 Rahel Sutterlütti
Rahel Sutterlütti Isabell König1
Isabell König1 Martin Jansen
Martin Jansen- 1Soil Science of Temperate Ecosystems, Busgen-Institute, Georg-August University of Göttingen, Göttingen, Germany
- 2Division of Grassland Science/Department of Crop Sciences, Georg-August-University of Göttingen, Göttingen, Germany
- 3Federal Research Center for Cultivated Plants, Julius Kühn-Institut (JKI), Institute for Crop and Soil Science, Braunschweig, Germany
Alley cropping systems are known as more sustainable land use alternatives compared to monoculture cropland. In addition to the improvement of above-ground structures and creation of biotopes relevant to nature conservation, the improvement of resource development through various root systems plays a major role. We studied the interaction of the root systems in an alley cropping system combining permanent grassland and willows and at a reference grassland site. The system was established 12 years prior to our study on a site with a shallow groundwater table at 130 cm depth. We measured carbon stocks in the topsoil and determined the share of root-bound carbon relative to the total carbon pool and extracted soil cores up to a depth of 150 cm along a distance gradient to the trees and at a reference grassland site with no tree influence. The maximal rooting depth of the grassland roots increased with increasing distance to the tree lines and total root biomass was higher than under the grassland reference up to a distance of 2.5 m from the tree line. Tree roots extended up to a distance of 5.5 m from the trees and we could distinguish zones of tree root dominance very close to the trees, zones of grassland root dominance at distances 8.5 m and an interaction zone in between those two extremes. We conclude that alley cropping increases belowground biomass as compared to grassland and has therefore a higher potential to store carbon in the subsoil.
1 Introduction
Alley cropping systems are considered a more sustainable land use alternative compared to monoculture cropland to increase carbon storage and ameliorate climate change (Quinkenstein et al., 2009). They combine woody perennials arranged in alleys on the same area as agricultural crops or grassland. Fast growing tree species were shown to increase soil organic carbon in the tree line (Tsonkova et al., 2011) and improve soil physical properties (Kahle and Janssen, 2020). Soil water availability for crops is likely increased by hydraulic lift (Richards and Caldwell, 1987), as shown by Gerjets et al. (2021) in a greenhouse experiment. Crops are likely to benefit from hydraulic lift in a zone where both tree and crop roots are present.
Planting rows of trees on grassland creates a zone of competition for resources between both systems (Jose et al., 2004). Interactions of root systems of woody perennials and agricultural crops in alley cropping are not yet completely understood, but documented e.g. by Mulia and Dupraz (2006) who found lower fine root densities (FRD) of trees in the upper soil horizons when annual crops were present. Fine root density relates the root mass to the volume of soil. Plant water and nutrient uptake is largely proportional to the surface area of fine roots (≤2 mm) (Coners and Leuschner, 2005) and therefore, quantifying root parameters is crucial to estimate below-ground interactions between the components of mixed cropping systems. The diameter-based definition does not consider functionality or root order and therefore limits our understanding of root processes (McCormack et al., 2015). Hajek et al. (2014) interpret higher fine root densities as dominance and advantage in below-ground competition between two poplar species. However, in a system where root architecture and especially the fine root diameter differs widely, using the root mass to estimate dominance might lead to the wrong conclusions. Fine root length density (FRLD) relates the total length of the roots to the volume of soil and is therefore less sensitive to differences in root mass. Therefore, we used fine root length density as a direct proxy of surface area after Weemstra et al. (2020) and estimate the interactions of two competitive root systems based on this parameter.
The subsoil is an important carbon sink with up to 50% of soil organic carbon (SOC) stored at depths >30 cm (Antony et al., 2022). New SOC in the subsoil is mainly added by the root system, whereas in more shallow soil horizons, both above- and belowground plant tissue contributes to soil carbon storage (Angst et al., 2016). An increased rooting depth potentially increases the depth at which C can be stored in the soil, but the process of root C sequestration is still poorly understood (Rumpel and Kögel-Knabner, 2011). The subsoil C storage capacity of alley cropping systems is not directly related to the fine root biomasses found in the system. Cardinael et al. (2015) found increased SOC under trees, but no effect of distance to the trees in a Mediterranean alley cropping system and Angst et al. (2016) report the same for a forest site. Similarly, Siegwart et al. (2023) found correlations between root C stocks and SOC for depths of up to 50 cm, but not for the subsoil. Direct predictions on root C input in the subsoil based on measurements of FRD are not possible with the present state of knowledge on the stability and turnover times of the C pools.
We conducted a root sampling study to understand the spatial dynamics between tree and grassland roots in an alley cropping system. We examined the influence of distance to the tree line on maximum rooting depth of grassland fine roots, grassland and total fine root density and fine root length density and determined the lateral extent of the tree root system below the grassland. We furthermore quantified root C stocks in absolute terms and relative to the C stock present in the topsoil. We hypothesized that total fine root biomass decreased with increasing distance to the tree line and that the maximum rooting depth and fine root density of grassland plants increased with increasing distance to the trees. Based on the fine root densities of trees and grassland species, we delineated zones of tree- and grassland root dominance and a zone where both root systems are present with similar densities (mixing zone).
2 Materials and methods
The study was conducted at an alley cropping site combining osier willow clones (Salix schwerinii x Salix viminalis) and permanent grassland close to Hannover (52°33’51”N 9°27’50”E). Trees were planted in alleys with a tree density of 15000 trees ha-1. The tree lines consisted of double rows (0.75 m distance to the next row) with a distance of 0.75 m between trees of one row. Spacing between double rows was 2 m and the total width of the tree line was 11 m. The width of the permanent grassland between the trees was 50 m at the experimental area (white rectangle, Figure S1). The willows were planted in 2008 and harvested the first time in winter 2015/16. Tree height measurements took place in spring 2019,when the trees had an average height of 5.3 m (personal communication Langhof, Julius-Kühn-Institut, Institute for Crop and Soil Science). The soil has not been ploughed since the onset of the study 2008, when the tree lines were established. It was formed from alluvial sand under a peat layer and the groundwater level was at an average of -130 ± 10 cm between July and December 2020. The organic layer had an average thickness of 47 ± 4 cm across the study site and the soil was described as Dystric Folic Histosol (Table 1) according to the World Reference Base for Soil Resources (IUSS working group WRB, 2015). The most dominant species in the grassland were Arrhenatherum elatius, Holcus lanatus, Ranunculus repens and Rumex acetosa.
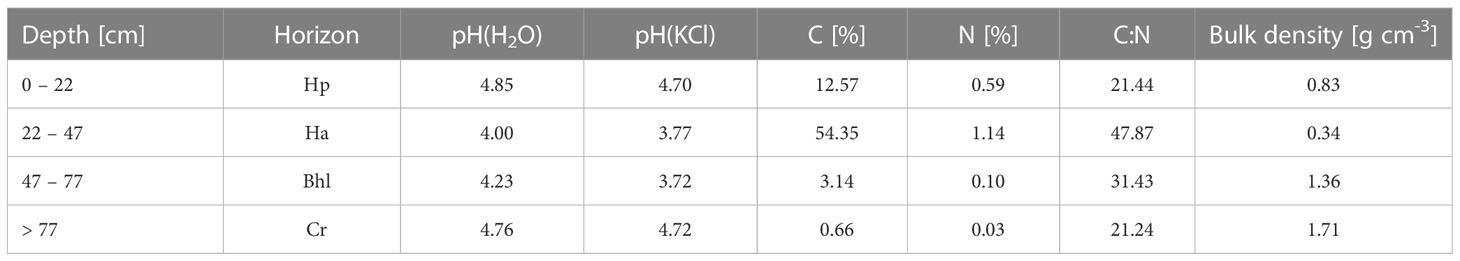
Table 1 Chemical properties (pH in water and KCl, carbon (C) and nitrogen (N)-content and C:N ratio) in the horizons with given bulk density.
Soil sampling was conducted in the year before root sampling with a Pürckhauer soil corer to a depth of 20 cm to quantify the carbon (C) contents in the topsoil in two intervals (0-10 cm and 10-20 cm). Samples were taken at distances of 0 m, 1.5 m, 5.5 m, 8.5 m and 25 m from the tree line with six replications at the locations indicated as orange transects in Figure S1 and the C contents were determined in the vario EL cube elemental analyzer (Elementar Analysensysteme GmbH, Langenselbold, Germany). C stocks were calculated for each interval. Soil cores for root sampling were taken in the end of June 2020 using a hydraulic soil corer with a maximum depth of 150 cm and a diameter of 6 cm at 7 distances to the tree line (0 m, 1.5 m, 2.5 m, 3.5 m, 4.5 m, 5.5 m, 8.5 m and 25 m) and at a reference site at > 30 m distance to the trees with three repetitions, resulting in a total of 27 cores. Transects for soil coring were located at the Northern and Southern edge of the experimental area (white rectangle) and one transect was established equidistantly (distance 22 m to the next transect) in between to avoid spatial auto-correlation between transects (Figure S1). Soil cores were divided into intervals of 10 cm up to 1 m and in 25 cm intervals between 1 m and 1.5 m depth, resulting in 12 depth intervals per core. Roots were washed manually and dead roots were excluded from the analysis. Live fine roots (diameter 2 mm) were separated into grassland- and tree roots based on morphological features to determine the maximum rooting depth of the grassland plants. All grassland roots showed diameters 2 mm and were treated as fine roots. Roots were scanned using the WinRhizo software (Regent Instruments) and root length densities (cm cm-3) were calculated. The dry weight of each sample was determined after drying at 60°C for 48 h in a forced-air oven and fine root densities (g m-3) were calculated as
and
Carbon content in roots was determined by C/N analysis at the Thünen Institute of Climate-Smart Agriculture. The C content of tree coarse and fine roots was determined and grass root samples were pooled to obtain enough biomass for analysis based on their distance to the tree line ( 3.5 m; 4.5 m and 5.5 m; 8.5 m and 25 m and reference site) to determine possible tree effects on C content.
The effect of distance to the trees on the maximum rooting depth of grassland plants, fine root densities of grassland and on the total fine root densities was evaluated by Kruskal-Wallis tests in the statistical software R (R core team, version 4.0.2). In case of significant effects, distances were compared with a Wilcoxon rank sum test.
The C stocks in coarse and fine tree roots and grassland roots along the profile were calculated based on the C contents and root biomasses. For the uppermost 20 cm, we calculated the share of root-bound carbon relative to the total carbon stock of our soil. We evaluated the effect of distance to the tree line on the carbon stock integrated over depth (kg C m-2) with a Kruskal-Wallis test and Wilcoxon rank sum tests in case of significant differences between distances as described above.
3 Results
The grassland root distribution was significantly (p = 0.019) influenced by the distance to the trees with a lower rooting depth close to the trees (Figure 1). Grassland maximum rooting depth differed significantly (p 0.05) between distances 4.5 m and 8.5 m. The grassland fine root density was significantly (p = 0.031) influenced by the distance to the trees, with lower grassland fine root densities at distances 3.5 m (p 0.05).

Figure 1 Mean fine root densities [g m-3] in depth intervals of 10 cm (0-100 cm) and 25 cm (100 – 150 cm) of trees and grassland species at different distances from the tree line and at a grassland reference site. Error bars represent minimum and maximum encountered fine root densities.
We showed that the lateral extension of tree roots was at least 5.5 m from the tree line and likely lower than 8.5 m. Corresponding to the lateral extension of the tree roots, we found that distance to the trees significantly (p = 0.026) influenced the total fine root density. The total fine root density was significantly (p = 0.05) higher at distances 2.5 m due to the presence of tree roots compared to the grassland reference.
If we interpret FRD as proxy for root dominance as in Hajek et al. (2014) of one functional group (trees or grassland) and define dominance as 66% of fine root biomass, we observed complementarity of root systems with areas with a clear spatial (vertical as well as horizontal) separation of the soil in tree- or grassland-dominated areas (Figure 2, top). The tree roots dominated in the uppermost 10 cm only up to 1.5 m distance from the tree line. In contrast, in the subsoil from about 40 cm soil depth, the trees opened up a much larger area up to 5.5 m. The dominance of the grass roots was limited to the top 10 cm in the range 2.5 to 4.5 m, while without the influence of the trees the grass rooting reaches 80 cm. The intermixing zone, with approximately equal proportions of roots of both trees and grassland species, was restricted to the 10-30 cm soil depth-range at 2.5 to 5.5 m intervals. The root systems tended to overlap (with similar proportions of fine root biomass) more frequently at larger distances to the tree line. The zones of co-dominance were comparatively small.
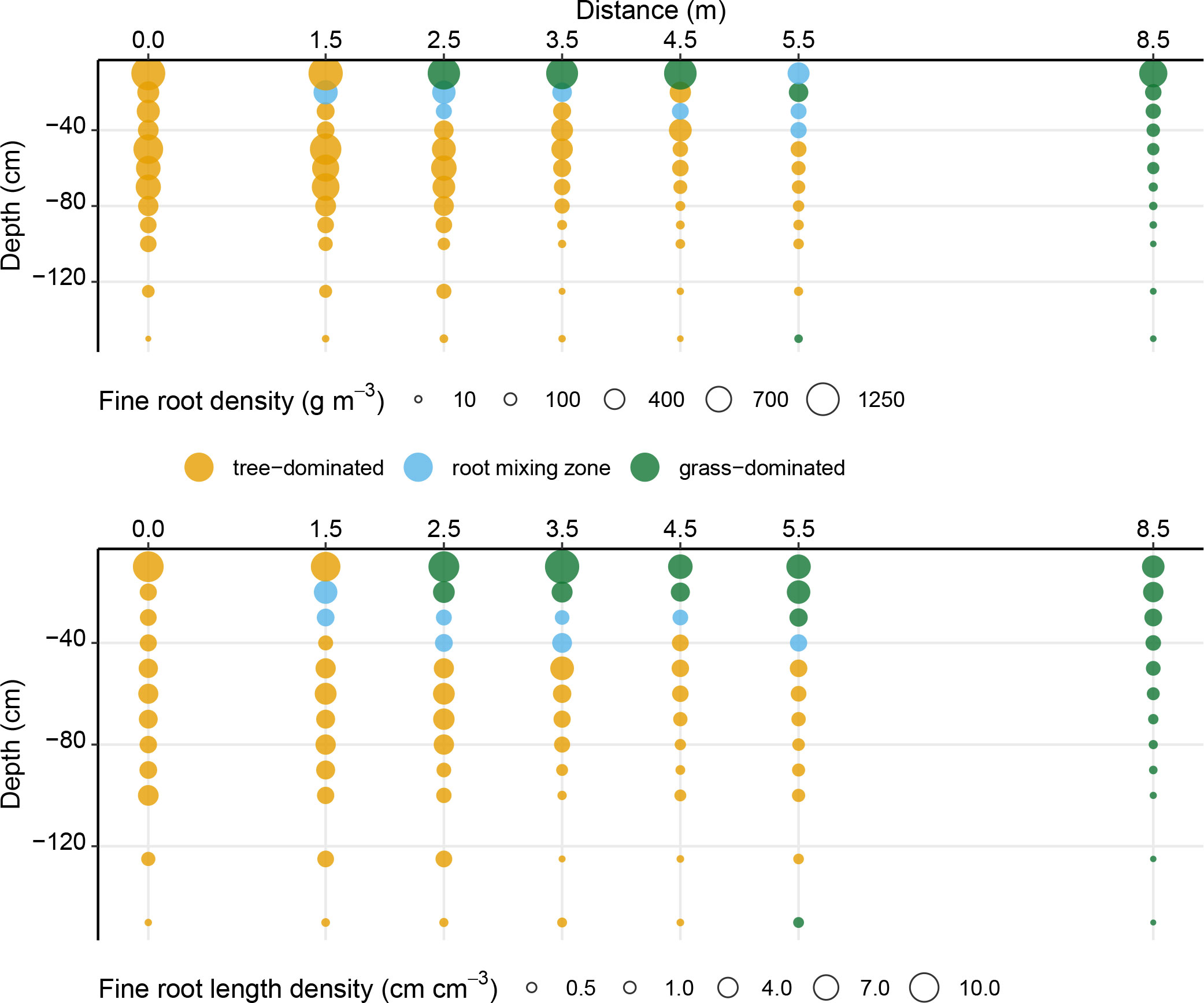
Figure 2 Group depending root dominance ( 66% of fine root densities [g m-3]) (top) and fine root length densities [cm cm-3] (bottom) in distance from tree line in an agroforestry system. Dots are scaled according to fine root density (top) and fine root length density (bottom).
The results regarding root dominance when evaluated based on FRLD were similar to the evaluation of dominance based on FRD. The zone of grassland dominance increased to depths of 20 cm for distances 1.5 m up to 4.5 m. At 5.5 m distance the dominance of grassland plants extended up to 30 cm. Furthermore, we observed that the areas of co-dominance are shifted to greater depths (Figure 2, bottom).
The soil at the present site is extremely rich in C (Table S1) because of post-glacial peatland growth. The background C contents are much too high to reflect the recent land use change by introducing the alley cropping system and we did not find a significant influence of the tree line on C contents in the topsoil (data not shown). The size of the C pool of the roots relative to the C pool of the soil is displayed in Figure 3. The tree roots account for a higher, but still very low percentage of carbon compared to the grassland. The total C stock integrated over the profile (compare Figure S2 for root C stocks along the profile) was significantly lower at distances 8.5 m compared to the C stock in the tree line. On the other hand, the C stock of the grassland reference was significantly lower than the C stock of the roots from distances 3.5 m.
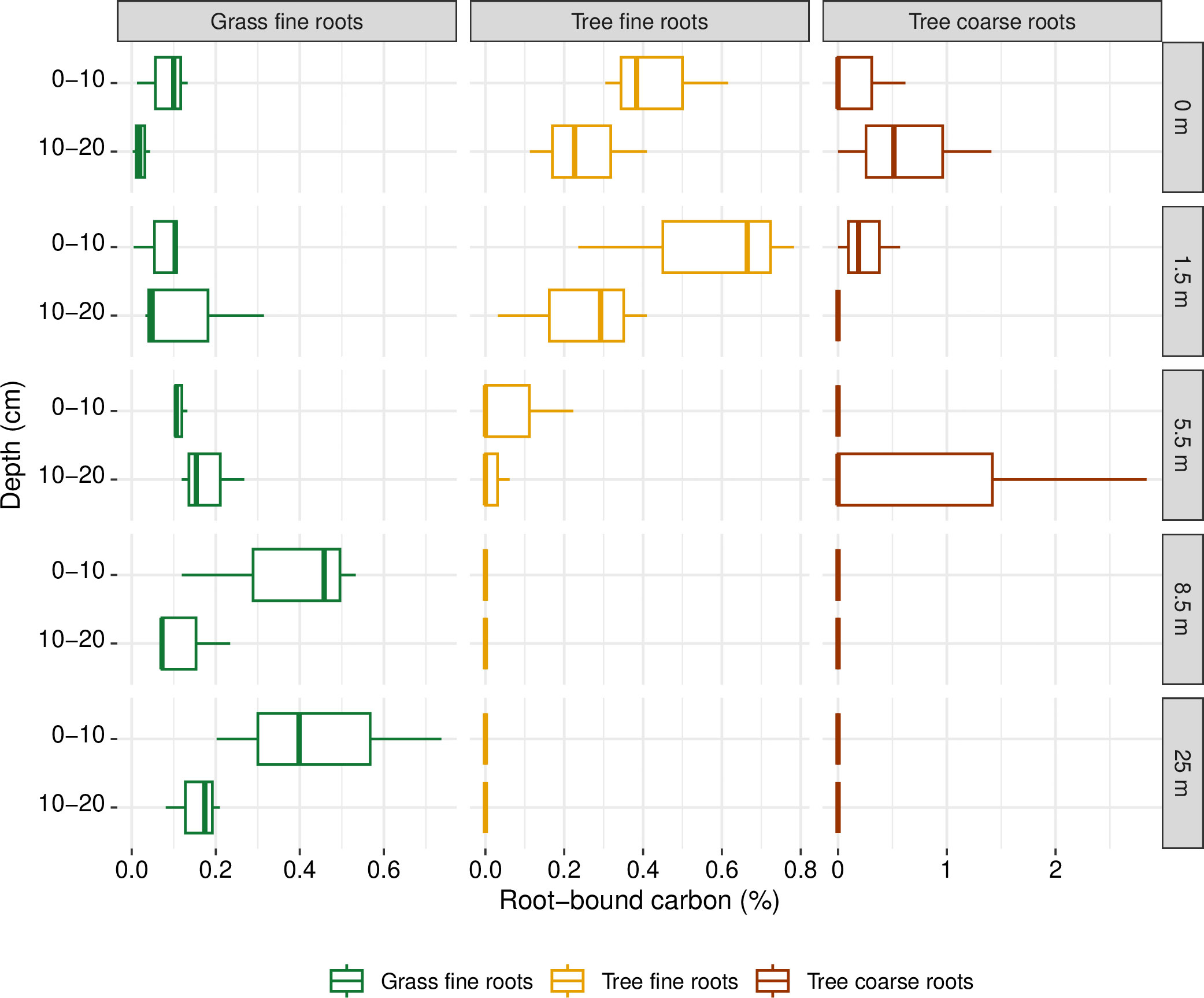
Figure 3 Percentage of root-bound carbon for grassland fine roots and tree fine and coarse roots relative to the total carbon stock of the topsoil in depths of 0-10 cm and 10-20 cm. Note the different scales of the x-axes to increase visibility.
4 Discussion
We found that the rooting depth of grassland species is affected by the distance to the tree line. The lower rooting depth of grassland species up to a distance of 4.5 m from the tree line is likely attributable to the interaction between the root systems also documented by Mulia and Dupraz (2006). The effects seem to depend heavily on the tree species as they found that wheat roots exhibit a lower rooting depth close to walnut trees, while the effect was much lower close to poplars. Especially the second maximum of the tree roots below the maximum grassland rooting depth (Figure 1) demands attention. The vertical displacement of tree roots was previously reported for walnut and poplar trees grown with an intercrop and with tillage up to 20 cm depth (Mulia and Dupraz, 2006). Here we show that the vertical displacement of tree roots in alley cropping systems might be independent of tillage and occur frequently in alley cropping systems. However, we cannot exclude an influence of the heterogeneous soil on this site. The soil conditions did not influence the grassland root biomass that declined exponentially with depth. Surprisingly, the second maximum of tree roots occurred below the organic horizon in a zone with more acid pH (Table 1) and less water storage capacity. According to the functional equilibrium hypothesis Poorter et al. (2012), plants increase their aboveground biomass relative to the belowground biomass in case of aboveground competition (e.g. by shading). Data not shown reported that aboveground biomasses are largely unaffected by the tree line. With equal aboveground biomass irrespective of distance to the trees, the belowground biomass is expected to be lower close to the trees to match the shift in aboveground relative to belowground biomass. The observed lower grassland belowground biomasses close to the trees up to a distance of 3.5 m correspond well to the functional equilibrium hypothesis.
Even though the root systems were very heterogeneous as reported by Mulia and Dupraz (2006), the lateral extent of the tree roots was similar to values found by Plante et al. (2014) for willow trees where the root distribution was studied in trench-profiles. This study did not detect a second rooting maximum even though the willows were grown with annual cereals. The authors found an influence of soil texture with higher fine root densities on lighter-textured soils. We therefore conclude that under the given circumstances with a shallow water table at 130 cm depth and a relatively coarse-grained substrate, willow roots extend laterally not more than 8.5 m.
When grown in mixtures, root systems tend to form patterns of spatial dominance (Leuschner et al., 2001). If we consider a root mass > 66% as dominant, a quite differentiated picture emerges. Our results are in accordance with research on tree species’ reactions to interspecific root competition as well as with research on competition effects on root distributions of grassland plants: under higher competitive pressure, they tend to increase root aggregation (Lepik et al., 2021), and consequently spatial separation of root systems. Even though the root systems of permanent grassland and trees overlapped in the mixing zone, the absolute root mass was not increased, but was within the range of tree- or grassroot-dominated areas.
Both fine root density and fine root length density are positively correlated with the root surface area and therefore with a plant’s ability to take up water. Given the large difference in radius between tree and grassland fine roots, we might overestimate tree water uptake ability if we only considered FRD. Therefore, we compared fine root length densities as proxies for dominance regarding plant water uptake. We find the biggest difference between the two parameters at 5.5 m from the tree line, where trees and grassland species are co-dominant on a root mass-, but not on a root length basis. Since we did not consider root functionality but only diameter, the root length is a better indicator of root function than the weight. The deviation between FRD and FRLD of tree roots is especially high in the peat layer. Tree roots in this area fall into our definition of fine roots, but without determining the root function as summarized by McCormack et al. (2015), we cannot determine if they absorb water (absorptive fine roots) or grow through the peat as transport fine roots. Hajek et al. (2014) considered only fine roots of the first root order as absorptive fine roots. Differences between the mass- and length-based dominance metrics might be of importance when modelling the interactions between the root systems, especially for root water and nutrient uptake. Grassland roots are most likely to profit from hydraulic lift in zones with similar root length densities of trees and grassland roots. The FRLD in the zones of co-dominance are consistently low (Figure 2, bottom). We estimate that the zone in which grassland roots benefit from hydraulic lift is largely limited to the zone of root co-dominance.
Alley cropping is often promoted as opportunity to increase above- and belowground C storage compared to agriculture (Drexler et al., 2021). For grasslands, the reviewed literature did not confirm significantly increased soil organic carbon (SOC) compared to alley cropping. In this meta-analysis, the average sampling depth for studies with a grassland control was 15.5 cm. Our results showed that even on a soil where the root zone is confined by ground water, this sampling depth is not sufficient to capture the C input by tree roots below the grassland roots. Choosing a low sampling depth will lead to an underestimation of below-ground C storage by tree roots (Figure S1). On the other hand, the C turnover in the subsoil is poorly understood and Siegwart et al. (2023) did not find a positive correlation between root C stocks and SOC for depths > 50 cm. Our results showed that the total C stock in the root system was higher up to a distance of 3.5 m compared to the grassland reference. Therefore, we assume that SOC stocks in the vicinity of the trees under alley cropping exceed the ones under cropland [as reported by Drexler et al. (2021)], but also the ones under permanent grassland in many cases. Table S1 and Figure 3 show that in our study, the soil C stocks were too high to detect the recent effect of the land use change and increased C input by trees directly in the soil. However, the fine root distribution measured in our study showed that the trees allocated carbon to the subsoil via their root system. The mechanism of subsoil C storage poses many open questions (Siegwart et al., 2023) and is especially interesting for mixed cropping systems that can potentially increase C storage in soils compared to cropland. High deviations between sampling points as seen in Figure 1 have to be taken into account when interpreting the results and show that a high number of repetitions is needed for studies on complex interactions of root systems.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
RS: formal analysis, investigation, visualization, writing - original draft preparation; IK: investigation, visualization, writing - review and editing; AS: methodology, investigation, resources, writing - review and editing; MJ: funding acquisition, formal analysis, investigation, methodology, resources, writing - review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was financed by the Federal Ministry of Education and Research as part of the SIGNAL (Sustainable Intensification of Agriculture through Agroforestry) project Grant number (031B0510A) within the BONARES network which is gratefully acknowledged. We acknowledge support by the Open Access Publication Funds of the Göttingen University.
Acknowledgments
We thank the department of Crop Science and especially Prof. Isselstein for his support. We are grateful to Maren Langhof for her support and feedback on the manuscript. We thank the staff of the department for assistance during root washing. Special thanks goes to Dirk Böttger and Julian Meyer for technical assistance and to Andreas Parth for assistance with WinRhizo. We thank the Friedrich Löffler Institute for their cooperation on the site.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2023.1200785/full#supplementary-material
References
Angst G., John S., Mueller C. W., Kögel-Knabner I., Rethemeyer J. (2016). Tracing the sources and spatial distribution of organic carbon in subsoils using a multi-biomarker approach. Sci. Rep. 6, 1–12. doi: 10.1038/srep29478
Antony D., Collins C. D., Clark J. M., Sizmur T. (2022). Soil organic matter storage in temperate lowland arable, grassland and woodland topsoil and subsoil. Soil Use Manage. 38, 1532–1546. doi: 10.1111/sum.12801
Cardinael R., Chevallier T., Barthès B. G., Saby N. P., Parent T., Dupraz C., et al. (2015). Impact of alley cropping agroforestry on stocks, forms and spatial distribution of soil organic carbon–a case study in a mediterranean context. Geoderma 259, 288–299. doi: 10.1016/j.geoderma.2015.06.015
Coners H., Leuschner C. (2005). In situ measurement of fine root water absorption in three temperate tree species–temporal variability and control by soil and atmospheric factors. Basic Appl. Ecol. 6, 395–405. doi: 10.1016/j.baae.2004.12.003
Drexler S., Gensior A., Don A. (2021). Carbon sequestration in hedgerow biomass and soil in the temperate climate zone. Regional Environ. Change 21, 74. doi: 10.1007/s10113-021-01798-8
Gerjets R., Richter F., Jansen M., Carminati A. (2021). Hydraulic redistribution by hybrid poplars (populus nigra x populus maximowiczii) in a greenhouse soil column experiment. Plant Soil 463, 145–154. doi: 10.1007/s11104-021-04894-0
Hajek P., Hertel D., Leuschner C. (2014). Root order-and root age-dependent response of two poplar species to belowground competition. Plant Soil 377, 337–355. doi: 10.1007/s11104-013-2007-3
IUSS working group WRB (2015). World reference base for soil resources 2014. international soil classification system for naming soils and creating legends for soil maps (Rome: FAO).
Jose S., Gillespie A., Pallardy S. (2004). Interspecific interactions in temperate agroforestry. Agroforestry Syst. 61, 237–255. doi: 10.1007/978-94-017-2424-1_17
Kahle P., Janssen M. (2020). Impact of short-rotation coppice with poplar and willow on soil physical properties. J. Plant Nutr. Soil Sci. 183, 119–128. doi: 10.1002/jpln.201900443
Lepik A., Abakumova M., Davison J., Zobel K., Semchenko M. (2021). Spatial mapping of root systems reveals diverse strategies of soil exploration and resource contest in grassland plants. J. Ecol. 109, 652–663. doi: 10.1111/1365-2745.13535
Leuschner C., Hertel D., Coners H., Büttner V. (2001). Root competition between beech and oak: a hypothesis. Oecologia 126, 276–284. doi: 10.1007/s004420000507
McCormack M. L., Dickie I. A., Eissenstat D. M., Fahey T. J., Fernandez C. W., Guo D., et al. (2015). Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 207, 505–518. doi: 10.1111/nph.13363
Mulia R., Dupraz C. (2006). Unusual fine root distributions of two deciduous tree species in southern France: what consequences for modelling of tree root dynamics? Plant Soil 281, 71–85. doi: 10.1007/s11104-005-3770-6
Plante P.-M., Rivest D., Vézina A., Vanasse A. (2014). Root distribution of different mature tree species growing on contrasting textured soils in temperate windbreaks. Plant Soil 380, 429–439. doi: 10.1007/s11104-014-2108-7
Poorter H., Niklas K. J., Reich P. B., Oleksyn J., Poot P., Mommer L. (2012). Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytol. 193, 30–50. doi: 10.1111/j.1469-8137.2011.03952.x
Quinkenstein A., Woellecke J., Böhm C., Grünewald H., Freese D., Schneider B. U., et al. (2009). Ecological benefits of the alley cropping agroforestry system in sensitive regions of Europe. Environ. Sci. Policy 12, 1112–1121. doi: 10.1016/j.envsci.2009.08.008
Richards J. H., Caldwell M. M. (1987). Hydraulic lift: substantial nocturnal water transport between soil layers by artemisia tridentata roots. Oecologia 73, 486–489. doi: 10.1007/BF00379405
Rumpel C., Kögel-Knabner I. (2011). Deep soil organic matter–a key but poorly understood component of terrestrial c cycle. Plant Soil 338, 143–158. doi: 10.1007/s11104-010-0391-5
Siegwart L., Jourdan C., Piton G., Sugihara S., Van den Meersche K., Bertrand I. (2023). Root distribution and properties of a young alley-cropping system: effects on soil carbon storage and microbial activity. Plant Soil 482, 601–625. doi: 10.1007/s11104-022-05714-9
Tsonkova P., Böhm C., Quinkenstein A., Freese D. (2012). Ecological benefits provided by alley cropping systems for production of woody biomass in the temperate region: a review. Agroforestry Syst. 85, 133–152.
Keywords: fine roots, alley cropping, agroforestry, root system, root competition, fine root density, maximal rooting depth, permanent grassland
Citation: Sutterlütti R, König I, Swieter A and Jansen M (2023) Spatial distribution of tree and grassland fine roots in an alley cropping system. Front. Agron. 5:1200785. doi: 10.3389/fagro.2023.1200785
Received: 05 April 2023; Accepted: 29 May 2023;
Published: 15 June 2023.
Edited by:
Nasim Ahmad Yasin, University of the Punjab, PakistanReviewed by:
Ian McIvor, The New Zealand Institute for Plant and Food Research Ltd, New ZealandLuca Bragazza, Agroscope, Switzerland
Copyright © 2023 Sutterlütti, König, Swieter and Jansen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rahel Sutterlütti, cmFoZWwuc3V0dGVybHVldHRpQHVuaS1nb2V0dGluZ2VuLmRl