 Po-Yun Teng
Po-Yun Teng Woo Kyun Kim
Woo Kyun Kim- Department of Poultry Science, University of Georgia, Athens, GA, United States
In recent years, prebiotics have been considered as potential alternatives to antibiotics. Mechanisms by which prebiotics modulate the ecosystem of the gut include alternation of the intestinal microbiota, improvement of the epithelium, and stimulation of the immune system. It is suggested that the administration of prebiotics not only influences these aspects but also regulates the interaction between the host and the intestinal microbiota comprehensively. In this review, we will discuss how each prebiotic ameliorates the ecosystem by direct or indirect mechanisms. Emphasis will be placed on the effects of prebiotics, including mannan oligosaccharides, β-glucans, and fructans, on the interaction between the intestinal microbiota, gut integrity, and the immunity of broilers. We will highlight how the prebiotics modulate microbial community and regulate production of cytokines and antibodies, improving gut development and the overall broiler health. Understanding the cross talk between prebiotics and the intestinal ecosystem may provide us with novel insights and strategies for preventing pathogen invasion and improving health and productivity of broilers. However, further studies need to be conducted to identify the appropriate dosages and better resources of prebiotics for refinement of administration, as well as to elucidate the unknown mechanisms of action.
Introduction
Since the use of antibiotic growth promoters was banned by the EU on January 1st, 2006, several feed additives have been studied as alternatives to antibiotics, such as probiotics, prebiotics, synbiotics, and herbal medicines (1). Among these feed additives, prebiotics have been studied and supplemented broadly into broiler diets in recent years. Gibson and Roberfroid (2) defined a prebiotic compound as a non-digestible food ingredient utilized by intestinal microbiota. It beneficially affects the host by selectively stimulating the growth and/or activity of one or a limited number of bacteria in the intestinal tract, consequently improving gut health and hosts' intestinal microbial balance. Gibson et al. (3) revised the definition and defined a prebiotic as a selectively fermented ingredient that allows specific changes in the composition and/or activity in the intestinal microbiota that confers benefits upon the host's well-being and health. Some researchers also confined prebiotics to indigestible oligosaccharides (4). Ideal characteristics of prebiotics were described by Patterson and Burkholder (5): (1) prebiotics should not be hydrolyzed by animal gastrointestinal enzymes, (2) prebiotics cannot be absorbed directly by cells in the gastrointestinal tracks, (3) prebiotics selectively enrich one or limited numbers of beneficial bacteria, (4) prebiotics alter the intestinal microbiota and their activities, and (5) prebiotics ameliorate luminal or systemic immunity against pathogen invasion.
The ecosystem of the gut is composed of three crucial elements: (1) microbial community, (2) intestinal epithelial cells, and (3) immune system (6). Generally, prebiotics can be fermented by health-promoting bacteria in the intestine, producing lactic acid, short-chain fatty acid (SCFA), or some antibacterial substances, such as bacteriocine against pathogenic species (7). These products may not only benefit the intestinal microbial structure but also improve the integrity of intestinal epithelial cells, which further increase the absorption of nutrients and enhance the growth performance of animals (8).
Intestinal microbiota are influenced by various factors, including diet, gender, background genotype, housing environment, litter, and also age of birds (9). These factors can alter the abundance of dominant bacterial phyla and families in each part of the intestine. For instance, gut microbiota in young chickens changed rapidly with increase of age. Clostridiaeae and Enterobacteriaceae are two dominant families in the ileum of 7 day-old chickens, whereas Lactobacillaceae and Clostridiacea represent the common families in the ileum of 35 day-old birds (9). However, the balance of intestinal microbiota is alterable. Application of prebiotics in diets could establish a healthy microbial community in the intestine of young broilers by enhancing the abundance of Lactobacilli and Bifidobacteria and reducing the titers of Coliform (10, 11).
Furthermore, the modulation of intestinal microbiota is associated with immune responses. On the one hand, inhibiting pathogen colonization by prebiotics can decrease detrimental molecules produced by pathogenic bacteria, which have been known as exogenous signals (12). These signals are also called pathogen-associated molecular patterns (PAMPs). The PAMPs can be recognized by pattern recognition receptors (PRR), including toll-like receptors (TLRs) and NOD-like receptors (NLRs), which are expressed on the surface of sentinel cells (13). Once PRRs recognize PAMPs, sentinel cells, such as epithelial cells, macrophages, mast cells, and dendritic cells, are activated, producing cytokines for the regulation of further innate immune responses. On the other hand, prebiotics can act as non-pathogenic antigens themselves. They can be recognized by receptors of immune cells, which consequently modulate host immunity beneficially.
Various prebiotics are composed of diverse sugar units. Therefore, each prebiotic may influence the animals differently. Here, we reviewed studies of broilers that discuss the effects of prebiotics on their underlying mechanisms of action. We will discuss the direct or indirect mechanisms by which prebiotics ameliorated the ecosystem of the chicken gut. Emphasis will be placed on the impacts of mannan oligosaccharides, β-glucans, and fructans on the interaction between the intestinal microbiota, immunity, and the integrity of the epithelial cells (Figures 1–3).
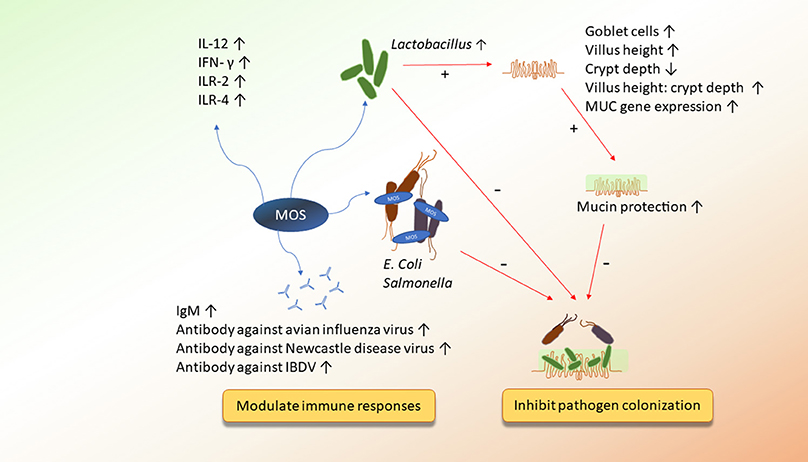
Figure 1. The potential mechanisms of action of MOS on improving immunity and inhibiting pathogen colonization.
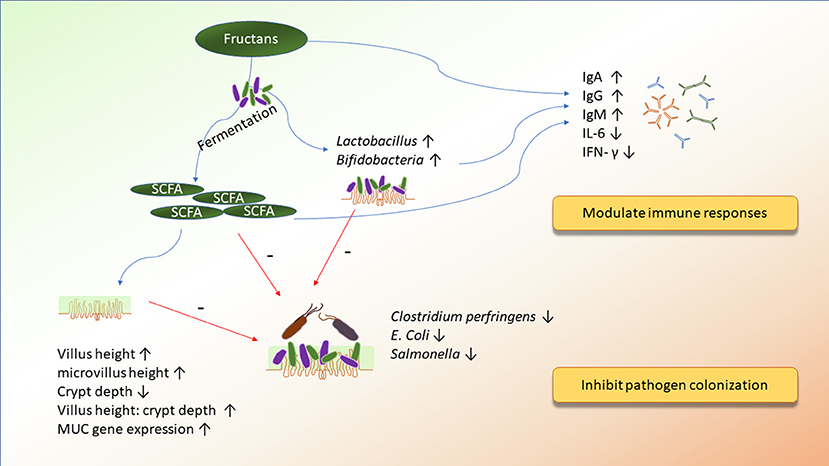
Figure 2. The potential mechanisms of action of fructans on improving immunity and inhibiting pathogen colonization.
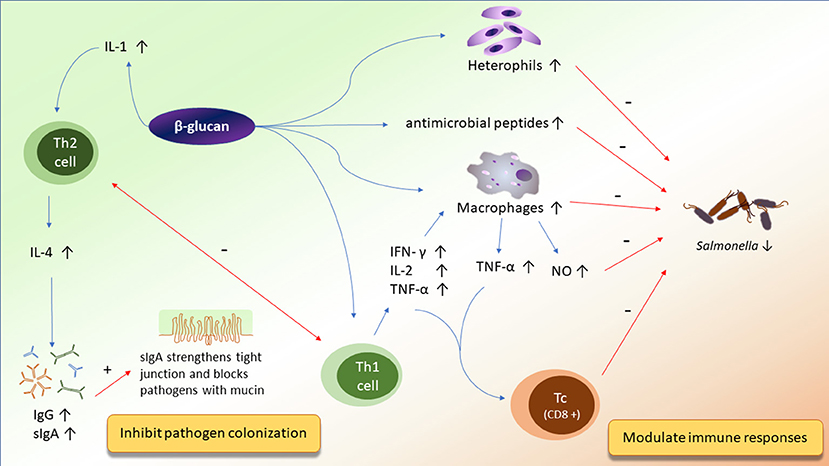
Figure 3. The potential mechanisms of action of β-glucan on improving immunity and inhibiting pathogen colonization.
Mannan Oligosaccharides (MOS)
Most of the mannan oligosaccharide (MOS) products are derived from yeast cell walls (Saccharomyces cerevisiae) and are rich in mannoproteins (12.5%), mannan (30%), and glucan (30%) (14, 15). Mannan oligosaccharides are known for their ability to bind pathogenic bacteria, which possess type-1 fimbriae, such as E. coli and Salmonella species (16). By blocking bacterial lectin, MOS could reduce colonization of these pathogens in the intestine of animals (17). Previous studies indicated that supplementation of MOS from 0.08 to 0.5% could alter cecal microbial community composition by increasing total anaerobic bacteria, Lactobacillus and Bifidobacterium, and decreasing Salmonella, E. coli, Clostridium perfringens, and Campylobacter (14, 16, 18–23). Apart from its effects on cecal microbiota, MOS also improved microbial community in other sections of the intestine, including the jejunum, the ileum, the jejunal mucosa, the ileal mucosa, and the ileocecal junction (11, 22, 24–26). It is interesting to note that MOS increased cecal Bacteroidetes in 7 and 35 day-old broilers (23, 27). Genus Bacteroides have been known for their strong metabolic activity. They can efficiently ferment indigestible polysaccharides to SCFA and, consequently, improve nutrient absorption and protect the host from pathogen infection (28). In previous studies, shown in Table 1, Lactobacillus species were the main species influenced by MOS. Mannan oligosaccharides increased the prevalence of ileal L. acetotolerans, L. delbrueckii subsp. lactis, L. sakei subsp. sakei. and cecal L. ingluviei, L. mucosae, L. salivarius, and L. crispatus (23, 29). Among these Lactobacillus species, L. crispatus was reported to have anti-E. coli and anti-Salmonella activities, whereas L. salivarius was mentioned to have the ability to limit Salmonella colonization (30, 31). The anti-pathogenic characteristics of Lactobacillus may be the reason why MOS reduced the numbers of E. coli or Salmonella in the intestine, ameliorating bacterial infection in pathogen-challenged broilers (14, 16, 19).
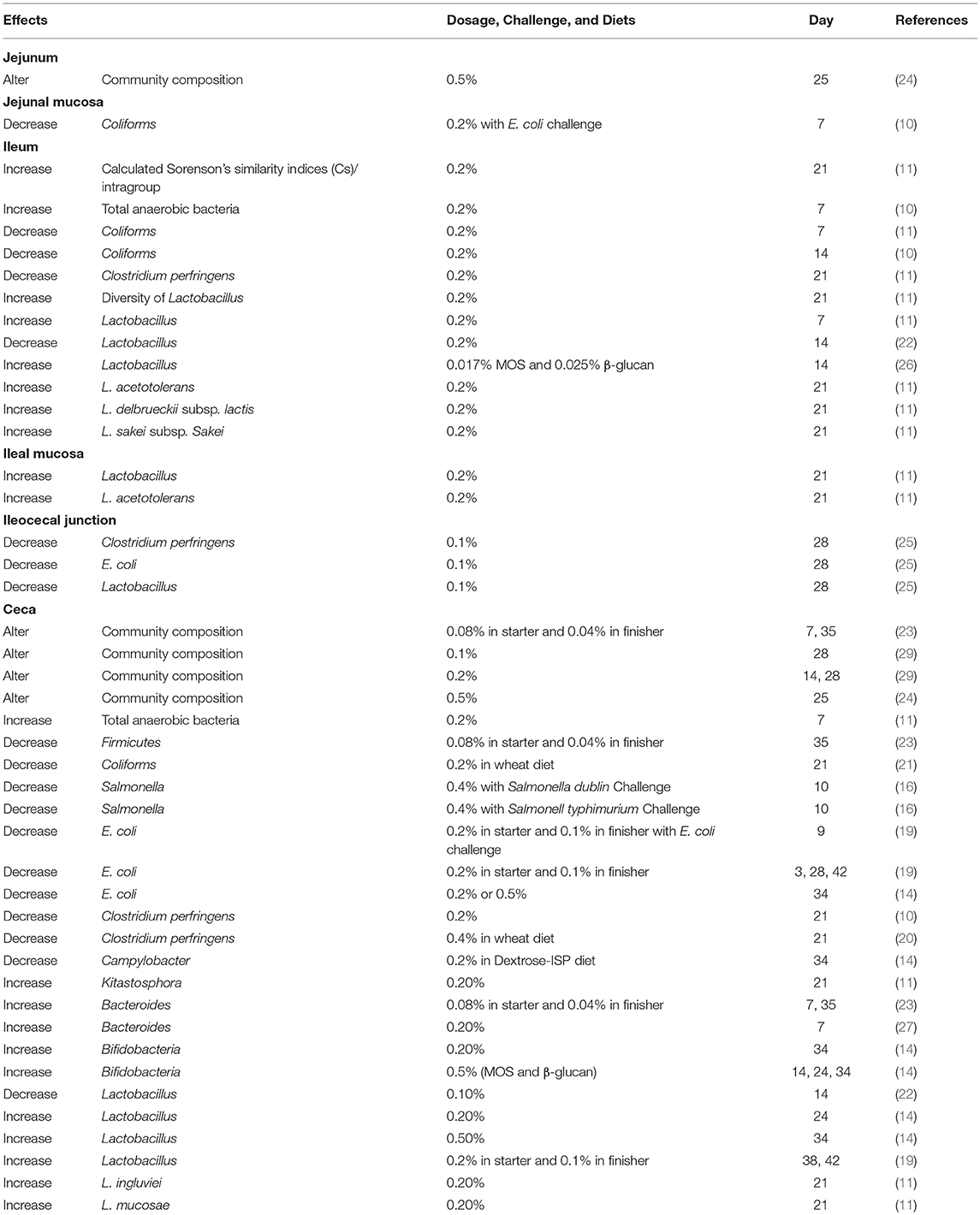
Table 1. Effects of mannan oligosaccharides on intestinal microbiota of broilers.
In addition, higher levels of intestinal Lactobacillus in birds fed with MOS may further result in the improvement of gut health status. Mannan oligosaccharides have been reported to increase villus height and surface area, decrease crypt depth, induce numbers of sulphated-acidic goblet cells, and upregulate gene expression of MUC, which is related to mucin secretion (10, 11, 14, 32–35) (Table 2). It has been reported that sulphated-acidic goblet cells are less degradable by the pathogen's glycosides (43, 44). Therefore, they can provide stronger protection against pathogens for the host. Similarly, Cheled-Shoval et al. (36) reported that in ovo administration of MOS enhanced villus area and proliferation of goblet cells. The greater numbers of goblet cells were able to increase the gene expression of MUC, synthesizing and secreting more mucin, which plays an important role as the first line of defense. Mucin can trap pathogens or impede them from invading epithelial cells (45). Thus, it is hypothesized that MOS establishes a bidirectional interaction: the increase of Lactobacillus counts may improve intestinal development, whereas mucin produced by goblet cells can conversely limit attachment of pathogens to epithelial cells.
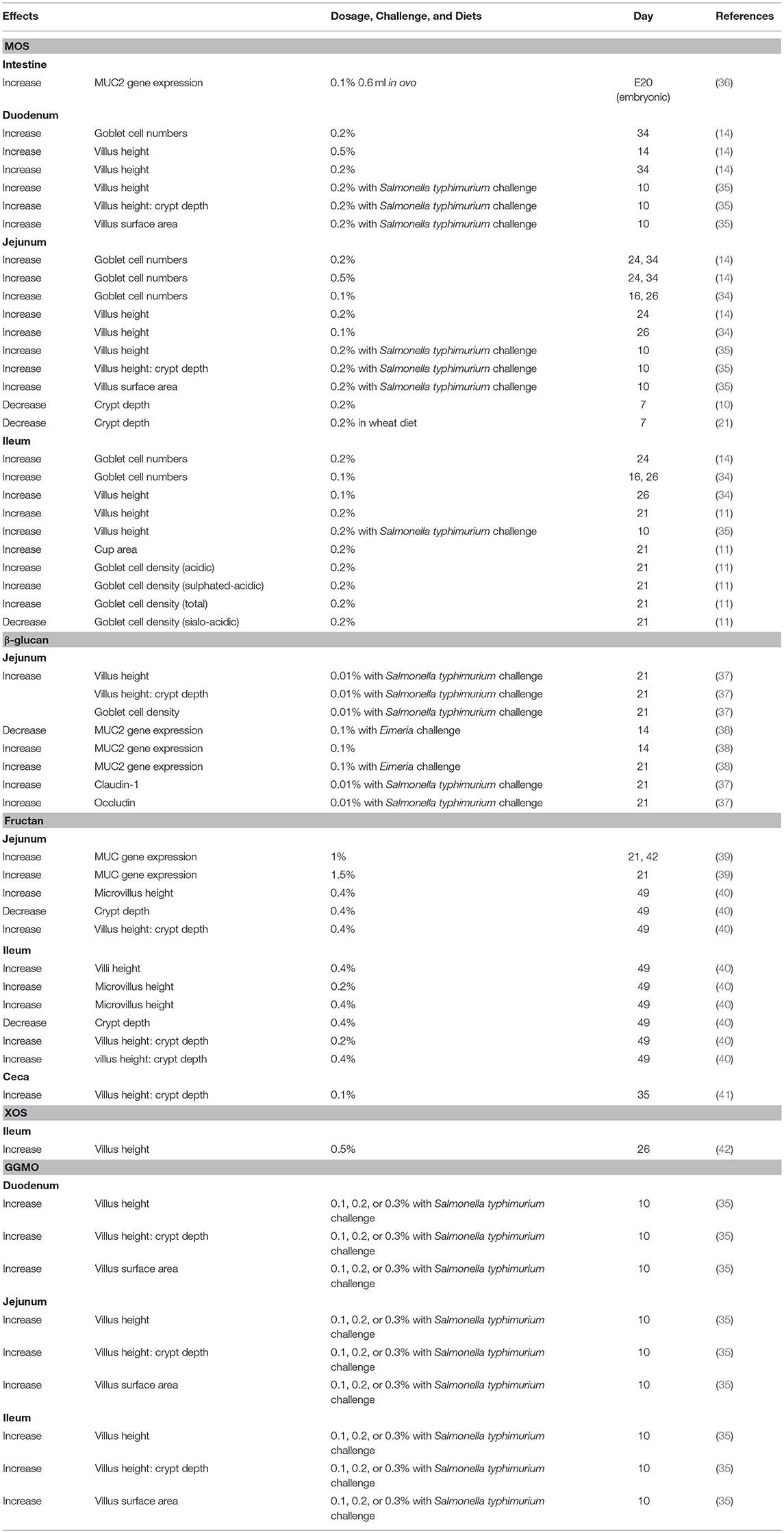
Table 2. Effects of prebiotics on intestinal morphology of broilers.
The effects of MOS on immunity of broilers are presented in Table 3. TLR4 and TLR2 were upregulated in the ileum or cecal tonsils by 0.2% MOS supplementation (50). It indicated that MOS could be recognized by both TLR4 and TLR2. Similar to mammalian TLR4, chicken TLR4 (chTLR4) mRNA has been found in a wide range of cells, particularly in macrophages and heterophils (61). TLR4 is a receptor that recognizes lipopolysaccharide (LPS) in mammals. After recognizing LPS, immune cells could produce high levels of nitric oxide and pro-inflammatory cytokines against pathogenic bacteria. Thus, it was suggested that reducing the exposure of LPS from E. coli by MOS could downregulate gene expression of chTLR4 and inhibit pro-inflammatory immunity (50). However, molecules of MOS can be recognized by TLR4 as well. It was reported that MOS may act as a pro-inflammatory factor that upregulates TLR4 gene expression and induces innate immune responses (62).
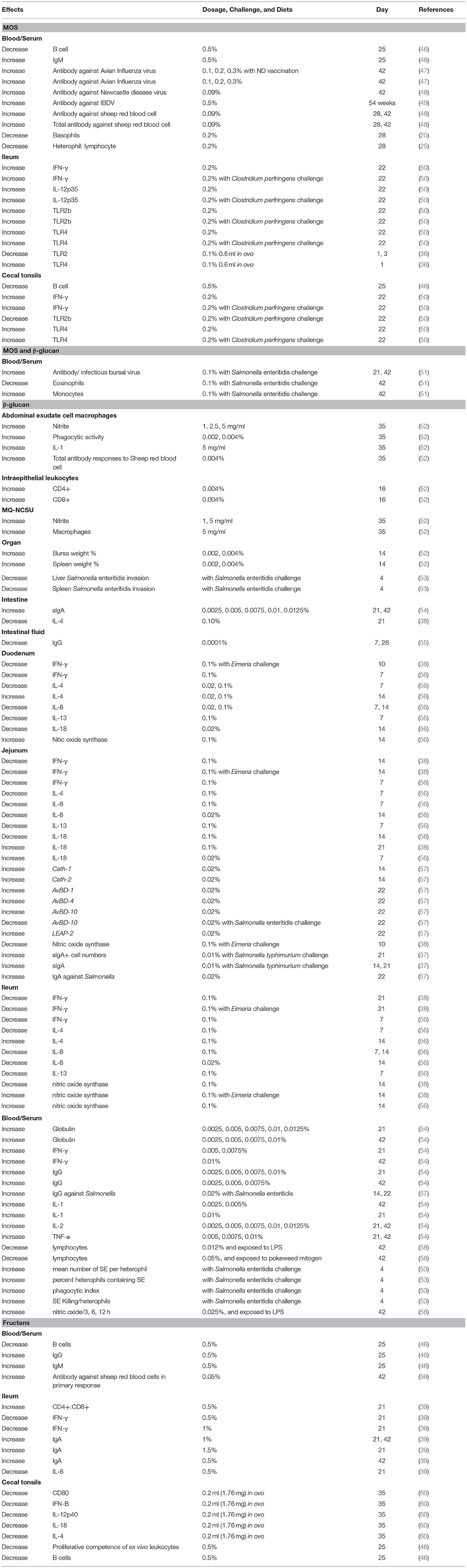
Table 3. Effects of mannan oligosaccharides, β-glucan, and fructans on immune responses of broilers.
However, chicken TLR2 (chTLR2) has approximately 50% amino acid identity to mammal TLR2, which can recognize a broad variety of PAMPs, including lipoproteins, aribinomannan, and peptidoglycan fugal zymosan (61). TLR2 may recognize MOS as well, which leads to the pro-inflammatory cytokines' cascade (63). A previous study demonstrated that supplementation of 0.2% MOS in broiler diets enhances ileal gene expression of interleukin-12 (IL-12) and interferon-γ (IFN-γ) (50). Interleukin-12 is a cytokine that stimulates T-helper type-1 cells (Th1 cells) and triggers IFN-γ to induce proliferation and cytotoxicity of immune cells, such as T cells, natural killer (NK) cells, and macrophages (12). Apart from the upregulation of innate immunity, MOS can impact humoral immune responses by acting as adjuvant of vaccines to enhance antibody titers. Previous studies have shown that MOS can strengthen antibody titers against sheep red blood cells, infectious bursal disease virus, Newcastle disease virus, and avian influenza virus (47–49). On the contrary, some reports have noted that antibody titers against Newcastle disease virus and infectious bursal disease virus failed to increase in chickens with MOS supplementation (64, 65). This discrepancy among studies may be based on whether or not broilers are infected with pathogens or the variations in MOS sources and environmental conditions (51).
The effects of MOS on intestinal microbiota have been reported broadly. Most of the MOS additions can significantly improve microbial community composition. However, there has been limited research on the impacts of MOS on mechanisms of immune responses in broilers. Although previous studies have found some auspicious results, further research is necessary to determine further antibody titers and gene expression of TLR or cytokines in order to elucidate how MOS improves the broiler's immunity.
β-glucan
β-glucan is a prebiotic derived from yeast or fungal cell walls. This long-chain polysaccharide is composed of D-glucose monomers with linkages of β-glycosidic 1-3 bonds, and its side-chains are linked by the 1–6 bonds. β-glucan can be recognized by receptors on sentinel cells, triggering production of cytokines and proliferation of lymphocytes (66). Lymphocytes are classified into three major types. The first type is NK cells, which play an important role in innate immunity. The second type is T cells, which regulate adaptive immunity. The third type is B cells, which produce antibodies against antigens. All types of lymphocytes can be modulated by β-glucan. The influences on immune responses of broilers are shown in Table 3.
Macrophages may be one of the sentinel cells that recognize β-glucan in the animal intestine. When macrophages are activated by β-glucan, they produce inducible nitric oxide synthase (iNOS) (56), an enzyme that produces large amounts of nitric oxide. Reacting with superoxide anion, nitric oxide is oxidized to a highly-toxic nitrogen dioxide radical that can kill a wide range of invading pathogens directly or block their DNA synthesis (12, 52, 58). Moreover, β-glucan exposure also triggers macrophage proliferation, enhances macrophage phagocytic ability, and induces macrophage-modulating gene expression of interleukin-1 (IL-1), interleukin-18 (IL-18), and tumor necrosis factor-α (TNF-α) (38, 52). Increasing TNF-α in birds fed with β-glucan may stimulate the incidence of CD8+ lymphocyte, a receptor expressed only on the cytotoxic T cell (Tc) (52, 54). Thus, it is hypothesized that β-glucans can regulate innate immune response by inducing proliferation of Tc cells to attack pathogen-infected cells.
Heterophils, recruited by sentinel cells, are the major granulocytes in most birds and work in a manner similar to neutrophils in mammals. Lowry et al. (53) showed the increases of heterophil phagocytosis in broilers fed with β-glucan, including enhancing the percentage of heterophils containing Salmonella enterica, mean numbers of Salmonella enterica per heterophil, and phagocytic index. One reasonable explanation that has been proposed is that the dectin-1 receptor involved in β-glucan recognition on the surface of macrophages may also be present on the surface of heterophils (67). Furthermore, heterophils stimulated by β-glucan can release nitric oxide and kill Salmonella enterica, resulting in the reduction of pathogenic organ invasion (53). Apart from heterophils, β-glucan receptors are also present on NK cells in humans (68). Therefore, activating NK cells by β-glucan may be another way to improve immune responses in broilers. On the contrary, Cox et al. (56) indicated that β-glucan could be an anti-inflammatory immunomodulator inhibiting interleukin-8 (IL-8) gene expression. Interleukin-8 is a cytokine produced by macrophages, which can recruit heterophiles to phagocytose pathogens at the site of inflammation (12). The inconsistent results may be attributed to whether or not the birds were challenged by pathogens. In a pathogen-challenging situation, pro-inflammatory immune responses may be enhanced by β-glucan supplementation, whereas in normal circumstances, β-glucan may be an anti-inflammatory modulator.
It was reported that the inclusion of β-glucan in diets could regulate the gene expression of antimicrobial peptides (AMPs) (57). Cathelicidins (Cath), avian β-defensins (AvBDs), and liver-expressed antimicrobial peptides (LEAP) are three major families of AMPs, which are expressed by the lung, intestine, immune, and reproductive organs in chickens (57). Antimicrobial peptides can penetrate the membrane of fungi or bacteria, leading to the death of pathogens. Among AMPs, Cath-1 and Cath-2 proteins have been shown to posses the capacity to bind to LPS, inhibiting LPS-mediated pro-inflammatory immune responses (61). On the other hand, AvBDs expressed in heterophils and the mucosal surface of the intestinal and respiratory tracts can damage pathogens, like Staphyloccocus aureas, E. coli, Candida albicans, S. Enteritidis, S. Typhimurium, Listeria monocytogenes, and Campylobacter jejuni (61). Shao et al. (57) reported that the gene expression of Cath-1, Cath-2, AvBD-1, AvBD-2, AvBD-4, AvBD-6, AvBD-9, and LEAP-2 were increased in Salmonella-challenged broilers with β-glucan addition. On the contrary, the same study showed that β-glucan reduced Cath-1, AvBD-4, and AvBD-9 in the spleen of birds without pathogen challenge. It could be concluded that if broilers were under pathogen infection, β-glucan would exhibit a strong protection against Salmonella and other pathogens in broilers.
After recognizing β-glucan, sentinel cells secrete cytokines that activate Th1 or Th2 cells. The Th1 cells drive the type-1 pathway attack against intracellular pathogens, whereas Th2 cells dominate the type-2 pathway triggering humoral immunity to upregulate antibody production (69). Although Th1 and Th2 cells could release cytokines to cross-inhibit each other, type-1 and type-2 pathways could both be triggered by β-glucan. In type-1 pathways, interleukin-12 (IL-12), produced by macrophages, is a key cytokine that enhances the proliferation of Th1 cells and the production of IFN-γ (12). Interferon-γ further reinforces with IL-18 in order to trigger the activation of Th1 cells and produce additional IFN-γ and IL-2 for the activation of NK cells, stimulation of macrophages and Tc cells, and inhibition against Th2 cells (12). Previous studies reported that β-glucan upregulates the gene expression of IL-2, IL-18, and IFN-γ (52, 54). Additionally, levels of the cytokines interleukin-4 (IL-4) and interleukin-13 involved in type-2 cell pathways are downregulated by β-glucan as well (56). These outcomes support the hypothesis that β-glucan can stimulate the type-1 pathway and inhibit the type-2 pathway.
However, gene expression of IL-1 involved in the type-2 pathway could also be induced by β-glucan (52). Increasing IL-1 found in abdominal exudate cell macrophages can activate Th2 cells and switch on the type-2 pathway. Once activated, Th2 cells release other cytokines to initiate the subsequent anti-inflammatory immune responses. For instance, IL-4 can suppress Th1 cells' activation, stimulate B cells' growth and differentiation, and activate mast cells to produce immunoglobulins (12). Owing to the suppression of Th1 cells, gene expression of IFN-γ was downregulated in duodenum, jejunum, and ileumthe duodenum, the jejunum, and the ileum by β-glucan in Eimeria-challenged broilers (38). On the other hand, enhancing immunoglobulins, including IgG and sIgA, in broilers were found by Zhang et al. (54). This is evidence showing that the type-2 pathway can be upregulated by β-glucan. Shao et al. (57) also reported that anti-Salmonella specific IgA levels in the jejunum and anti-Salmonella specific IgG levels in the serum were increased in birds fed with β-glucan. Similarly, Shao et al. (37) demonstrated that β-glucan could protect intestinal barrier function in Salmonella-challenged birds by increasing the amount of goblet cells and IgA-secreting cells, which enhance the sIgA production. sIgA is an important immunoglobulin that serves as the first line of defense (70). There are three major mechanisms of sIgA to protect the integrity of gut lining from pathogenic invasion (71). Firstly, sIgA interacts with non-pathogenic bacteria and epithelium, which consequently strengthens the tight junctions between intestinal epithelial cells and inhibits nuclear translocation of NF-κB (70). A previous study also confirmed that β-glucan enhanced the production of sIgA to ameliorate the damage of tight junction in the jejunum caused by Salmonella (37). Secondly, immune complexes that interact with sIgA are involved in the downregulation of gene expression of pro-inflammatory cytokines that include IFN-γ, TNF-α, and interleukin-6 (IL-6) (70). Thirdly, sIgA blocks pathogens within mucin, selecting and maintaining a favorable balance of microbiota in the intestine (70). Shao et al. (37) showed that increased sIgA by β-glucan was associated with the reduction of cecal Salmonella colonization and liver invasion.
In summary, β-glucan affects the broiler's immunity via either the type-1 or the type-2 pathway. The conflicting results among different studies may be attributed to the different dosages offered, different ages of the birds used, different parts of the tissue examined, or numerous resources of the β-glucan supplemented. Inconsistent results have also been demonstrated in other animals. For example, cytokines involved in the type-2 pathway of immune responses were downregulated by β-glucan in humans (72) but upregulated in mice (73). Therefore, additional investigation is needed to understand fully the effects of β-glucan on immune responses of broilers.
Fructans
Fructans, commonly extracted from different plants, hydrolyzed from polysaccharides, or produced by microorganism, have been administered recently in broiler diets. Fructans are classified into three distinct types: the inulin group, the levan group, and the branched group. Firstly, the inulin group, also known as fructooligosaccharides (FOS) can be divided into different categories based on degrees of polymerization (DP): Inulin, normally extracted from chicory roots (Cichorium intybus L.), consists of a DP of 3 to 60, and Oligofructose (OF), which can be generated by partial hydrolysis of inulin, enzymatic conversion of sucrose, or lactose, contains a DP of 2 to 10 (74, 75). Most of the inulin group can be found in plants, which comprise oligosaccharides with β-2,1 fructosyl-fructose linkage with a glucose terminal unit. Secondly, the levan group is another group of fructans, which are mostly linked by β-2,6 fructosyl-fructose bonds. Lastly, fructans, which belong to the branched group, contain both β-2,1 fructosyl-fructose and β-2,6 fructosyl-fructose bonds in fair amounts (76). It is the β-glycosidic bond in fructans that resists their breakdown by digestive enzymes in poultry and enhances the population of beneficial bacteria, such as Bifidobacteria and Lactobacilli, and suppresses levels of pathogenic bacteria, such as Clostridium pefringens and E. coli, in the intestine of broilers (25, 40, 77).
Saminathan et al. ( 78) evaluated the utilization of different oligosaccharides by 11 Lactobacillus species isolated from the gastrointestinal tract of chickens. This in vitro report showed that FOS were utilized by Lactobacillus more efficiently than MOS. The high availability of FOS may be associated with specific enzymatic activity and the oligosaccharide transport system of Lactobacillus species (79, 80). However, the intestinal microbiota of a broiler is far more complex than those in in vitro trials. The prebiotics may be fermented not only by Lactobacillus species but also by other microorganisms in the gastrointestinal tracts of animals. Thus, it cannot be assured that the utilization of FOS and MOS in in vitro trials is as efficient as in in vivo studies.
In addition, the more DP increased, the more residual FOS remained after fermentation by Bifidobacteria (81). A previous study indicated that almost 55 Bifidobacteria preferred to grow on short-chain FOS rather than long-chain FOS (75). Bifidobacteria could also ferment short-chain FOS to produce more acetic acid and lactic acid compared with long-chain FOS within 24 h (81). Similarly, Perrin et al. (82) reported that the population of Bifidobacteria and Lactobacilli increased earlier in fecal cultures containing OF instead of inulin. However, an increase in the production of formic acid, acetic acid, and lactic acid and a decrease in numbers of E. coli group and Cluster I clostridia were both observed in cultures containing OF or inulin after 24-h fermentation (82). The same research group also pointed out that butyric acid might be the major product in the inulin group, whereas more acetic acid and lactate acid could be produced from OF (75).
Long-chain fructans, which are degraded slowly in the animal gut, can pass through the small intestine and be fermented in the distal regions of the intestine. Therefore, the inulin group with higher DP might not affect the microbiota in the jejunum significantly (83), but, instead it might alter microbial structure and increase the concentration of SCFA or lactic acid in the ceca of broilers. Effects of FOS on intestinal microbiota are shown in Table 4. Park et al. (85) demonstrated that FOS increased the Shannon diversity of intestinal microbiome compared with the control treatment. Moreover, similar to in vitro results, Bifidobacteria and Lactobacillus are two major beneficial bacteria that were increased in broilers and hens fed with fructans (40, 41, 76, 84, 88). Bifidobacteria and Lactobacillus not only produced extracellular enzymes to degrade FOS but also competed with other species of intestinal microorganisms and suppressed the growth of pathogenic bacteria (75). For instance, Campylobacter titers in the ceca and large intestine were decreased in broilers fed with FOS (84). Regardless of the supplementation of long-chain FOS or short-chain FOS, a reduction in titers of C. perfringens was observed in the ileocecal junction or ceca of broilers (20, 25, 76). Similarly, colonization of cecal C. perfringens and Salmonella typhimurium was decreased by FOS or FOS combined with competitive exclusion products in E. coli or Salmonella- Typhimurium-challenged birds, respectively (10, 87). Additionally, diets containing different concentrations of FOS (from 0.25 to 1%) could decrease cecal E. coli and Salmonella in broilers (25, 40, 76, 84, 86). Besides the prevention of Salmonella colonization in the ceca of broilers, previous reports also demonstrated that FOS-supplemented diets decreased ovary, liver, and cecal Salmonella enteritidis in laying hens (89, 90). The reduction of these pathogenic bacteria might be attributed to cecal SCFA and lactic acid. Same as in vitro results, the concentration of cecal butyric acid and lactic acid was significantly higher in broilers fed with inulin (41, 83). Donalson et al. (89) also showed that 0.75 or 0.375% of FOS combined with alfalfa molt diets could increase the concentration of cecal isobutyric acid in hens. Short-chain fatty acids are important fuels in the intestine, and butyrate is the major one that is metabolized by epithelial cells, providing energy for the growth of mucosal epithelium (91). It is suggested that higher concentrations of butyric acid are associated with the improvement of mucosal structure. Previous studies reported that microvillus height in the jejunum and ileum and the ratio of villus to crypt depth in the ceca were increased by FOS (40, 41). Bogucka et al. (92) also reported that in ovo injection of inulin increased villus height in broilers at the first day after hatching. In addition, the use of inulin could increase jejunal mucin mRNA expression to produce more mucin, protecting intestinal epithelial cells in broilers (39). By improving intestinal morphology, FOS could further enhance activities of protease and amylase and nutrient absorption, leading to better growth performance (40).
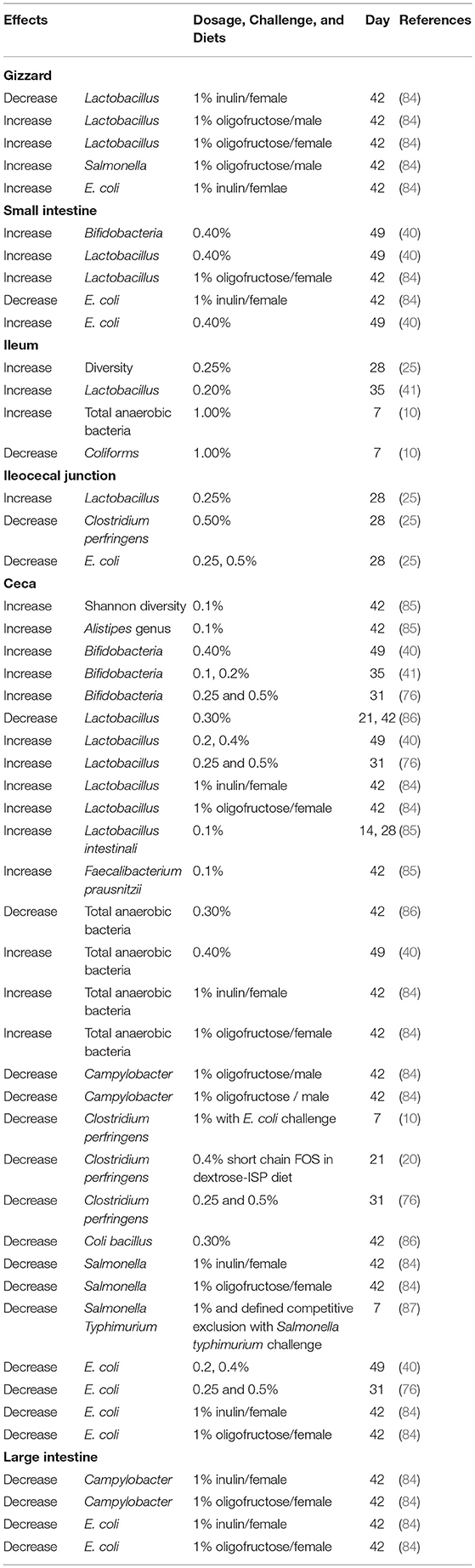
Table 4. Effects of fructans on intestinal microbiota of broilers.
However, adding high levels of fructans could result in negative impacts on broilers. Rapid fermentation by microbes in the intestine could produce too much SCFA, which damage intestinal mucosal barriers and increase intestinal permeability, consequently causing pathogen invasion, diarrhea, and poor growth performance (93, 94). Xu et al. (40) demonstrated that the addition of 0.2 or 0.4% of FOS in broiler diets could improve FCR and change cecal microbiota, but the supplementation of 0.8% of FOS had no significant differences compared with control treatment. It has been suggested that the supplementation of FOS above 0.5% is excessive; a previous report mentioned that birds fed with 0.5% FOS showed poorer growth performance and less intestinal Lactobacillus but higher titers of E. coli and C. perfringens compared with 0.25% FOS treatment (25). Furthermore, Biggs et al. (20) even showed that MEn and amino acid digestibility were reduced by 8% short-chain FOS or inulin addition.
Fructans improved the immune responses of gut-associated lymphoid tissue (GALT) and the systemic immune system through three major mechanisms. Firstly, increasing the levels of Bifidobacteria by fructans could modulate the production of cytokines or antibodies. Secondly, leukocytes could be activated after their receptors respond to fructans' metabolites, such as SCFA. Thirdly, fructans could be directly recognized by carbohydrate receptors on the surface of immune cells (95). Huang et al. (39) reported that inulin reduced the levels of IL-6 and IFN-γ, increased IgA, and tended to increase the ratio of CD4+/CD8+ cells in the ileum of broilers. Moreover, Janardhana et al. (46) found that FOS could lead to systemic immune responses by increasing the levels of plasma antibody titers of IgG and IgM. Similarly, primary antibody titers against sheep red blood cells increased in broilers fed with FOS, but antibody titers in the secondary immune response were not influenced by FOS (59). Likewise, FOS increased IgA+ cells and upregulated TLR-4 and IFN-γ in the ileum of laying hens (90). Interestingly, there is a hypothesis that fructans might modulate the development of the immune system during embryogenesis. In ovo administration of inulin (d 12) downregulated the gene expression of IL-4, IL-12p40, IL-18, CD80, and interferon-β in the cecal tonsils of broilers on day 35 after hatching (60). Furthermore, in ovo injection of inulin had no adverse effect on GALT development but stimulated more colonization of lymphoid tissue by T cells in the cecal tonsil of broilers (96). To our knowledge, there are only a few studies that evaluated the in ovo administration of prebiotics. Further research is needed to understand what causes the different results between in ovo administration and direct-fed supplementation of fructans in broilers. It could be concluded that owing to the various fructans groups and DP, supplementation of fructans in diets might have affected broilers inconsistently. However, in a general review, fructans could modulate intestinal microorganisms, levels of intestinal SCFA, mucosal morphology, and generate immune responses.
Other Prebiotics
Besides the three major prebiotics, MOS, β-glucan, and fructans, other oligosaccharides have been evaluated and considered as potential prebiotics, including chitosan oligosaccharides (COS), galacto-oligosaccharides (GOS), galactoglucomannan oligosaccharide (GGMO), and xylo-oligosaccharides (XOS).
Chitosan Oligosaccharides (COS)
Extracted from chitin, COS contain 2–10 sugar units of N-acetyl glucosamine with 1–4 β-linkages. It has been reported that the supplementation of COS in broiler diets could modulate immune responses and enhance nutrient digestibility and feed efficiency. Huang et al. (97) indicated that chicken with COS supplementation had higher weight of bursa of Fabricius and thymus, higher IgG, IgA, and IgM in serum and higher antibody titers against Newcastle disease vaccines. On the other hand, 0.01% of COS improved ileal digestibility of dry matter, energy, crude protein, and most of the amino acids in broilers (21 or 42 d) (98). The improved digestibility of nutrients was associated with better growth performance in the same study (98). However, supplementation of COS above 0.01% might be excessive because chickens fed with 0.015% COS had significantly less body weight than birds fed with 0.01% COS (98).
Galacto-Oligosaccharides (GOS)
Galacto-oligosaccharides, synthetic prebiotics with galactose with 1–4 or 1–6 β-linkages, are normally produced from lactose by the enzyme lactase with high galactosyltransferase activity (99). In ovo injection of GOS could increase body weight of broilers 34 days after hatching (100). Administration of GOS also influenced the intestinal microbiota. Park et al. (85) reported that GOS treatment exhibited higher levels of Alistipes genus, Lactobacillus intestinalis, and Faecalibacterium prausnitzii in the ceca of broilers compared with the control group. Although Biggs et al. (20) demonstrated that GOS had no effects on cecal Bifidobacteria and Lactobacillus population, it has been reported that the addition of GOS in broiler diets could increase counts of Bifidobacteria in feces (101). Moreover, broilers that received in ovo GOS injection also had higher concentrations of Bifidobacteria and Lactobacillus in feces (102). The author suggested that in ovo administration of GOS could replace prolonged water supplementation. Owing to the inconsistent results, future studies are needed to confirm the effects of GOS in modulating intestinal microbial structures and further affecting immune responses in broilers.
Galactoglucomannan Oligosaccharides (GGMO) and Galactoglucomannan Oligosaccharides-Arabinoxylan (GGMO-AX)
Galactoglucomannan oligosaccharides and galactoglucomannan oligosaccharides-arabinoxylan (GGMO-AX) are novel prebiotics extracted and processed from the wood chips of softwood trees (103). These oligosaccharides consist of mannose, glucose, and galactose monomers. An in vitro investigation showed that Lactobacillus could grow faster on GGMO than MOS (35). The same research also indicated that the supplementation of 0.2% GGMO in broiler diets could reduce colonization of Salmonella typhimurium in the ileum, ceca, and liver; as a consequence of clearing S. typhimurium infection, GGMO ameliorates intestinal morphology and growth performance compared with a Salmonella-challenged control treatment (35). The improvement might be attributed to the modulation of immune responses by GGMO. Faber et al. (104) reported that the Eimeria acervulina-challenged birds that received 4% GGMO-AX showed enhanced gene expression of pro-inflammatory cytokines, including IFN-γ, IL-1β, IL-6, and IL-12β, but also showed decreased levels of anti-inflammatory cytokines such as interleukin-15. Galactoglucomannan oligosaccharides-arabinoxylan might not only affect immune responses in broilers but also alter intestinal microbial population. It has been shown that the administration of 2% GGMO-AX increased counts of Bifidobactrium spp. in the ceca (104) and 4% GGMO-AX decreased the concentration of C. perfringens (105). Although the supplementation of GGMO-AX in high levels showed some positive effects on broilers, simultaneously, it could lead to poor growth performance (104). Therefore, further studies should evaluate the administration of GGMO or GGMO-AX in appropriate concentration to maintain growth performance and improve the health status of broilers at the same time.
Xylo-Oligosaccharides (XOS)
Xylo-oligosaccharides are oligosaccharides, which consist of xylose sugar units with β-linkages (42). Xylan, the main component of cereal fiber such as corn cobs, straws, hulls, and bran are the raw resources for XOS production (106). Xylan could be degraded to XOS by xylanase of fungi, steam, or diluted solutions of mineral acid (106). Similar to other prebiotics, XOS could improve growth performance, increase the intestinal villus height, increase the proportion of Lactobacillus, and enhance the levels of acetate, butyrate, and lactate in the ceca of broilers (42, 107, 108). It was suggested that XOS would improve humoral immunity in poultry. An increase in antibody titers against avian influenza H5N1 was observed in broilers by XOS addition (107). Furthermore, De Maesschalck et al. (42) speculated that XOS could lead to cross-feeding mechanisms between L. crispatus and Anaerostipes butyraticus in the gut of the broiler. Owing to XOS fermentation, L. crispatus produces lactate, which might be utilized by butyrate-producing bacteria that belong to members of Clostridium cluster XIVa. This hypothesis was further supported by the observation of increasing numbers of cecal Clostridium cluster XIVa and butyryl-CoA: acetate-CoA transferase, a marker indicating the butyrate-producing capacity of intestinal microbiota (42). As mentioned above, butyrate is a major energy source for intestinal epithelial cells. Apart from acting as an important fuel in the intestine, butyrate can stimulate MUC-2 gene expression, exert anti-inflammatory effects, and prevent necrotic enteritis from pathogenic infection (109–111). In summary, XOS supplementation would enhance cross-feeding mechanisms and produce butyrate, consequently leading to beneficial influences on broilers.
In Ovo Injection
Direct feeding and in ovo injection are two main strategies for applying prebiotics. Prebiotic can be administrated by injecting 0.2 ml aqueous solution into the air chamber of eggs on day 12 of embryonic incubation (112). In ovo injection of prebiotics can alter microbial community in embryonic guts, improve intestinal morphology, and directly promote robustness of both cellular and humoral immune responses in the GALTs of the neonate post hatching (96, 113, 114).
The embryonic microbiota is different from the intestinal microbiota of post hatching and adult birds. The dominant bacterial phylum is Proteobacteria, followed by Firmicutes, Bacteroidetes, and Actinobacteria in the chicken embryos (115). In addition, the embryonic microbial community is altered during the development of the embryos. The 19-day-old embryos exhibited more microbial diversity than the 4-day-old embryos. The proportion of Proteobacteria decreased, whereas Firmicutes, Bacteroidetes, and Actinobacteria increased in the 19-day-old embryos compared with the 4-day-old embryos (115). Even though Proteobacteria decreased in the late embryonic development period, this phylum dominated in early-age birds until Firmicutes became prominent after 7 days post hatching (116). However, the embryonic microbiota could be contaminated by pathogens directly from the yolk, yolk membranes, albumen, shell membranes originating from the reproductive organs of laying hens, or indirectly from the egg shells. Pathogens such as Salmonella located in the albumen were able to migrate and penetrate the vitelline membrane and grow in the yolk (117). On the other hand, it was suggested that spore forming bacteria such as Clostridium tertium were capable of surviving the disinfection process and penetrating eggs, resulting in contamination (118). To avoid extensive pathogen infection, prebiotics were delivered in ovo, which is likely fermented by the indigenous embryonic microbiota, inhibiting pathogen proliferation and regulating gene expression of immune responses (119). Villaluenga et al. (120) reported that injection of raffinose at day 12 of embryonic incubation had the highest amounts of Bifidobacteria in the ceca of 2 day-old broilers. Additionally, they indicated that 8.815 mg per egg of raffinose delivered in ovo reduced embryo weight. A later research showed that 4.5 mg of raffinose that was delivered in ovo had no significant effects on body weight but enhanced gene expression of CD3 and ChB6, which are associated with the activity of T cells and B cells (114). Moreover, villus height and villus height to crypt depth ratio of post hatching birds increased linearly with higher dosages of raffinose (114). In ovo injection of inulin and GOS also increased villus height in the jejunum of 1-day-old chickens (92). Moreover, administration of GOS in ovo showed differential gene expression in the ceca related to lymphocyte proliferation, activation, and differentiation and cytokine production (119). This study pointed out that GZMA (Granzyme A), a cytotoxic T cell-specific gene, was upregulated in the cecal tonsil of birds delivered with GOS in ovo. Similarly, other research has also demonstrated that GOS increased helper T cells in the cecal tonsil and B cells in the bursa of Fabricius (96). Furthermore, beta inhibin and lectin galactoside-binding soluble 3, which are related to regulation of T cell and innate immunity, were upregulated by GOS. On the other hand, GOS also downregulated the SERPING1 gene, which could inhibit part of the complement cascade system (119). It was suggested that the in ovo injection of GOS might not only regulate intestinal innate and adaptive immune system but also modulate gene expression of nutrient digestion and transportation. Firstly, chicken injected with GOS in ovo exhibited higher levels of sodium-dependent glucose co-transporters in the intestine, which are related to the absorption of monosaccharides (119). Secondly, birds delivered with GOS in ovo showed increased amylase and trypsin activity of the pancreas on embryonic day 21 and day 7 post hatching respectively (100). These studies led us to a conclusion that in ovo injection of prebiotics could affect the ecosystem of broilers, but, to our knowledge, little research has compared the difference between the direct-fed method and in ovo injection. A study reported that injection of galacto-oligosaccharides into eggs could increase Bifidobacteria and Lactobacillus in the feces of broilers. Though the author suggested that in ovo injection could replace prolonged supplementation via water system (102), more studies are needed to compare these two different approaches on the application of prebiotics.
Conclusion
The interaction between epithelium, microbiota, and immunity in animal gut is complicated. Recent data have demonstrated that prebiotics potentially alter the interaction between the host and gut microbiota and improve the health status of broilers. However, the interaction is sometimes induced by certain prebiotics or host species. Therefore, it is inevitable that prebiotics showed variable effects on animals. Still, most prebiotics can be fermented by beneficial bacteria, and the increased levels of Lactobacillus and Bifidobacteria or their metabolites may inhibit pathogen colonization and communicate with epithelial cells and immune cells. By improving gut environment or immune responses, prebiotics further provide resistance to pathogens and maintain efficient production. In addition, some prebiotics can be recognized by sentinel cells directly, triggering cytokines' cascade, which results in the upregulation of innate or humoral immunity. Although previous studies have discovered some mechanisms that participate in the cross talk between prebiotics and the ecosystem of the gut, there are still several hypotheses, which shall be confirmed in the future. In this context, administration of prebiotics presents tremendous influences on the broilers' gut health by the modulation of the gut microbial community and the interaction between the host immune system and gut microbiota. It is suggested that prebiotics delivered in ovo or fed directly can act as alternatives to antibiotics because of the significant improvement of microbial community, intestinal integrity, and immunity of the host.
Author Contributions
P-YT reviewed papers related to the topics and wrote the manuscript. WK reviewed papers related to the topic, gave directions and ideas to P-YT, and reviewed and revised the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Castanon JI. History of the use of antibiotic as growth promoters in European poultry feeds. Poult Sci. (2007) 86:2466–71. doi: 10.3382/ps.2007-00249
2. Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr. (1995) 125:1401–12.
3. Gibson GR, Probert HM, Loo JV, Rastall RA, Roberfroid MB. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr Res Rev. (2004) 17:259–75. doi: 10.1079/NRR200479
4. Kolida S, Tuohy K, Gibson GR. Prebiotic effects of inulin and oligofructose. Br J Nutr. (2002) 87:S193. doi: 10.1079/BJN/2002537
5. Patterson JA, Burkholder KM. Application of prebiotics and probiotics in poultry production. Poult Sci. (2003) 82:627–31. doi: 10.1093/ps/82.4.627
6. Lavelle EC, Murphy C, O'Neill LA, Creagh EM. The role of TLRs, NLRs, and RLRs in mucosal innate immunity and homeostasis. Mucosal Immunol. (2010) 3:17–28. doi: 10.1038/mi.2009.124
7. BogusŁAwska-Tryk M, Piotrowska A, Burlikowska K. Dietary fructans and their potential beneficial influence on health and performance parametrs in broiler chickens. J Cent Eur Agric. (2012) 13:270–88. doi: 10.5513/JCEA01/13.2.1045
8. Lan Y, Verstegen MWA, Tamminga S, Williams BA. The role of the commensal gut microbial community in broiler chickens. Worlds Poult Sci J. (2005) 61:95–104. doi: 10.1079/WPS200445
9. Pourabedin M, Zhao X. Prebiotics and gut microbiota in chickens. FEMS Microbiol Lett. (2015) 362:fnv122. doi: 10.1093/femsle/fnv122
10. Yang Y, Iji PA, Kocher A, Mikkelsen LL, Choct M. Effects of mannanoligosaccharide and fructooligosaccharide on the response of broilers to pathogenic Escherichia coli challenge. Br Poult Sci. (2008) 49:550–9. doi: 10.1080/00071660802290408
11. Chee SH, Iji PA, Choct M, Mikkelsen LL, Kocher A. Characterisation and response of intestinal microflora and mucins to manno-oligosaccharide and antibiotic supplementation in broiler chickens. Br Poult Sci. (2010) 51:368–80. doi: 10.1080/00071668.2010.503477
13. Kogut MH. The gut microbiota and host innate immunity: regulators of host metabolism and metabolic diseases in poultry? J Appl Poult Res. (2013) 22:637–46. doi: 10.3382/japr.2013-00741
14. Baurhoo B, Ferket PR, Zhao X. Effects of diets containing different concentrations of mannanoligosaccharide or antibiotics on growth performance, intestinal development, cecal and litter microbial populations, and carcass parameters of broilers. Poult Sci. (2009) 88:2262–72. doi: 10.3382/ps.2008-00562
15. Yang Y, Iji PA, Choct M. Dietary modulation of gut microflora in broiler chickens: a review of the role of six kinds of alternatives to in-feed antibiotics. Worlds Poult Sci J. (2009) 65:97. doi: 10.1017/S0043933909000087
16. Spring P, Wenk C, Dawson KA, Newman KE. The effects of dietary mannanoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of Salmonella-challenged broiler chicks. Poult Sci. (2000) 79:205–11. doi: 10.1093/ps/79.2.205
17. Ofek I, Beachey EH. Mannose binding and epithelial cell adherence of Escherichia coli. Infect Immun. (1978) 22:247–54.
18. Fernandez F, Hinton M, Van Gils B. Dietary mannan-oligosaccharides and their effect on chicken caecal microflora in relation to Salmonella Enteritidis colonization. Avian Pathol. (2002) 31:49–58. doi: 10.1080/03079450120106000
19. Baurhoo B, Phillip L, Ruiz-Feria CA. Effects of purified lignin and mannan oligosaccharides on intestinal integrity and microbial populations in the ceca and litter of broiler chickens. Poult Sci. (2007) 86:1070–8. doi: 10.1093/ps/86.6.1070
20. Biggs P, Parsons CM, Fahey GC. The effects of several oligosaccharides on growth performance, nutrient digestibilities, and cecal microbial populations in young chicks. Poult Sci. (2007) 86:2327–36. doi: 10.3382/ps.2007-00427
21. Yang Y, Iji PA, Kocher A, Mikkelsen LL, Choct M. Effects of dietary mannanoligosaccharide on growth performance, nutrient digestibility and gut development of broilers given different cereal-based diets. J Anim Physiol Anim Nutr. (2008) 92:650–9. doi: 10.1111/j.1439-0396.2007.00761.x
22. Yang Y, Iji PA, Kocher A, Thomson E, Mikkelsen LL, Choct M. Effects of mannanoligosaccharide in broiler chicken diets on growth performance, energy utilisation, nutrient digestibility and intestinal microflora. Br Poult Sci. (2008) 49:186–94. doi: 10.1080/00071660801998613
23. Corrigan A, de Leeuw M, Penaud-Frezet S, Dimova D, Murphy RA. Phylogenetic and functional alterations in bacterial community compositions in broiler ceca as a result of mannan oligosaccharide supplementation. Appl Environ Microbiol. (2015) 81:3460–70. doi: 10.1128/AEM.04194-14
24. Geier MS, Torok VA, Allison GE, Ophel-Keller K, Hughes RJ. Indigestible carbohydrates alter the intestinal microbiota but do not influence the performance of broiler chickens. J Appl Microbiol. (2009) 106:1540–8. doi: 10.1111/j.1365-2672.2008.04116.x
25. Kim GB, Seo YM, Kim CH, Paik IK. Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers. Poult Sci. (2011) 90:75–82. doi: 10.3382/ps.2010-00732
26. Wang X, Farnell YZ, Peebles ED, Kiess AS, Wamsley KG, Zhai W. Effects of prebiotics, probiotics, and their combination on growth performance, small intestine morphology, and resident Lactobacillus of male broilers. Poult Sci. (2016) 95:1332–40. doi: 10.3382/ps/pew030
27. Lee SI, Park SH, Ricke SC. Assessment of cecal microbiota, integron occurrence, fermentation responses, and Salmonella frequency in conventionally raised broilers fed a commercial yeast-based prebiotic compound. Poult Sci. (2016) 95:144–53. doi: 10.3382/ps/pev322
28. Wexler HM. Bacteroides: the good, the bad, and the nitty-gritty. Clin Microbiol Rev. (2007) 20:593–621. doi: 10.1128/CMR.00008-07
29. Corrigan A, Horgan K, Clipson N, Murphy RA. Effect of dietary supplementation with a Saccharomyces cerevisiae mannan oligosaccharide on the bacterial community structure of broiler cecal contents. Appl Environ Microbiol. (2011) 77:6653–62. doi: 10.1128/AEM.05028-11
30. Chen X, Xu J, Shuai J, Chen J, Zhang Z, Fang W. The S-layer proteins of Lactobacillus crispatus strain ZJ001 is responsible for competitive exclusion against Escherichia coli O157:H7 and Salmonella typhimurium. Int J Food Microbiol. (2007) 115:307–12. doi: 10.1016/j.ijfoodmicro.2006.11.007
31. Zhang G, Ma L, Doyle MP. Salmonellae reduction in poultry by competitive exclusion bacteria Lactobacillus salivarius and Streptococcus cristatus. J Food Prot. (2007) 70:874–8. doi: 10.4315/0362-028X-70.4.874
32. Mack DR, Michail S, Wei S, McDougall L, Hollingsworth MA. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression Am J Physiol. (1999) 276:941–50.
33. Gotteland M, Cruchet S, Verbeke S. Effect of Lactobacillus ingestion on the gastrointestinal mucosal barrier alterations induced by indometacin in humans. Aliment Pharmacol Ther. (2001) 15:11–7. doi: 10.1046/j.1365-2036.2001.00898.x
34. Pourabedin M, Xu Z, Baurhoo B, Chevaux E, Zhao X. Effects of mannan oligosaccharide and virginiamycin on the cecal microbial community and intestinal morphology of chickens raised under suboptimal conditions. Can J Microbiol. (2014) 60:255–66. doi: 10.1139/cjm-2013-0899
35. Rajani J, Dastar B, Samadi F, Karimi Torshizi MA, Abdulkhani A, Esfandyarpour S. Effect of extracted galactoglucomannan oligosaccharides from pine wood (Pinus brutia) on Salmonella typhimurium colonisation, growth performance and intestinal morphology in broiler chicks. Br Poult Sci. (2016) 57:682–92. doi: 10.1080/00071668.2016.1200013
36. Cheled-Shoval SL, Amit-Romach E, Barbakov M, Uni Z. The effect of in ovo administration of mannan oligosaccharide on small intestine development during the pre- and post hatch periods in chickens. Poult Sci. (2011) 90:2301–10. doi: 10.3382/ps.2011-01488
37. Shao Y, Guo Y, Wang Z. beta-1,3/1,6-Glucan alleviated intestinal mucosal barrier impairment of broiler chickens challenged with Salmonella enterica serovar Typhimurium. Poult Sci. (2013) 92:1764–73. doi: 10.3382/ps.2013-03029
38. Cox CM, Sumners LH, Kim S, McElroy AP, Bedford MR, Dalloul RA. Immune responses to dietary beta-glucan in broiler chicks during an Eimeria challenge. Poult Sci. (2010) 89:2597–607. doi: 10.3382/ps.2010-00987
39. Huang Q, Wei Y, Lv Y, Wang Y, Hu T. Effect of dietary inulin supplements on growth performance and intestinal immunological parameters of broiler chickens. Livest Sci. (2015) 180:172–6. doi: 10.1016/j.livsci.2015.07.015
40. Xu ZR, Hu CH, Xia MS, Zhan XA, Wang MQ. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult Sci. (2003) 82:1030–6. doi: 10.1093/ps/82.6.1030
41. Rebole A, Ortiz LT, Rodriguez ML, Alzueta C, Trevino J, Velasco S. Effects of inulin and enzyme complex, individually or in combination, on growth performance, intestinal microflora, cecal fermentation characteristics, and jejunal histomorphology in broiler chickens fed a wheat- and barley-based diet. Poult Sci. (2010) 89:276–86. doi: 10.3382/ps.2009-00336
42. De Maesschalck C, Eeckhaut V, Maertens L, De Lange L, Marchal L, Nezer C, et al. Effects of xylo-oligosaccharides on broiler chicken performance and microbiota. Appl Environ Microbiol. (2015) 81:5880–8. doi: 10.1128/AEM.01616-15
43. Robertson AM, Wright DP. Bacterial glycosulphatases and sulphomucin degradation. Can J Gastroenterol. (1997) 11:361–6. doi: 10.1155/1997/642360
44. Fontaine N, Meslin JC, Dore J. Selective in vitro degradation of the sialylated fraction of germ-free rat mucins by the caecal flora of the rat. Reprod Nutr Dev. (1998) 38:289–96. doi: 10.1051/rnd:19980309
45. Belley A, Göttke M, Keller K, Chadee K, Göettke M. Intestinal mucins in colonization and host defense against pathogens. Am J Trop Med Hyg. (1999) 60(4Suppl.):10–5.
46. Janardhana V, Broadway MM, Bruce MP, Lowenthal JW, Geier MS, Hughes RJ, et al. Prebiotics modulate immune responses in the gut-associated lymphoid tissue of chickens. J Nutr. (2009) 139:1404–9. doi: 10.3945/jn.109.105007
47. Tohid T, Hasan G, Alireza T. Efficacy of mannanoligosaccharides and humate on immune response to Avian Influenza (H9) disease vaccination in broiler chickens. Vet Res Commun. (2010) 34:709–17. doi: 10.1007/s11259-010-9444-8
48. Salehimanesh A, Mohammadi M, Roostaei-Ali Mehr M. Effect of dietary probiotic, prebiotic and synbiotic supplementation on performance, immune responses, intestinal morphology and bacterial populations in broilers. J Anim Physiol Anim Nutr. (2016) 100:694–700. doi: 10.1111/jpn.12431
49. Shashidhara RG, Devegowda G. Effect of dietary mannan oligosaccharide on broiler breeder production traits and immunity. Poult Sci. (2003) 82:1319–25. doi: 10.1093/ps/82.8.1319
50. Yitbarek A, Echeverry H, Brady J, Hernandez-Doria J, Camelo-Jaimes G, Sharif S, et al. Innate immune response to yeast-derived carbohydrates in broiler chickens fed organic diets and challenged with Clostridium perfringens. Poult Sci. (2012) 91:1105–12. doi: 10.3382/ps.2011-02109
51. Sadeghi AA, Mohammadi A, Shawrang P, Aminafshar M. Immune responses to dietary inclusion of prebiotic-based mannan-oligosaccharide and β-glucan in broiler chicks challenged with Salmonella enteritidis. Turk J Vet Anim Sci. (2013) 37:206–13. doi: 10.3906/vet-1203-9
52. Guo Y, Ali RA, Qureshi MA. The influence of beta-glucan on immune responses in broiler chicks. Immunopharmacol Immunotoxicol. (2003) 25:461–72. doi: 10.1081/IPH-120024513
53. Lowry VK, Farnell MB, Ferro PJ, Swaggerty CL, Bahl A, Kogut MH. Purified beta-glucan as an abiotic feed additive up-regulates the innate immune response in immature chickens against Salmonella enterica serovar Enteritidis. Int J Food Microbiol. (2005) 98:309–18. doi: 10.1016/j.ijfoodmicro.2004.06.008
54. Zhang B, Guo Y, Wang Z. The modulating effect of β-1, 3/1, 6-glucan supplementation in the diet on performance and immunological responses of broilercChickens. Asian Austral J Anim Sci. (2008) 21:237–44. doi: 10.5713/ajas.2008.70207
55. Revolledo L, Ferreira CSA, Ferreira AJP. Prevention of Salmonella Typhimurium colonization and organ invasion by combination treatment in broiler chicks. Poult Sci. (2009) 88:734–43. doi: 10.3382/ps.2008-00410
56. Cox CM, Stuard LH, Kim S, McElroy AP, Bedford MR, Dalloul RA. Performance and immune responses to dietary beta-glucan in broiler chicks. Poult Sci. (2010) 89:1924–33. doi: 10.3382/ps.2010-00865
57. Shao Y, Wang Z, Tian X, Guo Y, Zhang H. Yeast beta-d-glucans induced antimicrobial peptide expressions against Salmonella infection in broiler chickens. Int J Biol Macromol. (2016) 85:573–84. doi: 10.1016/j.ijbiomac.2016.01.031
58. Cheng Y-H, Lee D-N, Wen C-M, Weng C-F. Effects of β-glucan supplementation on lymphocyte proliferation, macrophage chemotaxis and specific immune responses in broilers. Asian Aust J Anim Sci. (2004) 17:1145–9. doi: 10.5713/ajas.2004.1145
59. Khodambashi Emami N, Samie A, Rahmani HR, Ruiz-Feria CA. The effect of peppermint essential oil and fructooligosaccharides, as alternatives to virginiamycin, on growth performance, digestibility, gut morphology and immune response of male broilers. Anim Feed Sci Tech. (2012) 175:57–64. doi: 10.1016/j.anifeedsci.2012.04.001
60. Plowiec A, Slawinska A, Siwek MZ, Bednarczyk MF. Effect of in ovo administration of inulin and Lactococcus lactis on immune-related gene expression in broiler chickens. Am J Vet Res. (2015) 76:975–82. doi: 10.2460/ajvr.76.11.975
61. Juul-Madsen HR, Viertlböeck B, Härtle S, Smith AL, Göbel TW. Innate Immune Responses. Boston, MA: Academic Press; Elsevier (2014) p. 121–147.
62. Tada H, Nemoto E, Shimauchi H, Watanabe T, Mikami T, Matsumoto T, et al. Saccharomyces cerevisiae- and Candida albicans-derived mannan induced production of tumor necrosis factor alpha by human monocytes in a CD14- and Toll-like receptor 4-dependent manner. Microbiol Immunol. (2002) 46:503–12. doi: 10.1111/j.1348-0421.2002.tb02727.x
63. Sato M, Sano H, Iwaki D, Kudo K, Konishi M, Takahashi H, et al. Direct binding of toll-like receptor 2 to zymosan, and zymosan-induced NF-κB activation and TNFα secretion are down-regulated by lung collectin surfactant protein A. J Immunol. (2003) 171:417–25. doi: 10.4049/jimmunol.171.1.417
64. Silva VK, da Silva JDT, Torres KAA, de Faria Filho DE, Hada FH, de Moraes VMB. Humoral immune response of broilers fed diets containing yeast extract and prebiotics in the prestarter phase and raised at different temperatures. J Appl Poult Res. (2009) 18:530–40. doi: 10.3382/japr.2009-00004
65. Houshmand M, Azhar K, Zulkifli I, Bejo MH, Kamyab A. Effects of prebiotic, protein level, and stocking density on performance, immunity, and stress indicators of broilers. Poult Sci. (2012) 91:393–401. doi: 10.3382/ps.2010-01050
66. SWiATkiewicz S, Arczewska-WŁOsek A, JÓZefiak D. Immunomodulatory efficacy of yeast cell products in poultry: a current review. Worlds Poult Sci J. (2014) 70:57–68. doi: 10.1017/S0043933914000051
67. Brown GD, Taylor PR, Reid DM, Willment JA, Williams DL, Martinez-Pomares L, et al. Dectin-1 is a major β-glucan receptor on macrophages. J Exp Med. (2002) 196:407–12. doi: 10.1084/jem.20020470
68. Duan X, Ackerly M, Vivier E, Anderson P. Evidence for involvement of beta-glucan-binding cell surface lectins in human natural killer cell function. Cell Immunol. (1994) 157:393–402. doi: 10.1006/cimm.1994.1236
69. Kidd P. Th1/Th2 balance: the hypothesis, its limitations, and implications for health and disease. Altern Med Rev. (2003) 8:223–46.
70. Mantis NJ, Rol N, Corthesy B. Secretory IgA's complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. (2011) 4:603–11. doi: 10.1038/mi.2011.41
71. Suzuki K, Ha SA, Tsuji M, Fagarasan S. Intestinal IgA synthesis: a primitive form of adaptive immunity that regulates microbial communities in the gut. Semin Immunol. (2007) 19:127–35. doi: 10.1016/j.smim.2006.10.001
72. Kirmaz C, Bayrak P, Yilmaz O, Yuksel H. Effects of glucan treatment on the Th1/Th2 balance in patients with allergic rhinitis: a double-blind placebo-controlled study. Eur Cytokine Netw. (2005) 16:128–34.
73. Chen Y, Dong L, Weng D, Liu F, Song L, Li C, et al. 1,3-beta-glucan affects the balance of Th1/Th2 cytokines by promoting secretion of anti-inflammatory cytokines in vitro. Mol Med Rep. (2013) 8:708–12. doi: 10.3892/mmr.2013.1553
74. Ritsema T, Smeekens S. Fructans: beneficial for plants and humans. Curr Opin Plant Biol. (2003) 6:223–30. doi: 10.1016/S1369-526600034-7
75. Rossi M, Corradini C, Amaretti A, Nicolini M, Pompei A, Zanoni S, et al. Fermentation of fructooligosaccharides and inulin by bifidobacteria: a comparative study of pure and fecal cultures. Appl Environ Microbiol. (2005) 71:6150–8. doi: 10.1128/AEM.71.10.6150-6158.2005
76. Zhao PY, Wang JP, Kim IH. Effect of dietary levan fructan supplementation on growth performance, meat quality, relative organ weight, cecal microflora, and excreta noxious gas emission in broilers. J Anim Sci. (2013) 91:5287–93. doi: 10.2527/jas.2012-5464
77. Ricke SC. Potential of fructooligosaccharide prebiotics in alternative and nonconventional poultry production systems. Poult Sci. (2015) 94:1411–8. doi: 10.3382/ps/pev049
78. Saminathan M, Sieo CC, Kalavathy R, Abdullah N, Ho YW. Effect of prebiotic oligosaccharides on growth of Lactobacillus strains used as a probiotic for chickens. Afr J Microbiol Res. (2011) 5:57–64. doi: 10.5897/AJMR10.700
79. Kaplan H, Hutkins RW. Metabolism of fructooligosaccharides by Lactobacillus paracasei 1195. Appl Environ Microbiol. (2003) 69:2217–22. doi: 10.1128/AEM.69.4.2217-2222.2003
80. Saulnier DM, Molenaar D, de Vos WM, Gibson GR, Kolida S. Identification of prebiotic fructooligosaccharide metabolism in Lactobacillus plantarum WCFS1 through microarrays. Appl Environ Microbiol. (2007) 73:1753–65. doi: 10.1128/AEM.01151-06
81. Perrin S, Fougnies C, Grill JP, Jacobs H, Schneider F. Fermentation of chicory fructo-oligosaccharides in mixtures of different degrees of polymerization by three strains of bifidobacteria. Can J Microbiol. (2002) 48:759–63. doi: 10.1139/w02-065
82. Pompei A, Cordisco L, Raimondi S, Amaretti A, Pagnoni UM, Matteuzzi D, et al. In vitro comparison of the prebiotic effects of two inulin-type fructans. Anaerobe (2008) 14:280–6. doi: 10.1016/j.anaerobe.2008.07.002
83. Rehman H, Hellweg P, Taras D, Zentek J. Effects of dietary inulin on the intestinal short chain fatty acids and microbial ecology in broiler chickens as revealed by denaturing gradient gel electrophoresis. Poult Sci. (2008) 87:783–9. doi: 10.3382/ps.2007-00271
84. Yusrizal N, Chen TC. Effect of adding chicory fructans in feed on fecal and intestinal microflora and excreta volatile ammonia. Int J Poult Sci. (2003) 2:188–94. doi: 10.3923/ijps.2003.188.194
85. Park SH, Perrotta A, Hanning I, Diaz-Sanchez S, Pendleton S, Alm E, et al. Pasture flock chicken cecal microbiome responses to prebiotics and plum fiber feed amendments. Poult Sci. (2017) 96:1820–30. doi: 10.3382/ps/pew441
86. Li X, Qiang L, Liu L, Xu CH. Effects of supplementation of fructooligosaccharide and:or Bacillus Subtilis to diets on performance and on intestinal microflora in broilers. Archive Tierzucht (2008) 51:64–70. Available online at: https://www.arch-anim-breed.net/51/64/2008/aab-51-64-2008.pdf
87. Telg BE, Caldwell DJ. Efficacy testing of a defined competitive exclusion product in combination with fructooligosaccharide for protection against Salmonella Typhimurium challenge in broiler chicks. J Appl Poult Res. (2009) 18:521–9. doi: 10.3382/japr.2009-00003
88. Rada V, Duskova D, Marounek M, Petr J. Enrichment of bifidobacteria in the hen caeca by dietary inulin. Folia Microbiol. (2001) 46:73–5. doi: 10.1007/BF02825891
89. Donalson LM, McReynolds JL, Kim WK, Chalova VI, Woodward CL, Kubena LF, et al. The influence of a fructooligosaccharide prebiotic combined with alfalfa molt diets on the gastrointestinal tract fermentation, Salmonella enteritidis infection, and intestinal shedding in laying hens. Poult Sci. (2008) 87:1253–62. doi: 10.3382/ps.2007-00166
90. Adhikari P, Cosby DE, Cox NA, Franca MS, Williams SM, Gogal RMJr, et al. Effect of dietary fructooligosaccharide supplementation on internal organs Salmonella colonization, immune response, ileal morphology, and ileal immunohistochemistry in laying hens challenged with Salmonella Enteritidis. Poult Sci. (2018) 97:2525–33. doi: 10.3382/ps/pey101
91. Topping DL, Clifton PM. Short-chain fatty acids and human colonic function: roles of resistant starch and nonstarch polysaccharides. Physiol Rev. (2001) 81:1031–64. doi: 10.1152/physrev.2001.81.3.1031
92. Bogucka J, Dankowiakowska A, Elminowska-Wenda G, Sobolewska A, Szczerba A, Bednarczyk M. Effects of prebiotics and synbiotics delivered in ovo on broiler small intestine histomorphology during the first days after hatching. Folia Biol. (2016) 64:131–43. doi: 10.3409/fb64_3.131
93. Wu TX, Dai XJ, Wu LY. Effects of fructooligosaccharide on the broiler production. Acta Agric Zhejiangensis (1999) 11:85–7.
94. Ten Bruggencate SJ, Bovee-Oudenhoven IM, Lettink-Wissink ML, Van der Meer R. Dietary fructooligosaccharides increase intestinal permeability in rats. J Nutr. (2005) 135:837–42. doi: 10.1093/jn/135.4.837
95. Seifert S, Watzl B. Inulin and oligofructose: review of experimental data on immune modulation. J. Nutr. (2007) 137(11Suppl.):2563–7. doi: 10.1093/jn/137.11.2563S
96. Madej JP, Bednarczyk M. Effect of in ovo-delivered prebiotics and synbiotics on the morphology and specific immune cell composition in the gut-associated lymphoid tissue. Poult Sci. (2016) 95:19–29. doi: 10.3382/ps/pev291
97. Huang R-L, Deng Z-Y, Yang C-B, Yin Y-L, Xie MY, Wu G-Y, et al. Dietary oligochitosan supplementation enhances immune status of broilers. J. Sci. Food Agric. (2007) 87:153–9. doi: 10.1002/jsfa.2694
98. Huang RL, Yin YL, Wu GY, Zhang YG, Li TJ, Li LL, et al. Effect of dietary oligochitosan supplementation on ileal digestibility of nutrients and performance in broilers. Poult Sci. (2005) 84:1383–8. doi: 10.1093/ps/84.9.1383
99. Alles MS, Hartemink R, Meyboom S, Harryvan JL, Van Laere KM, Nagengast FM, et al. Effect of transgalactooligosaccharides on the composition of the human intestinal microflora and on putative risk markers for colon cancer. Am J Clin Nutr. (1999) 69:980–91. doi: 10.1093/ajcn/69.5.980
100. Pruszynska-Oszmalek E, Kolodziejski PA, Stadnicka K, Sassek M, Chalupka D, Kuston B, et al. In ovo injection of prebiotics and synbiotics affects the digestive potency of the pancreas in growing chickens. Poult Sci. (2015) 94:1909–16. doi: 10.3382/ps/pev162
101. Jung SJ, Houde R, Baurhoo B, Zhao X, Lee BH. Effects of galacto-oligosaccharides and a Bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens. Poult Sci. (2008) 87:1694–9. doi: 10.3382/ps.2007-00489
102. Bednarczyk M, Stadnicka K, Kozlowska I, Abiuso C, Tavaniello S, Dankowiakowska A, et al. Influence of different prebiotics and mode of their administration on broiler chicken performance. Animal (2016) 10:1271–9. doi: 10.1017/S1751731116000173
103. Capek P, Kubackova M, Alfoldi J, Bilisics L, Liskova D, Kakoniova D. Galactoglucomannan from the secondary cell wall of Picea abies L. Karst Carbohydr Res. (2000) 329:635–45. doi: 10.1016/S0008-6215(00)00210-X
104. Faber TA, Dilger RN, Hopkins AC, Price NP, Fahey GC Jr. The effects of a galactoglucomannan oligosaccharide-arabinoxylan (GGMO-AX) complex in broiler chicks challenged with Eimeria acervulina. Poult Sci. (2012) 91:1089–96. doi: 10.3382/ps.2011-01993
105. Faber TA, Dilger RN, Iakiviak M, Hopkins AC, Price NP, Fahey GC Jr. Ingestion of a novel galactoglucomannan oligosaccharide-arabinoxylan (GGMO-AX) complex affected growth performance and fermentative and immunological characteristics of broiler chicks challenged with Salmonella typhimurium. Poult Sci. (2012) 91:2241–54. doi: 10.3382/ps.2012-02189
106. Mussatto SI, Mancilha IM. Non-digestible oligosaccharides: a review. Carbohydr Polym. (2007) 68:587–97. doi: 10.1016/j.carbpol.2006.12.011
107. Zhenping S, Wenting L, Ruikui Y, Jia L, Honghong L, Wei S, et al. Effect of a straw-derived xylooligosaccharide on broiler growth performance, endocrine metabolism, and immune response. Can J Vet Res. (2013) 77:105–9.
108. Pourabedin M, Guan L, Zhao X. Xylo-oligosaccharides and virginiamycin differentially modulate gut microbial composition in chickens. Microbiome (2015) 3:15. doi: 10.1186/s40168-015-0079-4
109. Willemsen LEM, Koetsier MA, van Deventer SJH, van Tol EAF. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E1 and E2 production by intestinal myofibroblasts. Gut (2003) 52:1442–7. doi: 10.1136/gut.52.10.1442
110. Hamer HM, Jonkers D, Venema K, Vanhoutvin S, Troost FJ, Brummer RJ. Review article: the role of butyrate on colonic function. Aliment Pharmacol Ther. (2008) 27:104–19. doi: 10.1111/j.1365-2036.2007.03562.x
111. Timbermont L, Lanckriet A, Dewulf J, Nollet N, Schwarzer K, Haesebrouck F, et al. Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils. Avian Pathol. (2010) 39:117–21. doi: 10.1080/03079451003610586
112. Bednarczyk M, Urbanowski M, Gulewicz P, Kasperczyk K, Maiorano G, Szwaczkowski T. Field and in vitro study on prebiotic effect of raffinose family oligosaccharides in chickens. Bull Vet Inst Pulawy (2011) 55:465–9.
113. Madej JP, Stefaniak T, Bednarczyk M. Effect of in ovo-delivered prebiotics and synbiotics on lymphoid-organs' morphology in chickens. Poult Sci. (2015) 94:1209–19. doi: 10.3382/ps/pev076
114. Berrocoso JD, Kida R, Singh AK, Kim YS, Jha R. Effect of in ovo injection of raffinose on growth performance and gut health parameters of broiler chicken. Poult Sci. (2017) 96:1573–80. doi: 10.3382/ps/pew430
115. Ding J, Dai R, Yang L, He C, Xu K, Liu S, et al. Inheritance and establishment of gut microbiota in chickens. Front Microbiol. (2017) 8:1967. doi: 10.3389/fmicb.2017.01967
116. Ballou AL, Ali RA, Mendoza MA, Ellis JC, Hassan HM, Croom WJ, et al. Development of the chick microbiome: how early exposure influences future microbial diversity. Front Vet Sci. (2016) 3:2. doi: 10.3389/fvets.2016.00002
117. Gantois I, Ducatelle R, Pasmans F, Haesebrouck F, Gast R, Humphrey TJ, et al. Mechanisms of egg contamination by Salmonella Enteritidis. FEMS Microbiol Rev. (2009) 33:718–38. doi: 10.1111/j.1574-6976.2008.00161.x
118. Kizerwetter-Swida M, Binek M. Bacterial microflora of the chicken embryos and newly hatched chicken. J Anim Feed Sci. (2008) 17:224–32. doi: 10.22358/jafs/66602/2008
119. Slawinska A, Plowiec A, Siwek M, Jaroszewski M, Bednarczyk M. Long-term transcriptomic effects of prebiotics and synbiotics dielivered in ovo in broiler chickens. PLoS One (2016) 11:e0168899. doi: 10.1371/journal.pone.0168899
Keywords: prebiotic, broilers, immunity, microbiota, mannan oligosaccharides, β-glucans, fructans
Citation: Teng P-Y and Kim WK (2018) Review: Roles of Prebiotics in Intestinal Ecosystem of Broilers. Front. Vet. Sci. 5:245. doi: 10.3389/fvets.2018.00245
Received: 18 May 2018; Accepted: 17 September 2018;
Published: 30 October 2018.
Edited by:
Rajesh Jha, University of Hawaii at Manoa, United StatesReviewed by:
Maria Siwek, University of Science and Technology (UTP), PolandAkshat Goel, Central Avian Research Institute (ICAR), India
Copyright © 2018 Teng and Kim. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Woo Kyun Kim, d2traW1AdWdhLmVkdQ==