 Doris K. Puozaa1,2
Doris K. Puozaa1,2 Sanjay K. Jaiswal
Sanjay K. Jaiswal Felix D. Dakora
Felix D. Dakora- 1Department of Crop Sciences, Tshwane University of Technology, Pretoria, South Africa
- 2CSIR-Savanna Agricultural Research Institute, Tamale, Ghana
- 3Chemistry Department, Arcadia Campus, Tshwane University of Technology, Pretoria, South Africa
Field studies on the effect of differing seedcoat colour on plant growth and symbiotic performance of three Bambara groundnut landraces using the 15N natural abundance technique and ureide analysis in the xylem stream and petiole extracts revealed a consistent pattern in nodule function between ureide levels and shoot δ15N values, as well as between ureide levels and percent N derived from fixation at Gbalahi, Kpalisogu, and Manga. At those three sites, where shoot δ15N was low and percent N derived from fixation high, ureide concentrations in the xylem sap and petiole extracts were also high, indicating that the two techniques were comparatively quite robust in estimating N2 fixation at a higher precision in the test Bambara groundnut landraces. A comparison of the effect of differing seedcoat pigmentation on the concentration of phenolics released by imbibed seeds showed that the Black landrace contained higher levels of flavonoids and anthocyanins, followed by the Red, and lowest in the Cream landrace. The Black landrace also recorded much higher nodule number and nodule fresh weight per plant, greater shoot biomass, lower shoot δ15N values, greater N derived from fixation, higher symbiotic N contribution, and increased water-use efficiency across all the five study sites, followed by the Red landrace, which produced the highest grain yield. Black seedcoat pigmentation in Bambara groundnut is therefore a marker for increased nodulation, N2 fixation, and plant growth that can be tapped in breeding programs for developing high N2-fixing, water-use efficient grain legumes for use by farmers in Africa.
Introduction
Bambara groundnut (Vigna subterranea L. Verdc.) is the third most important food legume in Africa after groundnut and cowpea, both in consumption and land area under cultivation. The crop is high in protein (20.6%) and carbohydrate (56.5%), and has 6.6% fat as well as 6.3% fibre, thus making it a complete meal (Mohale et al., 2013). It is cultivated by smallholder farmers either as a monoculture, in rotation with cereals, or mixed cultures with cereals, root, and tuber crops. Despite its importance, Bambara groundnut yields have remained low due to lack of improvement. Plant growth, symbiotic N2 fixation and grain yield of Bambara groundnut have been found to vary considerably across Africa (Nyemba and Dakora, 2005; Mabhaudhi et al., 2013; Mohale et al., 2013) due to environmental factors and the symbiotic efficacy of native rhizobia nodulating Bambara groundnut. Grain yield of Bambara groundnut has been reported to double with rhizobial inoculation (Gueye and Bordeleau, 1988), indicating that native soil rhizobia are sometimes inefficient in meeting the N demand of the crop. Applying adequate numbers of competitive and efficient rhizobia on legume seed at planting can be an insurance for enhanced nodulation and N2 fixation for increased legume crop yields.
A few field studies have evaluated N2 fixation in Bambara groundnut using the 15N natural abundance method (Nyemba and Dakora, 2005; Mohale et al., 2013). The 15N isotopic technique has been used to estimate N2 fixation in grain legumes, pasture species, and tree/shrub legumes (Unkovich et al., 2008; Pule-Meulenberg and Dakora, 2009; Belane and Dakora, 2010; Mohale et al., 2013; Muhaba and Dakora, 2020).
As the reproductive unit, seeds carry the genetic material of crop species over time and space, while the seedcoat plays a vital role in protecting the embryo, as well as determining seed dormancy and germination (Moise et al., 2005). The seedcoat also contains a host of novel compounds that are released during germination when the seed imbibes water. These molecules include flavonoids, proteins, peptides, amino acids, alkaloids, and terpenoids (Ndakidemi and Dakora, 2003) and play an important role in plant development, including plant defence against insect pests and pathogens, as well as induction of nodulation genes during symbiotic establishment in N2-fixing legumes (Ndakidemi and Dakora, 2003). The importance of these seedcoat compounds in legume nodulation has been demonstrated; Hungria and Phillips, 1993; Subramanian et al., 2007), and variation in seedcoat colour may be responsible for the differences in N2 fixation observed under field conditions among legume genotypes.
The seedcoat colour of Bambara groundnut landraces can vary distinctly due to the presence of different phenolic compounds (Ndakidemi and Dakora, 2003) and are often used as descriptors for landraces (Heller et al., 1997). Legume accessions are sometimes given local names based on the colour of the testa or seedcoat (Wang et al., 2008). Seedcoat compounds are also known to play a role in the nodulation of symbiotic legumes. In studies by Hungria et al. (1991) and Hungria and Phillips (1993), seedcoat pigmentation was found to strongly influence the nodulation of common bean. An isogenic line of common bean with black seedcoat colour formed more nodules than its cream mutant counterpart, due to differences in seedcoat flavonoid concentration and profile. In a recent study, Bambara groundnut landraces with different seedcoat colours also exhibited differences in grain yield and water-use efficiency under field conditions (Mabhaudhi et al., 2013). So far, however, the link between seedcoat colour and symbiotic performance of legumes has not been established under field conditions. Yet, seedcoat pigmentation could be a useful marker in breeding programs for selecting Bambara groundnut landraces for superior performance. The aim of this study was to assess plant growth, grain yield, N2 fixation, and water-use efficiency in field plants of three Bambara groundnut landraces with different seedcoat colours, using the 15N natural abundance technique.
Materials and Methods
Seed Source and Characteristics
Three Bambara groundnut landraces with different seedcoat colours (Black, Red, and Cream) were used in this study. Seed material of the Black and Red landraces were obtained from farmers at Tiza in the Upper West Region, while the Cream landrace was purchased from Tamale market in the Northern Region, Ghana.
The Bambara groundnut genotypes used in this study differed in seedcoat pigmentation. Otherwise, they exhibited similar growth habit, and phenology, recorded 50% flowering between 52 and 56 days after planting, and matured in 110–120 days.
Description of Field Study Sites
The field trials were set up in July 2013 at three locations within two contrasting agro-ecologies of Ghana. Three field experiments were sited in the Guinea savanna of Northern Region, at Savelugu (latitude 9.624722, longitude 0.827778), Gbalahi (latitude 9.433333, longitude 0.766667) and Kpalisogu (latitude 9.405066, longitude 1.002990), and two at Googo (latitude 10.7545041, longitude 0.4879915) and Manga (latitude 11.017331, longitude 0.264352) in the Sudano-sahelian savanna of the Upper East Region. The two regions have a unimodal rainfall. The annual rainfall in 2013 was 1,047.8 mm in Northern Region and 870 mm in Upper East Region (Table 1). Average daily maximum temperature was 30 to 35°C.
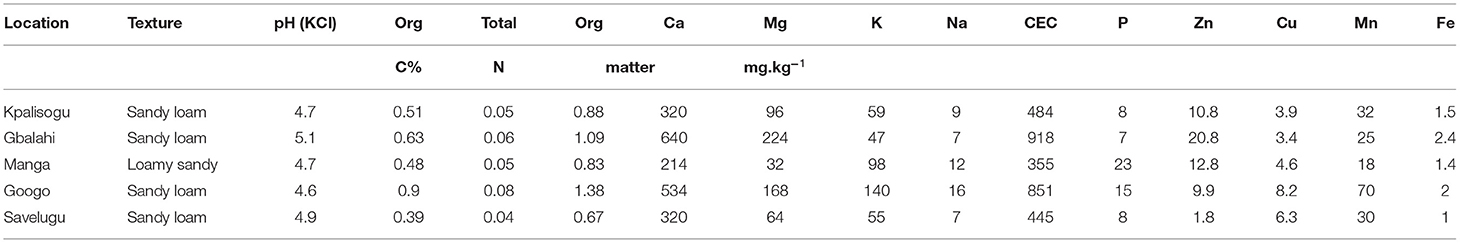
Table 1. Physical and chemical properties of bulk soil collected from each site prior to planting.
Experimental Design and Planting
In June 2013, Bambara groundnut landraces with black, red, and cream seedcoat pigmentation were evaluated for growth and symbiotic performance in five districts of the Northern and Upper East Regions of Ghana. Each field experiment was laid using a randomised complete block design with four replicate plots. Three seeds were planted per hole and latter thinned out to two seedlings per stand. To ensure optimal nodulation at each site, seeds of the three Bambara groundnut landraces were inoculated with Bradyrhizobium strain CB756 and planted at 50 cm between rows and 20 cm within rows. There were 12 plants per row and six rows per plot. A distance of 1.5 m was maintained between plots and 2 m between blocks. In June 2013, Bambara groundnut landraces with black, red and cream seedcoat pigmentation were evaluated for growth and symbiotic performance in five farming districts of the Northern and Upper East Regions of Ghana. Each field experiment was laid using a randomised complete block design with four replicate plots. Three seeds were planted per hole and latter thinned out to two seedlings per stand. To endure optimal nodulation at each site, seeds of the three Bambara groundnut landraces were inoculated with Bradyrhizobium strain CB756 and planted at 50 cm between rows and 20 cm within rows. There were 12 plants per row and six rows per plot. A distance of 1.5 m was maintained between plots in a row and 2 m between blocks.
Seed Inoculation
Seeds were inoculated with a peat-based inoculant of Bradyrhizobium strain CB756 at a rate of 10 g inoculant per kg seed of 108 rhizobial cells.gpeat−1. Inoculated seeds were allowed to dry under shade for a few minutes before planting. The uninoculated control seeds were planted first, followed by the inoculated treatment to avoid contamination. The plots were kept weed-free by hand hoeing, when necessary.
Soil Sampling and Analysis
Following land preparation but prior to planting at Savelugu, Gbalahi, Kpalisogu, Googo, and Manga, 20 soil samples were randomly collected from 0 to 20 cm depth from across each experimental plot using an augur. The soil samples were pooled per plot and thoroughly mixed. The composite soil sample was air-dried in the laboratory, and sieved (2.0 mm) for analysis of texture, organic matter, organic C, pH, CEC, P, Ca, K, Mg, Na, Fe, Zn, Cu, and Mn (ammonium acetate method), as well as total N using the Kjeldahl digestion method (Muhaba and Dakora, 2020).
Collection of Xylem Sap and Ureide Analysis
At flowering at 56 days after planting (DAP), the shoots of 10 to 15 plants per plot were deccapitated at the crown level, and the root bleeding sap collected into Eppindorf tubes using Pasteur pipets. The Eppindorf tubes containing xylem sap were stored in ice, transported to the laboratory, where they were analysed for ureides, as described by Unkovich et al. (2008).
Plant Sampling and Processing
The decapitated shoots of 10 to 15 plants per plot were placed in labelled paper bags. The roots were dug up and root nodules separated, countered, and weighed. The shoots of individual plants were oven-dried (70°C) for 48 h, weighed, and finely ground (0.85 mm) for 15N and 13C isotopic analysis. Shoots of non-legume plant species growing within the experimental plots were concurrently collected, oven-dried (70°C for 48 h) and similarly processed for 15N isotopic analysis in order to estimate soil N uptake by the legume.
15N/14N Isotopic Analysis, Percent N Derived From Fixation, and Amount of N-fixed
About 2.0–2.5 mg of ground sample of legume and reference plant shoots were fed into a mass spectrometer [Carlo Erba NA1500 elemental analyzer (Fisons Instruments SpA, Strada, Rivoltana, Italy) coupled to a Finnigan MAT252 mass spectrometer (Fisons Instrument SpA, Strada, Rivoltana, Italy) via conflo II open-split device] to measure 15N/14N isotopic composition. The δ15N of the shoot samples was computed as (Muhaba and Dakora, 2020):
where the 15N/14Nsample is the abundance ratio of 15N and 14N in the sample and 15N/14Natm is the abundance ratio of 15N and 14N in the atmosphere. The %N and %C of shoot samples were obtained directly from the mass spectrometer, and shoot C/N ratio computed as the ratio of C to N.
The N content of shoot samples was calculated as the product of shoot DM and %N. The %Ndfa of the test legumes was calculated as (Muhaba and Dakora, 2020):
where δ15Nref is the 15N natural abundance of reference plant, δ15Nleg is the 15N natural abundance of legume plant, and the B value is the 15N natural abundance of Bambara groundnut plants completely dependent on atmospheric N2 fixation for their N nutrition. The mean δ15N of all reference plants sampled from each study site was used to estimate the %Ndfa of landraces from that site.
The amount of N-fixed in Bambara groundnut plants was calculated as:
N-fixed = %Ndfa × shoot N content. The soil N uptake by Bambara groundnut was calculated as the difference between total shoot N content and amount of N-fixed.
13C/12C Isotopic Analysis
Percent C and 13C isotopic analysis of soybean plants was done using a mass spectrometer as done for 15N. To determine 13C/12C ratio, 2–3 mg plant sample was weighed into Al capsules, and fed into a Carlo Erba NA1500 elemental analyzer (Fisons Instruments SpA, Strada, Rivoltana, Italy) coupled to a Finnigan MAT252 mass spectrometer via conflo II open-split device. The ratio of 13C/12C in each shoot sample was used to calculate the 13C natural abundance (or δ13C) as (Muhaba and Dakora, 2020):
Where (13C/12C)sample is the isotopic ratio of soybean shoot and (13C/12C)standard is the isotopic ratio of Pee Dee Belemite (PDB) limestone formation, a universally accepted standard from IAEA (Muhaba and Dakora, 2020).
Determination of Flavonoids and Anthocyanins in Seed Extracts
The concentration of flavonoids and anthocyanin in the seeds of Bambara groundnut landraces was measured, as described by Makoi et al. (2010). Seeds of each landrace were weighed (50 g/landrace), counted to record seed number, and finely ground (0.85 mm sieve).Flavonoids and anthocyanins were extracted using 10 g powder in 50 ml of acidified methanol (79:20:1 MeOH H2O HCl). The mixture was incubated on a shaker for 48 h in darkness, filtered, and absorbance of the filtrate measured spectrometrically (UV-VIS) at 300, 530, and 657 nm using acidified methanol as control. The concentration of flavonoid was measured at 300 nm and expressed as Abs g.DM−1 or Abs. seed−1, while the anthocyanin was measured as Abs530 −1/3Abs657 and expressed as Abs g.DM−1 or Abs. seed−1.
Statistical Analysis
The data collected from individual sites were subjected to a one-way analysis of variance (ANOVA), while a 2-factorial arrangement comprising seedcoat colour of genotype and environment was used to analyse the data from across the five experimental sites. Where means were significantly different, they were separated using the Duncan multiple range test. Pearson's correlation analysis was carried out to determine the relationship between plant growth and N nutrition parameters as well as N nutrition and shoot δ13C. The analyses were carried out using STATISTIX version 10 software (StaSoft Inc., Tulsa, OK, USA) package.
Results
Soil Analysis
As shown in Table 1, the soils from the five sites were similar in texture (i.e., sandy loam) and pH, which ranged from pH 4.6 to 5.1. Gbalahi and Googo recorded the highest level of organic matter (1.09 and 1.38%, respectively) and %C (0.63 and 0.90%, respectively). As a result, Gbalahi and Googo had the highest CEC (918 and 851 mg kg−1, respectively), Ca (640 and 534 mg kg−1, respectively), Mg (224 and 168 mg kg−1, respectively), N (0.06 and 0.08 mg kg−1, respectively), Zn (20.8 and 9.9 mg kg−1, respectively) and Fe (2.4 and 2.0 mg kg−1, respectively).
Flavonoid and Anthocyanin Concentrations in Seed Extracts of Bambara Groundnut Landraces
The concentrations of flavonoids and anthocyanin released by imbibed seeds of the Bambara groundnut landraces in acidified methanol were higher in the landrace with black seedcoat (Black), followed by red seedcoat (Red) and least in the cream seedcoat (Cream), whether measured as absorbance per gramme of seed dry matter or on the basis of absorbance per single seed (Table 2). As would be expected, all the three test genotypes had more flavonoids than anthocyanins in the crude seed extract (Table 2). There was also a highly positive significant correlation (r = 0.85, p = 0) between flavonoid and anthocyanin levels in the test Bambara groundnut landraces (data not shown) implying that anthocyanin makes up a great proportion of the total seed flavonoid content.
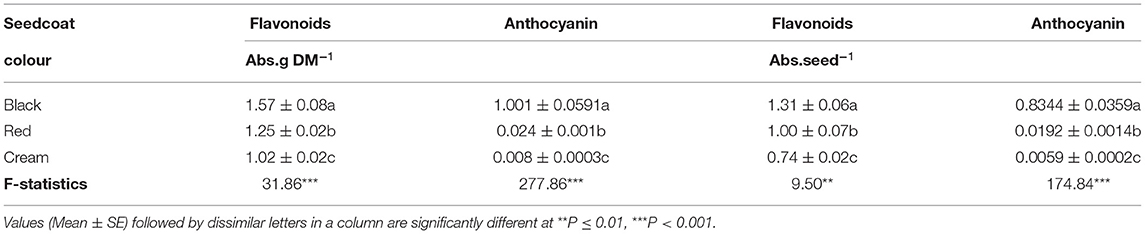
Table 2. Concentration of flavonoids and anthocyanin released by imbibing seeds of Bambara groundnut landraces in acidified MeOH.
δ15N of Reference Plants
At Gbalahi, five reference plant species were analysed, and the δ15N values ranged from +2.76 to +5.19‰, with a combined mean δ15N value of +3.75‰ (Table 3). The δ15N of the three reference plants sampled at Googo ranged from +4.97‰ to +5.39‰, with a mean value of +5.23‰. Four reference plant species were collected from each site at Kpalisogu and Manga, and their mean δ15N values were +3.95 and +4.30‰, respectively (Table 3). At Savelugu, the minimum and maximum δ15N values were +2.47‰ and +2.71‰, with a combined mean δ15N value of +2.61‰ for the six reference plant species (Table 3).
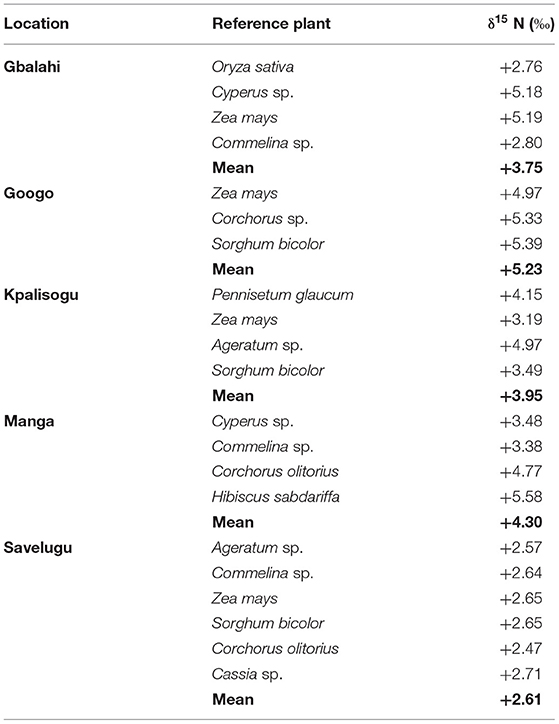
Table 3. Shoot δ15 N values of reference plants used for estimating soil N uptake by Bambara groundnut landraces.
Plant Growth, Symbiotic Performance, Water-Use Efficiency and Grain Yield of Bambara Landraces Planted at Different Experimental Sites
At Gbalahi, the Bambara groundnut landrace Black formed more nodules, with higher nodule fresh weight per plant compared to the Red and Cream landraces (Table 4). Plant growth (measured as shoot dry matter) and δ13C (water-use efficiency) were however similar for all three landraces. The Red landrace produced the highest grain yield relative to Black and Cream (Table 4).
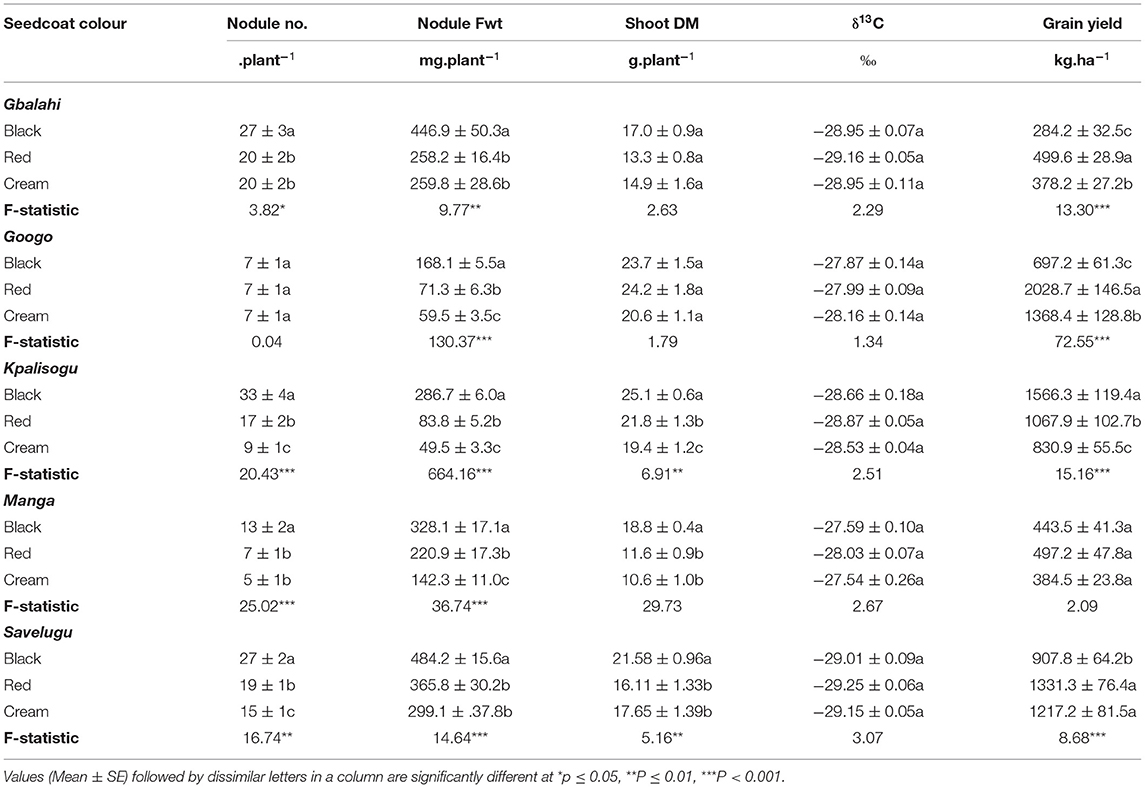
Table 4. Nodulation, shoot dry matter (DM), δ13C values and grain yield of Bambara groundnut landraces with different seedcoat colours sampled from five sites in Ghana.
The Black and Red landraces showed significantly increased shoot N concentration (%N) but not N content. Shoot δ15N was lowest in the Black landrace, and resulted in greater %Ndfa when compared to the Red and Cream landraces (Table 5). The Black landrace also showed the highest ureide concentration in the xylem stream.
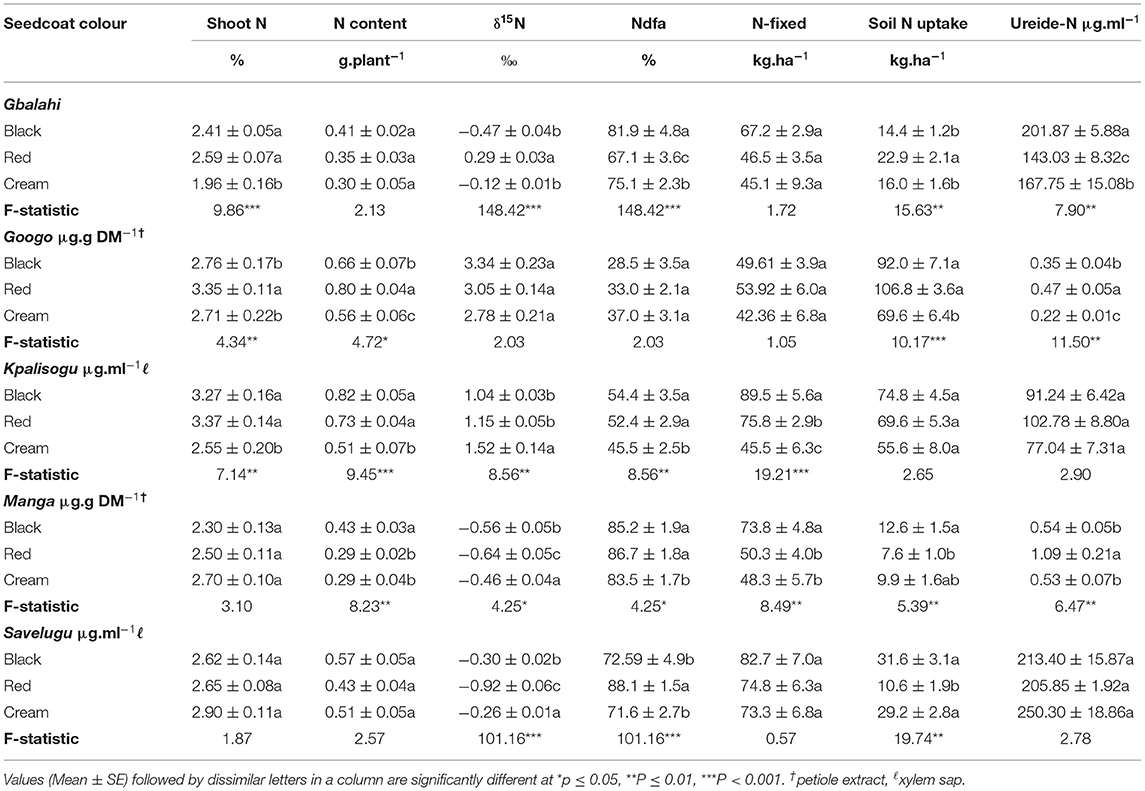
Table 5. Shoot N concentration, N-fixed, ureide concentration and soil N uptake of Bambara groundnut landraces with different seedcoat colours sampled at flowering from five sites in Ghana.
Although seedcoat colour had no effect on nodule number per plant at Googo, nodule fresh weight was greater in the Black landrace, followed by Red and least in the Cream landrace (Table 4). The three Bambara groundnut landraces revealed similar shoot DM and water-use efficiency (δ13C) at Googo. However, they differed in grain yield, with the Red landrace producing higher grain yield than the Cream and Black landraces (Table 4).
The shoot %N was highest in the Red landrace, which when combined with its slightly higher shoot DM resulted in much greater N content relative to the Black and Cream landraces (Table 5). The three landraces were however similar shoot δ15N, %Ndfa and N-fixed. Although soil N uptake was similar for the three landraces, the Red Banbara groundnut showed much higher ureide concentration compared to the others (Table 5).
At Kpalisogu, the Black landrace produced significantly higher nodule number and nodule fresh weight per plant, followed by the Red landrace and Cream (Table 4). Shoot dry matter was also markedly higher in the Black landrace, followed by Red, and lowest in the Cream landrace. Shoot δ13C values were however similar for all three landraces at the Kpalisogu site (Table 4). The Black landrace also recorded much higher shoot biomass and greater grain yield, which almost doubled the yield of the Cream landrace (Table 4).
Shoot N concentration and N content at Kpalisogu were markedly higher in the Black and Red landraces than the Cream (Table 5). The Black landrace also showed much lower shoot δ15N, followed by the Red. This was in contrast to the Cream landrace which recorded a much greater shoot δ15N (Table 5). As a result, the percent N derived from fixation and amounts of N-fixed were higher in the Black and Red compared to the Cream landrace (Table 5). Ureides in the xylem stream and petiole extracts was also much greater in the Black and Red than the Cream landrace, though not statistically significant (Table 5).
At Manga, nodule number and fresh weight per plant were both greater in the Black landrace, followed by Red, and least in the Cream landrace (Table 4). The Black landrace also recorded much higher shoot DM compared to the other two landraces, but shoot δ13C and grain yield were similar (Table 4).
Although shoot N concentration was similar for all three landraces, the N content was significantly higher in the Black landrace due its greater shoot biomass. Shoot δ15N values were much lower in the Black and Red landraces. As a result, the percent N derived from fixation and amount of N-fixed by Black and Red landraces were higher than the Cream. Ureides in the xylem sap of Red landrace was also much higher than the Cream. Soil N uptake by the Red landrace was less than the other two landraces (Table 5).
Nodule number and nodule fresh weight per plant were both greater in the Black landrace at the Savelugu, followed by Red, and much lower in the Cream landrace (Table 4). The Black landrace also produced significantly higher shoot DM but recorded much lower grain yield than the other two landraces.
The shoot N concentration and N content were similar for the three landraces at Savelugu. However, the shoot δ15N was lowest in the Red Bambara groundnut landrace, and resulted in significantly higher %Ndfa than the Black and Cream landraces (Table 5). The amount of N-fixed in the Black landrace was greater in magnitude, but not statistically. Soil N uptake was lower in the Red landrace relative to Black and Cream. Ureide-N in the xylem sap was however similar for all three landraces. As found at the other study sites, shoot δ13C was unaffected by seedcoat colour at Savelugu.
Effects of the Seedcoat Colour × Environment Interactions on Plant Growth, Nodulation, Grain Yield and Water-Use in Bambara Groundnut Landraces
A 2-way analysis of variance involving seedcoat colour and environment revealed marked differences in plant growth, δ13C and grain yield among the Bambara groundnut landraces in the different environments. Nodule number and nodule fresh weight were relatively higher at Gbalahi and Savelugu and lowest at Googo (Table 6). Shoot biomass was much greater at Googo and Kpalisogu, followed by Savelugu, and lowest at Manga and Gbalahi. Grain yield was also markedly higher at Googo, followed by Kpalisogu and lowest at Gbalahi and Manga (Table 6). Shoot δ13C was however greater at Manga in the Upper East Region, where rainfall was lower (867.6 mm), while Savelugu with high rainfall (1047.8 mm) recorded the lowest δ13C values.
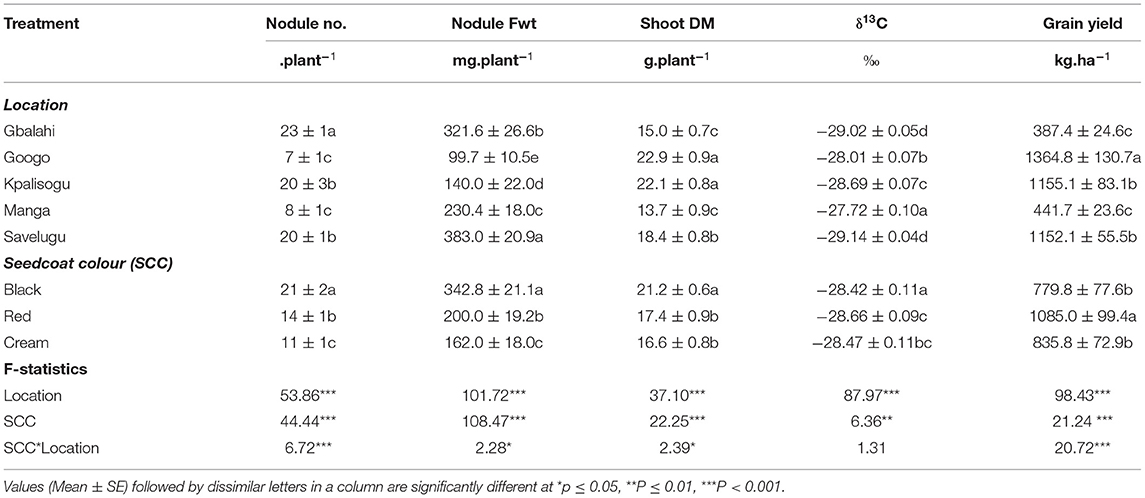
Table 6. Effects of environment and seedcoat colour (SCC) on nodulation, shoot dry matter (DM), δ13C values and grain yield of Bambara groundnut landraces.
The Black landrace produced significantly higher nodule number and nodule fresh weight per plant across all locations, followed by the Red and Cream landraces (which was the lowest). Grain yield was however higher in the Red landrace, but similar for the Black and Cream landraces (Table 6). Shoot δ13C was greater in the Black than the Red and Cream landraces.
Seedcoat colour × location interaction was significant for nodule number, nodule fresh weight, shoot biomass and grain yield, but not shoot δ13C (Table 6). Comparing the performance of each landrace across all five study sites revealed significant location effects. For example, with the Black landrace, nodule number was higher at Kpalisogu, followed by Gbalahi, and lowest at Googo (Figure 1A). With the Red and Cream landraces, nodule number was highest at Gbalahi, followed by Kpalisogu (Figure 1A). All three landraces recorded their highest nodule fresh weights at Savelugu, followed by Gbalahi, and their lowest at Googo and Kpalisogu (Figure 1B).
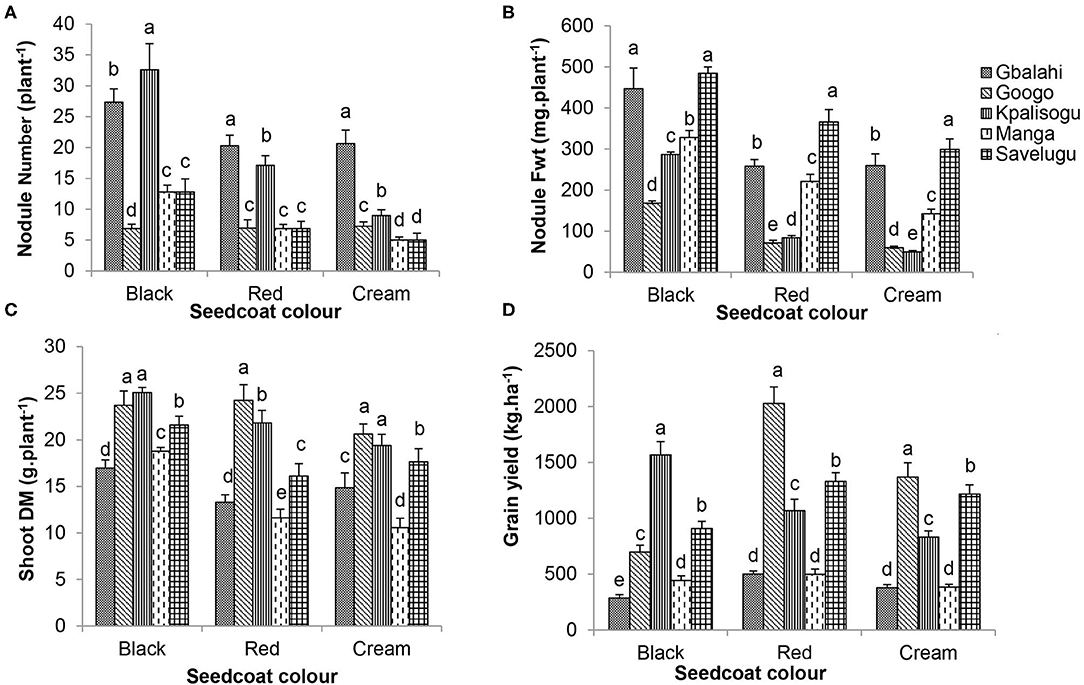
Figure 1. Interactive effects of seedcoat colour × environment on (A) nodule number per plant, (B) nodule Fwt, (C) shoot DM of Bambara groundnut plants sampled at flowering (56 DAP), and (D) grain yield at harvest. Bars with dissimilar letters are significantly different.
For all three landraces, shoot biomass was much higher at Googo and Kpalisogu, and the lowest for the Red and Cream at Manga (Figure 1C). Across all five study sites, grain yield of the Black landrace was highest at Kpalisogu, and lowest at Gbalahi (Figure 1D). With the Red and Cream landraces, grain yield was greater at Googo, followed by Savelugu, and lowest at Manga and Gbalahi (Figure 1D).
Effect of Seedcoat Colour × Environment Interaction on Symbiotic N Nutrition in Bambara Groundnut
The seedcoat colour × location interaction was significant for shoot N concentration and N content, δ15N, percent N derived from fixation, amount of N-fixed and soil N uptake (Table 7). A comparison of symbiotic performance of each landrace across the five study sites revealed significant location effects. For example, shoot %N of the Black and Red landraces was higher at Googo and Kpalisogu than the other three sites, while the Cream landrace showed higher shoot %N at Savelugu, and lowest at Gbalahi (Figure 2A). Shoot N content of the Black and Red landraces was similarly greater Googo and Kpalisogu, and much lower at Gbalahi and Manga, while the Cream landrace showed similar and higher N content at Googo, Kpalisogu, and Savelugu, and lower amounts at Gbalahi and Manga (Figure 2B). Shoot δ15N was highest at Googo, followed by Kpalisogu, and lowest at Manga and Savelugu for all the three landraces (Figure 2C).
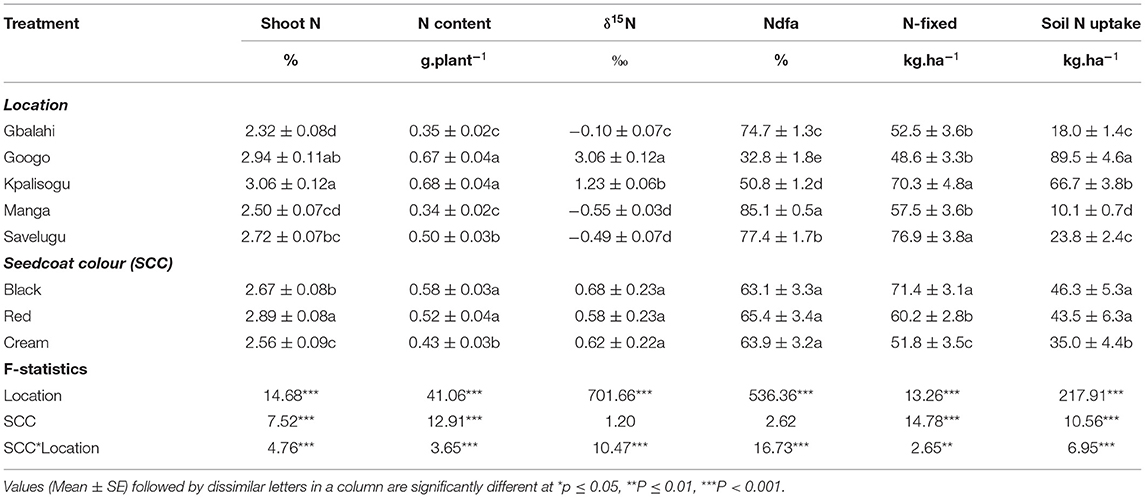
Table 7. Effects of seedcoat colour (SCC) and environment on the N nutrition of Bambara groundnut landrace sampled at flowering (56 DAP).
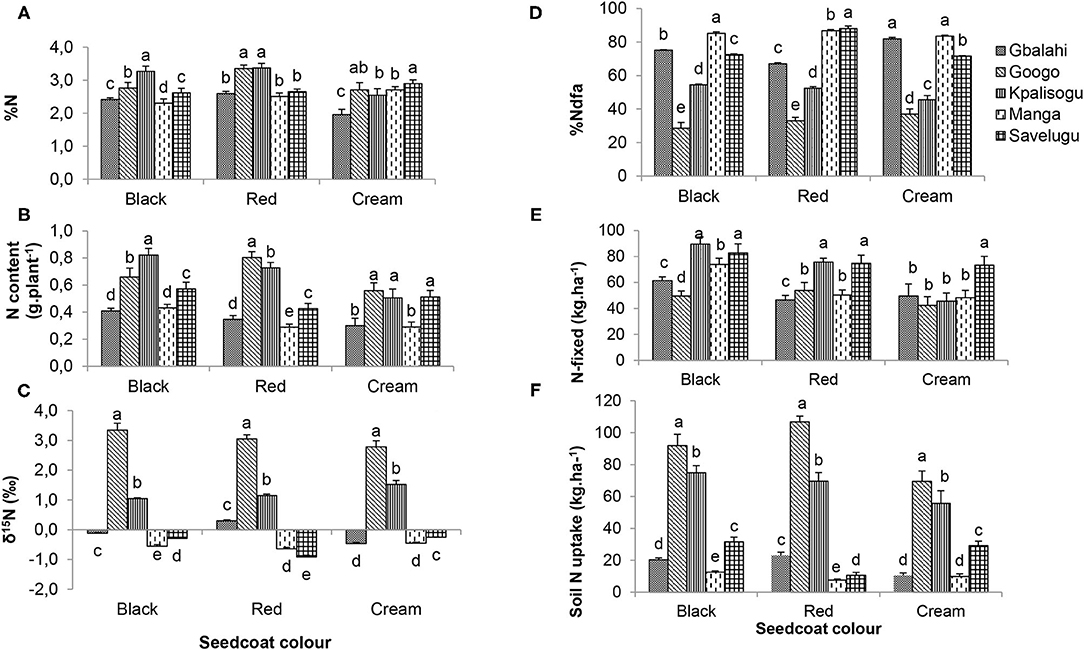
Figure 2. Interactive effects of seedcoat colour × environment on (A) shoot %N, (B) shoot N content, (C) shoot δ15N, (D) %Ndfa, (E) amount of N-fixed, and (F) soil N uptake of Bambara groundnut plants sampled at 56 DAP. Bars with dissimilar letters are significantly different.
The percent N derived from fixation was highest for the Black landrace at Manga, followed by Gbalahi and Savelugu, and lowest at Googo (Figure 2D). In contrast, the Red landrace showed much higher percent N derived from fixation at Savelugu, followed by Manga, and lowest at Googo. In contrast was much, shoot percent N obtained from fixation by the Cream landrace was much greater at Gbalahi and Manga, followed by Savelugu, and lowest at Googo and Kpalisogu (Figure 2D). Across all study sites, the amounts of N-fixed were generally higher for the Black landrace, followed by Red and then the Cream landrace (Figure 2E). However, for site-to- site comparison, the Black landrace recorded greater amounts of N-fixed at Kpalisogu and Savelugu, followed by Manga, and lowest at Googo and Gbalahi, which recorded much higher endogenous soil N concentrations that probably inhibited N2 fixation (Mbah and Dakora, 2018). The Red landrace similarly exhibited higher amounts of N-fixed at Kpalisogu and Savelugu, and the lowest at Gbalahi. In contrast, the amount of N-fixed by the Cream landrace was much greater at Savelugu than the other four sites, which showed similar N contribution (Figure 2E). Soil N uptake was highest at Googo (which had the highest soil N concentration), followed by Kpalisogu, and lowest at Manga for all three landraces (Figure 2F).
Correlation Analysis
There was an inverse relationship between nodule number and δ15N, between nodule fresh weight and δ15N, between nodule number and soil N uptake, as well as nodule fresh weight and soil N uptake (Figures 3A–D, respectively). Similarly, there was a significantly negative correlation between δ15N and %Ndfa, as well as %Ndfa and soil N uptake (Figures 4A,B). However, shoot δ13C (or water-use-efficiency) was positively correlated with shoot N content (p = 0.824), and also with soil N uptake (p = 0.013) (Figures 4C,D).
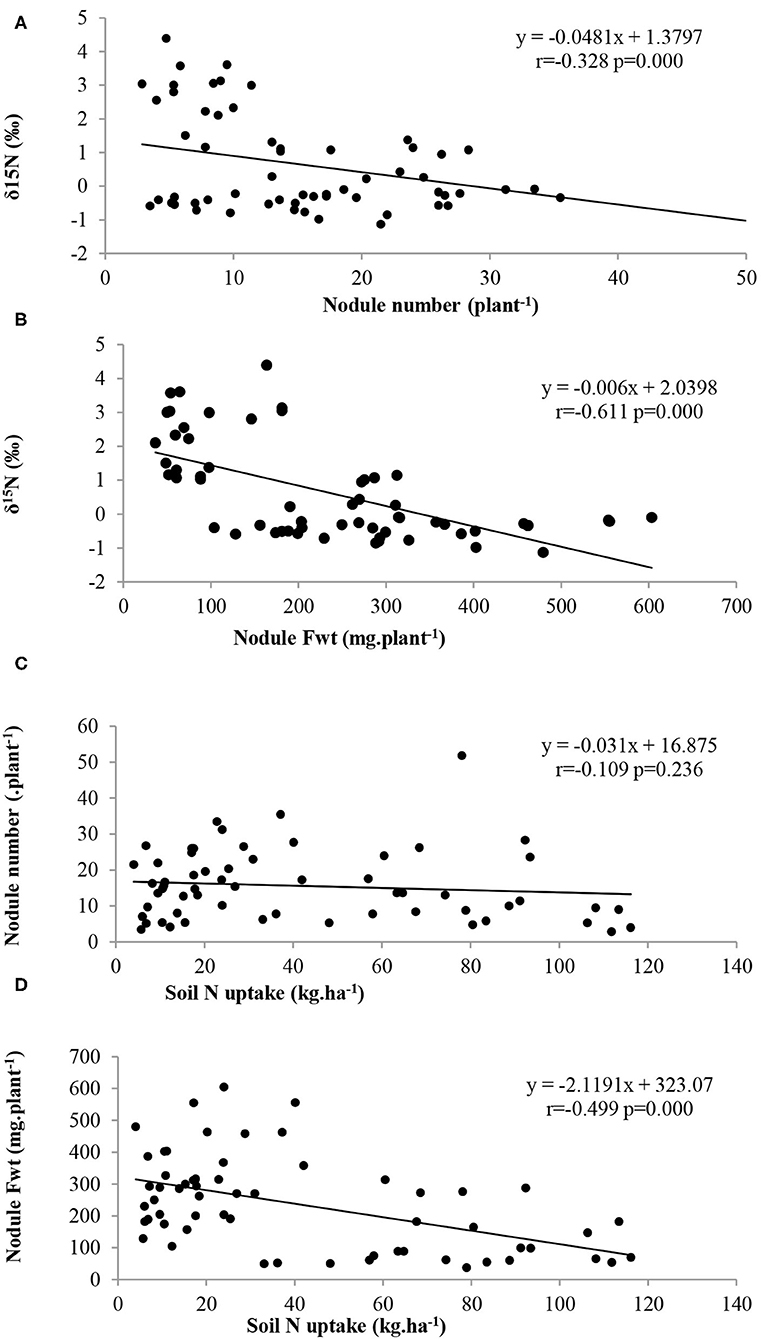
Figure 3. Relationships between (A) δ15N and nodule number, (B) δ15N and nodule Fwt, (C) nodule number and soil N uptake, and (D) nodule fresh weight and soil N uptake of Bambara groundnut landraces sampled at flowering at 65 DAP.
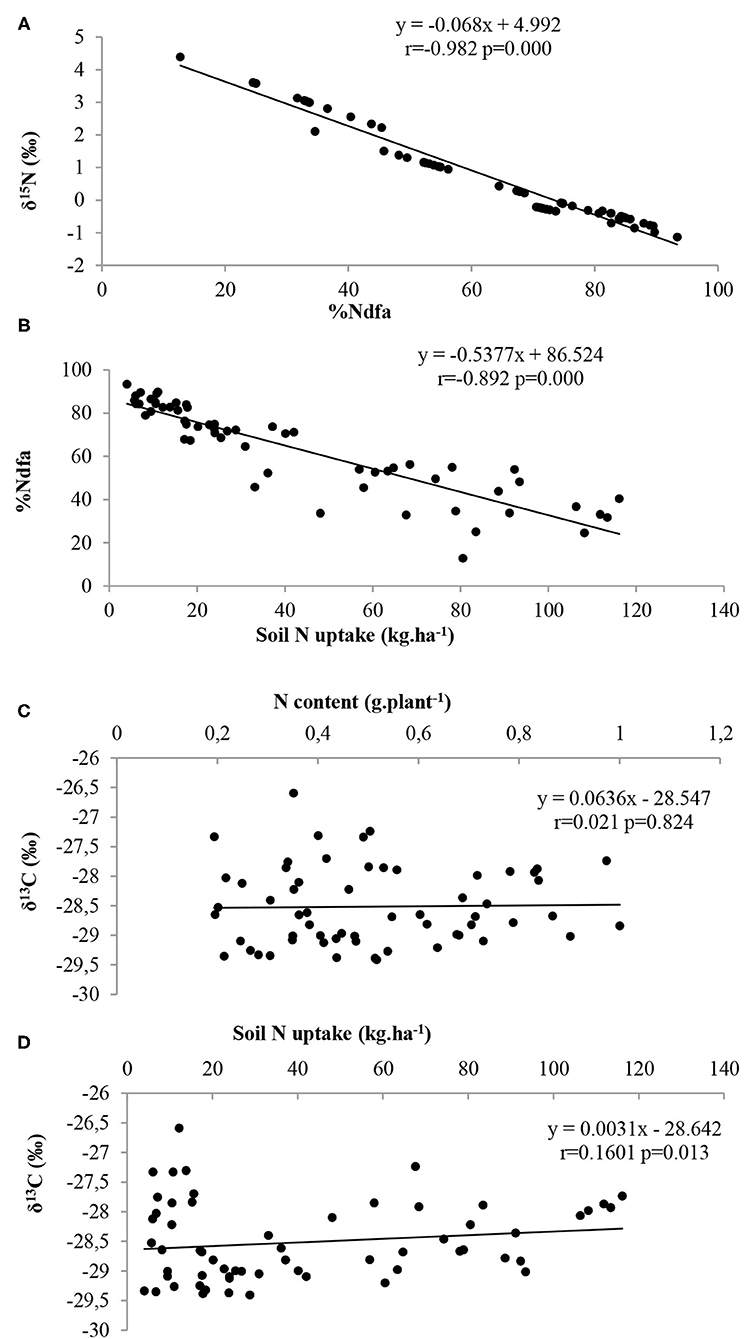
Figure 4. Relationships between (A) %Ndfa from ureide-N and δ15N, (B) %Ndfa and soil N uptake, (C) δ13C and shoot N content, and (D) δ13C and soil N uptake (D) of Bambara groundnut landrace at flowering at 56 DAP.
Discussion
The Black Seedcoat of Grain Legumes Is a Natural Source of Nutraceuticals and Anthocyanins for Useful Exploitation
In this study, the three Bambara groundnut landraces tested differed significantly in their levels of flavonoids and anthocyanins in the grain, a finding consistent with the report by Wang et al. (2008) who also found variations in the concentration of phenolic compounds in the seeds of the same legume species. The phenolic concentrations were much greater in the seeds of Black and Red landraces than the lighter Cream (Table 2); these results were confirmed using UPLC-qTOF-MS analysis (Tsamo et al., 2018). Earlier studies also found higher levels of phenolic compounds in the darker-coloured pea (Troszyńska and Ciska, 2002), azuki bean and soybean with black seedcoat (Lin and Lai, 2006). Ndakidemi and Dakora (2003) similarly reported higher concentrations of anthocyanins and flavonoids in a black-seeded Bambara groundnut accession relative to its cream counterpart. Seedcoat pigmentation is reported to be a major determinant of the profile and concentrations of flavonoids, anthocyanins and anthocyanidins in cowpea, Bambara groundnut and Kersting's groundnut seeds (Tsamo et al., 2018, 2019, 2021). Based on metabolite profiling of cowpea seed coats, as well as Bambara groundnut and Kersting's bean seeds, Tsamo et al. (2020) found black seeds of legumes to be a natural source of nutraceuticals for human consumption, and a reservoir of anthocyanins that can be exploited for developing cosmetic, food and pharmaceutical products.
However, consumer preference for cowpea and Bambara groundnut grain consumption across West Africa (where the most cowpea is produced in Africa) is guided by seed coat pigmentation. The majority of consumers in West Africa prefer cowpea and Bambara groundnut with cream or white seedcoat colour (Ira et al., 2019), which is in contrast to Brazil where “black beans” are most preferred by consumers possibly because they contain the high levels of total phenolics, flavonoids, anthocyanins, anthocyanidins, and antioxidant activity when compared to their counterparts with cream or white seedcoat colour (Hungria et al., 1991; Tsamo et al., 2018, 2019, 2021). The consumption of these antioxidant compounds have implications for human nutrition/health (Jiang et al., 2016; Panche et al., 2016).
Effect of Seedcoat Colour on Plant Growth, Water-Use Efficiency and Grain Yield of Bambara Groundnut Landraces
Relative to the Red and Cream landraces, the Black Bambara groundnut landrace showed greater plant growth (measured as shoot dry matter) at all five study sites, though this was significant at only three locations. The variation in plant growth among the landraces could be linked to varying efficiencies in intercepting solar radiation for conversion to biomass (Toure et al., 2012), inherent genetic differences in photosynthate production, and/or greater resistance to environmental stress factors that limit crop productivity. Improved seedling vigour is also important for early crop establishment, and might be linked to seedcoat colour, as Mabhaudhi et al. (2013) found differences in seedling vigour among Bambara groundnut landraces with varying seedcoat pigmentation. In this study, site-specific comparison showed that the Red landrace consistently produced higher grain yield than the Black and Cream landraces. In Cameroon and Tanzania, red seedcoat pigmentation among Bambara groundnut landraces was associated with increased grain yield (Collinson et al., 2000; Wamba et al., 2012).
Bambara groundnut is generally regarded as a drought-tolerant legume species. Although, in this study, the 13C isotopic analysis was used to measure water-use efficiency in the Bambara groundnut landraces (Mohale et al., 2013; Muhaba and Dakora, 2020), there was no direct relationship between seedcoat colour and water-use efficiency at individual experimental sites (Table 4). However, there were marked differences in shoot δ13C or water-use efficiency among the test Bambara groundnut landraces across the study sites (Table 6). The Black landrace, for example, showed much greater δ13C and therefore higher water-use efficiency than the Red and Cream landraces (Table 6). High tissue flavonoid concentration is known to protect the photosynthetic apparatus in plants under drought conditions (Nogués et al., 1998) and this probably explains the slightly higher shoot δ13C values of the Black landrace. However, enhanced N nutrition also seems to induce greater water-use efficiency (Cabrera-Bosquet et al., 2007). Thus, the significant correlation found between shoot δ13C and shoot N content, as well as shoot δ13C and soil N uptake (Figures 2C,D) in this study suggest greater water-use efficiency in the Black landrace, which was superior in N2 fixation relative to the Red and Cream landraces.
Effect of Seedcoat Pigmentation on Nodulation and Symbiotic N Nutrition of Bambara Groundnut Landraces
In this study, the Black Bambara groundnut landrace consistently showed greater nodule number and nodule fresh weight per plant at each site and across all sites, followed by the Red landrace (Tables 4, 6). The Black and Red landraces also generally recorded lower shoot δ15N values, derived a higher proportion of their N from fixation, and produced more symbiotic N. Anthocyanidins and their anthocyanin aglycones from black-seeded common bean have been reported to be potent inducers of nodulation genes in symbiotic rhizobia (Hungria et al., 1991). We therefore attribute the superior symbiotic performance of the Black Bambara groundnut landrace to its ability to produce and release higher concentrations of nod-gene-inducing anthocyanins and flavonoids than its Red and Cream counterparts (Table 2; Tsamo et al., 2018, 2019, 2020). This argument is supported by the findings of Hungria and Phillips (1993) who also showed that a black-seeded common bean genotype that released higher concentrations of nod-gene-inducing flavonoids elicited greater nodulation than its isogenic cream genotype with reduced biosynthesis of the flavonoid nod-gene inducers. Therefore, the markedly higher concentrations of anthocyanins and flavonoids found in the seedcoats of black cowpea and Bambara groundnut landraces (Tsamo et al., 2019, 2020) probably induce greater nod-gene expression in the microsymbionts of these species, leading to increased nodulation and N2 fixation, as found in this study (Tables 4, 5).
Although it can be argued that, besides seedcoat pigmentation, the observed differences in symbiotic functioning between the test Bambara groundnut landraces were probably due to their genetic makeup, the three landraces used in this study were similar in many traits except seedcoat colour. For example, they exhibited similar growth habit and phenology, recorded 50% flowering between 52 and 56 days after planting, and matured in 110 to 120 days. The seed colour trait, which differentiates them, must therefore be the dominant factor influencing gene expression in relation to N2 fixation.
There were marked differences in nodulation and N2 fixation of the test Bambara groundnut landraces within each location and across study sites. Variations in nodule formation and N2 fixation have been reported for Bambara groundnut in a number of studies (Egbe et al., 2013; Mohale et al., 2013). Possibly, due to the high levels of organic matter, soil %C, Ca, Mg, Zn, and Fe, and the relatively high pH (which all play a role in the nodulation process), the plants sampled from the Gbalahi site in this study had the highest nodule number per plant, followed by Savelugu, and lowest at Manga and Googo. However, nodule mass was much greater at Savelugu when compared to the other sites (Table 6). Furthermore, relative to the other sites, the plants from Manga formed fewer nodules, but derived over 80% of their N nutrition from symbiosis, an indication that though fewer in population, those nodules were highly efficient in N2 fixation due likely to the high concentration of plant-available P in the soil at Manga (23 mg.kg−1), which supported an increase in N2 fixation.
Although the Bambara groundnut symbiosis with Bradyrhizobium strain CB 756 has been reported to be tolerant of mineral N in the rooting medium (Dakora, 1998), the relatively high endogenous soil N at Googo (0.08%) appears to have suppressed nodulation and nodule functioning in Bambara groundnut plants at that site (Saito et al., 2014; Mbah and Dakora, 2018), as evidenced by the low N derived from fixation. Even then, there was greater nodule number and nodule fresh weight per plant of the Black landrace than its Red and Cream counterparts at all the study sites. Despite its greater shoot biomass and superior symbiotic performance, the Black landrace produced the least grain yield, in contrast to other studies which found a significantly strong correlation between N2 fixation and shoot biomass and/or grain yield (Belane et al., 2011; Mokgehle et al., 2014).
Because of the very high percent N derived from fixation at Manga, soil N uptake by all three landraces was lowest at that site, but much higher at Googo and Kpalisogu where N2 fixation levels were relatively lower. Although the soil population of microsymbionts nodulating Bambara groundnut was small at Manga due to the fewer nodules produced, they were highly efficient in N2 fixation, as evidenced by the lower shoot δ15N values and the higher percent N derived from fixation.
In this study, N2 fixation was measured using the 15N natural abundance technique and analysis of ureides (allanton and allantoic acid) in the xylem stream and petiole extracts. As found with the 15N methodology where shoot δ15N, percent N derived from fixation, and amounts of N-fixed differed significantly among the landraces, there were similarly marked variations in the ureide concentration of xylem sap and petiole extracts of the Bambara groundnut landraces (Table 5). A close scrutiny of the results revealed a consistent pattern in nodule function between ureide levels and shoot δ15N, as well as ureide levels and percent N derived from fixation at Gbalahi, Kpalisogu and Manga (Table 5). Where shoot δ15N values were low and percent N from fixation was high, as found at Gbalahi, Kpalisogu, and Manga, the ureide concentrations in xylem sap and petiole extract were also high (Table 5). This clearly shows that the two techniques were quite robust in estimating N2 fixation at a higher precision in the Bambara groundnut landraces (Unkovich et al., 2008). Where the trends among the landraces differed for the two techniques, this could possibly be attributed to the fact that, contrary to the 15N isotopic discrimination method which is cumulative over the life span of the plant, the ureide technique is an instantaneous measure that is easily influenced by environmental factors such as temperature and water, which could have affected transpiration rates at the time of sampling (Unkovich et al., 2008).
In conclusion, the darker seeds of Bambara groundnut landrace was found to contain higher levels of flavonoids, anthocyanin, anthocyanidins, and anti-oxidant activity relative to the Cream landrace. Seedlings from the black-seeded Bambara groundnut landrace also exhibited better plant growth, increased water-use efficiency, higher root nodulation and greater N contribution under field conditions. Black seedcoat pigmentation in Bambara groundnut is therefore a biomarker for increased nodulation and N2 fixation that can be tapped in breeding programs for developing high N2-fixing, water-use efficient grain legumes for use by farmers in Africa.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
DP collected samples and analysed the data. DP and SJ drafted the manuscript. FD conceived the idea, edited, and approved the final version of the paper. All authors contributed to the article and approved the submitted version.
Funding
Bill and Melinda Gates Foundation Project (OPP1028057) on Capacity Building in legume Sciences in Africa.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
This work was supported with grants from the Bill and Melinda Gates Foundation Project on Capacity Building in Legume Sciences in Africa, the South African Department of Science and Technology, the Tshwane University of Technology, the National Research Foundation in Pretoria, and the South African Research Chair in Agrochemurgy and Plant Symbioses. DP is grateful for a competitive doctoral fellowship from the Bill and Melinda Gates Foundation Project on Capacity Building in Legume Sciences in Africa.
References
Belane, A. K., Asiwe, J. A. N., and Dakora, F. D. (2011). Assessment of N2 fixation in 32nodulated field-grown cowpea (Vigna unguiculata L. Walp) genotypes at Taung in South Africa, using 15N natural abundance. Afr. J. Biotech. 10, 11450–11458. doi: 10.5897/AJB11.674
Belane, A. K., and Dakora, F. D. (2010). Symbiotic N2 fixation in 30 field-grown cowpea (Vigna unguiculata L. Walp.) genotypes in the Upper West Region of Ghana, measured using 15N natural abundance. Biol. Fertil. Soil 46, 191–198. doi: 10.1007/s00374-009-0415-6
Cabrera-Bosquet, L., Molero, G., Bort, J., Nogués, S., and Araus, J. L. (2007). The combined effect of constant water deficit and nitrogen supply on WUE, NUE and Δ 13C in durum wheat potted plants. Ann. Appl. Biol. 151, 277–289. doi: 10.1111/j.1744-7348.2007.00195.x
Collinson, S. T., Sibuga, K. P., Tarimo, A. J. P., and Azam-Ali, S. N. (2000). Influence of sowing date on the growth and yield of Bambara groundnut landraces in Tanzania. Exp. Agric. 36, 1–13. doi: 10.1017/S0014479700361038
Dakora, F. D. (1998). Nodule function in symbiotic Bambara groundnut (Vigna subterranea L) and Kerstings bean (Macrotyloma geocarpum L.) is tolerant of nitrate in the root medium. Ann. Bot. 82, 687–690. doi: 10.1006/anbo.1998.0720
Egbe, M. O., Alhassan, G. A., and Ijoyah, M. (2013). Nodulation, nitrogen yield and fixation by Bambara groundnut (Vigna subterranea L. Verdc.) landraces intercropped with cowpea and maize in Southern Guinea savanna of Nigeria. Agric. Sci. 1, 15–28. doi: 10.12735/as.v1i4p15
Gueye, M., and Bordeleau, L. M. (1988). Nitrogen fixation in Bambara groundnut (Voandzea subterranea L.Thouars). Mircen. J. 4, 365–375. doi: 10.1007/BF01096142
Heller, J., Begemann, F., and Mushonga, J. (1997). Bambara groundnut, Vigna subterranea (L.) Verdc. Prom. Conserv. Underutiliz. Neglect. 9:166. ISBN: 978-92-9043-299-9, ISBN: 92-9043-299-3
Hungria, M, Joseph, C. M., and Phillips, D. A. (1991). Anthocyanindins and flavonols, major nod-gene inducers from seeds of a black-seeded common bean (Phaseolus vulgaris L.). Plant Physiol. 97, 751–758. doi: 10.1104/pp.97.2.751
Hungria, M., and Phillips, D. (1993). Effects of a seed colour mutation on rhizobial nod-gene-inducing flavonoids and nodulation in common bean. Mol. Plant-Microbe Interact. 6, 418–422. doi: 10.1094/MPMI-6-418
Ira, A., Zhenyu, J., and Francis, K. (2019). Market preferences for cowpea (Vigna unguiculata L. Walp.) dry grain in Ghana. Afr. J. Agric. Res. 14, 928–934. doi: 10.5897/AJAR2019.13997
Jiang, N., Doseff, A. I., and Grotewold, E. (2016). Flavones: from biosybnthesis to health benefits. Plants 5:27. 5020027doi: 10.3390/plants5020027
Lin, P. Y., and Lai, H. M. (2006). Bioactive compounds in legumes and their germinated products. J. Agric. Food Chem. 54, 3807–3814. doi: 10.1021/jf060002o
Mabhaudhi, M., Modi, A. T., and Beletse, Y. G. (2013). Growth, phenological nd yield responses of a Bambara groundnut (Vigna subterranea L. Verdc.) landrace to imposed water stress: II: Rainfall shelter conditions. Water SA 39:22. Doi.org/10.4314/wsa.v39i2.2. doi: 10.4314/wsa.v39i2.2
Makoi, J. H. J. R., Belane, A. K., Chimphango, S. B. M., and Dakora, F. D. (2010). Seed flavonoids and anthocyanins as markers of enhanced plant defence in nodulated cowpea (Vigna unguiculata L. Walp.) Field Crop. Res. 118, 21–27. doi: 10.1016/j.fcr.2010.03.012
Mbah, G. C., and Dakora, F. D. (2018). Nitrate inhibition of N2 fixation and its effect on micronutrient accumulation in shoots of soybean (Glycine max L. Merr.), Bambara groundnut (Vigna subterranea L. Vedc.) and Kersting's groundnut (Macrotyloma geocarpum Harms.). Symbiosis 75, 205–216. doi: 10.1007/s13199-017-0531-2
Mohale, K. C., Belane, A. K., and Dakora, F. D. (2013). Symbiotic N nutrition, C assimilation, and plant water-use efficiency in Bambara groundnut (Vigna subterranea L. Verdc) grown in farmers' fields in South Africa, measured using 15N and 13C natural abundance. Biol. Fertil. Soil 50, 307–319. doi: 10.1007/s00374-013-0841-3
Moise, A. J., Han, S., Loreta Gudynaite-Savitch, L., Johnson, D. A., and Miki, B. L. A. (2005). Seed coats: structure, development, composition, and biotechnology. In Vitro Cell. Develop. Biol. Plant 41, 620–644. doi: 10.1079/IVP2005686
Mokgehle, S. N., Dakora, F. D., and Mathews, C. (2014). Variation in N2 fixation and N contribution by 25 groundnut (Arachis hypogaea L.) varieties grown in different agro-ecologies, measured using 15N natural abundance. Agric. Ecosys. Environ. 195, 161–172. doi: 10.1016/j.agee.2014.05.014
Muhaba, S. K., and Dakora, F. D. (2020). Symbiotic performance, shoot biomass and water-use efficiency of three groundnut (Arachis hypogea L.) genotypes in response to phosphorus supply under field conditions in Ethiopia. Front. Agric. Sci. Eng. 7, 455–466. doi: 10.15302/J-FASE-2020354
Ndakidemi, P. A., and Dakora, F. D. (2003). Legume seed flavonoids and nitrogenous metabolites as signals and protectants in early seedling development. Funct. Plant Biol. 30, 729–745. doi: 10.1071/FP03042
Nogués, S., Allen, D. J., Morison, I. L., and Baker, N. R. (1998). Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants. Plant. Physiol. 117, 173–81. doi: 10.1104/pp.117.1.173
Nyemba, R. C., and Dakora, F. D. (2005). Identifying suitable arable weeds as reference plants for measuring N2 fixation in grain legumes in Zambia using 15N natural abundance. Symbiosis 40, 79–86.
Panche, A. N., Diwan, A. D., and Chandra, S. R. (2016). Flavonoids: an overview. J. Nutr. Sci. 5, 1–15. doi: 10.1017/jns.2016.41
Pule-Meulenberg, F., and Dakora, F. D. (2009). Assessing the symbiotic dependency of grain and tree legumes on N2 fixation for their N nutrition in five agro-ecological zones of Botswana. Symbiosis 48, 68–77. doi: 10.1007/BF03179986
Saito, A., Tanabata, S., Tanabata, T., Tajima, S., Ueno, M., Ishikawa, S., et al. (2014). Effect of nitrate on nodule and root growth of soybean (Glycine max (L.) Merr.). Internat. J. Mol. Sci. 15, 4464–4480. doi: 10.3390/ijms15034464
Subramanian, S., Stacey, G., and Yu, O. (2007). Distinct crucial roles of flavonoids during legume nodulation. Trend Plant Sci. 12, 282–285. doi: 10.1016/j.tplants.2007.06.006
Toure, Y., Kone, M., Tanoh, H. K., and Kone, D. (2012). Agromorphological and phenological variability of 10 Bambara groundnut [Vigna subterranea L. Verdc. (Fabaceae)] landraces cultivated in the Ivory Coast. Tropicultura 30, 216–221.
Troszyńska, A., and Ciska, E. (2002). Phenolic compounds of seedcoats of white and coloured varieties of pea (Pisum sativum L.) and their total antioxidant activity. Czech J. Food Sci. 20, 15–22. doi: 10.17221/3504-CJFS
Tsamo, A. T., Mohammed, H., Mohammed, M., Ndibewu, P. P., and Dakora, F. D. (2019). Seed coat metabolite profiling of cowpea (Vigna unguiculata L. Walp.) accessions from Ghana using UPLC-PDA-QTOF-MS and chemometrics. Nat. Prod. Res. 28, 808–811. doi: 10.1080/14786419.2018.1548463
Tsamo, A. T., Mohammed, M., and Dakora, F. D. (2020). Metabolite fingerprinting of Kersting's groundnut [Macrotyloma geocarpum (Harms) Maréchal and Baudet] seeds using UPLC-qTOF-MS reveals the nutraceutical and antioxidant potentials of the orphan legume. Front. Nutr. 7:306. doi: 10.3389/fnut.2020.593436
Tsamo, A. T., Mohammed, M., Ndibewu, P. P, Dakora, F. D., Jaiswal, SK, and Dakora, FD (2021). Identification quantification of anthocyanins in seeds of Kersting's groundnut [Macrotyloma geocarpum (Harms) Marechal Baudet] landraces of varying seed coat pigmentation. J. Food Measur. Charact. 13, 2310–2317. doi: 10.1007/s11694-019-00150-3
Tsamo, A. T., Ndibewu, P. P., and Dakora, F. D. (2018). Phytochemical profile of seeds from 21 Bambara groundnut landraces via UPLC-qTOF-MS. Food Res. Internat. 112, 160–168. doi: 10.1016/j.foodres.2018.06.028
Unkovich, M., Herridge, D., Peoples, M., Cadisch, G., Boddey, R., Giller, K., et al. (2008). Measuring plant-associated nitrogen fixation in agricultural systems (ACIAR Mono). Canberra: Australian Centre for International Agricultural Research (ACIAR).
Wamba, O., Taffouo, V., Youmbi, E., Ngwene, B., and Amougou, A. (2012). Effects of organic and inorganic nutrient sources on growth, total chlorophyll and yield of three Bambara groundnut landraces in the Coastal Region of Cameroom. J. Agron., 11, 31–42. doi: 10.3923/ja.2012.31.42
Keywords: 15N natural abundance, shoot δ 15N, ureides, N-fixed, nodulation, phenolics, flavonoids and anthocyanins
Citation: Puozaa DK, Jaiswal SK and Dakora FD (2021) Black Seedcoat Pigmentation Is a Marker for Enhanced Nodulation and N2 Fixation in Bambara Groundnut (Vigna Subterranea L. Verdc.) Landraces. Front. Agron. 3:692238. doi: 10.3389/fagro.2021.692238
Received: 07 April 2021; Accepted: 16 July 2021;
Published: 13 August 2021.
Edited by:
Juan Imperial, Consejo Superior de Investigaciones Científicas (CSIC), SpainReviewed by:
Vinay Kumar, Central University of Punjab, IndiaDulce Nombre Rodriguez-Navarro, IFAPA Centro Las Torres Tomejil, Spain
Copyright © 2021 Puozaa, Jaiswal and Dakora. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Felix D. Dakora, ZGFrb3JhZmRAdHV0LmFjLnph