 James M. Villegas
James M. Villegas Blake E. Wilson
Blake E. Wilson Michael J. Stout
Michael J. Stout
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Agron. , 13 October 2021
Sec. Pest Management
Volume 3 - 2021 | https://doi.org/10.3389/fagro.2021.754673
This article is part of the Research Topic Advances in Crop Resistance for Insect Pest Control View all 6 articles
The rice water weevil, Lissorhoptrus oryzophilus Kuschel, is the most important early-season pest of rice in the U.S. Additionally, lepidopteran stem-boring pests particularly the invasive Mexican rice borer, Eoreuma loftini (Dyar), are increasingly becoming problematic in Louisiana rice. Although insecticidal seed treatments have been widely adopted and have proven highly effective against weevils and stemborers, alternative management tactics are needed. This study was conducted to evaluate the effects of flood timing and rice cultivar on rice water weevil and stemborer infestations. Field experiments were conducted in Crowley, Louisiana, from 2019–2020. In each year, early- and late-planted trials were established according to a split-split-plot randomized block design. Four commonly grown rice cultivars were selected and were either treated or not treated with a chlorantraniliprole seed treatment. Permanent flood was established at normal timing (approximately the five-leaf stage) or delayed by 2 weeks. Seed treatment reduced weevil densities and stemborer injury by 70–84% and 87–94%, respectively, across planting dates, years, cultivars, and flood timings. The rice cultivar ‘Jupiter' consistently supported the highest numbers of immature weevils, whereas low levels of stemborer injury were observed in ‘Jazzman-2'. Weevil densities were 72 and 20% lower in plots subjected to delayed flood compared to normal flood timing in the 2019 and 2020 late-planted trials, respectively. Reductions of 79 and 93% in stemborer injury in the 2019 early- and late-planted trials, respectively, were observed in delayed flooded plots. Weevil and stemborer infestations negatively affected rice yields, with losses among cultivars ranging from 14 to 49%. Yield losses were generally lower in plots subjected to delayed flood compared to normal flood timing but the difference was only significant in the 2020 late-planted trial. Our data suggest that the combination of cultivar resistance and delayed flooding can serve as a valuable component of an integrated pest management program for both rice water weevil and stemborers.
Rice, Oryza sativa L., is one of the most important crops globally and is consumed by more than half of the world's population (Mohanty, 2013). Rice in the southern U.S. is predominantly cultured by drill-seeding in dry soil followed by the establishment of permanent flood 4–6 weeks after planting. Of the ~180,000 ha of rice planted in Louisiana in the past 2 years, 70% was cultured in this manner (Harrell, 2020). In the U.S., the rice water weevil, Lissorhoptrus oryzophilus Kuschel (Coleoptera: Curculionidae), is the most destructive early-season pest of rice (Way, 1990; Aghaee and Godfrey, 2014). Adult weevils overwinter in leaf litter, bunch grasses, and stubble in and around rice fields and typically emerge from overwintering in early spring (Shang et al., 2004). When rice plants are available, adult weevils feed on the leaves, leaving narrow longitudinal scars parallel to the venation of the leaves. Injury from adult feeding is generally not considered economically important except under unusually heavy infestations (Stout et al., 2013). The establishment of permanent flood in rice fields triggers female weevils to lay eggs in rice leaf sheaths beneath the water surface (Stout et al., 2002b). The majority of economic loss is caused by the soil-dwelling, root-feeding weevil larvae which cause extensive injury to rice root systems, resulting in reductions in tiller numbers, grain weights, and grains per panicle (Shang et al., 2004; Zou et al., 2004a). Yield losses can exceed 25% if rice water weevil infestations are left unmanaged (Stout et al., 2000).
A complex of stem-boring lepidopteran pests (Lepidoptera: Crambidae) attacks U.S. rice from tillering to maturity with the predominant species in Louisiana being the Mexican rice borer, Eoreuma loftini (Dyar) (Way, 2003; Beuzelin et al., 2016). The Mexican rice borer typically lays eggs in folds of dry leaves or leaf sheaths in rice. Neonate larvae will initially feed on leaf sheaths and eventually burrow into the stem. When larval feeding occurs during the vegetative stages of rice growth and development, the affected tillers wither and die, a condition known as a deadheart (Pathak and Khan, 1994). Larval feeding in rice stems during the reproductive stages of rice plants disrupts flow of nutrients to the developing grains, resulting in blanked panicles or “whiteheads” (Chaudhary et al., 1984). Stemborer activity in the field is usually measured by evaluating whitehead density, which is negatively associated with rice yield (Way et al., 2006; Reay-Jones et al., 2007; Wilson et al., 2021a). Stemborers, particularly the invasive Mexican rice borer, are becoming increasingly problematic in southwestern Louisiana. The Mexican rice borer has become firmly established in the state's rice production area as indicated by continued expansion over the past decade and high population density in many regions (Wilson et al., 2015, 2017). Economic losses in Louisiana rice are predicted to reach up to US $40 million annually if infestations of the Mexican rice borer are not managed (Reay-Jones et al., 2008).
Insecticidal seed treatments are currently the most widely used control tactic against insect pests of rice in the southern U.S. (Wilson et al., 2019; Bateman et al., 2020). Seed treatments are applied primarily to control the rice water weevil (Stout et al., 2011a; Hummel et al., 2014; Villegas et al., 2019; Wilson et al., 2021a,b), but seed treatments containing chlorantraniliprole also reduce stemborer injury in rice (Sidhu et al., 2014; Villegas et al., 2019). Despite the effectiveness of insecticidal seed treatments against weevils and stemborers, alternative management strategies need to be investigated to reduce reliance on chemical control and mitigate resistance development. Recent studies reported that some rice cultivars grown in the southern U.S. were able to tolerate infestations by rice water weevil (Villegas et al., 2021a,b). Tolerant crop cultivars are generally able to withstand pest injury and produce adequate yields such that tolerant genotypes suffer lower yield losses than susceptible genotypes when subjected to similar levels of infestation or injury (Flinn et al., 2001; Smith, 2005). Previous studies also indicated that water management practices such as delaying application of permanent flood can reduce infestations and impacts of rice water weevil (Rice et al., 1999; Stout et al., 2001). When flooding is delayed until rice plants are older (after rice plants have begun tillering), the period of time in which rice plants are vulnerable to weevils is shortened, resulting in reductions in weevil infestations (Stout et al., 2013). The effect of delayed flood timing on stemborer injury in rice is unknown. Manipulation of planting date has also been investigated as a tactic for reducing weevil infestations. Rice planted earlier within the recommended range of planting dates is often subject to lower levels of weevil infestations (Stout et al., 2011b). Similarly, less stemborer injury was observed in early-planted rice compared to late-planted rice (Villegas, unpublished data). This study was conducted to evaluate the influence of rice cultivars and flood timing on the infestations of rice water weevil and stem borers and the corresponding impacts on rice yields.
Four field experiments were conducted from 2019 to 2020 at the LSU AgCenter H. Rouse Caffey Rice Research Station in Crowley, Louisiana. In each year, two separate trials were established – one planted early and the other late relative to recommended planting dates (Table 1). For each trial, field plots were laid out according to a split-split-plot randomized block design with four blocks. Each block contained a flood timing (whole-plot) × cultivar (sub-plot) × insecticidal seed treatment (sub-sub plot) field plot arrangement. Rice cultivars used in this study are commonly grown in Louisiana which include two long-grain cultivars (‘Cocodrie' and ‘Mermentau'), a specialty long-grain cultivar (‘Jazzman-2'), and a medium-grain cultivar (‘Jupiter'). The rice cultivar ‘Jupiter' was reported susceptible to the rice water weevil relative to the cultivars used in the study (Saad et al., 2018). Whereas, the rice cultivar ‘Jazzman-2' was reported moderately resistant to stemborer compared to the cultivars used in this study (Villegas et al., 2021a). Seeds were drill-planted at a seeding rate of 67 kg ha−1 in plots measuring 5.5 m in length with seven rows spaced 18 cm apart. Before planting, each cultivar was either treated or not treated with chlorantraniliprole seed treatment (Dermacor® X-100, Corteva Agriscience, Wilmington, Delaware) at a rate of 78.5 g a.i. ha−1 following the methods of Lanka et al. (2014). Dates of key production practices and data collection varied among trials (Table 1). After planting, fields were surface irrigated as needed to facilitate plant emergence and stand establishment. When the rice plants reached the five-leaf to early tillering developmental stage, permanent flood was applied to plots assigned to the normal flood timing treatment, whereas permanent flood was applied 2 weeks later to plots assigned to delayed flood timing. Field plots were maintained following the standard recommendations for weed control and fertilization for drill-planted rice in Louisiana (Saichuk, 2014).
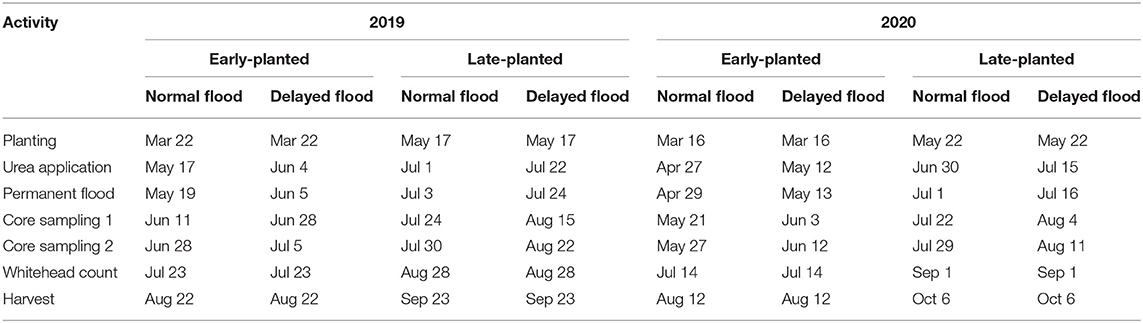
Table 1. Field activities and corresponding dates.
Immature rice water weevil (larvae and pupae) densities were evaluated 3–6 weeks after permanent flood according to established procedures (N'guessan and Quisenberry, 1992; Stout et al., 2001). Root-soil core samples were taken from each plot using a metal corer (9.2 cm diameter × 7.6 cm depth). Core sampling was conducted twice, and three core samples were taken in each plot per sampling time (768 samples total per year). Each core sample contained a minimum of one rice plant with intact roots, and each sample was processed by washing the soil from roots in a 40-mesh screen sieve bucket. The bucket was then placed in a basin with a salt-water solution. Rice water weevil larvae were counted as they floated to the surface of the solution and pupae were counted by inspecting bottoms of sieve buckets. Prior to analysis, the mean number of larvae and pupae across the three core samples from each plot was calculated.
Stemborer injury was evaluated when rice plants reached 100% heading. The number of whiteheads resulting from stemborer infestations was visually assessed and recorded in each plot. Whiteheads were collected and were brought back to the laboratory to retrieve and identify the stemborer species. Prior to analysis, the total number of whiteheads in each plot was converted to whiteheads per m2.
When rice plants reached grain maturity, entire plots were harvested using a small-plot combine (Wintersteiger Delta Plot Combine, Wintersteiger Inc., Salt Lake City, UT). Rough grain weights were adjusted to 12% moisture and converted to kg ha−1. Percentage yield loss was calculated for each cultivar using the formula: % yield loss = [(yieldtreated – yieldnon−treated)/yieldtreated] × 100.
The experimental design utilized in this study allowed the effects of cultivar, flood timing, and seed treatment to be analyzed statistically but did not allow statistical comparisons among planting dates because of insufficient replications. To avoid over-parameterization of the statistical model, data were analyzed separately by year and planting date. All analyses were performed in SAS version 9.4 (SAS Inst., Cary, NC). Rice water weevil and whitehead data were analyzed using generalized linear mixed-models (PROC GLIMMIX) with flood timing, cultivar, seed treatment, and their interactions as fixed effects and core date as a repeated-measure (weevil data only). Random effects were block, block × flood, block × flood × cultivar, and block × flood × cultivar × treatment (weevil only). Yield loss data were analyzed using the same model with cultivar, flood timing, and their interactions as fixed effects and block and block × flood as random effects. The analyses were modeled using a Gaussian distribution. Data residuals were examined (PROC UNIVARIATE) to ensure normality and homogeneity of variances. Weevil and whitehead data were log and square root transformed, respectively, prior to analyses but untransformed means and standard errors are presented. Kenward-Roger adjustment was used to calculate error degrees of freedom. Tukey's HSD post-hoc analysis (α = 0.05) was used for all mean separations.
Flood timing, cultivar, and insecticidal seed treatment influenced rice water weevil densities in all planting dates and years except cultivar and flood timing in the 2019 early-planted trial (Table 2). In the 2019 field experiments, insecticidal seed treatment reduced weevil densities by 83–84% across planting dates, cultivars, and flood timings (Figure 1A). Weevil densities were greater in plots planted with ‘Jupiter' than plots planted with ‘Cocodrie' or ‘Mermentau' in the late-planted trial only (Figure 1B). Weevil densities were reduced by ~72% in plots subjected to delayed flood compared to normal flood timing across insecticide treatments and cultivars in the late-planted trial (Figure 1C). Immature weevils captured in the second core sampling were greater than the first core sampling in the late-planted trial but no effects of core date were observed in the early-planted trial (Table 2; data not shown). In the 2020 field experiments, insecticidal seed treatment reduced weevil densities by 70–84% across planting dates, cultivars, and flood timings (Figure 1D). The rice cultivar ‘Jupiter' had greater weevil densities compared to other cultivars in both the early- and late-planted trials (Figure 1E). Delayed flood reduced weevil densities by 20% across insecticide treatments and cultivars in the late-planted trial but increased weevil densities by 16% in the early-planted trial (Figure 1F). Weevil counts on the second core sampling were higher compared to the first core sampling in the early-planted trial only (Table 2; data not shown).
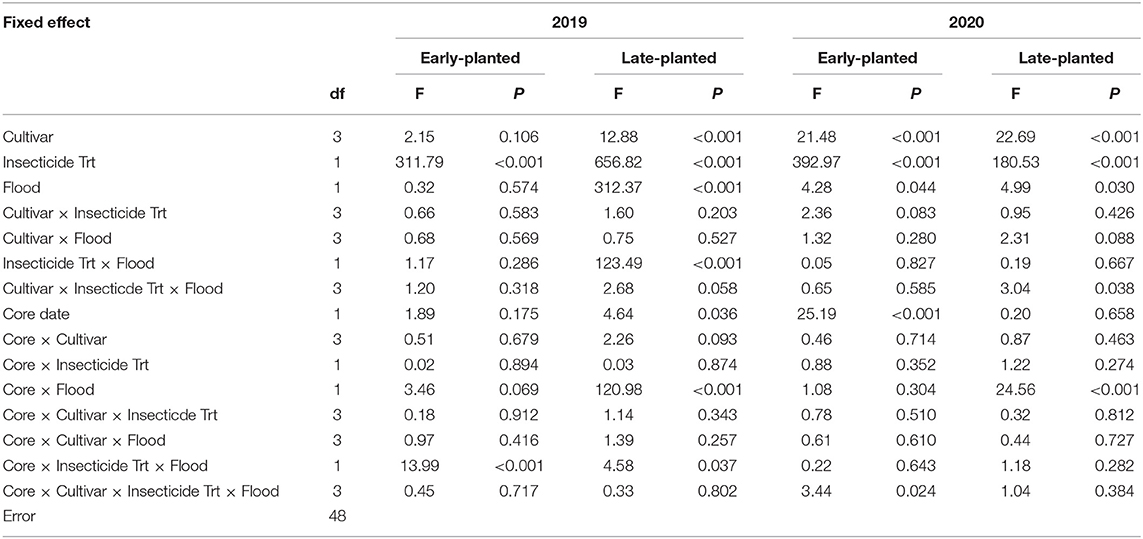
Table 2. Results of repeated measures general linear mixed models (repeated-GLIMMIX) for rice water weevil densities in 2019 and 2020 field trials at Crowley, Louisiana.
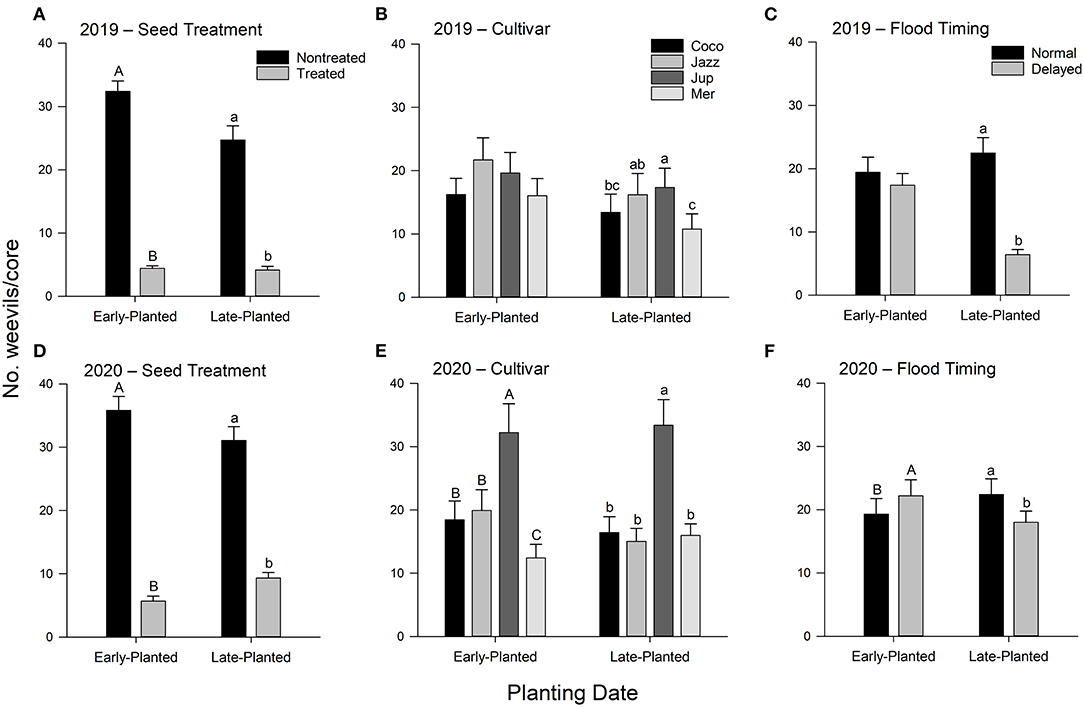
Figure 1. Mean (+SEM) number of immature rice water weevils per root-soil core as affected by (A,D) insecticidal seed treatment, (B,E) rice cultivar, and (C,F) flood timing main effects, Crowley, Louisiana, 2019–2020. Analyses were conducted separately by year and planting date. Bars accompanied by the same letter do not differ significantly (Tukey's HSD: α > 0.05).
No interactions were detected among cultivar, insecticide treatment, and flood timing in the 2019 and 2020 early-planted trials (Table 2; Figures 2A,C). There were significant insecticide treatment × flood interaction and marginally significant cultivar × insecticide treatment × flood interaction observed in the 2019 late-planted trial (Table 2). Weevil densities were greater in non-treated plots subjected to normal flood timing (Figure 2B). In the 2020 late-planted trial, cultivar × insecticide treatment × flood interaction was detected with the greatest weevil density observed on non-treated plots planted with ‘Jupiter' and subjected to normal flood timing (Figure 2D). Additional two- and three-way interactions involving core date were detected in some trials (Table 2; data not shown).
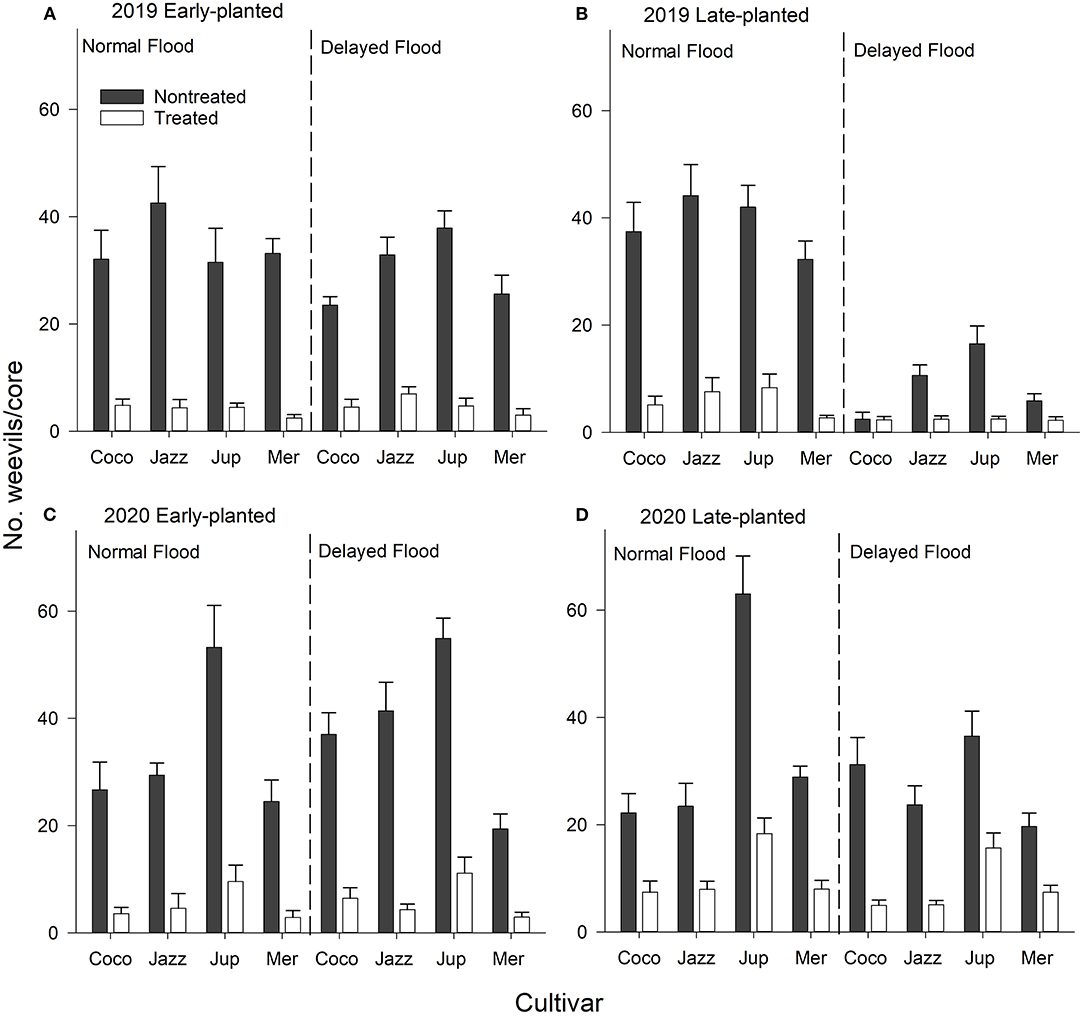
Figure 2. Mean (+SEM) number of immature rice water weevils per root-soil core sample as affected cultivar × seed treatment × flood timing interactions, (A,B) 2019 early- and late-planted trials and (C,D) 2020 early- and late-planted trials, Crowley, Louisiana. Analyses were conducted separately by year and planting date.
All stemborer larvae recovered from rice plants showing whitehead symptoms were determined to be Mexican rice borer. Insecticidal seed treatment influenced whitehead density in all trials (Table 3). Whitehead density was influenced by flood timing in the 2019 trials, but not in the 2020 trials. Cultivar influenced whitehead density in all trials except the 2019 early-planted trial (Table 3). In the 2019 field experiments, insecticidal seed treatment reduced whitehead density by 91–93% across planting dates, flood timings, and cultivars (Figure 3A). Whitehead densities were less in plots planted with ‘Jazzman-2' relative to other cultivars in the late-planted trial, but no differences were observed among cultivars in the early-planted trial (Figure 3B). Reductions of 79 and 93% in whitehead densities in early- and late-planted trials, respectively, were observed in plots subjected to delayed flood compared to normal flood timing across treatments and cultivars (Figure 3C). In the 2020 field experiments, seed treatment reduced whitehead density by 87–94% across planting dates, cultivars, and flood timings (Figure 3D). The rice cultivar ‘Jazzman-2' had reduced whiteheads relative to other cultivars in both early- and late-planted trials (Figure 3E). Flood timing did not affect whitehead density in either the early- or late-planted trials (Figure 3F).
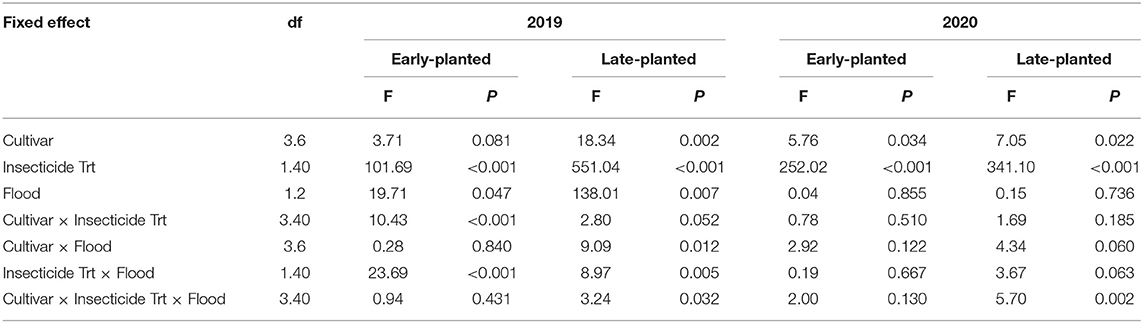
Table 3. Results of general linear mixed models (GLIMMIX) for stemborer injury (whitehead densities) in 2019 and 2020 field trials at Crowley, Louisiana.
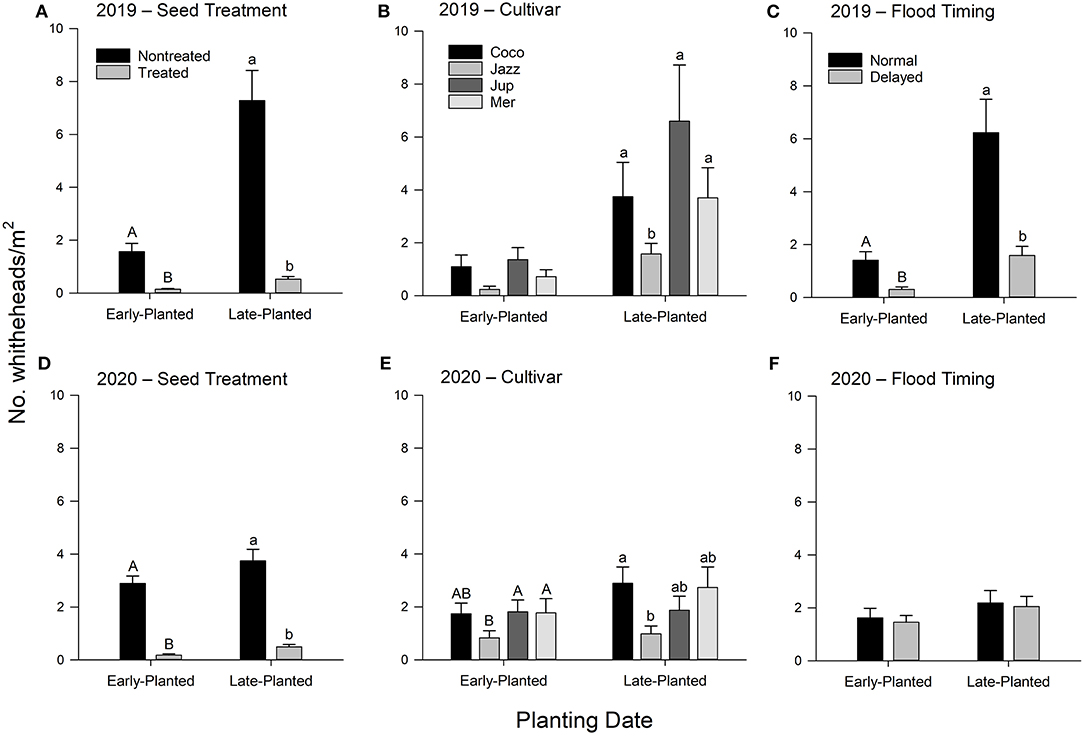
Figure 3. Mean (+SEM) whitehead (blanked panicles) densities as affected by (A,D) insecticidal seed treatment, (B,E) rice cultivar, and (C,F) flood timing main effects, Crowley, Louisiana, 2019–2020. Analyses were conducted separately by year and planting date. Bars accompanied by the same letter do not differ significantly (Tukey's HSD: α > 0.05).
There were significant cultivar × insecticide treatment and flood × insecticide treatment interactions for whiteheads in the 2019 early-planted trial (Table 3). Whitehead densities differed between treated and non-treated plots for all cultivars except ‘Jazzman-2' (Figure 4A). Non-treated plots subjected to normal flood timing had greater whitehead densities than non-treated plots subjected to delayed flood, but the same effect of flood timing was not observed for insecticide-treated plots (Figure 4A). Moreover, there was a significant cultivar × insecticide treatment × flood interaction in the 2019 late-planted trial with the greatest whitehead densities observed on non-treated ‘Cocodrie', ‘Mermentau', and ‘Jupiter' subjected to normal flood timing (Figure 4B). In the 2020 late-planted trial, a significant cultivar × insecticide treatment × flood interaction was detected with greater whitehead densities on non-treated ‘Jupiter' and ‘Mermentau' subjected to normal flood timing and non-treated ‘Cocodrie' subjected to delayed flood timing (Figure 4D). There were no interactions among cultivar, insecticide treatment, and flood timing in the 2020 early-planted trial (Figure 4C).
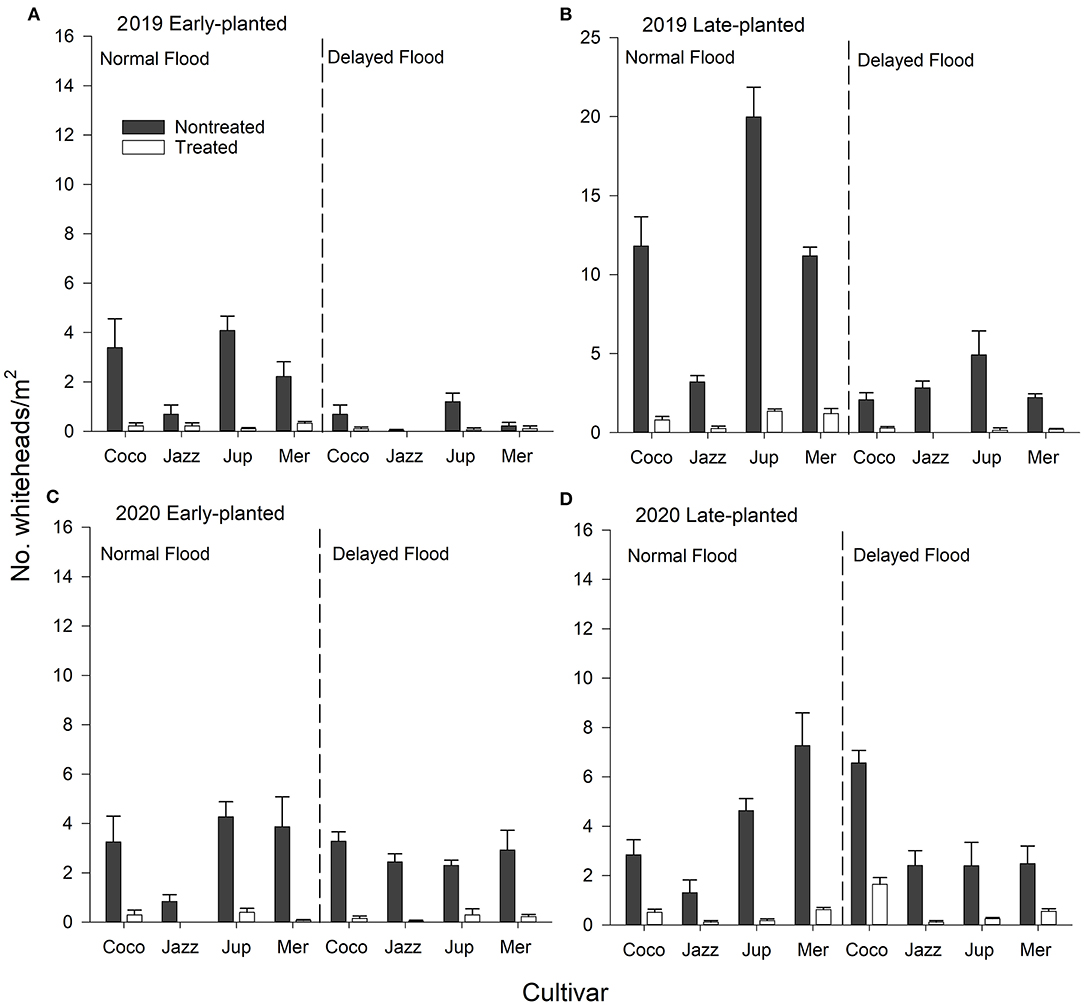
Figure 4. Mean (+SEM) whitehead (blanked panicles) densities as affected by cultivar × seed treatment × flood timing interactions, (A,B) 2019 early- and late-planted trials and (C,D) 2020 early- and late-planted trials, Crowley, Louisiana. Analyses were conducted separately by year and planting date.
Yield losses due to combined infestations of rice water weevil and Mexican rice borer ranged from 14 to 49% and were influenced by cultivar in all planting dates and years except in the 2020 early-planted trial. Flood timing influenced yield loss in the 2020 late-planted trial only. Flood timing × cultivar interactions were not detected in any trial (Table 4). In the 2019 field experiments, yield losses among cultivars across flood timings were 24–49% in the early-planted trial and 14–35% in the late-planted trial. In the early-planted trial, ‘Cocodrie' had greater yield loss compared to other cultivars (Figure 5A), whereas in the late-planted trial, yield loss was greater in ‘Jupiter' compared to ‘Mermentau' (Figure 5C). Although yield losses were reduced by 22 and 42% on plots subjected to delayed flood compared to normal flood timing in the early- and late-planted trials, respectively, the differences were only marginally significant in the late-planted trial (Figures 5B,D). In the 2020 field experiments, yield losses among cultivars were 22–28% in the early-planted trial and 22–34% in the late-planted trial. Yield loss was greater in 'Jupiter' compared to ‘Mermentau' in the late-planted trial (Figure 5G) but no differences were observed among cultivars in the early-planted trial (Figure 5E). Application of delayed flood significantly reduced yield loss by 38% compared to normal flood timing in the late-planted trial (Figure 5H). Despite the 36% reduction in yield loss in the delayed flooded plots relative to normal flooded plots in the early-planted trial, this reduction is not statistically significant (Figure 5F).
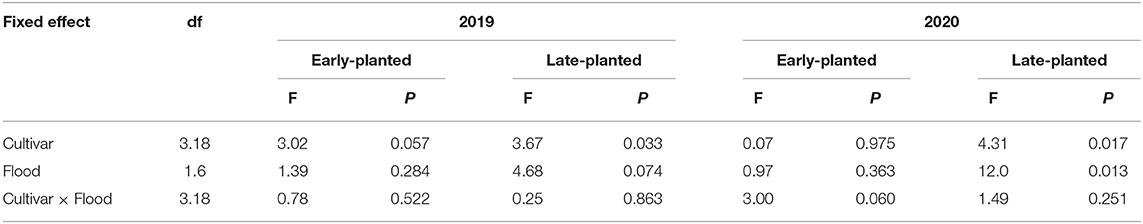
Table 4. Results of general linear mixed models (GLIMMIX) for yield loss in 2019 and 2020 field trials at Crowley, Louisiana.
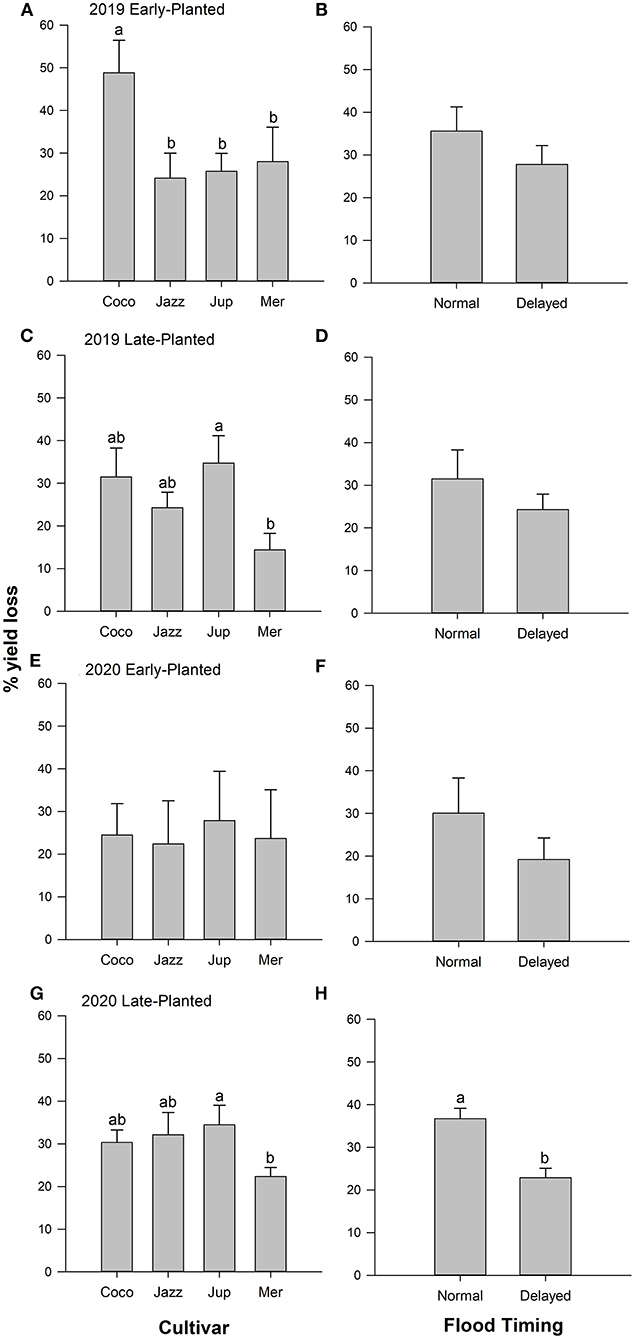
Figure 5. Mean (+SEM) yield loss as affected by (A,C,E,G) rice cultivar and (B,D,F,H) flood timing main effects, Crowley, Louisiana, 2019–2020. Analyses were conducted separately by year and planting date. Bars accompanied by the same letter do not differ significantly (Tukey's HSD α > 0.05).
This study examines alternative tactics to insecticidal seed treatments for managing infestations of rice water weevils and stemborers. The results from the experiments presented here demonstrate that rice cultivars vary in their susceptibility to infestations by rice water weevil and Mexican rice borer, and delaying flood application by only 2 weeks can reduce infestations and impact of these insect pests. Although delayed flood, rice cultivar, and insecticides have all been shown to affect rice water weevil impact individually, this is the first study to examine the interactions among these tactics. Further, this is the first study to examine the effect of flood timing on stemborer infestations and yield loss. No substantial antagonism was found among the tactics evaluated in this study. The significant interactions observed in this study arose from the persistent effect of the chlorantraniliprole seed treatment on weevil larvae and stemborer injury rather than the incompatibility of tactics.
Water management practices have a strong influence on the interaction between rice plants and the rice water weevil (Hesler et al., 1992; Rice et al., 1999; Tindall et al., 2013; Lanka et al., 2015). In particular, the presence of flood triggers the oviposition of adult rice water weevils in rice plants (Stout et al., 2002b). In this study, we found that delaying the application of permanent flood by 2 weeks reduced weevil densities in late-planted trials in both years. These results are consistent with a previous study that reported reductions in rice water weevil densities under a delayed flood system (2–4 weeks later than normal flood timing) (Rice et al., 1999). The field experiments conducted by Rice et al. (1999) were all established late in the planting season. We did not observe any differences in weevil densities between flood timings in the 2019 early-planted trial, whereas, in the 2020 early-planted trial, weevil densities were greater in the delayed flooded plots. Results observed from our early-planted trials were similar to results of studies conducted by Zou et al. (2004c) and Adams et al. (2015) that reported higher weevil densities on delayed flooded plots compared to normal flooded plots, while Stout et al. (2001) observed generally low weevil densities in delayed flooded relative to normal flooded plots but the differences were not statistically significant. The three studies mentioned conducted their field experiments earlier in the planting season relative to trials of Rice et al. (1999) and the late-planted trials reported herein.
The differences in the effects of delayed flood application on rice water weevil densities across planting dates may be explained, in part, by the biology and behavior of the rice water weevil and the susceptibility of rice plant age to weevils. Prior studies reported the emergence of rice water weevil from overwintering begins in late March with peak emergence occurring during April and May (Shang et al., 2004; Zou et al., 2004b). Rice water weevil may go through several generations if rice plants are continuously available in the fields. Although weevils can infest rice plants from the early vegetative to reproductive developmental stages, weevils express a clear preference for plants in the early-tillering stage of development (Stout et al., 2013). Plant phenology was not recorded in this study, but it can be assumed that the differences between flood timings would be greater in late-planted rice when growth is occurring more rapidly. Rice growth and development is affected by the amount of growing degree days (GDD), which is the daily accumulation of heat units during the crop's development, thus later-planted rice which receives more growing degree days in the early stages of development matures rapidly relative to early-planted rice (Cerioli et al., 2021). In this study, the application of delayed flood in late-planted rice allowed plants to receive more growing degree days before weevils start infesting the fields. When core sampling was conducted on delayed-flooded plots in the late-planted trials, most of the rice plants were at the boot to panicle emergence developmental stages. This indicates that the rice plants may have been at the late-tillering stage when permanent flood was established, a plant developmental stage less preferred by weevils (Stout et al., 2013). Alternatively, the reduction on weevil densities in delayed-flooded plots may have been due to the decrease in adult weevil populations in the rice fields. Shang et al. (2004) reported that rice water weevils return to overwintering sites starting late June until October. Weevil populations may be declining by July when the delayed-flooded application occurred in the late-planted trials as adults move to overwintering habitats. In contrast to the late-planted trials, we observed an increase in weevil densities on delayed-flooded plots in the 2020 early-planted trial. The result from this trial may be due to the presence of high adult weevil populations in the field because the permanent flood in the early-planted trials was applied during the peak emergence of the rice water weevil. Regardless of whether it resulted from differences in plant phenology or weevil dynamics, the greatest difference between flood timings was observed in the 2019 late-planted trial, which had our latest flooding date. This suggests the effect is maximized the later into the growing season.
The historically sporadic nature of stemborer infestations in Louisiana has made it challenging to study this pest complex under field conditions. However, due to the increasing prevalence of Mexican rice borer populations, this insect pest can be reliably studied. To our knowledge, this is the first study to document the effects of flood timing on stemborer infestations in general and Mexican rice borer in particular. Our results showed significant reductions in stemborer injury in delayed flooded plots in both early- and late-planted trials in 2019 but no differences in flood timings were observed in the 2020 trials. The effect of water management on stemborer infestation in rice is not well-studied. Nonetheless, two studies have reported reductions in stemborer and leaf folders infestations in rice under an alternate wetting and drying (AWD) water management regime (Chapagain et al., 2011; Hasan et al., 2016). Since flood timing only affected stemborer injury in the first year of this study, further investigation should be conducted.
Results from this study also showed substantial variation in susceptibility among rice cultivars to the Mexican rice borer. Low levels of whitehead densities (stemborer injury) were observed in ‘Jazzman-2' in this study, which is consistent with a recent study that reported this cultivar as moderately resistant to stemborers (Villegas et al., 2021a). Similarly, some variation in susceptibility to rice water weevil was observed among cultivars in this study. The high rice water weevil infestations in rice cultivar ‘Jupiter' reported herein is consistent with previous studies, and the cultivar is now used as a susceptible standard in cultivar resistance evaluations (Stout et al., 2001; Saad et al., 2018; Villegas et al., 2021a; Wilson et al., 2021a). Densities of immature weevils remained somewhat similar across years and planting dates, whereas stemborer injury was observed to be higher in late-planted trials. Wilson et al. (2021a) reported that the increase in yield loss due to stemborers in late- compared to early-planted rice may be attributed to the increase in stemborer infestation later in the planting season. The impact of planting dates, especially on stemborer infestations, needs to be further evaluated in replicated studies with several planting dates and multiple years.
Yield losses due to combined infestations of rice water weevil and Mexican rice borer in this study were generally lower in delayed flooded plots compared to normal flooded plots, but the differences were only significant in the 2020 late-planted trial. Previous studies reported a similar trend in which yield losses due to rice water weevils were reduced in delayed flooded plots (Stout et al., 2001; Zou et al., 2004c). Delaying flood application delays rice water weevil infestations until rice plants are older, which may enhance rice tolerance to this pest. Prior studies have documented that older rice plants can better tolerate injury by rice water weevil larval feeding compared to younger rice plants, and thus are less susceptible to yield losses due to weevil injury (Wu and Wilson, 1997; Stout et al., 2002a). One of the possible reasons is that older plants have highly developed root systems, and are able to quickly recover from weevil injury. Villegas et al. (2021b) proposed that rice plants that are more tolerant to weevil injury have vigorous root systems (particularly in the case of hybrid cultivars). Moreover, N'guessan et al. (1994a,b) reported that rice plants that have increased root regrowth after heavy weevil infestations are also less susceptible to yield losses. Rice cultivars also vary in their level of tolerance to insect pests. In this study, we found that the rice cultivar ‘Mermentau' had the lowest yield loss in the late-planted trials despite having intermediate stemborer injury and weevil infestations relative to other cultivars. This result is consistent with prior studies that documented some rice cultivars expressing varying levels of tolerance to rice water weevil injury (N'guessan et al., 1994a,b; Stout et al., 2001; Villegas et al., 2021a,b). Unfortunately, we were not able to distinguish yield losses from weevils and Mexican rice borer because the seed treatment we used in the study was effective in controlling both pests. Several studies have reported yield losses of 0.3–4.1% per weevil larvae per core (Stout et al., 2002a; Zou et al., 2004a; Adams et al., 2016; Villegas et al., 2021a,b; Wilson et al., 2021a). A recent study also reported a loss of 1.7% in yields per whitehead per m2 due to Mexican rice borer feeding (Wilson et al., 2021a).
The control of rice water weevil and stemborers can be best achieved by using insecticides but our results also indicate that these insect pests can be partly managed through the cultural practice of delaying flooding and the use of tolerant/resistant rice cultivars. Delaying permanent flood application until rice plants are older can be an effective strategy to reduce weevil larvae infesting rice when fields are planted later in the season. Delayed flood application can potentially reduce stemborer injury, but further experiments should be conducted to elucidate the factors that may contribute to this effect. This study, in addition to previous studies (Rice et al., 1999; Stout et al., 2001; Zou et al., 2004c), has shown reductions in yield losses in delayed flooded fields, thus providing a strong rationale for the cultural practice of delayed flooding. Water management may impact weed and disease control, thus effects of delayed flooding on other pest management should be explored. Results from this study also indicate that plant resistance to Mexican rice borer may be a useful strategy as this pest continues to establish its population in Louisiana rice. Furthermore, this study has shown that some rice cultivars may be able to tolerate infestations by both pests. Overall, delaying flood and host plant resistance are suitable alternative management approaches especially in situations where insecticide use is not desirable (i.e., proximity to crawfish production and organic rice production systems).
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
JV, BW, and MS designed the experiments and carried out all aspects of the experiments. JV analyzed the data and wrote the first draft of the manuscript. MS and BW contributed in interpreting the results. All authors edited the manuscript.
This work was supported by the Louisiana Rice Research Board and Southern Sustainable Agriculture Research and Education Graduate Student Grant (SUB00001789).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Kim Landry and numerous student workers both at the rice station and on-campus for their assistance in conducting the experiments. This manuscript was approved for publication by the director of the Louisiana Agricultural Experiment Station, Manuscript No. 2021-234-36582.
Adams, A., Gore, J., Musser, F., Cook, D., Catchot, A., Walker, T., et al. (2015). Impact of water management on the efficacy of insecticide seed treatments against rice water weevil (Coleoptera: Curculionidae) in Mississippi rice. J. Econ. Entomol. 108, 1079–1085. doi: 10.1093/jee/tov095
Adams, A., Gore, J., Musser, F., Cook, D., Catchot, A., Walker, T., et al. (2016). Efficacy of selected insecticide applied to hybrid rice seed. J. Econ. Entomol. 109, 200–206. doi: 10.1093/jee/tov310
Aghaee, M. A., and Godfrey, L. D. (2014). A century of rice water weevil (Coleoptera: Curculionidae): a history of research and management with an emphasis on the United States. J. Integr. Pest Manag. 5, D1–D14. doi: 10.1603/IPM14011
Bateman, N. R., Lorenz, G. M., Thrash, B. C., Gore, J., Way, M. O., Wilson, B. E., et al. (2020). 2017 Rice insect losses in the United States. Midsouth Entomol. 13, 24–32.
Beuzelin, J. M., Wilson, B. E., VanWeelden, M. T., Meszaros, A., Way, M. O., Stout, M. J., et al. (2016). Biology and management of the Mexican rice borer (Lepidoptera: Crambidae) in rice in the United States. J. Integr. Pest Manag. 7, 1–10. doi: 10.1093/jipm/pmw006
Cerioli, T., Gentimis, T., Linscombe, S. D., and Famoso, A. N. (2021). Effect of rice planting date and optimal planting window for Southwest Louisiana. Agron. J. 113, 1248–1257. doi: 10.1002/agj2.20593
Chapagain, T., Riseman, A., and Yamaji, E. (2011). Achieving more with less water: alternate wet and dry irrigation (AWDI) as an alternative to the conventional water management practices in rice farming. J. Agric. Sci. 3, 3–13. doi: 10.5539/jas.v3n3p3
Chaudhary, R. C., Khush, G. S., and Heinrichs, E. A. (1984). Varietal resistance to rice stem-borers in Asia. Int. J. Trop. Sci. 5, 447–463. doi: 10.1017/S1742758400004872
Flinn, M., Smith, M., Reese, J., and Gill, B. (2001). Categories of resistance to greenbug (Homoptera: Aphididae) biotyoe I in Agilops tauschii germplasm. J. Econ. Entomol. 94, 555–563. doi: 10.1603/0022-0493-94.2.558
Harrell, D. L. (2020). 2019 Louisiana Rice Acreage by Variety Survey. LSU AgCenter, 1. Available online at: https://www.lsuagcenter.com/topics/crops/rice/statistics/rice-varieties (accessed May 1, 2020).
Hasan, K., Habib, A., Abdullah, A., Bhattacharjee, D., and Afrad, M. S. S. (2016). Impact of alternate wetting and drying technique on rice production in drought prone areas of Bangladesh. Indian Res. J. Extention Educ. 16, 39–48.
Hesler, L. S., Grigarick, A. A., Oraze, M. J., and Palrang, A. T. (1992). Effects of temporary drainage on selected life history stages of the rice water weevil (Coleoptera: Curculionidae) in California. J. Econ. Entomol. 85, 950–956. doi: 10.1093/jee/85.3.950
Hummel, N. A., Meszaros, A., Ring, D. R., Beuzelin, J. M., and Stout, M. J. (2014). Evaluation of seed treatment insecticides for management of the rice water weevil, Lissorhoptrus oryzophilus Kuschel (Coleoptera: Curculionidae), in commercial rice fields in Louisiana. Crop Prot. 65, 37–42. doi: 10.1016/j.cropro.2014.06.025
Lanka, S. K., Blouin, D. C., and Stout, M. J. (2015). Integrating flood depth and plant resistance with chlorantraniliprole seed treatments for management of rice water weevil, Lissorhoptrus oryzophilus (Coleoptera: Curculionidae). Insect Sci. 22, 679–687. doi: 10.1111/1744-7917.12167
Lanka, S. K., Stout, M. J., Beuzelin, J. M., and Ottea, J. A. (2014). Activity of chlorantraniliprole and thiamethoxam seed treatments on life stages of the rice water weevil as affected by the distribution of insecticides in rice plants. Pest Manag. Sci. 70, 338–344. doi: 10.1002/ps.3570
N'guessan, F. K., and Quisenberry, S. S. (1992). Screening selected lines for resistance to the rice water weevil (Coleoptera: Curculionidae). Environ. Entomol. 23, 665–675. doi: 10.1093/ee/23.3.665
N'guessan, F. K., Quisenberry, S. S., and Croughan, T. P. (1994a). Evaluation of rice anther culture lines for tolerance to the rice water weevil (Coleoptera: Curculionidae). Environ. Entomol. 23, 331–336. doi: 10.1093/ee/23.2.331
N'guessan, F. K., Quisenberry, S. S., Thompson, R. A., and Linscombe, S. D. (1994b). Assessment of Louisiana rice breeding lines for tolerance to the rice water weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 87, 476–481. doi: 10.1093/jee/87.2.476
Pathak, M. D., and Khan, Z. R. (1994). Insect Pest of Rice. Los Banos, Philippines: International Rice Research Institute.
Reay-Jones, F. P. F., Way, M. O., and Reagan, T. E. (2007). Economic assessment of controlling stem borers (Lepidoptera: Crambidae) with insecticides in Texas rice. Crop Prot. 26, 963–970. doi: 10.1016/j.cropro.2006.09.002
Reay-Jones, F. P. F., Wilson, L. T., Reagan, T. E., Legendre, B. L., and Way, M. O. (2008). Predicting economic losses from the continued spread of the Mexican rice borer (Lepidoptera: Crambidae). J. Econ. Entomol. 101, 237–250. doi: 10.1093/jee/101.2.237
Rice, W. C., Croughan, T. P., Ring, D. R., Muegge, M. A., and Stout, M. J. (1999). Delayed flood for management of rice water weevil (Coleoptera: Curculionidae). Environ. Entomol. 28, 1130–1135. doi: 10.1093/ee/28.6.1130
Saad, M. M., Rahaman, M. M., and Stout, M. J. (2018). Varietal resistance against the rice water weevil in field and greenhouse studies. Environ. Entomol. 47, 388–395. doi: 10.1093/ee/nvx199
Shang, H., Stout, M. J., Zhang, Z., and Cheng, J. (2004). Rice water weevil (Coleoptera: Curculionidae) population dynamics in Louisiana. J. Entomol. Sci. 39, 623–642. doi: 10.18474/0749-8004-39.4.623
Sidhu, J. K., Hardke, J. T., and Stout, M. J. (2014). Efficacy of Dermacor-X-100 seed treatment against Diatraea saccharalis (Lepidoptera: Crambidae) on rice. Florida Entomol. 97, 224–232. doi: 10.1653/024.097.0129
Smith, C. M. (2005). Plant Resistance to Arthropod: Molecular and Conventional Approaches. Dordrecht, The Netherlands: Springer Verlag.
Stout, M. J., Hamm, J. C., Abbe, I., and Bergeron, C. (2013). The influence of rice plant age on susceptibility to the rice water weevil, Lissorhoptrus oryzophilus. J. Appl. Entomol. 137, 241–248. doi: 10.1111/j.1439-0418.2012.01746.x
Stout, M. J., Hamm, J. C., Frey, M. J., and Hummel, N. A. (2011a). “Comparison of the efficacies of CruisserMaxx and Dermacor X-100 against rice water weevil at three planting dates,” in LSU AgCenter Rice Research Station 100th Annual Research Report, 265–266.
Stout, M. J., Hummel, N. A., Frey, M. A., and Rice, W. C. (2011b). The impact of planting date on management of rice water weevil in Louisiana rice. Open Entomol. J. 5, 1–9. doi: 10.2174/1874407901105010001
Stout, M. J., Rice, W. C., Linscombe, S. D., and Bollich, P. K. (2001). Identification of rice cultivars resistant to Lissorhoptrus oryzophilus (Coleoptera: Curculionidae), and their use in an integrated management program. J. Econ. Entomol. 94, 963–970. doi: 10.1603/0022-0493-94.4.963
Stout, M. J., Rice, W. C., Riggio, R. M., and Ring, D. R. (2000). The effects of four insecticides on the population dynamics of the rice water weevil, Lissorhoptrus oryzophilus Kuschel. J. Entomol. Sci. 35, 48–61. doi: 10.18474/0749-8004-35.1.48
Stout, M. J., Rice, W. C., and Ring, D. R. (2002a). The influence of plant age on tolerance of rice to injury by the rice water weevil, Lissorhoptrus oryzophilus (Coleoptera Curculionidae). Bull. Entomol. Res. 92, 177–184. doi: 10.1079/BER2001147
Stout, M. J., Riggio, R. M., Zou, L., and Roberts, R. (2002b). Flooding influences ovipositional and feeding behaviour of the rice water weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 95, 715–721. doi: 10.1603/0022-0493-95.4.715
Tindall, K. V., Bernhardt, J. L., Stout, M. J., and Beighley, D. H. (2013). Effect of depth of flooding on rice water weevil, Lissorhoptrus oryzophilus, and yield of rice. J. Insect Sci. 13, 1–9. doi: 10.1673/031.013.6201
Villegas, J. M., Wilson, B. E., and Stout, M. J. (2019). Efficacy of reduced rates of chlorantraniliprole seed treatment on insect pests of irrigated drill-seeded rice. Pest Manag. Sci. 75, 3193–3199. doi: 10.1002/ps.5437
Villegas, J. M., Wilson, B. E., and Stout, M. J. (2021a). Assessment of tolerance and resistance of inbred rice cultivars to combined infestations of rice water weevil and stemborers. Entomol. Exp. Appl. 169:13054. doi: 10.1111/eea.13054
Villegas, J. M., Wilson, B. E., Way, M. O., Gore, J., and Stout, M. J. (2021b). Tolerance to rice water weevil, Lissorhoptrus oryzophilus Kuschel (Coleoptera: Curculionidae), infestations among hybrid and inbred rice cultivars in the Southern U.S. Crop Prot. 139:105368. doi: 10.1016/j.cropro.2020.105368
Way, M. O. (1990). “Insect pest management in rice in the United States,” in Pest Management in Rice, eds B. T. Grayson, M. B. Green, and L. G. Copping (Barking, UK: Elsevier Applied Science Publishers), 181–189.
Way, M. O. (2003). “Rice arthropod pests and their management in the United States,” in Rice: Origin, History, Technology, and Production, eds. C. W. Smith and R. H. Dilday (Hoboken, New Jersey: John Wiley and Sons Inc.), 437–456.
Way, M. O., Reay-Jones, F. P. F., and Reagan, T. E. (2006). Resistance to stem borers (Lepidoptera: Crambidae) among Texas cultivars. J. Econ. Entomol. 99, 1867–1876. doi: 10.1093/jee/99.5.1867
Wilson, B. E., Beuzelin, J. M., and Reagan, T. E. (2017). Population distribution and range expansion of the invasive Mexican rice borer (Lepidoptera: Crambidae) in Louisiana. Environ. Entomol. 46, 175–182. doi: 10.1093/ee/nvx036
Wilson, B. E., Hardy, T. N., Beuzelin, J. M., VanWeelden, M. T., Reagan, T. E., Miller, R., et al. (2015). Expansion of the Mexican rice borer into rice and sugarcane in Louisiana. Environ. Entomol. 44, 1–10. doi: 10.1093/ee/nvv016
Wilson, B. E., Villegas, J. M., and Kraus, E. (2019). Insect pests drive regional differences in rice seed treatment usage. Louisiana Agric. 62:17.
Wilson, B. E., Villegas, J. M., Stout, M. J., and Landry, K. J. (2021a). Relative yield loss from stem borers (Lepidoptera: Crambidae) and rice water weevil (Coleoptera: Curculionidae) in rice. J. Econ. Entomol. 114:toab046. doi: 10.1093/jee/toab046
Wilson, B. E., Villegas, J. M., Way, M. O., Pearson, R. A., and Stout, M. J. (2021b). Cyantraniliprole: a new insecticidal seed treatment for U.S. rice. Crop Prot. 140:105410. doi: 10.1016/j.cropro.2020.105410
Wu, G. W., and Wilson, L. T. (1997). Growth and yield response of rice to rice water weevil injury. Environ. Entomol. 26, 1191–1201. doi: 10.1093/ee/26.6.1191
Zou, L., Stout, M. J., and Dunand, R. T. (2004a). The effects of feeding by the rice water weevil, Lissorhoptrus oryzophilus Kuschel, on the growth and yield components of rice, Oryza sativa. Agric. For. Entomol. 6, 47–53. doi: 10.1111/j.1461-9555.2004.00203.x
Zou, L., Stout, M. J., and Ring, D. R. (2004b). Degree-day models for emergence and development of the rice water weevil (Coleoptera: Curculionidae) in Southwestern Louisiana. Environ. Entomol. 33, 1541–1548. doi: 10.1603/0046-225X-33.6.1541
Keywords: Lissorhoptrus oryzophilus, Eoreuma loftini, delayed flooding, IPM, water management, Mexican rice borer, rice water weevil
Citation: Villegas JM, Wilson BE and Stout MJ (2021) Integration of Host Plant Resistance and Cultural Tactics for Management of Root- and Stem-Feeding Insect Pests in Rice. Front. Agron. 3:754673. doi: 10.3389/fagro.2021.754673
Received: 06 August 2021; Accepted: 16 September 2021;
Published: 13 October 2021.
Edited by:
Cesar Rodriguez-Saona, Rutgers, The State University of New Jersey, United StatesReviewed by:
Jarrad R. Prasifka, United States Department of Agriculture, United StatesCopyright © 2021 Villegas, Wilson and Stout. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Blake E. Wilson, YndpbHNvbkBhZ2NlbnRlci5sc3UuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.