
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Vet. Sci. , 29 May 2019
Sec. Zoological Medicine
Volume 6 - 2019 | https://doi.org/10.3389/fvets.2019.00148
This article is part of the Research Topic Health and Disease in Free-Ranging and Captive Wildlife View all 41 articles
 Nuno Santos1*
Nuno Santos1* José Jambas2António Monteiro3Jorge Amaral3Nuno Martins4Javier Garcia5Ana Martinez Fernández6
José Jambas2António Monteiro3Jorge Amaral3Nuno Martins4Javier Garcia5Ana Martinez Fernández6 Kevin Morris Tyler7Tereza Almeida1,8
Kevin Morris Tyler7Tereza Almeida1,8 Joana Abrantes1Pedro J. Esteves1,8
Joana Abrantes1Pedro J. Esteves1,8Trichomonas gallinae is a pathogen of conservation relevance, whose main maintenance hosts are Columbiformes, but spillover to avian predators has been described. The goal of this study was to characterize the epidemiology of Trichomonas spp. in a community of free-ranging domestic and wild Columbiformes and an endangered predator, Bonelli's eagle Aquila fasciata. We surveyed 253 live-captured Rock doves, 16 nestling Bonelli's eagles and 41 hunted Columbiformes. Oro-esophageal swabs were incubated in culture media and Trichomonas spp. isolated from Bonelli's eagle (6.3%, CI95 1.1-28.3), Turtle dove Streptopelia turtur (56.3%, CI95 39.3–71.8), Wood pigeon Columba palumbus (83.3%, CI95 43.7–97.0) and Rock dove Columba livia (68.4%, CI95 62.4–73.8). Infected Rock doves showed significantly poorer body condition than uninfected ones (p = 0.022). From a subset of 32 isolates, 18S and ITS1/5.8S/ITS2 rRNA genes were sequenced and Maximum-Likelihood trees inferred. Four ribotypes of Trichomonas spp. were identified. In this study area Trichomonas spp. seem to persist in a multi-host system involving several species of Columbiformes. Conservation actions aimed at increasing the availability of trophic resources for Bonelli's eagles through Rock dove restocking should consider the risk of pathogen transmission and of introduction of alien strains.
Trichomonosis is an avian disease of conservation relevance, caused by infection with Trichomonas gallinae (1–3). Typical lesions in the upper digestive tract range from mild inflammation to caseous masses that can partially obstruct the digestive or respiratory tracts and progress to systemic disease (3). Trichomonosis can be present as subclinical to subacute, depending on the host species, immune status and the pathogenicity of the strain (1, 2, 4–8). Transmission can be direct, such as through “crop milk” or consumption of infected prey or indirect such as through contaminated water (3, 9–11).
Trichomonas gallinae is an important pathogen of Columbiformes, Falconiformes, and Passeriformes (3). It has been implicated in the decline of Band-tailed pigeons (Patagioenas fasciata) in western USA (2) and a highly pathogenic strain has recently emerged in North-Western Europe, causing population declines of finch species (3, 7, 12). Columbiformes are maintenance hosts for T. gallinae and the Rock dove (Columba livia) is considered the natural host of this pathogen (3). All species of Columbiformes occurring in Europe have been shown to harbor endemic Trichomonas gallinae infection (8, 13, 14). Particularly the Turtle dove (Streptopelia turtur) was shown to be heavily infected by Trichomonas gallinae (8, 15). European populations of Turtle doves heavily declined in the last decades and their migration routes pass through the Iberian Peninsula (16), with the potential of disseminating new strains. On the other hand, raptors are considered spillover hosts, acquiring infection from infected prey (3, 17).
Bonelli's eagle (Aquila fasciata) is a conservation priority species in Europe because of widespread declines in the last decades of the 20th century (18, 19). Most of the European population inhabits the Iberian Peninsula where its diet is based on Columbiformes, Red-legged partridge (Alectoris rufa) and Wild rabbit (Oryctolagus cuniculus) (20–22). Marginal populations of this species were shown to have low genetic variability (23), which could make them more vulnerable to emerging pathogens (24). Trichomonosis was shown to kill 2–14% of nestling Bonelli's eagles in several Iberian populations (25, 26). A marginal population of Bonelli's eagle comprising 22–24 pairs breeds in the Douro Internacional/Arribes del Duero natural parks and adjoining areas in northwestern Iberian Peninsula (19, 27, 28). This population is characterized by a low productivity, which has been linked to food shortage (28). Conservation management actions include supplementary feeding, either direct or through restocking of traditional pigeon lofts with Rock doves (27, 29).
Traditional pigeon lofts (TPL) are human constructions typical of Northwestern Iberian Peninsula (Figure 1), scattered across the landscape and originally aimed at producing Rock doves for human consumption and organic fertilizer for crops (30). These buildings provide nocturnal shelter and breeding sites for Rock doves, which are otherwise free-ranging (29, 30). Nowadays most TPL are abandoned but some have been recovered and restocked to increase prey availability for Bonelli's eagles (29).

Figure 1. Images of traditional pigeon lofts. (A) Landscape view; (B) outside view; (C) inside with Rock doves Columba livia.
The aim of this study was to investigate the epidemiology of Trichomonas spp. in the community of free-ranging domestic and wild Columbiformes and their endangered avian predator, Bonelli's eagle, in an area of Northwestern Iberian Peninsula. Additionally, we aimed to contribute to elucidating if restocking TPL with Rock doves could increase the exposure of Bonelli's eagles to infection by Trichomonas spp.
The study area consists of Douro Internacional/Arribes del Duero natural parks (comprising 193,000 hectares) and adjoining areas in Northwestern Iberian Peninsula (centroid 41° 11′ 36′ ′ N, 6° 45′ 49′′ W), encompassing both Portuguese and Spanish territory (Figure 2A). The area is an extensive plateau (600–800 m asl) where Douro river and several main tributaries (Águeda, Sabor, Côa, Tormes, and Huebra rivers) carved steep canyons in granite or shale rock. The landscape is highly fragmented, with oak (Quercus pyrenaica, Q. faginea, Q. rotundifolia, and Q. suber) and juniper (Juniperus oxycedrus) woods interspersed with shrub (mainly Cytisus sp. and Cistus sp.) and agricultural areas (mainly wheat Triticum sp., olive Olea europaea, vine Vitis vinifera, and pasture). The area is included in the Mesomediterranean and Supramediterranean bioclimatic zones showing Mediterranean-Subcontinental climate with large thermal amplitude, hot and dry summers and relatively cool and humid winters (31, 32).
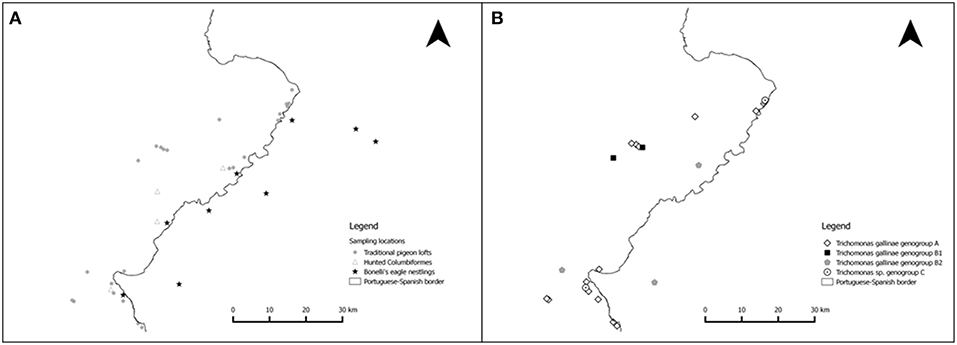
Figure 2. Location of the sample collection sites and of the Trichomonas sp. genogroups isolated. (A) Sampled traditional pigeon lofts (circles), wild Columbiformes (triangles), and Bonelli's eagle nests (stars). Due to the threatened status of Bonelli's eagles and their sensitivity to disturbance, the location of the sampling sites for this species is not exact. (B) Geographical sampling location of the Trichomonas sp. genogroups identified in the study area.
From August 2008 to October 2009 we surveyed Trichomonas spp. in 253 live-captured Rock doves and 41 hunted Columbiformes, comprising 32 Turtle doves, 6 Wood pigeons (Columba palumbus) and 3 Rock doves. Rock doves were live-captured on 24 TPL (Figure 2A) by closing the exits during the night and manually capturing them inside. To prevent repeated sampling of individuals, each TPL was sampled only once and Rock doves were kept in cages and released when all were sampled (<2 h of restraint). Samples from wild Columbiformes were obtained from animals hunted for recreational purposes, according to Portuguese and European legislation. No animals were killed for the purpose of this study. This study was carried out in accordance with national and European legislation and the recommendations of the “Guidelines on the care and use of wildlife” (33).
From May 2014 to June 2016 we surveyed Trichomonas spp. in 16 nestling Bonelli's eagles, sampled during regular monitoring for ringing (Figure 2A). Samples were obtained under permits 412/2014 and 316/2016 (Junta de Castilla y León, Spain) and 415/2014 (Instituto de Conservação da Natureza e Florestas, Portugal).
An oro-esophageal swab was obtained from each bird, immediately inoculated in InPouch™ media (Biomed Diagnostics Inc., White City, OR, USA) and kept at room temperature and away from sunlight until incubation (7–50 h post collection). From individual Rock doves, we collected data on sex, age class, weight, wing length, and plumage (wild-type or other). The residuals of a linear regression between log weight and log wing length of each Rock dove were used as indicators of body condition. The presence of macroscopic lesions compatible with trichomonosis was systematically investigated in the Bonelli's eagle nestlings but not in the Columbiformes.
Data were collected on each TPL during the sampling procedure and included geographical location, number of Rock doves, presence of other avian species, provision of water, location within occupied Bonelli's eagle territories (<3 km straight line from the center of a territory occupied by a pair during the previous breeding season) and the interval between collection and incubation of the biological samples.
Culture media were incubated at 37°C for 6 days and isolation of Trichomonas spp. was assessed by daily bright field microscopy at 100x magnification and considered positive when motile, flagellated protozoans were present in the culture media. Media containing Trichomonas were centrifuged at 1,430 × g for 10 min, the supernatant discarded, and the pellet re-suspended in PBS and stored at −20°C. DNA was extracted from a subset of 32 Trichomonas isolates, 1–2 per host species and TPL studied, with DNeasy Blood & Tissue Kit (Qiagen) and purified with Exo-AP Clean-up (Thermo Scientific), according to the manufacturer's instructions. For amplification of the 18S and ITS1/5.8S/ITS2 rRNA genes, previously established primers HM-LONG-F [5' AGGAAGCACACTATGGTCATAG 3'; (34)] and TFR1 [5' TGCTTCAGTTCAGCGGGTCTTCC 3'; (35)] were used. PCR amplification was performed with a PCR Master Mix (Phusion). Cycling parameters consisted of an initial denaturation at 98°C for 3 min, followed by 45 cycles of 98°C for 30 s, 61°C for 30 s and 72°C for 1 min, and 72°C for 5 min for the final extension step. products were purified and sequenced on an automatic sequencer PRISM 310 Genetic Analyzer (PE Applied Biosystems, Foster City, CA, USA) with the amplification primers and internal primer HM-LONG-R [5' CGTTACCTTGTTACGACTTCTCCTT 3'; (34)]. The sequences were deposited in GenBank under the following accession numbers: MK932769-773, MK932775-777. The ribotype nomenclature proposed by Gerhold et al. (36) and Grabensteiner et al. (37) is used throughout this paper.
Differences in prevalence between TPL were analyzed by non-parametric methods (Kruskal-Wallis test). We performed a binomial GLM with Trichomonas spp. infection status in individual Rock doves as dependent variable and individual traits (age class, sex, body condition, and plumage type) as independent variables (Table 1). We performed another GLM with Trichomonas spp. prevalence in TPL as dependent variable and environmental and sampling features as independent variables (number of Rock doves, presence of other avian species and water availability inside the TPL, location within an occupied Bonelli's eagle territory, and interval from collection of samples to start of incubation) (Table 1). We considered all TPL <3 km straight distance from an occupied nest as being located within an occupied territory, based on the home ranges of breeding Bonelli's eagles in the Iberian Peninsula (38, 39). Collinearity was checked by calculating the Variance Inflation Factor with a threshold of 4 and goodness of fit assessed by calculating the Hosmer-Lemeshow χ2. Statistical analyses were performed in R-3.3.2 (R Development Core Team). Maps were produced in QGIS 2.18.0 (QGIS Development Team). Maximum-Likelihood (ML) trees were inferred for the ITS1/5.8S/ITS2 and 18S rRNA regions (Table 3) in MEGA6 (40) under the best-fit nucleotide substitution model determined by the same software. Node support was determined from 500 bootstrap replicates of the ML trees.
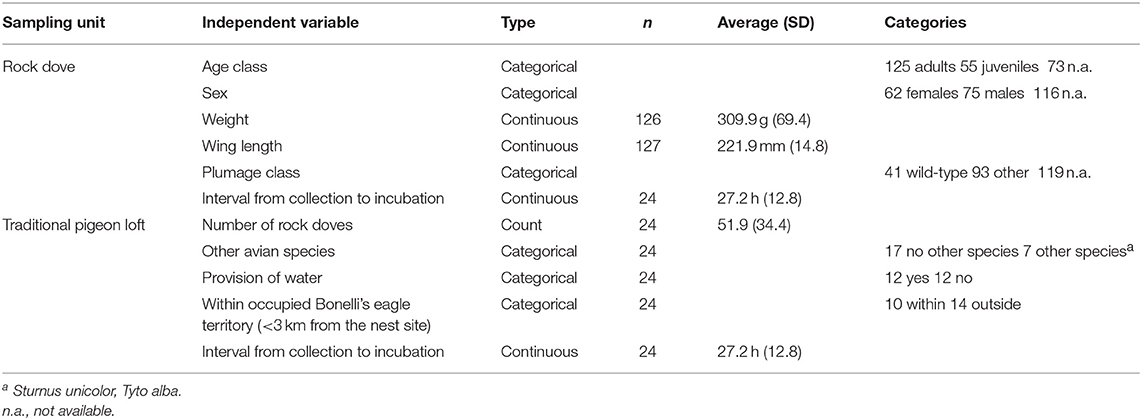
Table 1. Summary of the independent variables included in the statistical analysis.
Trichomonas spp. were isolated from 1/16 (6.3%, CI95 1.1–28.3) nestling Bonelli's eagles, 18/32 Turtle doves (56.3%, CI95 39.3–71.8, n = 32), 5/6 Wood pigeons (83.3%, CI95 43.7–97.0), and 175/256 Rock doves (68.4%, CI95 62.4–73.8) (Figure 3A). Prevalence of Trichomonas spp. in TPL ranged 14.3–100% (Figure 3B), showing statistically significant differences between TPL (p < 0.001, Kruskal-Wallis test).
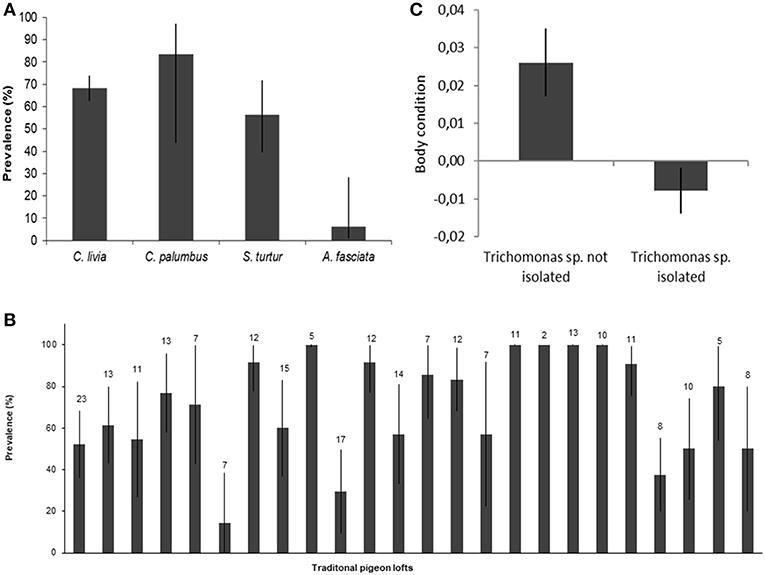
Figure 3. Determinants of Trichomonas sp. infection. Trichomonas sp. prevalence with 95% confidence intervals by (A) species; (B) traditional pigeon loft, with the sample size for each TPL; (C) Rock dove body condition by Trichomonas sp. infection status, with standard error of the mean.
In the binomial GLM with Trichomonas spp. isolation in individual Rock doves as dependent variable (Hosmer-Lemeshow χ2 = 6.445, p = 0.598), the only significant determinant of infection was the body condition, which was significantly lower (p = 0.021) in infected Rock doves (Table 2 and Figure 3C). In the GLM with Trichomonas spp. prevalence in TPL as dependent variable (Hosmer-Lemeshow χ2 = −1.2273, p = 1), no significant determinants of prevalence of infection were identified (Table 2).
Table 2. GenBank entries used in the phylogeny of the ITS1/5.8S/ITS2 and 18S rRNA regions.

Table 3. Results of the generalized linear models. Analysis of the potential determinants of Trichomonas sp. isolation in individual Rock doves and of prevalence in traditional pigeon lofts.
Three ribotypes of Trichomonas gallinae and one ribotype of Trichomonas sp. were obtained from the 32 isolates that were sequenced (Table 4 and Figure 4). Trichomonas gallinae ribotype 18S-VI, ITS-IV/B (n = 23) was distributed in TPL throughout the study area, while T. gallinae ribotype 18S-IV, ITS-I/D (n = 3) was detected in 2 TPL recently restocked with Rock doves acquired from a mixed-species avian collection. Both were only detected in Rock doves. Trichomonas gallinae ribotype 18S-II, ITS-I/D (n = 4) was detected in Rock doves from 2 TPL, 1 Wood pigeon and 1 nestling Bonelli's eagle, throughout the study area. Ribotype 18S-VIII, ITS-III of Trichomonas sp. (n = 3) was detected in 2 Rock doves from a single TPL and in 1 hunted Turtle dove in the northern and southern parts of the study area, respectively (Table 4 and Figure 2B).
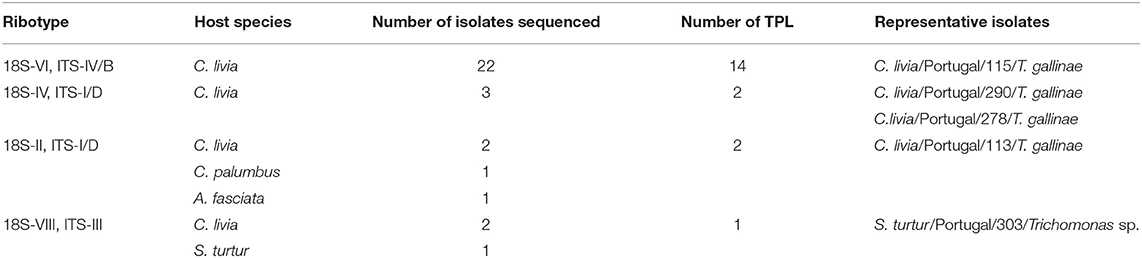
Table 4. Ribotypes of Trichomonas sp. identified in our sample based on ITS1/5.8S/ITS2 and 18S RNA sequences. Number of the isolates per species and traditional pigeon lofts (TPL). Ribotype nomenclature follows Gerhold et al. (36) and Grabensteiner et al. (37).
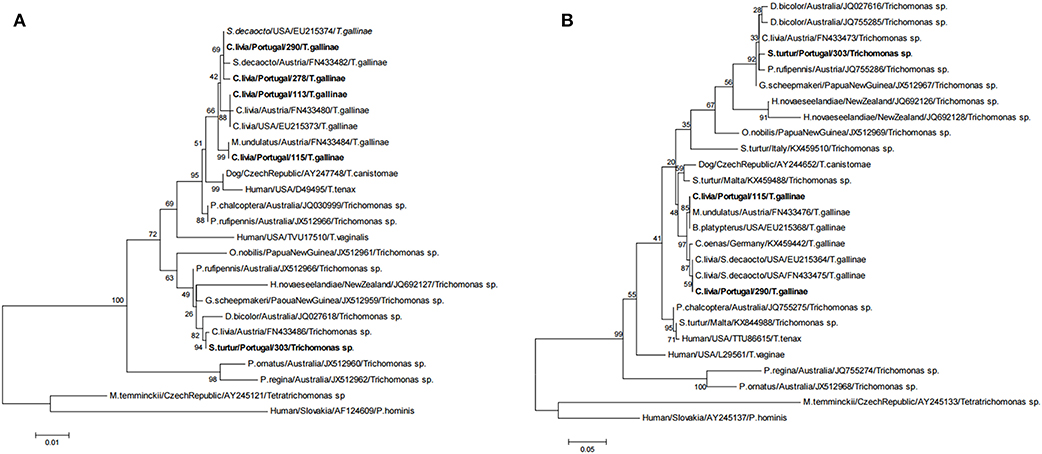
Figure 4. Phylogenetic tree of the Trichomonas isolates based on (A) 18S rRNA (1155bp) and (B) ITS1/5.8S/ITS2 (347bp). The phylogenetic analyses were estimated using the Maximum Likelihood (ML) method available in MEGA6 (40) under the best-fit nucleotide substitution model determined by the same software. Node support was determined from 500 bootstrap replicates of the ML trees. GenBank accession numbers of the retrieved sequences are indicated.
This study reports the epidemiology of Trichomonas spp. infection in the community of maintenance hosts for this pathogen and apparent spillover to an endangered avian predator. These results suggest that at least the Trichomonas gallinae ribotype 18S-II, ITS-227 I/D (n = 4), and ribotype 18S-VIII, ITS-III of Trichomonas sp. are maintained in the study area by a multi-host-pathogen system (41), where infection is likely transmitted between several species of free-ranging domestic and wild Columbiformes, as also documented in the United Kingdom (13). The inter-species transmission of Trichomonas spp. is supported by the high prevalence of infection in all studied Columbiformes species and particularly by the isolation of two genogroups in multiple species of Columbiformes.
Four distinct ribotypes of Trichomonas were detected in Rock doves, two of which are presumed to naturally occur in the study area (T. gallinae ribotypes 18S-VI, ITS-IV/B and 18S-II, ITS-I/D), as they were previously reported in this host species elsewhere in Europe (42). On the other hand, T. gallinae ribotype 18S-IV, ITS-I/D was previously reported only in Eurasian collared doves (36, 37), this being to our knowledge the first report of its isolation from Rock doves. Trichomonas gallinae ribotype 18S-IV, ITS-I/D was detected in the present study in two TPL recently restocked with Rock doves acquired in a mixed species collection. This observation highlights the risk of introducing new pathogen strains in local host communities by means of translocations for conservation purposes (43, 44). Here we report the first isolation in European wild Columbiformes of Trichomonas sp. ribotype 18S-VIII, ITS-III. This genogroup was previously detected in Europe only in 2007, in a domestic Rock dove in Austria (37).
Interestingly the most common and widespread T. gallinae ribotype 18S-VI, ITS-IV/B was only detected in Rock doves, while the less common T. gallinae ribotype 18S-II, ITS-I/D was also detected in Wood pigeon and Bonelli's eagle. Whether this corresponds to a greater ability of the latter for spillover to other host species is still to be determined. Differential inter-species transmission rates have been suggested for other T. gallinae genogroups, as well as differential pathogenicity (7, 15). As we only sequenced the 18S and ITS1/5.8S/ITS2 rRNA genes, more genetic diversity could have gone unnoticed in our sample of Trichomonas isolates. Furthermore, the opportunistic sampling performed in Bonelli's eagle, due to constraints in accessing nests of this endangered species, means there is no temporal overlap between samples collected from this species and those from Columbiformes.
Trichomonosis can be an important cause of mortality for nestling Bonelli's eagles; nevertheless, subclinical infections are more common than clinical ones (25, 26). In the present investigation only one nestling was shown to harbor Trichomonas gallinae, but no lesions were detected during handling in the nest and it survived to fledging. We could not isolate Trichomonas from two other nestlings with compatible lesions (oropharyngeal caseous masses), which could be due to a different etiology or a long interval between collection of samples and incubation. Hernández et al. (27) also estimated a low prevalence in this same population (10%). Nevertheless, the low sample size precludes drawing conclusions on the pathogenicity and potential impact of infection by Trichomonas gallinae on this Bonelli's eagle population.
Comparable studies in other populations of Bonelli's eagle report higher prevalence in nestlings, which could be due to the low sample size or to the relatively long interval between collection of the samples and incubation in our study. Trichomonas gallinae was isolated from 36 to 68.8% of nestlings in 3 populations from Southern Iberian Peninsula (5, 25, 26), where Columbiformes are 28.5–39.2% of the prey items of Bonelli's eagles (20, 45). In our study population, Columbiformes are 20.6% of the diet of this species, with a preponderance of Rock doves over Wood pigeons (27). Given the high prevalence of infection by Trichomonas gallinae in all species of domestic and wild Columbiformes and the importance of these prey on the diet of Bonelli's eagles across its Iberian range, most if not all nestlings should be exposed to Trichomonas sp. (26). The variable prevalence of infection in nestlings from different populations could be explained by individual, regional and temporal heterogeneities in the pathogenicity of Trichomonas gallinae strains present in the local population of Columbiformes or the immune status of Bonelli's eagle nestlings. Marginal Iberian populations of Bonelli's eagle were shown to have low genetic diversity (23), which could impair their immune response (24). Further large-scale studies on this predator-parasite-prey system, addressing the link between diet and Trichomonas gallinae prevalence, are needed to elucidate the reasons for this heterogeneity in prevalence between populations of Bonelli's eagles.
One of the drivers of this study was the concern that restocking TPL with Rock doves would increase the exposure of Bonelli's eagles to Trichomonas gallinae (46). While this could occur if the consumption of Rock doves increases in response to this conservation action, we show that Trichomonas spp. infection is widespread in the wild Columbiformes community. The single case of infection in a Bonelli's eagle nestling took place in a territory where no TPL restocking was carried out. Nevertheless, emerging Trichomonas spp. strains might be amplified in TPL, providing opportunity for spillover to Bonelli's eagles. Systematic disease surveillance in TPL should be implemented to detect the potential introduction of new strains and contingency plans prepared to prevent spillover to endangered avian predators.
Turtle doves in our sample showed lower prevalence than those reported elsewhere in Europe [67–100% - (8, 13–15)]. European populations of this species have undergone a sustained decline in the last decades, and trichomonosis has been suggested as contributing for this decline (8, 15, 16). The prevalence we determined in other species are comparable to those reported in the literature: 34–70% in Wood pigeon throughout Europe (8, 13, 14, 47) and 45–79% in Rock doves in Spain (5, 14).
Interestingly we observed that Rock doves from which Trichomonas spp. were isolated showed poorer body condition. It was shown in other host-pathogen systems that the physiological costs of immune system activation to fight infection trade-off with other physiological needs, such as reproduction, foraging and growth (48, 49). In our sample this translated into a lower body condition of infected Rock doves, as previously shown to occur in other free-ranging Columbiformes infected with T. gallinae (15, 47). The lower body condition of infected Rock doves could heighten the risk of predation, potentiating interspecies transmission, but being located within a Bonelli's eagle territory was not found to be a risk factor for prevalence in TPL. Studies including the whole community of avian predators of Rock doves are needed to address this question.
Trichomonas spp. seem to persist in the study area in a multi-host ecosystem, where intra- and inter-species transmission likely contribute to maintenance, making it demanding to individualize the role of each host species in the epidemiology of infection (41). More detailed molecular epidemiology tools have the potential to further contribute to unveiling the dynamics of pathogen transmission in this host community. Long-term studies will allow uncovering the origin of infection for avian predators such as the Bonelli's eagle, potentially highlighting ways to control spillover to this endangered species. Meanwhile, conservation actions aimed at increasing the availability of trophic resources for Bonelli's eagles should consider the potential risk of increased disease transmission and minimize the risk of introducing alien Trichomonas spp. strains.
This study was carried out in accordance with the recommendations of guidelines on the care and use of wildlife (33), Portuguese and European legislations. The study was conducted under permits 412/2014 and 316/2016 (Junta de Castilla y León, Spain) and 415/2014 (Instituto de Conservação da Natureza e Florestas, Portugal).
NS, JJ, AM, JG, JrA, and PE designed the study. NS, JJ, AM, JrA, NM, JG, AF, and TA performed the field and laboratory work. NS, TA, KT, JnA, and PE analyzed data and wrote the manuscript. All authors reviewed the manuscript upon submission.
This study was financed by the authors' institutions.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We acknowledge the collaboration of Parque Natural de los Arribes del Duero (Spain), Parque Natural do Douro Internacional, Associação Transumância e Natureza and Palombar (Portugal), as well as the owners of pigeon lofts and hunters for allowing collecting samples and the volunteers who helped with sample collection. FCT supported the post-doctoral fellowship of NS (SFRH/BPD/116596/2016).
1. Bunbury N, Jones CG, Greenwood AG, Bell DJ. Epidemiology and conservation implications of Trichomonas gallinae infection in the endangered Mauritian pink pigeon. Biol Conserv. (2008) 141:153–61. doi: 10.1016/j.biocon.2007.09.008
2. Robinson RA, Lawson B, Toms MP, Peck KM, Kirkwood JK, Chantrey J, et al. Emerging infectious disease leads to rapid population declines of common British birds. PLoS ONE. (2010) 5:e12215. doi: 10.1371/journal.pone.0012215
3. Amin A, Bilic I, Liebhart D, Hess M. Trichomonads in birds – a review. Parasitology. (2014) 141:733–47. doi: 10.1017/S0031182013002096
4. Höfle U, Gortázar C, Ortíz JA, Knispel B, Kaleta EF. Outbreak of trichomoniasis in a woodpigeon (Columba palumbus) wintering roost. Eur J Wildl Res. (2004) 50:73–7. doi: 10.1007/s10344-004-0043-2
5. Sansano-Maestre J, Garijo-Toledo MM, Gómez-Muñoz MT. Prevalence and genotyping of Trichomonas gallinae in pigeons and birds of prey. Avian Pathol. (2009) 38:201–7. doi: 10.1080/03079450902912135
6. Sansano-Maestre J, del Carmen Martínez-Herrero M, Garijo-Toledo MM, Gómez-Muñoz MT. RAPD analysis and sequencing of ITS1/5.8 S rRNA/ITS2 and Fe-hydrogenase as tools for genetic classification of potentially pathogenic isolates of Trichomonas gallinae. Res Vet Sci. (2016) 107:182–9. doi: 10.1016/j.rvsc.2016.05.016
7. Chi JF, Lawson B, Durrant C, Beckmann K, John S, Alrefaei AF, et al. The finch epidemic strain of Trichomonas gallinae is predominant in British non-passerines. Parasitology. (2013) 140:1234–45. doi: 10.1017/S0031182013000930
8. Marx M, Reiner G, Willems H, Rocha G, Hillerich K, Masello JF, et al. High prevalence of Trichomonas gallinae in wild columbids across western and southern Europe. Parasit Vectors. (2017) 10:242. doi: 10.1186/s13071-017-2170-0
9. Purple KE, Humm JM, Kirby RB, Saidak CG, Gerhold R. Trichomonas gallinae persistence in four water treatments. J Wildl Dis. (2015) 51:739–42. doi: 10.7589/2014-05-137
10. Sturgill F, Gerhold R. Trichomonas gallinae persistence in water with pH and organic material variance. Comp Parasitol. (2016) 83:237–9. doi: 10.1654/4799s.1
11. McBurney S, Kelly-Clark WK, Forzán MJ, Vanderstichel R, Teather K, Greenwood SJ. Persistence of Trichomonas gallinae in Birdseed. Avian Dis. (2017) 61:311–5. doi: 10.1637/11545-113016-RegR1
12. Lawson B, Robinson RA, Colvile KM, Peck KM, Chantrey J, Pennycott TW, et al. The emergence and spread of finch trichomonosis in the British Isles. Philos Trans Royal Soc B. (2012) 367:2852–63. doi: 10.1098/rstb.2012.0130
13. Lennon RJ, Dunn JC, Stockdale JE, Goodman SJ, Morris AJ, Hamer KC. Trichomonad parasite infection in four species of Columbidae in the UK. Parasitology. (2013) 140:1368–76. doi: 10.1017/S0031182013000887
14. Martínez-Herrero MC, Sansano-Maestre J, Márquez IL, Obón E, Ponce C, González J, et al. Genetic characterization of oropharyngeal trichomonad isolates from wild birds indicates that genotype is associated with host species, diet and presence of pathognomonic lesions. Avian Pathol. (2014) 43:535–46. doi: 10.1080/03079457.2014.967660
15. Stockdale JE, Dunn JC, Goodman SJ, Morris AJ, Sheehan DK, Grice PV, et al. The protozoan parasite Trichomonas gallinae causes adult and nestling mortality in a declining population of European Turtle Doves, Streptopelia turtur. Parasitology. (2015) 142:490–8. doi: 10.1017/S0031182014001474
16. Calderón L, Campagna L, Wilke T, Lormee H, Eraud C, Dunn JC, et al. Genomic evidence of demographic fluctuations and lack of genetic structure across flyways in a long-distance migrant, the European turtle dove. BMC Evol Biol. (2016) 16:237. doi: 10.1186/s12862-016-0817-7
17. Rogers KH, Girard YA, Woods L, Johnson CK. Avian trichomonosis in spotted owls (Strix occidentalis): indication of opportunistic spillover from prey. Int J Parasitol: Parasit Wildl. (2016) 5:305–11. doi: 10.1016/j.ijppaw.2016.10.002
18. Real J, Mañosa S. Demography and conservation of western European Bonelli's Eagle Hieraaetus fasciatus populations. Biol Conserv. (1997) 79:59–66. doi: 10.1016/S0006-3207(96)00100-0
19. Del Moral JC. El Águila Perdicera en España. Población en 2005 y Método De Censo. Madrid: SEO/BirdLife (2006).
20. Palma L, Beja P, Pais M, Fonseca LC. Why do raptors take domestic prey? The case of Bonelli's eagles and pigeons. J Appl Ecol. (2006) 43:1075–86. doi: 10.1111/j.1365-2664.2006.01213.x
21. Caro J, Ontiveros D, Pleguezuelos JM. The feeding ecology of Bonelli's eagle (Aquila fasciata) floaters in southern Spain: implications for conservation. Eur J Wildl Res. (2011) 57:729–36. doi: 10.1007/s10344-010-0480-z
23. Mira S, Arnaud-Haond S, Palma L, Cancela ML, Beja P. Large-scale population genetic structure in Bonelli's Eagle Aquila fasciata. Ibis. (2013) 155:485–98. doi: 10.1111/ibi.12065
24. Hawley DM, Sydenstricker KV, Kollias GV, Dhondt AA. Genetic diversity predicts pathogen resistance and cell-mediated immunocompetence in house finches. Biol Lett. (2005) 1:326–9. doi: 10.1098/rsbl.2005.0303
25. Höfle U, Blanco J, Palma L, Melo P. Trichomoniasis in Bonelli's eagles Hieraaetus fasciatus nestlings in south-west Portugal. In: eds Lumeij J, Remple J, Redig P, Lierz P, Cooper M. Raptor Biomedicine III (Lake Worth: Zoological Education Network) (2000). p. 45–52.
26. Real J, Mañosa S, Muñoz E. Trichomoniasis in a Bonelli's eagle population in Spain. J Wildl Dis. (2000) 36:64–70. doi: 10.7589/0090-3558-36.1.64
27. Hernández A, Resano J, Real J. Monitorización demográfica del Águila perdicera (Aquila fasciata) en Castilla y León, análisis de la dieta, de la calidad de los pollos y aplicaciones a la conservación - informe final. Technical Report Equip de Biologia de la Conservació, Departament de Biologia Animal, Universitat de Barcelona. Barcelona (2012).
28. Hernández-Matías A, Real J, Moleón M, Palma L, Sánchez-Zapata JA, et al. From local monitoring to a broad-scale viability assessment: a case study for the Bonelli's Eagle in western Europe. Ecol Monogr. (2013) 83:239–61. doi: 10.1890/12-1248.1
29. Sousa AJG. Caracterização ecológica e genética das populações de Pombo-da-rocha (Columba livia) no Parque Natural do Douro Internacional. Technical report, Instituto da Conservação da Natureza, Parque Natural do Douro Internacional, Figueira de Castelo Rodrigo. Portugal (2004).
30. García JEY. El Palomar Tradicional: Una Explotación Avícola Singular, Estúdio de Antecedentes Históricos y Aprovechamiento Humano. Zamora: Colegio Oficial de Veterinarios de Zamora (1997).
31. Rivas-Martínez S, Cantó P, Fernandez-González F, Navarro C, Pizarro JM, Sanchez-Mata D. Biogeografia de la Peninsula Iberica, Islas Baleares y Canarias. Publicaciones Deptartamento Biologia Vegetal (1990).
32. Costa JC, Aguiar C, Capelo JH, Lousã M, Neto C. Biogeografia de Portugal Continental. Quercetea (1998).
33. CCAC Canadian Council on Animal Care Guidelines on: The Care and Use of Wildlife. Ontario: Canadian Council on Animal Care (2003).
34. Hess M, Kolbe T, Grabensteiner E, Prosl H. Clonal cultures of Histomonas meleagridis, Tetratrichomonas gallinarum and a Blastocystis sp. established through micromanipulation Parasitology. (2006) 133:547–54. doi: 10.1017/S0031182006000758
35. Felleisen RS. Comparative sequence analysis of 5.8S rRNA genes and internal transcribed spacer (ITS) regions of trichomonadid protozoa. Parasitology. (1997) 115:111–9. doi: 10.1017/S0031182097001212
36. Gerhold RW, Yabsley MJ, Smith AJ, Ostergaard E, Mannan W, Cann JD, et al. Molecular characterization of the Trichomonas gallinae morphologic complex in the United States. J Parasitol. (2008) 94:1335–41. doi: 10.1645/GE-1585.1
37. Grabensteiner E, Bilic I, Kolbe T, Hess M. Molecular analysis of clonal trichomonad isolates indicate the existence of heterogenic species present in different birds and within the same host. Vet Parasitol. (2010) 172:53–64. doi: 10.1016/j.vetpar.2010.04.015
38. Bosch R, Real J, Tinto A, Zozaya EL, Castell C. Home-ranges and patterns of spatial use in territorial Bonelli's Eagles Aquila fasciata. Ibis. (2010) 152:105–17. doi: 10.1111/j.1474-919X.2009.00985.x
39. Pérez-García JM, Margalida A, Afonso I, Ferreiro E, Gardiazábal A, et al. Interannual home range variation, territoriality and overlap in breeding Bonelli's eagles (Aquila fasciata) tracked by GPS satellite telemetry. J Ornithol. (2012) 154:63–71. doi: 10.1007/s10336-012-0871-x
40. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. (2013) 30:2725–9. doi: 10.1093/molbev/mst197
41. Holt RD, Dobson AP, Begon M, Bowers RG, Schauber EM. Parasite establishment in host communities. Ecol Lett. (2003) 6:837–42. doi: 10.1046/j.1461-0248.2003.00501.x
42. Ganas P, Jaskulska B, Lawson B, Zadravec M, Hess M, Bilic I. Multi-locus sequence typing confirms the clonality of Trichomonas gallinae isolates circulating in European finches. Parasitology. (2014) 141:652–61. doi: 10.1017/S0031182013002023
43. Cunningham AA. Disease risks of wildlife translocations. Conserv Biol. (1996) 10:349–53. doi: 10.1046/j.1523-1739.1996.10020349.x
44. Walker SF, Bosch J, James TY, Litvintseva AP, Valls JAO, Piña S, et al. Invasive pathogens threaten species recovery programs. Curr Biol. (2008) 18:853–4. doi: 10.1016/j.cub.2008.07.033
45. Resano-Mayor J, Hernández-Matías A, Real J, Parés F, Inger R, Bearhop S. Comparing pellet and stable isotope analyses of nestling Bonelli's Eagle Aquila fasciata diet. Ibis. (2014) 156:176–88. doi: 10.1111/ibi.12095
46. Rollan À, Hernández-Matías A, Real J. Guidelines for the Conservation of Bonelli's Eagle Populations. Barcelona: Universitat de Barcelona. (2016). Available online at: http://hdl.handle.net/2445/69446 (accessed April 2, 2019).
47. Villanúa D, Höfle U, Pérez-Rodríguez L, Gortázar C. Trichomonas gallinae in wintering common wood pigeons Columba palumbus in Spain. Ibis. (2006) 148:641–8. doi: 10.1111/j.1474-919X.2006.00561.x
48. Van der Most PJ, de Jong B, Parmentier HK, Verhulst S. Trade-off between growth and immune function: a meta-analysis of selection experiments. Funct Ecol. (2011) 25:74–80. doi: 10.1111/j.1365-2435.2010.01800.x
Keywords: Trichomonas gallinae, Columbiformes, Aquila fasciata, Streptopelia turtur, Columba livia, multi-host-pathogen system, wildlife conservation
Citation: Santos N, Jambas J, Monteiro A, Amaral J, Martins N, Garcia J, Fernández AM, Tyler KM, Almeida T, Abrantes J and Esteves PJ (2019) Trichomonas Infection in a Community of Free-Ranging Domestic and Wild Columbiformes and Bonelli's Eagle (Aquila fasciata). Front. Vet. Sci. 6:148. doi: 10.3389/fvets.2019.00148
Received: 17 January 2019; Accepted: 29 April 2019;
Published: 29 May 2019.
Edited by:
Francesco Carlo Origgi, University of Bern, SwitzerlandReviewed by:
Bjørnar Ytrehus, Norwegian Institute for Nature Research (NINA), NorwayCopyright © 2019 Santos, Jambas, Monteiro, Amaral, Martins, Garcia, Fernández, Tyler, Almeida, Abrantes and Esteves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nuno Santos, bnVuby5zYW50b3NAY2liaW8udXAucHQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.