, no mar territorial brasileiro, na Zona Econômica Exclusiva – ZEE brasileira e águas internacionais (Diário Oficial da União, Brasília), 12 pp.
Bugoni L., Neves T. S., Leite N. O. Jr., Carvalho D., Sales G., Furness R. W., et al. (2008). Potential bycatch of seabirds and turtles in hook-and-line fisheries of the Itaipava Fleet, Brazil. Fisheries Res. 90, 217–224. doi: 10.1016/j.fishres.2007.10.013
Camiñas J. A. (1998). Capturas accidentales de tortuga boba (Caretta caretta, L. 1758) en el Mediterráneo occidental en la pesquería de palangre de superficie de pez espada (Xiphias gladius). Collective Volumes Sci. Papers ICCAT 50, 705–706.
Camiñas J. A., Báez J. C., Valeiras X., Real R. (2006). Differential loggerhead by-catch and direct mortality due to surface longlines according to boat strata and gear type. Scientia Marina 70, 661–665. doi: 10.3989/scimar.2006.70n4
Camiñas J. A., de la Serna J. M. (1995). The loggerhead distribution in the western Mediterranean Sea as deduced from captures by the Spanish long-line fishery. Scientia Herpetologica, 316–323.
Cantor M., Barreto A. S., Taufer R. M., Giffoni B., Castilho P. V., Maranho A., et al. (2020). High incidence of sea turtle stranding in the southwestern Atlantic Ocean. ICES J. Mar. Sci. 77, 1864–1878. doi: 10.1093/icesjms/fsaa073
Caracappa S., Persichetti M. F., Piazza A., Caracappa G., Gentile A., Marineo S., et al. (2018). Incidental catch of loggerhead sea turtles (Caretta caretta) along the Sicilian coasts by longline fishery. Peer J. 6, e5392. doi: 10.7717/peerj.5392
Casale P., Freggi D., Rocco M. (2008). Mortality induced by drifting longline hooks and branchlines in loggerhead sea turtles, estimated through observation in captivity. Aquat. Conserv. Mar. Freshw. Ecosyst. 18, 945–954. doi: 10.1002/aqc.894
Castilhos J. C., Coelho C. A., Argolo J. F., Santos E. A. P., Marcovaldi M. A., Santos A. S., et al. (2011). Avaliação do Estado de Conservação da Tartaruga Marinha Lepidochelys olivacea (Eschscholtz 1829) no Brasil. Biodiversidade Bras. Rev. Científica 1, 28–36. doi: 10.37002/biodiversidadebrasileira.v1i1.89
Chou W.-R., Wu P.-Y., Li T.-H. (2023). Assessing the relationship between sea turtle strandings and anthropogenic impacts in Taiwan. J. Mar. Sci. Eng. 11, 1962. doi: 10.3390/jmse11101962
Crespo-Picazo J. L., Parga M., Bernaldo De Quirós Y., Monteiro D., Marco-Cabedo V., Llopis-Belenguer C., et al. (2020). Novel insights into gas embolism in sea turtles: first description in three new species. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00442
Crouse D. T., Crowder L. B., Caswell H. (1987). A stage-based population model for loggerhead sea turtles and implications for conservation. Ecology 68, 1412–1423. doi: 10.2307/1939225
Dallagnolo R., Andrade H. A. (2008). Observações a respeito da pescaria sazonal de dourado (Coryphaena hippurus) com espinhel-de-superfície no sul do Brasil. Boletim do Instituto Pesca 34, 331–335.
Díaz-Figueroa O., Mitchell M. A. (2006). “Gastrointestinal anatomy and physiology,” in Reptile medicine and surgery. Ed. Mader D. R. (Saunders Elsevier, Missouri), 145–162.
Di Bello A., Freggi D., Ciraci I., Valastro C. (2007). “Using lesions to assess the impact of fisheries on loggerhead turtles,” in Proceedings of the 27th Annual Symposium on Sea Turtle Biology and Conservation. Eds. Rees A. F., Frick M., Panagopoulou A., Williams K. (NOAA Tech Memo NMFS-SEFSC-569, Myrtle Beach), 8–9.
Di Bello A., Valastro C., Freggi D., Lai O. R., Crescenzo G., Franchini D. (2013). Surgical treatment of injuries caused by fishing gear in the intracoelomic digestive tract of sea turtles. Dis. Aquat. Organisms 106, 93–102. doi: 10.3354/dao02641
Domingo A., Sales G., Giffoni B., Miller P., Laporta M., Maurutto G. (2006). Captura incidental de tortugas marinas con palangre pelagico en el Atlantico por las flotas de Brasil y Uruguay. Collective Volume Sci. Papers ICCAT 59, 992–1002.
Fiedler F. N., Sales G., Giffoni B. B., Maçaneiro L. R., Brito M. K., Nascimento C. J., et al. (2016). Characterization and comparison of Brazilian and foreign leased pelagic longline fleets in the Southwestern Atlantic Ocean between 2003 and 2014. How different are these fisheries? Rev. CEPSUL – Biodiversidade e Conservação Marinha 5, 1–16. doi: 10.37002/revistacepsul.vol5.5991-16
Fiedler F. N., Sales G., Giffoni B. B., Port D., Sant’Ana R., Barreto A. S., et al. (2015). Spatio-temporal distribution and target species in longline fisheries off the Southeastern/Southern Brazil between 2000 and 2011. Braz. J. Oceanography 63, 407–422. doi: 10.1590/S1679-87592015090706304
Franchini D., Valastro C., Ciccarelli S., Caprio F., Lenoci D., Di Bello A. (2018). Ultrasonographic detection of ingested fishing lines in loggerheads (Caretta caretta). J. Wildlife Dis. 54, 1–11. doi: 10.7589/2017-12-302
Gama L. R., Fuentes M. M. P. B., Trevizani T. H., Pellizzari F., Lemons G. E., Seminoff J. A., et al. (2021). Trophic ecology of juvenile green turtles in the Southwestern Atlantic Ocean: insights from stable isotope analysis and niche modelling. Mar. Ecol. Prog. Ser. 678, 139–152. doi: 10.3354/meps13868
García-Párraga D., Lorenzo T., Wang T., Ortiz J.-L., Ortega J., Crespo-Picazo J.-L., et al. (2018). Deciphering function of the pulmonary arterial sphincters in loggerhead sea turtles (Caretta caretta). J. Exp. Biol. 221, jeb179820. doi: 10.1242/jeb.179820
Geraci J. R., Lounsbury V. J., Yates N. (2005). Marine Mammals Ashore: A Field Guide for Strandings (Baltimore: National Aquarium in Baltimore).
Giffoni B. B., Sales G., Consulim C. E. N., Fiedler F. N., Peppes F., Swimmer Y. (2005). “Experimento com anzol circular na ZEE brasileira e em águas adjacentes, para mitigar a captura de tartarugas marinhas na pescaria de espinhel pelágico,” in II Jornada de Conservação e Pesquisa de Tartarugas Marinhas no Atlântico Sul Ocidental (Livro de Resumos, Rio Grande), 72–75.
Giffoni B. B., Sales G., Leite N. O. Jr., Brito M., Fiedler F. N., Olavo G. (2017). Fishery as administrative unit: Implications for sea turtle conservation. Collective Volumes Sci. Papers ICCAT 73, 3252–3268.
Gilman E., Clarke S., Brothers N., Alfaro-Shigueto J., Mandelman J., Mangel J., et al. (2008). Shark interactions in pelagic longline fisheries. Mar. Policy 32, 1–18. doi: 10.1016/j.marpol.2007.05.001
Gilman E., Zollett E., Beverly S., Nakano H., Davis B., Shiode D., et al. (2006). Reducing sea turtle bycatch in pelagic longline fisheries. Fish Fisheries 7, 2–23. doi: 10.1111/j.1467-2979.2006.00196.x
Guimarães S. M., Tavares D. C., Monteiro-Neto C. (2017). Incidental capture of sea turtles by industrial bottom trawl fishery in the Tropical South-western Atlantic. J. Mar. Biol. Assoc. United Kingdom 98, 1525–1531. doi: 10.1017/S0025315417000352
Hammer O., Harper D. A. T., Ryan P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4, 1–9.
Harms C. A., Mallo K. M., Ross P. M., Segars A. (2003). Venous blood gasses and lactates of wild loggerhead sea turtles (Caretta caretta) following two capture techniques. J. Wildlife Dis. 39, 366–374. doi: 10.7589/0090-3558-39.2.366
Hart K. M., Mooreside P., Crowder L. B. (2006). Interpreting the spatio-temporal patterns of sea turtle strandings: going with the flow. Biol. Conserv. 129, 283–290. doi: 10.1016/j.biocon.2005.10.047
Hoopes L. A., Landry A. M., Stabenau E. K. (2000). Physiological effects of capturing Kemp’s ridley sea turtles, Lepidochelys kempii, in entanglement nets. Can. J. Zool. 781941–1947. doi: 10.1139/z00–140
IBGE - Fundação Instituto Brasileiro de Geografia e Estatística (1990). Divisão Regional do Brasil em Mesorregiões e Microrregiões Geográficas (Rio de Janeiro: Diretoria de Geociências).
Laporta M., Miller P., Domingo A. (2013). Incidental capture of sea turtles in the Uruguayan trawl fishery. Munibe Monogr. Nat. Ser. 1, 43–50. doi: 10.21630/mmns.2013.1.06
Laurent L., Camiñas J., Casale P., Deflorio M., De Metrio G., Kapantagakis A., et al. (1999). Assessing marine turtle bycatch in European drifting longline and trawl fisheries for identifying fishing regulations (France: European Commission Directorate-General XIV-Fisheries), 17.
Leite N. O. Jr., Stein C. E., Thomé J. C. A., Neves T. (2005). “Monitoramento da pesca de currico e espinhel de superfície na localidade de Itaipava/ES e sua interação com tartarugas e aves marinhas,” in II Jornada de Conservação e Pesquisa de Tartarugas Marinhas no Atlântico Sul Ocidental (Livro de Resumos, Rio Grande), 64–67.
Lewison R. L., Crowder L. B. (2007). Putting longline bycatch of sea turtles into perspective. Conserv. Biol. 21, 79–86. doi: 10.1111/j.1523-1739.2006.00592.x
Lewison R. L., Crowder L. B., Read A. J., Freeman S. A. (2004). Understanding impacts of fisheries bycatch on marine megafauna. Trends Ecol. Evol. 19, 598–604. doi: 10.1016/j.tree.2004.09.004
Lucchetti A., Vasapollo C., Virgili C. (2017). An interview-based approach to assess sea turtle bycatch in Italian waters. Peer J. 5, e3151. doi: 10.7717/peerj.3151
Maçaneiro L. R., Nascimento C. J., Giffoni B. B., Fiedler F. N., Mazzoleni R. (2018). Caracterização da frota de espinhel modelo Itaipava operante no porto de Itajaí, Santa Catarina, Brasil, entre 2010 e 2015. In: Jornada de Pesquisa Conservação de Tartarugas Marinhas no Atlântico Sul Ocidental 8, Rio De Janeiro. https://www.tamar.org.br/publicacoes_html/publicacoes/2018.html.
Magalhães M. S., Santos A. J. B., Silva N. B., Moura C. E. B. (2012). Anatomy of the digestive tube of sea turtles (Reptilia: Testudines). Zoologia 29, 70–76. doi: 10.1590/S1984-46702012000100008
Marcovaldi M. A., Chaloupka M. (2007). Conservation status of the loggerhead sea turtle in Brazil: an encouraging outlook. Endangered Species Res. 3, 133–143. doi: 10.3354/esr003133
Marcovaldi M. A., Sales G., Thomé J. C. A., Silva A. C. C. D., Gallo B. M. G., Lima E. H. S. M., et al. (2006). Sea turtles and fishing interactions in Brazil: Identifying and mitigating potential conflicts. Mar. Turtle Newslett. 112, 4–8.
Monteiro D. S., Estima S. C., Gandra T. B. R., Silva A. P., Bugoni L., Swimmer Y., et al. (2016). Long-term spatial and temporal patterns of sea turtle strandings in southern Brazil. Mar. Biol. 163, 247. doi: 10.1007/s00227-016-3018-4
Ninou F. A., Melcón B. G., Falces P. M. (2008). Estudio clínico de las lesiones producidas por anzuelo circular de palangre en tracto digestivo de tortugas marinas (Barcelona: Fundación CRAM), 27pp.
Orós J., Calabuig P., Deniz S. (2004). Digestive pathology of sea turtles stranded in the Canary islands between 1993 and 2001. Veterinary Rec. 155, 169–174. doi: 10.1136/vr.155.6.169
Parga M. L. (2012). Hooks and sea turtles: a veterinarian’s perspective. Bull. Mar. Sci. 88, 731–741. doi: 10.5343/bms.2011.1063
Parga M. L., Mituhasi T., Rendón L., Parrales M., Hara Y., Alegre F., et al. (2010). “Study on hook related lesions of sea turtles incidentally captured by long-line fisheries, and assessment of hook-removal techniques,” in Proceedings of the 28th Annual Symposium on Sea Turtle Biology and Conservation. Eds. Dean K., Castro M. C. L. (NOAA Technical Memorandum NMFS-SEFSC-602, Miami), 116.
Petrobras. (2019). Projeto Executivo Integrado do PMP-BS (Rio de Janeiro: Petrobras), 84pp. Available at: https://comunicabaciadesantos.petrobras.com.br/sites/default/files/Projeto_Executivo_Monitoramento_Praias_Integrado.pdf.
Petrobras. (2022). Projeto de Monitoramento de Praias da Bacia de Santos (PMP-BS). Available online at: https://comunicabaciadesantos.petrobras.com.br/programa-ambiental/projeto-de-monitoramento-de-praias-pmp.html.
Phillips B. E., Cannizzo S. A., Godfrey M. H., Stacy B. A., Harms. C. A. (2015). Exertional myopathy in a juvenile green sea turtle (Chelonia mydas) entangled in a large mesh gillnet. Case Rep. Veterinary Med. 2015, 604320. doi: 10.1155/2015/604320
Phillott A. D., Godfrey M. H. (2019). When is a stranded turtle a bycatch turtle? Assessing potential cause of stranding in sea turtles. Indian Ocean Turtle Newslett. 30, 13–17.
Polovina J. J., Balazs G. H., Howell E. A., Parker D. M., Seki M. P., Dutton P. H. (2004). Forage and migration habitat of loggerhead (Caretta caretta) and olive ridley (Lepidochelys olivacea) sea turtles in the central North Pacific Ocean. Fisheries Oceanography 13, 36–51. doi: 10.1046/j.1365-2419.2003.00270.x
Ryder C. E., Conant T. A., Schroeder B. A. (2006). Report of the workshop on marine turtle longline post-interaction mortality (Silver Spring: NOAA Technical Memorandum NMFS-F/OPR-2936pp.
Sales G., Giffoni B. B., Barata P. C. R. (2008). Incidental catch of sea turtles by the Brazilian pelagic longline fishery. Journal of the Marine Biological. Assoc. United Kingdom 88, 853–864. doi: 10.1017/S0025315408000441
Sales G., Giffoni B. B., Fiedler F. N., Azevedo V. G., Kotas J. E., Swimmer Y., et al. (2010). Circle hook effectiveness for the mitigation of sea turtle bycatch and capture of target species in a Brazilian pelagic longline fishery. Aquat. Conserv. Mar. Freshw. Ecosyst. 20, 428–436. doi: 10.1002/aqc.1106
Santos A. S., Soares L. S., Marcovaldi M. A., Monteiro D. S., Giffoni B., Almeida A. P. (2011). Avaliação do Estado de Conservação da Tartaruga Marinha Caretta caretta (Linnaeus 1758) no Brasil. Biodiversidade Bras. 1, 3–11. doi: 10.37002/biodiversidadebrasileira.v1i1.87
Schofield G., Kopsida H., Dimopoulos D., Margaritoulis D. (2000). “Necrotic limbs: amputation and treatment,” in Proceedings of the 18th International Sea Turtle Symposium. Eds. Abreu-Grobois F. A., Briseño-Dueñas R., Márques-Millán R., Sarti-Martínez L. (NOAA Technical Memorandum NMFS-SEFSC-436, Mazatlan), 274–276.
Silva A. C. C. D., Castilhos J. C., Lopez G. G., Barata P. C. R. (2007). Nesting biology and conservation of the olive ridley sea turtle (Lepidochelys olivacea) in Brazil 1991/1992 to 2002/2003. J. Mar. Biol. Assoc. United Kingdom 87, 1047–1056. doi: 10.1017/S0025315407056378
Snoddy J. E., Landon M., Blanvillain G., Southwood A. (2009). Blood biochemistry of sea turtles captured in gillnets in the Lower Cape Fear River, North Carolina, USA. J. Wildlife Manage. 73, 1394–1401. doi: 10.2193/2008-472
Soares L., Bjorndal K., Bolten A., Wayne M., Castilhos J., Weber M., et al. (2021). Reproductive output, foraging destinations, and isotopic niche of olive ridley and loggerhead sea turtles, and their hybrids, in Brazil. Endangered Species Res. Vol. 44, 237–251. doi: 10.3354/esr01095
Stabenau E. K., Heming T. A., Mitchell J. F. (1991). Respiratory, acid-base and ionic status of Kemp’s ridley sea turtles (Lepidochelys kempi) subjected to trawling. Comp. Biochem. Physiol. 99107–111. doi: 10.1016/0300-9629(91)90243-6
Stabenau E. K., Vietti K. R. N. (2003). The physiological effects of multiple forced submergences in loggerhead sea turtles (Caretta caretta). Fishery Bulletin. 101, 889–899.
Tagliolatto A. B., Goldberg D. W., Godfrey M. H., Monteiro C. N. (2019). Spatiotemporal distribution of sea turtle strandings and factors contributing to their mortality in south-eastern Brazil. Aquat. Conservation: Mar. Freshw. Ecosyst. 30, 331–350. doi: 10.1002/aqc.3244
Thomson J. A., Burkholder D., Heithaus M. R., Dill L. M. (2009). Validation of a rapid visual-assessment technique for categorizing the body condition of green turtles (Chelonia mydas) in the field. Copeia 2009, 251–255. doi: 10.1643/CE-07-227
Tomas J., Dominici A., Nannarelli S., Forni L., Badillo F. J., Raga J. A. (2001). From hook to hook: the odyssey of a loggerhead sea turtle in the Mediterranean. Mar. Turtle Newslett. 92, 13–14.
Tomás J., Gozalbes P., Raga J. A., Godley B. J. (2008). Bycatch of loggerhead sea turtles: insights from 14 years of stranding data. Endang Species Res. 5, 161–169. doi: 10.3354/esr00116
Valente A. L. S., Parga M. L., Velarde. R., Marco I., Lavin S., Alegre F., et al. (2007). Fishhook lesions in loggerhead sea turtles. J. Wildlife Dis. 43, 737–741. doi: 10.7589/0090-3558-43.4.737
Wallace B. P., DiMatteo A. D., Bolten A. B., Chaloupka M. Y., Hutchinson B. J., Abreu-Grobois F. A., et al. (2011). Global conservation priorities for marine turtles. PloS One 6, 1–14. doi: 10.1371/journal.pone.0024510
Wallace B. P., Kot C. Y., DiMatteo A. D., Lee T., Crowder L. B., Lewison R. L. (2013). Impacts of fisheries bycatch on marine turtle populations worldwide: toward conservation and research priorities. Ecosphere 4, 1–49. doi: 10.1890/ES12-00388.1
Williard A., Parga M. L., Sagarminaga R., Swimmer Y. (2015). Physiological ramifications for loggerhead turtles captured in pelagic longlines. Biol. Lett. 11 (10), 2015060. doi: 10.1098/rsbl.2015.0607
Wilson S. M., Raby G. D., Burnett N. J., Hinch S. G., Cooke S. J. (2014). Looking beyond the mortality of bycatch: sublethal effects of incidental capture on marine animals. Biol. Conserv. 171, 61–72. doi: 10.1016/j.biocon.2014.01.020
Witzell W. N. (1999). Distribution and relative abundance of sea turtles caught incidentally by the U.S. pelagic longline fleet in western North Atlantic Ocean 1992–1995. Fishery Bull. 97, 200–211.
Work T. M., Balazs G. H. (2010). Pathology and distribution of sea turtles landed as bycatch in the Hawaii-based North Pacific pelagic longline fishery. J. Wildl. Dis. 46, 422–432. doi: 10.7589/0090-3558-46.2.422
Zamboni A. (2020). Auditoria da Pesca: Brasil 2020: uma avaliação integrada da governança, da situação dos estoques e das pescarias (Brasília: Oceana Brasil), 62. Available at: https://static.poder360.com.br/2021/04/auditoria-da-pesca-brasil-2020.pdf.
Keywords: loggerhead turtle (Caretta caretta), olive ridley turtle (Lepidochelys olivacea), longline fisheries, Southwest Atlantic ocean, coastal monitoring, bycath
Citation: Vanucci RM, Goldberg DW, Maranho A, Giffoni BdB, Boaventura ICdR, Dias RB, Leonardi SB, Neto HG, Silva BMG, Rogerio DW, Domit C, Barreto AS, Castilho PV, Koleniskovas C, Chupil H and Becker JH (2024) Impacts of pelagic longline fisheries on sea turtles in the Santos Basin, Brazil. Front. Amphib. Reptile Sci. 2:1385774. doi: 10.3389/famrs.2024.1385774
Received: 13 February 2024; Accepted: 06 May 2024;
Published: 31 May 2024.
Edited by:
Gabriela M. Vélez-Rubio, University of the Republic, UruguayReviewed by:
Marco Alberto Luca Zuffi, Natural History Museum University of Pisa, ItalyJill Awkerman, United States Environmental Protection Agency (EPA), United States
Copyright © 2024 Vanucci, Goldberg, Maranho, Giffoni, Boaventura, Dias, Leonardi, Neto, Silva, Rogerio, Domit, Barreto, Castilho, Koleniskovas, Chupil and Becker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Henrique Becker, Y3VydXBpcmFAdGFtYXIub3JnLmJy