 Alison R. Gerken
Alison R. Gerken William R. Morrison III
William R. Morrison III- United States Department of Agriculture (USDA), Agricultural Research Service, Center for Grain and Animal Health Research, Manhattan, KS, United States
Climate change is a danger to the agricultural system and will impact the availability of food to feed the world. While much attention has focused on the effects of climate change on pest management prior to harvest, much less attention and time has been devoted to determining the effects of climate change on pest management in the postharvest supply chain from farm to fork. Climate change may percolate to pest management at a macro level through compositional changes in which species attack commodities through distributional changes or what commodities are grown in a region (and thus processed in that area). However, climate change may also result in altered microclimates at food facilities, which can be tied to increased generation times, elevated damage and contamination potential, greater abundance of species, and greater need for external inputs. A variety of integrated pest management (IPM) strategies may help increase the resiliency and adaptation of management to climate change. Tactics susceptible to warming temperature changes in climate showing decreased efficacy include semiochemical-based, behaviorally-based tactics, a subset of insecticides (e.g., pyrethrins and pyrethroids), and those that rely on low temperature (e.g., grain aeration, grain chilling). Tactics at food facilities showing resilience to warming temperature changes in climate include packaging, other groups of insecticides, and likely sanitation. Data gaps include predicting changing distributions for stored product insects under climate change, translating macro climate changes into microclimate changes at food facilities, and rigorously investigating how IPM tactic efficacy varies under changing climate. Increased scrutiny of how climate change will affect pest management in the postharvest supply chain will deliver improved outcomes for the entire agricultural system.
Introduction
There has been an extensive and in-depth amount of attention paid to the effects of climate change on the biodiversity of the planet and ecosystem services (Mooney et al., 2009). Foremost among the concerns from climate change is how agriculture may be affected. For example, crop losses by 10–25% for rice, maize, and wheat are expected as a result of a warming climate (Deutsch et al., 2018). This is partly because both insect abundance and number of generations in a given time period is predicted to increase under a warming climate (e.g. Gu et al., 2018). However, the vast majority of literature has focused on how climate change will affect pre-harvest agriculture and associated pest management, while neglecting the post-harvest supply chain (Stathers et al., 2013a; Adler et al., 2022). Nonetheless, throughout the agricultural supply chain, climate change is expected to influence how grain is grown, stored, processed, and shipped. Food facilities (e.g., used in this contribution to mean any structure involved in the storage, processing, and marketing of commodities in the post-harvest supply chain up to the consumer) represent unique, buffered habitats in the landscape, which create special microclimates in which pests may respond differently to climate change than unbuffered landscapes (e.g., Milling et al., 2018). Storage by smallholder farmers also represents a range of postharvest management styles which could possibly increase the buffering of effects of climate change (Nyabako et al., 2021). Climate change and buffering may also affect pest management after harvest, with some strategies more or less successful in a warming climate. Thus, the goals of this review were to 1) determine how changes prior to harvest translate after harvest, 2) evaluate the evidence for how the buffered environments of food facilities may attenuate the effects of climate change, 3) elucidate how postharvest commodity quality may change under climate change, 4) synthesize the work on which postharvest integrated pest management tactics may be most effective under a warming climate, and 5) summarize the current research gaps in climate change research for stored products moving into the mid-21st Century.
Materials and Methods
We comprehensively reviewed the literature on climate change research after harvest. In particular, we searched Web of Science and Google Scholar with the following search terms and/or combinations thereof: “climate change”, “stored products” “postharvest”, “insects” “integrated pest management”, “grain”, “maize”, “rice”, “wheat”, “tactics”, “modeling”, “quality”, “microclimate”, “mycotoxin”, etc. We searched articles without respect to timeframe. We did not purposefully exclude articles on specific stored products, but acknowledge the existing bias in the primary literature towards postharvest studies on maize, rice, and wheat (Davis et al., 2021). However, we did not include literature on horticulture crops, as this would have made the review too cumbersome, horticulture crops are managed significantly differently than postharvest grains, and horticulture crops have also been recently reviewed by Malhotra (2017) and Moretti et al (2010). Among the IPM tactics, we excluded botanical insecticides (apart from spinosad and Bt) as there have been several recent reviews covering this topic (e.g., Campolo et al., 2018; Spochacz et al., 2018), there are hundreds of publications (mostly laboratory studies), and there are no widespread commercial products. We limited our search to English language journals. Anything that was explicitly pre-harvest without any connection to the postharvest supply chain was eliminated from consideration. We found a total of 172 articles that fit our criteria. We synthesize the remaining articles into the seven sections below.
Shifts From Climate Change in Where Crops Are Grown
The geographical location where crops are grown is expected to shift and will impact timing and survival of stored-product insect pests. In the US, the typical breadbasket of the country is the Great Plains (Nebraska, Kansas, Iowa, Indiana, Arkansas, Illinois, and Texas) (USDA-NASS, 2022a). The major crops grown there include wheat (primarily Kansas), corn, and rice (Arkansas and Texas) (USDA-NASS, 2022b). However, as temperatures increase under climate change, the highest latitude at which crops are currently grown may generally be expected to also increase (Hijmans, 2003). From 1981–2015, climate changes in North America resulted in substantial shifts in the dates of planting and harvesting of wheat, normally leading to an extension of the growing season, while higher temperatures in June and July negatively affected grain yield at most sites (Morgounov et al., 2018). In the Great Lakes region, long-season maize yield performed significantly better under climate change in the northern states, but much worse in the southern states (Southworth et al., 2000). Easterling et al. (1993) modeled wheat and small grain production in Missouri, Iowa, Nebraska, and Kansas under historical and future climate, and when factoring in then-available low-cost technologies with CO2 enrichment, there would be a small net positive impact of climate change on wheat production in this region by 2030. Conversely, Ortiz et al. (2008) projected that by 2050, the Indo-Gangetic Plains, which are part of a favorable, high potential, irrigated, low rainfall mega-environment and accounts for 15% of global wheat production, could transform into a heat-stressed, short-season region resulting in major yield reductions. In addition, the same study projected the area covered by the cool, temperate wheat-conducive mega-environment could expand as far north as 65°N in North America and Eurasia. In the Pacific Northwest of the US, Fei et al. (2017) predicted that warmer temperatures, drier seasons, and increased atmospheric CO2 will shift northwards in the Southern Great Plains and westward in the Northern Great Plains, which are moves to cooler conditions in both locations. In addition, there would be a decline in total wheat acreage from 6 million acres to 5.4–5.7 million acres in the Pacific Northwest by 2100. Overall, yields may decrease but cultivation may persist in the historical ranges of crops, and crops may also be grown in new areas. This continuity indicates minimal disruption to the postharvest supply chain as a result of crop switching, at least in the near and medium term.
Cereal crops in southern and central Europe have shown earlier ripening and this phenological shift could result in the grain entering storage at a more ideal time for infestation and insect population growth (Medina et al., 2015). This is more ideal for insects because grain is generally very warm when placed into storage, which is optimal for the development of insects, and when given a longer (more optimal) time to feed and develop, this may translate into more generations per year and a greater abundance of postharvest insects. Models that indicate an overall decline in yield (e.g., Batts et al., 1997; Basche et al., 2016; Deutsch et al., 2018) will only drive the need to protect post-harvest commodities to a greater extent (Nyabako et al., 2021). However, other models predict an increase in yield (Smith and Almaraz, 2004; Smith et al., 2013) as long as the temperature increases are not extreme (Lobell and Asner, 2003). In addition, drier conditions as well as an increase in extreme events create increased variability and uncertainty in yield predictions, especially when major droughts or storms occur (Kerr, 2003; Smith and Almaraz, 2004). Even if areas are expected to have increases in yield, these increases could be due to adaptation or changes in crop composition, resulting in net nutritional losses (Nuttall et al., 2017). Areas such as the western edge of the USA prairies, eastern Brazil, and western Australia are especially susceptible to decreases in crop yields while areas such as Canada and northern Europe and Russia are predicted to have increases in yield according to spatially explicit global circulation models (Gregory et al., 2005).
Climate Change-Induced Alterations in Postharvest Quality of Commodities
Climate change may induce postharvest impairments in the quality of commodities as they travel through the agricultural supply chain. Changes in quality may arise as a result of drought stress, heat stress, or elevated CO2. Many postharvest crop qualities can be linked to performance of the crop in the field prior to reaching a food facility (Figure 1). For example, drought stress was found to reduce the filling rate and duration of sorghum, thereby reducing final seed size (Chadalavada et al., 2022). Starch content was found to increase slightly under a small level of drought stress, but significantly decrease under high drought conditions in sorghum. Stress conditions like drought or high temperature typically result in increased protein content in sorghum as a result of an increase in the percent of nitrogen content (Weightman et al., 2008; Pang et al., 2018; Impa et al., 2019). In modeling changing rice quality under projected climate change in northern Italy, Cappelli and Bregaglio (2021) found commodity quality would decrease by 8–20%, with a loss of revenues to millers of 50–100 € per ton between 2030–2070. Heat stress and drought was found to decrease starch content (by 5–10%), but increase protein (by 10%) and mineral concentrations (by 10–20%) in a meta-analysis of C3 cereal grains (e.g., C3 is the most common photosynthetic pathway, including wheat, rice, barely, and oats, among 85% of other plant species; Mariem et al., 2021). Elevated CO2 was found to increase the carbohydrate content of rice (Goufo et al., 2014). Iron bioavailability decreased by 22% in brown rice, while calcium increased by 5–11% in the bran and husk (Goufo et al., 2014). Further, elevated CO2 also resulted in decreased concentrations of amino acids, including isoleucine, phenylalanine, and tyrosine (23–57%).
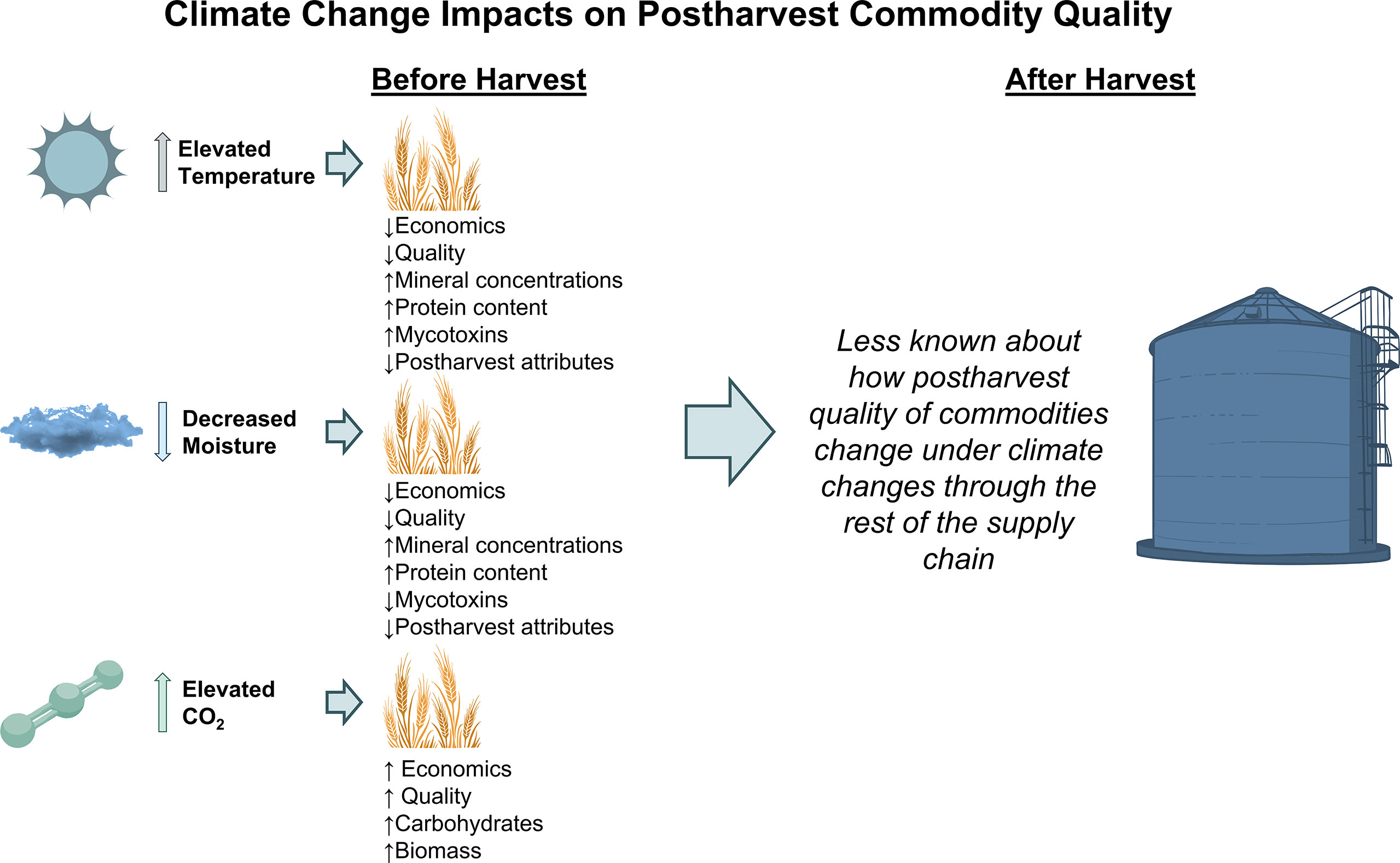
Figure 1 Climate change impacts (e.g. elevated temperature, changed moisture, elevated CO2) on postharvest commodity quality starting prior to harvest and continuing throughout the supply chain. Less is known about how commodity quality is affected after harvest compared to prior to harvest.
Importantly, climate change may directly affect milling and other important processing steps later in the postharvest supply chain. As temperatures are predicted to increase, higher temperatures are likely to reduce the shelf life of many commodities (Ashaye, 2018; Parajuli et al., 2019; Thole et al., 2021). In addition, the overall nutritional quality of food is greatly affected by climate change scenarios (Srivastava, 2019; Mariem et al., 2021), altering carbohydrate-to-protein and sugar-to-nutrient ratios (Ashaye, 2018) and even on milling qualities and cooking suitability (Cappelli and Bregaglio, 2021). More to the point, prior work has shown that 60% of the variation in technological and breadmaking parameters is influenced by the pre-harvest production season (Mastilović et al., 2018). The largest contributor within production season was environmental impact. That study found that a season simulating climate change conditions resulted in lower amylolytic enzymes, lower content of gluten, and higher extensograph dough resistance, among other parameters. Furthermore, prior work has shown a nonlinear effect of high temperature stress exposure on yield and milling quality of rice (Lyman et al., 2013). For instance, a 1°C increase in mean yearly temperature reduces paddy rice yield by 6.2%, but actually reduces total milled rice yield by 7.1–8%, head rice yield by 9–13.8%, and total rice milling revenue by 8.1-11%. By contrast, Zhao and Fitzgerald (2013) showed that low relative humidity and a high vapor pressure deficit in the dry season is associated with low chalkiness and high head rice yield in spite of higher maximum temperatures. That study demonstrated a combination of climate factors enabled rice to maintain a cool canopy. Furthermore, in a manipulative experiment, Giménez et al. (2021) found warmer nights during wheat growth ended up reducing yield of the plant, but also surprisingly increased the breadmaking quality parameters, including the amount of wet gluten.
In recent years, the food industry has been reporting increasing problems with off-odor grain (Liu et al., 2021). Much of this may be attributable to the increased prevalence of stored grain pathogens (Van Winkle et al., 2022). However, less is known about whether temperatures and the changes associated with climate change have a causal effect on the incidence of these problems in commodities after harvest. As recently as 2010, there was only one published study and one abstract which directly referred to the effects of climate change on mycotoxins (Paterson and Lima, 2010). Predictions at that time included that food grown in temperate regions would be more susceptible to aflatoxin contamination under higher temperatures, while food grown in tropical regions would become too inhospitable for conventional fungal growth and mycotoxin production (Paterson and Lima, 2010). Indeed, under the most probable scenario of warming (e.g., +2°C), aflatoxin B1 was predicted to become a food safety issue in maize in Europe within 100 years (Battilani et al., 2016). Improperly stored commodities may see increases in mycotoxins (Paterson and Lima, 2010; Magan et al., 2011; Medina et al., 2015; Akello et al., 2021). Increased weather extremes, including flooding or drought, greatly affects water quality in a region and can result in increased pathogen transfer in post-harvest handling (Atanda et al., 2011). More recently, Medina et al. (2017) found Aspergillus flavus was likely to be unaffected by climate change factors (elevated CO2, temperature, moisture), but that there was a significant stimulation in the production of its mycotoxin, aflatoxin B1 in vitro and in vivo in corn. However, cereal grains grown in drier and hotter areas of Zimbabwe had higher A. flavus contamination, while there was more fumonisin content in wetter years (Akello et al., 2021). In Malawi, warmer and drier conditions from climate change are predicted to make pre-harvest conditions more favorable to aflatoxin B1 contamination, with contamination risk increasing at more northern regions (Warnatsch et al., 2020). In addition, another study found aflatoxin contamination risk in maize in South and Central-Europe is highly likely in the next 30 years as a result of optimal climatic conditions for the growth of A. flavus (Moretti et al., 2019). In that study, deoxynivalenol contamination in wheat was also found to be an emerging problem in Northern Europe. Thermotolerant species will thrive under climate change, leading to a shift in dominance from Penicillium spp. to Aspergillus spp. (Perrone et al., 2020). Within Aspergillus spp., mycotoxin production is predicted to favor aflatoxins at the expense of other mycotoxins under climate change (Perrone et al., 2020). These recent studies suggest a strong effect of climate change factors on fungal growth, diversity, and mycotoxin production in post-harvest commodities.
Important gaps in the literature include knowing how degradation in postharvest commodity quality may vary under different longer storage durations given climate changes’ higher temperature, altered atmospheric mixture, and humidity changes. In addition, knowing how postharvest quality interacts with different types of buffered environments (e.g. food facilities) under different climate change scenarios is important to tease apart. Finally, better understanding the three-way interaction among mycotoxins, climate change, and postharvest quality will also be central.
Changing Microclimates at Food Facilities Under Climate Change
To be successful as pests, insects that infest post-harvest commodities must have the capacity to infest a wide range of habitats, including indoor and outdoor areas. Following food resources provided by stored commodities, these insects are often found within or adjacent to human-made food facilities (herein referred to as food facilities) and bulk grain storage. These areas can be buffered sources of microclimates, or small areas where the climatic conditions differ from a larger space (Rebaudo et al., 2016). As an example, sagebrush in a steppe environment in the western US can serve as a reasonable facsimile of the food facilities dotting agricultural landscapes in much of the central US, in terms of acting as an effective buffer for microclimate by lowering the maximum temperature reached and decreasing the magnitude of the range of temperatures (Milling et al., 2018; Figure 2). As ectotherms, insects rely on sources of heat to metabolically function (Sears and Angilletta, 2015) and finding areas of heat within stored products can create opportunities for population growth and other life processes. Even though they are buffered, as temperatures are predicted to warm, microclimates are expected to warm as well, often earlier than usual, which can increase population growth and infestation rates. These microclimates can also be influenced by both biotic and abiotic factors which can contribute to difficulties in prediction and increased variability within these areas (Andrew and Hill, 2017).
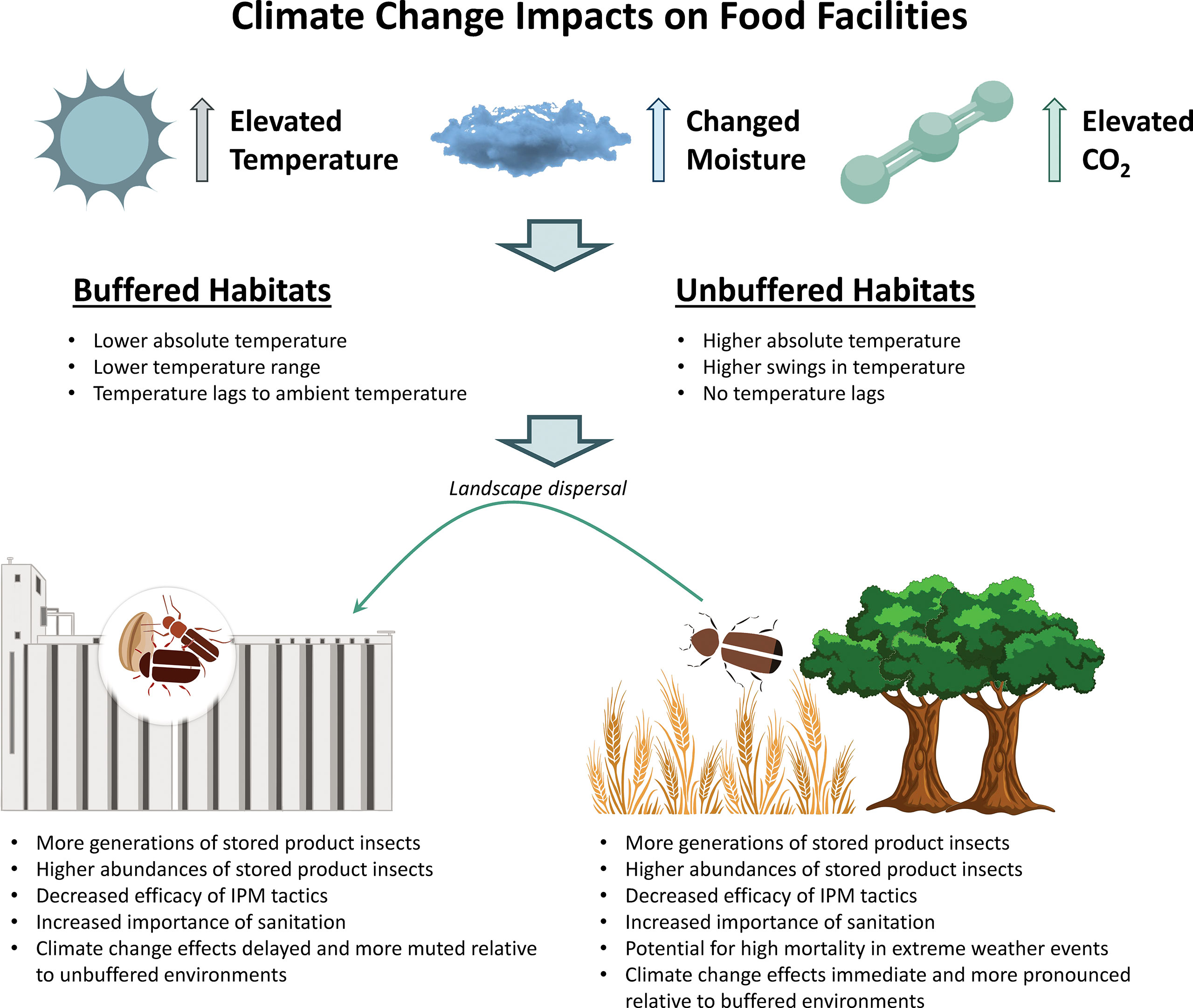
Figure 2 Climate change impacts (e.g. elevated temperature, changed moisture, elevated CO2) at food facilities, including buffered and unbuffered environments, and how this affects each environment differently.
Bulk grain itself is made of a myriad of microclimates (Elmouttie et al., 2010), with low temperatures being important to maintain integrity of the grain (Bradna et al., 2018). Low temperatures can also suppress insect population growth and lead to seasonal insect suppression during cooler months (Ileleji et al., 2007). Bulk grain changes consistently in response to outdoor thermal pressures and models can be developed to map different temperatures and moisture content throughout the grain (Casada and Young, 1994; Jian et al., 2005). The center of the internal mass of grain may be very warm shortly after loading into a silo and may present opportunities for rapid insect development (Arthur and Flinn, 2000). Humidity can also be a factor for bulk grain storage, as some insects congregate toward the top of the grain mass and can experience extreme drying conditions (Hagstrum et al., 1998; Cordeiro et al., 2016), often mimicking ambient conditions quickly (Sinha, 1973). Within the grain bulk, aggregations of insects create increased levels of relative humidity due to the metabolic activity of the insects (Ashaye, 2018), which can then drive attraction of insects searching for water. In addition, bagged grain is a microclimate that encourages insect growth (Omodara et al., 2020), at least when hermetic packaging is not used. The microclimates of bagged products change at a slower rate than the outside temperatures (Flinn et al., 2015). As climates continue to change, these rates inside packaged goods can become more variable and a reduction in the period of cool temperatures that suppress insect growth may result in earlier insect population growth and larger population sizes throughout the post-harvest supply chain (Plarre and Burkholder, 2009; Omodara et al., 2020).
Food facilities also often mirror the outdoor temperature but usually with some lag to ambient temperature, although the temperature range is compressed, and the magnitude of difference changes with season (Campbell et al., 2010; Arthur et al., 2014; Gerken and Campbell, 2019). These food facilities provide a buffer for the insects from extreme thermal events and can encourage cryptic populations to be present year-round. However, insects that primarily live in a food facility, such as mites, have still been shown to vary significantly by season, indicating that these environments are still greatly affected by outdoor climate conditions (Sinha, 1968; Moses et al., 2015; Tytar and Oksentyuk, 2019). As the climate changes, the ability to regulate indoor environments is predicted to change with each season and become more difficult to maintain. This could result in an increase in the energy required to regulate the seasonal and extreme temperature variation changes (Bradna et al., 2018). Natural cool periods are predicted to decrease in length and the cost to run a fan and other equipment needed to cool the grain could become prohibitive for some storage facilities (Bradna et al., 2018).
Storage and processing facilities have a wide variety of floor plans, including shipping and receiving areas, storage on shelving, machinery for processing the commodity, and office space for clerical work (Doud and Phillips, 2000; Campbell and Arbogast, 2004; Semeao et al., 2013). As with bulk grain, different spaces can lead to a variety of microclimates. Areas by doors and windows have more thermal variation including variation in air flow which can lead to differences in insect trap captures (Doud and Phillips, 2000; Arthur et al., 2014). Most insects will move to areas that are hidden or in cracks and crevices (Child, 2007). For example, shelving can provide crevices and areas where insects can reach both food and shelter (Toews et al., 2005) and these areas may not reflect the measured temperature or humidity of the larger space (Child, 2007). Microclimates can also be created by running machinery resulting in localized warmer regions where insects can thrive (Trematerra and Sciarretta, 2004). Thus, these variety of microclimates may have repercussions for postharvest commodity quality and insect populations throughout the postharvest supply chain, and food facilities may insulate them from the effects of climate change to varying degrees.
Smallholders throughout many tropical and semi-arid areas are also predicted to be impacted by climate change. Typically, smallholders have shifting storage of their grain from bulk storage in traditional free-standing granaries to polypropylene bags stacked in the living quarters in response to rising theft from food insecurity triggered by climate change (Nyabako et al., 2021). Generally, living quarters are smaller than large postharvest processing facilities in developed nations and are not climate-controlled, which may mean that microclimate may be buffered to a lower extent in response to climate change.
Changing Stored Product Insect Community in Response to Climate Change
Because insects are ectotherms and are susceptible to changes in temperature and humidity, climate change is predicted to impact agricultural insect pests greatly. Changes to insects can occur both physiologically and behaviorally and can result in significant adaptations over time (Estay et al., 2009). Phenological changes to ripening and harvesting brought on by climate change can also affect insect infestations. Climate variability has been shown to impact a third of crop yield variability on a global scale and climate change predictions indicate an increase in instability of crop yield predictions (Ray et al., 2015). Cereal crops in southern and central Europe have shown earlier ripening and this phenological shift could result in the grain entering storage at a more ideal time for infestation and insect population growth (Medina et al., 2015). Warm grain entering storage presents an ideal habitat for insect development, and when provided longer time periods at optimal conditions, more generations and higher abundance of postharvest insects are possible. Changes in phenology and yield variability can also be associated with adaptation or changes in crop composition, leading to changes in insect pest pressure (Smith and Almaraz, 2004), shifting habitats of insects, and changes in life history characteristics (Porter et al., 1991; Patterson et al., 1999; Stathers et al., 2013a).
Temperature is a significant driving force of lifetime fitness success in many insects, and predicted adaptations suggest that increasing temperatures will result in the ranges of insects shifting towards the poles or to higher altitudes (Bale et al., 2002). For those species that are more influenced by both biotic and abiotic factors, predicting changes may be more difficult. Species distribution models are a common tool for predicting potential areas of infestation but are also useful for projecting changes in habitat due to climate change. For example, Tribolium confusum (du Val) (Coleoptera: Tenebrionidae) is a common pest in northern Chile, and climate change models indicate that by 2071–2100, they could reach similar densities in southern Chile, where it is currently a non-pest species (Estay et al., 2009). Arthur et al. (2019) projected worldwide suitability of habitat for Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) given the climactic conditions of where it is known to occur, and the authors found there is the opportunity for it to expand polewards beyond its current range. Sitotroga cerealella (Olivier) also showed significant adaptation and changes in heat acclimation over multiple generations and may also use desiccation ability, along with P. truncatus, to expand its range over time (Machekano et al., 2018; Mutamiswa et al., 2021; Mpofu et al., 2022). It is likely that there have been dramatic shifts in the distributions of three weevil species, Sitophilus zeamais (Motschulsky) (Coleoptera: Curculionidae), Sitophilus oryzae (L.), and Sitophilus granaries (L.) in Europe over the last 20 years in response to climate change (Adler et al., 2022). Overall, increases in temperature increase reproductive potential and developmental rates and decrease generation time (Stange and Ayres, 2010).
In addition to warming, changes in cold stress can drive how insects will respond to climate change although this relationship is complex when considering extreme events and lower insulation levels from snowpack (e.g., layer of snow during winter; Stange and Ayres, 2010). Often the lower lethal limit of temperature is the driving habitat limitation for insects (Sharma, 2014). As temperatures increase during cooler months, winter mortality in insects is predicted to decrease with two caveats. First, insects rely on leaf litter and snow for overwintering insulation and may not be as protected from severe cold events and may not survive as easily (Lombardero et al., 2000). Even stored product insects are known to use wild hosts and refugia in the natural landscape (Jia et al., 2008; Mahroof et al., 2010) and can be trapped far from any nearby food facility (Scheff et al., 2021). It is also known that stored product insects will readily immigrate into food facilities from the natural landscape (Wilkins et al., 2021). Second, insect cold tolerance varies throughout the cold season and if extreme cold events occur at the end of the cold season when an insect has physiologically begun to prepare for spring, the insect could face greater mortality (Dukes et al., 2009). However, with a reduction in the length and severity of the cold season, there will be higher numbers of pests that will not die, increasing the opportunities for residual infestations (Stathers et al., 2013b).
The diversity of pest assemblages may also be impacted by climate change (e.g., Kansman et al., 2021). As some species relocate and establish in new climatic areas, interactions and competition can change species dynamics (Ladányi and Horváth, 2010; Sharma, 2014; Andrew and Hill, 2017). For example, Quellhorst et al. (2020) found P. truncatus was the superior competitor on maize at higher temperatures (e.g., 30 or 35°C), while S. zeamais was the superior competitor at cooler temperatures (e.g., 20–25°C). At 30°C, S. oryzae was the superior competitor relative to S. zeamais and S. granarius, regardless of whether competition was at low, medium, or high density (Athanassiou et al., 2017). The quarantine pest, khapra beetle, Trogoderma granarium Everts (Coleoptera: Dermestidae) performed better on rice than wheat and was the superior competitor at 32°C as opposed to 25°C compared to Trogoderma variabile Ballion (Coleoptera: Dermestidae) (Lampiri et al., 2022). Natural enemies of these agricultural pests may also be more prevalent in new locations which would be a positive outcome of the shifting distribution of the pest insect (Porter et al., 1991). In stored grain pests, the parasitoid Anisopteromalus calandrae (Howard) (Hymenoptera: Pteromalidae) has a lower limit for population growth of 20°C, while S. granarius can grow and survive as low as 15°C (Fields, 1992) again creating a thermal space where the insect pest can continue to infest with a reduction in parasitoid control. Overall, species distribution limits are key to understanding range expansions and release of predatory pressure as temperatures change. For example, Hansen and Steenberg (2007), demonstrated that Lariophagus distinguendus (Förster) (Hymenoptera: Pteromalidae) had a significantly greater control of S. granarium than did A. calandrae at 20°C. Knowing the thermal limits of these interactions will provide more efficient use of these biological control insects in the face of climate change.
Increased weather extremes, including flooding or drought, greatly affects water quality in a region and can result in increased pathogen transfer in post-harvest handling (Atanda et al., 2011). Overall, an increase in pathogens and mycotoxins can drive an increase in insect populations within post-harvest commodities by creating hotspots ideal for insect development (Srivastava, 2019; Ponce et al., 2021). If both mycotoxins and insect populations are set to increase, overall proportions of loss of post-harvest commodities are also predicted to increase. Drought can also significantly reduce yields (Eggen et al., 2019) and a combination of reduced yields plus increased potential of over-drying and cracking the cereal grain (Paterson and Lima, 2010) which can leave grains more susceptible to insect infestation and have a cumulative effect on total grain loss because of climate change. Conversely, flooding in a region can decrease drying of the grain before storage, increasing the moisture content and potential for insect infestations (Moses et al., 2015).
Changing Integrated Pest Management Efficacy After Harvest in Response to Climate Change
Effectiveness of IPM strategies also needs consideration when discussing how climate change will affect stored product pest insects. The effectiveness of synthetic pesticides in the face of changing climates has also received attention with differing results in effectiveness based on the properties of the pesticide (Tsaganou et al., 2021). Increased temperature leads to increased movement for insects which can lower the residual effectiveness of many insecticides (Arthur et al., 1992). However, increased movement can also increase the interaction with an insecticide, increasing mortality (Tsaganou et al., 2021). Laboratory experiments found the efficacy of diatomaceous earth (DE, e.g. Silicosec®) increased with temperature with S. oryzae were exposed, but mortality rates for T. confusum was lower at 32°C than 30°C for the 24 and 48 h exposure intervals (Athanassiou et al., 2005). Higher temperatures reduced the efficacy of DE against T. castaneum, but increased it against Cryptolestes ferrugineus (Stephens) (Coleoptera: Cucujidae) (Fields and Korunic, 2000). By contrast, regardless of insect species, the lower the moisture content of the grain, the greater the mortality inflicted. Efficacy of fipronil was hampered by application as a grain protectant at 30°C for Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae) and P. truncatus compared to 20 and 25°C (Kavallieratos et al., 2010). By contrast, spinetoram efficacy as a grain protectant increased with temperature against S. oryzae, but was unaffected against R. dominica and T. confusum (Vassilakos and Athanassiou, 2013).
Temperature and humidity had significant but complex effects on the efficacy of spinosad as a grain protectant against multiple stored product insects, but its efficacy was found to especially increase with higher temperatures (e.g. 30 vs 20°C) against S. oryzae (Athanassiou et al., 2008). The reason cited for this increased efficiency was the increased mobility of S. oryzae compared to the others and the fact that 30°C was near its thermal development optimum. Prior exposure to spinosad at 18 ppm or higher increased the mortality of R. dominica and S. oryzae when exposed to extreme high temperature (40°C) or low temperature (6-11°C) afterwards (Wijayaratne and Rajapakse, 2018). The efficacy of Bacillus thuringiensis (Bt) against Plodia interpunctella was decreased by a third after 42 weeks of exposure on wheat at 42°C compared to 25°C (Kinsinger and McGauhey, 1976). Short-term exposures to high temperatures (45 or 55°C) were found to have no appreciable effect on the efficacy of cyfluthrin or hydroprene when applied as a wettable powder on concrete against T. castaneum (Arthur and Dowdy, 2002). Hydroprene was equally effective at inducing mortality of T. castaneum and T. confusum larvae when applied at 27 or 32°C but was marginally more effective at 75% RH than at 57 or 40% RH. Residual application of pyrethrin + methoprene as an aerosol did not vary in efficacy against T. inclusum or T. variabile when tested between 25–40°C (Lanka et al., 2019). There was no consistent effect of temperature or humidity on the efficacy of pirimiphos-methyl as an emulsifiable concentrate and capsule formulation against seven stored product insect species (Rumbos et al., 2013). In general, the efficacy of synergized pyrethrins and most pyrethroids is negatively correlated with temperature (Musser and Shelton, 2005; Satpute et al., 2007; Arthur, 2012), including cyfluthrin, which is registered and commonly used as a residual insecticide in the US (Arthur, 1999). By contrast, the efficacy of most organophosphates, DE, carbamates, and neonicotinoids is positively correlated with temperature (Norment and Chambers, 1970; Kavallieratos et al., 2011; Arthur, 2012; Mansoor et al., 2015). The effect of relative humidity on conventional insecticides is generally mixed with both negative and positive effects, depending on species, and compound, except in the case of DE where efficacy usually declines with increased relative humidity. Thus, overall, climate change may have mixed effects on chemical control tactics, but with mostly negative impacts given the specific compounds that are most often used in stored product protection.
Fumigant efficacy may also vary with climate change parameters at food facilities. For example, the control of Liposcelis bostrychophila Badonnel (Pscoptera: Liposcelididae) with phosphine fumigation in grain increased from 20 to 30°C, but then dipped again by 35°C (Nayak et al., 2003). Similarly, Aulicky et al. (2015) found all adults and larvae by a susceptible and resistant strain of T. confusum were dead after a structural phosphine fumigation in a mill, even with temperature varying from 20–30°C and RH varying 44–78%. In testing phosphine fumigation efficacy against T. granarium from 25–39°C, full mortality of phosphine susceptible individuals was achieved by 34°C, whereas phosphine-resistant T. granarium required fumigation at 39°C (Ahmad et al., 2000). At 15°C, it took 1.5 d to kill 95% of the R. dominica eggs with phosphine, whereas it only took 0.7 d at 35°C (Flinn et al., 2001). In general, higher temperatures increased fumigation efficacy in stored products up to a point, likely because it increases respiration by insects.
Pheromone-based strategies may also vary in efficacy under climate change. Currently, the only semiochemical, behaviorally-based strategy commercially employed after harvest is mating disruption for a variety of stored product moths (Morrison et al., 2021). Semiochemicals may easily be impacted by many environmental factors including temperature, humidity, and wind (Heuskin et al., 2011; Cardé, 2021). High temperatures can often significantly decrease the half-life of the semiochemicals used (Cork et al., 2008) and understanding the timing of replacement of these lures or dispensers can provide the most consistent pest control options (Morrison et al., 2021). In addition, these lures and dispensers are often placed in areas that will have variable microclimates, which are expected to increase in variability as climates change (Andrew and Hill, 2017). However, the effect of climate change on pheromone-based tools has not been specifically evaluated after harvest. Nonetheless, a study evaluating decreased performance of the codlemone lure for mating disruption of Cydia pomonella (L.) (Lepidoptera: Tortricidae) found that there was an increase of 3°C in mean temperature to 35°C during the growing season in Germany, which led to significant decreases in response by males, and a correspondingly greater number of isomeric and chemical impurities in the codlemone compared to lures not experiencing the increased temperature (El-Sayed et al., 2021). While food facilities will be buffered to some extent from changes in temperature, it is likely that they may experience warmer temperatures for a greater portion of the season, which may result in lures being expended faster and/or having lower efficacy. Temperature was identified as a low severity challenge to the implementation of behaviorally-based management strategies at food facilities, with an additional consequence being more frequent maintenance of traps and lures (Morrison et al., 2021). One potential solution to the higher release rates in lures from higher temperatures during climate change may be to formulate lures in materials that slow diffusion of compounds. While information is still lacking on how pheromone-based tools may be affected at food facilities specifically under climate change, the little available evidence suggests climate change will have a negative impact on the efficacy and longevity of pheromone-based tools.
Biological control may also be affected by climate change at food facilities. For example, more temperate and wetter winters may give augmentative biological control agents a greater opportunity for success (Cook et al., 2004). Warmer temperatures would increase the activity of parasitic wasps, while higher humidity may improve the efficacy of entomopathogenic fungi (Cook et al., 2004). Studies on the functional response of stored product biocontrol agents to temperature support this point. For instance, Theocolax elegans (Westwood) (Hymenoptera: Pteromalidae) showed a Type-II functional response and parasitized the most R. dominica at 30°C, though this dipped slightly by 32.5°C (Flinn and Hagstrum, 2002). The search rate and handling time by T. elegans for R. dominica was also the shortest at these two highest temperatures compared to 25 or 20°C. Likewise, the highest developmental rate was observed for Lariophagus distinguendus, another biocontrol agent after harvest, at about 33°C (Ryoo et al., 1991). Thus, in the near and mid-term, there may be gains for biological control, but if warming progresses, these advantages may be lost.
Nonetheless, warmer temperatures may have pronounced negative effects on specific types of natural enemies. For example, higher temperatures may negatively affect conidial viability and germination of entomopathogenic fungi, with the optimum reported temperature for Beauveria bassiana somewhere between 25–30°C (Rumbos and Athanassiou, 2017a). Likewise, high temperatures and low humidity have a pronounced negative effect on the virulence of entomopathogenic nematodes at food facilities (reviewed in Rumbos and Athanassiou, 2017b). Thus, not all natural enemies may benefit.
However, parasitic wasps in food environments, like the pests they attack, prefer dry environments. Lariophagus distinguendus preferred environments with low RH (e.g. 32.5%) compared to those with higher RH (e.g. 53–97.5%; Steidle and Reinhard, 2003). The authors of that study concluded that the wasps use RH as a habitat cue. However, increased moisture content in maize kernels (from 11.6–14.6% MC) resulted in decreased development time and an increased intrinsic rate of increase by A. calandrae on S. zeamais, but this also favored the development of S. zeamais. As a result, if precipitation patterns are changed as a result of climate change at food facilities, this may disrupt host location and development by some parasitoids.
Although it has not been documented at food facilities, climate change may result in other destabilizing effects between parasitoids and hosts. For example, asynchrony in life history may arise between parasitoid populations and stored product pests. Asynchrony in life cycle has been documented in other systems (Logan and Wolesensky, 2007; Tylianakis and Binzer, 2014), which may result in a mismatch between susceptible host stages and parasitoid adults. In addition, increased temperatures may result in increased metabolic demand by parasitoids beyond what their consumptive capacity can provide (Binzer et al., 2012). This may be especially true if the parasitoid is the same or smaller than the prey and the prey is smaller than its plant resource (Binzer et al., 2012), which are both generally true in stored products. There may also be non-additive effects in combining climate change factors on the efficacy of biological control. For example, Romo and Tylianakis (2013) found separate warming and drought treatments increased parasitoid emergence success and offspring production, respectively, but when combined they observed the lowest parasitoid emergence of all the treatments. However, none of these aspects of climate change effects have been evaluated to determine how they affect biological control after harvest.
Less is also generally known about how cultural controls may vary under climate change after harvest in managing stored product insects. However, there is a fair bit known about how the performance of grain aeration may change under different climatic patterns. Grain aeration takes advantage of cooler external temperatures from a grain mass, then circulates that air inside at a rate of 3% that of grain drying techniques to lower the temperature of a grain mass below the developmental threshold for insects. This tactic performs very well in temperate regions (Arthur et al., 2020), and where cool external temperatures prevail for much of the year, additional pest management tactics may not be required. However, at more southern latitudes or in warmer areas, while grain aeration may still confer a benefit, it will need to be paired with other pest management tactics such as fumigation (Lopes and Steidle Neto, 2019; Morrison et al., 2020; Morrison et al., 2022) found that climate change reduced grain cooling potential, and increased energy costs, while making aeration of stored beans no longer practical in certain parts of Brazil. That study found other parts of Brazil could continue to use aeration under climate change but must pair it with good storage practices and efficient management procedures. A small study in the United Kingdom suggested that despite warmer than average temperatures in recent years, differential fan control and aeration can still achieve efficient cooling (Cook et al., 2004). However, it is likely that extreme shifts in temperature in the long-term, especially in autumn, may reduce aeration efficiency. As grain aeration becomes more difficult, it may become more necessary to implement grain chilling, whereby grain masses are actively cooled instead of passively cooled. Disadvantages to grain chilling include its high energy use and expense, and associated carbon emissions through its use. In the near and mid-term, grain aeration appears tenable as a pest management tactic that can continue to be used, albeit with attenuated efficacy.
There are a variety of other cultural controls commonly used against stored product insects, including extreme temperatures (heat and cold), packaging, sanitation, and grain chilling (mentioned above). Increasing temperatures can encourage increased movement of the insects (Stathers et al., 2013b), which can result in the spread of infestations into surrounding uninfested products or sites of spillage. This means that sanitation will become even more important going forward under climate change. In addition, higher temperatures from climate change may make sanitation even more critical for the proper functioning of a variety of other integrated pest tactics at food facilities (e.g., reviewed in Morrison et al., 2019). Research on hermetic bags showed that there was little effect of elevated temperature on the success of suppressing insect population growth in maize grain up to 38°C (Singano et al., 2020). Similar investigations with hermetic packaging showed increased mortality in the same treatments up to 50°C (Pražić Golić et al., 2016). Interestingly, higher temperatures are often recorded within the hermetic bags compared to the outside temperatures, which could also encourage insect mortality (Njoroge et al., 2014; Prasantha et al., 2014; Singano et al., 2020). While it is likely climate change has little impact on the efficacy of extreme temperatures, it is possible that climate change will make it more difficult to reach a specific temperature and/or hold the temperature for a given period. There may also be additional associated costs with the treatment from the added energy required to maintain temperature under inhospitable external ambient conditions. The use of extreme temperatures may remain easy to deploy in smaller, confined spaces, for example in quarantine applications.
In a comparison of multiple kinds of tactics, including neem leaf powder, Actellic Super dust, Zerofly bags, Purdue Improved Crop Storage Bags (PICS), Super Grain Bags, hermetic silos, and untreated grain in a polypropylene bag in the climate-risk prone Shire Valley in Malawi, (Singano et al., 2019) found that PICS resulted in the lowest damage to maize from insects under high mean temperatures (35.6°C) and high initial grain moisture (13.7%) for 32 weeks. However, in general, there have been few multi-tactic IPM assessments especially in relation to climate change after harvest at food facilities. This is certainly an area which could use more scrutiny in the future as the world moves to mitigation and adaptation to the changing climate.
Strategies for Adaptation by Food Facilities to Climate Change
While much work remains to be done in evaluating how the efficacy of many tactics may be impacted by climate change, there are several guiding principles and strategies that may help food facility managers prepare for working in a world with a changing climate. First among these is increased awareness that changing pest communities may result in new and unexpected pest problems. This is apparent from shifting pest distributions and is likely to become even more frequent. Particular pests to watch in North America include P. truncatus and T. granarium, a species of concern (Quellhorst et al., 2021), and quarantine threat, respectively (Athanassiou et al., 2018). The former resembles R. dominica, while the latter resembles many endemic dermestids, including T. variabile and T. inclusum (Lampiri et al., 2022).
Another relatively safe assumption may be that pest management for insects after harvest may be more expensive in the future, so may require a larger share of a food facility’s operating budget. This may result from a confluence of factors, including needing to run grain aeration for longer (Morrison et al., 2022; or switch to grain chilling), to replace pheromone lures more often, to reapply insecticides more often because they more quickly break down, and to deal with increased commodity spoilage under increased temperatures from climate change. Planning ahead financially will make sure food facilities are not caught unawares when additional expenses arise.
A strategy for adaptation may be to begin to implement comprehensive IPM programs for stored product pests with multiple hurdles to infestation at food facilities early. Already having effective programs in place will make it easier to make slight modifications, than to have to recreate the system from scratch under new more challenging circumstances. This may also include having a long-term monitoring program (Gerken and Campbell, 2021), which may provide an early detection system for new and different pest problems that may become an issue. In addition, establishing a collaborative network of pest managers that can share presence data or emergence timing of different species may create a better predictive system for monitoring and treatment efforts. Strategic planning of treatments that rely on monitoring data can save food facilities money and time in management efforts.
For smallholder farmers, postharvest adaptation must focus on making changes that benefit each farmer or postharvest storage systems (Stathers et al., 2013). A lack of investment and research in this area is a key detriment to protecting postharvest goods, especially in places like Sub-Saharan Africa (Nyabako et al., 2021). Filling in the knowledge gaps with data driven science will help guide farmers to developing plans to protect their commodities.
Finally, another key adaptation strategy may be to use IPM tactics favored under the projected climate change scenarios (Table 1). Very little seems known for certain about whether specific tactics would be of benefit under a warming climate, but there are many ones that might be, and several that work favorably under higher temperatures. Some residual insecticides will be more beneficial under climate change, while others will not be favored because of the speed at which they degrade (e.g. pyrethrins and pyrethroids). In terms of chemical control, fumigants generally increase in performance with increasing temperature, especially phosphine.
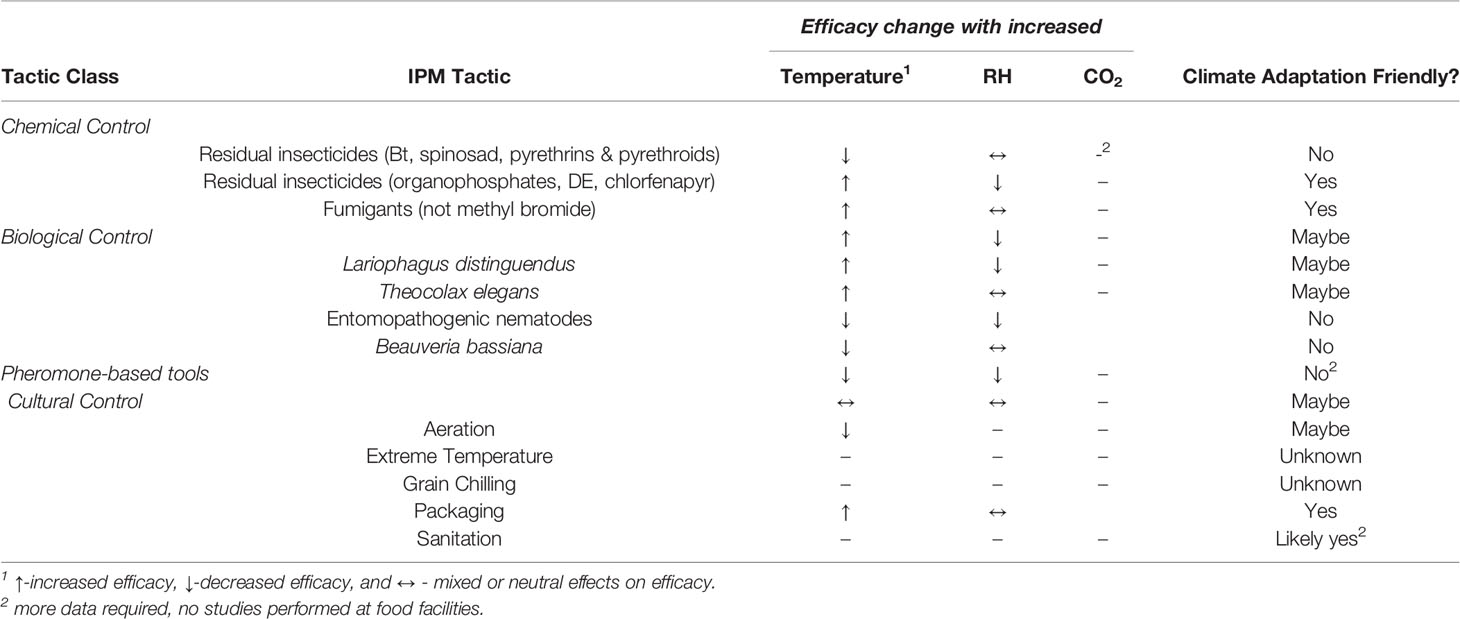
Table 1 Summary of climate adaptation-friendly IPM tactics at food facilities based on prior literature.
Research Gaps and Needs for the Mid-21st Century
The predicted impact on insects in the face of climate change is real and ongoing. Pest insects are well-known to adapt quickly to many biotic and abiotic changes, including insecticide resistance, thermal stress, and disturbance levels. Climate change has a particularly complex role for insect pests in stored products as they survive and thrive in both indoor and outdoor environments. There are still fundamental gaps in elucidating the risks posed by, optimal mitigation measures to, and improved resiliency strategies to climate change by food facilities. In better understanding risks, improved species forecasting for stored product insects under varying climate change scenarios should be executed to deliver better predictions of potential changes in distributions, especially for P. truncatus and T. granarium. These models should take into account not just bioclimatic variables, but niche components where possible and relevant. As species potentially shift distributions, it will become increasingly important to evaluate and predict species interactions (e.g. (Athanassiou et al., 2017; Quellhorst et al., 2020; Gourgouta et al., 2021; Lampiri et al., 2022; Baliota et al., 2022). However, the next step beyond individual pairwise tests of species in the lab will be modelling species interactions in 2-way or more interactions (Sutherst et al., 2007; Andrew and Hill, 2017) under projected scenarios of climate change. Few models have also attempted to predict how microclimate will specifically change at food facilities under climate change for the kinds of habitats that are present in these environments.
However, continued evaluation of insect response to different insecticides at different temperatures can provide thermal response curves for survivorship and effectiveness over time. By understanding effectiveness at different temperatures, managers may also be able to stave off adaptation and resistance to some insecticides by ensuring effectiveness. Properly knowing when to treat a commodity before it reaches a given thermal threshold can provide managers guidelines for successful use of insecticides. In addition, understanding the distribution of insecticides by fumigation or aerosols at different temperatures can provide predictions for spraying or treatment given a floor plan.
One neglected area of research has been to evaluate how IPM tactics at food facilities, including new potential insecticides, may contribute to greenhouse gas emissions so that climate friendly solutions can be incorporated into pest management programs. Bruce et al. 2022 recently showed how a greenhouse gas emission life cycle analysis can be incorporated early in the development of new potential insecticides using commercial alternatives as standards of comparison. Developing this sort of new dataset will be useful in order to ameliorate the ecological footprint of food facilities on their surrounding environments, while helping society reach its emissions targets.
While there has been a fair amount of work evaluating the effectiveness of certain IPM tactics (e.g. insecticides, biological control, grain aeration) under different temperatures and relative humidities as surrogates for climate change, this work should be expanded to a greater range of IPM tactics. Specific IPM tactics where more information is required about performance at elevated external temperatures under climate change include pheromone-based tools (monitoring and management tools), sanitation, grain chilling, and the application of extreme temperatures at food facilities. For example, it is unknown whether the increased stress on grain chilling equipment from higher external ambient temperatures may make grain chilling infeasible, or whether it may make it more difficult to use extreme temperatures. It is also unknown how exactly poor sanitation may interact with climate change factors like increased temperature; it is possible that insects may do better under poorly sanitized environments and higher temperatures because many of their essential processes would be sped up and there would be increased access to food. In addition, except for work evaluating CO2 as a modified atmosphere treatment (e.g. Navarro 2012; Vassilakos et al., 2019), no work has looked at elevated levels of CO2 (at levels relevant for climate change) on other IPM tactic efficacy after harvest. This represents a black box where very little is known about ramifications for stored product IPM tactics. It is possible that for some tactics such as contact insecticides there may be very little effect, while for others that depend on respiration such as fumigants or pheromone-based tools (e.g. where respiration, flight or diffusion of volatiles through air may be involved), there may be a much greater effect.
Along with monitoring for the presence and absence of individual species, population-level molecular data can provide information on migration and local adaptation in response to climate change (Cordeiro et al., 2016). Tracking populations with molecular markers can provide a more comprehensive monitoring picture and yield predictive patterns of emergence and infestation risk as climates change. Research on molecular markers associated with thermal tolerance can also provide information on the adaptation and changes associated with climate shifts and can provide useful information on long-term patterns of colonization of insect populations (Fallis et al., 2011).
There is also a lack of work detailing changes in microclimates within food facilities and bulk grain and understanding how climate change can impact stored product insect pests. Empirically collected data on rates of temperature change in bulk or bagged grain under different thermal regimes can be used as a baseline for modeling changes in temperature and risk of infestation by insects. Combined with thermal reaction norms and population growth demographics for stored product insects, these data can be modeled on various scales and climate change scenarios. Extensions of past programs like Stored Grain Advisor Pro (Flinn et al., 2007) can use these data to make better predictions about when to treat or when to expect populations to start growing and extended for use by stakeholders.
Precision agriculture may support climate adaptation throughout the postharvest supply chain. For example, in addition to improved forecasting for pest populations, it will become increasingly important to forecast successful fumigations to prevent the development of resistance. Extreme weather events may adversely affect fumigation, so being able to tie in local abiotic conditions to model outputs for pest managers in charge of fumigations will become increasingly important. Furthermore, monitoring traps that feed information instantly into the cloud so they are at the food facility manager’s fingertips will also be important, and other technology that automatizes processes that used to be labor intensive. Ultimately, information on the risks, strategies, and improvements to IPM will only be valuable insofar as it is regularly communicated in an easy-to-digest manner to stakeholders. Thus, throughout the process of developing new information on climate change and its impacts in the agricultural supply chain, there needs to be frequent dialogue between researchers, industry personnel, and stakeholders.
Author Contributions
AG and WM participated equally in the conception, outlining, writing, editing, and approval of this MS.
Funding
This work was funded, in part, by a United States Department of Agriculture, National Institute of Food and Agriculture, Crop Protection and Pest Management Grant#2020-70006-33000.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the excellent technical assistance of Shawn Chen (USDA), Samantha Gillette (KSU), Madison Lillich (KSU), and the rest of the Morrison and Gerken Laboratories. The use of trade names is for the purposes of providing scientific information only and does not constitute endorsement by the United States Department of Agriculture. The USDA is an equal opportunity employer.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2022.918845/full#supplementary-material
References
Adler C., Athanassiou C. G., Carvalho M. O., Emecki M., Gvozdenac S., Hamel D., et al. (2022). Changes in the Distribution and Pest Risk of Stored Product Insects in Europe Due to Global Warming: Need for Pan-European Pest Monitoring and Improved Food-Safety. J. Stor. Prod. Res. 97, 101977. doi: 10.1016/j.jspr.2022.101977
Ahmad F., Sarfraz M., Hassan M. (2000). Efficacy of Various Phosphine Concentrations and Temperature Combinations Against Different Strains of Trogoderma Granarium. Pakistan Entomol. 22, 91–94.
Akello J., Ortega-Beltran A., Katati B., Atehnkeng J., Augusto J., Mwila C. M., et al. (2021). Prevalence of Aflatoxin-And Fumonisin-Producing Fungi Associated With Cereal Crops Grown in Zimbabwe and Their Associated Risks in a Climate Change Scenario. Foods 10, 287. doi: 10.3390/foods10020287
Andrew N. R., Hill S. J. (2017). ““Effect of Climate Change on Insect Pest Management,”,” in Environmental Pest Management:Challenges for Agronomists, Ecologists, Economists and Policymakers. Eds. Coll M., Wajnberg E. (Hoboken, NJ: John Wiley & Sons Ltd), 195–223. doi: 10.1002/9781119255574.ch9
Arthur F. H. (1999). Effect of Temperature on Residual Toxicity of Cyfluthrin Wettable Powder. J. Econ. Entomol. 92, 695–699. doi: 10.1093/jee/92.3.695
Arthur F. H. (2012). Aerosols and Contact Insecticides as Alternatives to Methyl Bromide in Flour Mills, Food Production Facilities, and Food Warehouses. J. Pest Sci. 85, 323–329. doi: 10.1007/s10340-012-0439-9
Arthur F. H., Campbell J. F., Toews M. D. (2014). Distribution, Abundance, and Seasonal Patterns of Stored Product Beetles in a Commercial Food Storage Facility. J. Stored Prod. Res. 56, 21–32. doi: 10.1016/j.jspr.2013.11.003
Arthur F. H., Dowdy A. K. (2002). Impact of High Temperatures on Efficacy of Cyfluthrin and Hydroprene Applied to Concrete to Control Tribolium Castaneum (Herbst). J. Stored Prod. Res. 39, 193–204. doi: 10.1016/S0022-474X(01)00053-4
Arthur F. H., Flinn P. W. (2000). Aeration Management for Stored Hard Red Winter Wheat: Simulated Impact on Rusty Grain Beetle (Coleoptera: Cucujidae) Populations. J. Econ. Entomol. 93, 1364–1372. doi: 10.1603/0022-0493-93.4.1364
Arthur F. H., Morrison W. R., Morey A. (2019). Modeling the Potential Range Expansion of Larger Grain Borer, Prostephanus Truncatus (Coleoptera: Bostrichidae). Sci. Rep. 9, 6862. doi: 10.1038/s41598-019-42974-5
Arthur F. H., Morrison W. R., Trdan S. (2020). Feasibility of Using Aeration to Cool Wheat Stored in Slovenia: A Predictive Modeling Approach Using Historical Weather Data. Appl. Sci. 10, 6066. doi: 10.3390/app10176066
Arthur F. H., Throne J. E., Simonaitis R. A. (1992). Degradation and Biological Efficacy of Chlorpyrifos-Methyl on Wheat Stored at Five Temperatures and Three Moisture Contents. J. Econ. Entomol. 85, 1994–2002. doi: 10.1093/jee/85.5.1994
Ashaye O. A. (2018). “Post Harvest Handling of Food Crops in a Changing Climate : An Outlook on Food Research in Iar & T,” in 2018 Annual In-House Review Exercise of the Institute of Agricultural Research &Training (Moor- Plantation Ibadan), 1–41.
Atanda S. A., Pessu P. O., Isong I. U. (2011). The Concepts and Problems of Post–Harvest Food Losses in Perishable Crops. Afr. J. Food Sci. 5, 603–613. doi: 10.5897/AJFS.9000281
Athanassiou C. G., Kavallieratos N. G., Campbell J. F. (2017). Competition of Three Species of Sitophilus on Rice and Maize. PLoS One 12, e0173377. doi: 10.1371/journal.pone.0173377
Athanassiou C., Kavallieratos N. G., Yiatilis A. E., Vayias B. J., Mavrotas C. S., Tomanovic Z. (2008). Influence of Temperature and Humidity on the Efficacy of Spinosad Against Four Stored-Grain Beetle Species. J. Insect Sci. 8, 60. doi: 10.1673/031.008.6001
Athanassiou C. G., Phillips T. W., Wakil W. (2018). Biology and Control of the Khapra Beetle , Trogoderma Granarium, a Major Quarantine Threat to Global Food Security. Annu. Rev. Entomol. 64, 1–18. doi: 10.1146/annurev-ento-011118-111804
Athanassiou C. G., Vayias B. J., Dimizas C. B., Kavallieratos N. G., Papagregoriou A. S., Buchelos C. T. (2005). Insecticidal Efficacy of Diatomaceous Earth Against Sitophilus Oryzae (L.) (Coleoptera: Curculionidae) and Tribolium Confusum Du Val (Coleoptera: Tenebrionidae) on Stored Wheat: Influence of Dose Rate, Temperature and Exposure Interval. J. Stored Prod. Res. 41, 47–55. doi: 10.1016/j.jspr.2003.12.001
Aulicky R., Stejskal V., Frydova B., Athanassiou C. G. (2015). Susceptibility of Two Strains of the Confused Flour Beetle (Coleoptera: Tenebrionidae) Following Phosphine Structural Mill Fumigation: Effects of Concentration, Temperature, and Flour Deposits. J. Econ. Entomol. 108, 2823–2830. doi: 10.1093/jee/tov257
Bale J. S., Masters G. J., Hodkinson I. D., Awmack C., Bezemer T. M., Brown V. K., et al. (2002). Herbivory in Global Climate Change Research: Direct Effects of Rising Temperature on Insect Herbivores. Glob. Change Biol. 8, 1–16. doi: 10.1046/j.1365-2486.2002.00451.x
Baliota G. V., Scheff D. S., Morrison W. R. III, Athanassiou C. G. (2022). Competition Between Prostephanus Truncatus and Sitophilus Oryzae on Maize: The Species That Gets There First Matters. Bull. Entomol. Res.. doi: 10.1017/S000748532100105X
Basche A. D., Archontoulis S. V., Kaspar T. C., Jaynes D. B., Parkin T. B., Miguez F. E. (2016). Simulating Long-Term Impacts of Cover Crops and Climate Change on Crop Production and Environmental Outcomes in the Midwestern United States. Agric. Ecosyst. Environ. 218, 95–106. doi: 10.1016/j.agee.2015.11.011
Battilani P., Toscano P., van der Fels-Klerx H. J., Moretti A., Camardo Leggieri M., Brera C., et al. (2016). Aflatoxin B1 Contamination in Maize in Europe Increases Due to Climate Change. Sci. Rep. 6, 24328. doi: 10.1038/srep24328
Batts G. R., Morison J. I. L., Ellis R. H., Hadley P., Wheeler T. R. (1997). Effects of CO2 and Temperature on Growth and Yield of Crops of Winter Wheat Over Four Seasons. Eur. J. Agron. 7, 43–52. doi: 10.1016/S1161-0301(97)00022-1
Binzer A., Guill C., Brose U., Rall B. C. (2012). The Dynamics of Food Chains Under Climate Change and Nutrient Enrichment. Philos. Trans. R. Soc B Biol. Sci. 367, 2935–2944. doi: 10.1098/rstb.2012.0230
Bradna J., Šimon J., Hájek D., Vejchar D. (2018). The Impact of Weather Conditions on Microclimate in Storage Facilities. Agron. Res. 16, 1580–1589. doi: 10.15159/AR.18.178
Bruce A., Wilson A. N., Ranabhat S., Montgomery J., Nicholson S., Harris K., et al. (2022). A Biomass Pyrolysis Oil as a Novel Insect Growth Regulator Mimic for a Variety of Stored Product Beetles. J. Econ. Entomol 115 (3), 877–87. doi: 10.1093/jee/toac017
Campbell J. F., Arbogast R. T. (2004). Stored-Product Insects in a Flour Mill: Population Dynamics and Response to Fumigation Treatments. Entomol. Exp. Appl. 112, 217–225. doi: 10.1111/j.0013-8703.2004.00197.x
Campbell J. F., Toews M. D., Arthur F. H., Arbogast R. T. (2010). Long-Term Monitoring of Tribolium Castaneum in Two Flour Mills: Seasonal Patterns and Impact of Fumigation. J. Econ. Entomol. 103, 991–1001. doi: 10.1603/EC09347
Campolo O., Giunti G., Russo A., Palmeri V., Zappala L. (2018). Essential Oils in Stored Product Insect Pest Control. J. Food Qual 2018, 6906105. doi: 10.1155/2018/6906105
Cappelli G. A., Bregaglio S. (2021). Model-Based Evaluation of Climate Change Impacts on Rice Grain Quality in the Main European Rice District. Food Energy Secur. 10, e307. doi: 10.1002/fes3.307
Cardé R. T. (2021). Navigation Along Windborne Plumes of Pheromone and Resource-Linked Odors. Annu. Rev. Entomol. 66, 317–336. doi: 10.1146/annurev-ento-011019-024932
Casada M. E., Young J. H. (1994). Model for Heat and Moisture Transfer in Arbitrarily Shaped Two-Dimensional Porous Media. Trans. Am. Soc Agric. Biol. Eng. 94, 1927–1938. doi: 10.13031/2013.28285
Chadalavada K., Guna K., Kumari B. R., Kumar T. S. (2022). ““Drought Stress in Sorghum: Impact on Grain Quality,”,” in Climate Change and Crop Stress Eds. Shanker A. K., Shanker C., Anand A., Maheswari M. (Cambridge, MA: Academic Press), 113–134. doi: 10.1016/b978-0-12-816091-6.00003-1
Child R. E. (2007). ““Insect Damage as a Function of Climate,”,” in Museum Microclimates. Eds. Padfield T., Borchersen K. (Copenhagen, Denmark: National Museum of Denmark), 57–60. Available at: http://natmus.dk/fileadmin/user_upload/natmus/bevaringsafdelingen/billeder/far/Museum_Microclimate/Contributions_to_the_conference/child.pdf%5Cnhttp://eprints.sparaochbevara.se/158/1/child.pdf.
Cook D. A., Armitage D. M., Wildey K. B. (2004). What are the Implications of Climate Change for Integrated Pest Management of Stored Grain in the UK? In Proceedings of the Integrated Protection of Stored Products IOBC/wprs Bulletin, Vol. Vol. 27. Eds. Navarro S., Adler C., Riudavets J., Stejskal V. Kusadasi Turkey: IOBC wprs Bulletin, 1–11.
Cordeiro E. M. G., Campbell J. F., Phillips T. W. (2016). Movement and Orientation Decision Modeling of Rhyzopertha Dominica (Coleoptera: Bostrichidae) in the Grain Mass. Environ. Entomol. 45, 410–419. doi: 10.1093/ee/nvv232
Cork A., De Souza K., Hall D. R., Jones O. T., Casagrande E., Krishnaiah K., et al. (2008). Development of PVC-Resin-Controlled Release Formulation for Pheromones and Use in Mating Disruption of Yellow Rice Stem Borer, Scirpophaga Incertulas. Crop Prot. 27, 248–255. doi: 10.1016/j.cropro.2007.05.011
Davis K. F., Downs S., Gephart J. A. (2021). Towards Food Supply Chain Resilience to Environmental Shocks. Nat. Food. 2 (1), 54–65. doi: 10.1038/s43016-020-00196-3
Deutsch C. A., Tewksbury J. J., Tigchelaar M., Battisti D. S., Merrill S. C., Huey R. B., et al. (2018). Increase in Crop Losses to Insect Pests in a Warming Climate. Sci. (80-. ). 361, 916–919. doi: 10.1126/science.aat3466
Doud C. W., Phillips T. W. (2000). Activity of Plodia Interpunctella (Lepidoptera: Pyralidae) in and Around Flour Mills. J. Econ. Entomol. 93, 1842–1847. doi: 10.1603/0022-0493-93.6.1842
Dukes J. S., Pontius J., Orwig D., Garnas J. R., Rodgers V. L., Brazee N., et al. (2009). Responses of Insect Pests, Pathogens, and Invasive Plant Species to Climate Change in the Forests of Northeastern North America: What can We Predict? Can. J. For. Res. 39, 231–248. doi: 10.1139/X08-171
Easterling W. E., Crosson P. R., Rosenberg N. J., McKenney M. S., Katz L. A., Lemon K. M. (1993). Paper 2. Agricultural Impacts of and Responses to Climate Change in the Missouri-Iowa-Nebraska-Kansas (MINK) Region. Clim. Change 24, 23–61. doi: 10.1007/BF01091476
Eggen M., Ozdogan M., Zaitchik B., Ademe D., Foltz J., Simane B. (2019). Vulnerability of Sorghum Production to Extreme, Sub-Seasonal Weather Under Climate Change. Environ. Res. Lett. 14, 045005. doi: 10.1088/1748-9326/aafe19
Elmouttie D., Kiermeier A., Hamilton G. (2010). Improving Detection Probabilities for Pests in Stored Grain. Pest Manage. Sci. 66, 1280–1286. doi: 10.1002/ps.2009
El-Sayed A. M., Ganji S., Gross J., Giesen N., Rid M., Lo P. L., et al. (2021). Climate Change Risk to Pheromone Application in Pest Management. Sci. Nat. 108, 47. doi: 10.1007/s00114-021-01757-7
Estay S. A., Lima M., Labra F. A. (2009). Predicting Insect Pest Status Under Climate Change Scenarios: Combining Experimental Data and Population Dynamics Modelling. J. Appl. Entomol. 133, 491–499. doi: 10.1111/j.1439-0418.2008.01380.x
Fallis L. C., Fanara J. J., Morgan T. J. (2011). Genetic Variation in Heat-Stress Tolerance Among South American Drosophila Populations. Genetica 139, 1331–1337. doi: 10.1007/s10709-012-9635-z
Fei C. J., McCarl B. A., Thayer A. W. (2017). Estimating the Impacts of Climate Change and Potential Adaptation Strategies on Cereal Grains in the United States. Front. Ecol. Evol. 5. doi: 10.3389/fevo.2017.00062
Fields P. G. (1992). The Control of Stored-Product Insects and Mites With Extreme Temperatures. J. Stored Prod. Res. 28, 89–118. doi: 10.1016/0022-474X(92)90018-L
Fields P., Korunic Z. (2000). The Effect of Grain Moisture Content and Temperature on the Efficacy of Diatomaceous Earths From Different Geographical Locations Against Stored-Product Beetles. J. Stored Prod. Res. 36, 1–13. doi: 10.1016/S0022-474X(99)00021-1
Flinn P. W., Arthur F. H., Throne J. E., Friesen K. S., Hartzer K. L. (2015). Cold Temperature Disinfestation of Bagged Flour. J. Stored Prod. Res. 63, 42–46. doi: 10.1016/j.jspr.2015.06.001
Flinn P. W., Hagstrum D.W., Reed C.R., Phillips T. W. (2007). Stored Grain Advisor Pro: Decision Support System for Insect Management in Commercial Grain Elevators. J. Stored Prod. Res. 43, 375–383. doi: 10.1016/j.jspr.2006.09.004
Flinn P. W., Hagstrum D. W. (2002). Temperature-Mediated Functional Response of Theocolax Elegans (Hymenoptera: Pteromalidae) Parasitizing Rhyzopertha Dominica (Coleoptera: Bostrichidae) in Stored Wheat. J. Stored Prod. Res. 38, 185–190. doi: 10.1016/S0022-474X(01)00019-4
Flinn P., Phillips T., Hagstrum D., Arthur F., Throne J. (2001). “Modeling the Effects of Insect Stage and Grain Temperature on Phosphine-Induced Mortality for Rhyzopertha Dominica,” in International Conference Controlled Atmosphere and Fumigation in Stored Products, Fresno, CA, 29 Oct. - 3 Nov. 2000 Donahaye E. J., Navarro S., Leesch J. G. CA, U.S.A: Executive Printing Services, Clovis. 531–39.
Gerken A. R., Campbell J. F. (2019). Using Long-Term Capture Data to Predict Trogoderma Variabile Ballion and Plodia Interpunctella (Hübner) Population Patterns. Insects 10, 93. doi: 10.3390/insects10040093
Gerken A. R., Campbell J. F. (2021). Spatial and Temporal Variation in Stored-Product Insect Pest Distributions and Implications for Pest Management in Processing and Storage Facilities. Ann. Entomol. Soc Am. 115:239–52. doi: 10.1093/aesa/saab049
Giménez V. D., Serrago R. A., García G. A., Miralles D. J. (2021). How Milling and Breadmaking Quality are Modified by Warmer Nights in Wheat? J. Cereal Sci. 102, 103343. doi: 10.1016/j.jcs.2021.103343
Goufo P., Falco V., Brites C., Wessel D. F., Kratz S., Rosa E. A. S., et al. (2014). Effect of Elevated Carbon Dioxide Concentration on Rice Quality: Nutritive Value, Color, Milling, Cooking, and Eating Qualities. Cereal Chem. 91, 513–521. doi: 10.1094/CCHEM-12-13-0256-R
Gourgouta M., Baliota G. V., Morrison W. R., Domingue M. J., Athanassiou C. G. (2021). Comparative Capture of Trogoderma Granarium (Coleoptera: Dermestidae) and T. Variabile in Floor Traps in Single Species Releases With Previously Captured Conspecific or Heterospecific Individuals. J. Econ. Entomol. 114, 2591–2597. doi: 10.1093/jee/toab174
Gregory P. J., Ingram J. S. I., Brklacich M. (2005). Climate Change and Food Security. Philos. Trans. R. Soc B Biol. Sci. 360, 2139–2148. doi: 10.1098/rstb.2005.1745
Gu S., Han P., Ye Z., Perkins L. E., Li J., Wang H., et al. (2018). Climate Change Favours a Destructive Agricultural Pest in Temperate Regions: Late Spring Cold Matters. J. Pest Sci. 91, 1191–1198. doi: 10.1007/s10340-018-1011-z
Hagstrum D. W., Flinn P. W., Subramanyam B. (1998). Predicting Insect Density From Probe Trap Catch in Farm-Stored Wheat. J. Stored Prod. Res. 34, 251–262. doi: 10.1016/S0022-474X(98)00006-X
Hansen L. S., Steenberg T. (2007). Combining Larval Parasitoids and an Entomopathogenic Fungus for Biological Control of Sitophilus Granarius (Coleoptera: Curculionidae) in Stored Grain. Biol. Control 40, 237–242. doi: 10.1016/j.biocontrol.2006.09.012
Heuskin S., Verheggen F. J., Haubruge E., Wathelet J.-P., Lognay G. (2011). The Use of Semiochemical Slow-Release Devices in Integrated Pest Management Strategies. Biotechnol. Agron. Soc Environ. 15, 459–470.
Hijmans R. J. (2003). The Effect of Climate Change on Global Potato Production. Am. J. Potato Res. 80, 271–280. doi: 10.1007/BF02855363
Ileleji K. E., Maier D. E., Woloshuk C. P. (2007). Evaluation of Different Temperature Management Strategies for Suppression of Sitophilus Zeamais (Motschulsky) in Stored Maize. J. Stored Prod. Res. 43, 480–488. doi: 10.1016/j.jspr.2007.01.002
Impa S. M., Perumal R., Bean S. R., John Sunoj V. S., Jagadish S. V. K. (2019). Water Deficit and Heat Stress Induced Alterations in Grain Physico-Chemical Characteristics and Micronutrient Composition in Field Grown Grain Sorghum. J. Cereal Sci. 86, 124–131. doi: 10.1016/j.jcs.2019.01.013
Jian F., Jayas D. S., White N. D. G., Alagusundaram K. (2005). A Three-Dimensional, Asymmetric, and Transient Model to Predict Grain Temperatures in Grain Storage Bins. Trans. Am. Soc Agric. Biol. Eng. 48, 263–271. doi: 10.13031/2013.17927
Jia F., Toews M., Campbell J. F., Ramaswamy S. B. (2008). Survival and Reproduction of Lesser Grain Borer, Rhyzopertha Dominica (F.) (Coleoptera: Bostrichidae) on Flora Associated With Native Habitats in Kansas. J. Stor. Prod. Res. 44, 366–372. doi: 10.1016/j.jspr.2008.06.001
Kansman J. T., Crowder D. W., Finke D. L. (2021). Primacy of Plants in Driving the Response of Arthropod Communities to Drought. Oecologia 195, 833–842. doi: 10.1007/s00442-020-04844-0
Kavallieratos N. G., Athanassiou C. G., Hatzikonstantinou A. N. N. N., Kavallieratou H. N. (2011). Abiotic and Biotic Factors Affect Efficacy of Chlorfenapyr for Control of Stored-Product Insect Pests. J. Food Prot. 74, 1288–1299. doi: 10.4315/0362-028X.JFP-10-575
Kavallieratos N. G., Athanassiou C. G., Vayias B. J., Betsi P. C. C. (2010). Insecticidal Efficacy of Fipronil Against Four Stored-Product Insect Pests: Influence of Commodity, Dose, Exposure Interval, Relative Humidity and Temperature. Pest Manage. Sci. 66, 640–649. doi: 10.1002/ps.1923
Kerr R. A. (2003). A Perfect Ocean for Four Years of Globe-Girdling Drought. Science 299, 636. doi: 10.1126/science.299.5607.636
Kinsinger R. A., McGauhey W. H. (1976). Stability of Bacillus Thuringiensis and a Granulosis Virus of Plodia Interpunctella on Stored Wheat. J. Econ. Entomol. 69, 149–154. doi: 10.1093/jee/69.2.149
Ladányi M., Horváth L. (2010). A Review of the Potential Climate Change Impact on Insect Populations - General and Agricultural Aspects. Appl. Ecol. Environ. Res. 8, 143–152. doi: 10.15666/aeer/0802_143151
Lampiri E., Baliota G. V., Morrison W. R., Domingue M. J., Athanassiou C. G. (2022). Comparative Population Growth of the Khapra Beetle (Coleoptera: Dermestidae) and the Warehouse Beetle (Coleoptera: Dermestidae) on Wheat and Rice. J. Econ. Entomol. 115, 344–352. doi: 10.1093/jee/toab209
Lanka S. K., Arthur F. H., Campbell J. F., Yan Zhu K. (2019). Evaluation of Residual Efficacy of Pyrethrin + Methoprene Aerosol on Two Dermestids: Impact of Particle Size, Species, and Temperature. Insects 10, 142. doi: 10.3390/insects10050142
Liu K., Zhang C., Xu J., Liu Q. (2021). Research Advance in Gas Detection of Volatile Organic Compounds Released in Rice Quality Deterioration Process. Comp. Rev. Food Sci. 20, 5802–5828. doi: 10.1111/1541-4337.12846
Lobell D. B., Asner G. P. (2003). Climate and Management Contributions to Recent Trends in U.S. Agricultural Yields. Science 299, 1032. doi: 10.1126/science.1078475
Logan J. D., Wolesensky W. (2007). Accounting for Temperature in Predator Functional Responses. Nat. Resour. Model. 20, 549–574. doi: 10.1111/j.1939-7445.2007.tb00220.x
Lombardero M. J., Ayres M. P., Ayres B. D., Reeve J. D. (2000). Cold Tolerance of Four Species of Bark Beetle (Coleoptera: Scolytidae) in North America. Environ. Entomol. 29, 421–432. doi: 10.1603/0046-225X-29.3.421
Lopes D. C., Steidle Neto A. J. (2019). Effects of Climate Change on the Aeration of Stored Beans in Minas Gerais State, Brazil. Biosyst. Eng. 188, 155–164. doi: 10.1016/j.biosystemseng.2019.10.010
Lyman N. B., Jagadish K. S. V., Nalley L. L., Dixon B. L., Siebenmorgen T. (2013). Neglecting Rice Milling Yield and Quality Underestimates Economic Losses From High-Temperature Stress. PloS One 8, e72157. doi: 10.1371/journal.pone.0072157
Machekano H., Mvumi B. M., Nyamukondiwa C. (2018). “Sitotroga Cerealella (Olivier) Resilience to Extreme Temperature and Desiccation may Explain its Increasing Pest Status in Changing Climates,” in Proceedings of the 12th International Working Conference on Stored Product Protection (IWCSPP), Berlin, Germany, October 7-11, 2018 Eds. Adler C. S., Opit G., Fürstenau B., Müller-Blenkle C., Kern P., Arthur F. H., et al 463, 165–171.
Magan N., Medina A., Aldred D. (2011). Possible Climate-Change Effects on Mycotoxin Contamination of Food Crops Pre- and Postharvest. Plant Pathol. 60, 150–163. doi: 10.1111/j.1365-3059.2010.02412.x
Mahroof R., Edde P. A., Robertson B., Puckette J. A., Phillips T. W. (2010). Dispersal of Rhyzopertha Dominica (Coleoptera: Bostrichidae) in Different Habitats. Environ. Entomol. 39, 930–938. doi: 10.1603/EN09243
Malhotra S. K. (2017). Horticultural Crops and Climate Change: A Review. Indian J. Agric. Sci. 87, 12–22.
Mansoor M. M., Afzal M., Raza A. B. M., Akram Z., Waqar A., Afzal M. B. S. (2015). Post-Exposure Temperature Influence on the Toxicity of Conventional and New Chemistry Insecticides to Green Lacewing Chrysoperla Carnea (Stephens) (Neuroptera: Chrysopidae). Saudi J. Biol. Sci. 22, 317–321. doi: 10.1016/j.sjbs.2014.10.008
Mariem S. B., Soba D., Zhou B., Loladze I., Morales F., Aranjuelo I. (2021). Climate Change, Crop Yields, and Grain Quality of C3 Cereals: A Meta-Analysis of [CO2], Temperature, and Drought Effects. Plants 10, 1052. doi: 10.3390/plants10061052
Mastilović J., Živančev D., Lončar E., Malbaša R., Hristov N., Kevrešan Ž. (2018). Effects of High Temperatures and Drought During Anthesis and Grain Filling Period on Wheat Processing Quality and Underlying Gluten Structural Changes. J. Sci. Food Agric. 98, 2898–2907. doi: 10.1002/jsfa.8784
Medina A., Akbar A., Baazeem A., Rodriguez A., Magan N. (2017). Climate Change, Food Security and Mycotoxins: Do We Know Enough? Fungal Biol. Rev. 31, 143–154. doi: 10.1016/j.fbr.2017.04.002
Medina Á., Rodríguez A., Magan N. (2015). Climate Change and Mycotoxigenic Fungi: Impacts on Mycotoxin Production. Curr. Opin. Food Sci. 5, 99–104. doi: 10.1016/j.cofs.2015.11.002
Milling C. R., Rachlow J. L., Olsoy P. J., Chappell M. A., Johnson T. R., Forbey J. S., et al. (2018). Habitat Structure Modifies Microclimate: An Approach for Mapping Fine-Scale Thermal Refuge. Methods Ecol. Evol. 9, 1648–1657. doi: 10.1111/2041-210X.13008
Mooney H., Larigauderie A., Cesario M., Elmquist T., Hoegh-Guldbergh O., Lavorel S., et al. (2009). Biodiversity, Climate Change, and Ecosystem Services. Curr. Opin. Environ. Sust. 1, 46–54. doi: 10.1016/j.cosust.2009.07.006
Moretti A., Pascale M., Logrieco A. F. (2019). Mycotoxin Risks Under a Climate Change Scenario in Europe. Trends Food Sci. Technol. 84, 38–40. doi: 10.1016/j.tifs.2018.03.008
Moretti C. L., Mattos L. M., Calbo A. G., Sargent S. A. (2010). Climate Changes and Potential Impacts on Postharvest Quality of Fruit and Vegetable Crops: A Review. Food Res. Int. 43, 1824–1832. doi: 10.1016/j.foodres.2009.10.013
Morgounov A., Sonder K., Abugalieva A., Bhadauria V., Cuthbert R. D., Shamanin V., et al. (2018). Effect of Climate Change on Spring Wheat Yields in North America and Eurasia in 1981-2015 and Implications for Breeding. PLoS One 13, e0204932. doi: 10.1371/journal.pone.0204932
Morrison W. R., Arthur F. H., Athanassiou C. G., Lampiri E., Wilson L. T., Yang Y., et al. (2022). Modeling of Sitophilus Oryzae (L.) (Coleoptera: Curculionidae) Based on Historical Weather Data Indicates Aeration is Effective for Management of Wheat Stored in Greece. Comput. Electron. Agric 197, 106926. doi: 10.1016/j.compag.2022.106926
Morrison W. R., Arthur F. H., Wilson L. T., Yang Y., Wang J., Athanassiou C. G. (2020). Aeration to Manage Insects in Wheat Stored in the Balkan Peninsula: Computer Simulations Using Historical Weather Data. Agronomy 10, 1927. doi: 10.3390/agronomy10121927
Morrison III, W. R., Bruce A., Wilkins R. V., Albin C., Arthur F. H. (2019). Sanitation Improves Stored Product Pest Management. Insects 10, 77. doi: 10.3390/insects10030077
Morrison W. R., Scully E. D., Campbell J. F. (2021). Towards Developing Areawide Semiochemical-Mediated, Behaviorally-Based Integrated Pest Management Programs for Stored Product Insects. Pest Manage. Sci. 77, 2667–2682. doi: 10.1002/ps.6289
Moses J. A., Jayas D. S., Alagusundaram K. (2015). Climate Change and its Implications on Stored Food Grains. Agric. Res. 4, 21–30. doi: 10.1007/s40003-015-0152-z
Mpofu P., Cuthbert R. N., Machekano H., Nyamukondiwa C. (2022). Transgenerational Responses to Heat and Fasting Acclimation in the Angoumois Grain Moth. J. Stor. Prod. Res. 97, 101979. doi: 10.1016/j.jspr.2022.101979
Musser F. R., Shelton A. M. (2005). The Influence of Post-Exposure Temperature on the Toxicity of Insecticides to Ostrinia Nubilalis (Lepidoptera: Crambidae). Pest Manage. Sci. 61, 508–510. doi: 10.1002/ps.998
Mutamiswa R., Machekano H., Singano C., Joseph V., Chidawanyika F., Nyamukondiwa C. (2021). Desiccation and Temperature Resistance of the Larger Grain Borer, Prostephanus Truncatus (Horn) (Coleoptera: Bostrichidae): Pedestals for Invasion Success? Physiol. Entomol. 46, 157–166. doi: 10.1111/phen.12355
Navarro S. (2012). The Use of Modified and Controlled Atmospheres for the Disinfestation of Stored Products. J. Pest Sci. 85, 301–322. doi: 10.1007/s10340-012-0424-3
Nayak M. K., Collins P. J., Pavica H. (2003). “Effect of Fumigation Temperature on the Efficacy of Phospine Against Strongly Resistant Psocids Liposcelis Bostrychophila (Psocoptera: Liposcelididae),” in Proceedings of the 8th International Working Conference on Stored Product Protection, York, UK, 22-26 July 2002. Eds. Credland P. F., Armitage D. M., Bell C. H., Cogan P. M., Highley E. 1–2. doi: 10.1079/9780851996912.0654
Njoroge A. W., Affognon H. D., Mutungi C. M., Manono J., Lamuka P. O., Murdock L. L. (2014). Triple Bag Hermetic Storage Delivers a Lethal Punch to Prostephanus Truncatus (Horn) (Coleoptera: Bostrichidae) in Stored Maize. J. Stored Prod. Res. 58, 12–19. doi: 10.1016/j.jspr.2014.02.005
Norment B. R., Chambers H. W. (1970). Temperature Relationships in Organophosphorus Poisoning in Boll Weevils. J. Econ. Entomol. 63, 502–504. doi: 10.1093/jee/63.2.502
Nuttall J. G., O’Leary G. J., Panozzo J. F., Walker C. K., Barlow K. M., Fitzgerald G. J. (2017). Models of Grain Quality in Wheat—A Review. F. Crop Res. 202, 136–145. doi: 10.1016/j.fcr.2015.12.011
Nyabako T., Mvumi B. M., Stathers T. E., Machekano H. (2021). Smallholder Grain Postharvest Management in a Variable Climate: Practices and Perceptions of Smallholder Farmers and Their Service-Providers in Semi-Arid Areas. Environ. Dev. Sustain. 23, 9196–9222. doi: 10.1007/s10668-020-01019-y
Omodara M., Montross M., Mcneill S., Besson R. (2020). “Impact of Storage Environment on the Micro-Climate and Grain Moisture in Polypropylene Storage Bags,” in 2020 ASABE Annual International, Meeting Sponsored by ASABE Omaha, Nebraska, July 12–15, 2020. 1–10. doi: 10.13031/aim.202000071
Ortiz R., Sayre K. D., Govaerts B., Gupta R., Subbarao G. V., Ban T., et al. (2008). Climate Change: Can Wheat Beat the Heat? Agric. Ecosyst. Environ. 126, 46–58. doi: 10.1016/j.agee.2008.01.019
Pang Y., Tian Y., Fu C., Wang B., Li J., Ren Y., et al. (2018). Variability of Coastal Cephalopods in Overexploited China Seas Under Climate Change With Implications on Fisheries Management. Fish. Res. 208, 22–33. doi: 10.1016/j.fishres.2018.07.004
Parajuli R., Thoma G., Matlock M. D. (2019). Environmental Sustainability of Fruit and Vegetable Production Supply Chains in the Face of Climate Change: A Review. Sci. Total Environ. 650, 2863–2879. doi: 10.1016/j.scitotenv.2018.10.019
Paterson R. R. M., Lima N. (2010). How Will Climate Change Affect Mycotoxins in Food? Food Res. Int. 43, 1902–1914. doi: 10.1016/j.foodres.2009.07.010
Patterson D. T., Westbrook J. K., Joyce R. J. V., Lingren P. D., Rogasik J. (1999). Weeds, Insects, and Diseases. Clim. Change 43, 711–727. doi: 10.1023/A:1005549400875
Perrone G., Ferrara M., Medina A., Pascale M., Magan N. (2020). Toxigenic Fungi and Mycotoxins in a Climate Change Scenario: Ecology, Genomics, Distribution, Prediction and Prevention of the Risk. Microorganisms 8, 1496. doi: 10.3390/microorganisms8101496
Plarre R., Burkholder W. E. (2009). “Stored Products as Habitats,” in Encyclopedia of Insects (Cambridge, MA: Academic Press), 967–970. doi: 10.1016/B978-0-12-374144-8.00254-X
Ponce M. A., Kim T. N., Morrison W. R. (2021). A Systematic Review of the Behavioral Responses by Stored-Product Arthropods to Individual or Blends of Microbially Produced Volatile Cues. Insects 12, 391. doi: 10.3390/insects12050391
Porter J. H., Parry M. L., Carter T. R. (1991). The Potential Effects of Climatic Change on Agricultural Insect Pests. Agric. For. Meteorol. 57, 221–240. doi: 10.1016/0168-1923(91)90088-8
Prasantha B. D. R., Hafeel R. F., Wimalasiri K. M. S., Pathirana U. P. D. (2014). End-Use Quality Characteristics of Hermetically Stored Paddy. J. Stored Prod. Res. 59, 158–166. doi: 10.1016/j.jspr.2014.07.003
Pražić Golić M., Andrić G., Kljajić P. (2016). Combined Effects of Contact Insecticides and 50°C Temperature on Sitophilus Oryzae (L.) (Coleoptera: Curculionidae) in Wheat Grain. J. Stored Prod. Res. 69, 245–251. doi: 10.1016/j.jspr.2016.09.004
Quellhorst H. E., Athanassiou C. G., Bruce A., Scully E. D., Morrison III W. R. (2020). Temperature-Mediated Competition Between the Invasive Larger Grain Borer (Coleoptera: Bostrichidae) and the Cosmopolitan Maize Weevil (Coleoptera: Curculionidae). Environ. Entomol. 49, 255–264. doi: 10.1093/ee/nvz151
Quellhorst H., Athanassiou C. G., Zhu K. Y., Morrison III W. R. (2021). The Biology, Ecology, and Management of the Larger Grain Borer, Prostephanus Truncatus (Horn) (Coleoptera: Bostrichidae). J. Stored Prod. Res 94, 101860. doi: 10.1016/j.jspr.2021.101860
Ray D. K., Gerber J. S., Macdonald G. K., West P. C. (2015). Climate Variation Explains a Third of Global Crop Yield Variability. Nat. Commun. 6, 5989. doi: 10.1038/ncomms6989
Rebaudo F., Faye E., Dangles O. (2016). Microclimate Data Improve Predictions of Insect Abundance Models Based on Calibrated Spatiotemporal Temperatures. Front. Physiol. 7. doi: 10.3389/fphys.2016.00139
Romo C. M., Tylianakis J. M. (2013). Elevated Temperature and Drought Interact to Reduce Parasitoid Effectiveness in Suppressing Hosts. PLoS One 8, e58136. doi: 10.1371/journal.pone.0058136
Rumbos C. I., Athanassiou C. G. (2017a). Use of Entomopathogenic Fungi for the Control of Stored-Product Insects: Can Fungi Protect Durable Commodities? J. Pest Sci. 90, 839–854. doi: 10.1007/s10340-017-0849-9
Rumbos C. I., Athanassiou C. G. (2017b). The Use of Entomopathogenic Nematodes in the Control of Stored-Product Insects. J. Pest Sci. 90, 39–49. doi: 10.1007/s10340-016-0795-y
Rumbos C. I., Dutton A. C., Athanassiou C. G. (2013). Comparison of Two Pirimiphos-Methyl Formulations Against Major Stored-Product Insect Species. J. Stored Prod. Res. 55, 106–115. doi: 10.1016/j.jspr.2013.08.003
Ryoo M. I., Hong Y. S., Yoo C. K. (1991). Relationship Between Temperature and Development of Lariophagus Distinguendus (Hymenoptera: Pteromalidae), an Ectoparasitoid of Sitophilus Oryzae (Coleoptera: Curculionidae). J. Econ. Entomol. 84, 825–829. doi: 10.1093/jee/84.3.825
Satpute N. S., Deshmukh S. D., Rao N. G. V., Tikar S. N., Moharil M. P., Nimbalkar S. A. (2007). Temperature-Dependent Variation in Toxicity of Insecticides Against Earias Vitella (Lepidoptera: Noctuidae). J. Econ. Entomol. 100, 357–360. doi: 10.1603/0022-0493(2007)100[357:TVITOI]2.0.CO;2
Scheff D. S., Campbell J. F., Arthur F. H. (2021). Seasonal, Landscape, and Attractant Effects on Lesser Grain Borer, Rhyzopertha Dominica (F.), Captures in Northeast Kansas. Agronomy 12, 99. doi: 10.3390/agronomy12010099
Sears M. W., Angilletta M. J. (2015). Costs and Benefits of Thermoregulation Revisited: Both the Heterogeneity and Spatial Structure of Temperature Drive Energetic Costs. Am. Nat. 185, E94–E102. doi: 10.1086/680008
Semeao A., Campbell J. F., Hutchinson J. M. S., Whitworth R. J., Sloderbeck P. E. (2013). Spatio-Temporal Distribution of Stored-Product Insects Around Food Processing and Storage Facilities. Agric. Ecosyst. Environ. 165, 151–162. doi: 10.1016/j.agee.2012.11.013
Sharma H. C. (2014). Climate Change Effects on Insects: Implications for Crop Protection and Food Security. J. Crop Improv. 28, 229–259. doi: 10.1080/15427528.2014.881205
Singano C. D., Mvumi B. M., Stathers T. E. (2019). Effectiveness of Grain Storage Facilities and Protectants in Controlling Stored-Maize Insect Pests in a Climate-Risk Prone Area of Shire Valley, Southern Malawi. J. Stored Prod. Res. 83, 130–147. doi: 10.1016/j.jspr.2019.06.007
Singano C. D., Mvumi B. M., Stathers T. E., Machekano H., Nyamukondiwa C. (2020). What Does Global Warming Mean for Stored-Grain Protection? Options for Prostephanus Truncatus (Horn) Control at Increased Temperatures. J. Stored Prod. Res. 85, 101532. doi: 10.1016/j.jspr.2019.101532
Sinha R. N. (1968). Seasonal Changes in Mite Populations in Rural Granaries in Japan. Ann. Entomol. Soc Am. 61, 938–949. doi: 10.1093/aesa/61.4.938
Sinha R. N. (1973). “Interrelations of Physical, Chemical, and Biological Variables in the Deterioration of Stored Grains,” in Grain Storage: Part of a System Eds. R. N. Sinha and W.E. Muir (Westport CT: Avi Publishing Co.) 15–47.
Smith D. L., Almaraz J. J. (2004). Climate Change and Crop Production: Contributions, Impacts, and Adaptations. Can. J. Plant Pathol. 26, 253–266. doi: 10.1080/07060660409507142
Smith W. N., Grant B. B., Desjardins R. L., Kroebel R., Li C., Qian B., et al. (2013). Assessing the Effects of Climate Change on Crop Production and GHG Emissions in Canada. Agric. Ecosyst. Environ. 179, 139–150. doi: 10.1016/j.agee.2013.08.015
Southworth J., Randolph J. C., Habeck M., Doering O. C., Pfeifer R. A., Rao D. G., et al. (2000). Consequences of Future Climate Change and Changing Climate Variability on Maize Yields in the Midwestern United States. Agric. Ecosyst. Environ. 82, 139–158. doi: 10.1016/S0167-8809(00)00223-1
Spochacz M., Chowanski S., Walkowiak-Nowicka K., Szymczak M., Adamski Z. (2018). Plant-Derived Substances Used Against Beetles–Pests of Stored Crops and Food–and Their Mode of Action: A Review. Comp. Rev. Food Sci. Saf. 17, 1339–1366. doi: 10.1111/1541-4337.12377
Srivastava Y. (2019). “Climate Change: A Challenge for Postharvest Management, Food Loss, Food Quality, and Food Security,” in Climate Change and Agricultural Ecosystems: Current Challenges and Adaptation (Sawston, Cambridge: Woodhead Publishing, Elsevier Inc), 355–377. doi: 10.1016/B978-0-12-816483-9.00019-0
Stange E. E., Ayres M. P. (2010). “Climate Change Impacts: Insects,” in Encyclopedia of Life Sciences (Chichester: John Wiley & Sons, Ltd). doi: 10.1002/9780470015902.a0022555
Stathers T., Lamboll R., Mvumi B. M. (2013a). Postharvest Agriculture in Changing Climates: Its Importance to African Smallholder Farmers. Food Secur. 5, 361–392. doi: 10.1007/s12571-013-0262-z
Stathers T. E., Lamboll R., Mvumi B. M. (2013b). Post-Harvest Agriculture in a Changing Climate. Int. J. Rural Dev. 47, 1–3.
Steidle J. L. M., Reinhard J. (2003). Low Humidity as a Cue for Habitat Preference in the Parasitoid Lariophagus Distinguendus. BioControl 48, 169–175. doi: 10.1023/A:1022695912500
Sutherst R. W., Maywald G. F., Bourne A. S. (2007). Including Species Interactions in Risk Assessments for Global Change. Global Change Biol. 13, 1843–1859. doi: 10.1111/j.1365-2486.2007.01396.x
Thole V., Vain P., Martin C. (2021). Effect of Elevated Temperature on Tomato Post-Harvest Properties. Plants 10, 2359. doi: 10.3390/plants10112359
Toews M. D., Arthur F. H., Campbell J. F. (2005). Role of Food and Structural Complexity on Capture of Tribolium Castaneum (Herbst) (Coleoptera: Tenebrionidae) in Simulated Warehouses. Environ. Entomol. 34, 164–169. doi: 10.1603/0046-225X-34.1.164
Trematerra P., Sciarretta A. (2004). Spatial Distribution of Some Beetles Infesting a Feed Mill With Spatio-Temporal Dynamics of Oryzaephilus Surinamensis, Tribolium Castaneum and Tribolium Confusum. J. Stored Prod. Res. 40, 363–377. doi: 10.1016/S0022-474X(03)00027-4
Tsaganou F. K., Vassilakos T. N., Athanassiou C. G. (2021). Influence of Temperature and Relative Humidity on the Efficacy of Thiamethoxam for the Control of Three Stored Product Beetle Species. J. Stored Prod. Res. 92, 101784. doi: 10.1016/j.jspr.2021.101784
Tylianakis J. M., Binzer A. (2014). Effects of Global Environmental Changes on Parasitoid-Host Food Webs and Biological Control. Biol. Control 75, 77–86. doi: 10.1016/j.biocontrol.2013.10.003
Tytar V. M., Oksentyuk Y. R. (2019). Modelling the Bioclimatic Niche of a Cohort of Selected Mite Species (Acari, Acariformes) Associated With the Infestation of Stored Products. Vestn. Zool. 53, 399–416. doi: 10.2478/vzoo-2019-0036
USDA-NASS (2022a). Grain Stocks (January 2021) (Washington, D.C: United States Department of Agriculture, National Agricultural Statistics Service).
USDA-NASS (2022b) Quickstats V.2.0 (Washington, D.C: United States Department of Agriculture, National Agricultural Statistics Service). Available at: https://www.nass.usda.gov/Quick_Stats/.
Van Winkle T., Ponce M. A., Quellhorst H. E., Bruce A., Albin C. E., Kim T. N., et al. (2022). Microbial Volatile Organic Compounds From Tempered and Incubated Grain Mediate Attraction by a Primary But Not Secondary Stored Product Insect Pest in Wheat. J. Chem. Ecol. 48, 27–40. doi: 10.1007/s10886-021-01312-8
Vassilakos T. N., Athanassiou C. G. (2013). Effect of Temperature and Relative Humidity on the Efficacy of Spinetoram for the Control of Three Stored Product Beetle Species. J. Stored Prod. Res. 55, 73–77. doi: 10.1016/j.jspr.2013.06.004
Vassilakos T. N., Riudavets J., Castne C., Iturralde-Garcia R. D., Athanassiou C. G. (2019). Efficacy of Modified Atmospheres on Trogoderma Granarium (Coleoptera: Dermestidae) and Sitophilus Zeamais (Coleoptera: Curculionidae). J. Econ. Entomol. 112, 2450–2457. doi: 10.1093/jee/toz139
Warnatsch E. A., Reay D. S., Camardo Leggieri M., Battilani P. (2020). Climate Change Impact on Aflatoxin Contamination Risk in Malawi's Maize Crops. Front. Sust. Food Syst. 4, 591792. doi: 10.3389/fsufs.2020.591792
Weightman R. M., Millar S., Alava J., John Foulkes M., Fish L., Snape J. W. (2008). Effects of Drought and the Presence of the 1BL/1RS Translocation on Grain Vitreosity, Hardness and Protein Content in Winter Wheat. J. Cereal Sci. 47, 457–468. doi: 10.1016/j.jcs.2007.05.011
Wijayaratne L. K. W., Rajapakse R. H. S. (2018). Effects of Spinosad on the Heat Tolerance and Cold Tolerance of Sitophilus Oryzae L. (Coleoptera: Curculionidae) and Rhyzopertha Dominica F. (Coleoptera: Bostrichidae). J. Stor. Prod. Res. 77, 84–88. doi: 10.1016/j.jspr.2018.03.001
Wilkins R. V., Campbell J. F., Zhu K. Y., Starkus L. A., McKay T. (2021). Long-Lasting Insecticide-Incorporated Netting and Interception Traps at Pilot-Scale Warehouses and Commercial Facilities Prevents Infestation by Stored Product Beetles. Front. Sust. Food Syst. 4, 561820. doi: 10.3389/fsufs.2020.561820
Keywords: stored products, integrated pest management, microclimates, review, biosecurity, food security
Citation: Gerken AR and Morrison WR III (2022) Pest Management in the Postharvest Agricultural Supply Chain Under Climate Change. Front. Agron. 4:918845. doi: 10.3389/fagro.2022.918845
Received: 12 April 2022; Accepted: 31 May 2022;
Published: 01 July 2022.
Edited by:
Murray B. Isman, University of British Columbia, CanadaReviewed by:
Tanya Stathers, University of Greenwich, United KingdomBrighton Mvumi, University of Zimbabwe, Zimbabwe
Copyright © 2022 Gerken and Morrison. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alison R. Gerken, YWxpc29uLmdlcmtlbkB1c2RhLmdvdg==
†These authors have contributed equally to this work