
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Agron. , 18 October 2024
Sec. Disease Management
Volume 6 - 2024 | https://doi.org/10.3389/fagro.2024.1462323
 Karoline Barbosa Pontes1†
Karoline Barbosa Pontes1† Andressa Cristina Zamboni Machado2Alison Fernando Nogueira1Daniel Fernando Viana Fagundes1
Andressa Cristina Zamboni Machado2Alison Fernando Nogueira1Daniel Fernando Viana Fagundes1 Renato Barros de Lima Filho3Mirela Mosela4Silas Mian1Débora Cristina Santiago1
Renato Barros de Lima Filho3Mirela Mosela4Silas Mian1Débora Cristina Santiago1 Leandro Simões Azeredo Gonçalves1,3*
Leandro Simões Azeredo Gonçalves1,3*Root-knot nematodes (RKNs) are considered a limiting factor for tomato production, causing substantial economic losses. Several control measures have been used to control RKNs in infested areas, such as crop rotation or succession with non-host species, using resistant cultivars, and applying chemical and biological nematicides. Here, we examined the control effect of six microbiological products (Ag109 - Bacillus velezensis, Votivo Prime - B. firmus, Quartzo - B. subtilis and B. licheniformis, Veraneio - B. amyloliquefaciens, Nemat - Purpureocillium lilacinum, and Rizotec - Pochonia chlamydosporia) against different species of Meloidogyne in tomatoes cultivated in a greenhouse and evaluated these products under field conditions to verify their impact on root-associated nematode populations and agronomic and physicochemical traits of fruits from grafted and non-grafted tomato plants. For the greenhouse studies, higher pathogenicity was observed for tomato with M. javanica and M. enterolobii compared to M. incognita. The microbiological nematicides were more effective in controlling M. javanica and M. enterolobii, while for M. incognita, the evaluated products were ineffective. Regarding the field studies, most of the microbiological nematicides evaluated effectively increased production per plant. However, when analyzing the Meloidogyne population in the field, applying microbiological nematicides showed no nematode control in plants without rootstock. On the other hand, in the grafted plants, the Ag109, Veraneio, Rizotec, and Nemat treatments boosted the reduction in the Meloidogyne population in the roots, with average reductions of 68.47, 57.97, 77.29, and 56.70%, respectively. These results indicate the effectiveness of integrated management of microbiological nematicides and resistant rootstock in tomato. Furthermore, combining these two control measures resulted in no change in the nutritional quality of tomato fruits.
Tomato (Solanum lycopersicum L.) is among the most important vegetables in the world in terms of growing area, production, commercial use, and consumption. In addition to flavor, tomato fruits have high levels of compounds beneficial to health, such as phenolic compounds (phenolic acids and flavonoids), carotenoids (lycopene and α and β carotene), vitamins (ascorbic acid and vitamin A), and glycoalkaloids (tomatine), which act as antioxidant, antimutagenic, anti-inflammatory, and antiatherogenic agents (Chaudhary et al., 2018; Salehi et al., 2019). According to the FAO (2023), the estimated world production is 187 million t with an average growth of 3.0 million t year-1 in the last ten years.
Many factors challenge tomato production. Soil-borne diseases are a crucial limiting factor due to their strong influence on productivity and the difficulty in eradicating some pathogens in cropping systems (Cheng et al., 2021; Ma et al., 2023). Among these diseases, root-knot nematodes (RKNs) are considered one of the main limiting factors for tomato production, causing huge economic losses (Barbary et al., 2015; Shilpa et al., 2022). RKNs are obligate sedentary endoparasites that, upon penetrating by the second-stage juvenile (J2), migrate to the vascular cylinder of the host roots, where they initiate a series of changes resulting in the formation of galls and the development of specialized feeding cells, known as “giant cells” (Milligan et al., 1998; Castagnone-Sereno et al., 2013). These changes reduce water and nutrient absorption by the root system, decreasing growth and crop yield (Hofmann and Grundler, 2007). Furthermore, such plant infections increase susceptibility to other soil pathogens and form disease complexes with other pathogenic soil microorganisms (Oka, 2020; Back et al., 2002).
Most species of RKNs naturally infect tomato plants. However, the predominant Meloidogyne species that infect tomato plants worldwide are M. javanica, M. incognita, M. arenaria, M. hapla and, more recently, M. enterolobii (El-Sappah et al., 2019; Philbrick et al., 2020). Several control measures have been used to control RKNs in infested areas, such as crop rotation or succession with non-host species, using resistant cultivars, and applying chemical and/or biological nematicides (Barbary et al., 2015). The dominant gene Mi–1.2, introgressed from an accession of S. peruvianum L (Smith, 1944), has been widely used as a source of resistance to RKNs, conferring resistance to different species of Meloidogyne (Gabriel et al., 2022). However, its phenotypic expression can be impaired by high soil temperatures and the occurrence of virulent populations capable of suppressing and/or overcoming the action of this gene (El-Sappah et al., 2019; Gabriel et al., 2022; Hajihassani et al., 2022). Additionally, resistance in tomato cultivars mediated by Mi genes does not control M. enterolobii (Philbrick et al., 2020). In this context, integrated measures are considered an important control strategy.
Chemical nematicides are commonly used to control RKNs. However, most of these products (e.g., aldicarb and methyl bromide) have been withdrawn from the market due to their negative impact on human health and the environment (Oka, 2020; Mwamula et al., 2022). Therefore, using biological control agents to manage RKNs has been gaining prominence, with significant growth in this market (Saritha and Prasad Tollamadugu, 2019). In Brazil, 69 microbiological nematicides were registered with the Ministry of Agriculture, Livestock, and Supply (MAPA), which are based on different species of Bacillus (B. subtilis, B. amyloliquefaciens, B. firmus, B. methilotrophicus, and B. linheniformis) and fungi, especially Pochonia chlamydosporia and Purpureocillium lilacinum.
Bacillus spp. have different mechanisms to reduce nematodes, such as i) regulation of nematode behavior, interfering with host recognition; ii) competition for nutrients; iii) plant growth promotion; iv) induced systemic resistance; and v) production of metabolites that inhibit egg hatching, reduce juvenile survival, and/or directly kill nematodes (Engelbrecht et al., 2018; Aloo et al., 2019; Dimkić et al., 2022; Bhat et al., 2023). The species P. chlamydosporia and P. lilacinum are widely studied for the control of phytonematodes, mainly acting on the parasitism of eggs and females, inducing resistance in plants and promoting plant growth (Li et al., 2015; Ahmad et al., 2021).
In light of all that has been shown, the present study aimed to evaluate the control effect of different microbiological nematicides against different species of Meloidogyne in tomato cultivated in a greenhouse and to assess these products under field conditions to verify their effects on root-associated nematode populations and agronomic and physicochemical traits of fruits from grafted and non-grafted tomato plants.
The experiments were carried out in two stages. The first experiment was implemented in September 2021 in a greenhouse located at the Universidade Estadual de Londrina (UEL) in Londrina City, Paraná, where the population control of three nematodes (M incognita, M. javanica, and M. enterolobii) was based on different microbiological nematicides in tomato. The second stage of the study was conducted in situ at two distinct field locations, precisely location A in Uraí, Paraná, Brazil (23 °C, 14’ 28” S; 50 °C, 47’ 01” W) and Location B in Rolândia, Paraná, Brazil (23 °C, 20’ 58” S; 51 °C, 27’ 04” W), both of which identified with Meloidgyne sp. infestation. The objective was to assess the efficacy of the microbiological nematicides in enhancing tomato fruit yield and modifying its physicochemical attributes. Additionally, this phase evaluated the impact of the microbiological treatments on the biocontrol of Meloidogyne sp., aiming to elucidate their potential benefits for nematode management and soil health improvement in tomato cultivation.
The experiment was carried out in a greenhouse at UEL in September 2021. Six microbiological nematicides from different companies were evaluated: Ag109 (AgBio, B. velezensis), Votivo Prime® (Basf, B. firmus), Quartzo® (FMC, B. subtilis and B. licheniformis), Veraneio® (Koppert, B. amyloliquefaciens), Nemat® (Ballagro, P. lilacinum), and Rizotec® (Stoller, P. chlamydosporia) to control three nematode species (M. incognita, M. javanica, and M. enterolobii). A separate experiment was carried out for each Meloidogyne species. The concentrations and recommendations used for the products are described in Table 1.
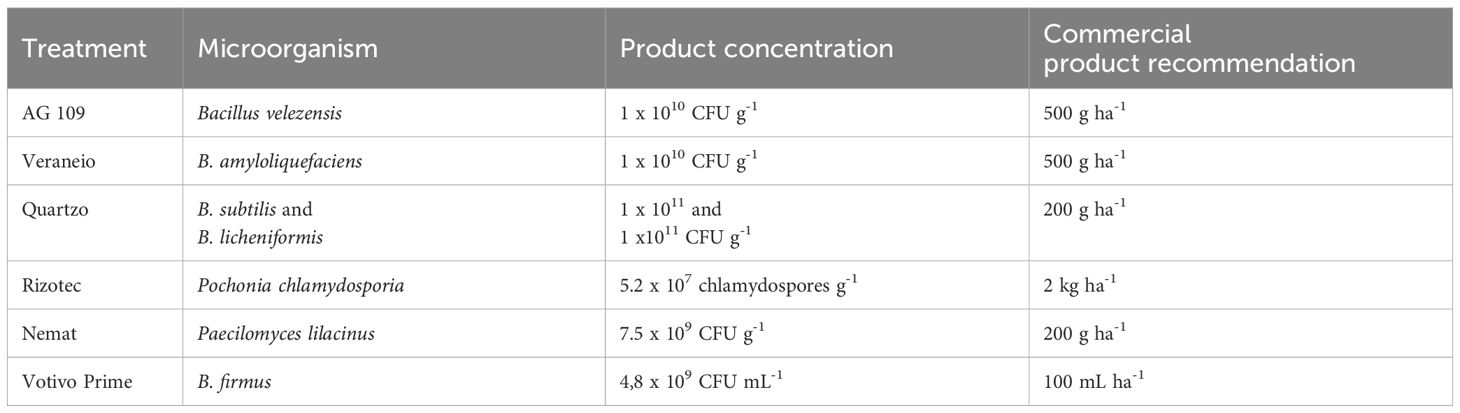
Table 1. Microbiological nematicides used to control root-knot nematodes in the tomato crop.
The experiments were conducted using a completely randomized design with ten repetitions. The experiment was implemented in 946-mL polystyrene pots containing sand and soil (7:1) previously sterilized and 3 g of Osmocote® (15% N, 9% P2O5, 12% K2O, 1% Mg, 2.3% S, 0.05% Cu, 0.45% Fe, 0.06% Mn, 0.02% Mo) per pot. Tomato seedlings of the Santa Clara cultivar were transplanted into the pots.
The inoculum of M. javanica used in the experiment comes from soybean plants from Londrina – Paraná, M. incognita from coffee plants from Altônia – Paraná, and that of M. enterolobii comes from guava plants from Carlópolis – Paraná. After purification from a single egg mass, all inoculums were multiplied in tomato plant cultivar Santa Clara in a greenhouse at the Institute of Rural Development of Paraná (IDR – Paraná), Londrina.
After transplanting the tomato seedlings, the microbiological products were applied by drenching, with 15 mL of syrup applied per pot. The volume of syrup applied was equivalent to 50 L ha-1. This treatment was repeated 30 days after the first application. The nematodes were inoculated seven days after transplanting the seedlings by pipetting 1 mL of suspension containing 1000 eggs and juveniles of M. javanica mL-1, 1000 eggs and juveniles of M. incognita mL-1, and 500 eggs of M. enterolobii mL-1 (in their respective experiments), which were extracted according to the methodology proposed by Boneti and Ferraz (1981) and quantified in a Peters chamber under a light microscope.
Forty-five days after transplanting, the plant roots were washed in running water, dried with absorbent paper, and weighed on a semi-analytical scale. Subsequently, they were processed with utmost care according to the methodology proposed by Boneti and Ferraz (1981) to extract the nematodes. Finally, these samples were quantified in a Peters chamber under a light microscope. For this experiment, the nematode reproduction factor (RF = final population of nematodes/initial population of inoculated nematodes) and the number of nematodes per gram of root were determined.
The field experiments were set up in two properties in the city of Uraí (Experiment 1) e Rolândia (Experiment 2) - Paraná, Brazil. Both properties adopted the organic farming system and have a history of losses caused by nematodes. Soil samples were collected from each location to determine the initial population of nematodes in the experimental areas. Nematode extraction was performed using the modified Baermann funnel method. The extracted nematodes were quantified in a Peters chamber under a light microscope (Eclipse Ci, Nikon).
The experiments were performed in a randomized block design, with three replications and twenty plants per plot. For each location, the following factors were analyzed: factor 1 (application of biological products) and factor 2 (grafted and non-grafted seeds on resistant rootstock). The cultivars Trinidade® (HM. Clause) and Graziani® (Sakata) were used for locations 1 and 2, respectively. For both experimental sites, the rootstock used was the resistant cultivar Woodstock® (Sakata). Ten harvests were carried out. Fruits were collected for physicochemical characterization in the fifth harvest. Microbiological nematicides were applied at one, 30, and 60 days after seedling transplantation, totaling three applications. The microbiological nematicides, concentrations, and recommendations used were the same as described above. Drench applications were carried out with 15 mL of product solution applied per plant (Figure 1). The volume of syrup applied was equivalent to 50 L ha-1.

Figure 1. Images of the experiments carried out in the municipality of Uraí e Rolândia (experiment 1 and 2, respectively), Paraná – Brazil, in the tomato crop applying different microbiological nematicides.
The characteristics evaluated were i) production (kg plant-1), ii) average fruit mass (AFM), iii) fruit length (FL), iv) fruit diameter (FD), v) fruit firmness (FIRM), vi) fruit pericarp thickness (PT), vii) total soluble solids (TSS) content, viii) reducing sugars (RS), ix) titratable acidity (TA), x) vitamin C (VITC), xi) total phenols (TP), and xii) antioxidant activity by the DPPH method (α, α-diphenyl-β-picrylhydrazyl).
FIRM was determined using a digital penetrometer with a 3 mm tip (FR-5120, Lutron, Taiwan) at two opposite points in the equatorial zone, and the results were expressed in Newtons (N). TSS was determined in a digital refractometer with automatic temperature compensation at 25°C (Atagor), according to ISO 2173, and expressed in 30 ◦Brix. RS was determined by the DNS method proposed by Maldonade et al. (2013). TA was quantified by titration with 0.1 mol L-1 sodium hydroxide (NaOH) according to the Association of Official Analytical Chemists (AOAC) method 942.15.
Vitamin C was quantified according to the standard AOAC method modified by Benassi and Antunes (1988) and expressed in mg ascorbic acid 100 g-1. The extract for quantifying total phenolic compounds and antioxidant activity was prepared from 1 g of fresh sample suspension in 10.0 mL of 70% ethanol (v/v), adapted from Vázquez et al. (2008). The quantification of total phenolic compounds was performed according to Swain and Hillis (1959) using gallic acid as an analytical standard, ranging from 10 to 100 mg L-1 (r=0.9960), expressed as equivalent mg of gallic acid 100 g-1. The antioxidant activity by the DPPH (2,2-diphenyl-1-picryl-hydrazyl) radical scavenging assay was quantified according to Brand-Williams et al. (1995). Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) was used as an analytical standard, ranging from 0.20 to 1.00 mmol L-1 (r=0.9992).
For the field experiment, roots were collected at the end of the experiment. For each treatment, six plants were collected per repetition, and the roots were removed and taken to the laboratory. Next, the roots were washed and processed according to the methodology proposed by Boneti and Ferraz (1981) for nematode extraction. Nematodes were then identified and quantified in a Peters chamber under a light microscope, calculating the number of juveniles of Meloidogyne spp. per gram of root.
Data were subjected to the assumptions of analysis of variance (normality of data, homogeneity of variances, and independence of errors). However, for the number of Meloidogyne per gram of root from the greenhouse and field experiments, these assumptions were not met; therefore, they were analyzed via non-parametric statistics. ANOVA-type statistics (ATS), which have an approximate F distribution under the null hypothesis based on asymptotic theory, were applied to the data. The Bonferroni test (p<0.05) was used to compare the treatments. For the other characteristics, analysis of variance was used using the following mathematical model: , where μ is the mean, is the effect of the k-th repetition within the j-th experiment, is the fixed effect of the i-th treatment, is the fixed effect of the j-th experiment, is the fixed effect of the interaction between treatments x experiments, and is the experimental error (random, independent, and normally distributed). Tukey’s test (p<0.05) was used to compare the treatments. Data were analyzed by the Genes program (Cruz, 2016) and the R program using the packages nparLD (Noguchi, 2011), agricolae (Mendiburu and Yaseen, 2015), and agroR (Shimizu et al., 2021).
By non-parametric ANOVA, a significant effect was observed for the sources of variation, experiments (E) and experiments x treatments (E x T), regarding reproduction factor and number of nematodes per gram of root (RF and NGR, respectively) (Table 2). For treatments, a significant effect was found only for the reproduction factor. The average RF in the experiments was 23.05, 125.74, and 130.83 for M. incognita, M. javanica, and M. enterolobii, respectively, while for NGR, it was 1790.94, 9615.08, and 3332.27, respectively.
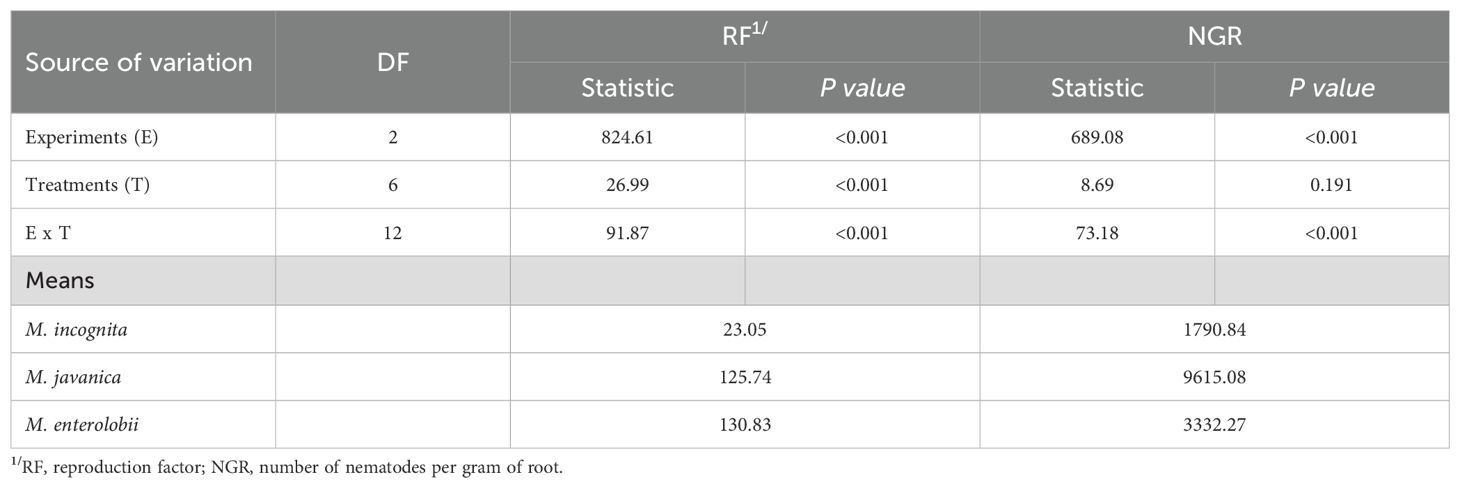
Table 2. ANOVA-type statistics and means regarding different microbiological nematicides for controlling Meloidogyne incognita, M. javanica, and M. enterolobii in a greenhouse.
For the control (inoculated, without nematicide treatment), higher RF and NGR were observed for the experiments with M. javanica and M. enterolobii than with M. incognita (Table 3). In the experiment with M. incognita, no control effect of microbiological nematicides was detected. In some cases, higher values of NGR were observed, as in the Ag109 and Nemat treatments. In the experiment with M. javanica, the lowest RF values came from the Ag109, Veraneio, and Rizotec treatments, with 53.12, 45.94, and 42.89% control percentages, respectively. For NGR, the lowest values were observed in the Ag109, Veraneio, Rizotec, and Nemat treatments, with control percentages of 40.09, 35.62, 35.62, and 20.81%, respectively. In the experiment with M. enterolobii, the lowest values of RF and NGR were obtained from the Nemat, Votivo, and Ag109 treatments; however, these treatments did not differ from the other microbiological nematicides evaluated. Control percentages ranged from 14.14 (Rizotec) to 29.49% (Nemat) for RF, while for NGR, these values varied from 18.77 (Rizotec) to 44.66% (Nemat).
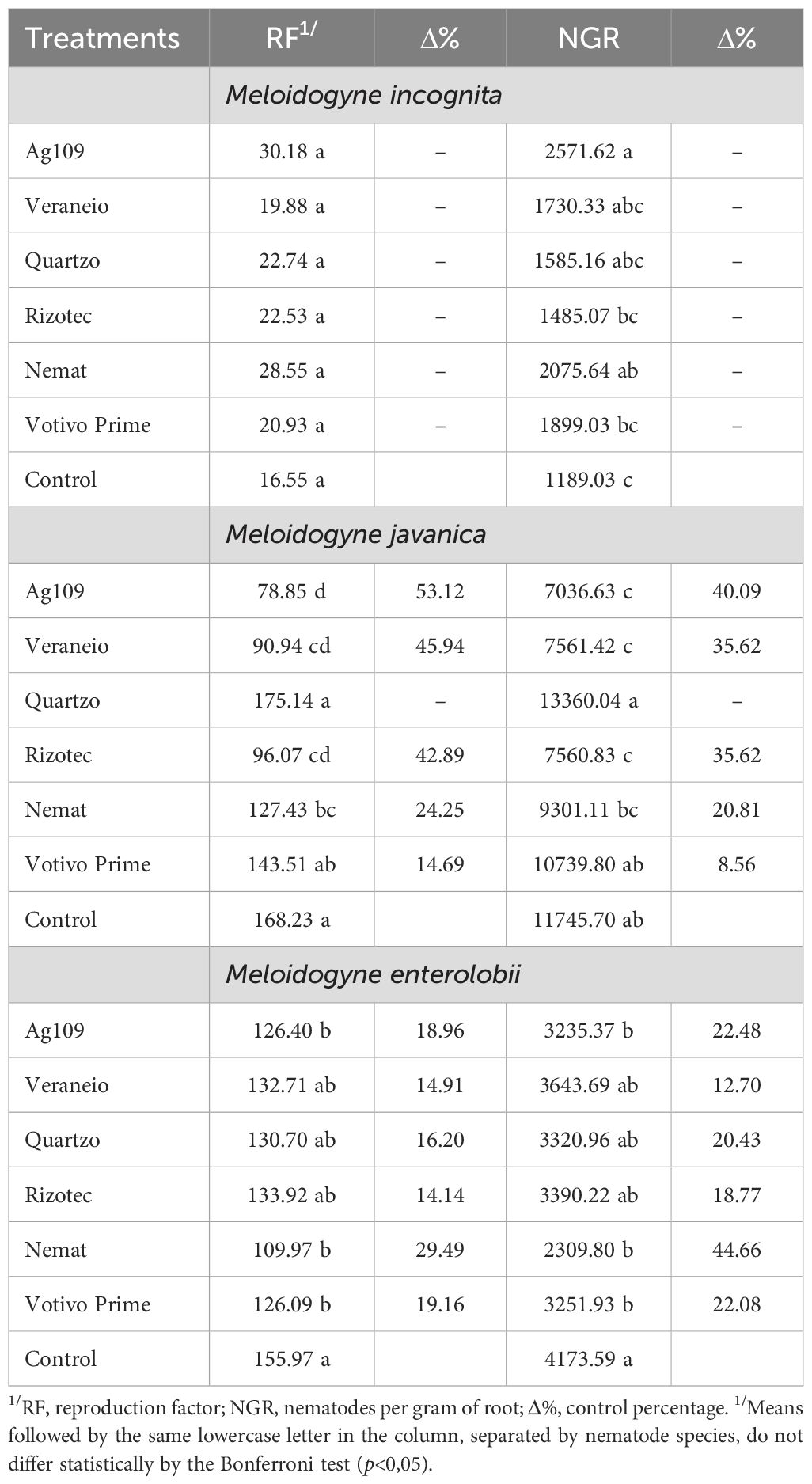
Table 3. Bonferroni test to evaluate different microbiological nematicides for controlling Meloidogyne incognita, M. javanica, and M. enterolobii in a greenhouse.
For production per plant (kg plant-1), a significant effect was observed in all sources of variation (grafting – G, treatments – T, and G x T) in experiment 1, while in experiment 2, this effect was observed only for treatments (Table 4). The coefficients of variation were 9.4 and 14.64% for experiments 1 and 2, respectively, indicating good experimental precision. In experiment 1, plants with rootstock showed a higher yield (3.94 kg ha-1) than those without rootstock (2.55 kg ha-1), representing an increase of 54.51%.
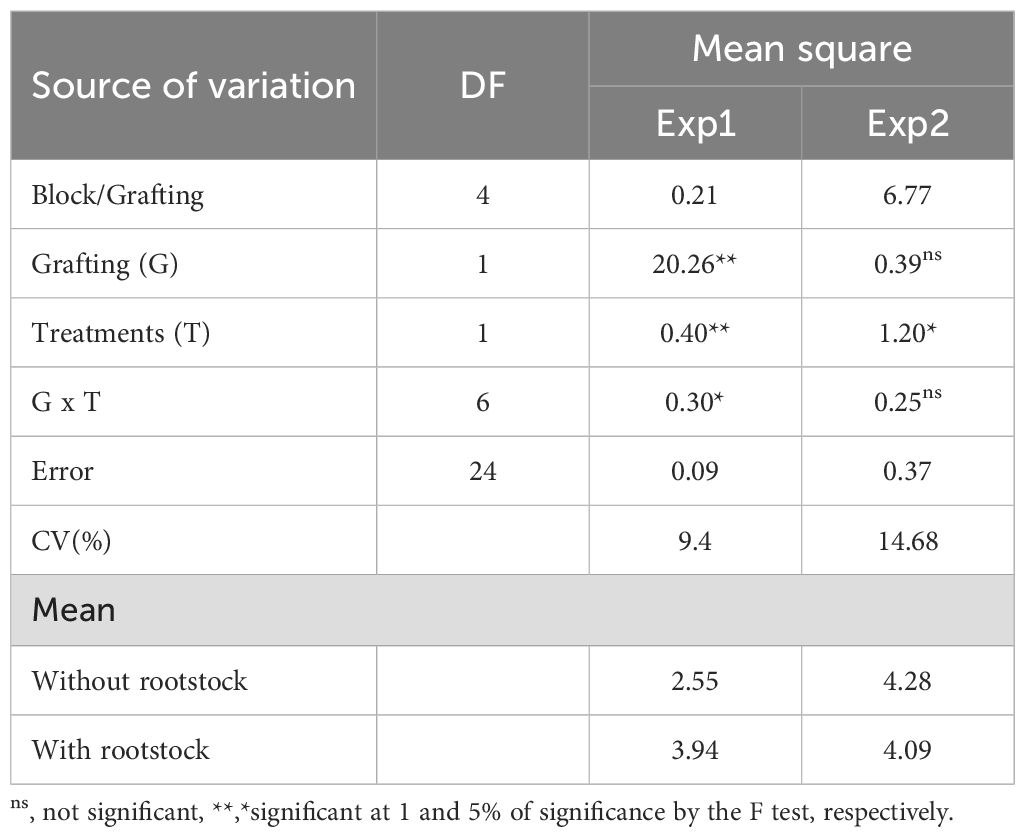
Table 4. Analysis of variance for production (kg plant-1) in tomato plants with and without resistant rootstock, treated with different microbiological nematicides, and evaluated in two locations.
In experiment 1 - without rootstock, the treatments that stood out were Ag109, Votivo, and Rizotec, with production of 3.25, 2.64, and 2.63 kg plant-1, respectively (Figure 2). These treatments provided 27.4, 3.5, and 3.1% increments in relation to the control. In the experiment with rootstock, no significant difference was found among treatments. In experiment 2, the highest production was observed in the Ag109 treatment (4.57 kg plant-1); however, it did not differ from the other microbiological treatments. The percentage of increase in relation to the control ranged from 11.7 to 28.0% in production per plant.
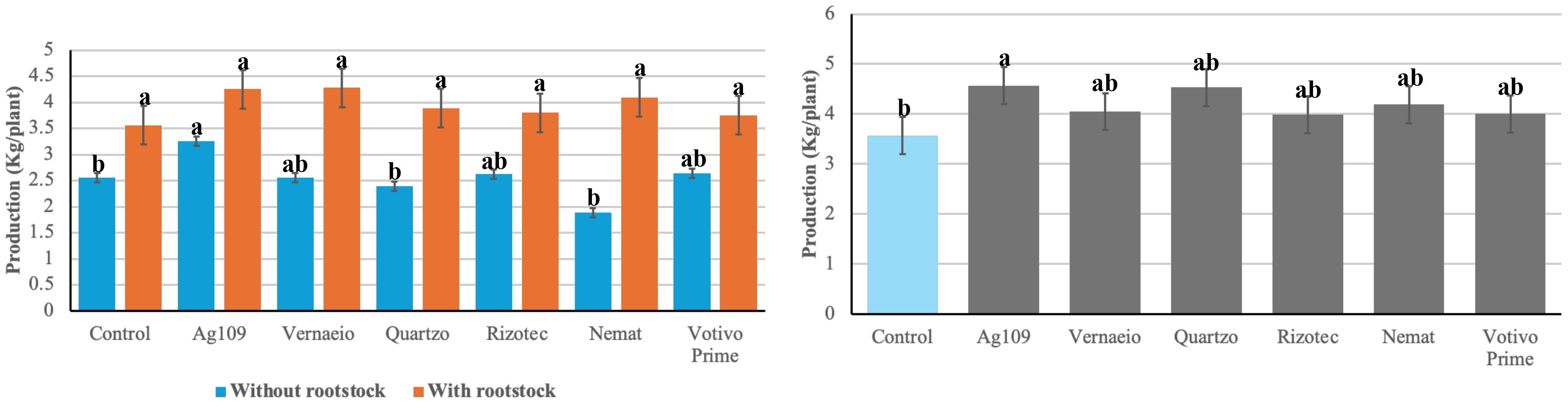
Figure 2. Comparison analysis of means for production (kg plant-1) of tomato plants with and without resistant rootstock, treated with different microbiological nematicides, and evaluated in two locations. Letters refer to separate mean comparisons for each y-axis. Means with differing letters indicate a significant difference between cutting treatments in analysis of variance (P < 0.05). Error bars represent standard deviation.
For the physicochemical traits in experiment 1, a significant effect was observed only in total phenols for grafting and DPPH for treatments (Supplementary Table S1). For total phenols, the highest values were observed in treatments without rootstock. For DPPH, no significant differences were observed between treatments by Tukey’s test (p<0.05). In experiment 2, a significant effect of fruit length (FL), total soluble solids content (TSS), total phenols (TP), and DPPH was observed for grafting, while for treatments, significant effects were observed only in DPPH (Supplementary Table S2). For the grafting x treatment interaction, only average fruit mass (AFM), fruit length (FL), and fruit diameter (FD) were significant. For FL, treatments with rootstock provided the highest values in relation to those without rootstock. In turn, for TSS, TP, and DPPH, treatments without grafting provided the highest values, especially for TP and DPPH.
For AFM and FL, the non-grafted treatments did not differ from each other by Tukey’s test (p<0.05), while for the treatments with rootstock, the highest values for these variables were observed for Rizotec, Ag109, Veraneio, Quartzo, Control, and Votivo. The highest FDs were obtained with Votivo, Quartzo, Veraneio, Control, Ag109, and Nemat for plants without rootstock. For rootstock treatments, there was no significant differentiation among them.
By non-parametric ANOVA, a significant effect of number of eggs + Meloidogyne juveniles per gram of root (MGR) was observed for grafting and grafting x treatments in both experiments (Table 5). Rootstock treatments had the lowest MGR values compared to those without rootstock, with 90.9 and 66.4% reductions in experiments 1 and 2, respectively. In experiments without rootstock, no significant difference was observed between treatments, with values varying from 267.96 (Nemat) to 523.68 (Rizotec) in experiment 1 and from 42.72 (Nemat) to 130.34 (Veraneio) in experiment 2 (Figure 3). The presence of eggs or juveniles of Meloidogyne sp. associated with roots was not detected in the treatment with Rizotec in experiment 1. In addition to Rizotec, the Ag109, Nemat, Votivo, and Veraneio treatments also differed significantly from the control treatment, presenting control percentages of 75.77, 62.08, 56.87, and 47.38%, respectively. In experiment 2, the treatments Veraneio, Ag109, Rizotec, and Nemat presented the lowest MGR values, with control percentages of 68.55, 61.17, 54.57, and 51.35%, respectively. Figures 4 and 5 show the images of roots from the two experiments with and without rootstock.
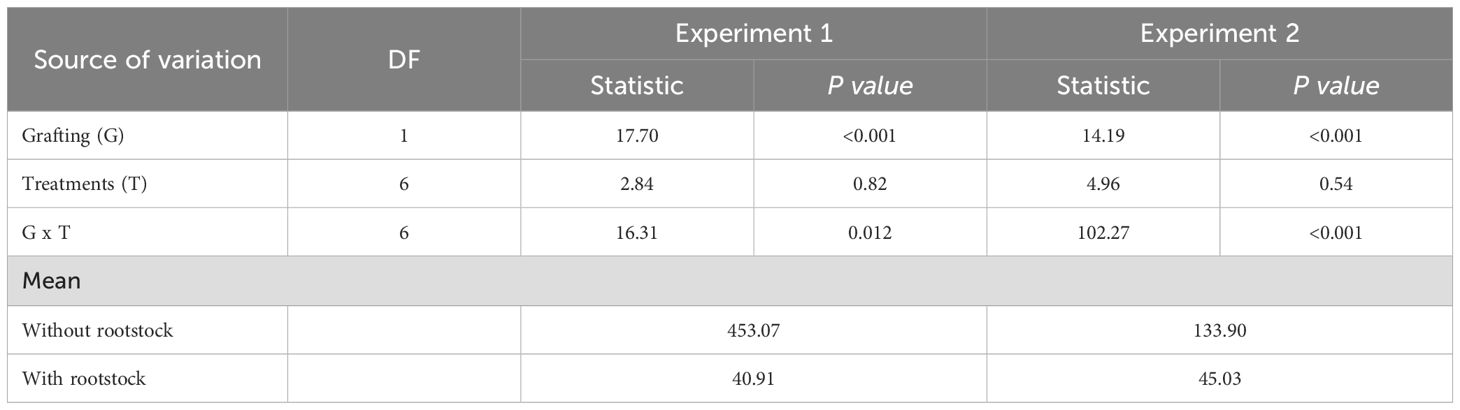
Table 5. ANOVA-type statistics for number of eggs + juveniles of Meloidogyne sp. per gram of root in tomato plants with and without resistant rootstock, treated with different microbiological nematicides, and evaluated in two locations.
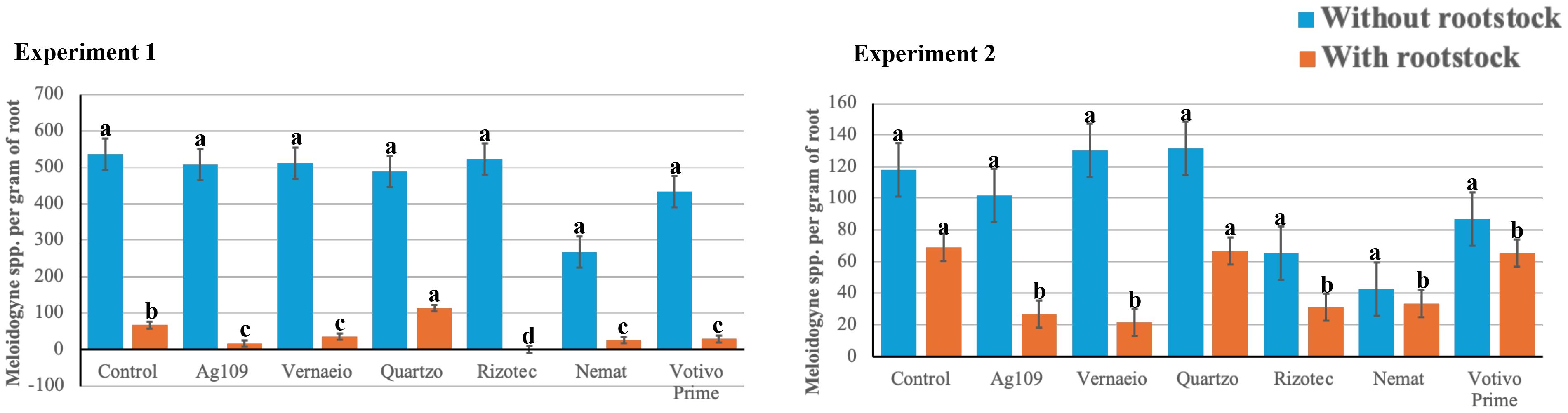
Figure 3. Number of eggs + juveniles of Meloidogyne spp. per gram of root in tomato plants with and without resistant rootstock, treated with different microbiological nematicides, and evaluated in two locations. 1/Letters refer to separate mean comparisons for each y-axis. Means with differing letters indicate a significant difference between cutting treatments in Bonferroni test (p<0.05). Error bars represent standard deviation.

Figure 4. Images of the root system of tomato plants (with and without resistant rootstock) with the application of different microbiological nematicides in experiment 1.

Figure 5. Images of the root system of tomato plants (with and without resistant rootstock) with the application of different microbiological nematicides in experiment 2.
Root-knot nematodes (RKNs) are among the most economically harmful plant parasitic nematode genera for many crops worldwide (Jones et al., 2013; Khan et al., 2023). Global damage from RKNs is estimated to cause an annual economic loss of over US$100 billion, accounting for approximately 12.6% of total crop losses (Singh et al., 2015). Therefore, several control measures have been adopted to minimize the impact of these pathogens, with biological control representing an important control measure for the integrated management of this disease (Radwan et al., 2012; Forghani and Hajihassani, 2020; Antil et al., 2023). In Brazil, several microbiological nematicides have been registered and commercialized, mainly based on Bacillus, P. chlamydosporia, and P. lilacinum species. Therefore, the present study aimed to evaluate the control effect of different microbiological nematicides for different species of Meloidogyne in tomato cultivated in a greenhouse. It also verified their effectiveness under field conditions in tomato plants with and without resistant rootstock.
In the tomato crop, different species and/or populations of the same Meloidogyne species may show different degrees of pathogenicity in a specific tomato cultivar (Seid et al., 2015). This study observed higher pathogenicity for M. javanica and M. enterolobii populations compared to M. incognita. The nematode M. enterolobii is an emerging pathogen and is considered a global threat to tomato production due to the lack of resistance in commercial cultivars and its aggressiveness (Philbrick et al., 2020; Collett et al., 2024). To date, few studies have been conducted to evaluate the efficiency of microbiological nematicides in controlling M. enterolobii under greenhouse conditions. Silva et al. (2020) evaluated two biological products (Serenade - B. subtilis, and NemOut™ - B. licheniformis, B. subtilis, and Trichoderma longibrachiatum). They observed no control effect on M. enterolobii in the tomato crop. Likewise, Almeida et al. (2022) did not observe a control effect of Trichoderma sp. strains on M. enterolobii in tomato. The best control results were obtained in the present study with the Nemat, Votivo, and Ag109 treatments.
For M. javanica and M. incognita, several studies have demonstrated the effectiveness of controlling different microbiological nematicides in tomato (Silva et al., 2017; Ghahremani et al., 2019; Girardi et al., 2022); (Krif et al., 2022); (Díaz-Manzano et al., 2023)). For M. javanica, the Ag109, Veraneio, and Rizotec treatments showed good control effectiveness. However, for M. incognita, the evaluated nematicides were not effective. Several factors may be related to this result, such as the population of M. incognita used, the environmental conditions of the experiment, the concentration of the products, and the inoculation methodology. Most of these products were registered to control M. javanica and M. incognita (agrofit.agricultura.gov.br). Redolfi (2014) found that Rizotec and Nemat products successfully controlled M. incognita in tobacco. Bontempo et al. (2017) also verified the effectiveness of Rizotec in controlling M. incognita in carrot. Therefore, further studies are needed to evaluate the efficiency of these commercial products in controlling M. incognita in tomato.
Under field conditions, most of the microbiological nematicides evaluated were effective for increasing production per plant, emphasizing the Ag109, Veraneio, Rizotec, and Votivo treatments. However, the population of Meloidogyne sp. in the field did not decrease in plants without resistant rootstock, even those treated with nematicides. Several factors may be related to this outcome, such as the time of root collection, root development, and the variation in nematode populations in the field. In the case of root collection time and development, higher root biomass was observed in some treatments with the application of microbiological nematicides about the control. Therefore, these treatments may present more nematode-feeding sites, especially at the end of the crop cycle. Several studies have demonstrated the effect of Bacillus, P. chlamydosporia, and P. lilacinum as root system growth promoters (Dallemole-Giaretta et al., 2015; Zavala‐Gonzalez et al., 2015; Baron et al., 2020). Mian et al. (2024) evaluated the Ag109 strain and observed its action as a root growth promoter in soybean. Through the genomic study of this strain, the authors identified several genes/gene groups associated with growth promotion, including volatile compounds and phytohormones.
The reduced Meloidogyne sp. population and increased production per plant in the control treatment of grafted plants about those without resistant rootstock indicates the effectiveness of this management system. Using resistant rootstock in tomato under protected cultivation systems has been widely adopted among farmers, allowing grafted plants to maintain high yields without using soil chemicals (Grieneisen et al., 2018). In the present study, the rootstock Woodstock® (Sakata seeds) was used, which, in addition to resistance to RKNs (M. javanica and M. incognita), also has genes that confer resistance to Verticillium dahliae, Fusarium oxysporum f. sp. lycopersici, Fusarium oxysporum f. sp. radicis-lycopersici, and Ralstonia solanacearum, important soil pathogens in the tomato crop.
The Ag109, Veraneio, Rizotec, and Nemat treatments boosted the reduction in the Meloidogyne population in the roots of grafted tomato, with average reductions of 68.47, 57.97, 77.29, and 56.70%, respectively, indicating the effectiveness of integrated management using microbiological nematicides and resistant rootstock in tomato. Furthermore, combining these two control measures did not change the nutritional quality of tomato fruits. Applying meta-analyses, Grieneisen et al. (2018) found that, in general, using rootstocks does not change fruit quality (pH, titratable acidity, total soluble solids, lycopene, vitamin C, firmness, and flavor).
The microbiological products associated with using Meloidogyne-resistant tomato rootstock successfully increased production and reduced the nematode population, indicating the effectiveness of integrated pest management.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
KP: Formal Analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. AM: Investigation, Methodology, Supervision, Writing – review & editing. AN: Investigation, Methodology, Writing – review & editing. DF: Investigation, Methodology, Writing – review & editing. RL: Investigation, Methodology, Writing – review & editing. MM: Investigation, Methodology, Writing – review & editing. SM: Investigation, Methodology, Writing – review & editing. DS: Investigation, Methodology, Writing – review & editing. LG: Investigation, Project administration, Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The authors thank CAPES (Coordination for the Improvement of Higher Education Personnel - financial code 01) for granting the master scholarship to the first author.
In memoriam the postgraduate student Karoline Barbosa Pontes and the farmer Keiko Mori who were the main pillars for this project.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2024.1462323/full#supplementary-material
Ahmad G., Khan A., Khan A. A., Ali A., Mohhamad H. I. (2021). Biological control: a novel strategy for the control of the plant parasitic nematodes. Antonie Van Leeuwenhoek 114, 885–912. doi: 10.1007/s10482-021-01577-9
Almeida S. F., Mello S. C. M., Cardoso A. L. B., Santos J. R. P., Carneiro R. M. D. G. (2022). Trichoderma spp. promote root growth and high populations of Meloidogyne enterolobii on tomato crop. Nematology 24, 509–520. doi: 10.1163/15685411-bja10147
Aloo B. N., Makumba B. A., Mbega E. R. (2019). The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol. Res. 219, 26–39. doi: 10.1016/j.micres.2018.10.011
Antil S., Kumar R., Pathak D. V., Kumari A. (2023). Recent advances in utilizing bacteria as biocontrol agents against plant parasitic nematodes emphasizing Meloidogyne spp. Biol. Control 183, 105244. doi: 10.1016/j.biocontrol.2023.105244
Back M. A., Haydock P. P. J., Jenkinson P. (2002). Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathol. 51, 683–697. doi: 10.1046/j.1365-3059.2002.00785.x
Barbary A., Djian-Caporalino C., Palloix A., Castagnone-Sereno P. (2015). Host genetic resistance to root-knot nematodes, Meloidogyne spp., in Solanaceae: from genes to the field. Pest Manag Sci. 71, 1591–1598. doi: 10.1002/ps.4091
Baron N. C., de Souza Pollo A., Rigobelo E. C. (2020). Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ 8, e9005. doi: 10.7717/peerj.9005
Benassi M. T., Antunes A. J. (1988). A comparison of metaphosphoric and oxalic acids as extractants solutions for the determination of vitamin C in selected vegetables. Arquivos Biol. e tecnologia 31, 507–513.
Bhat A. A., Shakeel A., Waqar S., Handoo Z. A., Khan A. A. (2023). Microbes vs. Nematodes: Insights into Biocontrol through Antagonistic Organisms to Control Root-Knot Nematodes. Plants 12, 451. doi: 10.3390/plants12030451
Boneti J. I. S., Ferraz S. (1981). Modificações no método de hussey & barker para extração de ovos de Meloidogyne exigua em raízes de cafeeiro. Fitopatol Bras. 6, 1–1.
Bontempo A. F., Lopes E. A., Fernandes R. H., Freitas L. G., Dallemole-Giaretta R. (2017). Dose-response effect of pochonia chlamydosporia against meloidogyne incognita on carrot under field conditions. Rev. Caatinga 30, 258–262. doi: 10.1590/1983-21252017v30n129rc
Brand-Williams W., Cuvelier M. E., Berset C. (1995). Use of a free radical method to evaluate antioxidant activity. LWT - Food Sci. Technol. 28, 25–30. doi: 10.1016/S0023-6438(95)80008-5
Castagnone-Sereno P., Danchin E. G. J., Perfus-Barbeoch L., Abad P. (2013). Diversity and evolution of root-knot nematodes, genus meloidogyne : new insights from the genomic era. Annu. Rev. Phytopathol. 51, 203–220. doi: 10.1146/annurev-phyto-082712-102300
Chaudhary P., Sharma A., Singh B., Nagpal A. K. (2018). Bioactivities of phytochemicals present in tomato. J. Food Sci. Technol. 55, 2833–2849. doi: 10.1007/s13197-018-3221-z
Cheng H., Zhang D., Ren L., Song Z., Li Q., Wu J., et al. (2021). Bio-activation of soil with beneficial microbes after soil fumigation reduces soil-borne pathogens and increases tomato yield. Environ. pollut. 283, 117160. doi: 10.1016/j.envpol.2021.117160
Collett R. L., Rashidifard M., Marais M., Daneel M., Fourie H. (2024). Insights into the life-cycle development of Meloidogyne enterolobii, M. incognita and M. javanica on tomato, soybean and maize. Eur. J. Plant Pathol. 168, 137–146. doi: 10.1007/s10658-023-02741-9
Cruz C. D. (2016). Genes Software - extended and integrated with the R, Matlab and Selegen. Acta Sci. Agron. 38, 547–552. doi: 10.4025/actasciagron.v38i3.32629
Dallemole-Giaretta R., Freitas L., Lopes E., Silva M., Kasuya M., Ferraz S. (2015). Pochonia chlamydosporia promotes the growth of tomato and lettuce plants. Acta Scientiarum Agronomy 37, 417–423.
Díaz-Manzano F. E., Amora D. X., Martínez-Gómez Á., Moelbak L., Escobar C. (2023). Biocontrol of Meloidogyne spp. in Solanum lycopersicum using a dual combination of Bacillus strains. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.1077062
Dimkić I., Janakiev T., Petrović M., Degrassi G., Fira D. (2022). Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms - A review. Physiol. Mol. Plant Pathol. 117, 101754. doi: 10.1016/j.pmpp.2021.101754
El-Sappah A. H. M. M. I., El-awady H. H., Yan S., Qi S, Liu J, et al. (2019). Tomato natural resistance genes in controlling the root-knot nematode. Genes (Basel) 10, 925. doi: 10.3390/genes10110925
Engelbrecht G., Horak I., Jansen van Rensburg P. J., Claassens S. (2018). Bacillus -based bionematicides: development, modes of action and commercialization. Biocontrol Sci. Technol. 28, 629–653. doi: 10.1080/09583157.2018.1469000
FAO (2023). Crops and Livestock Products. United Nation Food and Agriculture Organization. Rome: FAO.
Forghani F., Hajihassani A. (2020). Recent advances in the development of environmentally benign treatments to control root-knot nematodes. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.01125
Gabriel M., Kulczynski S. M., Santos M. F. A., Souza C. F. B., Muniz M. F. B., Boiteux L. S., et al. (2022). A novel virulent Brazilian pathotype of Meloidogyne javanica towards the tomato Mi-1.2 gene and pathogenicity to resistant rootstock. J. Plant Dis. Prot. 129, 1269–1276. doi: 10.1007/s41348-022-00618-3
Ghahremani Z., Escudero N., Saus E., Gabaldón T., Sorribas F. J. (2019). Pochonia chlamydosporia Induces Plant-Dependent Systemic Resistance to Meloidogyne incognita. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00945
Girardi N. S., Sosa A. L., Etcheverry M. G., Passone M. A. (2022). In vitro characterization bioassays of the nematophagous fungus Purpureocillium lilacinum: Evaluation on growth, extracellular enzymes, mycotoxins and survival in the surrounding agroecosystem of tomato. Fungal Biol. 126, 300–307. doi: 10.1016/j.funbio.2022.02.001
Grieneisen M. L., Aegerter B. J., Scott Stoddard C., Zhang M. (2018). Yield and fruit quality of grafted tomatoes, and their potential for soil fumigant use reduction. A meta-analysis. Agron. Sustain Dev. 38, 29. doi: 10.1007/s13593-018-0507-5
Hajihassani A., Marquez J., Woldemeskel M., Hamidi N. (2022). Identification of four populations of meloidogyne incognita in Georgia, United States, capable of parasitizing tomato-bearing mi -1.2 gene. Plant Dis. 106, 137–143. doi: 10.1094/PDIS-05-21-0902-RE
Hofmann J., Grundler F. (2007). How do nematodes get their sweets? Solute supply to sedentary plant-parasitic nematodes. Nematology 9, 451–458. doi: 10.1163/156854107781487305
Jones J. T., Haegeman A., Danchin E. G. J., Gaur H. S., Helder J., Jones M. G. K., et al. (2013). Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 14, 946–961. doi: 10.1111/mpp.12057
Khan A., Khan A., Ali A., Fatima S., Siddiqui M. A. (2023). Root-knot nematodes (Meloidogyne spp.): biology, plant-nematode interactions and their environmentally benign management strategies. Gesunde Pflanzen 75, 2187–2205. doi: 10.1007/s10343-023-00886-5
Krif G., Lahlali R., El Aissami A., Laasli S.-E., Mimouni A., Serderidis S., et al. (2022). Efficacy of authentic bio-nematicides against the root-knot nematode, Meloidogyne javanica infecting tomato under greenhouse conditions. Physiol. Mol. Plant Pathol. 118, 101803. doi: 10.1016/j.pmpp.2022.101803
Li J., Zou C., Xu J., Ji X., Niu X., Yang J., et al. (2015). Molecular mechanisms of nematode-nematophagous microbe interactions: basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 53, 67–95. doi: 10.1146/annurev-phyto-080614-120336
Ma M., Taylor P. W. J., Chen D., Vaghefi N., He J.-Z. (2023). Major soilborne pathogens of field processing tomatoes and management strategies. Microorganisms 11, 263. doi: 10.3390/microorganisms11020263
I. R. Maldonade, P. G. B. Carvalho, N. A. Ferreira (2013). Protocolo para determinação de açúcares totais em hortaliças pelo método de DNS (Brasilia: INFOTECA-E).
Mian S., Machado A. C. Z., Hoshino R. T., Mosela M., Higashi A. Y., Shimizu G. D., et al (2024). Complete genome sequence of Bacillus velezensis strain Ag109, a biocontrol agent against plant-parasitic nematodes and Sclerotinia sclerotiorum. BMC Microbiol. 24, 194.
Milligan S. B., Bodeau J., Yaghoobi J., Kaloshian I., Zabel P., Williamson V. M. (1998). The root knot nematode resistance gene mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich repeat family of plant genes. Plant Cell 10, 1307–1319. doi: 10.1105/tpc.10.8.1307
Mwamula A. O., Kabir M. F., Lee D. (2022). A review of the potency of plant extracts and compounds from key families as an alternative to synthetic nematicides: history, efficacy, and current developments. Plant Pathol. J. 38, 53–77. doi: 10.5423/PPJ.RW.12.2021.0179
Noguchi K. (2011). Package ‘nparLD’: Nonparametric analysis of longitudinal data in factorial experiments.
Oka Y. (2020). From old-generation to next-generation nematicides. Agronomy 10, 1387. doi: 10.3390/agronomy10091387
Philbrick A. N., Adhikari T. B., Louws F. J., Gorny A. M. (2020). Meloidogyne enterolobii, a major threat to tomato production: current status and future prospects for its management. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.606395
Radwan M. A., Farrag S. A. A., Abu-Elamayem M. M., Ahmed N. S. (2012). Biological control of the root-knot nematode, Meloidogyne incognita on tomato using bioproducts of microbial origin. Appl. Soil Ecol. 56, 58–62. doi: 10.1016/j.apsoil.2012.02.008
Redolfi A. (2014). Avaliação da eficiência de nematicidas biológicos à base de Pochonia chlamydosporia (Pc-10) e Purpureocillium lilacinum (Pae 10) no manejo de Meloidogyne incognita e M. javanica na cultura do tabaco (Porto Alegre, Brazil: Universidade Federal do Rio Grande do Sul).
Salehi B., Sharifi-Rad R., Sharopov F., Namiesnik J., Roointan A., Kamle M., et al. (2019). Beneficial effects and potential risks of tomato consumption for human health: An overview. Nutrition 62, 201–208. doi: 10.1016/j.nut.2019.01.012
Saritha M., Prasad Tollamadugu N. V. K. V. (2019). “The status of research and application of biofertilizers and biopesticides: global scenario,” in Recent developments in applied microbiology and biochemistry (Amsterdam: Elsevier), 195–207. doi: 10.1016/B978-0-12-816328-3.00015-5
Seid A., Fininsa C., Mekete T., Decraemer W., Wesemael W. M. L. (2015). Tomato (Solanum lycopersicum) and root-knot nematodes (Meloidogyne spp.) – a century-old battle. Nematology 17, 995–1009. doi: 10.1163/15685411-00002935
Shilpa, Sharma P., Thakur V., Sharma A., Rana R. S., Kumar P. (2022). A status-quo review on management of root knot nematode in tomato. J. Hortic. Sci. Biotechnol. 97, 403–416. doi: 10.1080/14620316.2022.2034531
Shimizu G. D., MArubayashi R. Y. P., Gonçalves L. S. A. (2021). AgroR: experimental statistics and graphics for agricultural sciences.
Silva J., Santana M. V., Freire L. L., Ferreira B., da Rocha M. R. (2017). Biocontrol agents in the management of Meloidogyne incognita in tomato. Ciec. Rural 47, e20161053. doi: 10.1590/0103-8478cr20161053
Silva J. O., Santana M. V., Carneiro F. A., Rocha M. R. (2020). Reaction of tomato genotypes to Meloidogyne enterolobii and effects of resistance inducing products. Nematology. 22, 213–220. doi: 10.1163/15685411-00003301
Singh S., Singh B., Singh A. P. (2015). Nematodes: A threat to sustainability of agriculture. Proc. Environ. Sci. 29, 215–216. doi: 10.1016/j.proenv.2015.07.270
Swain T., Hillis W. E. (1959). The phenolic constituents ofPrunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 10, 63–68. doi: 10.1002/jsfa.2740100110
Vázquez G., Fontenla E., Santos J., Freire M. S., González-Álvarez J., Antorrena G. (2008). Antioxidant activity and phenolic content of chestnut (Castanea sativa) shell and eucalyptus (Eucalyptus globulus) bark extracts. Ind. Crops Prod 28, 279–285. doi: 10.1016/j.indcrop.2008.03.003
Keywords: Solanum lycopersicum L., Meloidogyne, biological control, integrated pest management, bionematicide
Citation: Pontes KB, Machado ACZ, Nogueira AF, Fagundes DFV, de Lima Filho RB, Mosela M, Mian S, Santiago DC and Gonçalves LSA (2024) Efficacy of microbiological nematicides in controlling root-knot nematodes in tomato. Front. Agron. 6:1462323. doi: 10.3389/fagro.2024.1462323
Received: 09 July 2024; Accepted: 25 September 2024;
Published: 18 October 2024.
Edited by:
David Ezra, Agricultural Research Organization (ARO), IsraelReviewed by:
Tahir Sheikh, Sher-e-Kashmir University of Agricultural Sciences and Technology of Kashmir, IndiaCopyright © 2024 Pontes, Machado, Nogueira, Fagundes, de Lima Filho, Mosela, Mian, Santiago and Gonçalves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leandro Simões Azeredo Gonçalves, bGVhbmRyb3NhZ0B1ZWwuYnI=
†Deceased
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.