
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 27 November 2018
Sec. Evolutionary and Population Genetics
Volume 9 - 2018 | https://doi.org/10.3389/fpls.2018.01698
This article is part of the Research TopicPopulation Genetics and Evolutionary History of Plants in Mountainous RegionsView all 29 articles
 Javier Morente-López1*
Javier Morente-López1* Cristina García2,3
Cristina García2,3 Carlos Lara-Romero1,4
Carlos Lara-Romero1,4 Alfredo García-Fernández1
Alfredo García-Fernández1 David Draper5,6
David Draper5,6 José María Iriondo1*
José María Iriondo1*The study of the drivers that shape spatial genetic structure across heterogeneous landscapes is one of the main approaches used to understand population dynamics and responses in changing environments. While the Isolation-by-Distance model (IBD) assumes that genetic differentiation increases among populations with geographical distance, the Isolation-by-Resistance model (IBR) also considers geographical barriers and other landscape features that impede gene flow. On the other hand, the Isolation-by-Environment model (IBE) explains genetic differentiation through environmental differences between populations. Although spatial genetic studies have increased significantly in recent years, plants from alpine ecosystems are highly underrepresented, even though they are great suitable systems to disentangle the role of the different factors that structure genetic variation across environmental gradients. Here, we studied the spatial genetic structure of the Mediterranean alpine specialist Silene ciliata across its southernmost distribution limit. We sampled three populations across an altitudinal gradient from 1850 to 2400 m, and we replicated this sample over three mountain ranges aligned across an E-W axis in the central part of the Iberian Peninsula. We genotyped 20 individuals per population based on eight microsatellite markers and used different landscape genetic tools to infer the role of topographic and environmental factors in shaping observed patterns along the altitudinal gradient. We found a significant genetic structure among the studied Silene ciliata populations which was related to the orography and E-W configuration of the mountain ranges. IBD pattern arose as the main factor shaping population genetic differentiation. Geographical barriers between mountain ranges also affected the spatial genetic structure (IBR pattern). Although environmental variables had a significant effect on population genetic diversity parameters, no IBE pattern was found on genetic structure. Our study reveals that IBD was the driver that best explained the genetic structure, whereas environmental factors also played a role in determining genetic diversity values of this dominant plant of Mediterranean alpine environments.
Species resilience to changing environments largely depends on their genetic diversity (Lande and Shannon, 1996; Frankham et al., 2002). Thus, knowledge on the spatial distribution of species genetic diversity is an essential component for understanding the challenges brought about by climate change (Manel and Holderegger, 2013). This is especially important for species in habitats with marked environmental gradients, such as those found in mountain ecosystems. Projected climate warming rates are expected to have a great impact on mountain ecosystems (Beniston et al., 1997; Nogués-Bravo et al., 2007; Kohler et al., 2010; Rangwala and Miller, 2012), particularly on alpine plant communities (Thuiller et al., 2005; Gottfried et al., 2012; Steinbauer et al., 2018). It is, therefore, crucial to identify the main ecological drivers of genetic structure at different spatial scales to forecast the response of alpine plant species to climate change.
Several hypotheses have been proposed to generalize patterns of genetic variation across space. Gene flow limitation and drift caused by geographical isolation generate an Isolation-by-Distance (IBD) pattern (Wright, 1943). It assumes that linear relationship between genetic and Euclidean geographic distance per se is the main driver of genetic structure and considers that other factors like landscape features, range boundaries or environmental characteristics influencing gene flow are not relevant (Manel et al., 2003; McRae, 2006; Storfer et al., 2007). Nowadays, there is evidence that IBD may be too simplistic and that it should be contrasted with more complex models (Jenkins et al., 2010). Thus, the Isolation-by-Resistance (IBR) pattern incorporates range boundaries and landscape features, such as geographical barriers, as a cause of gene flow limitation and drift (McRae, 2006; Spear et al., 2010; Cushman et al., 2015). Both IBD and IBR are closely related to the underlying process of Isolation-by-Dispersal and indirectly related to environmental conditions (Orsini et al., 2013). Isolation-by-Environment (IBE) pattern is caused by environmental heterogeneity and local adaptation related to strong divergent selection (Sexton et al., 2014; Wang and Bradburd, 2014). In mountain ecosystems, high isolation produced by steep topography, barriers and environmental gradients facilitate various processes that generate different isolation patterns. Historical patterns of range expansion and contraction can also modify current isolation effects and affect genetic structure (Cushman et al., 2015). All these processes can act alone or in combination, and their importance may vary depending on the spatial scale of observation (Orsini et al., 2013).
Isolation processes related to geographic and environmental distances may produce profound demographic and genetic outcomes for plant populations within species distribution ranges (Eckert et al., 2008). Species range limits, characterized by increased genetic isolation (Sexton et al., 2009), often occur across ecological gradients where habitats become less suitable and environmental differentiation increases when moving toward the margins (Kawecki, 2008). Inside species distribution ranges, we can distinguish populations inhabiting environmentally central areas from populations occurring in marginal areas, i.e., those located at the edge of the species environmental and/or geographical distribution range (Soule, 1973; Brussard, 1984; Kawecki, 2008; Pironon et al., 2015; Papuga et al., 2018). It is important to consider that the geographical center of the species range is not necessarily associated to the areas with higher habitat quality (Sagarin et al., 2006). In the distribution margin, populations experience environmental conditions that differ from those found at the central areas and they tend to be prone to environmental fluctuations that ultimately reduce habitat quality and quantity and restrict resource availability, affecting population demography and thus genetic diversity (Hampe and Petit, 2005; Eckert et al., 2008; Kawecki, 2008). Thus, as we approach to the distribution margins it is expected that other processes such as IBE will influence the distribution of genetic diversity, in addition to IBD.
Plants from alpine ecosystems provide a very interesting context to disentangle the various mechanisms that structure genetic variation within and among populations at different scales related to patterns of isolation between populations (IBD/IBR vs. IBE). Several studies have shown adaptive genetic differentiation and local adaptation processes related with the strong environmental variability in these ecosystems using phenotypic, genetic, and/or genomic data (e.g., Gonzalo-Turpin and Hazard, 2009; Frei et al., 2012a, 2014; Di Pierro et al., 2017; Hämälä et al., 2018). In some instances, local adaptation persists despite the existence of significant gene flow between populations (Gonzalo-Turpin and Hazard, 2009; Kim and Donohue, 2013). However, in other cases, local adaptation has not been found (Ometto et al., 2015; Hirst et al., 2016; Hamann et al., 2017). Also the effect of geographical distance and landscape configuration have been documented as an important forces that hinder gene flow and promote genetic differentiation between populations (Kuss et al., 2008; Aægisdóttir et al., 2009; Geng et al., 2009; Wang et al., 2011; De Vriendt et al., 2017). However, few studies have combined different isolation processes in the context of landscape genetic approaches (e.g., Mosca et al., 2014; Wu et al., 2015; Noguerales et al., 2016) and focused on environmental variables causing the genetic isolation patters identification (e.g., Manel et al., 2012; Mosca et al., 2012). Furthermore, although landscape genetics studies have increased significantly in recent years, plants are still highly underrepresented (Storfer et al., 2010; DiLeo and Wagner, 2016).
Mediterranean mountain ranges encompass a territory with wide environmental and geographical heterogeneity where many species reach their southernmost distribution range limits. They are also rich in endemic species and are biogeographically important as glacier refuges (Nieto Feliner, 2014). In this context, we combined geographic, environmental and genetic data and applied landscape genetics tools to depict the genetic structure patterns of nine populations of a Mediterranean alpine plant, Silene ciliata Poiret (Caryophyllaceae), distributed along the elevational range of the species in central Spain. As an alpine species, the distribution range of S. ciliata has experienced historical environmental fluctuations related to glaciations (Hewitt, 2001; Nieto Feliner, 2011) and is presently challenged by environmental changes related to climate change (Körner, 2003, 2007). The species grows across an elevational gradient, where populations at the lowest elevation experience the most stressful conditions (due to water limitation in summer), constraining seedling establishment and reproductive performance (Giménez-Benavides et al., 2007a, 2011b; Lara-Romero et al., 2014). Our aim was to assess the role of geographic isolation (IBD), landscape features (IBR) and environmental heterogeneity (IBE) in shaping the genetic diversity and structure of S. ciliata. We specifically addressed three main questions: (i) Are S. ciliata populations genetically structured across mountain ranges and elevational gradients? (ii) If so, can observed genetic structure patterns be explained by isolation-by-distance (IBD), isolation by resistance (IBR) and/or isolation-by-environment (IBE)? and (iii) How do these factors affect genetic diversity?
Most genetic approaches are addressed to species with limited distribution ranges, whereas more widespread species have not deserved so much attention from the conservation genetics community. The results of this study will provide useful information for the conservation of the genetic variation of a dominant species of the endangered Mediterranean alpine ecosystems (Tutin et al., 1964; Kyrkou et al., 2015) in one of its southernmost distribution limits.
Silene ciliata Poiret (Caryophyllaceae) is a dwarf cushion perennial plant which inhabits Mediterranean alpine habitats with marked environmental gradients. It is pollinated by nocturnal insects, mainly belonging to the genus Hadena (Lepidoptera, Noctuidae), and diurnal insects (Giménez-Benavides et al., 2007b). The flowering period spans from the end of July to the end of August. Seeds are relatively small (mean ± SD: 1.53 ± 0.49 mm diameter, 0.59 ± 0.06 mg weight), and most of them are dormant and need cold stratification to germinate (Giménez-Benavides et al., 2005; García-Fernández et al., 2015). The species is essentially barochorous, i.e., seeds lack any specific structure to promote dispersal. Thus, effective seed dispersal distances are low and relatively invariant across populations (mean ± SE: 0.40 ± 0.08 m, Lara-Romero et al., 2014). The distribution range of S. ciliata comprises the mountain ranges of the Northern Mediterranean area from Spain to Bulgaria (Tutin et al., 1964; Kyrkou et al., 2015), reaching its southernmost limit in the Sistema Central of the Iberian Peninsula (Figure 1). S. ciliata populations from the Sistema Central have the same phylogenetic origin as shown by chloroplast DNA analysis (Kyrkou et al., 2015). In these areas the species grows from 1900 to 2590 m in dry cryophilic pastures above the tree line. This Mediterranean Alpine ecosystem presents a pronounced summer drought combined with high solar radiation which induces typical xerophilic characteristics in the inhabiting species (Rivas-Martínez et al., 1990). The study took place in three mountain ranges of the Sistema Central (Spain): Guadarrama (GDM), Béjar (BJR) and Gredos (GRD) (Figure 1 and Table 1). The Sistema Central is a southwest-northeast oriented mountain range of approximately 500 km located in the center of the Iberian Peninsula. The GDM, GRD and BJR mountain ranges are located in the western, central and eastern areas of the Sistema Central, respectively.
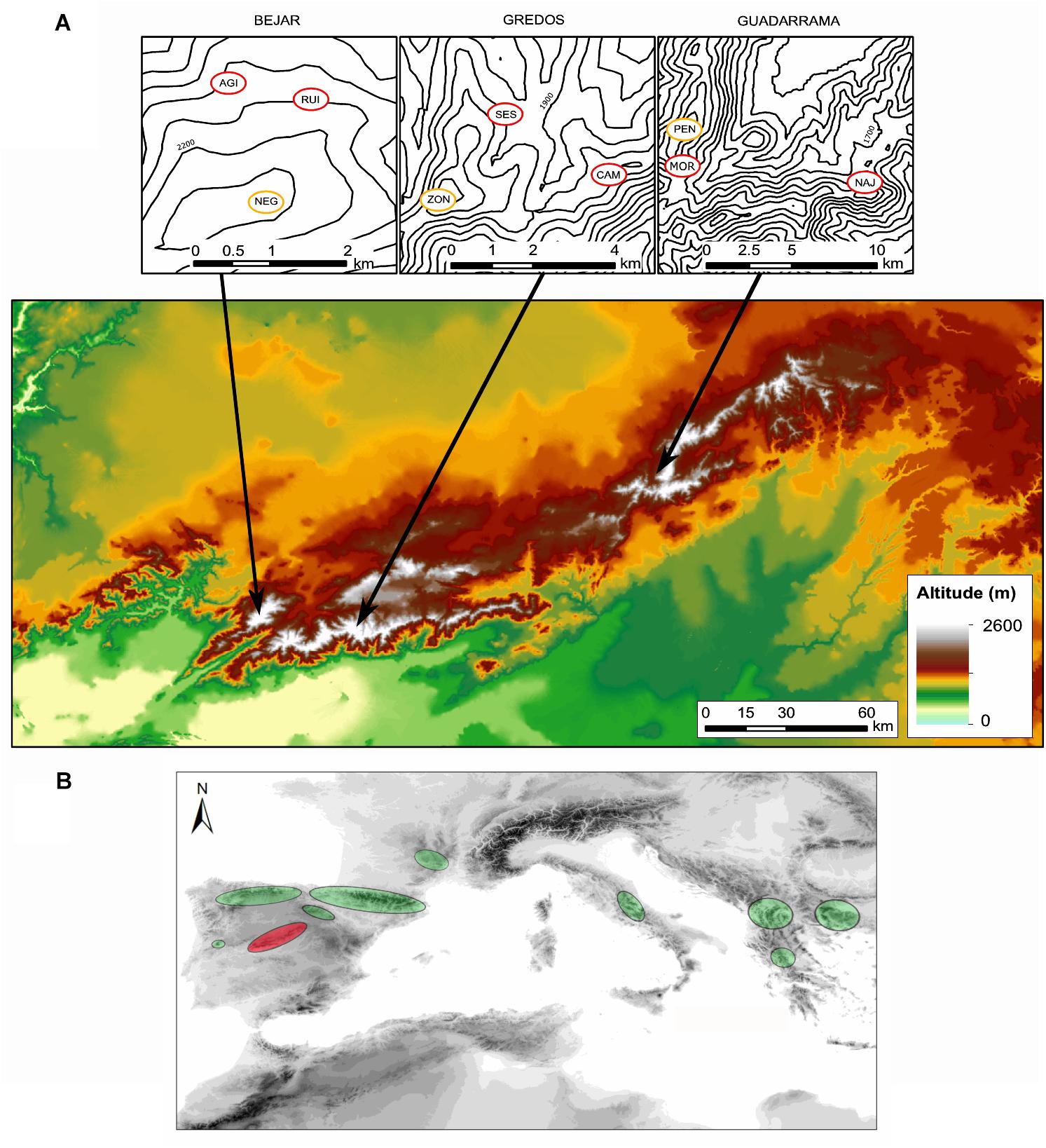
FIGURE 1. (A) Location of the study sites in the Sistema Central of the Iberian Peninsula. Yellow circles represent each of the three populations located in optimal areas and red dots represent each of the six populations located in marginal areas (see Table 1). NAJ, Najarra Baja; MOR, Morrena Peñalara; PEN, Pico Peñalara; SES, El Sestil; CAM, Los Campanarios; ZON, Altos del Morezón; RUI, Las Cimeras; AGI, Pico del Aguila; NEG, Canchal Negro. (B) Representation of Silene ciliata distribution area. Red circle indicates the Sistema Central of the Iberian Peninsula, where our study takes place.
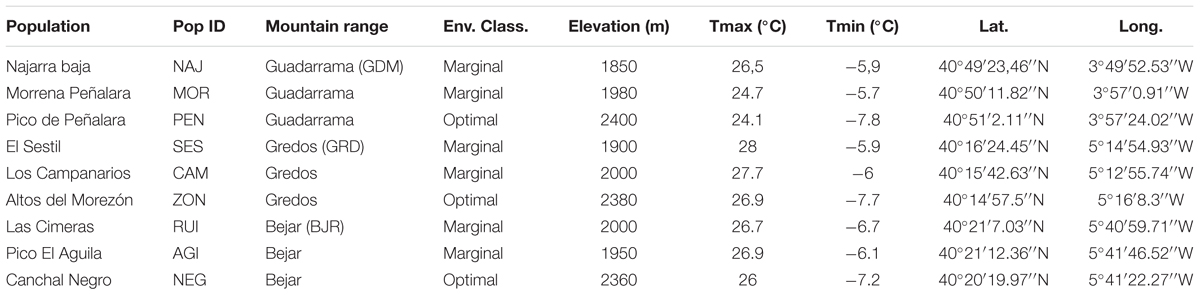
TABLE 1. Geographic and environmental features of nine sampled populations of Silene ciliata. Env. Class., populations environmental classification; Tmax, Annual maximum temperature; Tmin, annual minimum temperature.
We characterized the quality of the habitats of the study area by generating a niche model. We used MAXENT algorithm (Phillips et al., 2006) to generate a model for the Sistema Central according to the species environmental requirements (Morente-López et al., unpublished data). Minimum annual temperature (MAT), precipitation of the driest month (PPd), medium annual snowpack (SP) and medium annual potential evapotranspiration (PET) were used to build the model. We used climatic data from the Atlas Climático de la Península Ibérica with a 200-meter resolution (Ninyerola et al., 2005). Medium annual snowpack was calculated following the methodology proposed by López-Moreno et al. (2007) and the rest of the environmental variables were calculated using ENVIREM R package (Title and Bemmels, 2018). Thus, according to this model, we defined the higher-quality habitats as “Optimal” and the lower-quality habitats as “Marginal” following the definition of environmental marginality (Soule, 1973; Kawecki, 2008). “Optimal” areas were those with habitat suitability values in the highest 33rd percentile of the distribution. The lower values of MAT and PET and the higher values of SP and PPd define these areas. “Marginal” areas were defined as those with habitat suitability values in the lowest 33rd percentile of the distribution. This classification is congruent with demographic trends obtained by Giménez-Benavides et al. (2011a). We selected three S. ciliata populations in each of the three mountain ranges, one located in an optimal area and two in marginal areas. The names, location and ecogeographic characterization of the nine studied populations are shown in Table 1.
We collected S. ciliata leaf tissue from 20 individuals per population for genetic analysis (n = 180). DNeasy Plant minikit (QIAGEN, Valencia, CA, United States) was used for DNA extraction of 10–20 mg of dried S. ciliata tissue. Based on a previous study (García-Fernández et al., 2012a), we selected eight microsatellite loci for genotyping: Sci1224, Sci1208, Sci0106, Sci1443, EST-2HTS, EST-37HTS, EST-G34D06 and EST-G47A02. PCR protocols were performed as described in García-Fernández et al. (2012b). We genotyped all samples in an automated DNA sequencer (ABI PRISM 3730, Applied Biosystems, Foster City, CA, United States) in the Unidad de Genómica (Universidad Complutense de Madrid, Spain). GeneMarker version 1.85 (SoftGenetics, State College, PA, United States) was used to determine fragment size. We evaluated genotyping accuracy by re-amplifying and re-scoring 20% of the samples (N = 36). MICRO-CHECKER (Van Oosterhout et al., 2004) was used to assess the frequency of null alleles, and allelic dropout. No allelic dropout was found. We detected null alleles in most of the populations, but as they were not related to a particular locus across populations, we kept all the markers. We tested for linkage disequilibrium across loci and populations (Log-likelihood ratio G statistic based on 5000 permutations performed in GENEPOP v. 4.1, Rousset, 2008). Only one locus presented significant linkage disequilibrium across populations. However, we decided to include it in the analysis as the disequilibrium was only present in two of the nine populations.
To characterize the study populations, we calculated the following estimators of genetic diversity across populations: (i) total number of alleles (Na); (ii) number of private alleles (PA); (iii) mean number of alleles per locus (A); (iv) observed heterozygosity (Ho); (v) expected heterozygosity (He); and (vi) inbreeding coefficients Fis defined as [1-(Ho/He)] and (v) Fi, calculated as the probability that the two alleles at a locus are identical by descent following the definition of Malécot and Blaringhem (1948). We applied the methodology proposed by Chybicki and Burczyk (2008) using the Bayesian approach implemented in INEST 2.2 software to calculate Ho, He and inbreeding coefficient Fi corrected for null alleles. We also tested for deviances from Hardy–Weinberg equilibrium (HWE) per locus in each population following Haldane (1954) and for all populations using Fisher’s exact test. Significance of the latter was assessed with Monte Carlo tests using 2000 iterations. Analyses were performed with diveRsity v. 1.9.90 R package (Keenan et al., 2013) implemented in R (R Core Team, 2009).
In order to study the contribution to the genetic structuring of the different hierarchical levels in our data set (populations, mountain ranges and environments), we tested for genetic differentiation measured as Fst across three different hierarchical configurations: (i) genetic differentiation among and within the nine populations, (ii) genetic differentiation among the three mountain ranges considering the genetic variance among populations within mountain ranges and within populations and (iii) genetic differentiation between environments (optimal vs. marginal) considering the genetic variance among populations within environments and within populations (Table 1). Fst values and 95% confidence intervals were estimated by 1000 randomizations of bootstrapping distance matrices. We performed the hierarchical genetic structures analysis across these organization levels using Hierfstats v. 0.04-22 R package (Goudet and Jombart, 2015). Considering the hierarchical structure of our sampling design, we also performed an analysis of molecular variance (AMOVA) with 9999 permutations, using GeneAlEx 6.5 (Peakall and Smouse, 2012) to contrast the results of the hierarchical Fst analysis.
We searched for genetic clusters across spatial scales by performing a Discriminant Analysis of Principal Components (DAPC) (Jombart et al., 2010) as implemented in adegenet R package (Jombart, 2008). In DAPC, discriminant functions are linear combinations of the variables (principal components of PCA) which optimize the separation of individuals into pre-defined groups (Jombart and Ahmed, 2012) determined using the K-means clustering algorithm. We used the find.clusters function (adagenet R package) to study the optimal number of clusters regarding the maximum drop in BIC values. We used the a-score to set the number of PCs retained in the DAPC to control the possible overfit, which is a measure of the trade-off between power of discrimination and over-fitting the model (Jombart and Ahmed, 2012).
We also applied a Bayesian clustering method as implemented in STRUCTURE v 2.3.4 (Pritchard et al., 2000). We performed ten independent runs for each possible number of K clusters from one to nine. Each run assumed a burn-in period of 106 iterations, followed by 107 MCMC iterations considering the model of correlation frequencies and admixture origin. To elucidate the most plausible value of K, we followed the approach described in Evanno et al. (2005) implemented in Structure Harvester (Earl and vonHoldt, 2012). Then, we used Clumpp v. 1.1.2 (Jakobsson and Rosenberg, 2007) to obtain the permuted membership coefficient of each individual assigned to each cluster, joining the results of the 10 independents runs. The output from Clumpp was visualized by Distruct v 1.1 (Rosenberg, 2004).
A Geneland analysis (Guillot et al., 2005) was also developed considering five runs of 106 MCMC iterations after a burning process of 105 simulations, sampling each 1000 steps, ranging K values between 1 and 10 and applying spatial and null alleles corrections, to confirm analysis made with DAPC and STRUCTURE.
We wanted to elucidate whether the observed genetic structure was caused by geographical distance (IBD), environmental conditions (IBE), geographical conformation of the landscape (IBR) or a combination of these factors. To achieve this, we first obtained four different types of distance matrices:
(i) Genetic distance matrix, calculated as pairwise Fst distance between the nine populations using FreeNA (Chapuis and Estoup, 2007) to correct for the presence of null alleles; (ii) Geographic distance matrix, based on Euclidean distances between populations. Genetic and Euclidean distance matrices were transformed [Fst/(1-Fst)] and log(Euclidean distance), respectively, to linearize their relationship (Rousset, 1997); (iii) The environmental distance matrices were created with 200-meter resolution data from the Atlas Climático de la Península Ibérica (Ninyerola et al., 2005). As annual maximum and minimum temperatures (Tmax and Tmin, respectively) have been proposed as reference variables of environmental gradients in alpine ecosystems (Totland, 1999; Totland and Alatalo, 2002; Körner, 2003, 2007), we selected Tmax and Tmin as proxies of the environmental gradients of the study territory. We previously tested them for collinearity by checking if the variance inflation factor (VIF) was below 2 (Chatterjee and Hadi, 2015); and (iv) Least-cost distance matrix between each pair of populations was calculated in terms of the cost of effective migration from one population to another, using a digital elevation model (DEM) with 200-m spatial resolution (Ninyerola et al., 2005) as a proxy. Taking into account the biological features of this alpine species, effective gene flow was only considered to be possible along the areas of the Sistema Central with the highest elevations. Thus, 1280 m a.s.l. was selected as the threshold below which the species could not migrate. The selected threshold ensures connectivity among the three mountain ranges. Least-cost values were determinate by determined for each pair of populations as the accumulated cost value of all cells to be crossed. Cell cost values express cumulative cost of movement in terms of distance equivalence. Distances were measured according to the minimum amount of friction that must be accumulated to move from one population to another target population. Movements were allowed for the standard eight directions from any cell. Least-cost distances were calculated in IDRISI Selva V. 17 (Eastman, 2012).
Since the adequacy of simple Mantel tests has been considerably criticized in the past for its proneness to type I error (e.g., Manel and Holderegger, 2013) we used reciprocal causal modeling (RCM) to control for Mantel test proneness to spurious correlations (Cushman and Landguth, 2010) and to assess the relevance IBD, IBE and IBR patterns in our study case. We followed the methodology proposed by Cushman et al. (2006, 2013b) and applied by Ruiz-Gonzalez et al. (2015).
Reciprocal causal modeling uses pairs of reciprocal partial Mantel tests to study the relative support of alternative genetic configuration hypotheses (IBD, IBE, IBR). First, the partial Mantel correlation between one of the hypothesis (e.g., IBD) and the genetic distance (G.Dist), controlling for the effect of a second hypothesis (e.g., IBR) was calculated using partial Mantel test (G.Dist ∼ IBD| IBR). Second, a second partial Mantel test was developed but calculating the correlation between the genetic distance and the second hypothesis, controlling for the effect of the first hypothesis (G.Dist ∼ IBR| IBD). The relative support for IBD (focal model) relative to IBR (alternative model) is the difference between the partial correlations of the two tests (IBD| IBR-IBR| IBD) and vice versa (Cushman et al., 2013a,b). Thus, if IBD hypothesis is correct then IBD|IBR – IBR|IBD should be positive and IBR|IBD – IBD|IBR zero or negative, and, conversely, if IBR hypothesis is correct, then IBD|IBR – IBR|IBD should be zero or negative and IBR|IBD – IBD|IBR positive. Following this methodology a full matrix of all the possible hypothesis comparisons was calculated (reciprocal causal modeling matrix). If for a hypothesis all values in a column are positive and all associated values in a row are negative, then that model is fully supported and, thus, such hypothesis is the best compared to all alternatives. For each of the Mantel tests hypothesis combinations we also calculated the correlation values and significance through corrected permutation tests with 9999 permutations. Analyses were carried out using adegenet (Jombart and Ahmed, 2012), ade4 (Dray and Dufour, 2007), ecodist (Goslee and Urban, 2007) and stats (Pritchard et al., 2000) R packages.
To study the possible relationship between genetic diversity estimators and geographical and environmental variables, we also developed different geographically explicit generalized linear mixed models (spatial GLMM’s) using genetic, geographical and environmental information. The models were built using spaMM R package (Rousset and Ferdy, 2014). We used different genetic diversity estimators (Ho, He, Fis, Fi, A and PA) as dependent variables and annual Tmax and Tmin as independent variables, considering the geographical coordinates of the locations a random factor. To account for non-linear responses of the environmental variables, we also tested models including their squared values. We tested for normality and homoscedasticity of the model residuals and made the necessary transformations when required. We also tested the possible spatial autocorrelation of the residuals performing a Moran’s I test (Moran, 1950). For each model developed, a P-value of an associated likelihood ratio test (LRT) between the “full” model (including environmental variables) and the “null” model (only with spatial random effect) tested the effect of a given factor after applying the Bonferroni correction. We also performed an AIC rank test to select the best fitting model when more than one environmental variable had a significant effect on a genetic diversity estimator and to ensure the improvement of the “full” versus the “null” model.
The eight microsatellites scored a total of 107 different alleles across all individuals with an average of 5.2 alleles per locus. Number of alleles per population (Na) varied from 38 to 57 and number of alleles per locus from 4.30 to 6.05 (see Table 2). Populations from Guadarrama (MOR, NAJ and PEN) had the lowest number of private alleles (PA) compared to the populations from the other two mountain ranges (Kruskal–Wallis test, P = 0.02). Observed heterozygosity values (Ho) were lower than expected heterozygosity values (He) for all populations except MOR where Ho was higher than He. Populations NAJ and SES had similar Ho and He values. All populations except NAJ and PEN (GDM) departed from the H&W equilibrium, and they showed a significant excess of homozygotes across loci (Table 2). Inbreeding coefficients were significantly higher than zero in all populations (P < 0.05) except MOR (Fis = -0.08), NAJ (Fis = -0.01) and SES (Fis = -0.01), which were not significantly different from zero (P > 0.05 in all cases).
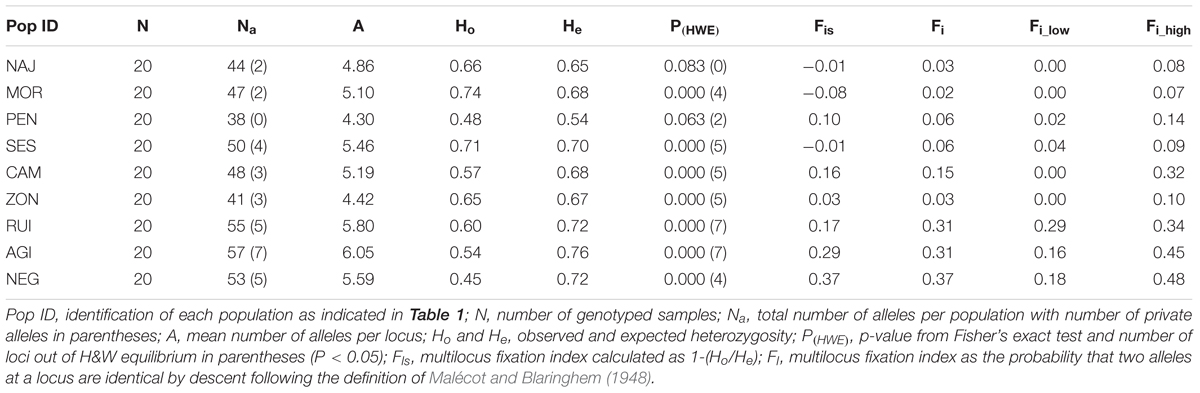
TABLE 2. Estimators of genetic diversity at the population level, fixation indexes, and Hardy-Weinberg exact tests in studied Silene ciliata populations.
When testing for population structure, hierarchical Fst analysis showed that genetic differentiation was higher among individuals within populations (FInd/Pop) (Mean [CI]: 0.26 [0.11, 0.41]) than among populations (FPop/T) (0.09 [0.06, 0.15]), although both were significant. The Fst hierarchical structure analyses of population nested within mountain ranges showed that all levels contributed to the hierarchical genetic structure with different magnitudes. Mountain range (FMt/T) (0.05 [0.006, 0.11]) and population nested within mountain range (FPop/Mt) (0.06 [0.04, 0.08]) showed a small but significant value. Individuals within populations showed the greatest genetic divergence (FInd/Pop/Mt) (0.26 [0.10, 0.42]). The Fst hierarchical structure analyses of population nested within environment found no significant effect of environment (FEnv/T) (0.0 [-0.02, 0.02]) and a small significant value for population nested in environment (FPop/Env) (0.09 [0.06, 0.15]). As in the Fst hierarchical structure analyses, individuals within populations had the greatest genetic divergence values (FInd/Pop/Env) (0.26 [0.11, 0.40]). Results are summarized in Table 3. AMOVA results showed very similar patterns of molecular variance partition (see Supplementary Table S1).
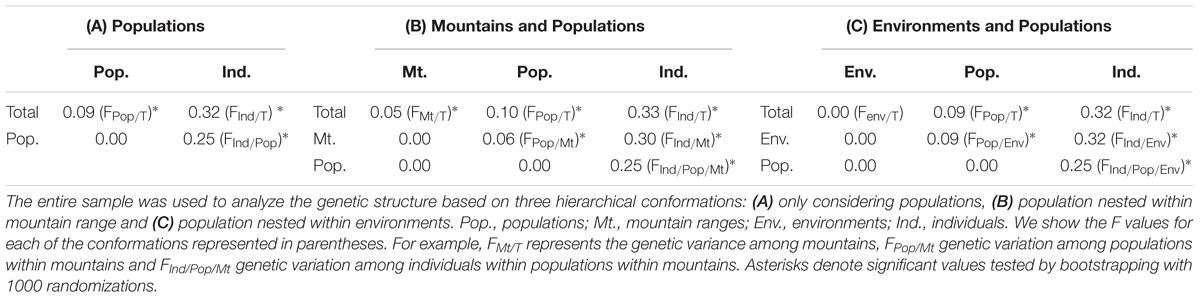
TABLE 3. Hierarchical Fst results.
In the DAPC clustering analysis, we selected K = 2 and K = 3 structures based on the BIC curve which represents the plausible number of clusters in the data (Supplementary Figure S1) and the biological meaning of our study case. In the K = 2 partition, individuals were neatly classified into two groups; one mainly composed of individuals from the GDM mountain range and the other of individuals from GRD and BJR (Figures 2A,B). K = 3 genetic data partition showed one group mostly composed of individuals from the three populations of the GDM mountain range. The other two groups were essentially a mixture of individuals from populations of the GRD and BJR mountain ranges (Figures 3A,B).
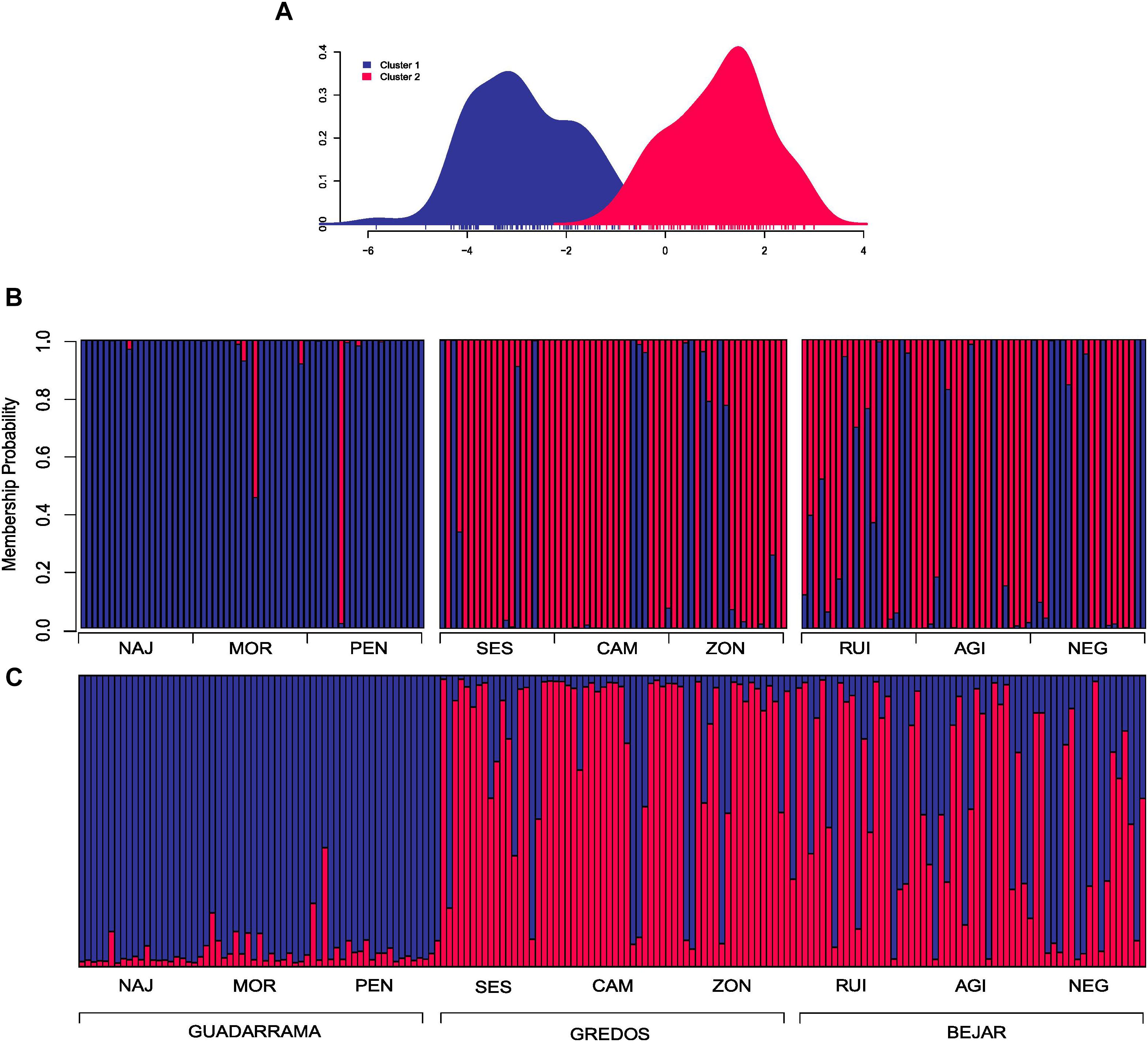
FIGURE 2. Discriminate analysis of principal components (DAPC) and Bayesian analysis of population structure (STRUCTURE) for two clusters conformation (K = 2). (A) Scatterplot of the first two principal components showing the differentiation between the two groups by colors. (B) DAPC composition plot (compoplot) of each individual grouped by mountain ranges. Colors represent the same clusters as the scatterplot. (C) STRUCTURE composition plot (compoplot) of each individual grouped by mountain ranges.
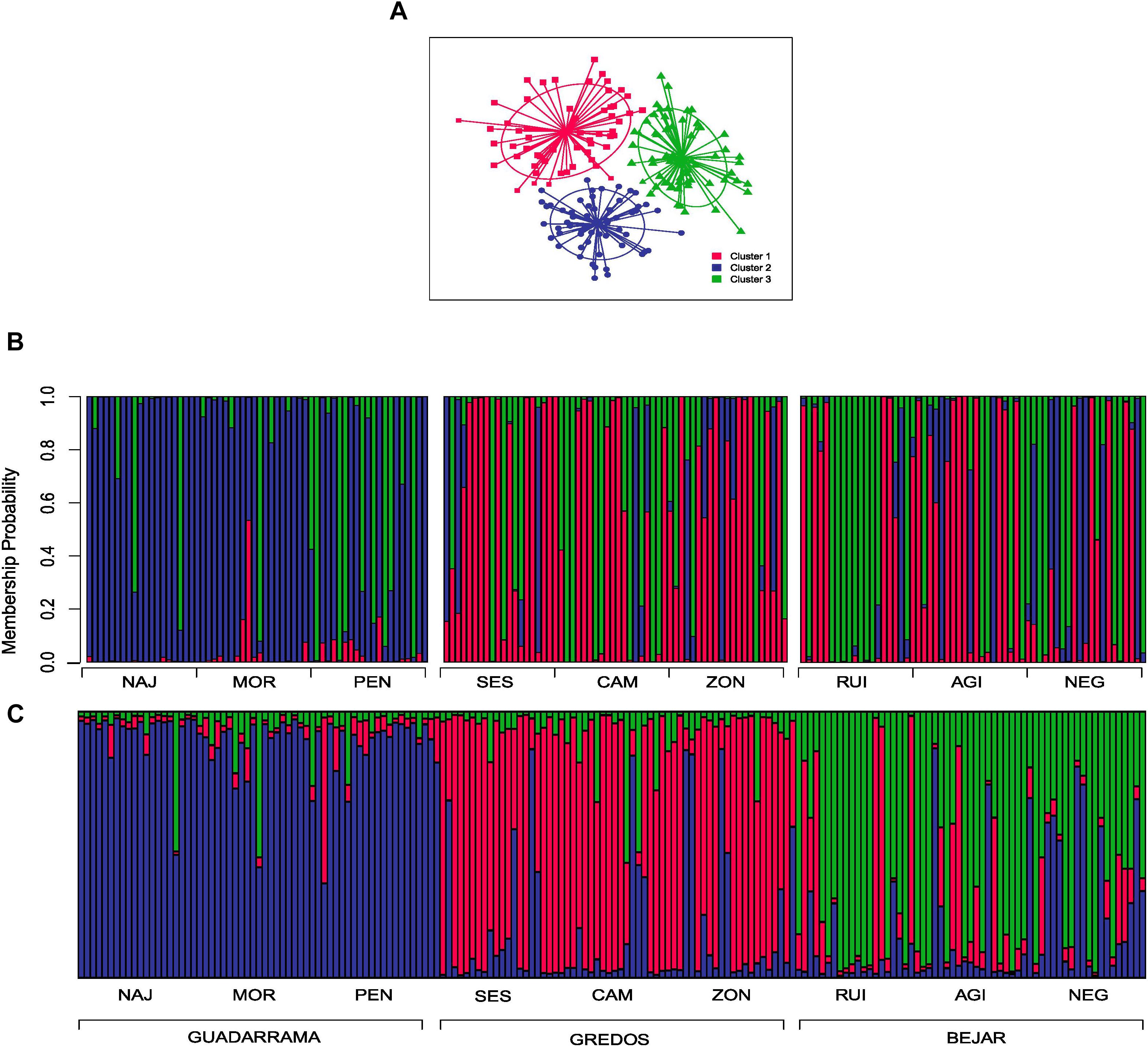
FIGURE 3. Discriminate analysis of principal components (DAPC) and Bayesian analysis of population structure (STRUCTURE) for K = 3 clusters. (A) Scatter plot of the first two principal components showing the differentiation between the three groups by colors and inertia ellipses. Dots, squares and triangles represent individuals of each cluster. (B) DAPC composition plot (compoplot) of each individual grouped by mountain ranges. Colors represent the same clusters as the scatterplot (C) STRUCTURE composition plot (compoplot) of each individual grouped by mountain ranges.
STRUCTURE showed that K = 2 best explained the genetic structure of the study data set (Supplementary Figure S2), separating individuals from the GDM mountain range and individuals from the GRD and BJR mountain ranges (Figure 2C). These results agree with those obtained in DAPC. The second most plausible structuring of the data was K = 3, with one group corresponding to each mountain range (GDM, BJR and GRD) (Figure 3C). Geneland results were consistent with the genetic structure founded with DAPC and STRUCTURE analysis (Supplementary Figure S3).
IDB arises as the strongest overall hypothesis regarding to the relative support values of the RCM matrix (Table 4A), presenting positive values in the entire column and negative values in the entire row. In a similar way, all partial Mantel tests values relates with this variable showed a significant and positive correlation between genetic and Euclidian distance (Table 4B). Regarding to the IBR hypothesis, negative relative support value arises when it was controlled by IBD hypothesis, but positive values emerge when it was controlled by IBE hypotheses (Table 4A). Weak and no significant correlation was found when partial Mantel test related with this variable was controlled by the Euclidean distance (Table 4B). Both IBE hypotheses are the ones with less support regarding to the RCM matrix and with none significant Mantel correlation values.
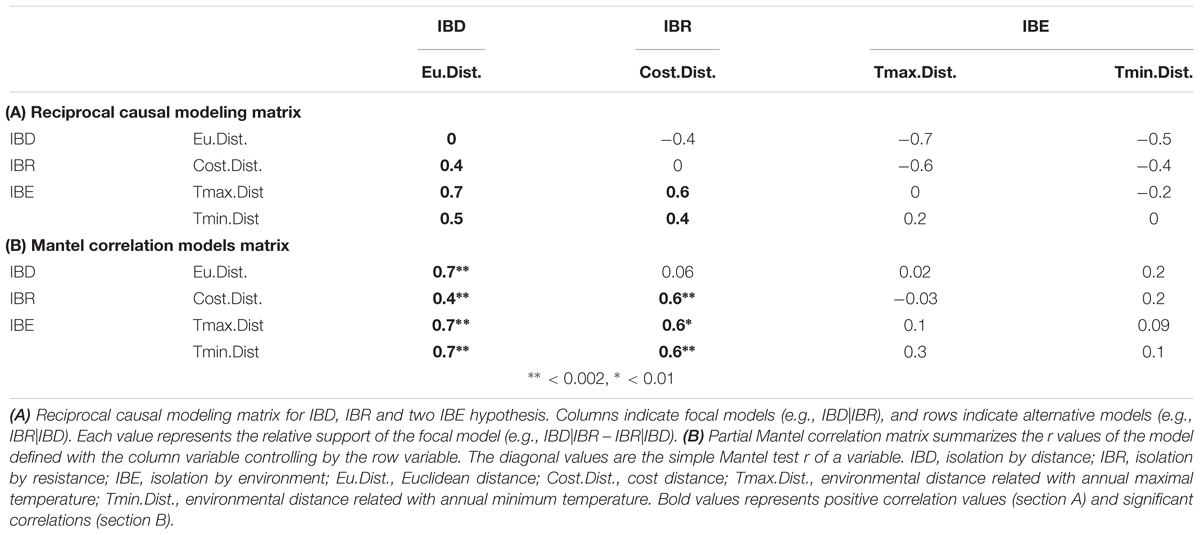
TABLE 4. Reciprocal causal modeling results.
Spatially explicit mixed models found a positive linear relationship between annual minimum temperatures (Tmin) and Ho (β = 0.009, df = 4.3, P < 0.05) and He (β = 0.004, df = 6.3, p < 0.05) and a quadratic relationship between Tmin and mean number of alleles per locus (A) (β = -0.0003, df = 4.6, P < 0.001) (Figure 4 and Table 5). Notably, intermediate levels of Tmin exhibited the highest levels of A. Maximum annual temperature (Tmax) was not related to Ho, He or A. The addition of Tmin as an explanatory variable significantly improved AIC values in the models compared to the “null” models (Table 5). No relationship was found between any of the environmental variables and Fi, Fis or PA. No significant spatial autocorrelation of the residuals was found.
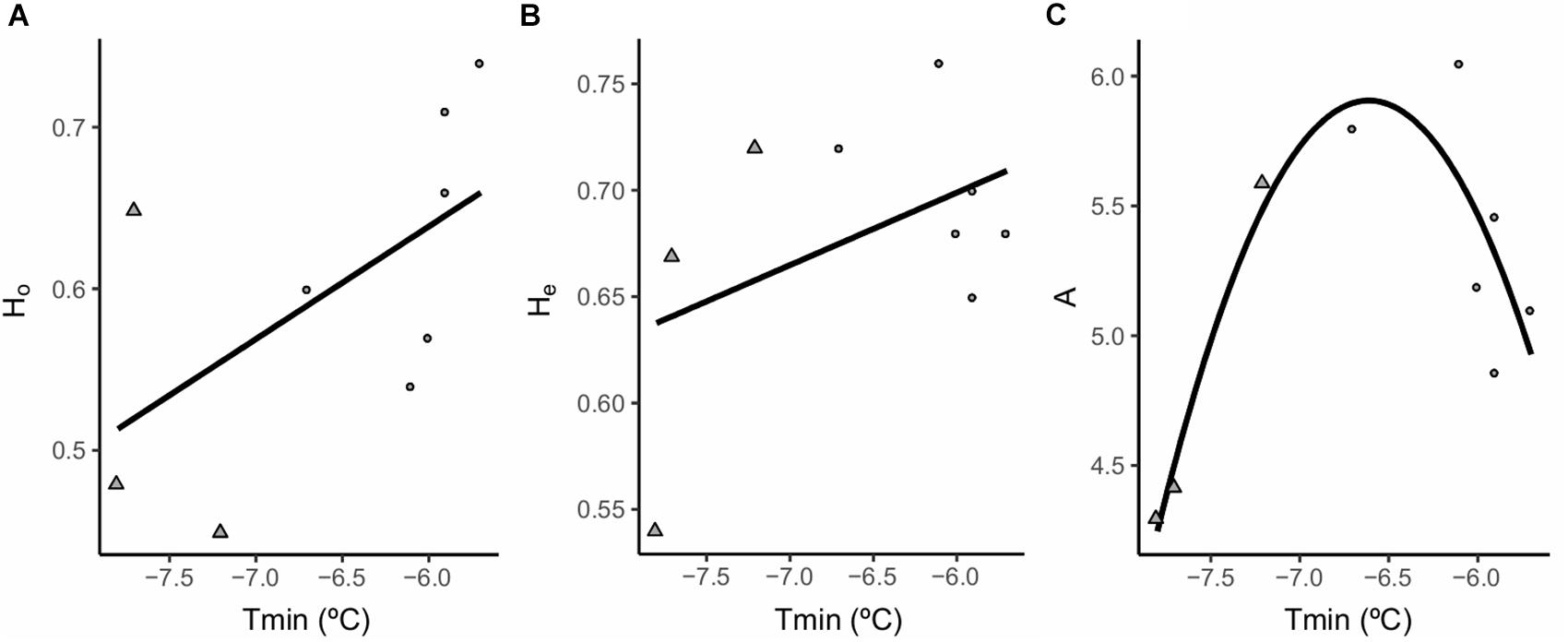
FIGURE 4. Significant relationships between the different genetic diversity estimators and the environmental variables used in the models. (A) Ho, observed heterozygosity; (B) He, expected heterozygosity; (C) A, mean allelic richness per locus (average number of alleles per locus). Tmin; minimum annual temperature. Dots represent populations classified as “marginal” and triangles populations classified as “optimal.”
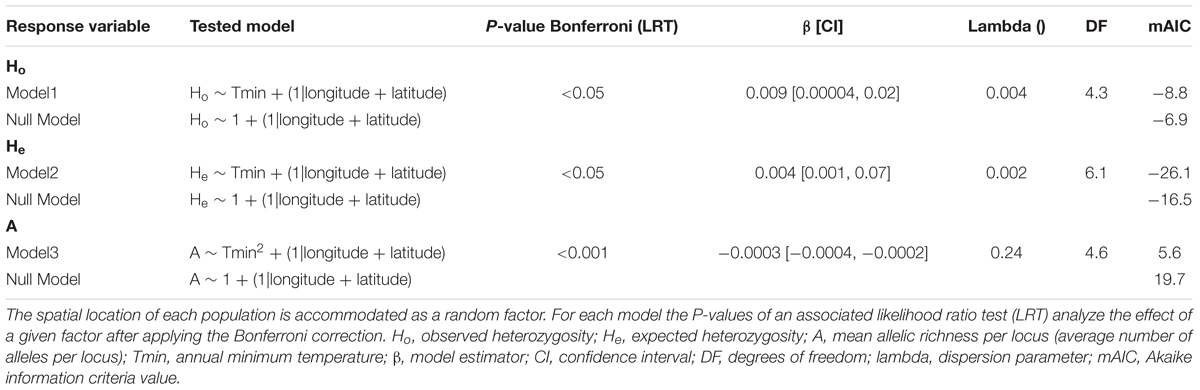
TABLE 5. Mixed effect models fitted to test the effect of environmental factors (fixed factors) in determining different estimates of genetic diversity across populations.
Results depict a complex scenario in which geographical and environmental factors influence the genetic structure and diversity of S. ciliata in the Central System of the Iberian Peninsula. S. ciliata presented a marked genetic structure, and IBD was the main factor shaping genetic patterns. No genetic structure was found between populations from optimal and marginal habitats suggesting no isolation effects related to environmental differences. However, a significant relationship was found between environmental variables and genetic diversity.
Hierarchical analyses showed that most of the genetic diversity resided within populations. This is probably due to the species’ breeding system which favors allogamous crossings (Giménez-Benavides et al., 2007b). It also reflects the presence of significant within-population genetic structure due to limited gene flow (Lara-Romero et al., 2016b), which is shaped by limited effective pollen and seed dispersal distances (Lara-Romero et al., 2014, 2016a,b). A significant genetic variation component between populations within mountain ranges was also observed, but no genetic differentiation was found when we contrasted populations inhabiting optimal and marginal habitats across mountain ranges. This suggests that genetic differentiation between populations within mountain ranges is found between marginal habitats. Hierarchical Fst found significant differentiation between mountain ranges of similar magnitude to that between populations within mountain ranges, which was further supported by clustering analyses. GDM was more differentiated than the other two mountain ranges (BEJ and GRE) as shown by the K = 2 conformation in the clustering analysis. On the other hand, the K = 3 conformation was clearly related to the three mountain ranges in STRUCTURE, and with an admixture between BEJ and GRE in DAPC. These results show the relevance of the spatial configuration of the landscape at the mountain range level on the genetic structure. This could be related with the colonization pattern of the species along the East-West oriented axis of the Sistema Central (see Figure 1). Patterns of genetic variation across landscapes in alpine ecosystems are diverse and context dependent. Geng et al. (2009) found short-distance genetic structure patterns related to limited gene dispersal along with substantial levels of gene flow and slow rates of genetic drift between topographically separated populations. Many other authors reported genetic structure at larger landscape scales (e.g., Aægisdóttir et al., 2009; Lega et al., 2014) related to natural barriers to gene flow. In our case, we detected both coarse-scale mountain range clustering and finer-scale short distance population differentiation. Our results suggest a prevalence of gene flow limitation between mountain ranges, with considerable genetic differentiation among populations and mountain ranges.
The significant relationship between geographical distance and genetic differentiation based on RCM and Mantel correlations suggested an IBD pattern between S. ciliata populations. IBD is the most common pattern of genetic differentiation in landscape genetics studies (Jenkins et al., 2010; Cushman et al., 2015), including alpine plants (e.g., Stöcklin et al., 2009; Wu et al., 2015). Nevertheless, the IBD hypothesis cannot be generalized in alpine ecosystems (Frei et al., 2012a,b). When we considered the topography of the study territory, we also found an IBR pattern showing a significant effect of cost distances in the genetic differentiation, although weaker than the IBD. This is not surprising since the IBR calculated based on the DEM includes a distance effect. The East-West alignment of the mountain ranges which conforms the Sistema Central (see Figure 1) includes passages of lower elevation that connect the mountain ranges and act as topographic barriers. This effect works in the same direction as the isolation effect imposed by the Euclidean distance (IBD). Thus, the East-West orientation of the mountain ranges and the associated barriers seem to be important drivers of the genetic structure at the landscape scale in our study.
No genetic structure associated to environmental differentiation was detected using RCM and partial Mantel correlations, or the genetic structure clustering and hierarchical Fst analyses approaches. Previous S. ciliata field studies carried out in similar areas found evidence of local adaptation in marginal populations, i.e., seed germination success and survival compared to optimal populations (Giménez-Benavides et al., 2007a,b). They also inferred a putative genetic isolation pattern between optimal and marginal populations because of mismatched flowering periods (Giménez-Benavides et al., 2007b, 2011b), measured under field conditions. Restriction of gene flow due to phenological mismatches, and possible differential selection along gradients, may cause genetic differentiation (Hirao and Kudo, 2004). However, a subsequent genetic study carried out in GDM populations (García-Fernández et al., 2012b) found significant gene flow along an elevational gradient and low genetic differentiation. Using AFPLs, Manel et al. (2012) showed that environmental variables are drivers of plant adaptation at the scale of a whole biome for a large number of alpine species. Furthermore, several studies in mountain ecosystems have found adaptive variation patterns using genomic approaches (e.g., Poncet et al., 2010; Mosca et al., 2012, 2014; Di Pierro et al., 2017). These previous results in our and other alpine species put us on the track of adaptive divergence between populations. The lack of an environmental signature in the genetic differentiation found in our study may result from the low number of neutral molecular markers used and does not preclude environmentally induced genetic signals in other areas of the S. ciliata genome. IBE patterns can be detected using neutral markers because the signature of selection extends to genome areas beyond the genes that are under selection (Shafer and Wolf, 2013; Sexton et al., 2014; Kern and Hahn, 2018), but they are certainly more difficult to find when the number of loci used is low. Further research using genomic and quantitative genetic approaches is needed in Mediterranean alpine ecosystems to provide insight into the identification of genetic differentiation patterns related to adaptation along environmental gradients.
In contrast with the lack of an environmental signature on genetic differentiation, we found a significant relationship between environmental variables and genetic diversity. Under an ecological niche perspective of the central-marginal model, optimal environmental conditions in the central areas allow larger and more stable populations with greater genetic diversity (Kawecki, 2008; Sexton et al., 2009). Conversely, populations in marginal habitats tend to be small and fluctuating in size and, therefore, prone to suffer bottleneck processes that causes genetic diversity erosion (Glémin et al., 2003; Whitlock, 2004; Kawecki, 2008). Consequently, genetic diversity is expected to be higher in optimal populations (Eckert et al., 2008; Pironon et al., 2015).
Contrary to our expectations, the lower Tmin values which entail optimal conditions for the species (Giménez-Benavides et al., 2007a, 2011a; Lara-Romero et al., 2014) were related to lower genetic diversity values. This apparent contradiction may be partially explained by a historical signature of genetic diversity patterns that overrides the effects of current conditions. The effects of glacial cycles have often been used to explain distributional shifts of species, as well as the contraction, fragmentation and connectivity of mountain populations (Marques et al., 2016). Thus, glacial pulses would have been responsible for shifting S. ciliata populations to lower elevations, allowing populations from nearby mountains to be connected (Hewitt, 2001; García-Fernández et al., 2013). As a result, these lower areas would have increased their genetic diversity and acted as a genetic reservoir for the species (Knowles, 2001; Tzedakis et al., 2002; Holderegger and Thiel-Egenter, 2009). Postglacial recolonization of higher altitudes from lower areas (Dechaine and Martin, 2004) would have originated a front advancing population edge by a subsample of the population with lower genetic diversity.
Ho and He values were the highest in the marginal populations at the lowest elevations, but allelic richness decreased sharply compared to populations at intermediate elevations and lower annual minimum temperatures fitting a quadratic response. The low population size and possible bottlenecks experienced by the most marginal populations, potentially associated with current high annual minimum temperatures may be responsible for this. This mismatch between allelic richness and heterozygosity values could be related to the impact of population bottlenecks on allelic diversity which is often greater and faster than on heterozygosity (Frankham et al., 2002).
We, thus, suggest that the observed genetic diversity pattern is a combination of present and past climatic factors and events. Present marginal conditions associated with lower elevations resulting from altitudinal shifts and climate warming may overlay a genetic diversity pattern that stems from the glacial pulses of the past involving both environmental and geographic factors. In the central-marginal model context, associated genetic diversity patterns can be highly context and scale dependent (Hampe and Petit, 2005; Hardie and Hutchings, 2010). Geographical, ecological and historical gradients act in conjunction creating diverse patterns that cannot be homogenized under a common rule (Sagarin and Gaines, 2002; Eckert et al., 2008; Duncan et al., 2015; Pironon et al., 2017).
Our results highlight the complexity of the patterns shaping the spatial distribution of genetic variation of plants inhabiting high mountain ecosystems. IBD arises as the main pattern shaping genetic structure in mountain ecosystems. In addition, IBR emerged as another important pattern shaping genetic structure although weaker than IBD when geographical distance and barriers works in the same direction. IBE should be considered as an important force in shaping genetic variation, especially in steep environmental ecosystems like mountainous regions. Present and past changes in environmental conditions inside distribution ranges strongly affect genetic diversity in alpine species.
The results of this study can be useful in a future comparison using populations inhabiting similar environmental gradients in other areas of the species distribution not represented here. This would warrant a more thorough perspective of the main drivers shaping plant populations genetics in widespread Mediterranean Alpine plants. Additional research using genomics and quantitative genetics arise as the path to further understand the variation patterns linked to Mediterranean alpine environments.
The dataset generated and used in this publication can be found in the Universidad Rey Juan Carlos data repository e-cienciaDatos (https://edatos.consorciomadrono.es/dataset.xhtml?persistentId=doi:10.21950/GSTZ26) doi: 10.21950/GSTZ26.
JM-L, AG-F, CL-R, and JI designed this study. JM-L and AG-F collected the samples and performed the laboratory work. JM-L, CG, AG-F, and DD analyzed the data. JM-L wrote the paper with the help of CG, CL-R, and JI. All authors reviewed the paper and approved the final manuscript.
This work was supported by the projects AdAptA (CGL2012-33528) and EVA (CGL2016-77377-R) of the Spanish Ministry of Science and Innovation. JM-L was supported by a fellowship for the pre-doctoral contracts for the training of doctors (BES-2013-064951) of the Spanish Ministry of Science and Innovation. CG was supported by the FCT Investigador Program (IF/01375/2012). CL-R was supported by a Juan de la Cierva post-doctoral fellowship (MINECO: FJCI-2015-24712). DD was supported by the FCT Post-doctoral Fellowships Program (SFRH/BPD/100384/2014).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Sandra Sacristan, Anaís Redruello, Rocío Garrido, and Ana Ramirez for their help in the laboratory work. We thank Silvia Matesanz and Yurena Arjona for their helpful discussions and Greta Carrete Vega for her help in making Figure 1. We also thank Lori De Hond for linguistic assistance. We also thank INEST 2.2 developer Igor J. Chubrichi for the new implementation of heterozygosity estimators. Finally, we thank the staff of the Parque Nacional de Guadarrama (Dirección General de Medio Ambiente, Comunidad de Madrid) and the Delegación Territorial de Salamanca and Ávila (Servicio Territorial de Medio Ambiente, Junta Castilla y León) for permission to work in the field area.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01698/full#supplementary-material
Aægisdóttir, H. H., Kuss, P., and Stöcklin, J. (2009). Isolated populations of a rare alpine plant show high genetic diversity and considerable population differentiation. Ann. Bot. 104, 1313–1322. doi: 10.1093/aob/mcp242
Beniston, M., Diaz, H. F., and Bradley, R. S. (1997). Climatic change at high elevation sites: an overview. Clim. Change 36, 233–251. doi: 10.1007/978-94-015-8905-5_1
Brussard, P. F. (1984). Geographic patterns and environmental gradients: the central-marginal model in Drosophila revisited. Annu. Rev. Ecol. Syst. 15, 25–64. doi: 10.1146/annurev.es.15.110184.000325
Chapuis, M. P., and Estoup, A. (2007). Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 24, 621–631. doi: 10.1093/molbev/msl191
Chatterjee, S., and Hadi, A. S. (2015). Regression Analysis by Example. Hoboken, NJ: John Wiley & Sons.
Chybicki, I. J., and Burczyk, J. (2008). Simultaneous estimation of null alleles and inbreeding coefficients. J. Hered. 100, 106–113. doi: 10.1093/jhered/esn088
Cushman, S., Storfer, A., and Waits, L. (2015). Landscape Genetics: Concepts, Methods, Applications. Hoboken, NJ: John Wiley & Sons.
Cushman, S. A., and Landguth, E. L. (2010). Spurious correlations and inference in landscape genetics. Mol. Ecol. 19, 3592–3602. doi: 10.1111/j.1365-294X.2010.04656.x
Cushman, S. A., McKelvey, K. S., Hayden, J., and Schwartz, M. K. (2006). Gene flow in complex landscapes: testing multiple hypotheses with causal modeling. Am. Nat. 168, 486–499. doi: 10.1086/506976
Cushman, S. A., Shirk, A. J., and Landguth, E. L. (2013a). Landscape genetics and limiting factors. Conserv. Genet. 14, 263–274. doi: 10.1007/s10592-012-0396-0
Cushman, S. A., Wasserman, T. N., Landguth, E. L., and Shirk, A. J. (2013b). Re-evaluating causal modeling with mantel tests in landscape genetics. Diversity 5, 51–72. doi: 10.3390/d5010051
De Vriendt, L., Lemay, M. A., Jean, M., Renaut, S., Pellerin, S., Joly, S., et al. (2017). Population isolation shapes plant genetics, phenotype and germination in naturally patchy ecosystems. J. Plant Ecol. 10, 649–659. doi: 10.1093/jpe/rtw071
Dechaine, E. G., and Martin, A. P. (2004). Historic cycles of fragmentation and expansion in Parnassius smintheus (Papilionidae) inferred using mitochondrial DNA. Evolution 58, 113–127. doi: 10.1111/j.0014-3820.2004.tb01578.x
Di Pierro, E. A., Mosca, E., González-Martínez, S. C., Binelli, G., Neale, D. B., and La Porta, N. (2017). Adaptive variation in natural alpine populations of Norway spruce (Picea abies [L.] Karst) at regional scale: landscape features and altitudinal gradient effects. For. Ecol. Manage. 405, 350–359. doi: 10.1016/j.foreco.2017.09.045
DiLeo, M. F., and Wagner, H. H. (2016). A landscape ecologist’s agenda for landscape genetics. Curr. Landsc. Ecol. Rep. 1, 115–126. doi: 10.1007/s40823-016-0013-x
Dray, S., and Dufour, A.-B. (2007). The ade4 package: implementing the duality diagram for ecologists. J. Stat. Softw. 22:9461. doi: 10.18637/jss.v022.i04
Duncan, S. I., Crespi, E. J., Mattheus, N. M., and Rissler, L. J. (2015). History matters more when explaining genetic diversity within the context of the core-periphery hypothesis. Mol. Ecol. 24, 4323–4336. doi: 10.1111/mec.13315
Earl, D. A., and vonHoldt, B. M. (2012). STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 4, 359–361. doi: 10.1007/s12686-011-9548-7
Eastman, J. R. (2012). IDRISI Selva Manual and Tutorial Manual Version 17. Worcester, MA: Clark University.
Eckert, C. G., Samis, K. E., and Lougheed, S. C. (2008). Genetic variation across species’ geographical ranges: the central-marginal hypothesis and beyond. Mol. Ecol. 17, 1170–1188. doi: 10.1111/j.1365-294X.2007.03659.x
Evanno, G., Regnaut, S., and Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Frankham, R., Briscoe, D. A., and Ballou, J. D. (2002). Introduction to Conservation Genetics. Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511808999
Frei, E. R., Hahn, T., Ghazoul, J., and Pluess, A. R. (2014). Divergent selection in low and high elevation populations of a perennial herb in the Swiss Alps. Alp. Bot. 124, 131–142. doi: 10.1007/s00035-014-0131-1
Frei, E. S., Scheepens, J. F., Armbruster, G. F. J., and Stöcklin, J. (2012a). Phenotypic differentiation in a common garden reflects the phylogeography of a widespread Alpine plant. J. Ecol. 100, 297–308. doi: 10.1111/j.1365-2745.2011.01909.x
Frei, E. S., Scheepens, J. F., and Stöcklin, J. (2012b). High genetic differentiation in populations of the rare alpine plant species Campanula thyrsoides on a small mountain. Alp. Bot. 122, 23–34. doi: 10.1007/s00035-012-0103-2
García-Fernández, A., Escudero, A., Lara-Romero, C., and Iriondo, J. M. (2015). Effects of the duration of cold stratification on early life stages of the Mediterranean alpine plant Silene ciliata. Plant Biol. 17, 344–350. doi: 10.1111/plb.12226
García-Fernández, A., Iriondo, J. M., and Escudero, A. (2012a). Inbreeding at the edge: does inbreeding depression increase under more stressful conditions? Oikos 121, 1435–1445. doi: 10.1111/j.1600-0706.2011.20219.x
García-Fernández, A., Segarra-Moragues, J. G., Widmer, A., Escudero, A., and Iriondo, J. M. (2012b). Unravelling genetics at the top: mountain islands or isolated belts? Ann. Bot. 110, 1221–1232. doi: 10.1093/aob/mcs195
García-Fernández, A., Iriondo, J. M., Escudero, A., Aguilar, J. F., and Feliner, G. N. (2013). Genetic patterns of habitat fragmentation and past climate-change effects in the Mediterranean high-mountain plant Armeria caespitosa (Plumbaginaceae). Am. J. Bot. 100, 1641–1650. doi: 10.3732/ajb.1200653
Geng, Y., Tang, S., Tashi, T., Song, Z., Zhang, G., Zeng, L., et al. (2009). Fine- and landscape-scale spatial genetic structure of cushion rockjasmine, Androsace tapete (Primulaceae), across southern Qinghai-Tibetan Plateau. Genetica 135, 419–427. doi: 10.1007/s10709-008-9290-6
Giménez-Benavides, L., Albert, M. J., Iriondo, J. M., and Escudero, A. (2011a). Demographic processes of upward range contraction in a long-lived Mediterranean high mountain plant. Ecography 34, 85–93. doi: 10.1111/j.1600-0587.2010.06250.x
Giménez-Benavides, L., García-Camacho, R., Iriondo, J. M., and Escudero, A. (2011b). Selection on flowering time in Mediterranean high-mountain plants under global warming. Evol. Ecol. 25, 777–794. doi: 10.1007/s10682-010-9440-z
Giménez-Benavides, L., Escudero, A., and Iriondo, J. M. (2007a). Local adaptation enhances seedling recruitment along an altitudinal gradient in a high mountain mediterranean plant. Ann. Bot. 99, 723–734. doi: 10.1093/aob/mcm007
Giménez-Benavides, L., Escudero, A., and Iriondo, J. M. (2007b). Reproductive limits of a late-flowering high-mountain Mediterranean plant along an elevational climate gradient. New Phytol. 173, 367–382. doi: 10.1111/j.1469-8137.2006.01932.x
Giménez-Benavides, L., Escudero, A., and Pérez-García, F. (2005). Seed germination of high mountain Mediterranean species: altitudinal, interpopulation and interannual variability. Ecol. Res. 20, 433–444. doi: 10.1007/s11284-005-0059-4
Glémin, S., Ronfort, J., and Bataillon, T. (2003). Patterns of inbreeding depression and architecture of the load in subdivided populations. Genetics 165, 2193–2212.
Gonzalo-Turpin, H., and Hazard, L. (2009). Local adaptation occurs along altitudinal gradient despite the existence of gene flow in the alpine plant species Festuca eskia. J. Ecol. 97, 742–751. doi: 10.1111/j.1365-2745.2009.01509.x
Goslee, S. C., and Urban, D. L. (2007). The ecodist package for dissimilarity-based analysis of ecological data. J. Stat. Softw. 22, 1–19. doi: 10.18637/jss.v022.i07
Gottfried, M., Pauli, H., Futschik, A., Akhalkatsi, M., Baranèok, P., Benito Alonso, J. L., et al. (2012). Continent-wide response of mountain vegetation to climate change. Nat. Clim. Change 2, 111–115. doi: 10.1038/nclimate1329
Goudet, J., and Jombart, T. (2015). Hierfstat: Estimation and Tests of Hierarchical F-statistics. R Packag. Version 0.04–22. Available at: https://CRAN.R-project.org/package=hierfstat
Guillot, G., Mortier, F., and Estoup, A. (2005). GENELAND: a computer package for landscape genetics. Mol. Ecol. Notes 5, 712–715. doi: 10.1111/j.1471-8286.2005.01031.x
Haldane, J. B. S. (1954). An exact test for randomness of mating. J. Genet. 52, 631–635. doi: 10.1007/BF02985085
Hämälä, T., Mattila, T. M., and Savolainen, O. (2018). Local adaptation and ecological differentiation under selection, migration and drift in Arabidopsis lyrata. Evolution 72, 1373–1386. doi: 10.1111/evo.13502
Hamann, E., Scheepens, J. F., Kesselring, H., Armbruster, G. F. J., and Stöcklin, J. (2017). High intraspecific phenotypic variation, but little evidence for local adaptation in Geum reptans populations in the Central Swiss Alps. Alp. Bot. 127, 121–132. doi: 10.1007/s00035-017-0185-y
Hampe, A., and Petit, R. J. (2005). Conserving biodiversity under climate change: the rear edge matters. Ecol. Lett. 8, 461–467. doi: 10.1111/j.1461-0248.2005.00739.x
Hardie, D. C., and Hutchings, J. A. (2010). Evolutionary ecology at the extremes of species’ ranges. Environ. Rev. 18, 1–20. doi: 10.1139/A09-014
Hewitt, G. M. (2001). Speciation, hybrid zones and phylogeography–or seeing genes in space and time. Mol. Ecol. 10, 537–549. doi: 10.1046/j.1365-294x.2001.01202.x
Hirao, A. S., and Kudo, G. (2004). Landscape genetics of alpine-snowbed plants: comparison along geographic and snowmelt gradients. Heredity 93, 290–298. doi: 10.1038/sj.hdy.6800503
Hirst, M. J., Sexton, J. P., and Hoffmann, A. A. (2016). Extensive variation, but not local adaptation in an Australian alpine daisy. Ecol. Evol. 6, 5459–5472. doi: 10.1002/ece3.2294
Holderegger, R., and Thiel-Egenter, C. (2009). A discussion of different types of glacial refugia used in mountain biogeography and phylogeography. J. Biogeogr. 36, 476–480. doi: 10.1111/j.1365-2699.2008.02027.x
Jakobsson, M., and Rosenberg, N. A. (2007). CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 23, 1801–1806. doi: 10.1093/bioinformatics/btm233
Jenkins, D. G., Carey, M., Czerniewska, J., Fletcher, J., Hether, T., Jones, A., et al. (2010). A meta-analysis of isolation by distance: relic or reference standard for landscape genetics? Ecography 33, 315–320. doi: 10.1111/j.1600-0587.2010.06285.x
Jombart, T. (2008). adegenet: a R package for the multivariate analysis of genetic markers. Bioinformatics 24, 1403–1405. doi: 10.1093/bioinformatics/btn129
Jombart, T., and Ahmed, I. (2012). Adegenet: An R Package for the Exploratory Analysis of Genetic and Genomic Data. R Packag. ‘Adegenet’ version, 1–3. Available at: https://github.com/thibautjombart/adegenet
Jombart, T., Devillard, S., and Balloux, F. (2010). Discriminant analysis of principal components: a new method for the analysis of genetically structured populations. BMC Genet. 11:94. doi: 10.1186/1471-2156-11-94
Kawecki, T. J. (2008). Adaptation to Marginal Habitats. Annu. Rev. Ecol. Evol. Syst. 39, 321–342. doi: 10.1146/annurev.ecolsys.38.091206.095622
Keenan, K., McGinnity, P., Cross, T. F., Crozier, W. W., and Prodöhl, P. A. (2013). diveRsity: an R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 4, 782–788. doi: 10.1111/2041-210X.12067
Kern, A. D., and Hahn, M. W. (2018). The neutral theory in light of natural selection. Mol. Biol. Evol. 65:054304. doi: 10.1093/molbev/msy092
Kim, E., and Donohue, K. (2013). Local adaptation and plasticity of Erysimum capitatum to altitude: its implications for responses to climate change. J. Ecol. 101, 796–805. doi: 10.1111/1365-2745.12077
Knowles, L. L. (2001). Did the pleistocene glaciations promote divergence? Tests of explicit refugial models in montane grasshopprers. Mol. Ecol. 10, 691–701. doi: 10.1046/j.1365-294X.2001.01206.x
Kohler, T., Giger, M., Hurni, H., Ott, C., Wiesmann, U., Wymann, et al. (2010). Mountains and climate change: a global concern. Mt. Res. Dev. 30, 53–55. doi: 10.1659/MRD-JOURNAL-D-09-00086.1
Körner, C. (2003). Alpine Plant Life. Functional Plant Ecology of High Mountain Ecosystems. Berlin: Springer-Verlag. doi: 10.1007/978-3-642-18970-8
Körner, C. (2007). The use of “altitude” in ecological research. Trends Ecol. Evol. 22, 569–574. doi: 10.1016/j.tree.2007.09.006
Kuss, P., Pluess, A. R., Aegisdottir, H. H., and Stocklin, J. (2008). Spatial isolation and genetic differentiation in naturally fragmented plant populations of the Swiss Alps. J. Plant Ecol. 1, 149–159. doi: 10.1093/jpe/rtn009
Kyrkou, I., Iriondo, J. M., and García-Fernández, A. (2015). A glacial survivor of the alpine Mediterranean region: phylogenetic and phylogeographic insights into Silene ciliata Pourr. (Caryophyllaceae). PeerJ 3:e1193. doi: 10.7717/peerj.1193
Lande, R., and Shannon, S. (1996). The role of genetic variation in adaptation and population persistence in a changing environment. Evolution 216, 434–437. doi: 10.1111/j.1558-5646.1996.tb04504.x
Lara-Romero, C., de la Cruz, M., Escribano-Ávila, G., García-Fernández, A., and Iriondo, J. M. (2016a). What causes conspecific plant aggregation? Disentangling the role of dispersal, habitat heterogeneity and plant–plant interactions. Oikos 125, 1304–1313. doi: 10.1111/oik.03099
Lara-Romero, C., García-Fernández, A., Robledo-Arnuncio, J. J., Roumet, M., Morente-López, J., López-Gil, A., et al. (2016b). Individual spatial aggregation correlates with between-population variation in fine-scale genetic structure of Silene ciliata (Caryophyllaceae). Heredity 116, 417–423. doi: 10.1038/hdy.2015.102
Lara-Romero, C., Robledo-Arnuncio, J. J., García-Fernández, A., and Iriondo, J. M. (2014). Assessing intraspecific variation in effective dispersal along an altitudinal gradient: a test in two Mediterranean high-mountain plants. PLoS One 9:e087189. doi: 10.1371/journal.pone.0087189
Lega, M., Fior, S., Li, M., Leonardi, S., and Varotto, C. (2014). Genetic drift linked to heterogeneous landscape and ecological specialization drives diversification in the alpine endemic columbine aquilegia thalictrifolia. J. Hered. 105, 542–554. doi: 10.1093/jhered/esu028
López-Moreno, J. I., Vicente-Serrano, S. M., and Lanjeri, S. (2007). Mapping snowpack distribution over large areas using GIS and interpolation techniques. Clim. Res. 33:257. doi: 10.3354/cr033257
Manel, S., Gugerli, F., Thuiller, W., Alvarez, N., Legendre, P., Holderegger, R., et al. (2012). Broad-scale adaptive genetic variation in alpine plants is driven by temperature and precipitation. Mol. Ecol. 21, 3729–3738. doi: 10.1111/j.1365-294X.2012.05656.x
Manel, S., and Holderegger, R. (2013). Ten years of landscape genetics. Trends Ecol. Evol. 28, 614–621. doi: 10.1016/j.tree.2013.05.012
Manel, S., Schwartz, M. K., Luikart, G., and Taberlet, P. (2003). Landscape genetics: combining landscape ecology and population genetics. Trends Ecol. Evol. 18, 189–197. doi: 10.1016/S0169-5347(03)00008-9
Marques, I., Draper, D., López-Herranz, M. L., Garnatje, T., Segarra-Moragues, J. G., and Catalán, P. (2016). Past climate changes facilitated homoploid speciation in three mountain spiny fescues (Festuca. Poaceae). Sci. Rep. 6, 1–11. doi: 10.1038/srep36283
Moran, P. A. P. (1950). Notes on continuous stochastic phenomena. Biometrika 37, 17–23. doi: 10.1093/biomet/37.1-2.17
Mosca, E., Eckert, A. J., Di Pierro, E. A., Rocchini, D., La Porta, N., Belletti, P., et al. (2012). The geographical and environmental determinants of genetic diversity for four alpine conifers of the European Alps. Mol. Ecol. 21, 5530–5545. doi: 10.1111/mec.12043
Mosca, E., González-Martínez, S. C., and Neale, D. B. (2014). Environmental versus geographical determinants of genetic structure in two subalpine conifers. New Phytol. 201, 180–192. doi: 10.1111/nph.12476
Nieto Feliner, G. (2014). Patterns and processes in plant phylogeography in the Mediterranean Basin: a review. Perspect. Plant Ecol. Evol. Syst. 16, 265–278. doi: 10.1016/j.ppees.2014.07.002
Ninyerola, M., Pons, X., and Roure, J. M. (2005). Atlas Climático Digital de la Península Ibérica: Metodología y Aplicaciones en Bioclimatología y Geobotánica. Bellaterra: Universitat Autònoma de Barcelona, Departament de Biologia Animal, Biologia Vegetal i Ecologia (Unitat de Botánica).
Noguerales, V., Cordero, P. J., and Ortego, J. (2016). Hierarchical genetic structure shaped by topography in a narrow-endemic montane grasshopper. BMC Evol. Biol. 16:96. doi: 10.1186/s12862-016-0663-7
Nogués-Bravo, D., Araújo, M. B., Errea, M. P., and Martínez-Rica, J. P. (2007). Exposure of global mountain systems to climate warming during the 21st Century. Glob. Environ. Change 17, 420–428. doi: 10.1016/j.gloenvcha.2006.11.007
Ometto, L., Li, M., Bresadola, L., Barbaro, E., Neteler, M., and Varotto, C. (2015). Demographic history, population structure, and local adaptation in alpine populations of Cardamine impatiens and Cardamine resedifolia. PLoS One 10:e0125199. doi: 10.1371/journal.pone.0125199
Orsini, L., Vanoverbeke, J., Swillen, I., Mergeay, J., and De Meester, L. (2013). Drivers of population genetic differentiation in the wild: isolation by dispersal limitation, isolation by adaptation and isolation by colonization. Mol. Ecol. 22, 5983–5999. doi: 10.1111/mec.12561
Papuga, G., Gauthier, P., Pons, V., Farris, E., and Thompson, J. D. (2018). Ecological niche differentiation in peripheral populations: a comparative analysis of eleven Mediterranean plant species. Ecography 41, 1650–1664. doi: 10.1111/ecog.03331
Peakall, P., and Smouse, R. (2012). GenAlEx 6.5: genetic analysis in excel. Population genetic software for teaching and research—an update. Bioinformatics 28, 2537–2539. doi: 10.1093/bioinformatics/bts460
Phillips, S. J., Anderson, R. P., and Schapire, R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Modell. 190, 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
Pironon, S., Papuga, G., Villellas, J., Angert, A. L., García, M. B., and Thompson, J. D. (2017). Geographic variation in genetic and demographic performance: new insights from an old biogeographical paradigm. Biol. Rev. 92, 1877–1909. doi: 10.1111/brv.12313
Pironon, S., Villellas, J., Morris, W. F., Doak, D. F., and García, M. B. (2015). Do geographic, climatic or historical ranges differentiate the performance of central versus peripheral populations? Glob. Ecol. Biogeogr. 24, 611–620. doi: 10.1111/geb.12263
Poncet, B. N., Herrmann, D., Gugerli, F., Taberlet, P., Holderegger, R., Gielly, L., et al. (2010). Tracking genes of ecological relevance using a genome scan in two independent regional population samples of Arabis alpina. Mol. Ecol. 19, 2896–2907. doi: 10.1111/j.1365-294X.2010.04696.x
Pritchard, J. K., Stephens, M., and Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959. doi: 10.1111/j.1471-8286.2007.01758.x
R Core Team (2009). A Language and Environment for Statistical Computing. Viena: R Core Team. Available at: https://www.R-project.org/
Rangwala, I., and Miller, J. R. (2012). Climate change in mountains: a review of elevation-dependent warming and its possible causes. Clim. Change 114, 527–547. doi: 10.1007/s10584-012-0419-3
Rivas-Martínez, S., Fernández-González, F., Sánchez-Mata, D., and Pizarro, J. M. (1990). Vegetación de la sierra de guadarrama. Itinera Geobot. 4, 3–132.
Rosenberg, N. A. (2004). DISTRUCT: a program for the graphical display of population structure. Mol. Ecol. Resour. 4, 137–138. doi: 10.1046/j.1471-8286.2003.00566.x
Rousset, F. (1997). Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance. Genet. Soc. Am. 145, 1219–1228.
Rousset, F. (2008). Genepop’007: a complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 8, 103–106. doi: 10.1111/j.1471-8286.2007.01931.x
Rousset, F., and Ferdy, J.-B. (2014). Testing environmental and genetic effects in the presence of spatial autocorrelation. Ecography 37, 781–790. doi: 10.1111/ecog.00566
Ruiz-Gonzalez, A., Cushman, S. A., Madeira, M. J., Randi, E., and Gómez-Moliner, B. J. (2015). Isolation by distance, resistance and/or clusters? Lessons learned from a forest-dwelling carnivore inhabiting a heterogeneous landscape. Mol. Ecol. 24, 5110–5129. doi: 10.1111/mec.13392
Sagarin, R., and Gaines, S. (2002). The ‘abundant centre’distribution: to what extent is it a biogeographical rule? Ecol. Lett. 5, 137–147. doi: 10.1046/j.1461-0248.2002.00297.x
Sagarin, R. D., Gaines, S. D., and Gaylord, B. (2006). Moving beyond assumptions to understand abundance distributions across the ranges of species. Trends Ecol. Evol. 21, 524–530. doi: 10.1016/j.tree.2006.06.008
Sexton, J. P., Hangartner, S. B., and Hoffmann, A. A. (2014). Genetic isolation by environment or distance: which pattern of gene flow is most common? Evolution 68, 1–15. doi: 10.1111/evo.12258
Sexton, J. P., McIntyre, P. J., Angert, A. L., and Rice, K. J. (2009). Evolution and ecology of species range limits. Annu. Rev. Ecol. Evol. Syst. 40, 415–436. doi: 10.1146/annurev.ecolsys.110308.120317
Shafer, A. B. A., and Wolf, J. B. W. (2013). Widespread evidence for incipient ecological speciation: a meta-analysis of isolation-by-ecology. Ecol. Lett. 16, 940–950. doi: 10.1111/ele.12120
Soule, M. (1973). The epistasis cycle: a theory of marginal populations. Annu. Rev. Ecol. Syst. 4, 165–187. doi: 10.1146/annurev.es.04.110173.001121
Spear, S. F., Balkenhol, N., Fortin, M. J., McRae, B. H., and Scribner, K. (2010). Use of resistance surfaces for landscape genetic studies: considerations for parameterization and analysis. Mol. Ecol. 19, 3576–3591. doi: 10.1111/j.1365-294X.2010.04657.x
Steinbauer, M. J., Grytnes, J. A., Jurasinski, G., Kulonen, A., Lenoir, J., Pauli, H., et al. (2018). Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 556, 231–234. doi: 10.1038/s41586-018-0005-6
Stöcklin, J., Kuss, P., and Pluess, A. R. (2009). Genetic diversity, phenotypic variation and local adaptation in the alpine landscape: case studies with alpine plant species. Bot. Helv. 119, 125–133. doi: 10.1007/s00035-009-0065-1
Storfer, A., Murphy, M. A., Evans, J. S., Goldberg, C. S., Robinson, S., Spear, S. F., et al. (2007). Putting the “landscape” in landscape genetics. Heredity 98, 128–142. doi: 10.1038/sj.hdy.6800917
Storfer, A., Murphy, M. A., Spear, S. F., Holderegger, R., and Waits, L. P. (2010). Landscape genetics: where are we now? Mol. Ecol. 19, 3496–3514. doi: 10.1111/j.1365-294X.2010.04691.x
Thuiller, W., Lavorel, S., Araújo, M. B., Sykes, M. T., and Prentice, I. C. (2005). Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. U.S.A. 102, 8245–8250. doi: 10.1073/pnas.0409902102
Title, P. O., and Bemmels, J. B. (2018). ENVIREM: an expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 41, 291–307. doi: 10.1111/ecog.02880
Totland, Ø. (1999). Effects of temperature on performance and phenotypic selection on plant traits in alpine Ranunculus acris. Oecologia 120, 242–251. doi: 10.1007/s004420050854
Totland,Ø., and Alatalo, J. M. (2002). Effects of temperature and date of snowmelt on growth, reproduction, and flowering phenology in the arctic/alpine herb, Ranunculus glacialis. Oecologia 133, 168–175. doi: 10.1007/s00442-002-1028-z
Tutin, T. G., Heywood, V. H., Burges, N. A., Valentine, D. H., Walters, S. M., and Webb, D. A. (1964). Flora Europaea. Cambridge: Cambridge University Press.
Tzedakis, P. C., Lawson, I. T., Frogley, M. R., Hewitt, G. M., and Preece, R. C. (2002). Buffered tree population changes in a quaternary refugium: evolutionary implications. Science 297, 2044–2047. doi: 10.1126/science.1073083
Van Oosterhout, C., Hutchinson, W. F., Wills, D. P. M., and Shipley, P. (2004). MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Resour. 4, 535–538. doi: 10.1111/j.1471-8286.2004.00684.x
Wang, I. J., and Bradburd, G. S. (2014). Isolation by environment. Mol. Ecol. 23, 5649–5662. doi: 10.1111/mec.12938
Wang, Y., Jiang, H., Peng, S., and Korpelainen, H. (2011). Genetic structure in fragmented populations of Hippophae rhamnoides ssp. sinensis in China investigated by ISSR and cpSSR markers. Plant Syst. Evol. 295, 97–107. doi: 10.1007/s00606-011-0466-7
Whitlock, M. C. (2004). “Selection and drift in metapopulations,” in Ecology, Genetics, and Evolution of Metapopulations, eds I. Hanski and O. E. Gaggiotti (Burlington, MA: Elsevier), 153–173. doi: 10.1016/B978-012323448-3/50009-X
Keywords: landscape genetics, isolation by distance, isolation by resistance, isolation by environment, genetic diversity, marginal populations, environmental gradient
Citation: Morente-López J, García C, Lara-Romero C, García-Fernández A, Draper D and Iriondo JM (2018) Geography and Environment Shape Landscape Genetics of Mediterranean Alpine Species Silene ciliata Poiret. (Caryophyllaceae). Front. Plant Sci. 9:1698. doi: 10.3389/fpls.2018.01698
Received: 25 June 2018; Accepted: 31 October 2018;
Published: 27 November 2018.
Edited by:
Jordi López-Pujol, Consejo Superior de Investigaciones Científicas (CSIC), SpainReviewed by:
Juli Caujapé Castells, Jardín Botánico Canario “Viera y Clavijo”, SpainCopyright © 2018 Morente-López, García, Lara-Romero, García-Fernández, Draper and Iriondo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javier Morente-López, amF2aW1vcmVudGVAZ21haWwuY29t José María Iriondo, am9zZS5pcmlvbmRvQHVyamMuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.