
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 05 March 2020
Sec. Plant Systematics and Evolution
Volume 11 - 2020 | https://doi.org/10.3389/fpls.2020.00127
This article is part of the Research TopicOrigins and Domestication of the GrapeView all 8 articles
 Jorge Cunha1,2*
Jorge Cunha1,2* Javier Ibáñez3
Javier Ibáñez3 Margarida Teixeira-Santos1
Margarida Teixeira-Santos1 João Brazão1,2
João Brazão1,2 Pedro Fevereiro2,4
Pedro Fevereiro2,4 José M. Martínez-Zapater3
José M. Martínez-Zapater3 José E. Eiras‐Dias1,2
José E. Eiras‐Dias1,2The domesticated grapevine spread along the Mediterranean basin from the primary Near East domestication area, where the greatest genetic diversity is found in its ancestor, the wild vine populations. Portuguese wild populations are on the southwestern fringe of the distribution of the Vitis vinifera L. ssp. sylvestris (C.C. Gmel.) Hegi in Europe. During the last Glacial Period they became isolated from the previous continuum that had been the territory of wild vine populations. Archaeological remains of domesticated vinifera grapevines in Portugal date back from 795 Before Common Era (BCE) in the lower Tagus river basin. In this work, 258 Portuguese vinifera varieties and sylvestris plants were characterized using 261 single nucleotide polymorphism (SNP) markers. The study of the genetic diversity of this local germplasm, its population structure and kinship, all framed in their historical and geographical backgrounds, revealed a complex network of first-degree relationships, where only Iberian varieties are involved. Some Iberian genotypes, like Alfrocheiro (Bruñal, in Spain), Sarigo (Cayetana Blanca), Mourisco Branco (Hebén), Amaral (Caiño Bravo), and Marufo (Moravia Dulce) are ancestors of a considerable fraction of all the autochthonous analyzed varieties. A part of the diversity developed was mostly local in some cases as shown by the closeness of several varieties (Vinhos Verdes) to the wild cluster in different analyses. Besides, several evidences of introgression of domesticated germplasm into wild vines was found, substantiating the high risk of genetic contamination of the sylvestris subspecies. All these findings together to the known matching between the wild maternal lineage of the Iberian Peninsula and an important number of Portuguese grapevine varieties (chlorotype A), point out that some of these varieties derive, directly or indirectly, from originally local wild populations, supporting the possible occurrence of secondary events of local domestication, or, at least, of an introgression process of wild into cultivated grapevines.
The domesticated grapevine (Vitis vinifera L. ssp. vinifera) is the most cultivated fruit crop of the Vitis genus, which contains about 60 inter-fertile wild species (This et al., 2006; Emanuelli et al., 2013). V. vinifera ssp. sylvestris is the only wild Vitis taxon native to Europe and the Near East, and it is believed to be the wild progenitor of almost 10,000 domesticated grapevine varieties today (This et al., 2006; Emanuelli et al., 2013). Genetic hybridizations among wild, cultivated and feral types make it difficult to untangle the history of current grapevine varieties (Zohary and Hopf, 1994).
Domestication results from selecting and propagating plants with desirable traits along generations. Given the dioecious behavior of the wild species, and its high heterozygosity, vegetative propagation is believed to have been adopted early in grapevine domestication (Zohary and Hopf, 1994), as the easiest way to maintain desirable traits. The domestication of grapevine generated notable morphological changes, including perfect hermaphrodite flowers, higher sugar content, and larger berry size together with others like altered seed morphology resulting from automatic selection (Olmo, 1995; This et al., 2006; Miller, 2008). For instance, since self-pollinating plants have improved fruit set and are more productive (Miller, 2008), hermaphrodite variant vines were selected compared to female plants. Domesticated grapevines were disseminated along with the spreading of wine culture from their primary domestication sites in the Near East in a process that lasted over 5000 years (McGovern et al., 2017) but likely a reduced number of sexual generations, given their vegetative multiplication (Arroyo-García et al., 2006). Along this time, introgression from local populations or even secondary domestication events could have taken place as suggested by Arroyo-García et al. (2006); Imazio et al. (2006); or Myles et al. (2011).
Relictic wild vine populations can still be found in Portugal in what can be considered as the southwestern fringe of the distribution of the V. vinifera ssp. sylvestris in Europe. During the late Ice Age, Iberian Vitis populations became isolated from the previous continuum that had been the territory of Mediterranean populations. Palynological evidences from Southern Portugal show the presence of pollen from V. vinifera in several river basins and lagoons in the Holocene (Leeuwaarden and Janssen, 1985; van der Knaap and van Leeuwen, 1995; Fletcher et al., 2007; Vis and Kasse, 2009; Schneider et al., 2016). Evidences of strong anthropogenic effects on the environment registered on the vegetation of this region after 2090 Before Common Era (BCE) suggest the possibility of domestication/cultivation of grapevine (Fletcher et al., 2007).
The oldest archeological remains of grapevine seeds in Portugal date back from the Chalcolithic (circa 3350–2250 BCE) and its presence continues until the Roman time, covering the Bronze and Iron Age and the Phoenician settlements (Ramil Rego and Aira Rodríguez, 1993; Aira Rodríguez and Ramil Rego, 1995; Arruda, 2008). Stummer (1911) introduced an index (width to length ratio) for the distinction between wild and cultivated grapevine seeds. Indexes between 0.76 and 0.83 indicate wild plants, while indices between 0.44 and 0.53 correspond to cultivated plants; values between 0.54 and 0.75 correspond to margins of overlap of the two subspecies. This index has been the main criterion used in the identification of archeological seeds (Renfrew, 1973).
In the Iberian Peninsula the production of wine is well establish since the 7th century BCE (Buxó, 2008). In Portugal, the production and use of grapes in a Phoenician urban context is established by the presence of grapevine seeds in Almada (near Lisbon), an archeological site from the 7th century BCE (Barros, 1998). Ceramic remains containing wine were found in a Phoenician archeological context in Santarem, Almada, and Lisbon (Barros, 1998; Sousa and Guerra, 2018). The 2nd century BCE the Greek author Polybius in his The Histories (book 34 chapter 8) refers to the low price of wine in Lusitania (“A metreta of wine costs a drachma”—circa 40 liters cost the day payment of a soldier). In the 1st century BCE the also Greek author Strabo in his Geography, book 3 chapter 3, refers to vineyards in the lower Tagus valley and in chapter 2 to wine exports from Tudertania. Roman remains (1st century BCE to 5th century CE) are ubiquitous, including grape seeds remains in archeological sites, oenological equipment and amphorae used to storage and transport of wine (Fabião, 1998; Tereso et al., 2011; Savo et al., 2016). Viticulture was not abandoned after the defeat of the Visigoths kingdom by Tariq (711 CE), and later in the Middle Ages, viticulture spread throughout the country likely related to the expansion of Christian kingdoms and also accompanied by the establishment of several monastic orders (Benedictines and Cistercians) with strong viticulture traditions (Fernandes, 1532; Martinez Tomé, 1991).
Ampelography (Aµπϵλος, “vine” and γραφος, “writing”) is the science of identifying, naming, and classifying grape varieties, mainly through its phenotypic characterization, and ampelographic data is the base of the description and identification of grapevines (Galet, 1979). Well-established ampelographic descriptors from the International Organization of Vine and Wine (OIV, 1983; OIV, 2009) identified numerous synonym and homonym varieties. However, the subjectivity of individual observations, as well as the variability generated by plant growing conditions, sanitary status and growing season, prevented unambiguous identifications. Single nucleotide polymorphisms (SNP) are one of the most powerful DNA markers recently developed. They have been adapted to a large number of applications in the study of grapevine, including genetic identification (Cabezas et al., 2011; Cunha et al., 2016), diversity studies, genetic structure, and domestication history of grape (Myles et al., 2011), or pedigree and phylogeny studies (Ibáñez et al., 2012; Zinelabidine et al., 2012). SNPs are alternative or complementary to other markers often used in grapevine, like nuclear microsatellites (nuclear simple sequence repeats SSR), or chloroplast microsatellites (cpSSR) (Bourquin et al., 1993; Sefc et al., 2000; Arroyo-García et al., 2006).
Chloroplasts are maternally inherited in grapevine, and the analysis of polymorphisms in its genome permits to follow the maternal lineage of any vine. The analyses of chloroplast microsatellites in a large sample of wild and cultivated grapevines along the Mediterranean area allowed to identify four major chlorotypes (A, B, C, and D) with a differential geographic distribution among the wild Vitis populations analyzed (Arroyo-García et al., 2006). Chlorotypes B, C, and D were detected in Near and Middle Eastern populations, while mostly A chlorotype was found in Western Mediterranean populations, including Iberian populations. Interestingly, over 75% of Iberian (Spanish and Portuguese) varieties contain the A chlorotype, (Arroyo-García et al., 2006; Cunha et al., 2009; Castro et al., 2013), coincident with the chlorotype detected in their natural sylvestris populations. In addition, varieties bearing chlorotypes B, C, and D, found more frequently in Eastern wine varieties (B and D) and table grape varieties (C) (Arroyo-García et al., 2006; Laucou et al., 2018) were also found in Portuguese varieties (Cunha et al., 2009; Cunha et al., 2010; Castro et al., 2013; Cunha et al., 2015) supporting their multiple origins, likely including introductions from the Near East and from the Maghreb. Secondary domestication events have been proposed for grapevine along the Mediterranean basin based on differences in chlorotype frequencies (Grassi et al., 2003; Arroyo-García et al., 2006) or on the increased expected heterozygosity values detected further from the putative center of origin of the species (De Lorenzis et al., 2019).
Several genetic studies based on molecular markers have been carried out with Portuguese grapevines, proving to be powerful tools for cultivar identification, and resulting in the discovering of many cases of synonyms, homonyms and misnames. The existence of genetic relationships within cultivated and wild plants was surveyed, as well as pedigrees between autochthonous and foreign cultivars (Lopes et al., 1999; Lopes et al., 2006; Almandanim et al., 2007; Cunha et al., 2007; Cunha et al., 2009; Cunha et al., 2010; Veloso et al., 2010; Castro et al., 2011; Castro et al., 2013; Cunha et al., 2015; Cunha et al., 2016; Moita Maçanita et al., 2018). In a previous study, we genotyped 288 Portuguese grapevine accessions using a set of 48 SNP markers to detect synonyms, homonyms, and mistakes within the national germplasm collection (Cunha et al., 2016). Here we have increased the information on those accessions by genotyping them up to 261 SNP markers to carry out deeper genetic analyses aiming to: i) evaluate the genetic diversity and genetic structure of Portuguese germplasm; ii) identify first-degree genetic relationships using the SNP database of the Instituto de Ciencias de la Vid y del Vino (ICVV), which includes many varieties and wild plants; and iii) combine this information with chloroplast genotype information to help integrating historical data.
The Portuguese plant material used in this work was collected in the Portuguese National Ampelographic Collection (PRT051), which is the reference collection for varieties allowed in Portugal for wine production (MAMAOT, 2012). It is hosted by the Instituto Nacional de Investigação Agrária e Veterinária (INIAV) within the Portuguese Ministry of Agriculture, Forestry, and Rural Development. This collection was established in 1988 and it is located at Quinta da Almoinha, Dois Portos, Torres Vedras, Portugal [39°02′34.03″N, −9°10′57.41″W]. All the collection is grafted onto SO4 (Selection Oppenheim 4) rootstock and the training system is bilateral cordon (Royat). Two hundred and thirty-one accessions of V. vinifera L. ssp. vinifera (legally authorized to produce wine in Portugal) and 27 of V. vinifera L. ssp. sylvestris (C.C. Gmel.) Hegi [collected from six populations (Cunha et al., 2016) and maintained in the collection] were used (Supplementary Table 1). These samples were previously characterized using a set of 48 SNPs (Cunha et al., 2016) as well as with agronomical, morphological, and microsatellite markers (six and nine microsatellites) recommended by OIV (Veloso et al., 2010; Eiras-Dias et al., 2011; Eiras-Dias et al., 2013). The locations of the Portuguese National Ampelographic Collection and of the wild vine populations are shown in Figure 1. The other material used is the SNP database from the ICVV that includes genotypes from several different sources, mainly from the ICVV grapevine collection in La Grajera (ESP217, Logroño) and the Vitis Germplasm Bank (VGB) from the Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA) in El Encín (ESP080, Alcalá de Henares). In addition, it also contains genotypes from several origins and collections: Algeria, Argentina, Australia, Belgium, Chile, France, Iran, Italy, Montenegro, Morocco, Portugal, Romania, Spain, and Tunisia.
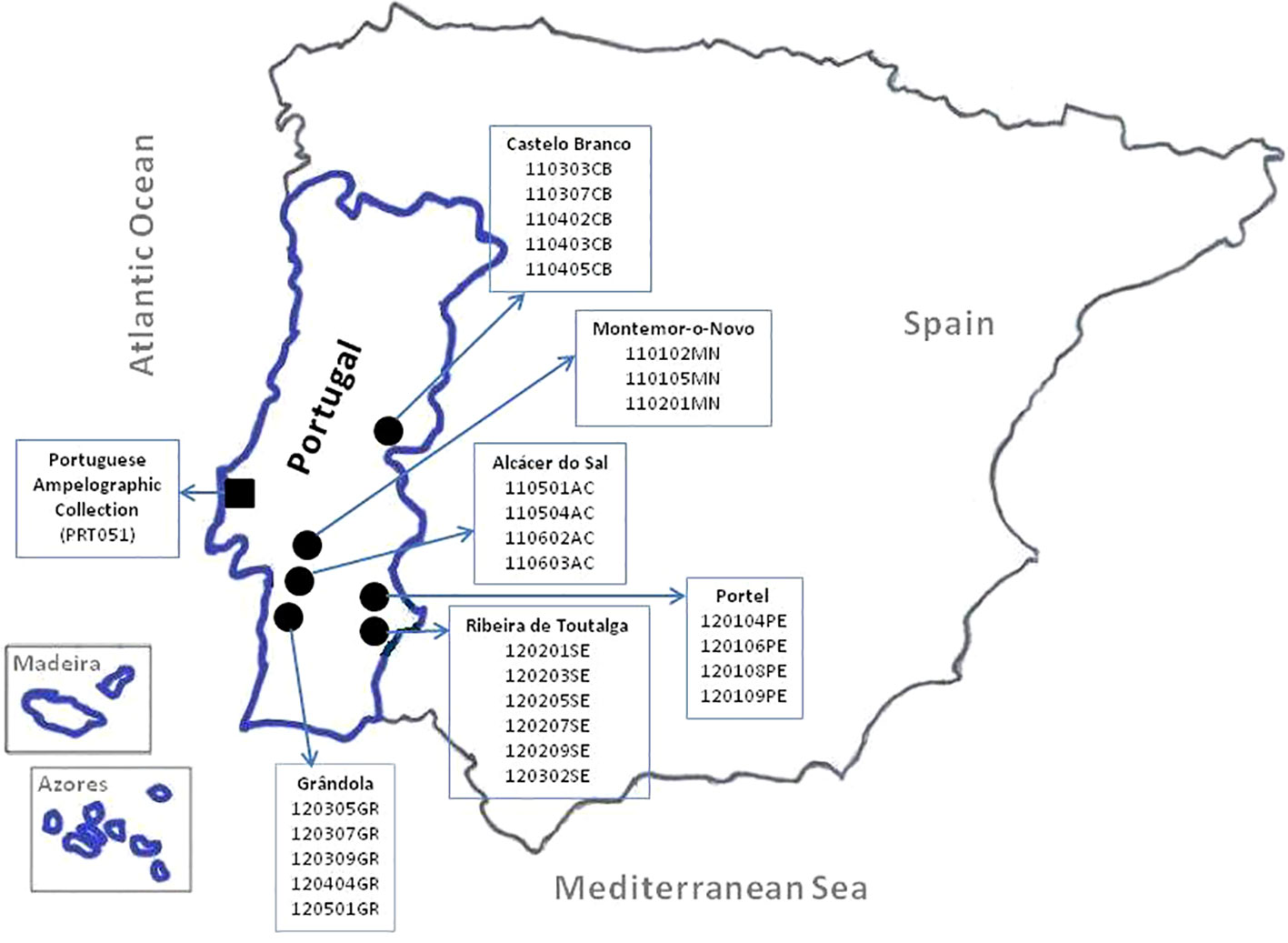
Figure 1 Map of Portugal with the locations of National Ampelographic Collection (◼) and wild vine populations in situ (●).
DNA was isolated from young leaves frozen at −80°C according to Thomas et al. (1993), with minor modifications. The quality and concentration of the DNA were determined by electrophoresis in agarose (0.8%) gels stained with ethidium bromide and visualized on a UV transilluminator. Concentration was calculated by comparing with known DNA concentrations (50, 100, and 200 ng/μl) of λDNA HindIII Fragments, 0.1 μg/μl, Invitrogen, Carlsbad, CA USA). NanoDrop 2000 C UV-Vis spectrophotometer (Thermo Scientific, Waltham, MA, USA) was used to check the quality and final concentration of each DNA sample.
DNA from grapevines was genotyped with 261 nuclear SNPs obtained by Lijavetzky et al. (2007) and Cabezas et al. (2011) (Supplementary Table 2). SNP genotyping was performed using SNPlex (Applied Biosystems, Waltham, MA USA) or Veracode (Illumina, San Diego, CA, USA) technologies as described in Zinelabidine et al. (2012; 2015). Genotyping services were provided in Spain by the National Genotyping Center1.
GeneAlEx 6.5 (Peakall and Smouse, 2012) was used to calculate the following genetic parameters: observed heterozygosity, expected heterozygosity, Shannon's information index, and hierarchical F-statistics. Calculations were performed on wild and cultivated groups separately using only polymorphic SNP markers in each group. The mean values of each parameter were analyzed using t-Student test (GraphPad Prism version 7.0, San Diego, CA, USA) to verify the statistical significance of the mean differences between the two groups. A p-value of <0.05 was considered statistically significant. Rarefaction on measures of allelic richness was performed to address the unbalanced sample size of wild and cultivated sets, using the program HP-RARE 1.0 (Kalinowski, 2005). GeneAlEx 6.5 was also used to test for deviation from the Hardy-Weinberg equilibrium (HWE) across all loci for each population.
Pairwise Euclidean distance was calculated for every pair of accessions using the genetic distance to assess the relationship among the wild and cultivated grapevine accessions, using the program GenAlEx 6.5.
MEGA 7 software, version 7.0.26 (Kumar et al., 2016), was used to generate a distance tree by the neighbor-joining (N-J) hierarchical clustering method (Saitou and Nei, 1987) based on the Pairwise Euclidean distance generated from the genetic distance obtained in GeneAlEx 6.5 software. Principal Coordinate analysis (PCoA) was performed on individual multilocus genotypes, with covariance standardized, using the same program.
Structure 2.3.4 (Pritchard et al., 2000) using the admixture model was employed to infer the number of genetic populations (K) existing in the sample and to assign individuals to their likely population of origin, with no prior information. A series of 10 independent runs was performed for each value of K between 1 and 15. An initial burn-in of 20,000 steps was used to minimize the effect of the starting configuration, followed by 100,000 Markov chain Monte Carlo steps, as recommended by Falush et al. (2007) and Ghaffari et al. (2014).
The software CERVUS 3.0 (Kalinowski et al., 2007), that uses a likelihood-based method for computation, was used to identify first-order kinship relationships: trios (mother-father-offspring) and duos (parent-offspring pairs) among the studied (wild and cultivated) accessions. Pedigree analysis was done using as candidate parents a total of 1,921 profiles of different origins, including those obtained here, present in the ICVV-SNP database. Natural logarithm of the overall likelihood ratio—logarithm-of-odds (LOD) score—was calculated for each trio and duo identified. Chlorotypes previously determined for the Portuguese genotypes (Cunha et al., 2009; Castro et al., 2013) were used whenever possible to determine the maternal progenitor in the trio (Arroyo-García et al., 2006).
A total of 258 accessions (27 wild vines and 231 grapevine varieties) from the Portuguese Ampelographic Collection bearing non-redundant genotypes when analyzed with 48 SNPs (Cunha et al., 2016) were genotyped with up to 261 SNP markers. Thirty of these SNP markers were discarded: 28 because data were missing in 66% of the samples and two because genotyping failed in all samples. The results of the remaining 231 SNP markers were used for subsequent analyses (Supplementary Table 1).
The analyses of the genetic diversity of the Portuguese wild vines and grapevine varieties are presented in Table 1. Overall the genetic diversity parameters showed higher values in the cultivated group than in the wild set. In all cases these differences were statistically significant (P < 0.05) (Table 1). The estimated fixation index F (also called inbreeding coefficient FIS) in the wild genotypes showed a positive value of 0.109 ± 0.017, indicating a heterozygote deficiency in the wild group. In grapevine varieties, a mean negative value (−0.030 ± 0.008) of index FIS was found, showing higher heterozygosity than in the wild samples. The analysis of allelic richness and private allelic richness using rarefaction methods, which take into account the differences in sample sizes, also showed a larger diversity in the cultivated set (Table 1).
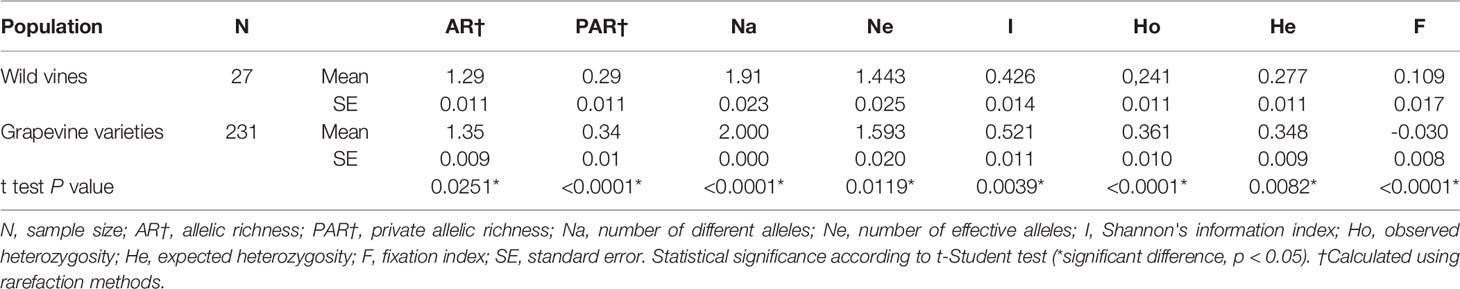
Table 1 Summary of genetic diversity parameters estimated for Vitis vinifera sylvestris (wild vines) and vinifera (grapevine varieties) from Portugal.
Furthermore, the FST statistic of overall SNP loci was used to analyze the genetic differentiation among wild and cultivated subspecies and was estimated to be 0.158 when the total of the 258 accessions were used for the calculation.
Deviation from the Hardy-Weinberg proportion (P < 0.05) was observed for 54 (23.38%) markers in the cultivated group and for 32 (13.85%) markers in the wild set.
A neighbor-joining (N-J) distance tree was constructed to investigate the genetic relationship among the 258 non-redundant genotypes from the 231 SNP matrix data (Figure 2). The hierarchical clustering of 258 unique genotypes produced three clusters (I, II, III). Cluster I mostly includes ancient Portuguese varieties and ancient Western and Central European varieties introduced and cultivated in Portugal time ago (e.g., Bastardo/Trousseau Noir, #12668; Branco Valente/Heunisch Weiss, #5374). Exceptions in cluster I are two wild vines, a male plant (120209SE) and a female plant (120302SE), from the population of Ribeira de Toutalga (Guadiana river basin, Figure 1). Cluster III groups mainly Portuguese varieties, including most of Marufo descendants and Bastardo descendants (see below). Apart from Fernão Pires, the most spread white variety in Portugal, most of the varieties included in this cluster are minority varieties. Cluster II includes four subgroups: subcluster IIa, with varieties obtained by the breeder Leão Ferreira de Almeida in the middle of 20th century; subcluster IIb, mostly with varieties from the Vinhos Verdes wine area (Northwestern Portugal); subcluster IIc, with almost all wild vines, and subcluster IId, with ancient varieties. Subcluster IIa also includes one wild vine genotype (110603AC) from the population of Alcácer do Sal (Sado river basin). In the subcluster IIc three varieties Barcelo, #980; Branjo, #17661 and Melhorio, #17225 cluster together with most wild vines. Two of these varieties, Barcelo and Melhorio, form duos, and thus are closely related, with Amaral, #818 (Supplementary Table 3). Amaral had already been mentioned in 1532 in the North of Portugal (Fernandes, 1532), while Barcelo was cited in the 18th century by Lacerda Lobo (1790).
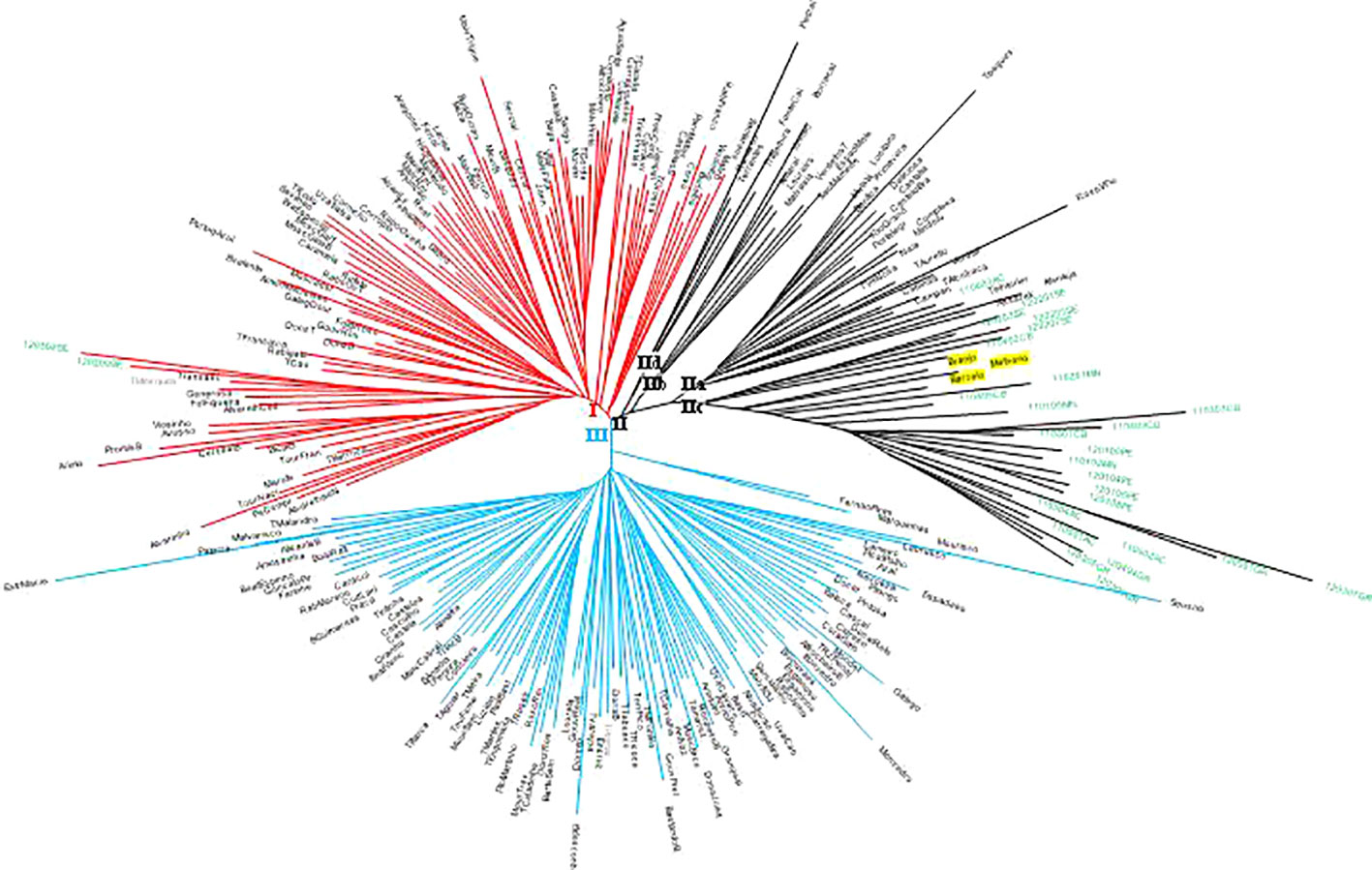
Figure 2 Neighbor-joining radiation tree showing genetic distance among Portuguese Vitis vinifera wild vines and varieties genotypes, based on 231 single nucleotide polymorphism (SNP) loci. Cluster I, solid lines in red color; cluster II, solid lines in black color; cluster III, solid lines in blue color; wild vine codes in green font; variety names in black font; cultivated grapevines with the wild group are highlighted in a yellow background.
A non-hierarchical PCoA based on the square distances' matrix was also used to analyze the relationships between wild and cultivated grapevines as revealed by SNP markers (Figure 3). The first two principal axes explain only 16.43% of the total variation (9.46 and 6.97%, respectively). PCoA provides a similar result to the N-J distance analysis, separating the grapevine varieties in two groups (A, C) and a third group (B) with wild vines and cultivated genotypes, although this last group is not as clearly separated. Accessions from wild vine populations 120207SE, 110603AC, and 110402CB are in the edge of one of the clusters of grapevine varieties, while Barcelo is the closest variety to the wild vines. Amaral, Branjo, and Melhorio are also in the vicinity of the wild plants.
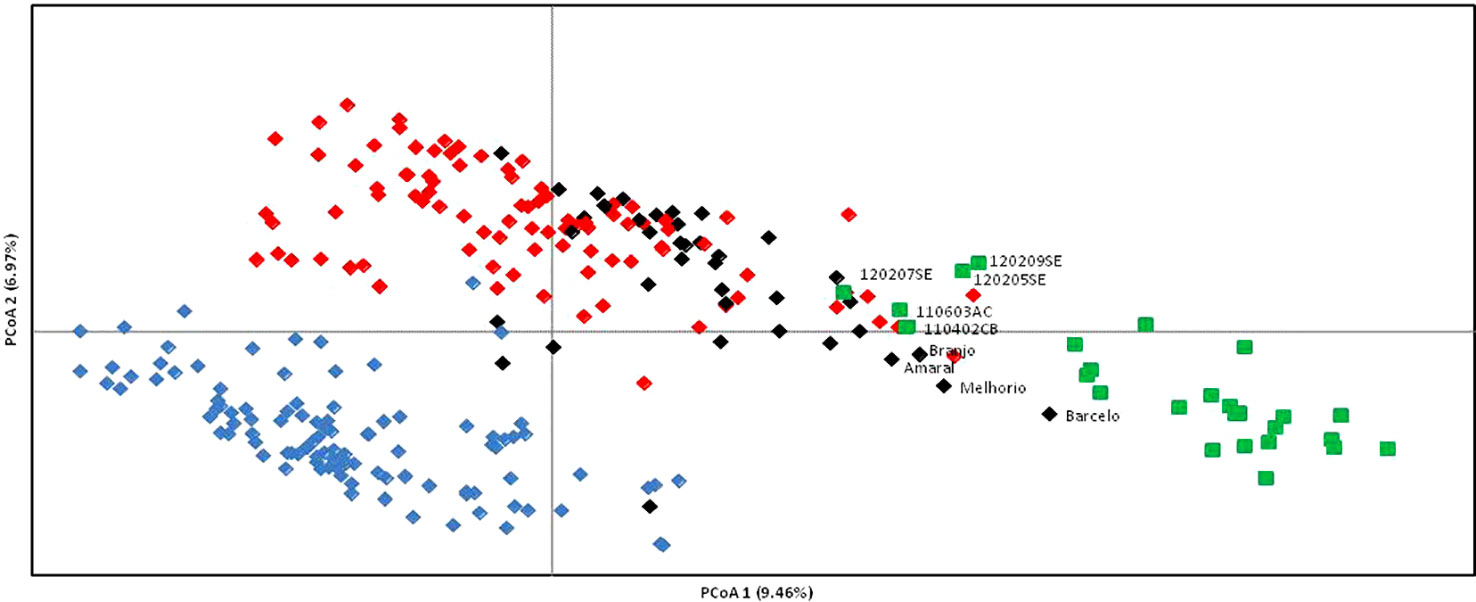
Figure 3 Plot of Portuguese Vitis vinifera, wild vines (green squares), and grapevine varieties (black, blue, and red diamonds) from a Principal Coordinate Analysis based on 231 single nucleotide polymorphism (SNP) markers and via covariance matrix with data standardization. Only wild and cultivated grapevines in intermediate positions are labeled. To facilitate the comparison with Figure 2 the varieties are colored according to their position in Figure 2 (red, black and blue for cluster I, II, and III respectively).
The existence of different number of genetic groups was also explored using Structure, from K = 1 to K = 15. Results for K = 2 and K = 3 were selected following Evanno et al. (2005) criterion and are displayed in Figure 4. In both cases 25 of the 27 wild plants were assigned to one of the subpopulations obtained (Pop2 and Pop3 respectively, Supplementary Table 4). The two accessions of wild vines that were not assigned to those groups were 120207SE and 110603AC. In the first scenario (K = 2), 39 out of the 231 grapevine varieties (17%) were assigned to Pop2, the “wild” subpopulation, including Amaral, Barcelo, Alvarinho (all with membership coefficients above 0.75), Branjo and Melhorio, and a few non-local varieties (Espadeiro Mole/Manseng Noir, #7340; Mondet/Durif, #3738; Santareno/Etraire de la Dui, #3993; Sevilhão/Corbeau, #2826 and Uva Salsa/Chasselas Cioutat, #2476). In the second scenario (K = 3), the subpopulation including most of the wild plants (Pop3) also included 12 cultivated genotypes, 11 of them Portuguese varieties such as the mentioned Amaral, Barcelo, and Alvarinho. The two other inferred subpopulations in K = 3 contain 54 and 41.6% of the grapevine varieties. Even though they do not match exactly, the groups assigned by structure (K = 3) mostly concord with the previously shown analyses of the N-J tree (Figure 2), and PCoA (Figure 3).
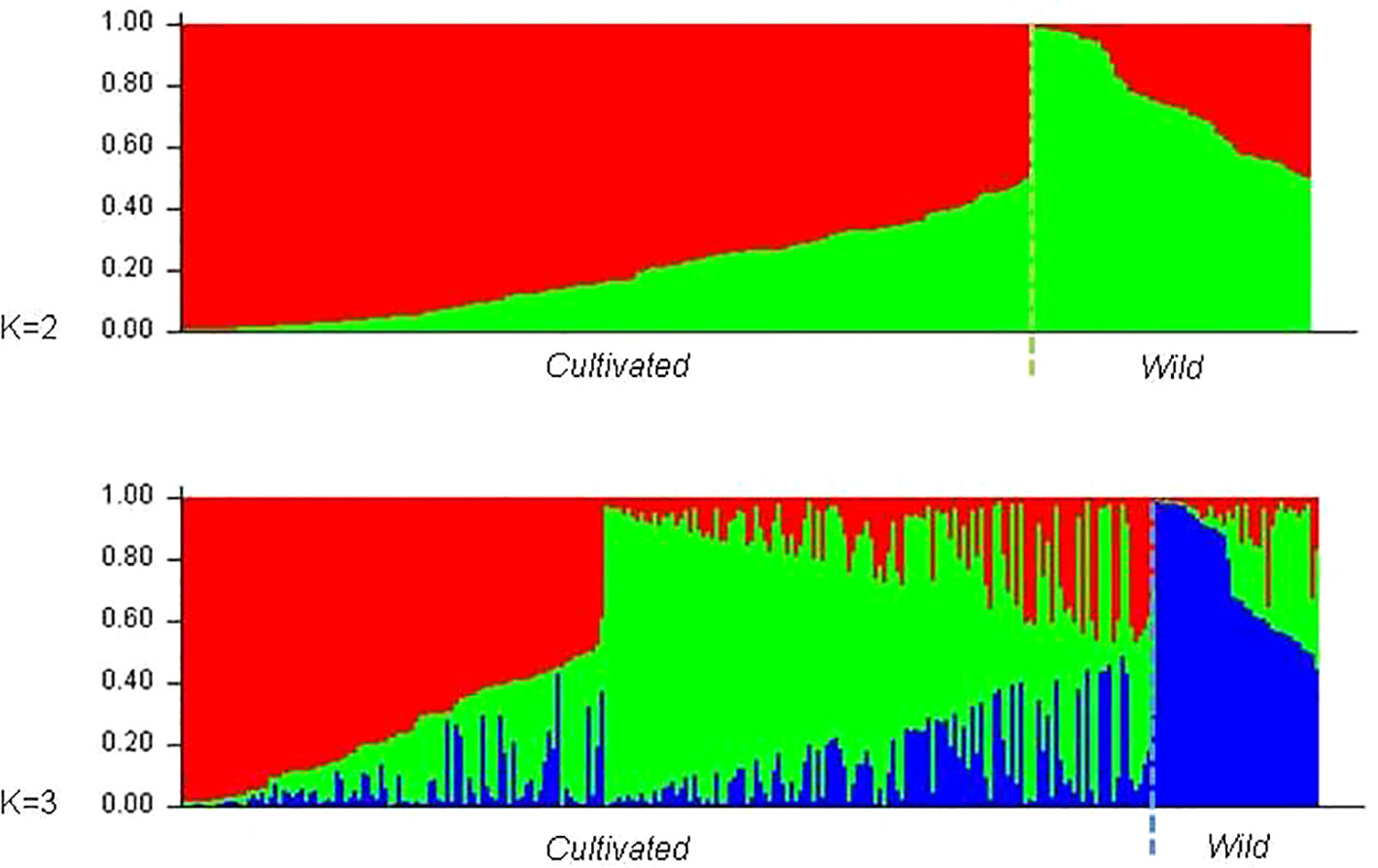
Figure 4 Barplot of the genetic population admixture in wild vines and cultivated varieties, as inferred by structure at K = 2 and K = 3. Each individual is represented by a single vertical bar broken into K color segments, with lengths proportional to the estimated probability of membership in each inferred cluster.
The data from 231 different varieties and 27 wild unique genotypes was added to the ICVV-SNP database, which was raised to 1,921 non-redundant genotypes, originated from the Near-East to Western European countries. A search for the possible first-order kinship relationships for these 258 non-redundant Portuguese genotypes was performed using all the 261 SNPs. One hundred and two trios (both parents and offspring) were identified (Supplementary Table 5). Thirty-two new trios are reported here for the first time, and all are supported by high LOD scores, that ranged from 52.70 to 80.70 (Figure 5A and Supplementary Table 5). Fifteen trios from the crosses made by the breeder Almeida (Ghira et al., 1982) were verified and all are supported by high LOD scores, that ranged from 61.5 to 85.3. Four of these bred varieties were the result of crosses between Castelão, #2324 and Alicante Bouschet, #304 (Figure 5B and Supplementary Table 5). Up to 55 trios previously described by other authors were confirmed (trios and references in Supplementary Table 5). Several hundred compatible duos (parent-offspring relationships) were also identified but, given the existence of large and close families among the studied accessions, it is difficult to conclude which ones correspond to real parent-offspring relationships. Only those considered more relevant are reported and discussed below (Supplementary Table 3).
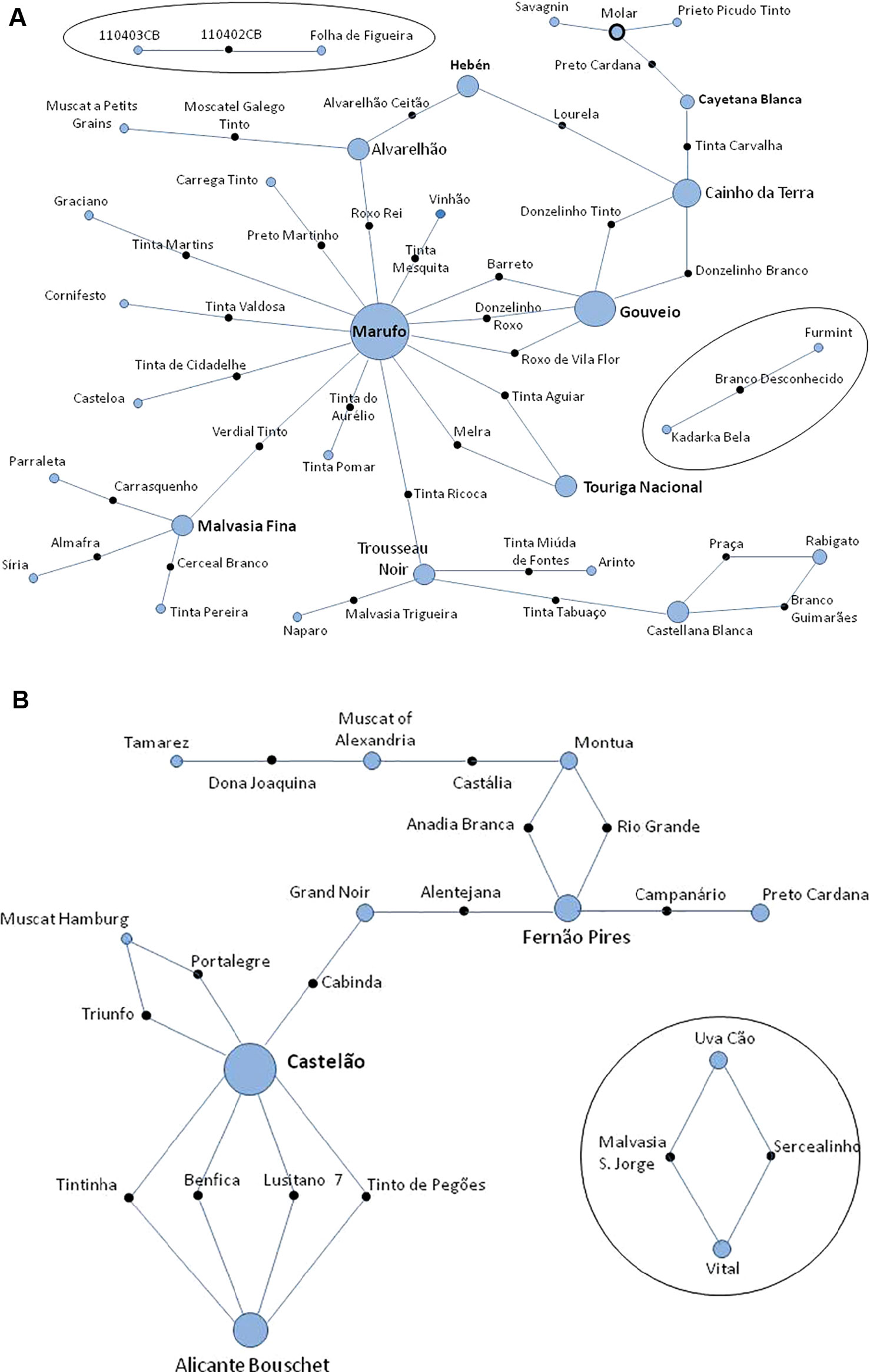
Figure 5 Networks of the proposed trios (parents and offspring) found in a parentage analysis of Portuguese set of Vitis vinifera germplasm using 261 single nucleotide polymorphism (SNP) markers. (A) Thirty-two previously unknown genetic relationships of varieties and/or wild plants; (B) confirmation of 15 pedigrees of new varieties reported by Almeida breeder in 1950. Solid black circles represent the offspring; blue circles represent the progenitors; circle diameter is proportional to the number of relationships where is involved; blue circles surrounded by a solid black line represent a variety that is an offspring and parent in the figure. The tree is not drawn to scale.
For 65% of the Portuguese genotypes none compatible trio were found, i.e., their two parents could not be identified. A number of varieties were found to have played an important role in the genetic network of the studied grapevine varieties. Together with Alfrocheiro, #277; Mourisco Branco/Hebén, #5335 and Sarigo/Cayetana Blanca, #5648, previously described (Zinelabidine et al., 2012; Zinelabidine et al., 2015; Cunha et al., 2015), Marufo, #8086, a female grapevine variety bearing chlorotype (chl) D, was found to be a parent (mother) in 18 trios (14 identified for the first time in this study) (Figure 5A, Supplementary Table 5). Gouveio, #12953 (chl A), an old variety from Douro wine region, is involved as a parent in five new trios. Bastardo/Trousseau Noir, #12668 (chl A), Cainho da Terra (chl D), and Malvasia Fina, #715 (chl A) are parents in four new trios (Figure 5A and Supplementary Table 5). Alvarelhão, #1650 (chl A) and Castellana Blanca, #26280 (chl A) participated in three new trios. Touriga Nacional, #12594 (chl A); Sarigo (chl A); Mourisco Branco (chl A); Rabigato, #9857 (chl A); Uva Cão, #12812 (chl A) and Vital, #13122 (chl A) are each involved in two previously unknown trios (Figure 5 and Supplementary Table 5).
A trio was found indicating gene flow from cultivated to wild population. The wild accession 110402CB, is a descendant of a cross between Folha de Figueira, a hermaphrodite variety from Castelo Branco region and 110403CB, a female wild plant (Figure 5A). The reliability of the trio is high as the LOD score was 69.80. The flowers of the accession 110402CB show a fully developed gynoecium and straight stamens shorter than the gynoecium (IPGRI, descriptor 6.2.1, note 4: female with straight stamens) in opposition to totally female plants that have reflexed stamens (IPGRI, descriptor 6.2.1, note 5). Folha de Figueira (VIVC #14142), is a white variety, which was first described by Lacerda Lobo in 1790. These wild plants were found in the margins of Ponsul river, a Northern tributary of the Tagus river, flowing close to the border with Spain in the Castelo Branco district.
Three consistent duos (0–1 mismatches, LOD > 25) involving wild vines were also identified. One duo is made up by the plants 110504AC and 110602AC from the Alcácer do Sal population, both males with different chloroplast haplotypes (A and B, respectively). The two other duos also involved cultivated varieties: one in the same Pônsul population between a female plant 110405AC and Frankenthal/Schiava Grossa, #10823 and the other in the Alcácer population between 110603AC and Castelão, #2324. Schiava Grossa is a spread variety from Italy, with 163 synonyms in the VIVC database (Maul et al., 2019). The oldest known reference in Portugal dates back from 1887, in the collection of Quinta da Viscondessa in Torres Vedras (Anonymous, 1887). Castelão is one of the most cultivated varieties in the south of Portugal (Cunha et al., 2015). Castelão wines are already mentioned in legal documents from the 14th century (Amaral, 1994).
The uneven sample sizes of the cultivated and wild sets of genotypes studied makes difficult the comparison of the genetic diversity parameters between them. Nevertheless, the results obtained, including those considering the sample sizes, indicate what other previous works have pointed out: the existence of a reduced diversity in the sylvestris subspecies (Marrano et al., 2017; Marrano et al., 2018). This unexpected situation (sylvestris subspecies is obligate outcrossing while most cultivars are self-pollinating) is caused by the small sizes commonly found in wild populations, due to their isolation by natural barriers, human actions, and the severe bottleneck that began in the 19th century with the pathogens introduced from North America (powdery mildew, downy mildew, and phylloxera) that has converted in relict the surviving populations.
Taken together, all the genetic analyses performed on the Portuguese grapevine germplasm based on SNP genotypes indicate the existence of three major clusters or genetic groups. Genetic group I and III contain mostly cultivated genotypes while genetic group II includes most wild vines as well as cultivated genotypes from the region of Vinhos Verdes. Genetic group I includes cluster I defined in the N-J distance analysis (Figure 2), corresponding to PCoA group A (Figure 3) and cluster 2 in the structure analysis at K3 (green color in Figure 4). It grouped the ancient Portuguese varieties, with unknown parentage, together with foreign varieties contributing to the Portuguese germplasm either from Western and Central Europe, like Bastardo (BastT/Trousseau Noir); or Northern Africa, like Ferral (Ferral/Ahmeur bon Ahmeur); and varieties from the Near East, like Moscatel Galego Branco (MoscGaleB/Muscat a Petits Grains Blancs). Genetic group III includes cluster III of the N-J distance analysis (Figure 2), corresponding to PCoA group C (Figure 3) and cluster 1 in the structure analysis at K3 (red color in Figure 4). It grouped Portuguese varietal families identified in this or in previous works (Lopes et al., 1999; Cunha et al., 2015). Finally, genetic group II includes cluster II of the N-J distance analysis (Figure 2), corresponding to PCoA group B (Figure 3) and includes almost all wild plants, varieties obtained by Leão Ferreira de Almeida and ancient varieties from the Vinhos Verdes wine region in the Northwest of Portugal, like Amaral, Barcelo, Branjo e Melhorio. When considering the structure analysis at K =3 the ancient varieties from Vinho Verde region are still assigned to the genetic group containing most wild plants (Supplementary Table 4). Although the correspondences are not complete, the genetic group I would be similar to the S-5.3 (wine—West and Central Europe), while the genetic group III would be closer to S-5.1 (wine and table—Iberian Peninsula and Maghreb) as defined by Bacilieri et al. (2013).
A deeper analysis of these genetic groups points out the existence of close parentage relationships within specific varietal families. In fact, about a quarter of the studied genotypes were found to have kinship with Portuguese or Iberian varieties, as part of duos and/or trios. These include 37 descendants of two female varieties: Marufo (chl D) and Mourisco Branco (Hebén) (chl A), with 19 and 18 offspring respectively (Figure 5A this work; Lacombe et al., 2013; Zinelabidine et al., 2015). The N-J tree, the PCoA and the population analyses grouped these plants with the vinifera genotypes. The female condition obliges them to cross-pollinate, contributing to generate higher genetic diversity and heterozygosity in their offspring and a concomitantly increased plant vigor. This fact probably favored them having a larger number of descendants becoming new cultivated varieties. Mourisco Branco (Hebén) is the female parent of Sarigo (Cayetana Blanca) which has several descendants with its own mother (Zinelabidine et al., 2012; Zinelabidine et al., 2015; Cunha et al., 2015).
A restricted number of the studied grapevine varieties (near 10%) have one or both parents probably coming from outside the Iberian Peninsula. One of these varieties is the unexpected case of Branco Desconhecido (Figure 5A), an offspring of Furmint and Kadarka Bela, two Hungarian grapevine varieties. A different case is that of Savagnin, a very old and disseminated variety (105 synonyms in VIVC) that has been cultivated at least during the last 900 years (Ramos-Madrigal et al., 2019). Savagnin has had a strong impact in the genetic composition of Western and Central European varieties producing many descendants (Lacombe et al., 2013). In addition, two offspring varieties of Savagnin, Alfrocheiro, and Bastardo, were found to contribute several offspring to the Portuguese germplasm (Cunha et al., 2015; Ramos-Madrigal et al., 2019; Figure 5A and Supplementary Table 5). The dissemination of Savagnin and Bastardo, the later already referred by Fernandes (1532) in the Douro region, could be explained by the extended medieval network formed by the Benedictines and Cistercians monasteries all over Europe. Cistercian monks had important and influential monasteries with vineyards in the Douro area (São João de Tarouca and Santa Maria de Salzedas) and its Portuguese headquarter was Santa Maria de Alcobaça (UNESCO world heritage monument), 110 km North of Lisbon. Remarkably, most of the Portuguese varieties that derive from Bastardo and Savagnin are found in the Douro and Lisbon regions. Direct germination of seeds and recollection of naturally growing plants was a documented practice in Portugal (Alarte, 1712; Fonseca, 1791) and it is likely in the origin of its present large grapevine genetic diversity. Variety names like Mourisco de Semente (VIVC #12471) and Barreto de Semente, synonym of the Barreto variety (VIVC #17655) further suggest the idea of plant multiplication through seed germination since “de Semente” means in Portuguese “from seed.”
Almost half of the varieties analyzed in this study could not be assigned to any trio or duo within the ICVV-SNP database. Since this database includes a large collection of Iberian (both Spanish and Portuguese) genotypes, we think that it is highly probable that their progenitors were not conserved either because they were lost during the phytopathological crisis of the 19th century or because they were minor varieties or individual plants lost along the evolution of viticulture.
It is very difficult to establish strong conclusions on the genetic relationships between cultivated and wild grapevine plants given the exiguous number of currently available wild populations. Still, several results from the described analyses suggest the existence of gene flow between wild populations and cultivated plants. In this way, grouping of varieties Barcelo, Branjo, Melhorio, and Amaral with sylvestris plants within cluster II (Figure 2), and/or in the PCoA analysis (Figure 3), and/or in the genetic groups containing most wild accessions for K=2 (Pop2) and for K=3 (Pop3) suggest a close genetic relationship among these cultivated varieties and wild plants. In fact, phenotypically, these four varieties share some trait similarity with wild plants: small and very loose bunches with many visible pedicels, small berries with blue black skin color, high acidity, and small orbicular leaves with one or three lobes. In addition, Barcelo and Amaral form a duo (LOD 45.76), as well as Melhorio and Amaral (LOD 25.18) (Supplementary Table 3). Interestingly, Amaral is clustered with wild plants in all studies and has several descendants among Portuguese varieties, as shown here (Supplementary Tables 3 and 5) and in previous works (Díaz-Losada et al., 2011; Lacombe et al., 2013). The association of these varieties used in the Vinhos Verdes region with the wild vines points out to the existence of introgression from wild plants into currently cultivated varieties in Portugal. Interestingly, wines from this region have low alcohol content, high acidity, and are naturally sparkling (http://www.vinhoverde.pt/en/demarcated-region, features characteristic of must or wine obtained from wild grapes: low sugar content, low pH, and high tartaric acid content (Arroyo-Garcia et al., 2006; Cunha et al., 2007). These results are in agreement with Riaz et al. (2018) whose findings indicated a considerable amount of gene flow between the two subspecies, which limited their differentiation, and the contribution of Western European wild populations to the development of Western European wine grapes.
The close genetic relationship between Vinhos Verdes varieties and wild plants is especially remarkable because the wild populations are not from that region, where wild plants have not been found. Anthropogenic pressure is probable the main cause of the current absence of wild vines populations in the Vinhos Verdes region, as it is the case in many other regions. At present, only in the south of Portugal it was possible to find wild populations surviving in riparian woods along small streams (Cunha et al., 2007).
On the other side, the degree of introgression from ancient and current cultivated varieties into the actual wild populations cannot be estimated but it is exemplified by the existence of wild plants grouped within clusters of cultivated ones (Figure 2), and by the relatively low values of membership coefficient to the wild group of the plants of the population from Ribera de Toutalga (Supplementary Table 4). The human use of V. vinifera ssp. sylvestris continued uninterrupted until the late 20th century to produce vinegar and folk medicines, as well as for other uses not related to grapes or wine (Ocete et al., 1999). This continuous use probably favored obtaining new varieties through selection of interesting individuals produced from seeds, in some cases perhaps from spontaneous hybrids between cultivated and sylvestris plants, such as those found in this work, which could inherit the hermaphrodite allele (50%) favoring their selection.
In our case a dubious, intermediate, flower phenotype was found in the accession 110402CB, which is a descendant of a cross between Folha de Figueira and 110403CB, a female wild plant from the Ponsul river population (Figure 5A). Apart from this full pedigree, two putative parent-offspring relationships (Supplementary Table 3) were also identified involving cultivated and wild plants: one in the same Ponsul population between a female plant 110405CB and Schiava Grossa and the other in the Alcácer do Sal population between female plant 110603AC (same type of flower as plant 110402CB) and Castelão. These three grapevines, sampled as “wild” (110402CB, 110405CB, and 110603AC) are grouped in cluster II (subcluster a) (Figure 2) or in intermediate positions close to vinifera genotypes (Figure 3) and represent clear cases of introgression into the sylvestris subspecies. There are other two grapevines collected in the wild populations (120209SE a male plant and 120302SE, female plant with straight stamens) that grouped in the cluster I (Figure 2), or in intermediate positions (Figure 3) and for which no kinship relationships were found. They could correspond to feral forms with morphological characteristics of wild plants. Introgression of cultivated into wild plants threatens the large potential of sylvestris as a source of resilience factors in future breeding programs to deal with climate change and the increasing demand of a sustainable viticulture (Marrano et al., 2018).
Grapevine domestication is probably a long process taking place along thousands of years and within a wide geographical area. Some authors defend the existence of a single domestication event, with later wild introgression events for local adaptation (Zhou et al., 2019) while others take the view of a primary domestication event in Transcaucasia and later secondary events of domestication in the West (Grassi et al., 2003; Arroyo-García et al., 2006). These two views are not so far, because both subspecies are not separate compartments in nature and gene flow has always occurred between them (Di Vecchi-Staraz et al., 2009) and because, given the relevance of vegetative multiplication in grapevine, the number of generations separating cultivated varieties from wild plants is likely very small (This et al., 2006; Arroyo-García et al., 2006). This is shown by the small genetic and phenotypic differences between wild and cultivated plants observed in some cases like those found in this work. The wild vines analyzed here belong to populations still present in the Tagus, Guadiana and Alcácer do Sal river basins, where palynological data of Paúl dos Patudos, Alpiarça (upstream the Tagus valley), showed a 33% increase of Vitis pollen in a stratum of the 7th century BCE compared to earlier strata. This led the authors, when related it to other evidences of anthropogenic changes in the palynological structure of the deposits, to consider it to be due to grapevine cultivation. Records predating 6050 BCE are certainly from subspecies sylvestris (Leeuwaarden and Janssen, 1985).
Moreover, seed remains found in northern part of Portugal have a Stummer index between 0.65 and 0.75 that could be assigned to a mix of both vinifera and sylvestris subspecies, as well as, seeds remains found in Iron Age site in Castro Marim (Algarve) (Ramil Rego and Aira Rodríguez, 1993; Queiroz and Mateus, 1994; Aira Rodríguez and Ramil Rego, 1995; Sanches, 1997; Queiroz and Mateus, 2007). The existence of a large particular gene pool of Portugal varieties that bear chlorotype A means that the seeds originating these cultivars (or her mothers, or her grandmothers) were produced in sylvestris plants, and thus human beings collected and multiplied plants grown from seeds of sylvestris plants, which had germinated and raised in the wild, and so they “domesticated” (select, collect, and cultivate) such plants. All these findings point out to the possible genetic contribution of wild to local cultivated grapevines, as it was shown in France for most archeological samples analyzed by Ramos-Madrigal et al. (2019) and it was indicated for wine Western European cultivars by Riaz et al. (2018).
The identification of kin relationships allowed the identification of major contributors to the present Portuguese germplasm, in particular the contribution of female genotypes. An important contribution for the Portuguese germplasm consists in the descendants of Savagnin (chl D) (Alfrocheiro, Bastardo, and Molar) probably introduced by Cistercian monks. Despite some foreign contributions, exemplified by the descendance of Marufo (chl D), about half of the Portuguese grapevine varieties analyzed in this study hold the A chlorotype, typical of the Iberian Peninsula and have no known kinship, while a quarter of the genotypes have some sort of kinship with Portuguese or Iberian varieties. The high number of varieties resulting from different parents seems to reflect a wide traditional use of seeds for vine propagation in Portugal.
The analysis of the SNP genotypes of the Portuguese cultivated and wild germplasm allowed to detect the existence of gene flow between both subspecies probably since historical times and until near present. This is corroborated by the disclosure of wild-cultivated kinship relations, especially in the South of Portugal where wild populations are still present in riparian woods. This subspecies has suffered a severe reduction in its genetic diversity due to the nineteenth century phytopathological crisis and to the anthropogenic pressure, hindering the study of its domestication. The introgression in the wild plants observed in this work illustrates the magnitude of the problem and reinforce the need to protect, study, and use the existing wild germplasm, which may be a source of useful characteristics to incorporate in the Portuguese germplasm.
The genetic relationships, ampelographic similarities, and must and wine characteristics share between the Vinhos Verdes region varieties and the wild vines, the specific genetic pool with chlorotype A existing in Portugal and different archaeological findings support the possible existence of secondary domestication events in Portugal, or, at least, of an introgression process of wild into cultivated grapevines.
All datasets generated and analyzed for this study are included in the article/Supplementary Material.
JC, JI, and JE-D conceived and designed the experiments. JC and JI performed the experiments. JC, JI, and MT-S analyzed the data. JC, JI, MT-S, JB, PF, JM-Z, and JE-D wrote the manuscript. JE-D, JC, and JB established vinifera and sylvestris collections and collected plant material. All authors contributed to editing the final version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by the funded projects: i) PTDC/AGR-PRO/4261/2014 and ii) PDR2020-784-042738. The authors acknowledge COST Action FA1003 (European Cooperation in the Field of Scientific and Technical Research) for a short scientific mission of JC in the ICVV to analyze and discuss some of the results presented in this paper and the financial support from Fundação para a Ciência e a Tecnologia (Lisbon, Portugal) through the research unit “GREEN-it: Bioresources for Sustainability” (UID/Multi/04551/2013). We want to thank all the people who have contributed to the development of the ICVV-SNP database. We would like to thank to the reviewers for useful suggestion to improve our manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2020.00127/full#supplementary-material
Aira Rodríguez, M. J., Ramil Rego, P. (1995). Datos paleobotánicos del Norte de Portugal (Baixo Minho). Estudio polínico y paleocarpológico. Lagascalia 18 (1), 25–38.
Alarte, V. (1712). Agricultura das vinhas, e tudo o que pertence a ellas até perfeito recolhimento do vinho, e relação das suas virtudes, e da cepa, vides, folhas, e borras (Lisboa: Officina Real Deslandesiana).
Almandanim, M. C., Baleiras-Couto, M. M., Pereira, H. S., Carneiro, L. C., Fevereiro, P., Eiras-Dias, J. E., et al. (2007). Genetic diversity of the grapevine (Vitis vinifera L.) cultivars most utilized for wine production in Portugal. Vitis 46 (3), 116–119.
Arroyo-García, R., Cantos, M., Lara, M., López, M. Á, Gallardo, A., Ocete, C. A., et al. (2016). Characterization of the largest relic Eurasian wild grapevine reservoir in Southern Iberian Peninsula. Spanish Journal of Agricultural Research 14 (3), e0708. doi: 10.5424/sjar/2016143-8929
Arroyo-García, R., Ruiz-García, L., Bolling, L., Ocete, R., López, M. A., Arnold, C., et al. (2006). Multiple origins of cultivated grapevine (Vitis vinifera L. ssp. sativa) based on chloroplast DNA polymorphisms. Mol. Ecol. 15 (12), 3707–3714. doi: 10.1111/j.1365-294X.2006.03049.x
Arruda, A. M. (2008). “Fenícios e Púnicos em Portugal: Problemas e Perspectivas,” in Nuevas Perspectivas II: la arqueología fenicia y púnica en la Península Ibérica. Eds. Vita, J. P., Zamora, J.Á. (Barcelona: Universidad Pompeu Fabra), 13–23.
Bacilieri, R., Lacombe, T., Le Cunff, L., Di Vecchi-Staraz, M., Laucou, V., Genna, B., et al. (2013). Genetic structure in cultivated grapevines is linked to geography and human selection. BMC Plant Biol. 13, 25. doi: 10.1186/1471-2229-13-25
Barros, L. (1998). Introdução à Pré e Proto-História de Almada (Almada: Câmara Municipal de Almada).
Bourquin, J. C., Sonko, A., Otten, L., Walter, B. (1993). Restriction fragment length polymorphism and molecular taxonomy in Vitis vinifera L. Theor. Appl. Genet. 87 (4), 431–438. doi: 10.1007/BF00215088
Buxó, R. (2008). The agricultural consequences of colonial contacts on the Iberian Peninsula in the first millennium B.C. Veg. Hist. Archaeobot. 17 (1), 145–154. doi: 10.1007/s00334-007-0133-7
Cabezas, J. A., Ibáñez, J., Lijavetzky, D., Vélez, D., Bravo, G., Rodríguez, V., et al. (2011). A 48 SNP set for grapevine variety identification. BMC Plant Biol. 11, 153. doi: 10.1186/1471-2229-11-153
Castro, I., Martín, J. P., Ortiz, J. M., Pinto-Carnide, O. (2011). Varietal discrimination and genetic relationships of Vitis vinifera L. cultivars from two major Controlled Appellation (DOC) regions in Portugal. Scientia Hortic. 127, 507–514. doi: 10.1016/j.scienta.2010.11.018
Castro, I., Pinto-Carnide, O., Ortiz, J. M., Martín, J. P. (2013). Chloroplast genome diversity in Portuguese grapevine (Vitis vinifera L.) varieties. Mol. Biotechnol. 54 (2), 528–540. doi: 10.1007/s12033-012-9593-9
Cunha, J., Baleiras-Couto, M., Cunha, J. P., Banza, J., Soveral, A., Carneiro, L. C., et al. (2007). Characterization of Portuguese Populations of Vitis vinifera L. ssp. sylvestris (Gmelin) Hegi. Genet. Resour. Crop Evol. 54 (5), 981–988. doi: 10.1007/s10722-006-9189-y
Cunha, J., Teixeira Santos, M., Carneiro, L. C., Fevereiro, P., Eiras-Dias, J. E. (2009). Portuguese traditional grapevine varieties and wild vines (Vitis vinifera L.) share morphological and genetic traits. Genet. Resour. Crop Evol. 56 (7), 975–989. doi: 10.1007/s10722-009-9416-4
Cunha, J., Teixeira-Santos, M., Veloso, M., Carneiro, L., Eiras-Dias, J., Fevereiro, P. (2010). The Portuguese Vitis vinifera L. Germplasm: Genetic Relations between Wild and Cultivated Vines. Ciência e Técnica Vitivinícola 25 (1), 25–37.
Cunha, J., Zinelabidine, L. H., Teixeira-Santos, M., Brazão, J., Fevereiro, P., Martínez-Zapater, J. M., et al. (2015). Grapevine variety ‘Alfrocheiro' or ‘Bruñal' plays a primary role in the relationship among Iberian grapevines. Vitis 54, 59–65.
Cunha, J., Ibáñez, J., Teixeira-Santos, M., Brazão, J., Fevereiro, P., Martínez-Zapater, J. M., et al. (2016). Characterisation of the Portuguese grapevine germplasm with 48 single-nucleotide polymorphisms. Aust. J. Grape Wine Res. 22 (3), 504–516. doi: 10.1111/ajgw.12225
Díaz-Losada, E., Tato Salgado, A., Ramos-Cabrer, A. M., Pereira-Lorenzo, S. (2011). Determination of genetic relationships of Albariño and Loureira varieties with the Caíño group by microsatellites. Am. J. Enol. Vitic. 62 (3), 371–375. doi: 10.5344/ajev.2011.10040
De Lorenzis, G., Mercati, F., Bergamini, C., Cardone, M. F., Lupini, A., Mauceri, A., et al. (2019). SNP genotyping elucidates the genetic diversity of Magna Graecia grapevine germplasm and its historical origin and dissemination. BMC Plant Biol. 19, 7. doi: 10.1186/s12870-018-1576-y
Di Vecchi-Staraz, M., Laucou, V., Bruno, G., Lacombe, T., Gerber, S., Bourse, T., et al. (2009). Low level of pollen-mediated gene flow from cultivated to wild grapevine: consequences for the evolution of the endangered subspecies Vitis vinifera L. subsp. silvestris. J. Heredity 100 (1), 66–75. doi: 10.1093/jhered/esn084
Eiras-Dias, J. E., Faustino, R., Clímaco, P., Fernandes, P., Cruz, A., Cunha, J., et al. (2011). Catálogo das castas para vinho cultivadas em Portugal Vol. 1 (Lisboa: Instituto da Vinha e do Vinho I.P., Chaves Ferreira – Publicações).
Eiras-Dias, J. E., Faustino, R., Clímaco, P., Fernandes, P., Cruz, A., Cunha, J., et al. (2013). Catálogo das castas para vinho cultivadas em Portugal Vol. 2 (Lisboa: Instituto da Vinha e do Vinho I.P., Chaves Ferreira – Publicações)).
Emanuelli, F., Lorenzi, S., Grzeskowiak, L., Catalano, V., Stefanini, M., Troggio, M., et al. (2013). Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 13, 39. doi: 10.1186/1471-2229-13-39
Evanno, G., Regnaut, S., Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Fabião, C. (1998). O vinho na Lusitânia: reflexões em torno de um problema arqueológico. Rev. Portuguesa Arqueologia 1 (1), 169–198.
Falush, D., Stephens, M., Pritchard, J. K. (2007). Inference of population structure using multilocus genotype data: dominant markers and null alleles. Mol. Ecol. Notes 7 (4), 574–578. doi: 10.1111/j.1471-8286.2007.01758.x
Fernandes, R. (1532). Descrição do terreno ao redor de Lamego duas léguas, [1531-1532]. Ed. Barros, A.J.M. (Casal de Cambra: Caleidoscópio — Edição e Artes Gráficas). (2012)
Fletcher, W. J., Boski, T., Moura, D. (2007). Palynological evidence for environmental and climatic change in the lower Guadiana valley, Portugal, during the last 13 000 Years. Holocene 17 (4), 481–494. doi: 10.1177/0959683607077027
Fonseca, F. P. R. (1791). “Memória sobre o assunto proposto pela Academia Real das Ciências para o ano de 1790. Qual é o método mais conveniente, e cautelas necessárias para o cultivo das vinhas em Portugal; para a vindima; extracção e fermentação do mosto; conservação, e bondade do vinho, e para a melhor reputação, e vantagem deste importante ramo do nosso comércio?,” in Memórias de Agricultura premiadas pela Academia Real das Ciências de Lisboa ed. Tomo, II. (Lisboa: Academia Real das Ciências de Lisboa), 1–273.
Ghaffari, S., Hasnaoui, N., Zinelabidine, L. H., Ferchichi, A., Martínez-Zapater, J. M., Ibáñez, J. (2014). Genetic diversity and parentage of Tunisian wild and cultivated grapevines (Vitis vinifera L.) as revealed by single nucleotide polymorphism (SNP) markers. Tree Genet. Genomes 10 (4), 1103–1112. doi: 10.1007/s11295-014-0746-9
Ghira, J. C., Carneiro, L. C., Carvalho, H. P., Garcia, I. S., Vinagre, J. S. (1982). Estudo vitícola e enológico de castas novas da EAN Vol. 9 (Lisboa: D.G. Extensão Rural).
Grassi, F., Labra, M., Imazio, S., Spada, A., Sgorbati, S., Scienza, A., et al. (2003). Evidence of a secondary grapevine domestication centre detected by SSR analysis. Theor. Appl. Genet. 107 (7), 1315–1320. doi: 10.1007/s00122-003-1321-1
Ibáñez, J., Muñoz-Organero, G., Zinelabidine, L. H., De Andrés, M. T., Cabello, F., Martínez-Zapater, J. M. (2012). Genetic origin of the grapevine variety Tempranillo. Am. J. Enol. Vitic. 63 (4), 549–553. doi: 10.5344/ajev.2012.12012
Imazio, S., Labra, M., Grassi, F., Scienza, A., Failla, O. (2006). Chloroplast microsatellites to investigate the origin of grapevine. Genet. Resour. Crop Evol. 53 (5), 1003–1011. doi: 10.1007/s10722-004-6896-0
Kalinowski, S. T., Taper, M. L., Marshall, T. C. (2007). Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 16 (5), 1099–1106. doi: 10.1111/j.1365-294X.2007.03089.x
Kalinowski, S. T. (2005). HP – Rare: a computer program for performing rarefaction mensures of allelic diversity. Mol. Ecologie Notes 5, 187–189. doi: 10.1111/j.1471-8286.2004.00845.x
Kumar, S., Stecher, G., Tamura, K. (2016). MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 33 (7), 1870–1874. doi: 10.1093/molbev/msw054
Lacerda Lobo, C. (1790). “Memória sobre a cultura das vinhas de Portugal,” in Memórias Económicas da Academia Real das Ciências de Lisboa, para o adiantamento da agricultura, das artes, e da indústria de Portugal, e suas conquistas (1789-1815) ed. Tomo, II. (Lisboa: Academia Real das Ciências de Lisboa, 13–162.
Lacombe, T., Boursiquot, J. M., Laucou, V., Di Vecchi-Staraz, M., Péros, J. P., This, P. (2013). Large-scale parentage analysis in an extended set of grapevine varieties (Vitis vinifera L.). Theor. Appl. Genet. 126 (2), 401–414. doi: 10.1007/s00122-012-1988-2
Laucou, V., Launay, A., Bacilieri, R., Lacombe, T., Adam-Blondon, A.-F., Bérard, A., et al. (2018). Extended diversity analysis of cultivated grapevine Vitis vinifera with 10K genome-wide SNPs. PloS One 13 (2), e0192540. doi: 10.1371/journal.pone.0192540
Leeuwaarden, V. W., Janssen, C. R. (1985). “A preliminary palynological study of peat deposits near an oppidum in the Lower Tagus Valley,” in Actas da I Reunião do Quaternário Ibérico, vol. II. (Lisboa: Grupo de Trabalho Português para o Estudo do Quaternário), 226–236.
Lijavetzky, D., Cabezas, J., Ibáñez, A., Rodríguez, V., Martínez-Zapater, J. M. (2007). High throughput SNP discovery and genotyping in grapevine (Vitis vinifera L.) by combining a re-sequencing approach and SNPlex technology. BMC Genomics 8, 424. doi: 10.1186/1471-2164-8-424
Lopes, M. S., Sefc, K. M., Eiras Dias, E., Steinkellner, H., Laimer Câmara Machado, M., Câmara Machado, A. (1999). The use of microsatellites for germplasm management in a Portuguese grapevine collection. Theor. Appl. Genet. 99 (3–4), 733–739. doi: 10.1007/s001220051291
Lopes, M. S., Santos, M. R., Eiras-Dias, J. E., Mendonça, D., Câmara Machado, A. (2006). Discrimination of Portuguese grapevines based on microsatellite markers. J. Biotechnol. 127 (1), 34–44. doi: 10.1016/j.jbiotec.2006.06.006
MAMAOT (2012). Portaria N° 380/2012, de 22 de Novembro. Ministério da Agricultura, do Mar, do Ambiente e do Ordenamento do Território (MAMAOT). Diário da República, 1.a Série - N.° 226. Lisboa, Portugal.
Marrano, A., Birolo, G., Prazzoli, M. L., Lorenzi, S., Valle, G., et al. (2017). SNP-Discovery by RAD-Sequencing in a Germplasm Collection of Wild and Cultivated Grapevines (V. vinifera L.). PloS One 12, 1–19. doi: 10.1371/journal.pone.0170655
Marrano, A., Micheletti, D., Lorenzi, S., Neale, D., Grando, M. S. (2018). Genomic signatures of different adaptations to environmental stimuli between wild and cultivated Vitis vinifera L. Hortic. Res. 5, 12. doi: 10.1038/s41438-018-0041-2
Martinez Tomé, A. (1991). El monasterio cisterciense en el origen de los vinos españoles (Madrid: Servicio de Extensión Agraria).
Maul, et al. (2019). Vitis International Variety Catalogue - www.vivc.de (accessed June 2019).
McGovern, P., Jalabadze, M., Batiuk, S., Callahan, M. P., Smith, K. E., Hall, G. R., et al. (2017). Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. 114, 48) E10309–E10318. doi: 10.1073/pnas.1714728114
Miller, N. F. (2008). Sweeter than wine? The use of the grape in early western Asia. Antiquity 82 (318), 937–946. doi: 10.1017/S0003598X00097696
Moita Maçanita, A., Santos, R., Catarina Gomes, A. (2018). Unravelling the origin of Vitis vinifera L. Verdelho. Aust. J. Grape Wine Res. 24, 450–460. doi: 10.1111/ajgw.12353
Myles, S., Boyko, A. R., Owens, C. L., Brown, P. J., Grassi, F., Aradhya, et al. (2011). Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. U. S. A. 108 (9), 3530–3535. doi: 10.1073/pnas.1009363108
Ocete, R., López, M. A., Pérez, M. A., Del Tío, R., Lara, M. (1999). Las poblaciones españolas de vid silvestre: Características de un recurso fitogenético a conservar. Monografías INIA: Agrícola 3, 1–52.
OIV (1983). Descriptor list for grapevine varieties and Vitis species (Paris: Organisation Internationale de la Vigne et du Vin).
OIV (2009). Descriptor list for grapevine varieties and Vitis species. 2nd Ed (Paris: Organisation Internationale de la Vigne et du Vin).
Olmo, H. P. (1995). “The origin and domestication of vinifera grape,” in The origin and ancient history of wine, McGovern, P., Fleming, S. J., Katz, S. H. (New York: Gordon & Breach), 31–43.
Peakall, R., Smouse, P. E. (2012). GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28 (19), 2537–2539. doi: 10.1093/bioinformatics/bts460
Pritchard, J. K., Stephens, M., Donnelly, P. J. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959.
Queiroz, P. F., Mateus, J. E. (1994). Preliminary palynological investigation on the Holocene deposits of Lagoa de Albufeira and Lagoa de Melides, Alentejo (Portugal). Revista de Biologia Lisboa 15(1/4), 15–27.
Queiroz, P. F., Mateus, J. E. (2007). Acerca das Graínhas de Uva da Idade do Ferro de Castro Marim (Lisboa: CIPA – IPA).
Ramil Rego, P. R., Aira Rodríguez, M. J. (1993). A palaeocarpological study of Neolithic and Bronze Age levels of the Buraco da Pala rock-shelter (Bragança, Portugal). Veg. Hist. Archaeobot. 2 (3), 163–172. doi: 10.1007/BF00198587
Ramos-Madrigal, J., Runge, A. K. W., Bouby, L., Lacombe, T., Castruita, J. A. S., Adam-Blondon, A.-F., et al. (2019). Palaeogenomic insights into the origins of French grapevine diversity. Nat. Plants 5 (6), 595–603. doi: 10.1038/s41477-019-0437-5
Renfrew, J. M. (1973). Paleoethnobotany. The prehistoric food plants of the Near East and Europe. (Nueva York: Columbia University Press).
Riaz, S., De Lorenzis, G., Velasco, D., Koehmstedt, D., Maghradze, D., Bobokashvili, Z., et al. (2018). Genetic diversity analysis of cultivated and wild grapevine (Vitis vinifera L.) accessions around the Mediterranean basin and Central Asia. BMC Plant Biol. 18, 137. doi: 10.1186/s12870-018-1351-0
Saitou, N., Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4 (4), 406–425. doi: 10.1093/oxfordjournals.molbev.a040454
Sanches, M. J. (1997). Pré-história recente de Trás-os-Montes e Alto Douro: o Abrigo do Buraco da Pala (Mirandela) no contexto regional. Vol. 2 (Porto: Sociedade Portuguesa de Antropologia e Etnologia).
Savo, V., Kumbaric, A., Caneva, G. (2016). Grapevine (Vitis Vinifera L.) symbolism in the ancient Euro-Mediterranean cultures. Econ. Bot. 70 (2), 190–197. doi: 10.1007/s12231-016-9347-x
Schneider, H., Höfer, D., Trog, C., Mäusbacher, R. (2016). Holocene landscape development along the Portuguese Algarve coast – a high resolution palynological approach. Quat. Int. 407, 47–63. doi: 10.1016/j.quaint.2016.02.039
Sefc, K. M., Lopes, M. S., Lefort, F., Botta, R., Roubelakis-Angelakis, K. A., Ibáñez, J., et al. (2000). Microsatellite variability in grapevine varieties from different European regions and evaluation of assignment testing to assess the geographic origin of varieties. Theor. Appl. Genet. 100 (3–4), 498–505. doi: 10.1007/s001220050065
Sousa, E., Guerra, S. (2018). A presença fenícia em Lisboa: novos vestígios descobertos no alto da colina do Castelo de São Jorge. SAGVNTVM (P.L.A.V.) 50, 57–88. doi: 10.7203/SAGVNTVM.50.10636
Stummer, A. (1911). Zur Urgeschichte der Rebe und des Weinbaues. Mitt. Anthropologischen Ges. in Wien 41, 283–296.
Tereso, J., Rego, R. P., da Silva, A.R. (2011). “A Exploração de Recursos Alimentares Silvestres e Seu Enquadramento Nas Dinâmicas Económicas e Sociais Das Comunidades Agrícolas Desde a Pré-História à Época Romana,” in Florestas do Norte de Portugal: História, Ecologia e Desafios de Gestão. Eds. Tereso, J. P., Honrado, J. P., Pinto, A. T., Rego, F. C. (InBio - Rede de Investigação em Biodiversidade e Biologia Evolutiva), 55–83.
This, P., Lacombe, T., Thomas, M. R. (2006). Historical origins and genetic diversity of wine grapes. Trends Genet. 22, 511–519. doi: 10.1016/j.tig.2006.07.008
Thomas, M. R., Matsumoto, S., Cain, P., Scott, N. S. (1993). Repetitive DNA of grapevine: classes present and sequences suitable for variety identification. Theor. Appl. Genet. 86 (2–3), 173–180. doi: 10.1007/BF00222076
van der Knaap, W. O., van Leeuwen, J. F. N. (1995). Holocene vegetation succession and degradation as responses to climatic change and human activity in the Serra de Estrela, Portugal. Rev. Palaeobot. Palynol. 89 (3–4), 153–211. doi: 10.1016/0034-6667(95)00048-0
Veloso, M. M., Almandanim, M. C., Baleiras-Couto, M., Sofia Pereira, H., Carneiro, L. C., Fevereiro, P., et al. (2010). Microsatellite database of grapevine (Vitis Vinifera L.) varieties used for wine production in Portugal. Ciência e Técnica Vitivinícola 25 (2), 53–61.
Vis, G.-J., Kasse, C. (2009). Late Quaternary valley-fill succession of the Lower Tagus Valley, Portugal. Sediment. Geol. 221 (1–4), 19–39. doi: 10.1016/j.sedgeo.2009.07.010
Zhou, Y., Muyle, A., Gaut, B. S. (2019). “Evolutionary genomics and the domestication of grapes,” in The grape genome. Eds. Cantu, D., Walker., M. A. (Cham: Springer International Publishing), 39–55.
Zinelabidine, L. H., Haddioui, A., Rodríguez, V., Cabello, F., Eiras-Dias, J. E., Martínez Zapater, J. M., et al. (2012). Identification by SNP analysis of a major role for Cayetana Blanca in the genetic network of Iberian Peninsula grapevine varieties. Am. J. Enol. Vitic. 63 (1), 121–126. doi: 10.5344/ajev.2011.11052
Zinelabidine, L. H., Cunha, J., Eiras-Dias, J. E., Cabello, F., Martínez-Zapater, J. M., Ibáñez, J. (2015). Pedigree analysis of the Spanish grapevine variety ‘Hebén'. Vitis 54, 81–86.
Keywords: grapevine varieties, genetic relationships, Iberia, introgression, pedigrees, sylvestris, wild
Citation: Cunha J, Ibáñez J, Teixeira-Santos M, Brazão J, Fevereiro P, Martínez-Zapater JM and Eiras‐Dias JE (2020) Genetic Relationships Among Portuguese Cultivated and Wild Vitis vinifera L. Germplasm. Front. Plant Sci. 11:127. doi: 10.3389/fpls.2020.00127
Received: 13 September 2019; Accepted: 28 January 2020;
Published: 05 March 2020.
Edited by:
Fabrizio Grassi, University of Bari Aldo Moro, ItalyReviewed by:
Jean-Pierre Péros, Institut National de la Recherche Agronomique Centre Montpellier, FranceCopyright © 2020 Cunha, Ibáñez, Teixeira-Santos, Brazão, Fevereiro, Martínez-Zapater and Eiras‐Dias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jorge Cunha, am9yZ2UuY3VuaGFAaW5pYXYucHQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.