
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Psychiatry , 10 July 2020
Sec. Schizophrenia
Volume 11 - 2020 | https://doi.org/10.3389/fpsyt.2020.00680
This article is part of the Research Topic Psychoneuroendocrinology of Psychosis Disorders View all 14 articles
 Anton Iftimovici1,2
Anton Iftimovici1,2 Oussama Kebir1,3
Oussama Kebir1,3 Qin He1
Qin He1 Thérèse M. Jay1
Thérèse M. Jay1 ICAAR Study Group
ICAAR Study Group Guy A. Rouleau4
Guy A. Rouleau4 Marie-Odile Krebs1,3
Marie-Odile Krebs1,3 Boris Chaumette1,3,5*
Boris Chaumette1,3,5*Introduction: The emergence of psychosis in at-risk individuals results from interactions between genetic vulnerability and environmental factors, possibly involving dysregulation of the hypothalamic-pituitary-adrenal axis. Hypercorticism was indeed described in schizophrenia and ultra-high-risk states, but its association with clinical outcome has yet to be demonstrated. The impact of stress through cortisol may vary depending on the expression level of genes related to the stress pathway.
Methods: To test this hypothesis, we selected NR3C1, the gene encoding the glucocorticoid receptor, and modeled through logistic regression how its peripheral expression could explain some of the risk of psychosis, independently of peripheral cortisol levels, in a French longitudinal prospective cohort of 133 at-risk individuals, adjusted for sex, age, cannabis, and antipsychotic medication intake. We then performed a genome-wide association analysis, stratified by sex (55 females and 78 males), to identify NR3C1 expression quantitative trait loci to be used as instrumental variables in a Mendelian randomization framework.
Results: NR3C1 expression was significantly associated with a higher risk of conversion to psychosis (OR = 2.03, p = 0.03), independently of any other factor. Cortisol was not associated with outcome nor correlated with NR3C1. In the female subgroup, rs6849528 was associated both with NR3C1 mRNA levels (p = 0.015, Effect-Size = 2.7) and conversion (OR = 8.24, p = 0.03).
Conclusions: For the same level of cortisol, NR3C1 expression increases psychotic risk, independently of sex, age, cannabis, and antipsychotic intake. In females, Mendelian randomization confirmed NR3C1’s effect on outcome to be unbiased by any environmental confounder.
The concept of schizophrenia has moved from a chronic to a progressive illness that typically emerges during late adolescence and goes through several stages: early vulnerability, at-risk mental state, first episode of psychosis (FEP), and finally, chronic disease (1, 2). The at-risk state includes subjects with psychotic symptoms that are either attenuated or not frequent enough to allow a diagnosis of FEP, or who have a genetic risk and present with nonspecific functional decline. Only up to a third of at-risk subjects might convert to FEP after 3 years and the reasons for this differential outcome are yet to be understood (3). According to the main hypothesis in the field, the emergence of psychotic symptoms could imply an interaction between genes and environment, and could be mediated through epigenetic (4) and transcriptomic processes (5). The biological response to stress has been hypothesized to play a role in pathophysiology. Abnormal cortisol levels have indeed been suggested at each stage: increased basal cortisol in subjects at-risk (6) or with schizophrenia (7), a possibly blunted cortisol awakening response in FEP and in schizophrenia (8), and an attenuated cortisol response to acute psychosocial stress in at-risk subjects (9). Cortisol levels also have been proposed to predict the prognosis in the at-risk individuals (10). However, relatively small sample sizes and heterogeneous measures and outcomes mean that these findings need to be interpreted with caution. Moreover, our recent meta-analysis did not confirm the association between morning salivary cortisol levels and conversion to psychosis (6). The frequency of stress-related dysregulation in the at-risk group as a whole might indeed not allow cortisol by itself to discriminate between future converters and non-converters.
Further investigations of the stress pathway, at a molecular and genetic levels, are therefore warranted. To this purpose, confounders such as sex or environmental factors need to be controlled, as they might account for cortisol’s observed weak predictive power. Cortisol awakening response could be significantly lower in males with FEP compared to male controls and affected females (11). Sex differences have also been suggested in morning cortisol levels in the at-risk stage (12). This moderating effect of sex might further be seen at the molecular level, on the glucocorticoid nuclear receptor NR3C1, which acts as a transcription factor that binds to glucocorticoid response elements and regulates gene expression upon stress. Sex-specific effects of negative environment on NR3C1 regulatory regions’ methylation have been observed (13), as well as sex-specific upregulation of NR3C1 transcription under acute stress in animal models (14). In humans, it has recently been suggested that epigenetic mechanisms, including NR3C1 regulation, might differ between males and females (15), with a sex-dependent role of NR3C1’s methylation in depression, but also a sex-specific effect of allelic variation of the NR3C1 gene (16, 17).
Moreover, NR3C1 expression could be of interest in psychosis, not only through its relationship with cortisol, but also as a possible direct marker. One study, although limited in sample size, suggested that NR3C1 mRNA might be decreased in the dorso-lateral prefrontal cortex of schizophrenia cases, relative to controls, while among schizophrenia cases, it might be increased in suicide-positive vs suicide-negative subjects (18). Furthermore, anxiety and depression are important prodromal symptoms in the emergence of psychosis (2), and allelic variations in the NR3C1 gene have been found associated with depression, with or without psychotic features (19, 20), as well as with cognitive deterioration, independently of cortisol levels (20). However, there is not, to our knowledge, any NR3C1 gene expression (mRNA or protein) study in the context of emergence of psychosis.
In this context, we hypothesized that, in at-risk individuals, NR3C1 peripheral expression may explain some of the risk of psychosis, independently of cortisol levels, and adjusted by sex, age, cannabis and antipsychotic medication intake. To shore up this argument, we considered a Mendelian randomization analysis in order to account for all possible unknown environmental factors. In this analysis, the effect of an “exposure” variable (here, the level of gene expression) on an “outcome” variable (here, conversion to psychosis) is tested through the use of an “instrument” that explains exposure independently of confounding factors (e.g. environmental biases): stratifying a population using the instrumental variable thus allows to test the unbiased effect of the exposure on the outcome (21). A genetic variant representing an expression quantitative trait locus (eQTL) of this gene could serve as such an instrumental variable, because it does not depend on the environment (Figure 1) (22). In our data, if an eQTL explained the variation of NR3C1 expression, and if this expression level was associated with conversion to psychosis, then stratifying at-risk individuals on this eQTL could highlight the differences in conversion rates that are only due to NR3C1 expression levels, independently of any non-genetic confounders.
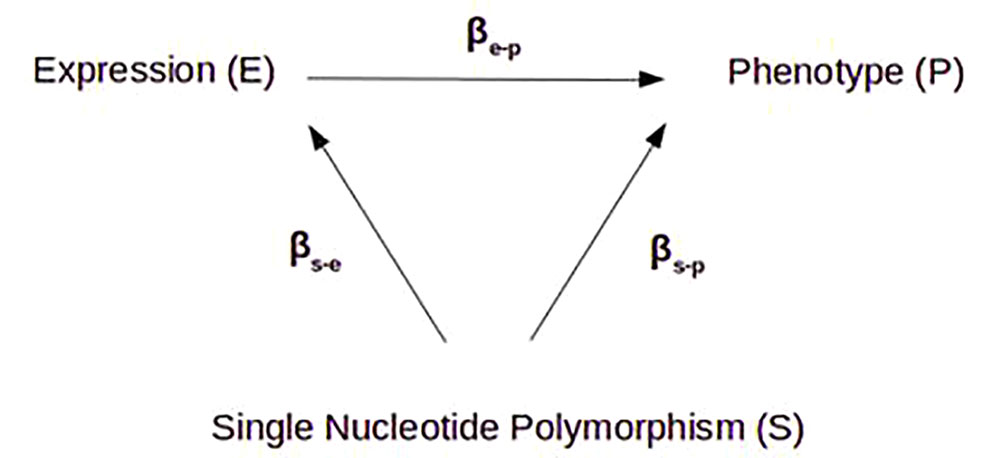
Figure 1 Principle of Mendelian randomization. In a purely causal model, the variation of gene expression (e) is fully determined by an eQTL SNP (s), which itself has no effect (β) on phenotype (p) except through that gene’s expression, so βs-e x βe-p = βs-p. By identifying the different β, we can estimate the effect of gene expression on the phenotype (explained variance), free of other potential confounding factors.
Using a French longitudinal prospective cohort of individuals at-risk for psychosis, presenting attenuated or prodromal symptoms [ICAAR, previously described (23)], with data on cortisol levels, NR3C1 expression levels, and whole-genome genotyping, we thus tested the following hypotheses in a Mendelian randomization framework (Figure 2). i) At the hormonal and molecular level, are basal cortisol and NR3C1 expression reliable biomarkers of conversion to psychosis? ii) At the genetic level, does a genome-wide analysis study (GWAS), stratified by sex to account for genotype-by-sex interactions (16, 17), identify any eQTL that can explain NR3C1 mRNA levels? iii) Finally, would such an eQTL be associated with psychosis, thus confirming the environmentally unbiased effect of NR3C1 on risk of psychosis? Last, as secondary outcomes, we tested if NR3C1 expression is associated with functional or depressive outcomes or more specifically with psychotic features.
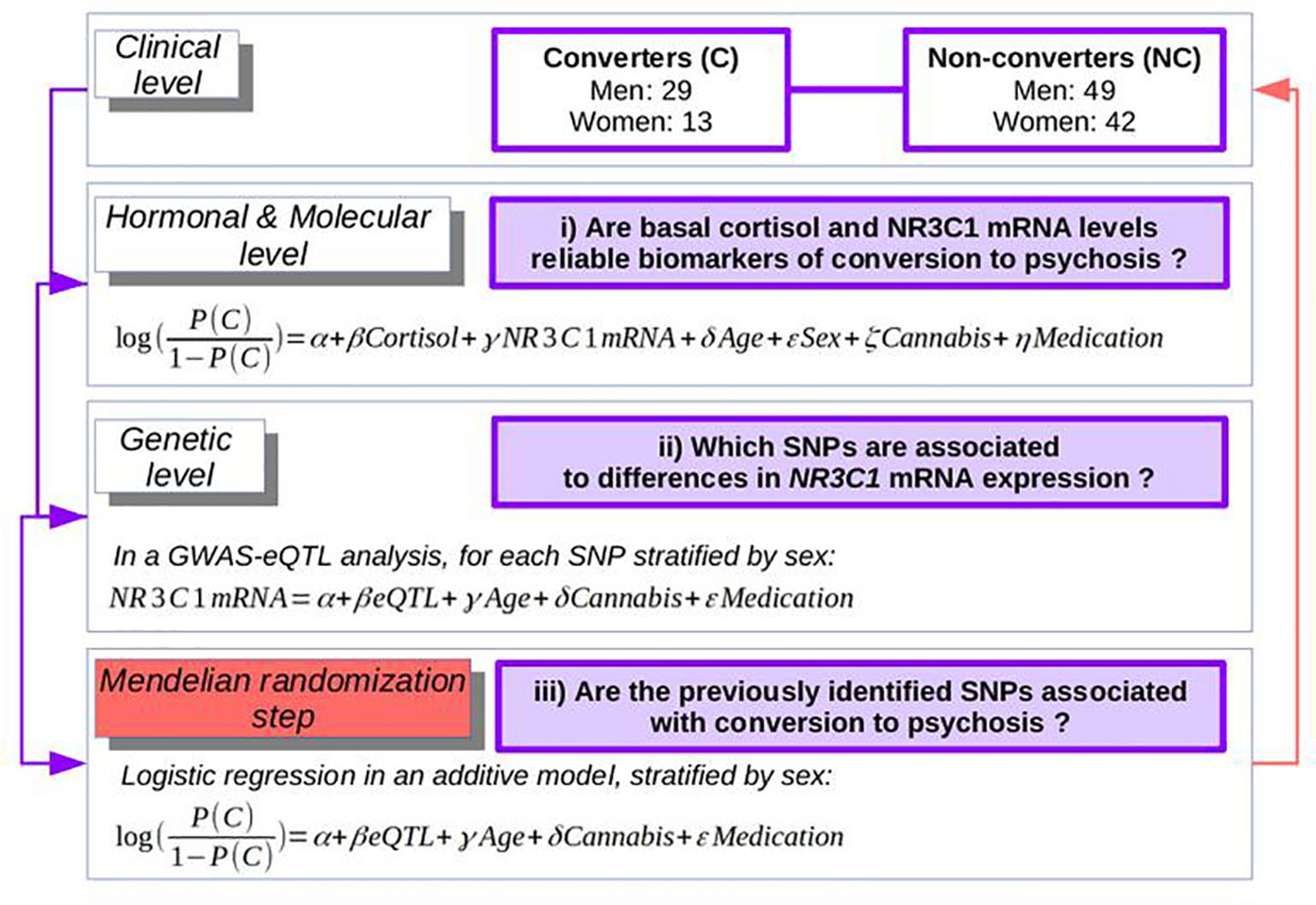
Figure 2 The strategy of the analyses: from clinical to hormonal, molecular, and genetics levels.
Participants were recruited through the French ICAAR cohort (PHRC AOM-07-118, promoted by Hôpital Sainte-Anne) among help-seeking individuals (16 to 30 years old) consecutively referred to the Adolescent and Young Adult Assessment Centre (Service Hospitalo-Universitaire, Hôpital Sainte-Anne, Paris, France) between 2009 and 2014 (23). All help-seeking individuals were examined at baseline and after 1 year follow-up with the Comprehensive Assessment of at-risk mental state, CAARMS (24), in its translated version (25), by specifically trained psychiatrists. After baseline assessment, a consensus meeting for best estimated diagnosis was held, and help-seekers were classified as at-risk for psychosis stage IA or stage IB according to the staging model distinction (26). Were included in the analysis all at-risk stage IA and stage IB individuals, for which clinical, biological and genetic data was available. Stage IA included patients with mild or non-specific symptoms of psychosis or severe mood disorder, and mild functional change. Stage IB included patients with moderate subthreshold symptoms and moderate functional change. Inclusion criteria were alterations in global functioning (Social and Occupational Functioning Assessment Scale score < 70) during the past year, which were associated with psychiatric symptoms and/or subjective cognitive complaints. Exclusion criteria included manifest symptoms of psychosis (fulfilling DSM-IV criteria), or other established psychiatric diagnoses (pervasive developmental disorder, bipolar disorder, obsessive compulsive disorder), serious or non-stabilized somatic and neurological disorders, head injury and IQ below 70. Psychotic conversion was characterized using the CAARMS-defined psychosis onset threshold (i.e., supra-threshold psychotic symptoms—thought content, perceptual abnormalities and/or disorganized speech—present for more than 1 week). Individuals who reached the threshold during the follow-up were considered as converters and individuals who recovered or displayed persistent subthreshold symptoms were called non-converters. Additionally, each individual underwent clinical assessments including the Social and Occupational Functioning Assessment Scale (SOFAS), the Positive And Negative Syndrome Scale (PANSS) and the Montgomery-Åsberg depression rating scale (MADRS), cannabis intake in the last month, and antipsychotic treatment summarized by the chlorpromazine equivalent doses (references for the computation of chlorpromazine equivalent doses are available in Supplementary Table 1). The population demographic and clinical characteristics at baseline are described in Table 1. We used the conversion status available after 1-year follow-up in order to have an outcome that was closest to biological sampling.
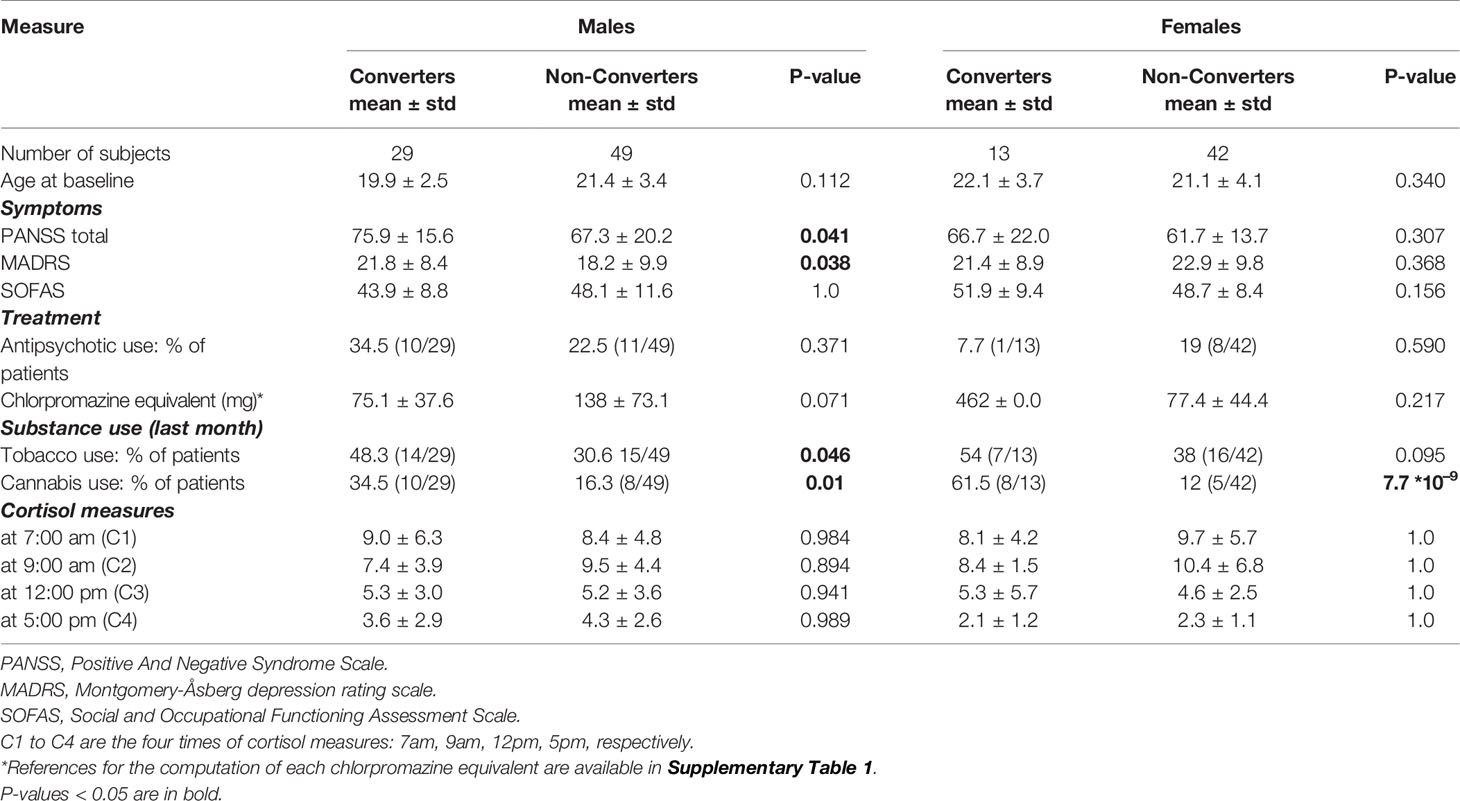
Table 1 Demographic, clinical characteristics, and cortisol measures of the male and female datasets at baseline.
Salivary cortisol was collected using a synthetic swab at four time-points during the day after initial enrollment. Cortisol was measured at 7:00 am (C1), 9:00 am (C2), 12:00 pm (C3), and 5:00 pm (C4). A range of ± 1 h was accepted for each time of sampling. None of the subjects worked night shifts. The saliva samples were stored at -20°C until analysis. After thawing, saliva samples were centrifuged at 2000 g for 10 min, which resulted in a clear supernatant of low viscosity; 100 μl of saliva were used for duplicate analysis of each sample. Cortisol measurement was done using a competitive solid phase time-resolved fluorescence immunoassay with fluorometric end point detection (DELFIA) conducted by Cortisollabor, University of Trier, Department of Clinical and Physiological Psychology, Trier, Germany (27). The inter-assay coefficient of variation was 8.6% and intra-assay coefficient of variation was 4.3% as previously reported (6).
Total RNA was extracted and purified from blood samples (PAXgene tubes) using a QIAcube robot and PAXgene Blood RNA kit (QIAGEN) according to the manufacturer’s protocol. Quality control was done using LabChip GX (Perkin Elmer, Waltham USA). The full quantitative PCR (qPCR) protocol has been described in Chaumette et al. (5). Briefly, complementary DNA (cDNA) synthesis was performed using Reverse Transcription Master Mix from Fluidigm® according to the manufacturer’s protocol with random primers using a Nexus thermocycler (Eppendorf). Specific target pre-amplification was performed using a Fluidigm® PreAmp Master Mix at 12 cycles. Real time PCR was performed on the qPCR-HD-Genomic Paris Centre platform, using BioMark™ HD System, GE Dynamic Arrays (Fluidigm) and TaqMan® Gene Expression assays (Life Technologies, ThermoFisher). Thermal conditions for qPCR were: 25°C for 30 min and 70°C for 60 min for thermal mix; 50°C for 2 min and 95°C for 10 min for hot start; 40 cycles at 95°C for 15 s and 60°C for 1 min. Data were processed by automatic threshold, with linear derivative baseline correction using BioMark Real-Time PCR Analysis Software 4.0.1 (Fluidigm). The quality threshold was set at the default setting of 0.65. Normalization was done using the GAPDH rate followed by a livak normalization with a transformation by the 2ΔΔCT method (28) providing the relative mRNA expression level of NR3C1 in each sample. Moreover, we checked on the Genevestigator platform (https://genevestigator.com), using the RefGenes tool (29), that GAPDH was among the top 10 genes with an expression that was both stable, and in NR3C1 ranges, for it to be a good reference for normalization (Supplementary Figure 1).
In the ICAAR cohort, 102 Caucasian individuals have been genotyped using the Infinium PsychArray-24 v1.2 BeadChip (Illumina). This chip was designed by the Psychiatric Genomic Consortium and is enriched for polymorphisms relevant for psychiatric diseases. Single Nucleotide Polymorphisms (SNPs) annotation was given by the Illumina annotation file. Plink v2.0 (www.cog-genomics.org/plink/2.0/) was used for quality control and association analyses. The quality control excludes samples with less than 90% genotyping rate (mind > 0.1), SNPs with a minor allele frequency less than 1% (maf < 0.01) and SNPs that were not genotyped in at least 60% of the sample (geno > 0.4). No sample was excluded during the quality control; after filtering, 306,841 SNPs remained to be analyzed in male samples and 300,732 in female samples. Linkage disequilibrium and haplotype blocks were analyzed with Haploview.
Statistical analyses of basal cortisol level and gene expression in ICAAR cohort were performed using Python 3.7.2 and R 3.6.2. Group distributions of quantitative values were compared with a non-parametric Mann-Whitney Wilcoxon rank sum test. Comparisons of multiple ordinal categorical groups were made with a Chi-squared test for proportions. Correlations were calculated using Spearman’s test. For demographic and clinical comparisons between groups, the Bonferroni corrected threshold was at 0.004. However, to be more stringent in detecting the potential confounders at baseline, we considered the uncorrected p-value threshold of 0.05 in order not to ignore any clinical difference that could bias the association between NR3C1 expression and conversion, and for which Mendelian randomization would be needed. All measures were standardized using the mean and standard deviation in all the cohort. The reported effect-sizes (ES) were calculated with Hedge’s g. In our cohort of 133 subjects, we first regressed the odds of conversion, OR, as follows (Figure 2):
A second model using a linear regression on the same combination of explanatory variables was applied to test their association with depression (MADRS scale) and functional outcomes (SOFAS scale). Then for each SNP, stratified by sex, we applied a linear regression as follows:
Finally, we applied a logistic regression to model the odds of conversion in the female group as follows:
In order to derive robust non-parametric p-values for each Wilcoxon test, we randomly permuted the assignment of values to the groups, and repeated the statistical test 10,000 times. We then computed how many times a p-value was smaller or equal to the observed one. The reported p-value was calculated as the ratio of this number to the total number of tests done (10,000). The 95% confidence intervals (95% CI) were computed by bootstrapping, where the variance of means from each group was estimated by random sampling with replacement. This prevents any inference on the statistical distribution of the population.
The Mendelian randomization analysis follows the steps of a two-stage least squares (2SLS) regression analysis. First, the effect of gene expression (e) on phenotype (p) was estimated by >logistically regressing the log-odds of conversion on cortisol levels, NR3C1 mRNA levels, age, sex, cannabis and antipsychotic intake. This gives a βe-p parameter for the effect of NR3C1 mRNA on conversion. Second, using Plink v2.0 (30), we performed a GWAS eQTL analysis stratified by sex. NR3C1 mRNA levels were used as the quantitative trait, linearly regressed on the alleles of each SNP in the GWAS, giving a βs-e parameter that estimates the effect of SNP eQTLs on expression (e). The usual false discovery rate (FDR) threshold of 0.05 was lowered to 0.025 to account for the sex stratification. Last, we measured the effect of the SNP we found (s) on phenotype (p) by applying logistic regression to explain the log-odds of conversion by the alleles of the SNP, giving a βs-p parameter (Figure 1). Its significance would confirm the effect of mRNA levels on the risk of conversion, independently from non-genetic confounders (Figure 2). An additive model was applied for the SNPs’ alleles.
At baseline, converters and non-converters were comparable in age, antipsychotic treatment and clinical scales, except for male converters who exhibited higher total PANSS and MADRS scores than male non-converters (p = 0.041 and 0.038, respectively). In both male and female groups, converters showed a higher cannabis use than non-converters (p = 0.010 and p = 7.7*10⁻⁹ respectively). Male converters smoked more tobacco than male non-converters (p = 0.046). None of the cortisol levels at any 4 times of the day were significantly different between converters and non-converters in either group (Table 1). None of the cortisol levels were significantly associated with NR3C1 expression (Supplementary Figure 2).
In model (I), an increase in NR3C1 mRNA levels significantly increased the odds of conversion, independently of cortisol, age, sex, cannabis use, and antipsychotic intake (OR = 2.03, p = 0.03), with an explained variance of 11.6% (pseudo-R²) (Table 2, Figure 3, Supplementary Figure 3).
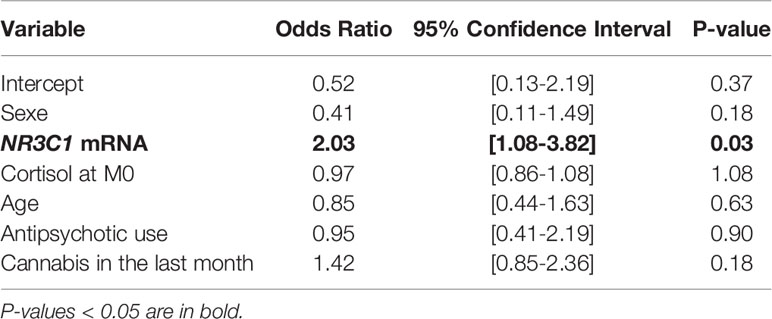
Table 2 NR3C1 expression explains the risk of conversion independently of cortisol, age, sex, cannabis, and antipsychotic intake.
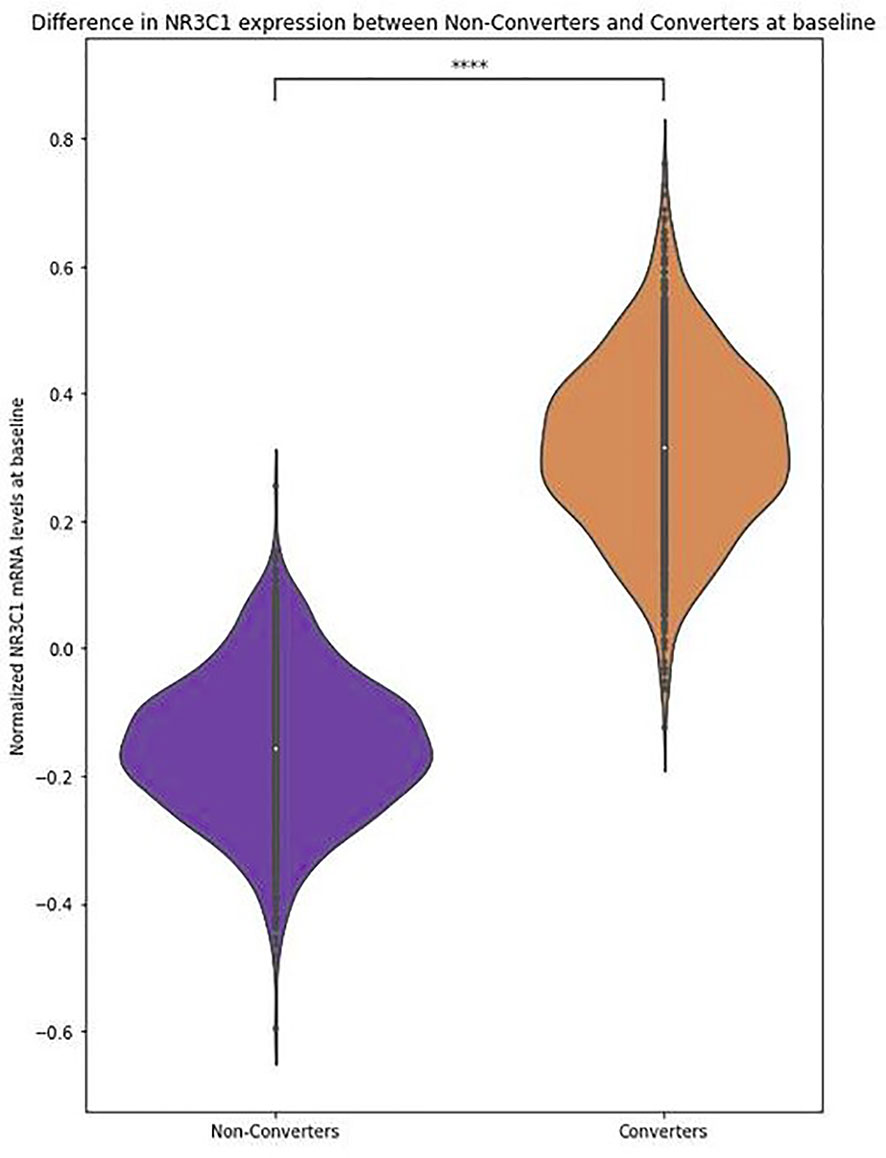
Figure 3 NR3C1 expression levels are higher in converters than in non-converters.
When applying the same combination of variables from model (I) to explain dimensional outcomes, no significant association was found between depression, or functional outcome, and NR3C1 mRNA or cortisol, adjusted for age, sex, cannabis use, and antipsychotic intake.
After FDR correction, correction for stratification, and adjustment for age, cannabis, and antipsychotic intake, only one SNP, rs6849528, remained associated with NR3C1 expression (p = 0.015, ES = 2.7) and explained 43% of its variance in the female group. The mean of NR3C1 mRNA levels was robustly higher in subjects with at least one allele A (genotype AG or AA, 95% CI = [0.12‑0.15]) than in subjects with genotype GG (95%CI = [0.07 − 0.09]), with a significant non-parametric p-value = 2*10-4 (Supplementary Figures 4 and 5). No significant eQTL was found in males with the same correction and adjustments.
rs6849528 could therefore be used as an instrumental variable for Mendelian randomization in the female group. Because the probability of distribution of its alleles is not related to environmental factors such as cannabis or tobacco use, randomizing the female cohort on rs6849528 would account for any unknown environmental confounder that could bias the association between NR3C1 expression and conversion. Accounting for age, cannabis, and antipsychotic intake, the risk of conversion to psychosis was significantly associated with rs6849528, in an additive model, with an odds-ratio of 8.24 (p = 0.03) (Figure 4).
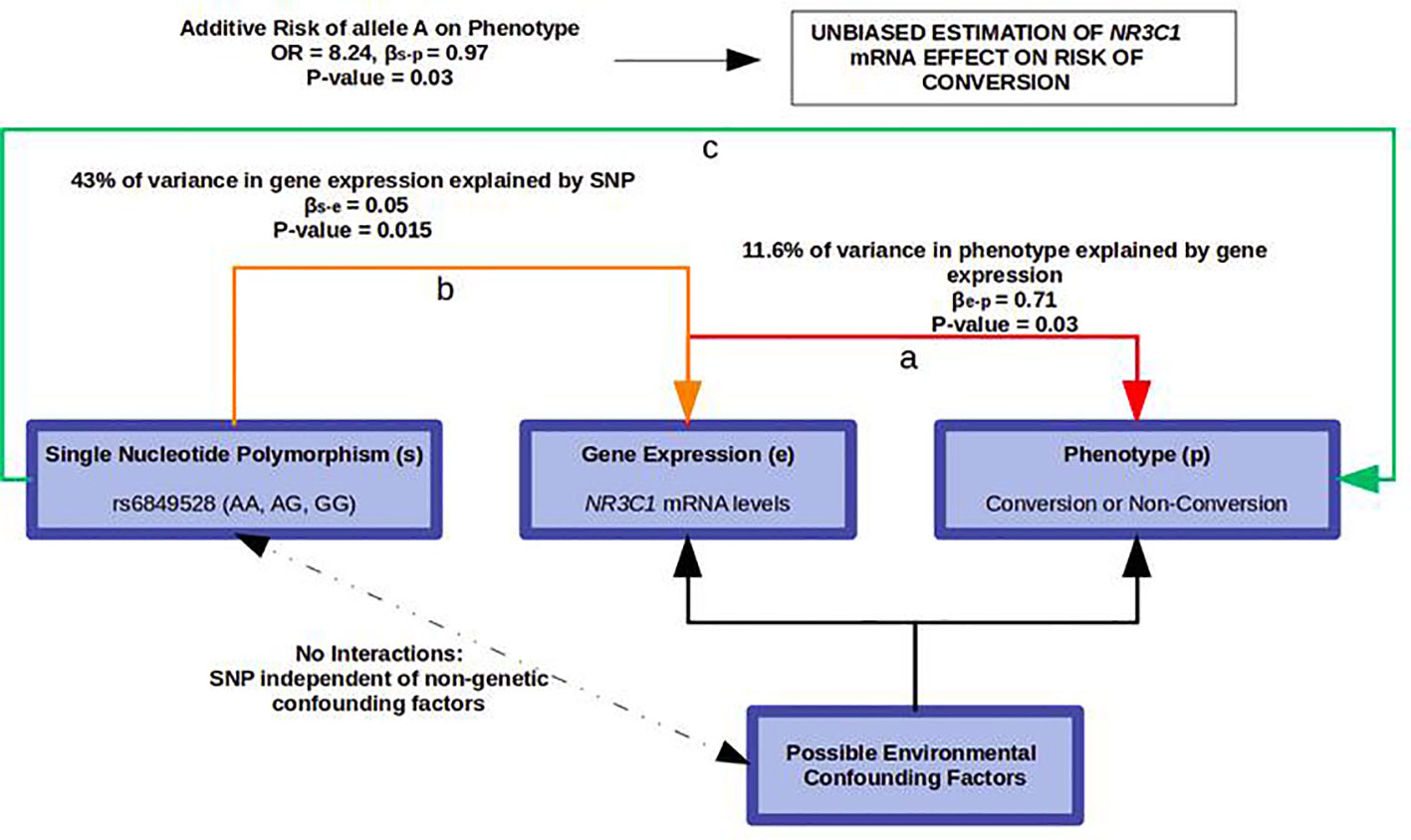
Figure 4 Mendelian Randomization using rs6849528 as an instrumental variable. a) NR3C1 mRNA levels are associated with psychosis. b) rs6849528 alleles lead to differential NR3C1 expression. c) Because the probability of distribution of allele A is independent of environmental factors, its association with psychosis is in favour of the involvement of NR3C1 mRNA in this risk, independently of non-genetic confounding factors.
We applied multiple levels of analysis (hormonal, molecular and genetic) to test the association of conversion to psychosis with potential biomarkers from the biological stress pathway (NR3C1 expression level and cortisol levels at different times of the day), while adjusting for age, sex, cannabis use, and antipsychotic intake, in a longitudinal cohort of 133 subjects at-risk for psychosis. Both stage IA and stage IB were included to be representative of daily clinical practice.
First, we showed that after adjusting for cortisol levels, an increase in NR3C1 expression was significantly associated with a higher risk of conversion to psychosis. Cortisol itself was not associated with conversion, which is in line with our previous meta-analysis assessing the morning levels of cortisol (6). This negative result was extended to other times of the day (waking time, noon, afternoon). As cortisol levels were neither correlated with NR3C1 expression, this may suggest cortisol’s effects in psychosis might be conditional to the underlying biological and genetic background.
Second, to strengthen the validity of NR3C1 mRNA’s association with psychosis, we performed a Mendelian randomization analysis. This allowed to account for any possibly uncontrolled environmental confounding factor that could have biased the association. Because of reported NR3C1 genotype-by-sex effects, we performed this GWAS eQTL analysis stratifying by sex. In females, we identified a SNP (rs6849528) strongly associated with NR3C1 expression, and the odds of conversion to psychosis appeared significantly increased in the group with the minor allele A. Because the probability of distribution of alleles is independent of environmental factors, this SNP’s association with psychosis confirmed the involvement of NR3C1 expression in the risk of psychosis, independently of any non-genetic confounder (Figure 4).
Whereas we initially postulated that the environment regulated gene expression, through the mediation of biological stress, we found that NR3C1 expression, one of the genes most implicated in the stress pathway, was not dependent on cortisol levels, but rather on genetic variability, in females. Given the same level of cortisol, presuming the same level of stress, female individuals with higher NR3C1 expression levels appeared thus more vulnerable than others to the risk of conversion.
The GWAS analysis did not find any significant eQTL associated with NR3C1 expression in males, so Mendelian randomization could not be applied to bolster the association between NR3C1 and psychosis in males. The absence of significant eQTL in males might either be due to a lack of power in a sample not large enough to detect the small effect-sizes of common polymorphisms, either to the aforementioned genotype-by-sex specificity of NR3C1 regulation (16, 17).
In this Mendelian randomization framework, we used rs6849528 as an independent instrumental variable for randomization. This SNP, located in an intron of the acyl-CoA oxidase 3 gene (ACOX3) on chromosome 4, acted as a trans-eQTL regulator of NR3C1, located on chromosome 5. We wish to highlight the fact that we relied on its association with NR3C1 expression, regardless of any actual pathophysiological mechanism, and with the unique purpose of randomizing the cohort in an environmentally unbiased way. It is indeed not possible to draw any conclusion regarding rs6849528’s real biological effect, especially as it may be associated with psychosis through a pleiotropic effect rather than causal association. ACOX3 participates in peroxisomal fatty acid beta-oxidation. Peroxisomal dysregulation has been described in relation with psychotic symptoms, in the context of inborn errors of metabolism (31), so a possible direct effect of ACOX3 gene on conversion to psychosis could not be ruled out. rs6849528 might affect NR3C1 gene expression on the one hand, and phenotype, independently of NR3C1 expression, on the other. In a purely causal model, the variation of gene expression (e) is fully determined by an eQTL SNP (s), which itself has no effect on phenotype (p) except through that gene’s expression, and βs-e x βe-p = βs-p (22). However, in our study, βs-e x βe-p << βs-p (0.04 < 0.97) suggesting pleiotropy. This was expected as psychosis is a complex phenotype resulting from the effect of many genetic and environmental factors. This also explained the limited variance of risk of conversion explained by NR3C1 gene expression (11.6%) compared to the variance of gene expression determined by the eQTL (43%) (Figure 4).
Moreover, the causal biological effect may be driven by other SNPs in linkage disequilibrium with rs6849528, which could act as a proxy for its entire haplotype block in our eQTL analysis. None of the SNPs in rs6849528 haplotype block have been associated with schizophrenia in a GWAS. However, the haplotype block around rs6849528 comprises several SNPs acting as eQTLs of transfer RNA methyltransferase 44 (TRMT44), according to the GTEx portal (https://www.gtexportal.org/home/). One of these eQTL variants, rs6845969, impacts the expression of TRMT44 in regions such as the hypothalamus or the pituitary gland, involved in the stress pathway. Other variants in this haplotype act as eQTLs for expression of TRMT44 in the nucleus accumbens, relevant to motivation through reward and reinforcement (rs2386223; rs12503034, rs1880025). None of these SNPs were available for testing in our eQTL GWAS, but they could constitute pathophysiologically relevant factors; tRNAs have a major role in translation, decoding mRNA sequence into protein, and methylation of tRNA contributes to its stabilization (32). Conversely, hypomethylation leads to cleavage of tRNAs into tRNA-derived small RNA fragments, like microRNAs, which have been shown to activate stress pathways (33). This might explain how genetic variants could be associated with the expression levels of a distant mRNA (trans eQTL). Through their eQTL effect on TRMT44 activity, they might indirectly lead to an increase in specific miRNAs which in turn regulate gene expression. For instance, NR3C1 has been reported to contain in its 3’ untranslated region a target of miR-124 (34).
Our study has several limitations. First, sampling times varied up to 1 h around the theoretical times for cortisol sampling. This could explain why cortisol measurements did not correlate with outcome. Second, a limitation inherent to Mendelian randomization analysis is that it is not possible to account for genetic confounders. In the same haplotype block, one SNP might only be associated with NR3C1 expression level, while another might only correlate with phenotype. Their linkage disequilibrium would therefore act as a genetic confounding factor that cannot be disentangled by such analysis (35). Third, we only considered the variance of gene expression explained by the eQTL, while other non-genetic factors are also known to regulate NR3C1 expression. For instance, CpG-specific methylation of NR3C1 promoter regions has been robustly implicated in its expression and correlated with psychosocial stress (36). However, this restriction to genetic factors was necessary in order to ensure the unbiasedness of Mendelian randomization. Fourth, the eQTL analysis did not find any of the previously reported SNPs that are in the NR3C1 gene (19). However, such studies used a candidate gene approach to analyze SNPs, while we looked for eQTL genome-wide. Any eQTL we found had to have therefore an important effect-size strengthening its interest for Mendelian randomization. Also, all the data came from peripheral measures: gene expression in blood and cortisol level in saliva. These data may imperfectly reflect the levels in brain tissue. However, blood-brain correlation has been reported to range from 0.25 to 0.64 and to be greater for genes highly expressed in both tissues (37). Finally, we cannot exclude that some non-converters at 1 year did convert later, but the conversion rate is maximal during the first year (3).
Further work is required to replicate our results, but our multimodal approach, using phenotypic, hormonal, transcriptomic and genetic data, illustrates how the implementation of statistical methods, e.g. with Mendelian randomization analyses, could help to detect biomarkers in psychiatry by adjusting for environmental factors. This also suggests that the impact of stress should be investigated in a comprehensive way, where cortisol levels would only be one of these factors.
The datasets presented in this article are not readily available because of an ethical issue. The consent form signed by the participants to the ICAAR study did not indicate that the genetic data would be shared or deposited in a repository.
This study involved human participants and was reviewed and approved by Comité de protection des personnes, Ile-de-France III, Paris, France. Written informed consent to participate in this study was provided by the participants or their legal guardians.
AI, BC and M-OK designed the study. AI, OK, M-OK, GR, and BC obtained the funding and supervised the study. AI, OK, the ICAAR study group and BC collected the data. AI and BC analyzed the data. AI, OK, QH, TJ, GR, M-OK, and BC interpreted the data. AI, OK, and BC drafted the manuscript. All authors contributed to the article and approved the submitted version.
This work has been supported by the French government’s “Investissements d’Avenir” programme, which is managed by the Agence Nationale de la Recherche (ANR), under the reference ANR-18-RHUS-0014 (“Project PsyCARE”). This work was also supported by the French Ministry grant PHRC AOM07-118 (for the ICAAR cohort), Institut National de la Santé et de la Recherche Médicale (INSERM), Université Paris Descartes (recurrent funding), the Canadian Institutes of Health research (GR), the Fondation Bettencourt Schueller (BC), and the Fondation pour la Recherche Médicale (AI). The Centre Hospitalier Sainte-Anne promoted the study. The sponsors had no role in the design and conduct of the study, in the collection, management, analysis or interpretation of the data, in the preparation, review or approval of the manuscript, or in the decision to submit the manuscript for publication. All the authors declare they have no competing interests related to this work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank all the patients and parents who participated in the ICAAR study, the staff from the C’JAAD team and the staff from the Clinical Evaluation and Research Center at the Service Hospitalo-Universitaire, Centre Hospitalier Sainte-Anne. A special thanks to Yannick Morvan for his help with the data management and his statistical advice. We would like to thank the contributors of the ICAAR Study Group: Isabelle Amado, Julie Bourgin, Claire Daban Huard, Célia Jantac Mam-Lam-Fook, Marion Plaze, Fabrice Rivollier.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2020.00680/full#supplementary-material
1. Nelson B, McGorry PD, Wichers M, Wigman JTW, Hartmann JA. Moving From Static to Dynamic Models of the Onset of Mental Disorder: A Review. JAMA Psychiatry (2017) 74(5):528. doi: 10.1001/jamapsychiatry.2017.0001
2. Howes OD, Murray RM. Schizophrenia: An Integrated Sociodevelopmental-Cognitive Model. Lancet (2014) 383:1677–87. doi: 10.1016/S0140-6736(13)62036-X
3. Fusar-Poli P, Borgwardt S, Bechdolf A, Addington J, Riecher-Rössler A, Schultze-Lutter F, et al. The Psychosis High-Risk State: A Comprehensive State-of-the-Art Review. JAMA Psychiatry (2013) 70(1):107. doi: 10.1001/jamapsychiatry.2013.269
4. Kebir O, Chaumette B, Rivollier F, Miozzo F, Lemieux Perreault LP, Barhdadi A, et al. Methylomic Changes during Conversion to Psychosis. Mol Psychiatry (2017) 22(4):512–18. doi: 10.1038/mp.2016.53
5. Chaumette B, Kebir O, Pouch J, Ducos B, Selimi F, ICAAR study group, et al. Longitudinal Analyses of Blood Transcriptome During Conversion to Psychosis. Schizophr Bull (2019) 45(1):247–55. doi: 10.1093/schbul/sby009
6. Chaumette B, Kebir O, Mam-Lam-Fook C, Morvan Y, Bourgin J, Godsil BP, et al. Salivary Cortisol in Early Psychosis: New Findings and Meta-Analysis. Psychoneuroendocrinology (2016) 63:262–70. doi: 10.1016/j.psyneuen.2015.10.007
7. Girshkin L, Matheson SL, Shepherd AM, Green MJ. Morning Cortisol Levels in Schizophrenia and Bipolar Disorder: A Meta-Analysis. Psychoneuroendocrinology (2014) 49:187–206. doi: 10.1016/j.psyneuen.2014.07.013
8. Berger M, Kraeuter AK, Romanik D, Malouf P, Amminger GP, Sarnyai Z. Cortisol Awakening Response in Patients with Psychosis: Systematic Review and Meta-Analysis. Neurosci Biobehav Rev (2016) 68:157–66. doi: 10.1016/j.neubiorev.2016.05.027
9. Pruessner M, Béchard-Evans L, Boekestyn L, Iyer SN, Pruessner JC, Malla AK. Attenuated Cortisol Response to Acute Psychosocial Stress in Individuals at Ultra-High Risk for Psychosis. Schizophr Res (2013) 146:79–86. doi: 10.1016/j.schres.2013.02.019
10. Walker EF, Trotman HD, Pearce BD, Addington J, Cadenhead KS, Cornblatt BA, et al. Cortisol Levels and Risk for Psychosis: Initial Findings From the North American Prodrome Longitudinal Study. Biol Psychiatry (2013) 74(6):410–7. doi: 10.1016/j.biopsych.2013.02.016
11. Pruessner M, Vracotas N, Joober R, Pruessner JC, Malla AK. Blunted Cortisol Awakening Response in Men with First Episode Psychosis: Relationship to Parental Bonding. Psychoneuroendocrinology (2013) 38:229–40. doi: 10.1016/j.psyneuen.2012.06.002
12. Carol EE, Spencer RL, Mittal VA. Sex Differences in Morning Cortisol in Youth at Ultra-High-Risk for Psychosis. Psychoneuroendocrinology (2016) 72:87–93. doi: 10.1016/j.psyneuen.2016.06.013
13. Jaric I, Rocks D, Cham H, Herchek A, Kundakovic M. Sex and Estrous Cycle Effects on Anxiety- and Depression-Related Phenotypes in a Two-Hit Developmental Stress Model. Front Mol Neurosci (2019) 12:74. doi: 10.3389/fnmol.2019.00074
14. Bourke CH, Raees MQ, Malviya S, Bradburn CA, Binder EB, Neigh GN. Glucocorticoid Sensitizers Bag1 and Ppid Are Regulated by Adolescent Stress in a Sex-Dependent Manner. Psychoneuroendocrinology (2013) 38:84–93. doi: 10.1016/j.psyneuen.2012.05.001
15. Hill J, Pickles A, Wright N, Quinn JP, Murgatroyd C, Sharp H. Mismatched Prenatal and Postnatal Maternal Depressive Symptoms and Child Behaviours: A Sex-Dependent Role for NR3C1 DNA Methylation in the Wirral Child Health and Development Study. Cells (2019) 8:943. doi: 10.3390/cells8090943
16. Sarubin N, Hilbert S, Naumann F, Zill P, Wimmer AM, Nothdurfter C, et al. The Sex-Dependent Role of the Glucocorticoid Receptor in Depression: Variations in the NR3C1 Gene Are Associated with Major Depressive Disorder in Women but Not in Men. Eur Arch Psychiatry Clin Neurosci (2017) 267:123–33. doi: 10.1007/s00406-016-0722-5
17. Kumsta R, Entringer S, Koper JW, van Rossum EFC, Hellhammer DH, Wüst S. Sex Specific Associations between Common Glucocorticoid Receptor Gene Variants and Hypothalamus-Pituitary-Adrenal Axis Responses to Psychosocial Stress. Biol Psychiatry (2007) 62:863–69. doi: 10.1016/j.biopsych.2007.04.013
18. Sinclair D, Fullerton JM, Webster MJ, Weickert CS. Glucocorticoid Receptor 1B and 1C MRNA Transcript Alterations in Schizophrenia and Bipolar Disorder, and Their Possible Regulation by GR Gene Variants. PloS One (2012) 7:e31720. doi: 10.1371/journal.pone.0031720
19. Schatzberg AF, Keller J, Tennakoon L, Lembke A, Williams G, Kraemer FB, et al. HPA Axis Genetic Variation, Cortisol and Psychosis in Major Depression. Mol Psychiatry (2014) 19:220–27. doi: 10.1038/mp.2013.129
20. Keller J, Gomez R, Williams G, Lembke A, Lazzeroni L, Murphy GM, et al. HPA Axis in Major Depression: Cortisol, Clinical Symptomatology and Genetic Variation Predict Cognition. Mol Psychiatry (2017) 22:527–36. doi: 10.1038/mp.2016.120
21. Davey Smith G, Ebrahim S. ’Mendelian Randomization’: Can Genetic Epidemiology Contribute to Understanding Environmental Determinants of Disease? Int J Epidemiol (2003) 32:1–22. doi: 10.1093/ije/dyg070
22. Zhu Z, Zhang F, Hu H, Bakshi A, Robinson MR, Powell JE, et al. Integration of Summary Data from GWAS and EQTL Studies Predicts Complex Trait Gene Targets. Nat Genet (2016) 48:481–87. doi: 10.1038/ng.3538
23. Oppetit A, Bourgin J, Martinez G, Kazes M, Mam-Lam-Fook C, Gaillard R, et al. The C’JAAD: A French Team for Early Intervention in Psychosis in Paris: An Early Intervention Team in Paris. Early Interv Psychiatry (2018) 12:243–49. doi: 10.1111/eip.12376
24. Yung AR, Yuen HP, Mcgorry PD, Phillips LJ, Kelly D, Dell’Olio M, et al. Mapping the Onset of Psychosis: The Comprehensive Assessment of At-Risk Mental States. Aust N Z J Psychiatry (2005) 39:964–71. doi: 10.1080/j.1440-1614.2005.01714.x
25. Krebs MO, Magaud E, Willard D, Elkhazen C, Chauchot F, Gut A, et al. Évaluation des états mentaux à risque de transition psychotique : validation de la version française de la CAARMS. L’Encéphale (2014) 40:447–56. doi: 10.1016/j.encep.2013.12.003
26. McGorry PD, Purcell R, Hickie IB, Yung AR, Pantelis C, Jackson HJ. Clinical staging: a heuristic model for psychiatry and youth mental health. Med J Aust (2007) 187(S7):S40–2. doi: 10.5694/j.1326-5377.2007.tb01335.x
27. Dressendörfer RA, Kirschbaum C, Rohde W, Stahl F, Strasburger CJ. Synthesis of a Cortisol-Biotin Conjugate and Evaluation as a Tracer in an Immunoassay for Salivary Cortisol Measurement. J Steroid Biochem Mol Biol (1992) 43:683–92. doi: 10.1016/0960-0760(92)90294-S
28. Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2–ΔΔCT Method. Methods (2001) 25:402–8. doi: 10.1006/meth.2001.1262
29. Hruz T, Wyss M, Docquier M, Pfaffl MW, Masanetz S, Borghi L, et al. RefGenes: identification of reliable and condition specific reference genes for RT-qPCR data normalization. BMC Genomics (2011) 12:156. doi: 10.1186/1471-2164-12-156
30. Chang CC, Chow CC, Tellier LCAM, Vattikuti S, Purcell SM, Lee JJ. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. GigaScience (2015) 4:7. doi: 10.1186/s13742-015-0047-8
31. Trakadis YJ, Fulginiti V, Walterfang M. Inborn errors of metabolism associated with psychosis: literature review and case-control study using exome data from 5090 adult individuals. J Inherit Metab Dis (2018) 41:613–21. doi: 10.1007/s10545-017-0023-9
32. Swinehart WE, Jackman JE. Diversity in Mechanism and Function of TRNA Methyltransferases. RNA Biol (2015) 12:398–411. doi: 10.1080/15476286.2015.1008358
33. Blanco S, Dietmann S, Flores JV, Hussain S, Kutter C, Humphreys P, et al. Aberrant Methylation of t RNA s Links Cellular Stress to Neuro-developmental Disorders. EMBO J (2014) 33:2020–39. doi: 10.15252/embj.201489282
34. Kozuka T, Omori Y, Watanabe S, Tarusawa E, Yamamoto H, Chaya T, et al. MiR-124 Dosage Regulates Prefrontal Cortex Function by Dopaminergic Modulation. Sci Rep (2019) 9:3445. doi: 10.1038/s41598-019-38910-2
35. Bandres-Ciga S, Noyce AJ, Traynor BJ. Mendelian Randomization—A Journey From Obscurity to Center Stage With a Few Potholes Along the Way. JAMA Neurol (2019) 77(1):7–8. doi: 10.1001/jamaneurol.2019.3419
36. Palma-Gudiel H, Córdova-Palomera A, Tornador C, Falcón C, Bargalló N, Deco G, et al. Increased Methylation at an Unexplored Glucocorticoid Responsive Element within Exon 1 D of NR3C1 Gene Is Related to Anxious-Depressive Disorders and Decreased Hippocampal Connectivity. Eur Neuropsychopharmacol (2018) 28:579–88. doi: 10.1016/j.euroneuro.2018.03.015
Keywords: ultra-high risk of psychosis, stress, cortisol, hypothalamic–pituitary–adrenal axis, Mendelian randomization, expression quantitative trait locus, genome-wide analysis study
Citation: Iftimovici A, Kebir O, He Q, Jay TM, ICAAR Study Group , Rouleau GA, Krebs M-O and Chaumette B (2020) Stress, Cortisol and NR3C1 in At-Risk Individuals for Psychosis: A Mendelian Randomization Study. Front. Psychiatry 11:680. doi: 10.3389/fpsyt.2020.00680
Received: 01 May 2020; Accepted: 29 June 2020;
Published: 10 July 2020.
Edited by:
Jerome Brunelin, INSERM U1028 Centre de Recherche en Neurosciences de Lyon, FranceReviewed by:
Romain Rey, Centre Hospitalier Le Vinatier, FranceCopyright © 2020 Iftimovici, Kebir, He, Jay, ICAAR Study Group, Rouleau, Krebs and Chaumette. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Boris Chaumette, Ym9yaXMuY2hhdW1ldHRlQGluc2VybS5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.