 Minghui Lv
Minghui Lv Kai He
Kai He Jia Li
Jia Li- Ministry of Education Key Laboratory of Cell Activities and Stress Adaptation, School of Life Sciences, Lanzhou University, Lanzhou, China
Brassinosteroids (BRs) are essential phytohormones mainly perceived by a single-pass transmembrane receptor-like protein kinase (RLK), BRASSINOSTEROID INSENSITIVE 1 (BRI1). bri1-5 and bri1-9, two distinct mutants with point mutations in the extracellular domain of BRI1, show weak defective phenotypes. Previous studies indicated that bri1-5 and bri1-9 mutated proteins can be recognized and eliminated via an endoplasmic reticulum quality control (ERQC) mechanism. Most of these two proteins, therefore, cannot reach their destination, plasma membrane. Here, we report our functional characterization of bri1-301, another BRI1 mutant protein with an amino acid substitution in the cytoplasmic kinase domain. bri1-301 is a partially functional BR receptor with significantly decreased protein abundance. Interestingly, protein stability and subcellular localization of bri1-301 are temperature-sensitive. At 22∘C, an optimal temperature for indoor Arabidopsis growth, bri1-301 shows a weak defective phenotype. At a lower temperature condition such as 18∘C, bri1-301 exhibits subtle morphological defects. At a higher temperature condition such as 28∘C, on the other hand, bri1-301 displays an extremely severe phenotype reminiscent to that of a null bri1 mutant due to greatly increased bri1-301 internalization and degradation. Our detailed analyses suggest that bri1-301 stability is controlled by ERQC and plasma membrane quality control (PMQC) systems. Since PMQC has not been well studied in plants, bri1-301 can be used as a model mutant for future genetic dissection of this critical process.
Introduction
Brassinosteroids (BRs) are a group of polyhydroxylated phytohormones widely identified in plant kingdom. BRs regulate various physiological processes during growth and development (Grove et al., 1979; Clouse and Sasse, 1998; Zhang et al., 2009; Vilarrasa-Blasi et al., 2014; Lee et al., 2015). Arabidopsis mutants with defects in BR biosynthesis, perception, or signal transduction usually display similar phenotypic abnormalities including dwarfism, dark green and compact rosette leaves, delayed senescence, reduced male fertility, and de-etiolation in the dark (Clouse et al., 1996; Szekeres et al., 1996; Li et al., 2001).
Brassinosteroids are perceived by a protein complex consisting of BRASSINOSTEROID INSENSITIVE1 (BRI1) and BRI1-ASSOCIATED RECEPTOR KINASE1 (BAK1), both of which are single-pass transmembrane leucine-rich repeat receptor-like protein kinases (LRR-RLKs) (Li and Chory, 1997; Li et al., 2002; Nam and Li, 2002). Binding of BRs to the extracellular domain of BRI1 relieves its kinase domain from an inhibitory state caused by both the interference of its C-terminal tail as well as an inhibitory binding protein, BRI1 KINASE INHIBITOR 1 (BKI1) (Wang et al., 2005b; Wang and Chory, 2006; Jaillais et al., 2011). Physical interaction between BRs and the extracellular domain of BRI1 generates a docking platform for the recruitment of the co-receptor, BAK1 (Hothorn et al., 2011; She et al., 2011; He et al., 2013; Santiago et al., 2013). Only when BRI1-BR-BAK1 is formed, both BRI1 and BAK1 are activated through a reciprocal transphosphorylation mechanism (Wang et al., 2008). Fully activated BRI1 then triggers downstream signaling cascade predominantly via protein phosphorylation and dephosphorylation (Li and Nam, 2002; Mora-Garcia et al., 2004; Tang et al., 2008; Kim et al., 2009, 2011; Tang et al., 2011). Transcription factors, including well-characterized BZR1 and BES1, are subsequently activated and thousands of their target genes are transcriptionally regulated (Wang et al., 2002; Yin et al., 2002; He et al., 2005; Sun et al., 2010; Yu et al., 2011).
As the major BR receptor, BRI1 has been extensively studied and over 30 unique bri1 mutant alleles have been identified through a number of independent genetic screens (Clouse, 1996; Li and Chory, 1997; Noguchi et al., 1999; Friedrichsen et al., 2000; Xu et al., 2008; Belkhadir et al., 2010; Shang et al., 2011; Gou et al., 2012; Sun et al., 2017). Severe phenotypes of bri1 null alleles revealed the crucial roles of BRI1 on growth and development (Clouse, 1996; Li and Chory, 1997; Friedrichsen et al., 2000; Gou et al., 2012). Meanwhile, studies on bri1 weak alleles contributed to our better understanding of several BRI1-associated cellular processes. For example, bri1-9 is a semi-dwarfed bri1 mutant with an S662F substitution within the BR binding domain of BRI1. It was therefore predicted that this mutation might have disrupted bri1-9 to bind its ligand (Friedrichsen et al., 2000). A genetic suppressor screen discovered that bri1-9 can be suppressed by a loss-of-function mutant of EBS1, which encodes a UDP-glucose:glycoprotein glucosyltransferase essential to endoplasmic reticulum-mediated protein quality-control (ERQC) (Jin et al., 2007). ERQC is a highly conserved mechanism that monitors the protein folding process, allowing export of only correctly folded proteins, but retaining misfolded proteins (e.g., bri1-9) (Smith et al., 2011). Loss-of-function mutant of EBS1 in bri1-9 reduces ERQC stringency and allows misfolded bri1-9 protein to export to the cell surface, resulting in a phenotypic suppression of the mutant (Jin et al., 2007). The wild-type-like morphology of ebs1-1 bri1-9 suggests that the defective phenotype of bri1-9 is likely caused by lack of plasma-membrane-localized BR receptor due to ER retention rather than disrupted BR binding of bri1-9 (Jin et al., 2007). Similar to bri1-9, another extracellular-domain mutant of BRI1, bri1-5, is also retained in the ER by the ERQC mechanism and is degraded by the ERAD process (Noguchi et al., 1999; Hong et al., 2008).
bri1-301 is a weak bri1 allele that contains two tandem nucleotide mutations but only one single amino acid substitution of G989I within the VIa kinase subdomain (Vert et al., 2005; Xu et al., 2008). In vitro autophosphorylation analysis indicated that bri1-301 does not show any kinase activity. In addition, bri1-301 is unable to phosphorylate BAK1, the co-receptor and also one of the known substrates of BRI1 (Xu et al., 2008). Nevertheless, molecular mechanisms explaining the weak phenotype of bri1-301 have never been elucidated. Our initial interest is to investigate why such a kinase inactive bri1 mutant shows a weak instead of a strong phenotype. Is the kinase activity truly non-essential for the biological functions of BRI1? If it is true, how can we explain the severe phenotypes of all other kinase-dead bri1 mutants identified so far?
Here, we show that bri1-301 possesses kinase-activity in vivo, although greatly reduced. More interestingly, we report that the morphological severity of bri1-301 and the protein accumulation of bri1-301 are temperature-dependent. In addition, the PM-localization of bri1-301 is normal at 22°C but is greatly disrupted at 28°C. Our results suggest a possible PMQC mechanism which is involved in recognition and removal of non-native membrane proteins such as bri1-301.
Materials and Methods
Plant Materials, Growth Conditions, and Phenotypic Analysis
Arabidopsis thaliana mutants used in this study include bri1-301, det2-1, cpd91 (Du et al., 2016), bri1-705 (Sun et al., 2017), bin2-1, cpd (Du et al., 2016) and bri1-701, all of which are Col-0 background. Plants were grown under long-day light condition (16 h light and 8 h dark) or darkness at 18°C, 22°C, and 28°C, respectively.
For seedlings, 7-day-old light-grown and 4-day-old dark-grown seedlings on 1/2 MS plates were photographed. For adults, the rosettes and leaves were photographed at indicated time points. All measurements were carried out using ImageJ1. Three independent biological replicates were carried out and at least 20 seedlings were used for each measurement. The statistical significance was evaluated by Student’s t-test.
BL Treatment
After surface sterilization with 30% (v/v) bleach for 10 min and washing several times with sterile deionized water, seeds were sown on 1/2 MS medium plates containing 0.8% (w/v) agar and 1% (w/v) sucrose supplemented with or without 1 μM 24-epibrassinolide (Sigma). The plates were vernalized for 3 days at 4°C and then were transferred to growth chamber. Photos were taken at the indicated time points and the root length were measured as described above. Three independent biological replicates were carried out and the statistical significance was evaluated by Student’s t-test.
RNA Extraction and Quantitative Real-Time PCR
11/9/7-day-old seedlings of Col-0 and bri1-301 grown on 1/2 MS plates at 18°C/22°C/28°C were treated with or without 1 μM 24-epibrassinolide in deionized water for 1.5 h at same temperature condition. RNA extraction, reverse transcription and quantitative real-time PCR were performed as previously described (Zhao et al., 2016). Primers used in this study are listed in Supplementary Table S1. Three independent biological replicates were carried out for all the quantitative analyses.
Vector Construction and Transgenic Plant Generation
The full-length BAK1, BRI1-301 and other mutated BRI1 coding sequence were cloned into destination vector pBIB-35S-GWR-FLAG (Gou et al., 2010) through two-step Gateway technology (Invitrogen, Life Technologies). The constructs were then transformed into Col-0, bri1-301, and bri1-701 background through floral dip method (Clough and Bent, 1998). The 35S::BRI1-FLAG and 35S::BAK1-GFP transgenic plants were described in a previous report (Zhao et al., 2016).
Protein Extraction, Immunoprecipitation, and Western Blot Analysis
For BES1 and BRI1/bri1 detection, the materials were prepared as described in RNA analysis. Total proteins were isolated with extraction buffer as described previously (Zhou et al., 2017). The lysates were centrifuged at 16,000 ×g for 10 min at 4°C after vortexing and the supernatant was then separated on a 12% (for BES1) and 7% (for BRI1) Bis-Tris SDS-PAGE gel and analyzed by immunoblotting with anti-BES1 serum, anti-BRI1 antibody (Agrisera) or anti-FLAG (Sigma) and anti-Tubulin (Sigma). The gray values of signal bands were measured by ImageJ 1.4.3 software (see footnote 1).
For the in vivo phosphorylation assay, transgenic plants were grown on 1/2 MS plates for 11/9/7 days at 18°C/22°C/28°C and then were treated with or without 1 μM BL for 1.5 h at the same temperature. After grinding in liquid nitrogen, the powder was lysed with extraction buffer (10 mM HEPES (pH 7.5), 100 mM NaCl, 1 mM EDTA, 10% glycerol, 0.5% Triton X-100 and 1:100 protease inhibitor cocktail from Roche). After vortexing vigorously for 30 s, the samples were centrifuged at 16,000 ×g for 10 min at 4°C, and the supernatant was then incubated with anti-FLAG (Sigma) agarose beads for 2 h at 4°C with gentle shaking. The immunoprecipitated proteins were separated on a 7% Bis-Tris SDS-PAGE gel and were analyzed by immunoblotting with anti-FLAG (Abmart), or anti-Pi-Thr antibody (Cell Signaling Technology).
Confocal Microscopy
Five-day-old pBRI1::BRI1-GFP and pBRI1::bri1-301-GFP transgenic seedlings grown at 22°C and 28°C were used for imaging. For protein abundance observation, seedlings were directly mounted and scanned. For subcellular localization observation, seedlings were treated with propidium iodide (aqueous solution, 10 μg/mL) for 10 min, then were mounted in water and immediately observed under a Zeiss LSM 510 confocal microscope. The BRI1-GFP, bri1-301-GFP and PI were excited with 488-nm wavelength. The fluorescence emissions were detected with spectral detector set BP 505-560. Laser intensity and detection settings were kept constant except a higher detector gain value for bri1-301-GFP signal during its subcellular localization study.
Endo H Treatment
11/9/7-day-old Col-0, bri1-301 and bri1-5 seedlings grown on 1/2 MS agar plates at 18°C/22°C/28°C were harvested and the total protein extracts were prepared as described previously (Hong et al., 2012). Briefly, seedlings were ground in liquid N2, dissolved in 2× SDS buffer containing 125 mM Tris (pH 6.8), 4% (w/v) SDS, 20% (v/v) glycerol, 200 mM DTT, 0.02% (w/v) bromophenol blue. After boiling and centrifugation, supernatants were incubated with or without 1,000 U of Endo H in 1× G5 buffer (New England Biolabs) for 1 h at 37°C. The Western blot analyses were performed as above.
Results
Phenotypic Severity of bri1-301 Is Temperature-Dependent
bri1-301 was originally characterized as a weak bri1 allele carrying two tandem nucleotide mutations, resulting in one amino-acid substitution, G989I (Xu et al., 2008). A previous report indicated that bri1-301 does not show any detectable activity either for autophosphorylating itself or phosphorylating its known substrate, BAK1, in vitro (Xu et al., 2008). The appeared weak phenotype of bri1-301 is contradictory with other known BRI1 kinase-dead mutants, which always show extreme morphological defects. Therefore, we are interested in studying whether the kinase activity of BRI1 is essential to the BR signaling pathway using bri1-301 as a studying material. During our investigation, however, we unintentionally observed that the phenotypic defects of soil-grown bri1-301 mutants largely rely on their growing temperatures. In a relatively low temperature growth chamber (18°C), bri1-301 only shows a subtle defective phenotype compared to wild type (Figure 1). In a growth chamber normally used for Arabidopsis growth (22°C), bri1-301 displays a weak bri1 defective phenotype, similar to several other well-characterized bri1 mutants such as bri1-5, bri1-120, and bri1-702 (Noguchi et al., 1999; Shang et al., 2011; Sun et al., 2017) (Figure 1). Surprisingly, in a warmer growth chamber (28°C), bri1-301 exhibits a severe defective phenotype, similar to null bri1 mutants, such as bri1-701, bri1-4, and bri1-709 (Noguchi et al., 1999; Gou et al., 2012; Sun et al., 2017) (Figure 1).
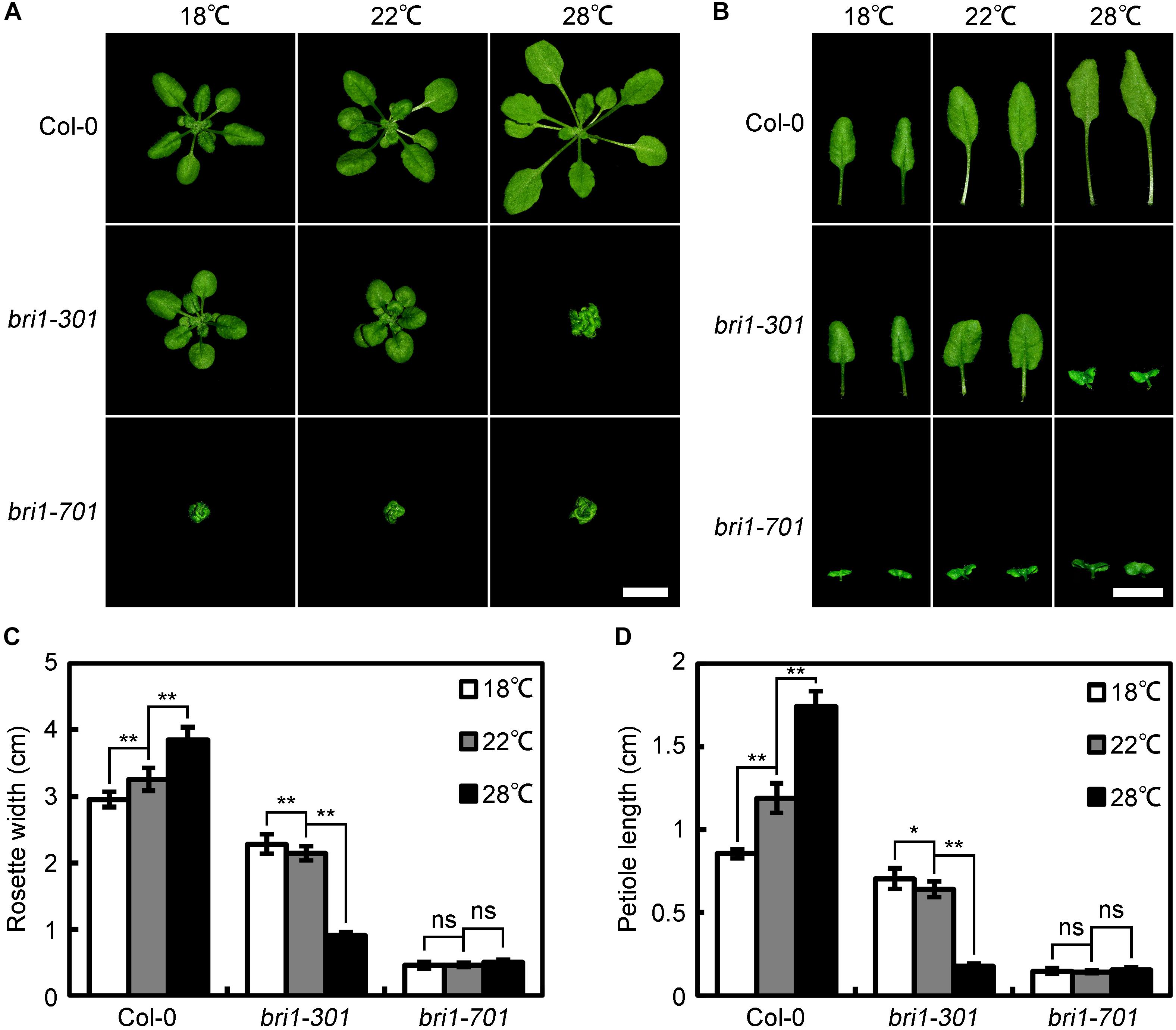
FIGURE 1. bri1-301 growth is significantly retarded by elevated ambient temperature. (A) Rosette phenotypes of wild type and different bri1 alleles grown at 18°C, 22°C, and 28°C. (B) Leaf phenotypes of wild type and different bri1 alleles grown at 18°C, 22°C, and 28°C. Photographs of the rosettes and the third and fourth true leaves were taken 4 weeks after germination. Scale bars represent 1 cm. (C) Measurements of rosette width of plants as shown in (A). (D) Measurements of petiole length of the third and fourth true leaves as present in (B). In (C,D), the data shown are means and standard deviations. The asterisks indicate statistical significance evaluated by Student’s t-test (∗P < 0.05, ∗∗P < 0.01), ns represents not significant. Three independent biological replicates were carried out. Similar results were obtained. One of the representative results is shown.
We also examined the phenotypes of seedlings grown on half strength of Murashige and Skoog (1/2 MS) media at three different temperatures, 18°C, 22°C, and 28°C. Under a long-day photoperiod condition (16 h light/8 h dark), warmer temperature inhibits the elongation of cotyledon petioles of bri1-301, resulting in compact and curled cotyledons (Supplementary Figure S1A). In darkness, hypocotyl elongation of Col-0 is stimulated by warmer temperature (Supplementary Figures S1B,C). Although bri1-301 shows more elongated hypocotyls at 22°C, further raising temperature to 28°C can significantly inhibit the hypocotyl elongation (Supplementary Figures S1B,C).
Warmer Temperature-Inhibited Growth of bri1-301 Is Caused by G989I Substitution
Previous studies indicated that some Arabidopsis mutants, including zed1-D (hopz-eti-deficient 1-dominant) (Wang et al., 2017), eta1 (enhancer of tir1-1 auxin resistance 1) (Quint et al., 2005), scd1-1 (stomatal cytokinesis-defective 1-1) (Falbel et al., 2003), exhibit growth retardation under a comparatively warmer condition. Hence, it is possible that another point mutation other than G989I in bri1-301 is responsible for the warm-enhanced phenotype. To examine this possibility, we crossed bri1-301 with Col-0 and a newly identified subtle bri1 mutant, bri1-705, and analyzed the phenotypes of F1 individuals at 18°C, 22°C, and 28°C, respectively (Sun et al., 2017). We found that bri1-301 × Col-0 F1 heterozygous individuals show fully complemented phenotype similar to wild type, Col-0, whereas bri1-301 ×bri1-705 F1 individuals show a slightly retarded growing phenotype at 28°C (Figures 1A,C, 2A,B). We also generated transgenic plants harboring bri1-301 coding sequence driven by a BRI1 native promoter in a bri1-701 background. As expected, the transgenic lines recapitulated a temperature-dependent phenotype as bri1-301 (Figures 2C,D). These observations suggest that the warmer temperature-inhibited growth of bri1-301 is indeed caused by the G989I substitution within BRI1 but unlikely by other unknown mutations within the bri1-301 genome.
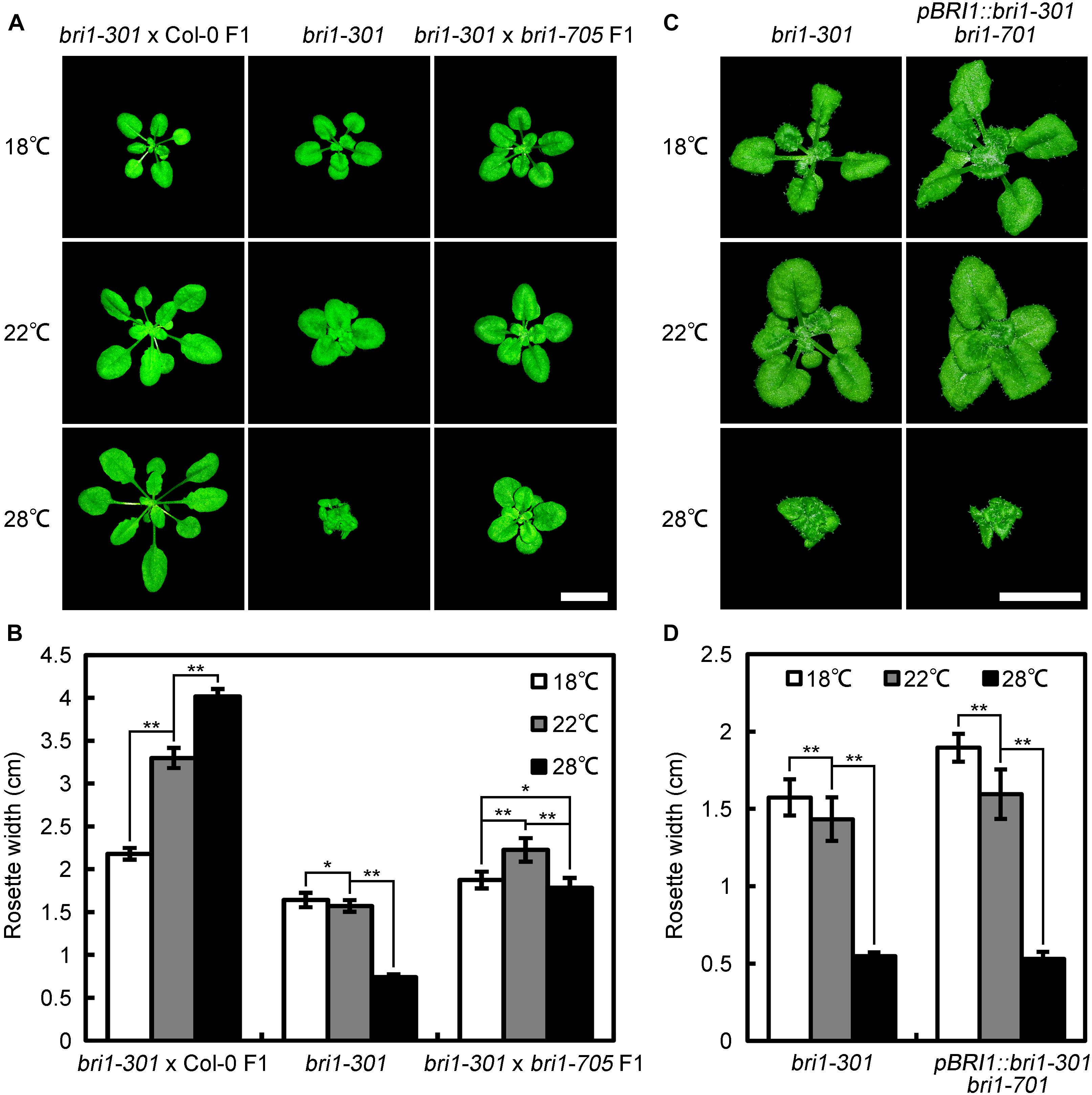
FIGURE 2. Genetic analyses demonstrated that the point mutation shown in bri1-301 is responsible for retarded growth phenotype at elevated temperature. (A) Phenotypes of 4-week-old bri1-301 and F1 hybrids grown at 18°C, 22°C, and 28°C. Scale bar represents 1 cm. (B) Measurements of rosette width of the plants as shown in (A). (C) Phenotypes of 5-week-old bri1-301 and pBRI1:bri1-301 bri1-701 grown at 18°C, 22°C, and 28°C. Scale bar represents 1 cm. (D) Measurements of rosette width of the plants as shown in (A). In (B,D), the data shown are means and standard deviations. The asterisks indicate statistical significance evaluated by Student’s t-test (∗P < 0.05, ∗∗P < 0.01). Three biological replicates were carried out. Similar results were obtained. One of the representative results is shown.
Temperature-Dependent bri1-301 Severity Is Allele Specific
To test whether the temperature-dependent bri1-301 severity is allele specific, we examined the phenotypes of a number of BR deficient and signaling mutants at 18°C, 22°C, and 28°C. We found that warmer temperature generally promotes rosette growth of all other weak BR mutants tested at varying degrees, but it appears no obvious effect on the growth of null bri1 alleles (Figure 3). We also mutated the G989 residue of BRI1 to other amino acids through site-directed mutagenesis and introduced the mutated bri1 coding sequences driven by the BRI1 native promoter into bri1-701. We successfully generated 11 G989-C, D, E, F, H, I, L, M, Q, S, or Y transgenic plants. To our surprise, all of the G989- C, D, E, F, H, L, M, Q, S, or Y transgenic plants failed to mimic the warm-inhibited growth phenotype of bri1-301 except for the G989I transgenic plants (Supplementary Figure S2). These results suggest that warm-enhanced growth defects of bri1-301 is both allele and amino acid specific.
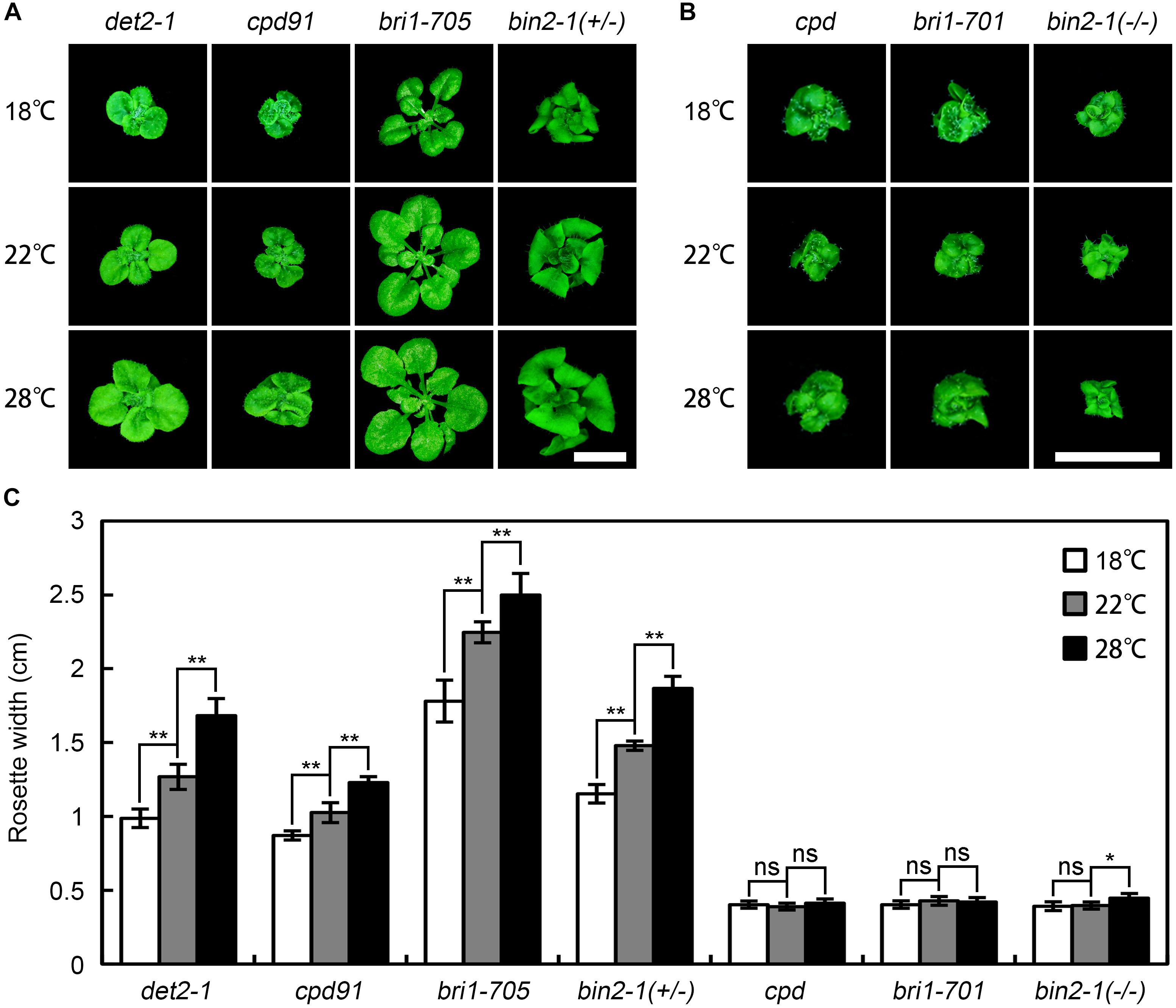
FIGURE 3. Effects of elevated ambient temperature on growth of other BR mutants. (A,B) Phenotypes of 4-week-old BR mutants grown at 18°C, 22°C, and 28°C. Scale bars represent 1 cm. (C) Measurements of rosette width of the plants as shown in (A,B). The data shown are means and standard deviations. The asterisks indicate statistical significance evaluated by Student’s t-test (∗P < 0.05, ∗∗P < 0.01), ns represents not significant. Three independent biological replicates were carried out. Similar results were obtained. Only one of the representative results is shown.
bri1-301 Is Sensitive to the BL Treatment at Lower Temperature but Not at Warmer Temperature
The phenotypic resemblance of 28°C-grown bri1-301 to bri1-701 suggests that BR signal transduction pathway in bri1-301 is disrupted at 28°C. To test this hypothesis, we compared the BR responses of Col-0 and bri1-301 under various temperature conditions. Root growth inhibition assay indicated that when grown at 18°C (Clouse et al., 1996), the primary root growth of bri1-301 is inhibited by exogenously applied BL, the most active form of BRs (Figures 4A,B). However, such inhibitory effect was not observed at 22°C and 28°C (Figures 4A,B). Consistently, quantitative RT-PCR assay showed that the expression levels of CPD and SAUR-AC1 are significantly increased and decreased, respectively, in bri1-301 compared to those in wild type at 28°C (Supplementary Figure S3). Meanwhile, when raising temperature, the feedback regulations of CPD and SAUR-AC1 by exogenously applied BL were gradually attenuated (Goda et al., 2002) (Figures 4C,D). Western blot analyses revealed that the BL induced accumulation of unphosphorylated BES1 in bri1-301 can be detected at 18°C and 22°C but not at 28°C (Yin et al., 2002) (Figure 4E). In summary, these results demonstrated that warmer temperature at 28°C inhibits BR signaling in bri1-301.
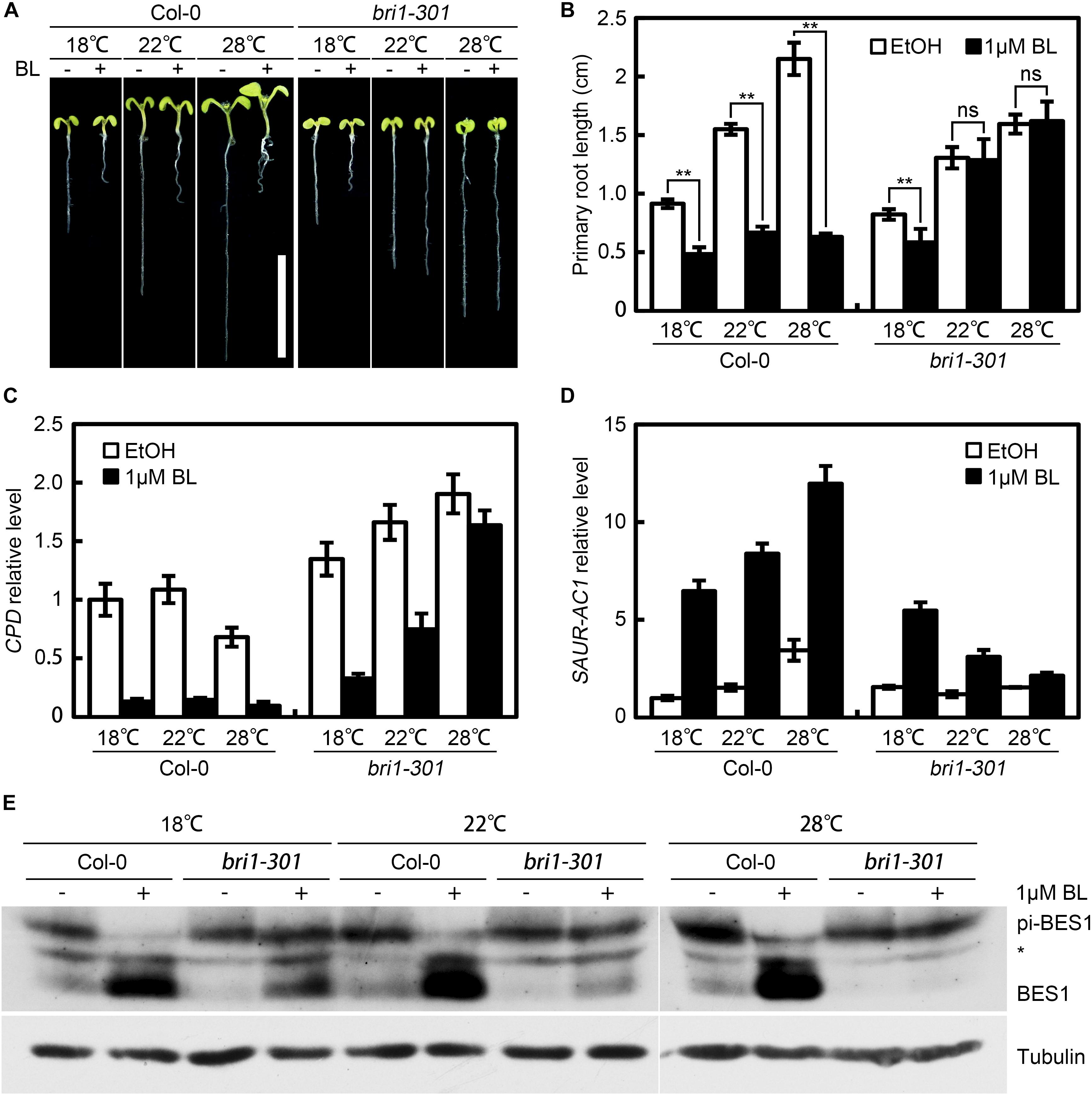
FIGURE 4. bri1-301 is sensitive to BL at lower temperature but insensitive to BL at higher temperature. (A) Root growth inhibition assay of Col-0 and bri1-301 at 18°C, 22°C, and 28°C. Seven-day-old seedlings grown on 1/2 MS media supplemented with or without 1 μM 24-epiBL were photographed. Scale bar represents 1 cm. (B) Measurements of primary root length of the seedlings as shown in (A). The asterisks indicate statistical significance evaluated by Student’s t-test (∗∗P < 0.01), ns represents not significant. Three independent biological replicates were carried out. Similar results were obtained. One of the representative results is shown. (C,D) Transcriptional responses of CPD (C) and SAUR-AC1 (D) to 24-epiBL treatment at three different temperatures. ACTIN2 was used as an internal control. Three independent biological replicates were carried out. Similar results were obtained. One of the representative results is shown. (E) BES1 phosphorylation analysis in response to exogenous 24-epiBL at different temperatures. The upper and lower bands represent phosphorylated and dephosphorylated BES1, respectively. The middle bands marked by an asterisk indicate non-specific signals generated by anti-BES1 serum. Tubulin was used as the loading control.
At Lower Temperature but Not at Warmer Temperature, BAK1 Can Promote the Growth of bri1-301
As co-receptors, BAK1 and its homologs play indispensable roles in the initiation of BR signal transduction (Gou et al., 2012). Overexpression of BAK1 can suppress the defective phenotype of weak but not null bri1 alleles (Li et al., 2002), suggesting the requirement of at least a partially functional BRI1 for the function of BAK1. To test whether bri1-301 is functional at warmer temperature, we overexpressed BAK1 in bri1-301 and analyzed the phenotypes of the transgenic plants at three aforementioned different temperatures. Our results indicated that overexpression of BAK1 can partially suppress the defective phenotype of bri1-301 at both 18°C and 22°C but not at 28°C (Figure 5). These genetic results confirmed that bri1-301 lost most of its biological function at 28°C.
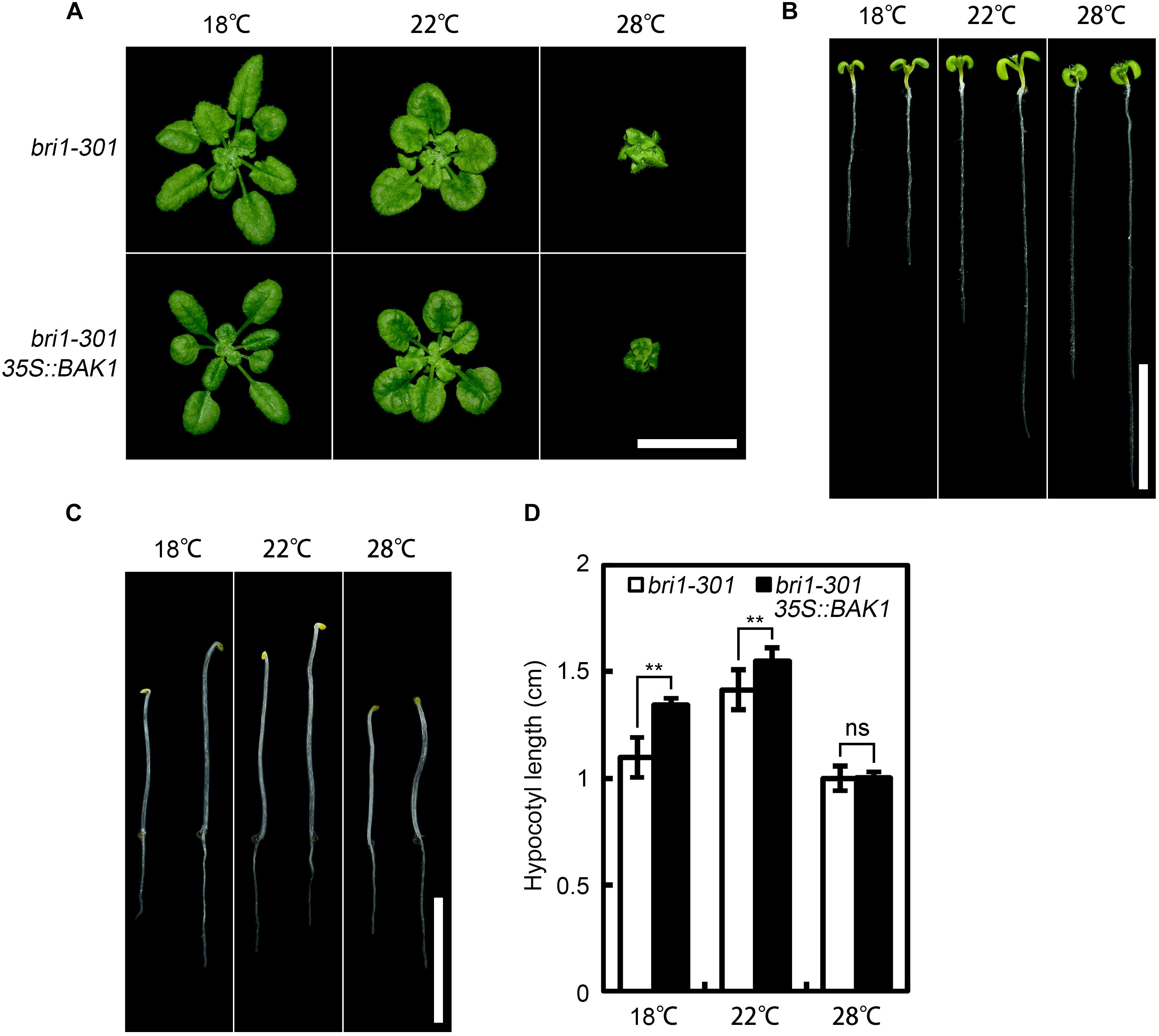
FIGURE 5. Overexpression of BAK1 can suppress bri1-301 at lower temperature but not at higher temperature. (A) Phenotypes of 4-week-old bri1-301 and 35S:BAK1 bri1-301 plants grown at 18°C, 22°C, and 28°C. (B) Phenotypes of 7-day-old light-grown seedlings on 1/2 MS plates at 18°C, 22°C, and 28°C. (C) Phenotypes of 4-day-old dark-grown seedlings on 1/2 MS plates at 18°C, 22°C, and 28°C. For each temperature panel shown in (B,C), the left seedling represents a bri1-301 and the right one represents a 35S:BAK1 bri1-301 transgenic plant. Scale bars represent 1 cm. (D) Measurements of hypocotyl length of the seedlings as shown in (C). The data shown are means and standard deviations. The asterisks indicate statistical significance evaluated by Student’s t-test (∗∗P < 0.01), ns represents not significant.
BR Signaling Initiation in bri1-301 Is Impaired Under a Warmer Temperature Condition
Successful BR signal initiation depends on the activation of the BR receptor BRI1 and co-receptor BAK1, which can be analyzed by their phosphorylation status after the treatment of the ligand, BL. We tested the phosphorylation levels of BRI1, bri1-301, and BAK1 in response to exogenous applied BL at different temperatures. The phosphorylation levels of BRI1 and BAK1 from wild type can be significantly induced by the BL treatment (Figure 6), as previously reported (Wang et al., 2005a). The phosphorylation levels of bri1-301 and BAK1 from bri1-301 can be induced by BL at 18°C. The degree of induction, however, is greatly reduced compared to their corresponding proteins from wild type. The induction of phosphorylation status of bri1-301 and BAK1 in bri1-301 is almost invisible at 28°C (Figure 6). These results further demonstrated that warmer temperature blocks the BR signaling initiation in bri1-301.
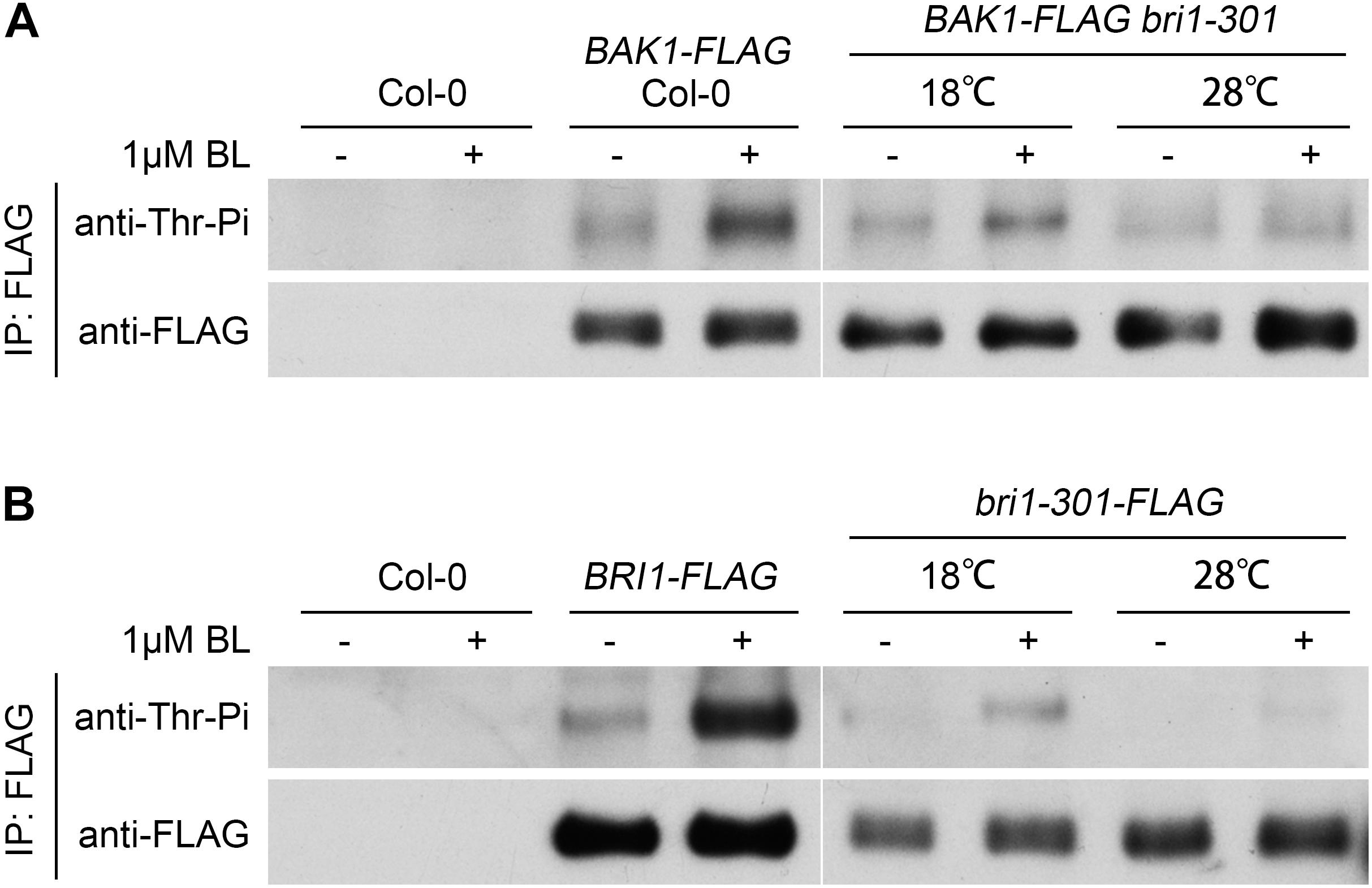
FIGURE 6. Exogenous application of 24-epiBL can induce the phosphorylation levels of both BAK1 and bri1-301 at 18°C, but at 28°C such effect is significantly reduced. (A) Western blotting analyses of BL-induced phosphorylation of BAK1 in Col-0 and bri1-301 at two indicated temperatures. (B) Western blotting analyses of BL-induced phosphorylation of BRI1 and bri1-301 at two indicated temperatures. Transgenic seedlings were treated with or without 1 μM 24-epiBL for 1.5 h at the same temperature conditions they grown.
bri1-301 Is Less Abundant Than BRI1 and Its Stability Is Greatly Reduced at 28°C
To elucidate the molecular mechanisms regulating bri1-301 functionality at various temperatures, we first evaluated the expression levels of BRI1 and bri1-301 and their responses to elevated temperatures. BRI1 and bri1-301 showed similar expression levels at 18°C and 22°C. At 28°C, bri1-301 is expressed even more than BRI1 (Supplementary Figure S4A). These results indicated that the severe phenotype of bri1-301 at 28°C is not caused by reducing transcription levels of bri1-301. The regulation is therefore likely at a post-transcriptional level. To test whether the protein abundance of the BR receptor has been altered in bri1-301 compared to wild type, we carried out a semi-quantitative Western blot analysis to detect the BR receptor proteins in wild type and bri1-301 by using an anti-BRI1 antibody. We found that the abundance of bri1-301 is much lower than BRI1, suggesting a reduced stability of bri1-301 than BRI1 (Supplementary Figure S4B). Due to low signal intensity of bri1-301 as shown in a Western blot analysis using an anti-BRI1 antibody (Supplementary Figure S4B), we therefore analyzed the impact of different temperatures on FLAG-tagged versions of BRI1 and bri1-301 proteins by using an anti-FLAG antibody. Our data showed that wild-type BRI1 protein level does not respond to elevated ambient temperature, whereas bri1-301 protein abundance is dramatically reduced at 28°C relative to those at 18°C or 22°C (Figures 7A,B and Supplementary Figure S4C). Transferring bri1-701 pBRI1::bri1-301-FLAG from 18 to 28°C for only 1 h can significantly reduce the abundance of bri1-301-FLAG (Figure 7C), suggesting a negative role of warmer temperature on bri1-301 protein stability. To further confirm this hypothesis, we evaluated BRI1/bri1-301-GFP protein abundance in BRI1-GFP and bri1-301-GFP transgenic seedlings treated with cycloheximide (CHX), a protein biosynthesis inhibitor, at three different temperatures. Our results showed that warmer temperature does not affect BRI1 stability. BRI1 abundance drops in similar patterns at different temperatures when protein biosynthesis is blocked (Figure 7D). In comparison, bri1-301 abundance drops much quicker under warmer temperature conditions (Figure 7E). Moreover, confocal microscopic analysis showed that the bri1-301 protein was accumulated more in the endosomal compartments compared to BRI1 when exposed to a warmer condition (Figure 8). In conclusion, bri1-301 appeared less stable than BRI1 and warmer temperature could further destabilize bri1-301.
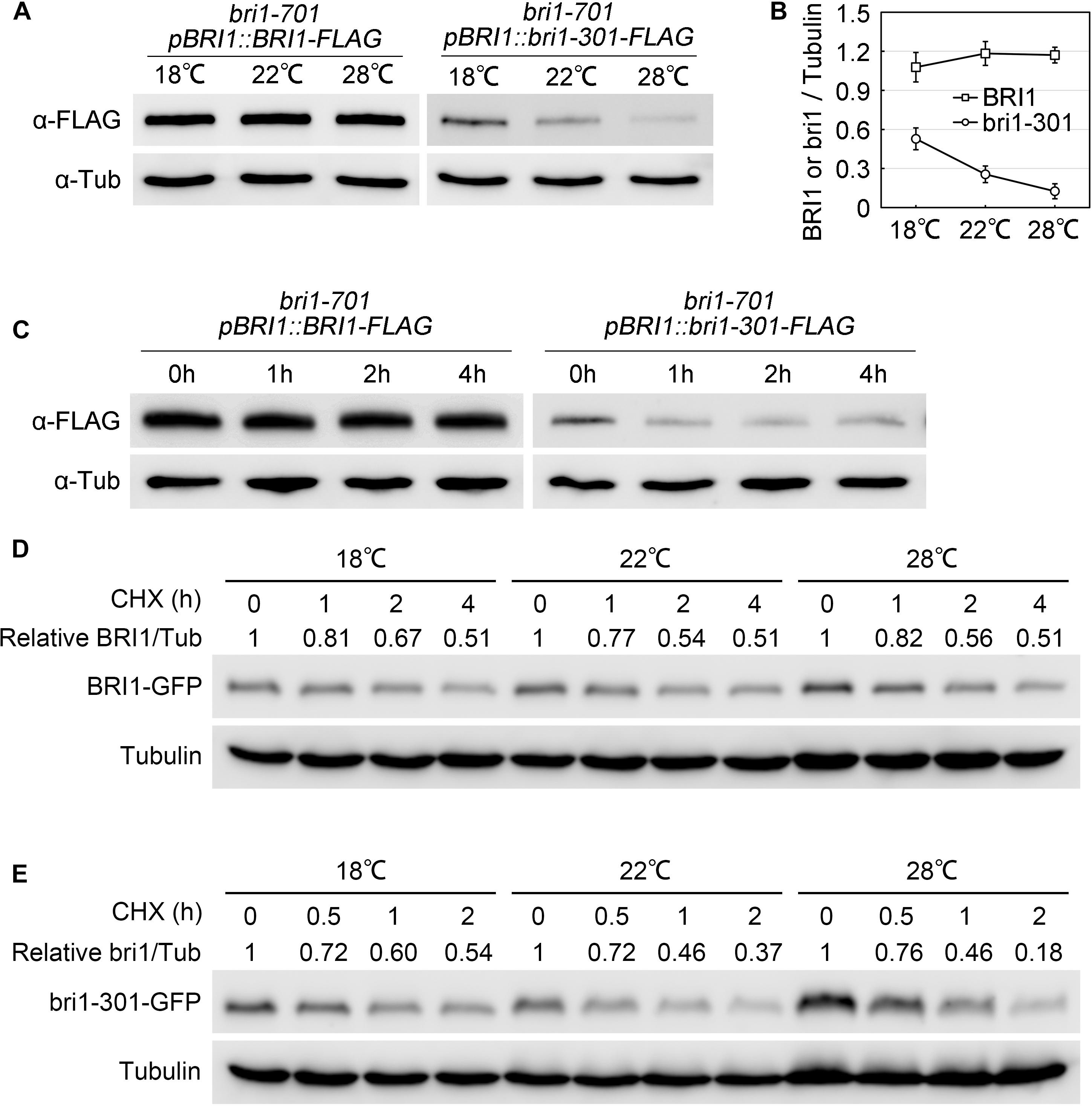
FIGURE 7. Stability of bri1-301 protein is negatively regulated by elevated temperature. (A) Western blot analyses of total protein extracts from aerial parts of 3-week-old pBRI1::BRI1-FLAG bri1-701 and pBRI1::bri1-301-FLAG bri1-701 transgenic plants. Tubulin was used as the loading control. (B) Relative BRI1 or bri1-301 protein levels were calculated as the average ratios (±SD) of Bri1/Tubulin signals in (A) from three independent experiments. (C) BRI1 and bri1-301 protein accumulation under short-term warm treatment. Three-week-old 18°C-grown pBRI1::BRI1-FLAG bri1-701 and pBRI1::bri1-301-FLAG bri1-701 transgenic plants were transferred into a 28°C growth chamber and the aerial parts were harvested at the indicated time points. Tubulin was used as the loading control. Three independent biological replicates were carried out. Similar results were obtained. One of the representative results is shown. (D,E) Abundance of BRI1 (D) and bri1-301 (E) in response to a time-course CHX treatment at three temperatures. Seven-day-old pBRI1::BRI1-GFP bri1-701 and pBRI1::bri1-301-GFP bri1-701 transgenic seedlings were used for the CHX treatment, total protein extracts were used for Western blot analyses. Tubulin was used as the loading control. Relative BRI1 or bri1-301 protein levels were calculated as ratios of BRI1/Tubulin.
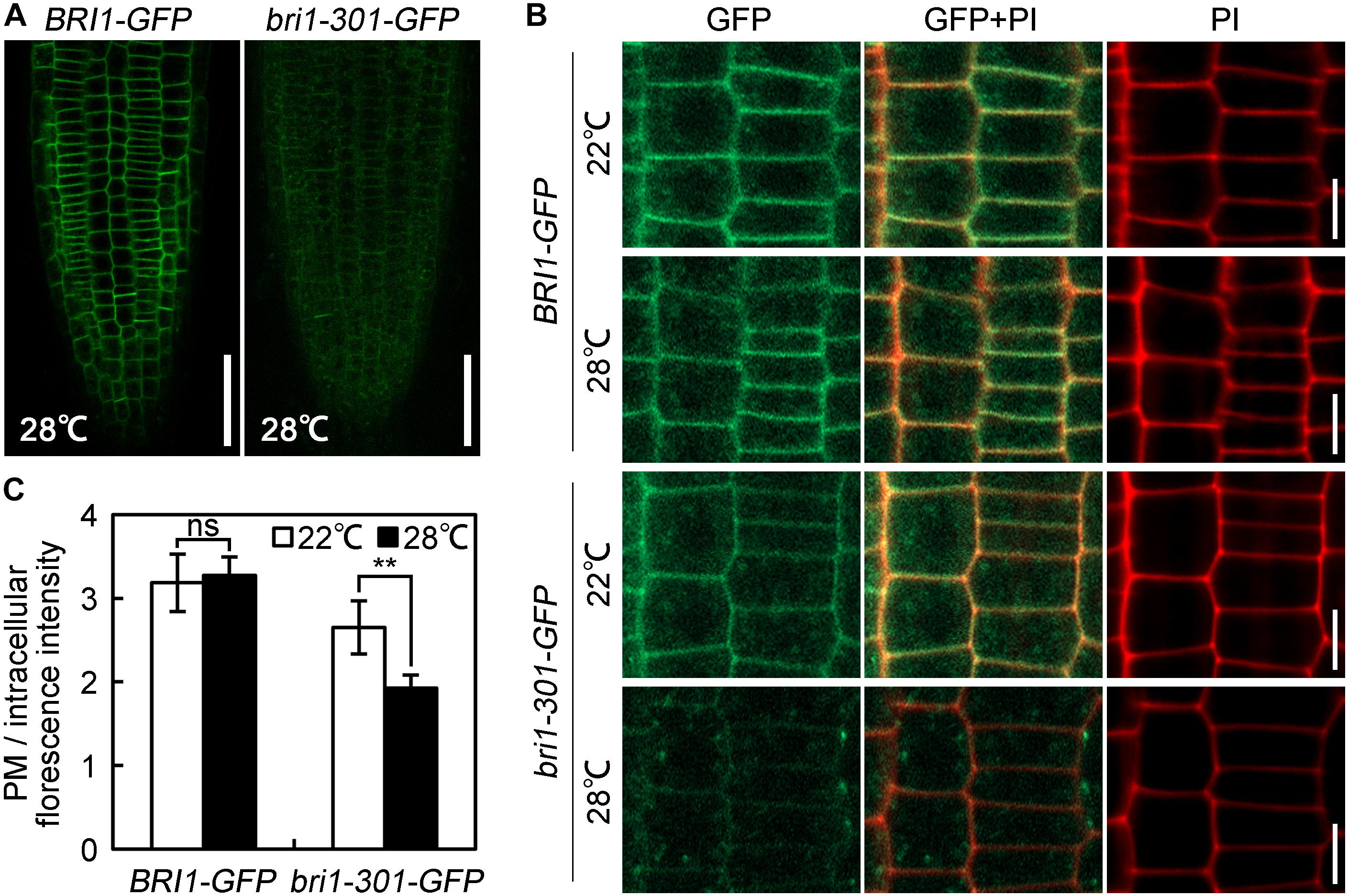
FIGURE 8. Warm-enhanced endocytosis of bri1-301 protein. (A) Confocal microscopic images of 28°C-grown seedlings. Roots were immediately scanned by confocal microscope after mounting. Scale bars represent 50 μm. (B) The subcellular localization of BRI1 and bri1-301 protein. Roots were stained with PI and then were immediately scanned by confocal microscope. Scale bar represents 10 μm. (C) Quantification of the ratio between the PM and the intracellular BRI1/bri1-301-GFP signal intensities from images shown in (B). The data shown are means and standard deviations. The asterisks indicate statistical significance evaluated by Student’s t-test (∗∗P < 0.01), ns represents not significant. At least 10 roots for each sample were analyzed. Similar results were obtained. One of the representative picture for each treatment is shown.
Discussion
Kinase Activity of bri1-301 Is Crucial for Its Biological Function
As a receptor kinase, BRI1 kinase activity is essential to the initiation of successful BR signaling (Friedrichsen et al., 2000; Wang et al., 2008; Oh et al., 2009; Jaillais et al., 2011). The crucial role of kinase activity for BRI1 function has been demonstrated by severe morphological defects of null bri1 mutant alleles, such as bri1-1, bri1-101, bri1-117, and bri1-703 (Clouse et al., 1996; Li and Chory, 1997; Friedrichsen et al., 2000; Sun et al., 2017). Consistently, mutants with partial BRI1 kinase activities, such as bri1-702, usually show weak defective phenotypes (Sun et al., 2017). Interestingly, in vitro kinase autophosphorylation assay indicated that bri1-8/108 and bri1-301 proteins do not show any kinase activities, but bri1-8/108 and bri1-301 plants exhibit even weaker defective phenotypes than bri1-702 (Noguchi et al., 1999; Xu et al., 2008; Sun et al., 2017). Because of these exceptions, there was an argument debating whether the kinase activity of BRI1 is truly essential to plant growth and development (Xu et al., 2008). In one of our previous reports, we showed that phosphorylation level of BAK1 in bri1-301 is much higher than that in bri1-701, a null T-DNA insertional mutant of BRI1 (Sun et al., 2017). In addition, the phosphorylation level of BAK1 in bri1-301 can be significantly induced by exogenously applied BL. These results indirectly suggested bri1-301 is a partially functional receptor in vivo (Sun et al., 2017). Here, we provide additional evidence to show that, under normal Arabidopsis laboratory growth conditions (22°C, 16 h light/8 h dark), bri1-301 mutant is able to respond to exogenously applied BL, indicated by ligand-induced both target gene feedback regulation and dephosphorylated BES1 accumulation analyses (Figures 4C–E). In vivo phosphorylation analysis showed that the weak phosphorylation level of bri1-301 can be detected and elevated by exogenous BL treatment (Figure 6). In addition, although the mRNA levels of bri1-301 and BRI1 are equivalent in bri1-301 and wild type (Supplementary Figure S4A), the bri1-301 protein abundance in bri1-301 is significantly reduced compared to BRI1 in wild type (Figures 7A,B and Supplementary Figure S4B), suggesting that bri1-301 is a highly unstable protein. These data imply that the reduced protein abundance and partially impaired kinase activity likely have caused the weak phenotype of bri1-301. Our analyses clarified the early question regarding the importance of BRI1 kinase activity (Xu et al., 2008).
Warmer Condition Destabilizes bri1-301 Receptor, Leading to Severer Defective Phenotype of bri1-301
During our phenotypic analyses of bri1-301, we accidentally identified that the morphological defects of bri1-301 is temperature-dependent. When grown at lower temperature (18°C), bri1-301 shows a subtle defective phenotype. Although the BRI1 protein level in bri1-301 is reduced to approximately 50% that of wild type, such amount of bri1-301 seems sufficient to maintain normal growth and development (Figures 1, 7A,B). When grown at warmer temperature (28°C), however, bri1-301 shows a severer defective phenotype similar to that of a strong bri1 allele, such as bri1-701 (Figure 1). The bri1-301 abundance is further reduced to about 10% the level of BRI1 in wild type (Figure 7). In addition, warmer temperature also increases internalization of bri1-301 (Figures 8B,C). Therefore, these results suggest that warm-temperature-induced bri1-301 protein internalization and degradation is likely the cause of the severer phenotype of bri1-301.
The bri1-301 Protein Partially Undergoes the ERQC Process
BRI1 was reported to undergo a highly conserved endoplasmic reticulum-mediated protein quality control (ERQC) mechanism before further trafficking to plasma membrane. Some missense BRI1 receptors with mutations in the extracellular domain, such as bri1-5 and bri1-9, are ER-retained and are subjected to ER-associated degradation (ERAD), resulting in low protein abundance (Liu and Li, 2014). Although bri1-301 also shows a decreased stability and low protein abundance, the quality control mechanisms of bri1-301 seem to be different from those of bri1-5 and bri1-9. Firstly, the amino acid substitution in bri1-301 occurs in the cytoplasmic domain, whereas the mutations in bri1-5 and bri1-9 are in the extracellular domain. The extracellular domain of BRI1 is toward to the lumen side of ER during trafficking. Secondly, bri1-301 is partially retained by glycan-dependent mechanism. Our results showed that bri1-301 displays a partial cleavage pattern when treated with endoglycosidase H (Endo H) (Supplementary Figure S5), an enzyme can remove high-mannose-type glycans of ER-localized glycoproteins but cannot cleave Golgi-processed glycans (Maley et al., 1989). The Endo H susceptibility of bri1-301 is much lower than bri1-5 and bri1-9, two well characterized Endo H substrates (Hong et al., 2009; Jin et al., 2009; Liu and Li, 2014). Thirdly, previous studies indicated that inactivating ERQC by disrupting EBS1/UGGT can suppress the defective phenotype of bri1-9 but not bri1-301 (Jin et al., 2007). In addition, our confocal analysis indicated that at 22°C, bri1-301-GFP, like BRI1-GFP, is mainly localized on the plasma membrane instead of retaining in ER (Figures 8B,C). Our observation suggests that the instability of bri1-301 is likely controlled by multiple mechanisms including the glycan-dependent ERQC.
Thermal-Enhanced bri1-301 Instability Suggests a Plasma Membrane Quality Control System in Plants
According to studies in yeast and mammalian cells, non-native membrane proteins are normally eliminated by ERQC systems. By contrast, the non-native membrane proteins escaped from the ERQC are preferentially eliminated by plasma membrane quality control (PMQC) mechanisms (Okiyoneda et al., 2011; Apaja and Lukacs, 2014). Many mutant variants of membrane proteins such as PM H+-ATPase (Pma1), α factor receptor (Ste2-3), and cystic fibrosis transmembrane conductance regulator (Δ508CFTR) are rapidly internalized and degraded from cell surface in a temperature-sensitive manner (Chang and Fink, 1995; Jenness et al., 1997; Sharma et al., 2001). The destabilizing point mutations in these membrane proteins are predominantly localized in the cytoplasmic and transmembrane segments (Okiyoneda et al., 2011). Our analysis indicated that bri1-301 can be partially recognized by ERQC machinery. As a result, a great deal of bri1-301 can escape from the ER and reach its destination, the plasma membrane under a normal Arabidopsis growing condition. Some similarities can be found between bri1-301 and well-studied animal PMQC proteins: (1) they share cytoplasmic mutations; (2) these mutations can decrease protein stability; and (3) they have thermal-enhanced mutant phenotype, protein internalization and degradation properties. Based on these findings, we propose the existence of PMQC system for BRI1 and possibly other RLKs in plants.
bri1-301 Can Serve as a Model to Study PMQC Mechanism in Plants
Although a number of PMQC proteins have been found in yeast and mammalian cells, it remains unclear how unfolded membrane proteins are recognized and removed from the plasma membrane (Okiyoneda et al., 2011). Recent studies proposed a general PMQC mechanism for removing the conformationally unstable proteins from the PM via ubiquitination, endocytosis, and lysosomal degradation (Okiyoneda et al., 2011; Apaja and Lukacs, 2014). In plants, PM-localized BRI1 has been reported to be ubiquitinated, internalized, and sorted to recycling or degradation machinery (Russinova et al., 2004; Geldner et al., 2007; Martins et al., 2015; Zhou et al., 2018). These processes implicate two plant U-box E3 ubiquitin ligases, PUB12 and PUB13 (Zhou et al., 2018); several components involved in endocytosis, AP-2, TPLATE, and GNOM (Irani et al., 2012; Di Rubbo et al., 2013; Gadeyne et al., 2014); and ALIX1, a cytoplasmic protein participating in vacuolar sorting (Cardona-Lopez et al., 2015). Therefore, it is intriguing to investigate the functional relevance between BRI1 membrane trafficking and its conformational change during BR perception and signaling initiation. As an unstable BR receptor, especially at warmer temperature, bri1-301 can be used as a model to study PMQC and its function in maintaining BRI1 integrity.
Author Contributions
JL conceived the research plans, designed the experiments, and edited the manuscript. MLv, MLi, WC, YW, CS and HY performed the experiments. MLv analyzed the data and prepared the manuscript draft. KH provided comments for the manuscript.
Funding
This project was supported by the National Natural Science Foundation of China (31530005 and 31470380 to JL).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank JL lab member Yao Xiao for preparing the anti-BES1 serum, and Professor Guang Wu from Shanxi Normal University for providing pBRI1::BRI1-GFP and pBRI1::bri1-301-GFP transgenic plants.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01620/full#supplementary-material
Footnotes
References
Apaja, P. M., and Lukacs, G. L. (2014). Protein homeostasis at the plasma membrane. Physiology 29, 265–277. doi: 10.1152/physiol.00058.2013
Belkhadir, Y., Durbak, A., Wierzba, M., Schmitz, R. J., Aguirre, A., Michel, R., et al. (2010). Intragenic suppression of a trafficking-defective brassinosteroid receptor mutant in Arabidopsis. Genetics 185, 1283–1296. doi: 10.1534/genetics.109.111898
Cardona-Lopez, X., Cuyas, L., Marin, E., Rajulu, C., Irigoyen, M. L., Gil, E., et al. (2015). ESCRT-III-associated protein ALIX mediates high-affinity phosphate transporter trafficking to maintain phosphate homeostasis in Arabidopsis. Plant Cell 27, 2560–2581. doi: 10.1105/tpc.15.00393
Chang, A., and Fink, G. R. (1995). Targeting of the yeast plasma membrane H + -ATPase: a novel gene AST1 prevents mislocalization of mutant Atpase to the vacuole. J. Cell Biol. 128, 39–49. doi: 10.1083/jcb.128.1.39
Clough, S. J., and Bent, A. F. (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. doi: 10.1046/j.1365-313x.1998.00343.x
Clouse, S. D. (1996). Molecular genetic studies confirm the role of brassinosteroids in plant growth and development. Plant J. 10, 1–8. doi: 10.1046/j.1365-313x.1996.10010001.x
Clouse, S. D., Langford, M., and Mcmorris, T. C. (1996). A brassinosteroid-insensitive mutant in Arabidopsis thaliana exhibits multiple defects in growth and development. Plant Physiol. 111, 671–678. doi: 10.1104/pp.111.3.671
Clouse, S. D., and Sasse, J. M. (1998). Brassinosteroids: essential regulators of plant growth and development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 427–451. doi: 10.1146/annurev.arplant.49.1.427
Di Rubbo, S., Irani, N. G., Kim, S. Y., Xu, Z. Y., Gadeyne, A., Dejonghe, W., et al. (2013). The clathrin adaptor complex AP-2 mediates endocytosis of brassinosteroid insensitive1 in Arabidopsis. Plant Cell 25, 2986–2997. doi: 10.1105/tpc.113.114058
Du, J. B., Zhao, B. L., Sun, X., Sun, M. Y., Zhang, D. Z., Zhang, S. S., et al. (2016). Identification and characterization of multiple intermediate alleles of the key genes regulating brassinosteroid biosynthesis pathways. Front. Plant Sci. 7:1893. doi: 10.3389/fpls.2016.01893
Falbel, T. G., Koch, L. M., Nadeau, J. A., Segui-Simarro, J. M., Sack, F. D., and Bednarek, S. Y. (2003). SCD1 is required for cell cytokinesis and polarized cell expansion in Arabidopsis thaliana. Development 130, 4011–4024. doi: 10.1242/dev.00619
Friedrichsen, D. M., Joazeiro, C. A., Li, J. M., Hunter, T., and Chory, J. (2000). Brassinosteroid-insensitive-1 is a ubiquitously expressed leucine-rich repeat receptor serine/threonine kinase. Plant Physiol. 123, 1247–1256. doi: 10.1104/pp.123.4.1247
Gadeyne, A., Sanchez-Rodriguez, C., Vanneste, S., Di Rubbo, S., Zauber, H., Vanneste, K., et al. (2014). The TPLATE adaptor complex drives clathrin-mediated endocytosis in plants. Cell 156, 691–704. doi: 10.1016/j.cell.2014.01.039
Geldner, N., Hyman, D. L., Wang, X. L., Schumacher, K., and Chory, J. (2007). Endosomal signaling of plant steroid receptor kinase BRI1. Genes Dev. 21, 1598–1602. doi: 10.1101/gad.1561307
Goda, H., Shimada, Y., Asami, T., Fujioka, S., and Yoshida, S. (2002). Microarray analysis of brassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 130, 1319–1334. doi: 10.1104/pp.011254
Gou, X. P., He, K., Yang, H., Yuan, T., Lin, H. H., Clouse, S. D., et al. (2010). Genome-wide cloning and sequence analysis of leucine-rich repeat receptor-like protein kinase genes in Arabidopsis thaliana. BMC Genomics 11:19. doi: 10.1186/1471-2164-11-19
Gou, X. P., Yin, H. J., He, K., Du, J. B., Yi, J., Xu, S. B., et al. (2012). Genetic evidence for an indispensable role of somatic embryogenesis receptor kinases in Brassinosteroid signaling. PLoS Genetics 8:e1002452. doi: 10.1371/journal.pgen.1002452
Grove, M. D., Spencer, G. F., Rohwedder, W. K., Mandava, N., Worley, J. F., Warthen, J. D., et al. (1979). Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature 281, 216–217. doi: 10.1038/281216a0
He, J. X., Gendron, J. M., Sun, Y., Gampala, S. S. L., Gendron, N., Sun, C. Q., et al. (2005). BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science 307, 1634–1638. doi: 10.1126/science.1107580
He, K., Xu, S. B., and Li, J. (2013). BAK1 directly regulates brassinosteroid perception and BRI1 activation. J. Integr. Plant Biol. 55, 1264–1270. doi: 10.1111/jipb.12122
Hong, Z., Jin, H., Fitchette, A. C., Xia, Y., Monk, A. M., Faye, L., et al. (2009). Mutations of an alpha1,6 mannosyltransferase inhibit endoplasmic reticulum-associated degradation of defective brassinosteroid receptors in Arabidopsis. Plant Cell 21, 3792–3802. doi: 10.1105/tpc.109.07028
Hong, Z., Jin, H., Tzfira, T., and Li, J. M. (2008). Multiple mechanism-mediated retention of a defective brassinosteroid receptor in the endoplasmic reticulum of Arabidopsis. Plant Cell 20, 3418–3429. doi: 10.1105/tpc.108.061879
Hong, Z., Kajiura, H., Su, W., Jin, H., Kimura, A., Fujiyama, K., et al. (2012). Evolutionarily conserved glycan signal to degrade aberrant brassinosteroid receptors in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 109, 11437–11442. doi: 10.1073/pnas.1119173109
Hothorn, M., Belkhadir, Y., Dreux, M., Dabi, T., Noel, J. P., Wilson, I. A., et al. (2011). Structural basis of steroid hormone perception by the receptor kinase BRI1. Nature 474, 467–471. doi: 10.1038/nature10153
Irani, N. G., Di Rubbo, S., Mylle, E., Van Den Begin, J., Schneider-Pizon, J., Hnilikova, J., et al. (2012). Fluorescent castasterone reveals BRI1 signaling from the plasma membrane. Nat. Chem. Biol. 8, 583–589. doi: 10.1038/nchembio.958
Jaillais, Y., Hothorn, M., Belkhadir, Y., Dabi, T., Nimchuk, Z. L., Meyerowitz, E. M., et al. (2011). Tyrosine phosphorylation controls brassinosteroid receptor activation by triggering membrane release of its kinase inhibitor. Genes Dev. 25, 232–237. doi: 10.1101/gad.2001911
Jenness, D. D., Li, Y., Tipper, C., and Spatrick, P. (1997). Elimination of defective alpha-factor pheromone receptors. Mol. Cell. Biol. 17, 6236–6245. doi: 10.1128/MCB.17.11.6236
Jin, H., Hong, Z., Su, W., and Li, J. M. (2009). A plant-specific calreticulin is a key retention factor for a defective brassinosteroid receptor in the endoplasmic reticulum. Proc. Natl. Acad. Sci. U.S.A. 106, 13612–13617. doi: 10.1073/pnas.0906144106
Jin, H., Yan, Z. Y., Nam, K. H., and Li, J. M. (2007). Allele-specific suppression of a defective brassinosteroid receptor reveals a physiological role of UGGT in ER quality control. Mol. Cell. 26, 821–830. doi: 10.1016/j.molcel.2007.05.015
Kim, T. W., Guan, S. H., Burlingame, A. L., and Wang, Z. Y. (2011). The CDG1 kinase mediates brassinosteroid signal transduction from BRI1 receptor kinase to BSU1 phosphatase and GSK3-like kinase BIN2. Mol. Cell. 43, 561–571. doi: 10.1016/j.molcel.2011.05.037
Kim, T. W., Guan, S. H., Sun, Y., Deng, Z., Tang, W., Shang, J. X., et al. (2009). Brassinosteroid signal transduction from cell-surface receptor kinases to nuclear transcription factors. Nat. Cell Biol. 11, 1254–1260. doi: 10.1038/ncb1970
Lee, H. S., Kim, Y., Pham, G., Kim, J. W., Song, J. H., Lee, Y., et al. (2015). Brassinazole resistant 1 (BZR1)-dependent brassinosteroid signalling pathway leads to ectopic activation of quiescent cell division and suppresses columella stem cell differentiation. J. Exp. Bot. 66, 4835–4849. doi: 10.1093/jxb/erv316
Li, J., Wen, J. Q., Lease, K. A., Doke, J. T., Tax, F. E., and Walker, J. C. (2002). BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 110, 213–222. doi: 10.1016/S0092-8674(02)00812-7
Li, J. M., and Chory, J. (1997). A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction. Cell 90, 929–938. doi: 10.1016/S0092-8674(00)80357-8
Li, J. M., and Nam, K. H. (2002). Regulation of brassinosteroid signaling by a GSK3/SHAGGY-like kinase. Science 295, 1299–1301. doi: 10.1126/science.1065769
Li, J. M., Nam, K. H., Vafeados, D., and Chory, J. (2001). BIN2, a new brassinosteroid-insensitive locus in Arabidopsis. Plant Physiol. 127, 14–22. doi: 10.1104/pp.127.1.14
Liu, Y., and Li, J. M. (2014). Endoplasmic reticulum-mediated protein quality control in Arabidopsis. Front. Plant Sci. 5:162. doi: 10.3389/fpls.2014.00162
Maley, F., Trimble, R. B., Tarentino, A. L., and Plummer, T. H. (1989). Characterization of glycoproteins and their associated oligosaccharides through the use of endoglycosidases. Anal. Biochem. 180, 195–204. doi: 10.1016/0003-2697(89)90115-2
Martins, S., Dohmann, E. M., Cayrel, A., Johnson, A., Fischer, W., Pojer, F., et al. (2015). Internalization and vacuolar targeting of the brassinosteroid hormone receptor BRI1 are regulated by ubiquitination. Nat. Commun. 6:6151. doi: 10.1038/ncomms7151
Mora-Garcia, S., Vert, G., Yin, Y. H., Cano-Delgado, A., Cheong, H., and Chory, J. (2004). Nuclear protein phosphatases with Kelch-repeat domains modulate the response to bras sino steroids in Arabidopsis. Genes Dev. 18, 448–460. doi: 10.1101/gad.1174204
Nam, K. H., and Li, J. M. (2002). BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell 110, 203–212. doi: 10.1016/S0092-8674(02)00814-0
Noguchi, T., Fujioka, S., Choe, S., Takatsuto, S., Yoshida, S., Yuan, H., et al. (1999). Brassinosteroid-insensitive dwarf mutants of Arabidopsis accumulate brassinosteroids. Plant Physiol. 121, 743–752. doi: 10.1104/pp.121.3.743
Oh, M. H., Wang, X. F., Kota, U., Goshe, M. B., Clouse, S. D., and Huber, S. C. (2009). Tyrosine phosphorylation of the BRI1 receptor kinase emerges as a component of brassinosteroid signaling in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 106, 658–663. doi: 10.1073/pnas.0810249106
Okiyoneda, T., Apaja, P. M., and Lukacs, G. L. (2011). Protein quality control at the plasma membrane. Curr. Opin. Cell Biol. 23, 483–491. doi: 10.1016/j.ceb.2011.04.012
Quint, M., Ito, H., Zhang, W., and Gray, W. M. (2005). Characterization of a novel temperature-sensitive allele of the CUL1/AXR6 subunit of SCF ubiquitin-ligases. Plant J. 43, 371–383. doi: 10.1111/j.1365-313X.2005.02449.x
Russinova, E., Borst, J. W., Kwaaitaal, M., Cano-Delgado, A., Yin, Y., Chory, J., et al. (2004). Heterodimerization and endocytosis of Arabidopsis brassinosteroid receptors BRI1 and AtSERK3 (BAK1). Plant Cell 16, 3216–3229. doi: 10.1105/tpc.104.025387
Santiago, J., Henzler, C., and Hothorn, M. (2013). Molecular mechanism for plant steroid receptor activation by somatic embryogenesis co-receptor kinases. Science 341, 889–892. doi: 10.1126/science.1242468
Shang, Y., Lee, M. M., Li, J. M., and Nam, K. H. (2011). Characterization of cp3 reveals a new bri1 allele, bri1-120, and the importance of the LRR domain of BRI1 mediating BR signaling. BMC Plant Biol. 11:8. doi: 10.1186/1471-2229-11-8
Sharma, M., Benharouga, M., Hu, W., and Lukacs, G. L. (2001). Conformational and temperature-sensitive stability defects of the Delta F508 cystic fibrosis transmembrane conductance regulator in post-endoplasmic reticulum compartments. J. Biol. Chem. 276, 8942–8950. doi: 10.1074/jbc.M009172200
She, J., Han, Z. F., Kim, T. W., Wang, J. J., Cheng, W., Chang, J. B., et al. (2011). Structural insight into brassinosteroid perception by BRI1. Nature 474, 472–476. doi: 10.1038/nature10178
Smith, M. H., Ploegh, H. L., and Weissman, J. S. (2011). Road to ruin: targeting proteins for degradation in the endoplasmic reticulum. Science 334, 1086–1090. doi: 10.1126/science.1209235
Sun, C., Yan, K., Han, J. T., Tao, L., Lv, M. H., Shi, T., et al. (2017). Scanning for new BRI1 mutations via TILLING analysis. Plant Physiol. 174, 1881–1896. doi: 10.1104/pp.17.00118
Sun, Y., Fan, X. Y., Cao, D. M., Tang, W. Q., He, K., Zhu, J. Y., et al. (2010). Integration of brassinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis. Dev. Cell 19, 765–777. doi: 10.1016/j.devcel.2010.10.010
Szekeres, M., Nemeth, K., Koncz-Kalman, Z., Mathur, J., Kauschmann, A., Altmann, T., et al. (1996). Brassinosteroids rescue the deficiency of CYP90, a cytochrome P450, controlling cell elongation and de-etiolation in Arabidopsis. Cell 85, 171–182. doi: 10.1016/S0092-8674(00)81094-6
Tang, W. Q., Kim, T. W., Oses-Prieto, J. A., Sun, Y., Deng, Z. P., Zhu, S. W., et al. (2008). BSKs mediate signal transduction from the receptor kinase BRI1 in Arabidopsis. Science 321, 557–560. doi: 10.1126/science.1156973
Tang, W. Q., Yuan, M., Wang, R. J., Yang, Y. H., Wang, C. M., Oses-Prieto, J. A., et al. (2011). PP2A activates brassinosteroid-responsive gene expression and plant growth by dephosphorylating BZR1. Nat. Cell Biol. 13, 124–131. doi: 10.1038/ncb2151
Vert, G., Nemhauser, J. L., Geldner, N., Hong, F., and Chory, J. (2005). Molecular mechanisms of steroid hormone signaling in plants. Annu. Rev. Cell Dev. Biol. 21, 177–201. doi: 10.1146/annurev.cellbio.21.090704.151241
Vilarrasa-Blasi, J., Gonzalez-Garcia, M. P., Frigola, D., Fabregas, N., Alexiou, K. G., Lopez-Bigas, N., et al. (2014). Regulation of plant stem cell quiescence by a brassinosteroid signaling module. Dev. Cell 30, 36–47. doi: 10.1016/j.devcel.2014.05.020
Wang, X., Goshe, M. B., Soderblom, E. J., Phinney, B. S., Kuchar, J. A., Li, J., et al. (2005a). Identification and functional analysis of in vivo phosphorylation sites of the Arabidopsis brassinosteroid-insensitive1 receptor kinase. Plant Cell 17, 1685–1703. doi: 10.1105/tpc.105.031393
Wang, X., Li, X., Meisenhelder, J., Hunter, T., Yoshida, S., Asami, T., et al. (2005b). Autoregulation and homodimerization are involved in the activation of the plant steroid receptor BRI1. Dev. Cell 8, 855–865. doi: 10.1016/j.devcel.2005.05.001
Wang, X. F., Kota, U., He, K., Blackburn, K., Li, J., Goshe, M. B., et al. (2008). Sequential transphosphorylation of the BRI1/BAK1 receptor kinase complex impacts early events in brassinosteroid signaling. Dev. Cell 15, 220–235. doi: 10.1016/j.devcel.2008.06.011
Wang, X. L., and Chory, J. (2006). Brassinosteroids regulate dissociation of BKI1, a negative regulator of BRI1 signaling, from the plasma membrane. Science 313, 1118–1122. doi: 10.1126/science.1127593
Wang, Z., Cui, D., Liu, J., Zhao, J., Liu, C., Xin, W., et al. (2017). Arabidopsis ZED1-related kinases mediate the temperature-sensitive intersection of immune response and growth homeostasis. New Phytol. 215, 711–724. doi: 10.1111/nph.14585
Wang, Z. Y., Nakano, T., Gendron, J., He, J. X., Chen, M., Vafeados, D., et al. (2002). Nuclear-localized BZR1 mediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis. Dev. Cell 2, 505–513. doi: 10.1016/S1534-5807(02)00153-3
Xu, W. H., Huang, J., Li, B. H., Li, J. Y., and Wang, Y. H. (2008). Is kinase activity essential for biological functions of BRI1? Cell Res. 18, 472–478. doi: 10.1038/cr.2008.36
Yin, Y. H., Wang, Z. Y., Mora-Garcia, S., Li, J. M., Yoshida, S., Asami, T., et al. (2002). BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 109, 181–191. doi: 10.1016/S0092-8674(02)00721-3
Yu, X. F., Li, L., Zola, J., Aluru, M., Ye, H. X., Foudree, A., et al. (2011). A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Arabidopsis thaliana. Plant J. 65, 634–646. doi: 10.1111/j.1365-313X.2010.04449.x
Zhang, L. Y., Bai, M. Y., Wu, J., Zhu, J. Y., Wang, H., Zhang, Z., et al. (2009). Antagonistic HLH/bHLH transcription factors mediate brassinosteroid regulation of cell elongation and plant development in rice and Arabidopsis. Plant Cell 21, 3767–3780. doi: 10.1105/tpc.109.070441
Zhao, B. L., Lv, M. H., Feng, Z. X., Campbell, T., Liscum, E., and Li, J. (2016). Twisted dwarf 1 associates with brassinosteroid-insensitive 1 to regulate early events of the brassinosteroid signaling pathway. Mol. Plant 9, 582–592. doi: 10.1016/j.molp.2016.01.007
Zhou, J. G., Liu, D. R., Wang, P., Ma, X. Y., Lin, W. W., Cheng, S. X., et al. (2018). Regulation of Arabidopsis brassinosteroid receptor BRI1 endocytosis and degradation by plant U-box PUB12/PUB13-mediated ubiquitination. Proc. Natl. Acad. Sci. U.S.A. 115, E1906–E1915. doi: 10.1073/pnas.1712251115
Keywords: Arabidopsis, BRI1, protein folding, endoplasmic reticulum quality control, plasma membrane quality control
Citation: Lv M, Li M, Chen W, Wang Y, Sun C, Yin H, He K and Li J (2018) Thermal-Enhanced bri1-301 Instability Reveals a Plasma Membrane Protein Quality Control System in Plants. Front. Plant Sci. 9:1620. doi: 10.3389/fpls.2018.01620
Received: 22 July 2018; Accepted: 18 October 2018;
Published: 06 November 2018.
Edited by:
Laigeng Li, Shanghai Institutes for Biological Sciences (CAS), ChinaReviewed by:
Hong-Wei Xue, Shanghai Institutes for Biological Sciences (CAS), ChinaJianming Li, University of Michigan, United States
Copyright © 2018 Lv, Li, Chen, Wang, Sun, Yin, He and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jia Li, lijia@lzu.edu.cn